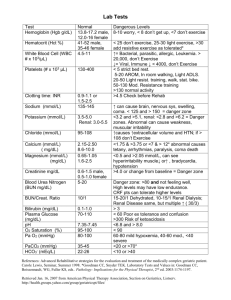
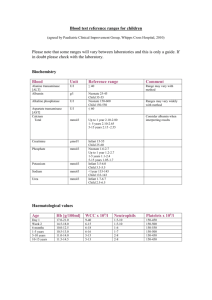
Supplementary Data
advertisement

CVR-2011-1235R2 Supplemental Material and Methods 1. Cell death detection, DNA fragmentation, and cell proliferation assay Cell death was measured by cytoplasmic-associated DNA fragments using Cell Death Detection ELISA (Roche, Mannheim, Germany). Apoptotic DNA-Ladder was detected using Apoptotic DNA Ladder Kit (Roche). Cell growth was evaluated using Cell Proliferation ELISA, BrdU (colorimetric) (Roche). These assays were performed according to the manufacturer’s instructions. 2. Real-time quantitative RT-PCR analysis Transcript levels of target genes were identified by reverse transcription and quantitative PCR using High Capacity RNA-to-cDNA Kit (Applied Biosystems, NY, USA) , TaqMan PCR Master Mix (Applied Biosystems,), and FAM-labeled TaqMan probes (Assays-on-Demand, Applied Biosystems) for human preproET-1 (Applied Biosystems, Hs00174961_m1) , HIF-1α (Applied Biosystems, Hs00153153_m1), low-density lipoprotein glyceraldehyde-3-phosphate receptor (LDL-R) dehydrogenase (Applied (GAPDH) Biosystems, (Applied Hs00181192_m1), Biosystems, and Hs00266705_g1). Expression data were normalized for GAPDH levels. 3. Western blot assay Whole-cell lysates were subjected to Western blot analyses (10% SDS-PAGE or 5–20% gradient SDS-PAGE; 20 μg protein per lane) using anti-HIF-1α (sc-10790, Santa Cruz Biotechnology, CA, USA) at 1 : 5000 dilution, anti-HIF-1β (sc-5580, Santa Cruz Biotechnology) at 1 : 3000 dilution, anti-VHL 1 CVR-2011-1235R2 (sc-17780, Santa Cruz Biotechnology) at 1 : 1000 dilution, anti-HA antibody (Clontech, Mountain View, CA, USA) at 1 : 5000 dilution, and anti-β-actin (Sigma) at 1 : 10000 dilution. Subsequently, the membranes were incubated with a secondary antibody conjugated to horseradish peroxidase (GE Healthcare, Little Chalfont, UK) at 1 : 10000 dilution. The proteins were visualized and quantified using a chemiluminescence method (ECL Plus Western Blotting Detection System, GE Healthcare). 4. Chromatin immunoprecipitation (ChIP) assay VSMC (2 × 106 cells) were cross-linked using 1% formaldehyde for 10 min at 37°C. Cross-linking was stopped by adding glycine (final concentration, 150 mmol/L) to the medium and incubating for 5 min at room temperature. After suspension with ice-cold phosphate buffered saline containing protease inhibitors and centrifugation, the cell pellets were subsequently resuspended in 900 μL lysis buffer (0.1% SDS, 300 mmol/L NaCl, 1 mmol/L EDTA, 0.5 mmol/L EGTA, 0.5% N-lauroylsarcosine, and 10 mmol/L Tris–HCl [pH 8.0]) and incubated on ice for 10 min. The lysates were sonicated using a Bioruptor Ultrasonicator UCD-250 (Cosmo-Bio, Tokyo, Japan) and the chromatin in each lysate was sheared to an average size of 500 bp. After addition of 90 μL of 10% Triton X-100 and centrifugation at 12,000 xg for 20 min, the supernatants were collected and incubated with rotation at 4°C overnight in the presence of Dynabeads M-280 for anti-rabbit IgG (Invitrogen), which were pre-bound with 4 μg anti-HIF-1α antibody or rabbit IgG (Santa Cruz Biotechnology). After incubation, the beads were washed once with low-salt wash buffer (0.1% SDS, 150 mmol/L NaCl, 1% Triton X-100, 2 mmol/L EDTA, and 20 mmol/L Tris, [pH 8.0]), twice with high-salt wash buffer (0.1% SDS, 400 mmol/L NaCl, 1% Triton X-100, 2 mmol/L 2 CVR-2011-1235R2 EDTA, and 20 mmol/L Tris [pH 8.0]), five times with LiCl wash buffer (0.5 M LiCl, 1% Nonidet P-40, 0.7% deoxycholate, 1 mmol/L EDTA, and 50 mmol/L HEPES [pH 7.6]), and finally once with Tris–EDTA buffer. After centrifugation, immune complexes were resuspended in 200 μL elution buffer (1% SDS, 50 mmol/L Tris–HCl [pH 8.0], and 10 mmol/L EDTA) and incubated for 15 min at 65°C. Cross-linking of immune complexes was reversed in 200 mmol/L NaCl at 65°C overnight, after which complexes were digested with proteinase K at 45°C for 1 h. Genomic DNA fragments were recovered by phenol–chloroform extraction, ethanol precipitation, and resuspension in sterile H2O. Human genomic sequences containing HRE from −204 bp to +173 bp relative to the transcription start site of the ET-1 gene were amplified using the following primers: ET-1 forward, 5′-CTGCCCCGAATTGTCAGACG-3′; ET-1 reverse, 5′-CAAAGCGATCCTTCAGCCC-3′. Supplemental Discussion Previous studies reported that statin affects the function of the mammalian target of rapamycin1, an important factor in HIF-1α synthesis2. However, there was no significant effect of fluvastatin on HIF-1α protein level in the presence of MG-132 or CoCl2, as shown in Figures 4A and 5. Therefore, the main inhibitory effect of statin on HIF-1α protein level is the acceleration of protein degradation. The relationships between HIF-1 and other transcription factors, including NF-κB-dependent HIF-1α transcription and the interaction between HIF-1 and AP-1, have been investigated3,4. Several studies showed the inhibitory effect of statin on these transcription factors5,6. The effect of statin on these transcription factors may interfere with or enhance HIF-1-dependent ET-1 regulation. However, the 3 CVR-2011-1235R2 pattern of inhibition of HIF-1 protein by fluvastatin was similar to that of hypoxia-dependent ET-1 expression and there was no significant effect on HIF-1α mRNA level in our study. Therefore, the crosstalk between HIF-1 and these transcriptional factors may not have a great influence. Oxidative stress reportedly affects HIF-1 function: Sirtuin 3 attenuates HIF-1α accumulation in hypoxia by suppression of mitochondrial reactive oxygen species in carcinoma cell lines7. Previous investigations reported that statins inhibited cellular oxidative stress, including inhibition of nicotinamide adenine dinucleotide phosphate hydrogen (NADPH) oxidase 1 expression and changes in superoxide dismutase (SOD) levels as pleiotropic effects8,9, and it is possible that this involves the attenuation of hypoxia-dependent ET-1 induction. On the other hand, fluvastatin has the unique structure of a fluorophenyl-substituted indole ring that exerts an additional antioxidant effect10, but other statins, such as pravastatin and simvastatin, do not have this structure. Therefore, there may be a difference between fluvastatin and other statins in the effect on cellular oxidative stress. Nevertheless, the inhibitory effect on HIF-1-dependent ET-1 expression was similar among the three types of statin (fluvastatin, pravastatin, and simvastatin), as shown in Figure 2B, Supplemental Figure 3A, and 3C. Based on these findings, the influence of statin on cellular oxidative stress may not involve hypoxia-induced ET-1 expression. Previous studies in vascular endothelial cells have revealed that statins inhibit preproET-1 mRNA expression and preproET-1 promoter activity using 0.65 kb, 1.5 kb, and 5.2 kb of the preproET-1 5’ flanking region under normoxic conditions11. These studies also stated that high concentrations of statins are required for these inhibitory effects: ≥2 μM of simvastatin or ≥5 μM of atorvastatin with more than 12 h of incubation in bovine aortic endothelial cells12, and ≥1 μM of fluvastatin with more than 6 h 4 CVR-2011-1235R2 of incubation in human umbilical vein endothelial cells 13 . Our findings in vascular smooth muscle cells under normoxic conditions almost agree with these previous findings, although there are minor differences in time course and statin concentrations. Treatment with lower concentration (100 nM) of fluvastatin for >8 h affected preproET-1 mRNA expression, as shown in Figure 2C. Moreover, the previous studies did not analyze hypoxia-induced preproET-1 gene expression. Our study is the first report, as per our knowledge, of the inhibitory effects of fluvastatin on preproET-1 expression under both normoxic and hypoxic conditions. Supplementary References 1. Wagner RJ, Martin KA, Powell RJ, Rzucidlo EM. Lovastatin induces VSMC differentiation through inhibition of Rheb and mTOR. Am J Physiol Cell Physiol 2010;299:C119-27. 2. Nakamura H, Makino Y, Okamoto K, Poellinger L, Ohnuma K, Morimoto C, et al. TCR engagement increases hypoxia-inducible factor-1 alpha protein synthesis via rapamycin-sensitive pathway under hypoxic conditions in human peripheral T cells. J Immunol 2005;174:7592-9. 3. Diebold I, Djordjevic T, Hess J, Gorlach A. Rac-1 promotes pulmonary artery smooth muscle cell proliferation by upregulation of plasminogen activator inhibitor-1: role of NFkappaB-dependent hypoxia-inducible factor-1alpha transcription. Thromb Haemost 2008;100:1021-8. 4. Laderoute KR. The interaction between HIF-1 and AP-1 transcription factors in response to low oxygen. Semin Cell Dev Biol 2005;16:502-13. 5. Hattori Y, Matsumura M, Kasai K. Vascular smooth muscle cell activation by C-reactive protein. 5 CVR-2011-1235R2 Cardiovasc Res 2003;58:186-95. 6. Viedt C, Shen W, Fei J, Kamimura M, Hansch GM, Katus HA, et al. HMG-CoA reductase inhibition reduces the proinflammatory activation of human vascular smooth muscle cells by the terminal complement factor C5b-9. Basic Res Cardiol 2003;98:353-61. 7. Bell EL, Emerling BM, Ricoult SJ, Guarente L. SirT3 suppresses hypoxia inducible factor 1alpha and tumor growth by inhibiting mitochondrial ROS production. Oncogene 2011;30:2986-96. 8. Briones AM, Rodriguez-Criado N, Hernanz R, Garcia-Redondo AB, Rodrigues-Diez RR, Alonso MJ, et al. Atorvastatin prevents angiotensin II-induced vascular remodeling and oxidative stress. Hypertension 2009;54:142-9. 9. Verreth W, De Keyzer D, Davey PC, Geeraert B, Mertens A, Herregods MC, et al. Rosuvastatin restores superoxide dismutase expression and inhibits accumulation of oxidized LDL in the aortic arch of obese dyslipidemic mice. Br J Pharmacol 2007;151:347-55. 10. Blum CB. Comparison of properties of four inhibitors of 3-hydroxy-3-methylglutaryl-coenzyme A reductase. Am J Cardiol 1994;73:3D-11D. 11. Hernandez-Perera O, Perez-Sala D, Soria E, Lamas S. Involvement of Rho GTPases in the transcriptional inhibition of preproendothelin-1 gene expression by simvastatin in vascular endothelial cells. Circ Res 2000;87:616-22. 12. Hernandez-Perera O, Perez-Sala D, Navarro-Antolin J, Sanchez-Pascuala R, Hernandez G, Diaz C, et al. Effects of the 3-hydroxy-3-methylglutaryl-CoA reductase inhibitors, atorvastatin and simvastatin, on the expression of endothelin-1 and endothelial nitric oxide synthase in vascular endothelial cells. J 6 CVR-2011-1235R2 Clin Invest 1998;101:2711-9. 13. Ozaki K, Yamamoto T, Ishibashi T, Matsubara T, Nishio M, Aizawa Y. Regulation of endothelial nitric oxide synthase and endothelin-1 expression by fluvastatin in human vascular endothelial cells. Jpn J Pharmacol 2001;85:147-54. 7