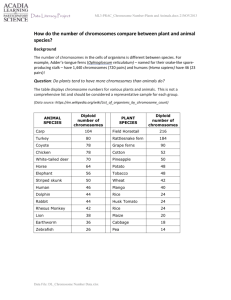
An Introduction to Genetic Analysis Chapter 18 Chromosome
advertisement

An Introduction to Genetic Analysis Chapter 18 Chromosome Mutation II: Changes in Chromosome Number Chapter 18 Chromosome Mutation II: Changes in Chromosome Number Key Concepts Organisms with multiple chromosome sets (polyploids) are generally larger than diploid organisms, but meiotic pairing anomalies make some polyploid organisms sterile. An even number of polyploid sets is generally more likely to result in fertility. Then the single-locus segregation ratios are different from those of diploids. Crosses between two different species followed by the doubling of the chromosome number in the hybrid produces a special kind of fertile interspecific polyploid. Variants in which a single chromosome has been gained or lost generally arise by nondisjunction (abnormal chromosome segregation at meiosis or mitosis). Such variants tend to be sterile and show the abnormalities attributable to gene imbalance. When fertile, such variants show abnormal gene segregation ratios for the misrepresented chromosome only. Introduction The second major type of chromosome mutation is change in chromosome number. Few aspects of genetics impinge on human affairs quite so directly as this one. Chromosome numbers change spontaneously as accidents within cells, and this process has been going on for as long as life has existed on the planet. In fact, changes in chromosome number have been instrumental in molding genomes during evolution. For examples of such changes, we have to look no farther than the food on our dining tables because many of the plants (and some of the animals) that we eat arose through spontaneous changes in chromosome number in the course of the evolution of those species. Today, breeders emulate this process by manipulating chromosome number to improve productivity or some other useful feature of the organism. But perhaps the main relevance of chromosome numbers is for members of our own species in that a large proportion of genetically determined ill health in humans is caused by abnormal chromosome numbers. In this chapter, we shall investigate the processes that produce new chromosome numbers, the diagnostic tests for detecting such changes, and the properties of cells and individual organisms carrying the different kinds of variant chromosomal complements. As with any area of cytogenetics, the techniques are a combination of genetics and microscopy. Changes in chromosome number are usually classified into two types—changes in whole chromosome 1 勇者并非无所畏惧,而是能判断出有比恐惧更重要的东西. An Introduction to Genetic Analysis Chapter 18 Chromosome Mutation II: Changes in Chromosome Number sets and changes in parts of chromosome sets—and these two types are dealt with in the next two sections. Aberrant euploidy The number of chromosomes in a basic set is called the monoploid number (x). Organisms with multiples of the monoploid number of chromosomes are called euploid. We learned in earlier chapters that eukaryotes normally carry either one chromosome set (haploids) or two sets (diploids). Haploids and diploids, then, are both cases of normal euploidy. Euploid types that have more than two sets of chromosomes are called polyploid. The polyploid types are named triploid (3x), tetraploid (4x), pentaploid (5x), hexaploid (6x), and so forth. Polyploids arise naturally as spontaneous chromosomal mutations and, as such, they must be considered aberrations because they differ from the previous norm. However, many species of plants and animals have clearly arisen through polyploidy, so evidently evolution can take advantage of polyploidy when it arises. It is worth noting that organisms with one chromosome set sometimes arise as variants of diploids; such variants are called monoploid (1x). In some species, monoploid stages are part of the regular life cycle, but other monoploids are spontaneous aberrations. The haploid number (n), which we have already used extensively, refers strictly to the number of chromosomes in gametes. In most animals and many plants with which we are familiar, the haploid number and monoploid number are the same. Hence, n or x (or 2n or 2x) can be used interchangeably. However, in certain plants, such as modern wheat, n and x are different. Wheat has 42 chromosomes, but careful study reveals that it is hexaploid, with six rather similar but not identical sets of seven chromosomes. Hence, 6x = 42 and x = 7. However, the gametes of wheat contain 21 chromosomes, so n = 21 and 2n = 42. Monoploids Male bees, wasps, and ants are monoploid. In the normal life cycles of these insects, males develop parthenogenetically—that is, they develop from unfertilized eggs. However, in most species, monoploid individuals are abnormal, arising in natural populations as rare aberrations. The germ cells of a monoploid cannot proceed through meiosis normally, because the chromosomes have no pairing partners. Thus, monoploids are characteristically sterile. (Male bees, wasps, and ants bypass meiosis in forming gametes; here, mitosis produces the gametes.) If a monoploid cell does undergo meiosis, the single chromosomes segregate randomly, and the probability of all chromosomes going to one pole is (1/2)x−1 where x is the number of chromosomes. This formula estimates the frequency of viable (whole-set) gametes, which is a small number if x is large. Monoploids play an important role in modern approaches to plant breeding. Diploidy is an inherent nuisance when breeders want to induce and select new gene mutations that are favorable and to find new combinations of favorable alleles at different loci. New recessive mutations must be made homozygous before they can be expressed, and favorable allelic 2 勇者并非无所畏惧,而是能判断出有比恐惧更重要的东西. An Introduction to Genetic Analysis Chapter 18 Chromosome Mutation II: Changes in Chromosome Number combinations in heterozygotes are broken up by meiosis. Monoploids provide a way around some of these problems. In some plant species, monoploids can be artificially derived from the products of meiosis in a plant's anthers. A cell destined to become a pollen grain can instead be induced by cold treatment to grow into an embryoid, a small dividing mass of cells. The embryoid can be grown on agar to form a monoploid plantlet, which can then be potted in soil and allowed to mature (Figure 18-1). Plant monoploids can be exploited in several ways. In one, they are first examined for favorable traits or allelic combinations, which may arise from heterozygosity already present in the parent or induced in the parent by mutagens. The monoploid can then be subjected to chromosome doubling to achieve a completely homozygous diploid with a normal meiosis, capable of providing seed. How is this achieved? Quite simply, by the application of a compound called colchicine to meristematic tissue. Colchicine—an alkaloid drug extracted from the autumn crocus—inhibits the formation of the mitotic spindle, so cells with two chromosome sets are produced (Figure 18-2). These cells may proliferate to form a sector of diploid tissue that can be identified cytologically. Another way in which the monoploid may be used is to treat its cells basically like a population of haploid organisms in a mutagenesis-and-selection procedure. A population of cells is isolated, their walls are removed by enzymatic treatment, and they are treated with mutagen. They are then plated on a medium that selects for some desirable phenotype. This approach has been used to select for resistance to toxic compounds produced by one of the plant's parasites and to select for resistance to herbicides being used by farmers to kill weeds. Resistant plantlets eventually grow into haploid plants, which can then be doubled (with the use of colchicine) into a pure-breeding, diploid, resistant type (Figure 18-3). These powerful techniques can circumvent the normally slow process of meiosis-based plant breeding. The techniques have been successfully applied to several important crop plants, such as soybeans and tobacco. The anther technique for producing monoploids does not work in all organisms or in all genotypes of an organism. Another useful technique has been developed in barley, an important crop plant. Diploid barley, Hordeum vulgare, can be fertilized by pollen from a diploid wild relative called Hordeum bulbosum. This fertilization results in zygotes with one chromosome set from each parental species. In the ensuing somatic cell divisions, however, the chromosomes of H. bulbosum are eliminated from the zygote, whereas all the chromosomes of H. vulgare are retained, resulting in a haploid embryo. (The haploidization appears to be caused by a genetic incompatibility between the chromosomes of the different species.) The chromosomes of the resulting haploids can be doubled with colchicine. This approach has led to the rapid production and widespread planting of several new barley varieties, and it is being used successfully in other species too. MESSAGE 3 勇者并非无所畏惧,而是能判断出有比恐惧更重要的东西. An Introduction to Genetic Analysis Chapter 18 Chromosome Mutation II: Changes in Chromosome Number To create new plant lines, geneticists produce monoploids with favorable genotypes and then double the chromosomes to form fertile, homozygous diploids. Polyploids In the realm of polyploids, we must distinguish between autopolyploids, which are composed of multiple sets from within one species, and allopolyploids, which are composed of sets from different species. Allopolyploids form only between closely related species; however, the different chromosome sets are homeologous (only partly homologous)—not fully homologous, as they are in autopolyploids. Triploids Triploids are usually autopolyploids. They arise spontaneously in nature or are constructed by geneticists from the cross of a 4x (tetraploid) and a 2x (diploid). The 2x and the x gametes unite to form a 3x triploid. Triploids are characteristically sterile. The problem, like that of monoploids, lies in pairing at meiosis. Synapsis, or true pairing, can take place only between two chromosomes, but one chromosome can pair with one partner along part of its length and with another along the remainder, which gives rise to an association of three chromosomes. Paired chromosomes of the type found in diploids are called bivalents. Associations of three chromosomes are called trivalents, and unpaired chromosomes are called univalents. Hence in triploids there are two pairing possibilities, resulting either in a trivalent or in a bivalent plus a univalent. Paired centromeres segregate to opposite poles, but unpaired centromeres pass to either pole randomly. We see in Figure 18-4 that the net result of both the pairing possibilities is an uneven segregation, with two chromosomes going in one direction and one in the other. This happens for every chromosome threesome. If all the single chromosomes pass to the same pole and simultaneously the other two chromosomes pass to the opposite pole, then the gametes formed will be haploid and diploid. The probability of this type of meiosis will be (1/2)x−1, and this proportion is likely to be low. All other possibilities will give gametes with chromosome numbers intermediate between the haploid and diploid number; such genomes are aneuploid—“not euploid.” It is likely that these aneuploid gametes will not lead to viable progeny; in fact, this category is responsible for the almost complete lack of fertility of triploids. The problem is one of genome imbalance, a phenomenon that we shall encounter repeatedly in this chapter. For most organisms, the euploid chromosome set is a finely tuned set of genes in relative proportions that seem to be functionally significant. Multiples of this set are tolerated because there is no change in the relative proportions of genes. However, the addition of one or more extra chromosomes is nearly always deleterious because the proportions of genes in those extra chromosomes are altered. Although the action of some genes can be regulated to compensate for extra gene “dosage,” the overall effect of the extra genetic material seems too great to be overcome by gene regulation. The deleterious effect can be expressed at the level of gametes, making them nonfunctional, or at the level of the zygote, resulting in lethality, sterility, or lowered fitness. 4 勇者并非无所畏惧,而是能判断出有比恐惧更重要的东西. An Introduction to Genetic Analysis Chapter 18 Chromosome Mutation II: Changes in Chromosome Number In triploids, it is possible that some haploid or diploid gametes will form, and some may unite to form a euploid zygote, but the likelihood of this possibility is inherently low. Consider bananas. The bananas that are widely available commercially are triploids with 11 chromosomes in each set (3x = 33). The probability of a meiosis in which all univalents pass to the same pole is (1/2)x−1, or (1/2)10 = 1/1024, so bananas are effectively sterile. The most obvious expression of the sterility of bananas is that there are no seeds in the fruit that we eat. Another example of the commercial exploitation of triploidy in plants is the production of triploid watermelons. For the same reasons that bananas are seedless, triploid watermelons are seedless, a phenotype favored by some for its convenience. MESSAGE Some types of chromosome mutations are themselves aneuploid; other types produce aneuploid gametes or zygotes. Aneuploidy is nearly always deleterious because of genetic imbalance—the ratio of genes is different from that in euploids and interferes with the normal operation of the genome. Autotetraploids Autotetraploids arise naturally by the spontaneous accidental doubling of a 2x genome to a 4x genome, and autotetraploidy can be induced artificially through the use of colchicine. Autotetraploid plants are advantageous as commercial crops because, in plants, the larger number of chromosome sets often leads to increased size. Cell size, fruit size, flower size, stomata size, and so forth, can be larger in the polyploid (Figure 18-5). Here we see another effect that must be explained by gene numbers. Presumably the amount of gene product (protein or RNA) is proportional to the number of genes in the cell, and this number is higher in the cells of polyploids compared with diploids. MESSAGE Polyploid plants are often larger and have larger organs than their diploid relatives. Because 4 is an even number, autotetraploids can have a regular meiosis, although this is by no means always the case. The crucial factor is how the four homologous chromosomes, one from each of the four sets, pair and segregate. There are several possibilities, as shown in Figure 18-6. Pairings between four chromosomes are called quadrivalents. In tetraploids, the two-bivalent and the quadrivalent pairing modes tend to be most regular in segregation, but even here there is no guarantee of a 2:2 segregation. If all chromosome sets segregate 2:2 as they do in some species, then the gametes will be functional and a formal genetic analysis can be developed for such autotetraploids. Let's consider the genetics of a fertile tetraploid. We can consider an experiment in which colchicine is used to double the chromosomes of an A/a plant to form an A/A/a/a auto-tetraploid, which we will assume shows 2:2 segregation. We now have a further concern because polyploids such as tetraploids give different phenotypic ratios in their progeny, depending on whether the locus in question is tightly linked to the centromere. First, we 5 勇者并非无所畏惧,而是能判断出有比恐惧更重要的东西. An Introduction to Genetic Analysis Chapter 18 Chromosome Mutation II: Changes in Chromosome Number consider a centromere-linked gene. The three possible pairing and segregation patterns are presented in Figure 18-7; these patterns occur by chance and with equal frequency. As Figure 18-7 shows, the 2x gametes produced are A/a, A/A, or a/a, in a ratio of 8:2:2, or 4:1:1. If such a plant is selfed, the probability of an a/a/a/a phenotype in the offspring is 1/6 × 1/6 =1/36. In other words, a 35:1 phenotypic ratio of A/–/–/–:a/a/a/a will be observed if A is fully dominant over three a alleles. If, in the same kind of plant, a genetic locus having the alleles B and b is very far removed from the centromere, crossing-over must be considered. This example forces us to think in terms of chromatids instead of chromosomes; there are four B chromatids and four b chromatids (Figure 18-8). Because the number of crossovers in such a long region will be large, the genes will become effectively unlinked from their original centromeres. The packaging of genes two at a time into gametes is very much like grabbing two balls at random from a bag of eight balls: four of one kind and four of another. The probability of picking two b genes is then So, in a selfing, the probability of a b/b/b/b phenotype is 2/4 × 3/14 = 9/196, which is approximately 1/22. Hence, there will be a 21:1 phenotypic ratio of B/–/–/–:b/b/b/b. For genetic loci of intermediate position, intermediate ratios will result. Allopolyploids The “classic” allopolyploid was synthesized by G. Karpechenko in 1928. He wanted to make a fertile hybrid that would have the leaves of the cabbage (Brassica) and the roots of the radish (Raphanus). Each of these species has 18 chromosomes, and they are related closely enough to allow intercrossing. A viable hybrid progeny individual was produced from seed. However, this hybrid was functionally sterile because the nine chromosomes from the cabbage parent were different enough from the radish chromosomes that pairs did not synapse and disjoin normally: However, one day a few seeds were in fact produced by this (almost) sterile hybrid. On planting, these seeds produced fertile individuals with 36 chromosomes. All these individuals were allopolyploids. They had apparently been derived from spontaneous, accidental chromosome doubling to 2n1 + 2n2 in the sterile hybrid, presumably in tissue that 6 勇者并非无所畏惧,而是能判断出有比恐惧更重要的东西. An Introduction to Genetic Analysis Chapter 18 Chromosome Mutation II: Changes in Chromosome Number eventually became germinal and underwent meiosis. Thus, in 2n1 + 2n2 tissue, there is a pairing partner for each chromosome and balanced gametes of the type n1 + n2 are produced. These gametes fuse to give 2n1 + 2n2 allopolyploid progeny, which also are fertile. This kind of allopolyploid is sometimes called an amphidiploid, which means “doubled diploid” (Figure 18-9). (Unfortunately for Karpechenko, his amphidiploid had the roots of a cabbage and the leaves of a radish.) When the allopolyploid was crossed with either parental species, sterile offspring resulted. The offspring of the cross with radish were 2n1 + n2 constituted from an n1 + n2 gamete from the allopolyploid and an n1 gamete from the radish. The n2 chromosomes had no pairing partners, so sterility resulted. Consequently, Karpechenko had effectively created a new species, with no possibility of gene exchange with its parents. He called his new species Raphanobrassica. Today, allopolyploids are routinely synthesized in plant breeding. Instead of waiting for spontaneous doubling to occur in the sterile hybrid, the plant breeder adds colchicine to induce doubling. The goal of the breeder is to combine some of the useful features of both parental species into one type. This kind of endeavor is very unpredictable, as Karpechenko learned. In fact, only one synthetic amphidiploid has ever been widely used. This amphidiploid is Triticale, an amphidiploid between wheat (Triticum, 2n = 6x = 42 and rye (Secale, 2n = 2x = 14 Triticale combines the high yields of wheat with the ruggedness of rye. Figure 18-10 shows the procedure for synthesizing Triticale. In nature, allopolyploidy seems to have been a major force in speciation of plants. There are many different examples. One particularly satisfying one is shown by the genus Brassica, as illustrated in Figure 18-11. Here three different parent species have hybridized in all possible pair combinations to form new amphidiploid species. A particularly interesting natural allopolyploid is bread wheat, Triticum aestivum(2n = 6x = 42). By studying various wild relatives, geneticists have reconstructed a probable evolutionary history of bread wheat (Figure 18-12). In a bread wheat meiosis, there are always 21 pairs of chromosomes. Furthermore, it has been possible to establish that any given chromosome has only one specific pairing partner (homologous pairing)—not five other potential partners (homeologous pairing). The suppression of such homeologous pairing (which would make the species more unstable) is maintained by an allele, Ph, on the long arm of chromosome 5 of the B set. Thus, Ph ensures a diploidlike meiotic behavior for this hexaploid species. Without Ph, bread wheat could probably never have arisen. It is interesting to speculate about whether Western civilization could have begun or progressed without this species—in other words, without the Ph mutation. Somatic allopolyploids from cell hybridization Another innovative approach to plant breeding is to try to make allopolyploid-like hybrids by fusing asexual cells. Theoretically, such a technique would allow us to combine widely 7 勇者并非无所畏惧,而是能判断出有比恐惧更重要的东西. An Introduction to Genetic Analysis Chapter 18 Chromosome Mutation II: Changes in Chromosome Number differing parental species. The technique does indeed work, but the only allopolyploids that have been produced so far can also be made by the sexual methods that we have considered already. In the cell-fusion procedure, cell suspensions of the two parental species are prepared and stripped of their cell walls by special enzyme treatments. The stripped cells are called protoplasts. The two protoplast suspensions are then combined with polyethylene glycol, which enhances protoplast fusion. The parental cells and the fused cells proliferate on an agar medium to form colonies (in much the same way as microbes). If these colonies, or calluses as they are called, are examined, a fair percentage of them are found to be allopolyploid-like hybrids with chromosome numbers equal to the sum of the parental numbers. Thus, not only do the protoplast cell membranes fuse to form a kind of heterokaryon, but the nuclei fuse, too, to give rise to a 2n1 + 2n2 amphidiploid. Another good example of an allopolyploid-like hybrid is commercial tobacco, Nicotiana tabacum, which has 48 chromosomes. This species of tobacco was originally found in nature as a spontaneous amphidiploid. The two probable parents are N. sylvestris and N. tomentosiformis, each of which has 24 chromosomes. A sexual cross between N. tabacum and either of these two probable parents give a 36-chromosome hybrid containing 12 chromosome pairs plus 12 unpaired chromosomes. A cross between N. sylvestris and N. tomentosiformis yields a 24-chromosome hybrid in which there is no pairing at all. Hence, it appears that part of the N. tabacum genome is from N. sylvestris and part is from N. tomentosiformis. This amphidiploid can be recreated either sexually, by using colchicine as described previously, or somatically by cell fusion. When cells of the prospective parental species are fused, a 48-chromosome hybrid cell line is produced, from which plants identical in behavior to N. tabacum may be grown. (Note that, in the latter method, colchicine is not required, because the fusion product is already amphidiploid.) The recovery of somatic hybrids may be enhanced if a selective system is available. In one example in N. tabacum, two monoploid lines were fused to form a diploid hybrid culture by using complementation as the selection system. The first line had whitish, light-sensitive leaves due to a recessive mutation w. The other had yellowish, light-sensitive leaves due to a mutation y at a separate locus. When the cells were combined in a petri dish, the diploid w+/w · y+/y calluses could be selected by their resistance to light and their normal green color. The calluses can be grown into plantlets, which then are either grafted onto a mature plant to develop or potted themselves. The protocol for this experiment is illustrated in Figure 18-13. Two allotetraploids of Petunia, one produced by sexual hybridization and the other by somatic hybridization, are compared in Figure 18-14. Note that the two are identical in appearance and produce the same range of progeny types. MESSAGE Allopolyploids can be synthesized either by crossing related species and doubling the chromosomes of the hybrid or by asexually fusing the cells of different species. Polyploidy in animals 8 勇者并非无所畏惧,而是能判断出有比恐惧更重要的东西. An Introduction to Genetic Analysis Chapter 18 Chromosome Mutation II: Changes in Chromosome Number You may have noticed that most of the discussion of polyploidy so far has concerned plants. Indeed, polyploidy is more common in plants than in animals; nevertheless there are many cases of polyploid animals. Examples are found in flatworms, leeches, and brine shrimp. In these cases, reproduction is by parthenogenesis, the development of a special type of unfertilized egg into an embryo, without the need for fertilization. However, examples are not confined to these so-called lower forms. Polyploid amphibians and reptiles are surprisingly common. They show several modes of reproduction. The males and females of polyploid frogs and toads participate in their sexual cycles, whereas polyploid salamanders and lizards are parthenogenetic. Some fish also are polyploid; in two cases, it appears that a single polyploid event gave rise to an entire taxonomic family in evolution. This situation contrasts with that of amphibians and reptiles because in those cases the polyploids all have closely related diploid species, and, hence, the polyploid events do not seem to have been important in the evolution of the group as a whole. The Salmonidae family of fishes (containing salmon and trout) is a familiar example of a group that appears to have originated through polyploidy. Salmonids have twice as much DNA as have related fish. Different salmonid species have different chromosome numbers, but the group has an almost invariant number of chromosome arms (some fused in some species) that is twice the number of arms in related groups. Hence, the evidence points to the salmonids' having evolved from a single event that gave rise to a tetraploid. You might also be interested to know that the sterility of triploids has been commercially exploited in animals as well as plants. Triploid oysters have been developed, and they have a commercial advantage over their diploid relatives. The diploids go through a spawning season, during which they are unpalatable. Triploids, however, because of their sterility, do not spawn and are palatable the whole year round. Human polyploid zygotes do arise through various kinds of mistakes in cell division. Most die in utero. Occasionally, triploid babies are born, but none survive. Aneuploidy Aneuploidy is the second major category of chromosome mutations in which chromosome number is abnormal. An aneuploid is an individual organism whose chromosome number differs from the wild type by part of a chromosome set. Generally, the aneuploid chromosome set differs from wild type by only one or a small number of chromosomes. Aneuploids can have a chromosome number either greater or smaller than that of the wild type. Aneuploid nomenclature is based on the number of copies of the specific chromosome in the aneuploid state. For example, the aneuploid condition 2n − 1 is called monosomic (meaning “one chromosome”) because only one copy of some specific chromosome is present instead of the usual two found in its diploid progenitor. The aneuploid 2n + 1 is called trisomic,2n − 2 is nullisomic, and n + 1 is disomic. Nullisomics (2n − 2) 9 勇者并非无所畏惧,而是能判断出有比恐惧更重要的东西. An Introduction to Genetic Analysis Chapter 18 Chromosome Mutation II: Changes in Chromosome Number Although nullisomy is a lethal condition in diploids, an organism such as bread wheat, which behaves meiotically like a diploid although it is a hexaploid, can tolerate nullisomy. The four homoeologous chromosomes apparently compensate for a missing pair of homologs. In fact, all the possible 21 bread wheat nullisomics have been produced; they are illustrated in Figure 18-15. Their appearances differ from the normal hexaploids; furthermore, most of the nullisomics grow less vigorously. Monosomics (2n − 1) Monosomic chromosome complements are generally deleterious for two main reasons. First, the missing chromosome perturbs the overall gene balance in the chromosome set. (We encountered this effect earlier). Second, having a chromosome missing allows any deleterious recessive allele on the single chromosome to be hemizygous and thus to be directly expressed phenotypically. Notice that these are the same effects produced by deletions. Nondisjunction in mitosis or meiosis is the cause of most aneuploids. Disjunction is the normal separation of homologous chromosomes or chromatids to opposite poles at nuclear division. Nondisjunction is a failure of this disjoining process, and two chromosomes (or chromatids) go to one pole and none to the other. Nondisjunction occurs spontaneously; it is another example of a chance failure of a basic cellular process. In meiotic nondisjunction, the chromosomes may fail to disjoin at either the first or second division (Figure 18-16). Either way, n + 1 and n − 1 gametes are produced. If an n − 1 gamete is fertilized by an n gamete, a monosomic(2 n − 1) zygote is produced. The fusion of an n + 1 and an n gamete yields a trisomic 2n + 1 The precise molecular processes that fail in nondisjunction are not known, but, in experimental systems, the frequency of nondisjunction can be increased by interference with microtubule action. It appears that disjunction is more likely to go awry in meiosis I. This likelihood may not be surprising, because normal anaphase I disjunction requires that proper homologous associations be maintained during prophase I and metaphase I. In contrast, proper disjunction at anaphase II or at mitosis requires that the centromere splits properly but does not require nearly as elaborate a process during prophase and metaphase. Meiosis I nondisjunction can be viewed as the failure to form or maintain a tetrad (a group of four chromatids) until anaphase I. Crossovers are implicit in this process normally. In most organisms, the amount of crossing-over is sufficient to ensure that all tetrads will have at least one exchange per meiosis. In Drosophila, many of the nondisjunctional chromosomes in newly arising disomic gametes are nonrecombinant, with one nondisjunctional homolog carrying the markers of one input chromosome and the other homolog carry- ing the markers of the other chromosome. Similar observations have been made in human trisomies. In addition, in several different experimental organisms, mutations that interfere with recombination have the effect of massively increasing the frequency of meiosis I nondisjunction. This effect points to an important role of crossing-over in maintaining 10 勇者并非无所畏惧,而是能判断出有比恐惧更重要的东西. An Introduction to Genetic Analysis Chapter 18 Chromosome Mutation II: Changes in Chromosome Number chromosomal associations in the tetrad; in the absence of these associations, chromosomes are vulnerable to anaphase I nondisjunction. In humans, a sex-chromosome monosomic complement of 44 autosomes + 1 X produces a phenotype known as Turner syndrome. Affected people have a characteristic, easily recognizable phenotype: they are sterile females, are short in stature, and often have a web of skin extending between the neck and shoulders (Figure 18-17). Although their intelligence is near normal, some of their specific cognitive functions are defective. About 1 in 5000 female births have this monosomic chromosomal complement. Monosomics for all human autosomes die in utero. Geneticists have used viable plant nullisomics and monosomics to identify the chromosomes that carry the loci of newly found recessive mutant alleles. For example, a geneticist may obtain different monosomic lines, each of which lacks a different chromosome. Homozygotes for the new mutant allele are crossed with each monosomic line, and the progeny of each cross are inspected for expression of the recessive phenotype. The phenotype appears in some of the progeny of the parent that is monosomic for the locus-bearing chromosome and thus identifies it. Figure 18-18 shows that this test works because meiosis in the monosomic parent produces some gametes that lack the chromosome bearing the locus. When one of these gametes forms a zygote, the single chromosome contributed by the other homozygous recessive parent determines the phenotype. Genetic analysis of humans occasionally reveals a similar unmasking of a recessive phenotype by an n − 1gamete. For example, two people whose vision is normal may produce a daughter who has Turner syndrome and who is also red-green colorblind. This coincidence is interpreted as follows. First, because the father is not colorblind, the mother must be heterozygous for the recessive allele and must have passed this allele on to the Turner daughter. Nondisjunction must have occurred in the father, resulting in an n − 1 sperm, which combined with an egg bearing the colorblind allele from the mother. MESSAGE Monosomics show the deleterious effects of genome inbalance, as well as unexpected expression of recessive alleles carried on the monosomic chromosome. Trisomics (2n + 1) The trisomic condition also is one of chromosomal imbalance and can result in abnormality or death. However, there are many examples of viable trisomics. You might remember that we studied the trisomics of the Jimson weed Datura stramonium in Chapter 3 (see Figure 3-7). Furthermore, trisomics can be fertile. When cells from some trisomic organisms are observed under the microscope at the time of meiotic chromosome pairing, the trisomic chromosomes are seen to form a trivalent, an associated group of three, whereas the other chromosomes form regular bivalents. What genetic ratios might we expect for genes on the trisomic chromosome? Let us consider a gene, A, that is close to the centromere on that chromosome, and let us 11 勇者并非无所畏惧,而是能判断出有比恐惧更重要的东西. An Introduction to Genetic Analysis Chapter 18 Chromosome Mutation II: Changes in Chromosome Number assume that the genotype is A/a/a. Furthermore, if we postulate that two of the centromeres disjoin to opposite poles as in a normal bivalent and that the other chromosome passes randomly to either pole, then we can predict the three equally frequent segregations shown in Figure 18-19. These segregations result in an overall gametic ratio of 1A:2A/a:2a:1a/a. This ratio and the one corresponding to a trisomic of genotype A/A/a are observed in practice. If a trisomic tester set is available (much like the nullisomic tester set described earlier), then a new mutation can be located to a chromosome by determining which of the testers gives the special ratio. There are several examples of viable trisomics in humans. The combination XXY (1 in 1000 male births) results in Klinefelter syndrome, males with lanky builds who are mentally retarded and sterile (Figure 18-20). Another combination, XYY, also occurs in about 1 in 1000 male births. Attempts have been made to link the XYY condition with a predisposition toward violence. This linkage is still hotly debated, although it is now clear that an XYY condition in no way guarantees such behavior. Nevertheless, several enterprising lawyers have attempted to use the XYY genotype as grounds for acquittal or compassion in crimes of violence. The XYY males are usually fertile. Their meioses are of the XY type; the extra Y is not transmitted, and their gametes contain either X or Y, never YY or XY. The most common type of viable human aneuploid is Down syndrome (Figure 18-21), occurring at a frequency of about 0.15 percent of all live births. We have already encountered the translocation form of Down syndrome in Chapter 17. However, by far the most common type of Down syndrome is trisomy 21, caused by nondisjunction of chromosome 21 in a parent who is chromosomally normal. Like any mechanism, chromosome disjunction is error prone and sometimes produces aneuploid gametes. In this type of Down syndrome, there is no family history of aneuploidy, unlike the translocation type described earlier. Down syndrome is related to maternal age; older mothers run a greatly elevated risk of having Down-syndrome children (Figure 18-22). For this reason, fetal chromosome analysis (by amniocentesis or by chorionic villus sampling) is now recommended for older mothers. A less pronounced paternal-age effect also has been demonstrated. Even though the maternal-age effect has been known for many years, its cause is still not known. Nonetheless, there are some interesting biological correlations. It is possible that one aspect of the strong maternal-age effect on nondisjunction is an age-dependent decrease in the probability of keeping the chromosomal tetrad together during prophase I of meiosis. Meiotic arrest of oocytes (female meiocytes) in late prophase I is a common phenomenon in many animals. In female humans, all oocytes are arrested at diplotene before birth. Meiosis continues only at menstruation, which means that proper chromosome associations in the tetrad must be maintained for decades. If we speculate that, by accident through time, these associations have an increasing probability of breaking down, we can envision a mechanism contributing to increased maternal nondisjunction with age. Consistent with this speculation, most nondisjunction related to the maternal-age effect is due to nondisjunction at anaphase I, not anaphase II. 12 勇者并非无所畏惧,而是能判断出有比恐惧更重要的东西. An Introduction to Genetic Analysis Chapter 18 Chromosome Mutation II: Changes in Chromosome Number The multiple phenotypes that make up Down syndrome include mental retardation, with an IQ in the 20 to 50 range; broad, flat face; eyes with an epicanthic fold; short stature; short hands with a crease across the middle; and a large, wrinkled tongue. Females may be fertile and may produce normal or trisomic progeny, but males have never reproduced. Mean life expectancy is about 17 years, and only 8 percent survive past age 40. The only other human autosomal trisomics to survive to birth are afflicted with either trisomy 13 (Patau syndrome) or trisomy 18 (Edwards syndrome). Both show severe physical and mental abnormalities. The generalized phenotype of trisomy 13 includes a harelip; a small, malformed head; “rockerbottom” feet; and a mean life expectancy of 130 days. That of trisomy 18 includes “faunlike” ears, a small jaw, a narrow pelvis, and rockerbottom feet; almost all babies with trisomy 18 die within the first few weeks after birth. Chromosome mutation in general plays a prominent role in determining genetic ill health in humans. Figure 18-23 summarizes the surprisingly high levels of various chromosome abnormalities at different human developmental stages. In fact, the incidence of chromosome mutations ranks close to that of gene mutations in human live births (Table 18-1). This fact is particularly surprising when we realize that virtually all the chromosome mutations listed in Table 18-1 arise anew with each generation. In contrast, gene mutations (as we shall see in Chapter 24) owe their level of incidence to a complex interplay of mutation rates and environmental selection that spans many human generations. When the frequencies of various chromosome mutations in live births are compared with the corresponding frequencies found in spontaneous abortions (Table 18-2), it becomes clear that the chromosome mutations that we know about as clinical abnormalities are just the tip of an iceberg of chromosome mutations. First, we see that many more types of abnormalities are produced than survive to birth; for example, trisomies of chromosome 2, 16, and 22 are relatively common in abortuses but never survive to birth. Second, the specific aberrations that survive are part of a much larger number that do not survive; for example, Down syndrome (trisomy 21) is produced at almost 20 times the frequency in live births. The comparison is even more striking for Turner syndrome (XO). An estimated minimum of 10 percent of conceptions have a major chromosome abnormality; our reproductive success depends on the natural weeding-out process that eliminates most of these abnormalities before birth. Incidentally, no evidence suggests that these aberrations are produced by environmental insult to our reproductive systems or that the frequency of the aberrations is increasing. MESSAGE Trisomics show the deleterious effects of genome inbalance and produce chromosome-specific modified phenotypic ratios. Disomics (n + 1) A disomic is an aberration of a haploid organism. In fungi, they can result from meiotic nondisjunction. In the fungus Neurospora (a haploid), an n − 1 meiotic product aborts and 13 勇者并非无所畏惧,而是能判断出有比恐惧更重要的东西. An Introduction to Genetic Analysis Chapter 18 Chromosome Mutation II: Changes in Chromosome Number does not darken like a normal ascospore; so we may detect MI and MII nondisjunctions by observing asci with 4:4 and 6:2 ratios of normal to aborted spores, respectively, as shown here. These diagrams correspond exactly to the outcomes of the chromosomal events shown in Figure 18-16. In these organisms, the disomic (n + 1) meiotic product becomes a disomic strain directly. The abortion patterns themselves are diagnostic for the presence of disomics in the asci. Another way of detecting disomics in fungi is to cross two strains with homologous chromosomes bearing multiple auxotrophic mutations; for example: From such a cross, large populations of ascospores are plated onto minimal medium. Only ascospores of genotype ++++++ can grow and form colonies. Most of these colonies are found to be disomics and not multiple crossover types. MESSAGE Disomics in fungi can be selected from asci showing special spore abortion patterns or as meiotic progeny that must contain homologous chromosomes from both parents. Somatic aneuploids Aneuploid cells can arise spontaneously in somatic tissue or in cell culture. In such cases, the initial result is a genetic mosaic of cell types. Human sexual mosaics—individuals whose bodies are a mixture of male and female tissue—are good examples. One type of sexual mosaic, (XO)(XYY), can be explained by postulating an XY zygote in which the Y chromatids fail to disjoin at an early mitotic division, so both go to one pole: 14 勇者并非无所畏惧,而是能判断出有比恐惧更重要的东西. An Introduction to Genetic Analysis Chapter 18 Chromosome Mutation II: Changes in Chromosome Number The phenotypic sex of such individuals depends on where the male and female sectors end up in the body. In the type of nondisjunction being considered, nondisjunction at a later mitotic division would produce a three-way mosaic (XY)(XO)(XYY), which contains a clone of normal male cells. Other sexual mosaics have different explanations; as examples, XO/XY is probably due to early X-chromosome loss in a male zygote (Figure 18-24), and (XX)(XY) is probably the result of a double fertilization (fused twins). In general, sexual mosaics are called gynandromorphs. Geneticists working with many species of experimental animals occasionally find gynandromorphs among their stocks. A classic example is the Drosophila gynandromorph shown in Figure 18-25. In this case, the zygote started out as a female heterozygous for two X-linked genes, white eye and miniature wing (w+m+/w m). Loss of the wild-type allele–bearing X chromosome at the first mitotic division resulted in the two cell lines and ultimately in a fly differing from one side to the other in sex, eye color, and size of wing. A similar gynandromorph in the Io moth is shown in Figure 18-26. MESSAGE Mitotic nondisjunction and other types of aberrant mitotic chromosome behavior can give rise to mosaics consisting of two or more chromosomally distinct cell types, including aneuploids. Somatic aneuploidy and its resulting mosaics are often observed in association with cancer. People suffering from chronic myeloid leukemia (CML), a cancer of the white blood cells, frequently harbor cells containing the so-called Philadelphia chromosome. This chromosome was once thought to represent an aneuploid condition, but it is now known to be a 15 勇者并非无所畏惧,而是能判断出有比恐惧更重要的东西. An Introduction to Genetic Analysis Chapter 18 Chromosome Mutation II: Changes in Chromosome Number translocation product in which part of the long arm of chromosome 22 attaches to the long arm of chromosome 9. However, CML patients often show aneuploidy in addition to the Philadelphia chromosome. In one study of 67 people with CML, 33 proved to have an extra Philadelphia chromosome and the remainder had various aneuploidies; the most common aneuploidy was trisomy for the long arm of chromosome 17, which was detected in 28 people. Of 58 people with acute myeloid leukemia, 21 were shown to have aneuploidy for chromosome 8; 16 for chromosome 9; and 10 for chromosome 21. In another study of 15 patients with intestinal tumors, 12 had cells with abnormal chromosomes, at least some with trisomy for chromosome 8, 13, 15, 17, or 21. Such studies merely established correlations, and it is not clear whether the abnormalities are best thought of as a cause or as an effect of cancer. MESSAGE Aneuploids are produced by nondisjunction or some other type of chromosome misdivision at either meiosis or mitosis. Mechanisms of gene imbalance In considering aberrant euploidy, we noted that an increase in the number of full chromosome sets correlates with increased organism size but that the general shape and proportions of the organism remain very much the same. In contrast, autosomal aneuploidy typically alters the shape and proportions in characteristic ways. Plants tend to be somewhat more tolerant of aneuploidy than are animals. Studies of the jimsonweed, Datura, provide a classic example of the effects of aneuploidy and polyploidy. In the jimsonweed, the haploid chromosome number is 12. As is expected, the polyploid jimsonweed is proportioned like the normal diploids, only larger. In contrast, each of the 12 possible trisomics is disproportionate, but in ways different from one another, as exemplified by changes in the shape of the seed capsule (see Figure 3-7). The 12 different trisomies lead to 12 different and characteristic shape changes in the capsule. Indeed, these and other characteristics of the individual trisomies are so reliable that the phenotypic syndrome can be used to identify plants carrying a particular trisomy. Similarly, the 12 monosomies are themselves different from one another and from each of the trisomies. In general, a monosomic for a particular chromosome is more severely abnormal than is the corresponding trisomic. We see similar trends in aneuploids of animals as well. In the fruit fly, Drosophila, the only autosomal aneuploids that survive to adulthood are trisomics and monosomics for chromosome 4, which is the smallest Drosophila chromosome, representing from only about 1 to 2 percent of the genome. Trisomics for chromosome 4 are only very mildly affected and are much less severely abnormal than are monosomic-4 flys. In humans, no autosomal monosomic survives to birth, whereas three autosomal trisomies survive, as mentioned earlier. As is true with aneuploid jimsonweed, the three surviving trisomies produce unique phenotypic syndromes, owing to the special effects of altered dosages of each of these chromosomes. Why are aneuploids so much more abnormal than polyploids? Why do aneuploids for different chromosomes each have their own characteristic phenotypic effects? And why are 16 勇者并非无所畏惧,而是能判断出有比恐惧更重要的东西. An Introduction to Genetic Analysis Chapter 18 Chromosome Mutation II: Changes in Chromosome Number monosomics typically more severely affected than the corresponding trisomics? The answers seem certain to be a matter of gene balance. In a euploid, the ratio of genes on any one chromosome to genes on other chromosomes is 1:1 (that is, 100 percent), regardless of whether we are considering a monoploid, diploid, triploid, or tetraploid. In contrast, in an aneuploid, the ratio of genes on the aneuploid chromosome to genes on the other chromosomes differs from wild type by 50 percent (50 percent for monosomics; 150 percent for trisomics). Thus, we can see that the aneuploid genes are out of balance. How does this help us to answer the questions raised? A key fact is that, in general, the amount of transcript produced by a gene is directly proportional to the number of copies of that gene in a cell. That is, for a given gene, the rate of transcription is directly related to the number of DNA templates. Thus, the more copies of that gene, the more transcripts are produced. Because of this gene-dosage relation, segmental aneuploids, in which pieces of individual chromosomes are trisomic or monosomic (effectively, duplications and deletions), have proved to be very useful in locating the positions of genes encoding various cellular enzymes. The approach is to look for segments of the genome that change the amount of an enzyme proportionally to the dosage of that genomic segment. This approach has been exploited extensively in Drosophila, where there has been about a 90 percent success rate in identifying enzymecoding gene loci by this method. We can infer that normal physiology in a cell depends on the proper ratio of gene products in the euploid cell. This ratio is the normal gene balance. If the relative dosage of certain genes changes—for example, owing to the removal of one of the two copies of a chromosome or a segment thereof—physiological imbalances in cellular pathways can arise. In some cases, the imbalances of aneuploidy are due to a few “major” genes. Such genes can be viewed as haplo-abnormal or triploabnormal or both and contribute significantly to the aneuploid phenotypic syndrome. For example, the study of persons trisomic for only part of human chromosome 21 has made it possible to localize determinants specific to Down syndrome to various regions of chromosome 21, hinting that some aspects of the phenotype might be due to trisomy for single major genes in these chromosomal regions. In addition to these major-gene effects, other aspects of aneuploid syndromes are likely to be due to cumulative effects of aneuploidy for numerous genes whose products are all out of balance. Undoubtedly, the entire aneuploid phenotype is a synthesis of the imbalance effects of a few major genes, together with a cumulative imbalance of many minor genes. However, the gene-balance idea does not tell us why having too few gene products (monosomy) is much worse for an organism than having too many gene products (trisomy). Along the same lines, in well-studied organisms, there are many more haploabnormal genes than triploabnormal ones. An important factor in explaining the abnormality of monosomics is that any deleterious recessives present on the autosome will be automatically expressed. This same effect is relevant to deletion mutations. 17 勇者并非无所畏惧,而是能判断出有比恐惧更重要的东西. An Introduction to Genetic Analysis Chapter 18 Chromosome Mutation II: Changes in Chromosome Number How do we apply the idea of gene balance to cases of sex-chromosome aneuploidy? Gene balance holds for sex chromosomes as well, but we also have to take into account the special properties of the sex chromosomes. In organisms with X-Y sex determination, the Y chromosome seems to be a degenerate X chromosome in which there are very few functional genes other than some involved in sex determination itself or in sperm production or both. On the other hand, the X chromosome contains many genes involved in basic cellular processes (“housekeeping genes”) that just happen to reside on the chromosome that eventually evolved into the X chromosome. X-Y sex-determination mechanisms have probably evolved independently from 10 to 20 times in different taxonomic groups. Thus, there appears to be one sex-determination mechanism for all mammals, but it is completely different from the mechanism governing X-Y sex determination in fruit flies. In a sense, X chromosomes are naturally aneuploid. Females have two of them, whereas males have only one. Nonetheless, it has been found that the X chromosome's housekeeping genes are expressed to equal extents per cell in both females and males. How is this accomplished? The answer depends on the organism. In fruit flies, the male's X chromosome appears to be hyperactivated, allowing it to be transcribed at twice the rate of either X chromosome in the female. In mammals, in contrast, the rule is that no matter how many X chromosomes are present, there is only one transcriptionally active X chromosome in each somatic cell. This dosage compensation is achieved by random Xchromosome inactivation. (A person with two or more X chromosomes is a mosaic of two cell types in which one or the other X is active.) Thus, XY and XX mammals produce the same amount of X-chromosome housekeeping gene products. X-chromosome inactivation also explains why triplo-X humans are phenotypically normal, inasmuch as only one of the three X chromosomes is transcriptionally active in a given cell. Similarly, an XXY male is only moderately affected because only one of his two X chromosomes is active in each cell. Why are XXY persons abnormal at all, given that triplo-X persons are phenotypically normal? It turns out that a small part of the X chromosome near one telomere—the pseudoautosomal region—is not inactivated by the mechanism of dosage compensation. In XXY males, this region is active at twice the level of the pseudoautosomal region in XY males. This level appears to have the consequence of slightly feminizing the phenotype of XXY males, although exactly which pseudoautosomal genes contribute to this effect is currently unknown. In XXX females, on the other hand, the pseudoautosomal region is active at only 1.5 times the level that it is in XX females. This lower level of functional aneuploidy in XXX than that in XXY plus the fact that the pseudoautosomal genes appear to lead to feminization may explain the feminized phenotype of XXY males. The severity of XO, Turner syndrome, can be interpreted as being due to the considerable deleterious effects of monosomy for the pseudoautosomal region of the X. As is usually observed for aneuploids, monosomy for this segment of the X chromosome produces a more abnormal phenotype than does having an extra copy of the same region (triplo-X females or XXY males). As noted earlier, occasionally human polyploids are born, but none survive. This fact seems to violate the principle discussed earlier in this section—namely, that polyploids are more normal 18 勇者并非无所畏惧,而是能判断出有比恐惧更重要的东西. An Introduction to Genetic Analysis Chapter 18 Chromosome Mutation II: Changes in Chromosome Number than aneuploids. The explanation for this violation seems to lie with X-chromosome dosage compensation. Part of the rule for the single active X seems to be that there is one active X for every two copies of the autosomal chromosome complement. Thus, some cells in triploid mammals are found to have one active X, whereas others have two. Neither situation is in balance with autosomal genes. Presumably this functional underactivation of housekeeping genes (in 3n cells with one active X) or functional overactivation (in 3n cells with two active X's) leads to substantial functional aneuploidy and the inviability of triploid humans. Chromosome mechanics in plant breeding What we know about chromosome mutation can be used in the type of genetic engineering that produces and maintains new crop types in our hungry world. The classic example, performed by E. R. Sears in the 1950s, concerns the transfer of a gene for leaf-rust resistance from a wild grass, Aegilops umbellulata, to bread wheat, which is highly susceptible to this disease, to offset a major problem in the wheat industry (Figure 18-27). The first problem that Sears encountered was that these two species (Figure 18-28) are not interfertile, so the feat of gene transfer seemed impossible. Sears sidestepped this problem with a bridging cross, in which he crossed A. umbellulata to a wild relative of bread wheat called emmer, Triticum dicoccoides. (Follow the process in Figure 18-29.) A. umbellulata is a diploid, 2n = 2x = 14. We will call its chromosome sets CC. T. dicoccoides is a tetraploid, 2n = 4x = 28, with sets AA BB. From this cross, the resulting sterile hybrid A B C was doubled into a fertile amphidiploid AA BB CC with 42 chromosomes. This amphidiploid was fertile in crosses with bread wheat (T. aestivum), which is represented as 2n = 6x = 42, AA BB DD. The offspring, AA BB CD, were almost completely sterile owing to pairing irregularities between the C and D sets. However, backcrosses to wheat did produce a few rare seeds, some of which grew into plants that were resistant to leaf rust, like the wild parent in the first cross. Some of these plants were almost the desired type, having 43 chromosomes (42 of which were wheat, plus one Aegilops chromosome bearing the resistance gene). Thus, in the AA BB CD hybrid, some aberrant form of chromosome assortment had produced a gamete with 22 chromosomes: A B D plus one from the C group. Unfortunately, the extra chromosome carried just too many undesirable Aegilops genes along with the good one, and the plants were weedy and low producers. So the Aegilops gene linkage had to be broken. Sears did so by using irradiated pollen from these plants to pollinate wheat. He was looking for translocations of part of the Aegilops chromosome tacked onto the wheat chromosomes. These translocations were quite common, but only one turned out to be ideal—a very small, insertional, unidirectional translocation (Figure 18-30). When bred to homozygosity, the resistant plants were indistinguishable from wheat. The study of chromosome mutation is of great importance—not only in pure biology, where there are ramifications in many areas (especially in evolution), but also in applied biology. The application of such studies is especially important in agriculture and medicine. Rather curiously, very little is known about the mechanisms of chromosome mutation, especially at 19 勇者并非无所畏惧,而是能判断出有比恐惧更重要的东西. An Introduction to Genetic Analysis Chapter 18 Chromosome Mutation II: Changes in Chromosome Number the molecular level. More details are needed about the normal chemical architecture of chromosomes. Summary When chromosome mutation changes the number of whole chromosome sets present, aberrant euploids result; when chromosome mutation changes parts of sets, aneuploids result. Polyploids such as triploids (3x) and tetraploids (4x) are common in the plant kingdom and are even represented in the animal kingdom. An odd number of chromosome sets makes an organism sterile because there is not a partner for each chromosome at meiosis, whereas even numbers of sets can produce standard segregation ratios. Allopolyploids (polyploids formed by combining sets from different species) can be made by crossing two related species and then doubling the progeny chromosomes through the use of colchicine; they can also be made through somatic cell fusion. These techniques have important applications in crop breeding, because allopolyploids combine features of both parental species. Polyploidy can result in an organism of larger dimensions; this discovery has permitted important advances to be made in horticulture and in crop breeding. Aneuploids have also been important in the engineering of specific crop genotypes, although aneuploidy itself usually results in an unbalanced genome with an abnormal phenotype. Examples of aneuploids include monosomics (2n − 1), trisomics (2n + 1), and disomics n + 1).Aneuploid conditions are well studied in humans. Down syndrome (trisomy 21), Klinefelter syndrome (XXY), and Turner syndrome (XO) are well-documented examples. The spontaneous level of aneuploidy in humans is quite high and produces a major percentage of genetically based ill health in human populations. Most instances of aneuploidy result from chromosome nondisjunction at meiosis or chromatid nondisjunction at mitosis. Mitotic nondisjunction can lead to somatic mosaics. Solved Problems 1. There is a controversy about the type of chromosome pairing seen in autotetraploids formed between two geographical races of a certain plant. It is known that chromosomes associate by pairs, but three hypotheses are put forward concerning how pairs form: a. Pair formation is random. b. Pairs form only between chromosomes of the same race. c. Pairs form only between chromosomes of different races. Consider a locus, A, that is closely linked to the centromere. The following cross is made: The autotetraploid is then selfed. What ratio of phenotypes can be expected under each hypothesis of chromosome pairing? Explain your answer. See answer 20 勇者并非无所畏惧,而是能判断出有比恐惧更重要的东西. An Introduction to Genetic Analysis Chapter 18 Chromosome Mutation II: Changes in Chromosome Number Solution a. Under random pairing, all possible pairing combinations are equally likely. If we label the four homologous chromosomes 1, 2, 3, and 4, then the equally probable combinations are 1-2/3-4, 1-3/2-4, and 1-4/2-3. This kind of situation in an A/A/a/a tetraploid was examined in the chapter (see page 560). We saw that a/a gametes are produced at a frequency of 1/6 and that the frequency of a/a/a/a progeny from selfing must therefore be 1/36 All other progeny types contain at least one A allele. Hence, the expected ratio is 35 A/–/–/–:1 a/a/a/a. b. In this alternative, the chromosome pairs look like this: because both the chromosomes carrying A come from race 1 and both the chromosomes carrying a come from race 2. This diagram shows how being methodical can help in problem solving, because it makes it very clear that the only possible segregation is that of one A and a to each pole. Hence, all gametes are A/a and, on self-fertilization, all genotypes would be A/A/a/a; only the A phenotype would be seen. c. Here the pairing looks like this: Because segregation of the pairs is independent, the gametic population can be represented as follows: Self-fertilization would produce a/a/a/a progeny at a frequency of 1/4 × 1/4 = 1/16. Therefore, 15/16 of the progeny are A/–/–/– and a 15:1 phenotypic ratio is predicted. In conclusion, the ratio of phenotype A to phenotype a in each case is a. 35:1 b. 1:0 c. 15:1 2. Owing to the small size of the Drosophila chromosome 4, monosomics and trisomics for this chromosome are viable, but tetrasomics and nullisomics are not. A fly that is trisomic for chromosome 4 and carries the recessive gene for bent bristles (b) on all copies of chromosome 4 is crossed to a phenotypically normal fly that is monosomic for chromosome 4. a. What genotypes and phenotypes can be expected in the progeny and in what proportions? b. If trisomics from these progeny are interbred, what phenotypic ratio can be expected in the next generation? Assume that only one copy of b+ is needed to produce normal (nonbent) bristles, that unpaired chromosomes pass to either pole at random, and that 21 勇者并非无所畏惧,而是能判断出有比恐惧更重要的东西. An Introduction to Genetic Analysis Chapter 18 Chromosome Mutation II: Changes in Chromosome Number aneuploid gametes of any kind survive. See answer Solution a. The cross is From the bent-bristled parent, the gametes will be 1/2b/b and 1/2b From the monosomic parent, the gametes will be 1/2b+ and 1/20 (the latter contain no chromosome 4). Hence, the progeny will be 1/4b+/b/b, 1/4b/b, 1/4b+/b, and 1/4b, which provides a phenotypic ratio of 1:1. b. The trisomics referred to are of genotype b+/b/b. If we label these chromosomes 1, 2, and 3, then and segregation produces Fertilization can be represented as follows: The ×'s represent tetrasomics, which, we are told, are not viable. Of the remaining 27/36, 8/36, are of b phenotype; therefore, a ratio of 19:8 is predicted. Problems 1. Distinguish between Klinefelter, Down, and Turner syndromes in humans. See answer 2. List two ways in which you could make an allotetraploid between two related diploid plant species 2n = 28. 3. a. Suppose that nondisjunction of Neurospora chromosome 3 occurs at the second division of meiosis. Show the content of each of the eight resultant ascospores with regard to chromosome 3. b. Neurospora normally has seven chromosomes. How many chromosomes are present in each of the ascospores in part a? See answer 22 勇者并非无所畏惧,而是能判断出有比恐惧更重要的东西. An Introduction to Genetic Analysis Chapter 18 Chromosome Mutation II: Changes in Chromosome Number 4. a. How would you synthesize a pentaploid (5x)? b. How would you synthesize a triploid (3x) of genotype A/a/a? c. You have just obtained a rare recessive mutation a* in a diploid plant, which Mendelian analysis tells you is A/a*. From this plant, how would you synthesize a tetraploid (4x) of genotype A/A/a*/a*? d. How would you synthesize a tetraploid of genotype A/a/a/a? e. How would you synthesize a plant that is resistant to a chemical herbicide? (Assume that mutation to this trait is very infrequent, and you are lucky to get one mutant.) 5. Which of the following phrases correctly completes the sentence? Allopolyploids are (a) not fertile at all; (b) fertile only among themselves; (c) fertile with one parent only; (d) fertile with both parents only; (e) fertile with both parents and themselves. See answer 6. Tetraploid yeast can be created by fusing two diploid cells. These tetraploids undergo meiosis like any other tetraploid and produce four diploid products of meiosis. Assume that homologous chromosomes synapse randomly in pairs and that there is no crossing-over in the region between the B locus and the centromere. What nonlinear tetrads are produced by a tetraploid of genotype B/B/b/b? What are the frequencies of the ascus types? (Note: This question requires tetrad analysis of tetraploid cells instead of the usual diploid cells.) 7. Consider a tetraploid of genotype A/A/a/a in which there is no pairing between chromosomes from the same parent. In another tetraploid of genotype B/B/b/b, pairs form only between chromosomes from the same parent. What phenotypic ratios result from selfing in each of these cases? (Assume that one parent carries the dominant allele and that the other carries the recessive allele in each case.) See answer 8. In the plant genus Triticum, there are many different polyploid species, as well as diploid species. Crosses were made between some different species, and hybrids were obtained. The meiotic pairing was observed in each hybrid and is recorded in the following table. (A bivalent is two homologous chromosomes paired at meiosis, and a univalent is an unpaired chromosome at meiosis.) Explain these results and, in doing so, a. deduce the somatic chromosome number of each species used. 23 勇者并非无所畏惧,而是能判断出有比恐惧更重要的东西. An Introduction to Genetic Analysis Chapter 18 Chromosome Mutation II: Changes in Chromosome Number b. state which species are polyploid, and whether they are auto- or allopolyploids. c. account for the chromosome pairing pattern in the three hybrids. 9. The New World cotton species Gossypium hirsutum has a 2n chromosome number of 52. The Old World species G. thurberi and G. herbaceum each have a 2n number of 26. Hybrids between these species show the following chromosome pairing arrangements at meiosis: Draw diagrams to interpret these observations phylogenetically, clearly indicating the relations between the species. How would you go about proving that your interpretation is correct? (Problem 9 after A. M. Srb, R. D. Owen, and R. S. Edgar, General Genetics, 2d ed. W. H. Freeman and Company, 1965.) 10. The genotype of an autotetraploid that is heterozygous at two loci is F/F/f/f ; G/G/g/g. Each locus affects a different character, and the two loci are located very close to the centromere on different (nonhomologous) chromosomes. a. What gametic genotypes are produced by this individual and in what proportions? b. If the individual is self-fertilized, what proportion of the progeny will have the genotype F/F/F/f ; G/G/g/g? the genotype f/f/f/f ; g/g/g/g? 11. Which of the following abnormalities are not caused by meiotic nondisjunction? (a) Turner syndrome; (b) cri du chat syndrome; (c) Down syndrome; (d) Klinefelter syndrome; (e) XYY syndrome; (f) achondroplastic dwarfism; (g) Edwards syndrome; (h) Marfan syndrome. See answer 12. A woman with Turner syndrome is found to be colorblind. Both her mother and father have normal vision. How can her colorblindness be explained? Does this outcome tell us whether nondisjunction occurred in the father or in the mother? If the colorblindness gene were close to the centromere (it is not, in fact), would the available facts tell us whether the nondisjunction occurred at the first or at the second meiotic division? Repeat the question for a colorblind man with Klinefelter syndrome. See answer 13. Individuals have been found who are colorblind in one eye but not in the other. What would this finding suggest if these individuals were (a) only or mostly females? (b) only or mostly males? (Assume that this trait is X-linked recessive.) 24 勇者并非无所畏惧,而是能判断出有比恐惧更重要的东西. An Introduction to Genetic Analysis Chapter 18 Chromosome Mutation II: Changes in Chromosome Number 14. When geneticists treat human sperm with quinacrine dihydrochloride, about half the sperm show a fluorescent spot thought to be the Y chromosome. About 1.2 percent of normal human sperm show two fluorescent spots. Geneticists examined the sperm of some industrial workmen who were exposed for about 1 year to the chemical dibromochloropropane. They found the frequency of sperm with two spots to be on average 3.8 percent. Propose an explanation for this result, and explain how you would test your explanation. 15. A person with Turner syndrome is also found to have the phenotype nystagmus (an eye disorder). a. What event or events can explain the coincidence of the two hereditary conditions? b. In which parent and at which stage did the event or events occur? 16. In British Columbia, the mean age of women giving birth to Down syndrome babies fell from 34 years to 28 years between 1952 and 1972. What are two possible causes of this population trend, and how could the two hypotheses be tested? 17. Several kinds of sexual mosaicism are well documented in humans. Suggest how each of the following examples may have arisen: a. (XX)(XO) (that is, there are two cell types in the body, (XX and XO) b. (XX)(XXYY) c. (XO)(XXX) d. (XX)(XY) e. (XO)(XX)(XXX) See answer 18. The discovery of chromosome banding in eukaryotes has greatly improved the ability to distinguish various cytogenetic events. Particularly useful are banding polymorphisms, because the “morphs” can be used as chromosome markers. These morphs make chromosomes cytologically distinguishable by virtue of minor variation in band size, position, and so forth. Consider chromosome 21 in humans. Assume that one set of parents is 21a21b ♀ × 21c21d ♂, where a, b, c, and d represent morphs of a polymorphism for this chromosome. Also assume that fetuses of the following types are produced (where 42A stands for the rest of the autosomes): 1. 42A + 21b21b21c + XY 2. 42A + 21a21b21d + XX 3. 42A + 21b21d + XY 25 勇者并非无所畏惧,而是能判断出有比恐惧更重要的东西. An Introduction to Genetic Analysis Chapter 18 Chromosome Mutation II: Changes in Chromosome Number 4. 42A + 21a21c21c + XX 5. 42A + 21a21b + XY 6. 42A + 21a21c + XYY 7. (42A + 21a + XY)(42A + 21a21c21c + XY) (mosaic) 8. (42A + 21a21c + XY)(42A + 21b21d + XX) (mosaic) In each case: a. State the genetic term for the condition. b. Diagram the events that give rise to the condition. c. State the individual in which the events took place. 19. Suppose that you have a line of mice that has cytologically distinct forms of chromosome 4. The tip of the chromosome can have a knob (called 4K), or a satellite (4S), or neither (4). Here are sketches of the three types: You cross a 4K 4S female with a 4 4 male and find that most of the progeny are 4K 4 or 4S 4, as expected. However, you occasionally find some rare types as follows (all other chromosomes are normal): a. 4K 4K 4 b. 4K 4S 4 c. 4K d. 4/4K 4K 4 mosaic e. 4K 4/4S 4 mosaic Explain the rare types that you have found. Give as precisely as possible the stages at which they originate, and state if they are present in the male parent, the female parent, or the zygote. (Give reasons briefly.) See answer 20. In Drosophila, a cross (cross 1) is made between two mutant flies, one homozygous for the recessive mutation bent wing (b) and the other homozygous for the recessive mutation eyeless (e). The mutations e and b are alleles of two different genes that are known to be very closely linked on the tiny autosomal chromosome 4. All the progeny were wild-type phenotype. One of the female progeny was crossed to a male of genotype b e/b e; call this cross 2. The progeny of cross 2 were mostly of the expected types, but there was also one rare female of wild-type phenotype. a. Explain what the common progeny are expected to be from cross 2. b. Could the rare wild-type female have arisen by (1) crossing-over? (2) nondisjunction? Explain. c. The rare wild-type female was testcrossed to a male of genotype b e/b e (cross 3). The progeny were 26 勇者并非无所畏惧,而是能判断出有比恐惧更重要的东西. An Introduction to Genetic Analysis Chapter 18 Chromosome Mutation II: Changes in Chromosome Number Which of the explanations in part b are compatible with this result? Explain the genotypes and phenotypes of progeny of cross 3 and their proportions. Unpacking the Problem 1. Define homozygous, mutation, allele, closely linked, recessive, wild type, crossing-over, nondisjunction, testcross, phenotype, and genotype. 2. Does this problem concern sex linkage? Explain. 3. How many chromosomes does Drosophila have? 4. Draw a clear pedigree summarizing the results of crosses 1, 2, and 3. 5. Draw the gametes produced by both parents in cross 1. 6. Draw the chromosome 4 constitution of the progeny of cross 1. 7. Is it surprising that the progeny of cross 1 are wild-type phenotype? What does this outcome tell you? 8. Draw the chromosome 4 constitution of the male tester used in cross 2, and the gametes that he can produce. 9. With respect to chromosome 4, what gametes can the female parent in cross 2 produce in the absence of nondisjunction? Which would be common and which rare? 10. Draw first- and second-division meiotic nondisjunction in the female parent of cross 2, as well as the resulting gametes. 11. Are any of the gametes from part 10 aneuploid? 12. Would you expect the aneuploid gametes to give rise to viable progeny? Would these progeny be nullisomic, monosomic, disomic, or trisomic? 13. What progeny phenotypes would be produced by the various gametes considered in parts 9 and 10? 14. Consider the phenotypic ratio in the progeny of cross 3. Many genetic ratios are based on halves and quarters, but this ratio is based on thirds and sixths. What might this point to? 15. Could there be any significance to the fact that the crosses concern genes on a very small chromosome? When is chromosome size relevant in genetics? 16. Draw the progeny expected from cross 3 under the two hypotheses, and give some idea of relative proportions. See answer 21. A cross is made in tomatoes between a female plant that is trisomic for chromosome 6 and a normal diploid male plant that is homozygous for the recessive allele for potato leaf (p/p). a. A trisomic F1 plant is backcrossed to the potato-leaved male. What is the ratio of normal-leaved plants to potato-leaved plants when you assume that p is on chromosome 6? b. What is the ratio of normal- to potato-leaved plants when you assume that p is not 27 勇者并非无所畏惧,而是能判断出有比恐惧更重要的东西. An Introduction to Genetic Analysis Chapter 18 Chromosome Mutation II: Changes in Chromosome Number located on chromosome 6? 22. A tomato geneticist attempts to assign five recessive genes to specific chromosomes using trisomics. She crosses each homozygous mutant (2n) to each of three trisomics—chromosomes 1, 7, and 10. From these crosses, the geneticist selects trisomic progeny (which are less vigorous) and backcrosses them to the appropriate homozygous recessive. The diploid progeny from these crosses are examined. Her results follow, in which the ratios are wild type:mutant. Which of the genes can the geneticist assign to which chromosomes? (Explain your answer fully.) 23. Petunia plants have four loci, A, B, C, and D, that are known to be very closely linked. A plant of genotype a B c D/A b C d is irradiated with γ rays and then crossed to an a b c d/a b c d plant. In the progeny, plants of phenotype A B C D are occasionally found. What are two possible modes of origin, and which is the more likely? See answer 24. A cross in Neurospora between strains carrying the multiply marked chromosomes a b+c d+e and a+b c+d e+ produces one product of meiosis that grows on minimal medium. (Assume that a, b, c, d, and e determine nutritional requirements.) When the rare colony grows up, some asexual spores are a b+c d+e in genotype, some are a+b c+d e+, and the remainder grow on minimal medium. Explain the origin of the rare product of meiosis and the origin of the three types of asexual spores. 25. Two Neurospora auxotrophs are crossed: pan1 × leu2. These loci are linked on the same chromosome arm, with leu2 between the centromere and pan1. Most octads from this cross were of the type expected, but the following unexpected types also were obtained: a. fully prototrophic fully prototrophic fully prototrophic fully prototrophict abort abort abort abort b. pan-requiring pan-requiring 28 勇者并非无所畏惧,而是能判断出有比恐惧更重要的东西. An Introduction to Genetic Analysis Chapter 18 Chromosome Mutation II: Changes in Chromosome Number abort abort Leu-requiring Leu-requiring Leu-requiring Leu-requiring c. fully prototrophic fully prototrophic abort abort Leu-requiring Leu-requiring pan-requiring pan-requiring Give explanations for these three types. *26. The ascomycete Sordaria brevicollis has two closely linked complementing markers, b1 and b2, that result in buff-colored (light-brown) ascospores. In the cross b1 × b2, if one or both centromeres divide and separate precociously at the first division of meiosis, what patterns of spore colors are produced in those asci? How can these asci be distinguished from normal asci and from asci in which nondisjunction has occurred? (Note: Ascospores in this fungus are normally black, and nullisomic ascospores are white; assume that there is no crossing-over between b1 and b2.) 27. Design a test system for detecting agents in the human environment that are potentially capable of causing aneuploidy in eukaryotes. See answer 28. Two alleles determine flower color in the plant Datura: P determines purple; and p, white. The P locus is on the smallest chromosome, number 3. In plants trisomic for this chromosome, n + 1 pollen is never functional, and only half the n + 1 eggs are functional. Assume that P is always completely dominant to any number of p's, and that the locus is close to the centromere. What is the expected ratio of purple:white in the progeny of the following crosses? a. P/P/p ♀ × P/P/p ♂ b. P/p/p ♀ × P/p/p ♂ c. P/p/p ♀ × p/p ♂ 29. There are six main species in the Brassica genus: B. carinata, B. campestris, B. nigra, B. oleracea, B. juncea, and B. napus. You can deduce the interrelationships between these six species from the following table: 29 勇者并非无所畏惧,而是能判断出有比恐惧更重要的东西. An Introduction to Genetic Analysis Chapter 18 Chromosome Mutation II: Changes in Chromosome Number a. Deduce the chromosome number of B. campestris, B. nigra, and B. oleracea. b. Show clearly any evolutionary relationships between the six species that you can deduce at the chromosomal level. 30. In the fungus Ascobolus (similar to Neurospora), ascospores are normally black. The mutation f, producing fawn ascospores, is in a gene just to the right of the centromere on chromosome 6, whereas mutation b, producing beige ascospores, is in a gene just to the left of the same centromere. In a cross of fawn with beige parents (1f × b 1) most octads show four fawn and four beige ascospores, but three rare exceptional octads were found as shown here. In the sketch, black represents the wild-type phenotype, a vertical line represents fawn, a horizontal line represents beige, and an empty circle represents an aborted (dead) ascospore. a. Provide reasonable explanations for these three exceptional octads. b. Diagram the meiosis that gave rise to octad 2. 31. The life cycle of the haploid fungus Ascobolus is similar to that of Neurospora. A mutational treatment produced two mutant strains, 1 and 2, both of which when crossed to wild type gave unordered tetrads all of the following type (fawn is a light-brown color; 30 勇者并非无所畏惧,而是能判断出有比恐惧更重要的东西. An Introduction to Genetic Analysis Chapter 18 Chromosome Mutation II: Changes in Chromosome Number normally, crosses produce all black ascospores): a. What does this result show? Explain. The two mutant strains were crossed. Most of the unordered tetrads were of the following type: b. What does this result suggest? Explain. When large numbers of unordered tetrads were screened under the microscope, some rare ones that contained black spores were found. Four cases are shown here: (Note: Ascospores with extra genetic material survive, but those with less than a haploid genome abort.) c. Propose reasonable genetic explanations for each of these four rare cases. d. Do you think the mutations in the two original mutant strains were in one single gene? Explain. 32. In a certain tropical plant there are two genes, each of which produces color pigment. The R (red color;>r) and B (blue color; > b loci are centromere linked and on separate chromosomes. Tetraploid plants are made in which the chromosomes pair in twos at meiosis. The following cross is made: and an F1 is obtained and then selfed to give an F2. If only one dominant allele is required to give the dominant phenotype at each locus, what phenotypes are expected in the F2 and in what proportions? 33. A tomato geneticist working on Fr, a dominant mutant allele that causes rapid fruit ripening, decides to find out which chromosome contains this gene by using a set of lines, each of which is trisomic for one chromosome. To do so, she crosses a homozygous diploid mutant to each of the wild-type trisomic lines. a. A trisomic F1 plant is crossed to a diploid wild-type plant. What is the ratio of fast- to slow-ripening plants in the diploid progeny of this second cross if Fr is on the trisomic chromosome? Use diagrams to explain. 31 勇者并非无所畏惧,而是能判断出有比恐惧更重要的东西. An Introduction to Genetic Analysis Chapter 18 Chromosome Mutation II: Changes in Chromosome Number b. What is the ratio of fast- to slow-ripening plants in the diploid progeny of this second cross if Fr is not located on the trisomic chromosome? Use diagrams to explain. c. Here are the results of the crosses. On which chromosome is Fr and why? (Problem 33 from Tamara Western.) Chapter 18* 1. Klinefelter syndrome XXY male Down syndrome trisomy 21 Turner syndrome XO female 3. a. 3, 3, 3, 3, 3/3, 3/3, 0, 0 b. 7, 7, 7, 7, 8, 8, 6, 6 5. b 7. If there is no pairing between chromosomes from the same parent, then all pairs are A/a. The gametes are 1 A/A : 2 A/a : 1 a/a. With selfing, the progeny are 1/16 A/A/A/A, 4/16 A/A/A/a, 6/16 A/A/a/a, 4/16 A/a/a/a, 1/16 a/a/a/a. If there is no pairing between chromosomes from different parents, then all pairs are B/B and b/b and all gametes are B/b. Thus 100% of the progeny are B/B/b/b. 11. b, f, and h, and sometimes c 12. Colorblindness appears to be an X-linked recessive disorder. That means that the woman with Turner syndrome had to have obtained her sole X from her mother. She did not obtain a sex chromosome from her father, which indicates that nondisjunction occurred in him. The nondisjunction could have occurred at either MI or MII. If the colorblind patient had Klinefelter syndrome (XXY), then both X's must carry the allele for colorblindness. Therefore, nondisjunction had to occur in the mother. Remember that during meiosis I, given no crossover between the gene and the centromere, allelic 32 勇者并非无所畏惧,而是能判断出有比恐惧更重要的东西. An Introduction to Genetic Analysis Chapter 18 Chromosome Mutation II: Changes in Chromosome Number alternatives separate from each other. During meiosis II, identical alleles on sister chromatids separate. Therefore, the nondisjunctive event had to occur during meiosis II because both alleles are identical. 17. a. Loss of one X in the developing fetus after the two-celled stage. b. Nondisjunction leading to Klinefelter syndrome (XXY) followed by a nondisjunctive event in one cell for the Y chromosome after the two-celled stage, leading to XX and XXYY. c. Nondisjunction for X at the one-celled stage. d. Either fused XX and XY zygotes or fertilization of an egg and polar body by one sperm bearing an X and another bearing a Y, followed by fusion. e. Nondisjunction of X at the two-celled stage or later. 19. a. The extra chromosome must be from the mother. Because the chromosomes are identical, nondisjunction had to have occurred at MII. b. The extra chromosome must be from the mother. Because the chromosomes are not identical, nondisjunction had to have occurred at MI. c. The mother correctly contributed one chromosome, but the father did not contribute any chromosome 4. Therefore, nondisjunction occurred in the male at either meiotic division. d. One cell line lacks a maternal contribution, whereas the other has a double maternal contribution. Because the two lines are complementary, the best explanation is that nondisjunction occurred in the developing embryo in mitosis. e. Each cell line is normal, indicating that nondisjunction did not occur. The best explanation is that the second polar body was fertilized and was subsequently fused with the developing embryo. 20. a. The common progeny are b e+/b e and b+e/b e. b. The rare female could have come from crossing-over, which would have resulted in a gamete that was b+e+. The rare female also could have come from nondisjunction that resulted in a gamete that was b e+/b+e. Such a gamete might give rise to viable progeny. c. The female was a product of nondisjunction. 23. Radiation could have caused point mutations or induced recombination, but nondisjunction is a more likely explanation. 27. Aneuploidy is the result of nondisjunction. Therefore, any system that will detect nondisjunction will work. One of the easiest follows. Cross white-eyed females with red-eyed males and look for the reverse of X linkage in the progeny. Compare populations unexposed to any environmental pollutants with those exposed to different suspect agents. 33 勇者并非无所畏惧,而是能判断出有比恐惧更重要的东西.