GNPL_Supplementary Material_Template_Word_XP_2003
advertisement

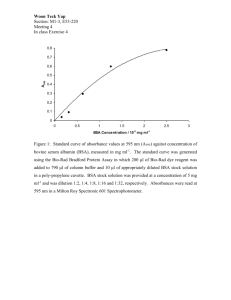
SUPPLEMENTARY MATERIAL Decreased glycation and structural protection properties of γglutamyl-S-allyl-cysteine peptide isolated from fresh garlic scales (Allium sativum L.) Dehong Tana, Yao Zhangb, Lulu Chena, Ling Liua, Xuan Zhanga, Zhaoxia Wua, Bing Baia* and Shujuan Jia a College of Food Science, Shenyang Agricultural University, Shenyang, Liaoning Province 110866, P.R.China b Shenyang Normal University, Shenyang, Liaoning Province 110034, P.R.China Contact details of authors: Dehong Tan: tandehongsy@126.com Yao Zhang: sudongbaihe@163.com Lulu Chen: 412246090@qq.com Ling Liu: liuling4568@sina.com Xuan Zhang: zhxllm@163.com Zhaoxia Wu: wuzxsau@163.com Bing Bai: baibing@hotmail.com Shujuan Ji: jsjsyau@sina.com This work was supported by the [National Natural Science Foundation of China] under Grant [31101220]; [Key Projects in the National Science and Technology Pillar Program of China] under Grant [2012BAD38B07]. Running headers of author names: D. Tan, Y. Zhang, L. Chen, L. Liu, X. Zhang, Z. Wu, B. Bai, S. Ji *Corresponding author. Email: baibing@hotmail.com Decreased glycation and structural protection properties of γglutamyl-S-allyl-cysteine peptide isolated from fresh garlic scales (Allium sativum L.) The antiglycative effect of γ-glutamyl-S-allyl-cysteine (GSAC) peptide isolated from fresh garlic scales was investigated in the bovine serum albumin (BSA)/glucose system. GSAC inhibited the increase of fluorescence intensity at about 440 nm in a concentration-dependent manner and reduced reacted free lysine side chains by 10.9%, 24.7% and 37.7%, as the GSAC concentrations increased from 0.1 to 2.5 mg ml-1. Glycation specific decline in BSA -helix content (from 61.3% to 55.6%) and increase in β-sheet (from 2.1% to 5.4%) were prevented by GSAC (2.5 mg ml-1) in vitro, implying its stabilization effect. GSAC treatment (2.5 mg ml-1) suppressed protein crosslinking to form polymers. Additionally, GSAC (10, 40, and 160 g ml-1) showed radical-scavenging and metal-chelating capacities. In conclusion, GSAC has an antiglycative effect, which may involve its radical-scavenging and metal chelating capacities. Keywords: Maillard reaction; advanced glycation end products; chelating; radical-scavenging; organosulfur peptide 1. Experimental 1.1. Plant Material The sample of Allium sativum L. was identified by Prof Baodong Wei of the College of Food Science, Shenyang Agricultural University. Fresh garlic bulbs harvested in May 2013 were obtained from a local market of Xinmin, Shenyang, Liaoning Province, China. They were freeze dried and stored at –70°C until used for analysis. A voucher specimen (GAR 5109) was deposited at the Key Laboratory of Fruits and Vegetables of the Shenyang Agricultural University. 1.2. Isolation and identification of GSAC from fresh garlic scales GSAC was isolated from fresh garlic scales according to the methods of Lawson et al. (1991) and Kubec & Musah (2005) with slight modifications. Briefly, whole garlic scales (1000 g) were carefully cleaned and incubated in ethyl alcohol (800 ml) at 70 °C for 5 min. The pieces were homogenized using a blender and the slurries obtained were filtered. The crude extracts were concentrated to about 300 ml by vacuum evaporation (40 °C). The pH value of extracts was adjusted to 2.0 by addition of 2 M HCl. The precipitate was filtered off and the filtrate was loaded on a column (60 cm × 5.0 cm) of Dowex 50 × 4 - 400 cation-exchange resin (SigmaAldrich, Shanghai, China) saturated with H+. After washed the column with H2O (1000 ml), the dipeptide fraction was eluted with 1000 ml of 1 M NH4OH. The yellowish eluent obtained was concentrated to about 300 mL by vacuum evaporation at 60 °C. The eluate was adjusted to pH 7.0 with glacial acetic acid and subjected to a column (40 cm × 5.0 cm) of Dowex 2 × 8 - 200 anion-exchange resin (SigmaAldrich, Shanghai, China) saturated with OH+. The column was washed with 800 ml of H2O and the dipeptide fraction was eluted with 10% acetic acid (800 ml). The eluate fraction was lyophilized to a white powder which was further purified using a preparative liquid chromatography (Hitachi Elite D-2000, Japan), equipped with a C-18 prep column (250 mm × 9.4 mm i.d. 5 μm, Agilent, USA) and with water/MeOH (2:3, v/v) as the mobile phase. The fraction eluting at 9.3 min was repeatedly collected. From 1 kg of fresh garlic scales, 377 mg dry weight of GSAC was obtained by freeze-drying as a white hygroscopic powder. The identity of GSAC was confirmed by comparisons of HPLC-MS (1100LC/MSD Agilent, USA), 1 H NMR and 13C NMR (AVANCE III HD Spectrometer, Brucker, Germany) data with published data (Mütsch-Eckner et al. 1992). The powder obtained in the dipeptide-containing fraction was confirmed to yield the expected [M-H] –of 288.8 (GASC, C11H18O5N2S, 290.0937) on HPLC-MS (Figure S1) analysis. The 1H and 13C NMR spectra (Figure S2) of GASC was confirmed as follows. 1H NMR (600 MHz, D2O), δ: 2.32 (1H, dd, J=7.2, 9.0 Hz, CH2CH2CH-a), 2.43 (1H, dd, J=6.0, 8.4 Hz, CH2CH2CH-b), 2.66 (2H, s, CH2CH2CH), 2.74~2.78 (H, m, SCH2CH-a), 2.82~2.85 (H, m, SCH2CH-b), 3.21 (2H, d, J=7.2 Hz, SCH2CH=CH2), 3.45~3.47 (1H, m, Glu-CH), 4.72~4.83 (1H, m, Cys-CH), 5.17~5.22 (2H, m, SCH2CH=CH2), 5.82~5.89 (1H, m, SCH2CH=CH2). 13 C NMR (600 MHz; D2O),δ:27.8 (CH2CH2CH), 32.6 (CH2CH2CH), 36.4 (SCH2CH), 37.9 (SCH2CH=CH2), 57.7 (Glu-CH, Cys-CH), 120.2 (SCH2CH=CH2), 136.9 (SCH2CH=CH2), 182.5 (Glu-COOH), 183.2 (NHCOCH2), 184.4 (CysCOOH). 1.3. Preparation of the BSA/glucose reaction system The reaction system was prepared as described previously (Peng et al. 2008). Briefly, 0.2 mg ml-1 BSA was incubated at 37 °C in a solution of 2 mg ml-1 D-glucose in 0.1 M phosphate buffer (pH 7.4) in the absence (control) or presence of different concentrations (0,1, 0,5, 2.5 mg ml-1) of GSAC, with added NaN3 (final concentration 0.2%). After 7 d, BSA-glucose solutions were filtered through low protein-binding filters (0.22-μm filter unit, Merck-Millipore, Billerica, MA) and dialyzed against autoclaved phosphate-buffered saline (PBS) at 4 °C to remove free glucose molecules. BSA solution (0.2 mg ml-1) incubated under the same conditions was used as the blank. Protein concentrations were determined by the Pierce-BCA protein assay (Thermo Scientific, China) calibrated with a BSA standard. Optical density of glycation samples were recorded by measuring the absorbance at wavelength of 190–600 nm in a PERSEE TU-1810 spectrophotometer. 1.4. Analysis of AGE formation Using methods described by Wu et al. (2009), AGE-related autofluorescence of relevant samples (0.2 mg ml-1) was analyzed after excitation at 370 nm and monitoring the emission at 400–500 nm and at 360–380 nm using a spectrofluorometer (Hitachi F-4600, Japan). 1.5. Determination of free lysine residues Determination of free lysine residues was accomplished using the 2-4-6trinitrobenzene sulfonic acid (TNBSA) method (Sattarahmady et al. 2008). Briefly, 4% NaHCO3 and 0.1% TNBSA were added to the protein solution, reacted at 37°C for 1 h, and solubilized with 10% sodium dodecyl sulfonate (SDS); 1 M HCl was added to stop the reaction. Ultraviolet absorbance was then measured at 335 nm using a spectrophotometer. 1.6. Congo Red assay The Congo Red binding assay was performed according to methods described by Miroliaei et al. (2011). In brief, 800 μl of protein solution (100 μM) was incubated with 200 μl of Congo Red solution (100 μM Congo Red in 10% PBS-ethanol). Absorbance was recorded at 530 nm with a spectrophotometer. 1.7. Circular dichroism (CD) spectropolarimetry A quantitative analysis of the secondary structure elements upon glycation was performed within far-UV CD spectra (190–240 nm) at room temperature on a JASCO-810 spectropolarimeter (Japan) using solutions with a BSA concentration of 0.2 mg ml-1. Sample groups with the exception of native BSA were incubated in Water-jacket constant temperature incubator at 37 °C for 7 d to glycation. Results are expressed as molar ellipticity [Θ] (mdeg). The molar ellipticity was determined as [Θ] = mdeg/(cl), where l, is the light path length in millimeter, c, is the protein concentration in mM, and Θ is the measured ellipticity in degrees at the relevant wavelength. The relative percentages of the secondary structure elements were estimated using SELCON3 software. 1.8. Gel electrophoresis assay Protein cross-linking was determined by SDS polyacrylamide gel electrophoresis (SDS-PAGE, 10% polyacrylamide gels) (Ahmad et al. 2007). Briefly, protein samples (native BSA, glycated BSA control, and glycated BSA in the 10 mg/ml GSAC treatment group) were diluted with loading buffer (containing 0.1% 2mercaptoethanol) and boiled for 5 min. Samples (containing 10 μg protein) were loaded into the wells and subjected to electrophoresis (Bio-Rad PowerPace Basic, Hercules, USA). The gels were removed and stained in 2.5 g kg-1 Coomassie Blue R solution (50% methanol and 10% acetic acid) for 0.5 h and then destained in a solution containing 25% methanol and 7% acetic acid for more than 3 h. 1.9. Evaluation of 2,2-diphenyl-1-picrylhydrazyl (DPPH) radical–scavenging capacity The DPPH radical-scavenging capacity was assessed using the microplate method (Ho et al. 2010). Briefly, different concentrations (10, 40, and 160 g ml-1) of GSAC (20 l) were reacted with 180 l of 0.2 mM DPPH methanolic solution in a 96-well microplate for 5 min. Absorbance at 520 nm was then measured using a microtiter plate reader. 1.10. Evaluation of metal-chelating capacity Metal-chelating capacity was determined according to methods described by Ho et al. (2010). Different concentrations (10, 40, and 160 g ml-1) of GSAC were reacted with 40 M ferrous chloride and 200 M ferrozine at ambient temperature for 10 min. Absorbance at 540 nm was then determined using a microtiter plate reader. 1.11. Statistical analysis The experiment was performed in 3 independent replicates. Data were analyzed using the statistical analysis system software package SPSS 16.0. Analyses of variance were performed using ANOVA. Significant differences between means were determined using Duncan’s multiple range tests. References Ahmad MS, Pischetsrieder M, Ahmed N. 2007. Aged garlic extract and S-allyl cysteine prevent formation of advanced glycation endproducts. Eur J Pharmacol. 561:32–38. Ho SC, Wu SP, Lin SM, Tang YL. 2010. Comparison of anti-glycation capacities of several herbal infusions with that of green tea. Food Chem. 122:768–774. Kubec R, Musah RA. 2005. γ-Glutamyl dipeptides in Petiveria alliacea. Phytochem. 66:2494–2497. Lawson LD, Wang Z-yJ, Hughes BG. 1991. γ-Glutamyl-S-alkylcysteines in garlic and other Allium spp.: Precursors of age-dependent trans-1-propenyl thiosulfinates. J Nat Prod. 54:436–444. Miroliaei M, Khazaei S, Moshkelgosha S, Shirvani M. 2011. Inhibitory effects of Lemon balm (Melissa officinalis L.) extract on the formation of advanced glycation end products. Food Chem. 129:267–271. Mütsch-Eckner M, Meier B, Wright AD, Sticher O. 1992. γ-glutamyl peptides from Allium sativum bulbs. Phytochem. 31:2389–2391. Peng X, Zheng Z, Cheng K, Shan F, Ren G, Chen F, Wang M. 2008. Inhibitory effect of mung bean extract and its constituents vitexin and isovitexin on the formation of advanced glycation endproducts. Food Chem. 106:475–481. Sattarahmady N, Moosavi-Movahedi AA, Habibi-Rezaei M, Ahmadian S, Saboury AA, Heli H, Sheibani N. 2008. Detergency effects of nanofibrillar amyloid formation on glycation of human serum albumin. Carbohyd Res. 343:2229–2234. Wu JW, Hsieh CL, Wang HY, Chen HY. 2009. Inhibitory effects of guava (Psidium guajava L.) leaf extracts and its active compounds on the glycation process of protein. Food Chem. 113:78–84. Figure S1. HPLC-MS-MS of GSAC. a: MS of GSAC, b: MS2 of GSAC with precursor ion of 288.7. isolated from fresh garlic scales. Figure S2. 1H NMR (a) and 13C NMR (b) spectra of GSAC. 1 2 Figure S1. Figure S2.