Supplementary material - Springer Static Content Server
advertisement

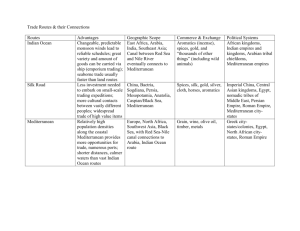
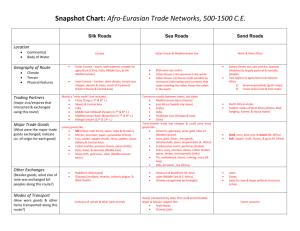
Supplementary material Supplementary text A: The Adriatic Sea was formed with the final rise of the Alps 20 Million Years ago (Ma). At the same time, the Apulian Plate collided with the Adriatic Plate (Battaglia et al. 2004). 10 Ma ago, there was still a connection between the Eastern and the Western Mediterranean Sea which was closed, before the Messinian crisis occurred. During the Messinian age (7.2-5.3 Ma), the Mediterranean Sea almost dried up because of the closure of the Strait of Gibraltar (Hsü et al. 1973, Hsü 1974). At the beginning of the following Pliocene (5.3 Ma), the Strait of Gibraltar reopened which resulted in a flooding of the Mediterranean Sea (Hsü 1974, McKenzie 1999, Bianchi and Morri 2000) and a re-immigration of species from the Atlantic Ocean. These species are assumed to be the basic forerunners of the current Mediterranean benthos, which presents a strong similarity with that currently populating the Eastern Atlantic Ocean (for review see Boudouresque 2004). Supplementary text B: Patterns of speciation and histories of colonization of certain Mediterranean basins are obviously unique for any taxon. As these colonizations took place at different times and interacted in a highly complex way, it appears often difficult or even impossible to reconstruct the ancestral courses of immigration and final isolation of putative stem species from which the recent stock of Mediterranean endemics evolved. Nevertheless, there are more examples of Mediterranean macrozoobenthic ‘neo-endemics’, besides spadellid Chaetognatha. In principle, some boreal immigrants of the Pleistocene survived periodic warming of epipelagic and neritic waters by sticking to bathybenthic grounds. One example for this deep-sea migration is the recent presence of the gastropod Buccinum undatum in the Western Mediterranean basin on muddy sediment below 1.000 m depth (Por and Dimentman 2006). Many other taxa, among them brachiopods and deep-water fishes, had same habitat segregation strategies, but became however extinct and can be found in deposits of the Pleistocene Mediterranean (e.g., Álvarez and Emig 2000, Girone 2002). Changes in thermal conditions of the Mediterranean Sea are also known to drive the reorganization and fragmentation of populations in other benthic fully or moderately boreal metozoans, such as stylasterid hydrozoans (Zibrowius and Cairns 1992), scleractinian anthozoans (Zibrowius and Cairns 1992), polychetes (Arvanitidis et al. 2002), decapod crustaceans (e.g., Almaca 1985), brachiopods (Por and Dimentman 2006, cf. Logan 1979), holothurian echinoderms (Tortonese 1985), and fishes (Tortonese 1985, Rahel and Hubert 1991, Ben Rais Lasram and Mouillot 2009, Ben Rais Lasram et al. 2010). Captions supplementary movies: Suppl. movie 1. Structure and macrozoobenthic inhabitants of the type locality of S. valsalinae sp. nov. at Valsaline Bay, Pula (Croatia), as filmed with a HD underwater camera. The Bay of Valsaline is known to house some interesting but rather bizarre hard substrate biocoenoses the classification of which appears rather problematic, since equivalents are hard or even impossible to find in common textbooks dealing with marine biota of the Mediterranean Sea (e.g. the Cystoseira amentacea – paguroid – Gobius facies: see account of Müller and Schubart 2007). The lack of a broad belt of Posidonia oceanica meadows - a species which normally represents a dominant component of the Mediterranean infralittoral - causes direct converging and mixing of faunal elements of the upper and lower infralittoral macrozoobenthos. Some species identified in the investigated habitat, such as the Pilgrim’s Scallop Pecten jacobeus (Linnaeus, 1758) or the hermit crab Paguristes eremita (Linnaeus, 1767) (see also Fig. 1C), usually indicate muddy or silty bottoms below depth levels of at least 30 metres (e.g. Poppe and Goto 1993, d’Udecem d’Acoz 1999), belonging to the ‘coastal detritic assemblage’ in the sense of Pérès (1985). Other species found in the type locality, such as the lancelet Branchiostoma lanceolatum (Pallas, 1774), are typical inhabitants of coarse sandy sediments in the upper infralittoral (e.g. Ros et al. 1985, Heß 2002). When also regarding Vatova`s (1935) classification of Mediterranean marine zoocoenoses modified for the unique conditions along the coastal waters of the Istrian Peninsula, it can be stated that S. valsalinae sp. nov. belongs to a soft bottom facies combining elements of the ‘Branchiostoma’ (also called ‘Amphioxus sand’) and ‘Venus verrucosa’ type. Suppl. movie 2. Living specimen of Spadella valsalinae sp. nov. collected in the Bay of Pula (Croatia) in July 2009. The movie was shot through one ocular of a stereo light microscope (40 times magnification). At first the head is shown by zooming through the transparent animal from dorsal (sharp vision of the eyes and corona cilata) to ventral surface (sharp vision of oesophagus and mouth cuticle). The hood (praeputium) covers the head, the grasping spine apparatus is compact and located at either side of the head. Afterwards, the camera zooms along the trunk and tail region, the again starting from dorsal surface. Peristaltic movements of the midgut are well visible. Please note that the lateral fins appear somewhat damaged and that the small, thin posterolateral protuberances on the head are missed, what is most probably a sampling artefact. The seminal vesicles are emptied and therefore appear rather inconspicuous, but the clear separation from tail and lateral fins is still visible. Supplementary references Álvarez, F. & Emig, C.C. (2000). Brachiopoda from the Luso-Iberian zone. II. Geographic and bathymetric distribution. Abstract Proceedings of the Millenium Brachiopod Congress (London), 1 p. Almaca, C. (1985). Evolutionary and zoogeographical remarks on the Mediterranean fauna of brachyuran crabs. In Moraitou-Apostolopoulou, M. & Kiortsis, V. (Eds.), Mediterranean marine ecosystems. NATO Conference Series I. Ecology. Vol. 8. (pp. 347-366). New York, London: Plenum Press. Battaglia, M., Murray, M.H., Serpelloni, E. & Burgmann, R. (2004). The adriatic region: an independent microplate within the Africa-Eurasia collision zone. Geophysical Research Letters, 31(9), 9605. Ben Rais Lasram, F. & Mouillot, D. (2009). Increasing southern invasion enhances congruence between endemic and exotic Mediterranean fish fauna. Biological Invasions, 11, 97-711. Ben Rais Lasram, F., Guilhaumon, F., Albouy, C., Somot, S., Thuiller, W. & Mouillot, D. (2010). The Mediterranean Sea as a ‘cul-de-sac’ for endemic fishes facing climate change. Global Change Biology, 16, 3233-3245. Bianchi, C.N. & Morri, C. (2000). Marine biodiversity of the Mediterranean Sea: situation, problems and prospects for future research. Marine Pollution Bulletin, 40, 367-376. Boudouresque, C.F. (2004). Marine biodiversity in the mediterranean status of species, populations and communities. Scientific Reports of Port-Cros national Park, 20, 97-146. Girone, A. (2002). Aspetti Paleobiogeogerafici della fauna de Teleostei (otoliti) del Pleistocene del Mediterraneo. Giornale di Paleontologia. Societa Paleontologica Italiana. Abstract. Heß, M. (2002). 6. Lebensräume und Lebensgemeinschaften. Die Sedimentböden. In Hofrichter, R (Ed.). Das Mittelmeer – Fauna, Flora, Ökologie. Band I: Allgemeiner Teil. (pp. 384-400). Heidelberg: Spektrum Akademischer Verlag. Hsü, K.J. (1974). The Miocene desiccation of the Mediterranean and its climatical and zoogeographical implications. Naturwissenschaften, 61, 137-142. Hsü, K.J., Ryan, W.B.F. & Cita, M.B. (1973). Late Miocene desiccation of the Mediterranean. Nature, 242, 240. Logan, A. (1979). The recent Brachiopoda of the Mediterranean Sea. Bulletin de l’Institut Océanographique de Monaco, 72, 1-112. McKenzie, J.A. (1999). From desert to deluge in the Mediterranean. Nature, 400, 613-614. Müller, C.H.G. & Schubart, C.D. (2007). Insights into the Crustacea of the Adriatic Sea. Observations from four sampling locations along the Croation coast. Rostocker Meeresbiologische Beiträge, 18: 117-135. Pérès, J.M. (1985). History of the Mediterranean biota and the colonization of the depths.Chapter 7. In Margalef, R. (Ed.). Western Mediterranean (Key environments). (pp. 198-232). Oxford: Pergamon Press. Poppe, G.T. & Goto, Y. (1993). European seashells. Volume II (Scaphopoda, Bivalvia, Cephalopoda). Wiesbaden: Verlag Christa Hemmen, 221 p. Por, F.D. & Dimentman, C. (2006). Mare nostrum. Neogene and anthropic natural history of the Mediterranean basin, with emphasis on the Levant. Sofia: Pensoft Publishers, 349 p. Rahel, F.J. & Hubert, W.A.. (1991). Fish assemblages and habitat gradients in a Rocky Mountain – Great Plains stream: biotic zonation and additive patterns of community change. Transactions of the American Fisheries Society, 120, 319332. Ros, J., Romero, J., Ballesteros, E. & Gili, J.M. (1985). Diving in blue water. The benthos. Chapter 8. In: Margalef, R. (Ed.). Western Mediterranean (Key environments). (pp. 233-295). Oxford: Pergamon Press. Tortonese, E. (1985). Distribution and ecology of endemic elements in the Mediterranean fauna (fishes and echinoderms). In Moraitou-Apostolopoulou, M. & Kiortsis, V. (eds.), Mediterranean marine ecosystems. NATO Conference Series I. Ecology. Vol. 8. (pp. 57-83). Ney York, London: Plenum Press. d’Udecem d’Acoz, C. (1999). Inventaire et distribution des crustacés décapodes de l’Atlantique nord-oriental, de la Méditerranée et des eaux continentales adjacentes au nord de 25°N. Bruxelles : Museum National d’Histoire Naturelle (Institut d’Ècologie et de Gestion de la Biodiversité service du Patrimoine Naturel), 383 p. Vatova, A. (1935). Richerche preliminari sulle biocenosi del Golfo di Rovigno. Thalassia, 2: 1-30. Zibrowius, H. & Cairns, S.D. (1992). Revision of the northeast Atlantic and Mediterranean Stylasteridae (Cnidaria: Hydrozoa). Mémoires du Muséum National d’Histoire Naturelle (Paris) Série Zoologie Tome, 153, 1-136.