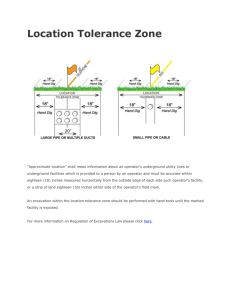
The Triune Brain as Neurobehavioral Basis of God Concept
advertisement

M. ERNANDES & M. WINKELMAN Theopoiesis or The Production of “God” Concepts by Brain Structures and Operators in Early Humans Michele (Michael) Ernandes: Department of Historical-Archaeological, Socio-Anthropological and Geographical Heritage, Palermo University, Scientific Centre of the Polytechnic of the Mediterranean, Italy. (i. e., Dipartimento di Beni Culturali Storico-Archeologici, Socio-Antropologici e Geografici, Università di Palermo, Italia) Email: ernandes@unipa.it Michael Winkelman: School of Human Evolution & Social Change Arizona State University Email: michael.winkelman@asu.edu Short Abstract Religious ideas have the peculiarity of being universal, so we may suspect yhat they have some kind of genetic basis, or that religion may be rooted in genetic predisposition, that operates through the development of encephalic structures. MacLean elaborated the “Triune Brain” model, and located in it three principal phylogenetic structures, termed as R-complex, Limbic System and Neomammalian brain. The R-complex is fundamental for genetically constituted behaviors as imprinting, or forming social hierarchies. In a behavioral sense, religious activities consist in submissive displays performed to appease a dominant individual. According to evolutionary psychology, all human behaviors are products of internal mechanisms, in conjunction with inputs that trigger their activation. In this paper we indicate the acquisition of the consciousness of own mortality as the input, and in the action of R-complex on the Neocortex as the main mechanism by which, in the human evolution, the “God” concept emerged. Long Abstract. Religious ideas have the peculiarity of being universal, so we may suspect that they may be rooted in genetic predisposition, that operates through the development of encephalic structures. MacLean described primate’s brain as formed by three principal philogenetic structure that have been super- imposed and have been integrated during evolution, and termed them as R – complex, Limbic System, and Neomammalian Brain. The R-complex is fundamental for genetically constituted forms of behavior as hunting, homing, mating, breeding, imprinting and forming social hierarchies. The Limbic Ssystem may be seen as a regulator of the R–complex, and most of this regulation seems to be inhibitory, while the Neomammalian brain is the main seat of mind capabilities as, in humans, selfconsciousness or the connections of causality. The most important religious displays consist in the gathering of several human groups who perform repeated and prolonged demonstrations of submission toward dominant individuals. These dominant individuals take different forms in each culture but they share some characteristics including an immense power. For what concerns the genesis of the ideas of such immense power beings, some scholars (as Freud or Morris) felt that such beings could be the projection result of the figure of the dominant male of primate’s “Single Male” breeding group. But, on the basis of the sexual dimorphism noticed in fossils, and inferring social behaviour from it, most scholars think that in ancient groups of Hominini endowed with low sexual dimorphism (as in genus Homo), there wasn’t any individual that could act as realistic example of a “Being with immense power”. How did it happen, therefore, that among human beings with a low male-male competition social system we can find a projection in the super- human world of a being with immense power that should have to be associated with a high male-male competition social system? A possible answer is that the human brain preserved structures and hierarchy forming functions in its R-complex. Consequently we can deduce that at some time in the course of human evolution the hierarchy forming structure of the R-complex has been set free from the inhibitory action of the Limbic System; this was presumably a consequence of a strong external stimulus which, causing a psychic trauma, weakened the inhibitory action of the Limbic System. Indeed, according to evolutionary psychology, all human behaviors are a product of internal mechanisms in conjunction with inputs that cause activation of those mechanisms. We may presume that the “input” for the Prehistoric man consisted in the acquisition of the awareness of his own mortality. Homo sapiens is conscious of being mortal, but he does not recognize this fact as a natural datum. Instead he feels that death is a violence he has to suffer: we may presume that in this way death could have been considered by early humans. As Neocortex seeks to determine agents who cause phenomena, early humans tried to find the cause of death, but failed to find an empirical cause. Because of this shock, the Limbic System activity on R-complex would have had a variation that caused the activation of hierarchic R-complex structures and that led the Neocortical structures to accept the idea, proposed by the R-complex, that a “Powerful, but unseen, Being” was the agent of death. After this, the Neocortical association areas, in relations to environment, developed various systems of religion. § I. Definition, observations and assumption concerning the Biological Basis of Religion Tylor (1871) defined religion as belief in immaterial and extraordinary beings as souls, ghosts, saints, angels, demons, and gods. This definition has been emphasized by many authors, such as M. Harris (1989), who characterized religion in terms of belief in the existence of one or more of such beings. There are other broader definitions of religion, but the supernatural agent concept is invariably central, as it is for our present purpose. Since the 16th century, ethnographic studies have suggested that such religious ideas are universal. In 1744 Giambattista Vico noticed that all cultures have had three institution: Religion, marriage, and burial. Contemporary scholars also asserted the universality of religious ideas, such as A. Brelich (1970): “No society has been found, even among the most ‘primitive,’ which was devoid of any faith in divine person-like beings.” Such sentiments were elaborated by Rappaport (1971, p. 23), who claimed “Neither history nor anthropology knows of societies from which religion has been totally absent” and echoed more recently by Boyer (2003, p. 119) who asserts that “Religious beliefs and practices are found in all human groups”. Many anthropologists would concur that when an aspect of behavior is universal to human societies, it is likely that it has some kind of genetic basis. It seems reasonable to suppose that religion has a biological basis given that it is universal, and surmise that some aspects of religious behavior and thought may be hardwired in a genetic predisposition. Spirits seem naturally conceivable, thinkable, to the human mind - and the human brain must have certain anatomic and physiologic structures which have favored the birth of religious ideas that subsequently become a part of the cultural heritage. In this vein, Boyer (2003, p. 123) notes that “Cognitive science and neuroscience suggests a less dramatic but perhaps more empirically grounded picture of religion as a probable, although by no means inevitable by-product of the normal operation of human cognition.” Religious thoughts and behaviors may be studied at various levels: a first level may regard the biological bases, neurological or ethological, of religious thought (Mandell 1980, Gazzaniga 1985, Alper 1996, Burkert 1996, Ernandes & Giammanco 1998, Winkelman 2000, W. and Baker 2008, Boyer 2001, 2003, Hamer 2004); a second level, phenomenologic-structural, the subordinate relationship of human beings to divine beings (Hubert and Mauss 1898, van der Leew 1933, Widengren 1969, Burkert 1972), as well as semeiotic values of religious thoughts and behaviors; a third, the relationships between social structures and peculiar shapes of credence and rituals (Weber 1904/05, Durkheim 1912, and their sociological schools) or the social value of religious rites (Sosis and Bressler 2003); a fourth, the economic-ecological one, the material goods used for religious rituals (Firth 1963, Harris 1977). These four approaches can coexist and interact synchronically with a fifth one, the historical or diachronic aspect. Obviously, for a given religion, results of studies carried out on one level must be coherent with the results obtained for other levels, since a comprehensive study must be non-contradictory. Until now the tendency to favor a particular type of study to the exclusion of others has prevailed (Pals 1996). This may have been reasonable in the past, as these various approaches were discovered little by little, but it is less excusable now. In 1966 C. Geertz noted stagnation in the field of anthropology of religion and foreboded the utilization of other disciplines such as philosophy, history or hard sciences with the aim of achieving a thick description of religion. As noted by Sosis and Alcorta (2003, p. 264), “the multiple roles and complex functions of religion render it difficult to capture within a single theoretical approach”, so it requires different methodological tools. In the present article we start from biological evolution, neurobiology and ethology with the aim of furnishing the ground for a multilevel study of religion. Returning to Tylor's definition, religious thought consists of belief in extra-natural power beings. We conceptualize an “Immense Power Being” as the most general and comprehensive of these beings, defined as a supernatural being with immense power over human lives. As has been established by Kant (1781), it is impossible to prove rationally the existence or the non-existence of any “Immense Power Being” (or, in Kant’s words, “God”). Consequently, our purpose is not to know if “God” exists or does not, but rather consists in an effort to understand why “God” concepts exists so naturally and comfortably in the human mind, why such ideas exist universally in human beings. It is pertinent to investigate such a question by means of the natural sciences, because “Thoughts and beliefs are necessarily dependent on neurophysiological activity of the brain” (Delgado 1969). According to evolutionary psychology, all human (or animal) behavior is a product of mechanisms internal to the person (or to the animal), in conjunction with inputs that trigger the activation of those mechanisms. No mechanisms, no behavior; no input, no behavior (adapted from Buss & Shackelford 1997, p. 607). If we consider the production of ideas as a particular kind of behavior (an “internal” brain behavior), our aim then consists in investigating the internal mechanisms and the input that had lead human mind to conceive the “Immense Power Being” concept. § II. MacLean’s Triune Model of the Brain Genetic predisposition operates through the development of encephalic structures. In Vertebrates the Central Nervous System grows up as a dorsal tube that, subsequently, is surrounded by vertebrae (the spinal cord) and by skull (the encephalon). A canal goes along the tube and forms at first three vesicle with surrounding swellings (forebrain, midbrain and hindbrain). These formations then become five ones: telencephalon (endbrain), diencephalon (interbrain), mesencephalon (midbrain), metencephalon (pons and cerebellum), and myelencephalon (medulla oblongata). (Fig. II. 1). At the end of the development, the hindbrain vesicle is termed fourth ventricle, the midbrain vesicle becomes thin and is called aquaeductus mesencephali (or cerebri), the interbrain vesicle forms the third ventricle. The telencephalic vesicle, in growing forward impeded by the frontal bone, in its development overturns and covers the diencephalon, splitting and forming two lateral ventricles. In a section of each telencephalic vesicle we may note four parts: the lateral and medial basal ones, and the lateral and medial dorsal ones. (Fig. II. 2 a). In the basal parts neurons form some masses (called nuclei or ganglia), while in the dorsal parts neurons are disposed in layers (normally three). P. D. MacLean (1970-1990) elaborated a model of brain structure and evolution. He described it as a “Triune brain”, because he located in it three principal phylogenetic structures that have been superimposed and that have become integrated during evolution. He termed these three basic types reptilian (Protoreptilian, R-complex), old mammalian (Paleomammalian, Limbic System) and new mammalian (Neomammalian). The heuristic value of this model, for a long time underappreciated, has been revalued by many scholars (Winkelman 2000; Cory & Gardner 2002; Oatley 2004; Whybrow 2005). Here we examine the triune brain model, reappraising it in relation with recent knowledge in neurobiological and evolutionary sciences, and in terms of its applicability to explaining the neurological basis of the “Immense Power Being.” The R-complex The protoreptilian brain represents a fundamental core of the nervous system: it consists in the upper spinal cord, parts of the midbrain and the diencephalons, and the lateral basal part of the telencephalic vesicle. The lateral basal telencephalon consists in the striatum and pallidum as main components, and in the substantia nigra, ventral tegmental area and subthalamic nucleus as smaller components. The striatum comprises dorsally the caudate nucleus and putamen, ventrally the nucleus accumbens and the olfactory tubercle. The globus pallidus (or pallidum) consists in three parts: the internal and the external segments, and the ventral pallidum (fig. II. 3). The ventral part of the striatum and the ventral pallidum are also called “paleostriatum”, the dorsal striatum “neostriatum”: however there is no evolutionary evidence supporting these names, that must be considered old-fashioned (fig. II.2 c). The medial basal part of the telencephalon gives rise to the septum, while the dorsal part forms the pallium (or cortex): these structures are not included in the R-complex, not because they are absent, but because they are less developed and are not indispensable in determining the fundamental behaviors in Reptiles and Birds (so that the term Striatal complex is more accurate but less suggestive than R-complex). The presence of the R-complex as a stage in the brain evolution from Fish to Mammals has been corroborated by subsequent research in anatomy and physiology. Reiner, Medina and Veenman (1998) have found that both striatum and pallidum are typically much more cell poor in Anamniotes (Fish and Amphibians) than they are in Amniotes (Reptiles, Birds and Mammals). Moreover, neural pathways connecting basal ganglia and the telencephalic cortex are less important in Reptiles and Birds than in Mammals (Striedter 2005), as it is shown in fig II. 5 f. According to MacLean, the R-complex in the brains of existing mammals derives from a form of mammal-like reptiles that, in Permian and Triassic geologic periods, populated the earth in large numbers. Even if for comparative neurobehavioral study, no existing reptiles are directly in line with mammals, it has been noticed that most ancient mammal-like reptiles were lizard-like in appearance, so that MacLean and following scholars have favored the use of lizards in their comparative studies. Moreover, the lizards chose are ones in which the cortical part of telencephalon is small, and not those ones provided with evident pallial gyri, as the tegu lizard (Tupinambis teguixin), which has a paleo and an archeo cortical gyri surrounding a neocortical gyrus. MacLean (1973a, p. 8) wrote that “the counterpart of the reptilian brain in mammals is fundamental for genetically constituted forms of behavior as selecting homesites, establishing territory engaging in various types of display, hunting, homing, mating, breeding, imprinting, forming social hierarchies, and selecting leaders”. In mammals, “the R-complex is necessary for ritualistic displays and the averbal communication associated with them. At the human level … certain behavioral tendencies are due to an inheritance of dispositions mediated by this same, primal brain region. These include certain violent reactions, the preference for routine or “ritualistic” actions, and some forms of displacement activities” (Isaacson 1982, p. 246). The Limbic System. From Fish to Reptiles, the basal parts of the telencephalon are more abounding in neurons than the dorsal parts, which appear as a thin mantle (the pallium) or cortex that covers the lateral vesicle. In Mammals the cortex expands a lot, wrapping the basal part of the telencephalon. Ariëns Kappers (1909) termed the lateral cortex (whose main derived region is called piriform cortex) as paleopallium, and the medial cortex (whose main structure is the hyppocampus) as archipallium. In these nouns the paleo- and archi- prefixes have no more the temporal or evolutionary significance that first XX century authors intended give them. Their significance, related to pallium, is now only spatial (fig. II 2 b, c and d). Paleo- and archipallium neurons are disposed in three layers: this disposition is termed allocortex. In the phylogenetically subsequent neocortex, derived from the dorsal pallium, neurons are disposed in six layers to form the isocortex. Paleo- and archi-pallium form the largest regions of the Limbic System, which also includes the olfactory bulb, septum, fornix, amygdala, and cingulated gyrus. “The paleomammalian brain, or the limbic system, represents an advance in neural tissue because it represents a device for providing the animals that have this tissue with better means of coping with the environment. Parts of the limbic system are concerned with primal activities related to food and sex; others are related to emotions and feelings; and still others combine messages from the external world with those from inside” (Isaacson 1982, p. 246). MacLean (1990) has identified three main limbic subdivisions located respectively in the amygdale, septum, and thalamus that are a source of afferents to the limbic cortex (fig. II. 4). I) The amygdalar division comprises the amygdala together with the frontotemporal limbic cortex; the amygdala is also connected with the hippocampus. This division is primarily involved with fear, aggression, and mimic judgment (Morris et al. 1998; Adolphs et al. 1998). II) The septal division projects to the entire hippocampal formation and also includes the entorhinal cortex (gyrus hippocampi). This division is mainly connected with sex activities. The amygdala and septum serve as telencephalic internodes for neural circuits relating the rostrally located olfactory apparatus: all together they form the rhinencephalon. III) The thalamocingulate division is comprised of the mesocortical cingulated areas receiving afferents from the anterior and other thalamic nuclei. It also comprises mammillary bodies of the hypothalamus. Differently from other parts of Limbic System, the thalamocingulate division seems to have no representation in the reptilian brain (MacLean 1990, p. 247). This division is concerned with three cardinal behavioral developments that, in addition to endothermy, characterize the evolutionary transition from ancient Reptiles to Mammals. Namely: 1) Nursing, in conjunction with maternal care; 2) audio-vocal communication for maintaining maternal-offspring contact; and 3) play (MacLean 1990, p. 380). Nursing is due to the action of oxytocin, the mammalian hypothalamic hormone that derives from the reptilian hormone mesotocin. This change may be considered the biochemical border marking the transition from the primordial Limbic System of Reptiles to the regular Limbic System of Mammals (Insel & Young 2000). Another important modification has affected the vagus nerve. Only Mammals have a myelinated, and therefore faster, vagus, so that heart activity may be quickly linked to the Limbic system’s emotions. According to the polyvagal theory (Porges 2001), in mammals, both neocortical and reptilian strucutres, but primarily Limbic ones, control the activity of the vagus and other cranial nerves that govern facial expressions and vocalizations in social interactions: such non verbal communications of unconscious emotions and conscious intentions constitute the bases of Primates’ social life. The Limbic System may be seen as a regulator of the R-complex. Most of this regulation seems to be inhibitory and acted via the serotonergic system. Laboratory stimulation of the Limbic System often produces a suppression of R-complex ongoing behaviors, whereas lesions made in it often seem to “release” various reptilian activities (adapted from Isaacson 1982, p. 246). This means that the mammalian R-complex is not exactly the same as the reptilian R-complex: in fact the mammalian one receives new inputs from limbic structures and it has also fit receptors (often different compared with reptilian ones) for neuromodulators or neurotransmitters that mediates these inputs. Furthermore, as early noted by MacLean (1970), alterations of the usual activities of the Limbic System, e. g. by epileptic episodes, can produce various experiences and feelings, as those associated with knowledge of fundamental truths, feeling of depersonalization, hallucinations, and paranoid feelings. Other observations about the action of the Limbic System on the R-complex will be shown in the subsequent section (§ III). The cerebral cortex is the most evident part of human encephalon, and it is divided in two hemispheres, which are connected by three fibrous structures: the corpus callosum, and the anterior and posterior commissures. Each hemisphere is constituted by numerous convex formations called gyri, divided each other by sulchi. In each hemisphere we may also note four regions called lobes: the frontal lobe comprises the upon ocular socket area and behind the forehead up to near the top of the brain, where the central sulcus divides it from the parietal lobe; the temporal lobe is located under frontal and parietal lobes and it covers a little lobe, the insular one; the occipital lobe forms the back of the brain (fig II. 5 a). In a side view of a cross section of the brain (fig. II. 5 b), we can see a large gyrus surrounding the corpus callosum: it is called cingulate gyrus and is constituted by mesocortex, which is intermediate between allo- and isocortex. The cingulate gyrus, in the basal brain, continues as piriform cortex (derived from the ancient lateral pallium). Inside this latter is a structure derived from the ancient medial pallium (archicortex), which is called hippocampus. Cingulate gyrus, piriform cortex and hippocampus constitute the limbic cortex. Cingulate and rhinal sulchi divide the cerebral limbic cortex from the Neocortex, that in apes and humans constitutes the major part of the cerebral hemispheres. The Neomammalian brain The Neomammalian brain consists of the Neocortex and structures of the brainstem with which it is connected, as lemnisci, pyramidal tracts, and neothalamus. The Neocortex is, on the human level, the seat of language and, in general, it is the seat of those behaviors that allow a person to tackle new and unexpected situations. The ability to foresee the future resides in it. We owe conscious thought to the Neocortex: it is the main seat of mind capabilities as self-consciousness or the connections of causality, as we’ll see in the following III §. The whole brain. As MacLean (1973a: 7; 1973b: 114) has made clear, the three basal brain types show differences both in structure (1) and chemistry (2), and so they are capable of functioning somewhat independently; but they are in no sense separate, autonomous entities: they must intermesh and function together as a triune brain. (1) As regards the structure, we have seen that in the R-complex neurons form masses (nuclei or ganglia); in the Limbic System besides nuclei there are pallial (or cortical) structures, in which neurons are disposed in three (allocortex) or more layers (mesocortex). Neocortex has neurons disposed in six layers (isocortex). (2) As regards the chemistry, researchers have found less marked differences: while, for example, acetylcholine is typical, but not exclusive, of the R-complex (MacLean 1990: 38-43), dopamine and serotonin are well allocated in all three brain types. Dopamine is synthesized by hydroxylation and subsequent decarboxylation of the amino acid tyrosine (that may also derive from phenylalanine). In Mammals there are four main dopaminergic pathways: (a) the nigro-striatal, (b) the infundibular, (c) the mesolimbic and (d) the mesocortical ones (fig II 5 d, modified from Kandel et al. 1996). They function respectively most in (a) tuning of sensori-motor programs, (b) regulation of hormone release in the pituitary gland, (c) modulation of sensory perception, maintaining reward values from life experiences and, thus, motivation up to addiction or aversion, (d) regulation of frontal cortex activity (Kandel et al. 1996). Dopamine reward system originates in the ventral tegmental area of the midbrain and projects to the nucleus accumbens of the ventral striatum. This system is termed “mesolimbic” by Kandel and colleagues, who don’t use the triune tripartition. In the triune brain conception, it is more properly a reptilian subsystem (so in fig. II. 5 d we have emphasized this pathway). Serotonin (or 5-Hydroxytryptamine or 5-HT) is a compound produced in some nervous centers starting from the amino acid tryptophan, and it exerts its effects by acting as a classic neurotransmitter or as a neuromodulator (these effects are due to postsynaptic receptors: 5-HT receptors are many and diversify serotonin’s effects in the various encephalic regions. See Baumgarten & Göthert 1997). Serotoninergic neurons are present in all animals that possess nervous systems, both Protostoma and Deuterostoma . In Vertebrates, nervous centers in which 5-HT is produced are raphe nuclei. In fig. II. 5 e the various mammalian raphe nuclei and their main projections are shown. According to ten Donkelaar (1998: 1417) “In reptiles, only two raphe nuclei can be distinguished: i. e. the rostral, mainly small-celled nucleus raphes superior and the large caudal nucleus raphes inferior containing medium-sized to very large cells”. The caudal part of the inferior raphes nucleus may correspond to the raphes pallidus of mammals, the rostral part to the mammalian nucleus raphes magnus. In the reptilian nucleus raphes superior may be hidden a primordium of the mammalian nucleus raphes dorsalis. It seems that the mammalian raphes obscurus has no homologue in the reptilian brain. Following MacLean’s model, in the serotonergic system we can distinguish a reptilian part (i.e. of the R-complex) and a mammalian part (i.e. of the Limbic System), the latter being formed in particular by raphes obscurus and dorsal raphes nuclei. Ascending fibers extend from the pontine nuclei to several telencephalic regions, among which are basal ganglia. So, the serotonergic system’s evolution is an example of the evolution of the brain according to MacLean’s model. In fact we may distinguish in it a reptilian part which is a regulator system inherent to the R-complex itself and to the spinal cord; and a limbic part which greatly increases control over the R-complex and moreover has connections with the Neocortex, in particular with prefrontal cortex which, according to MacLean (1978), helps us to see deeply into other peoples’ feelings. Given its numerous projections, the serotonergic system provides the most powerful system of integration and coordination among MacLean’s three brain types identified till now. § III. Brain and Mind, Modules and Operators. According to evolutionary psychologists, the human “mind is organized into modules or mental organs, each with a specialized design that makes it an expert in one arena of interaction with the world. The modules’ basic logic is specified by our genetic program. Their operation was shaped by natural selection to solve the problems of the hunting and gathering life led by our ancestors in most of our evolutionary history” (Pinker 1997, p. 21). We obviously agree with the main lines of evolutionary psychology, going over our hunter and gathering ancestors’ past: in the evolutionary landscape we have depicted we search the biological bases of god concepts in steps, first in Reptiles, ancient Mammals and finally Primates. Considering the global functioning of the Brain (i. e. the Mind), d'Aquili and Newberg (1999) have considered its primary functional components, which they have referred to as cognitive operators, which have specific functions that are localized in specific regions of the brain and perform activities that underlie the capacities of the mind. Newberg and d’Aquili (2001) have described eight cognitive operators: the holistic, the reductionist, the causal, the abstractive, the binary, the quantitative, the emotional, and the existential ones. As these authors noticed (2001, p. 187), the cognitive operator concept is clearly “similar to the concept of cognitive modules in that both are functions and are localizable to one or more specific areas of the brain.” … However “Cognitive operators differ from cognitive modules”, because cognitive modules represent more specific functions that are localized to particular brain structures, whereas cognitive operators refer to more generalized functions of many areas of the brain: modules are concerned more the anatomy of the brain, operators more the physiology of the brain (or of the encephalon). However brain functions also produce behaviors and unconscious knowledge. Let us extend the operator concept from cognitive ones to encephalic ones. Encephalic operators may be defined as behavioral, emotive or cognitive operators that are specific functions performed by specific parts of the encephalon. Proceeding along the evolutionary scheme of the triune brain, we may define, in the R-complex, some behavioral operators, such as: 1) the specific operator, that allows the acquisition of species identity by the imprinting (i. e. a fast form of learning that occurs only during a critical period in the development of the organism); 2) the sexual operator (for male or female distinct behaviors): its primary component may be located in the hypothalamic medial preoptic nucleus, which produces gonadotropine releasing factors, that has different size in male and female individuals, and that is “crucially involved in the consummatory phase of masculine copulatory behavior” … and “also plays a prominent role in feminine sexual behavior” (Voogd et al. 1998, p. 1871); another behavioral important component of this operator lies in the nucleus accumbens. 3) The territorial operator: animals mark their territory with smells or other signals of boundaries, warning other (conspecific) animals not to intrude; 4) the isopraxic (or mimetic) operator: it allows the animals to act in a like manner and it is “implicated in conspecific recognition and in most forms of communication involved in self-preservation and in the procreation of the species” (MacLean 1990, p. 144) and, in the opposite sense, it serves to promote species isolation; it is evident when conspecific animals behave in the same way at the same time (group or mass isopraxis); 5) the hierarchic operator, whose primary component most likely lies in the pallidum, and that is fundamental for engaging in various types of display. R- complex may be also produce cognitive operators such as: 6) the space operator, that allows an animal to know its territory; 7) the time operator, that permits the chronology of events; 8) the sequence operator: from the interaction of the former two operators, an animal can know that after an event a specific one follows or may follow; 9) the semiotic operator: it allows the right understanding, often species-specific, of behaviors and non verbal communications between individuals inside the same species (and to some extent, interaction with different species). It moreover permits a symbolic codification of some behavior (for example, in Primates, the penis display made by a male towards another male is not a sexual presentation, but a challenge display), and let the animal receiving the message to decode and correctly interpret the behavior it sees (e.g., the male to whom penis display is directed must understand it as a challenge display and not as a sexual presentation). Summarizing, the semiotic operator allows the correct coordination of behavior operators’ actions listed above. The main site of these cognitive operators seems to be the dorsal striatum (Parent and Hazrati 1995). The Limbic System may be considered the main site of several emotional operators, as the follows: 10) the fear operator. It allows graded behavioral outputs in regard to safe, unsafe and life threatening environments. Its main nervous center is the amygdale, which acts in connection with the cortex, the hypothalamus, the parasympathetic (mainly the vagus nerve) and the sympathetic systems. Porges (2001) has fully discussed the connections among these nervous and endocrine organs. When sensations received by sensory cortex are judged by the amygdale as caused by a safe environment, the amygdale stimulates vagus action both directly and by hypothalamic oxytocin, which is released to the sensory and motor portions of the vagal complex and systemically to visceral organs. Oxytocin fosters a calm or “anti-stress” state, and the sympathetic nervous system is not stimulated. When the amygdale judges the environment as unsafe, it stimulates the hypothalamus to release the hormone vasopressin: the vagus is inhibited, the sympathetic system stimulated, and the animal is ready for fight or flight behaviors. A life-threatening environment may elicit immobilization (i.e., feigning death or passive avoidance) and fight-flight behaviors are inhibited. According to Porges’ polyvagal theory, the neural regulation of the autonomic nervous system is the result of three evolutionary stages, each with a connected behavioral strategy. The first stage is characterized by immobilization behaviors due to the action of a primitive unmyelinated vagus that responds to threat by depressing metabolic activity (and this causes immobilization). The second stage is characterized by the sympathetic nervous system that is capable of increasing metabolic outputs and inhibiting the visceral vagus to foster mobilization behaviors necessary for fight or flight. These two stages of the “fear operator” characterize the reptilian limbic system. The third stage, unique to mammals, is characterized by a myelinated vagus that can rapidly regulate cardiac output to foster engagement and disengagement with the environment. As a rule, mammals are active in the second and third stage of this evolutionary operator, but in threat environment may regress to the first stage, and subsequent immobilization. 11) The aggression operator: it seems that we may distinguish various aggression operators, as, at first, between predatory or interspecific aggression and the intraspecific one. The last is linked to the hierarchic operator of the R-complex and has its limbic component mainly in the amygdala. 12) Also the mimic judgment operator has its focus in the amygdalar complex. It is very important for those mammals that are able to change their face expression, as mainly primates do. Face expressions are linked to the Autonomic Nervous System, as Porges (2001, p. 123) has remarked: “The neomammalian vagus is neuroanatomically linked to the cranial nerves that regulate social engagement via facial expression and vocalization”. A primordium of the mimic judgment operator also exists in the brain of vertebrates that are each other stimulated by visual signals. In reptiles and birds emotional or intentional (as aggressive, sexual and other) signals are not given by facial muscles’ movements, but by skin color changes, ruffling up of feathers, shows of cutaneous appendixes (i. e. skin capes or fans as in the genus Amphibolurus). Humans employ plumages, mantles, headpieces and other various outfits to show their rank or their well or ill disposition towards other humans: the human R-complex unconsciously and immediately decodes and interprets these signals. 13) The “falling in love” operator allows pair bonds establishing. The neurobiological bases of this behavior have been studied only recently: Pair bonds are not very frequent in mammals, but, in the species in which they exist, seem to be linked to the activity of the rhinencepha-lon, and to the each other smell recognition of the individuals. It seems that in the human pair bond instauration there is the involvement both of the serotonergic (Marazziti et al. 1999) and dopamine (Aron et al. 2005) systems. The initial serotonin tone deficiency (observed by Marazziti et al.) may have the role of paving the way to the reward function of the subsequent dopaminergic action (observed by Aron et al.). 14) The call operator activity is triggered when the pup feels to be neglected by its parents and in turn triggers in them: 15) the nursing operator activity, that, as we have said above, depends on the oxytocin release. The call and nursing operators are not in action only, or towards, infancy: in some emotional circumstances they are active even and towards adults (all the more reason it happens in Homo sapiens that is a neothenic species, in which infancy features and behaviors persist or could reappear along the life). 16) The attachment operator allows the establishment of a peculiar bond between pups and parents. It does not operate from birth in all the species, and this makes possible the adoption of a forsake (or an orphan) pup by other adults. Its action becomes evident when the pup is able to distinguish one another its neighbors, preferring some of them and fearing others. 17) the play operator allows pups to learn their specific behaviors according to the various circumstances. So behaviors of Mammals are less fixed and stereotyped than behaviors of most Reptiles and Birds. (Ethologists have observed play-like behaviors in some birds). Acquired behaviors and knowledge are, as a rule, proportional to the playing age period, which varies among the species, and within a species, among individuals (in the human species, experimental scientists may be considered boys that are playing despite their adult feature). The Limbic System may be considered the primarily seat of a cognitive operator, the existential one. 18) The existential (or ontological) operator assigns a sense of existence or non existence to the sensory information processed by the brain. It likely gives also the sense of existence to the thoughts elaborated by other parts of the encephalon, such as the R-complex or the Neocortex. In other terms, this operator gives a sense of reality to beliefs, regardless of whether they might be non contradictory or contradictory, or counterintuitive, according to Neocortical operators. Emotional and existential operators may be assumed as generally functioning in Mammals. In humans, the existential operator, that is cognitive but not necessarily rational, is linked to limbic emotional operators. MacLean wrote (1973 b, p. 123) concerning the feelings associated with knowledge of fundamental truths: “It seems that the ancient limbic system provides the ingredients for the strong affective feeling or conviction that we attach to our beliefs, regardless of whether they are true or false!” The Neocortex is the main site of most cognitive operators. The following list summarizes data from d’Aquili and Newberg (1999) and Newberg et al. (2001) for human mind operators, from the holistic to the quantitative one. To these we add the linguistic, the insight, and the metacognition operators. 19) the holistic operator enables the organism to view reality as a whole (gestalt). It likely rises from the activity of the parietal lobe in the non dominant hemisphere (i. e., in most humans, the right one); 20) the reductionist operator allows the mind to see the whole broken down into its component parts; it functions in the opposite manner to that of the holistic operator, and it resides primarily in the dominant parietal lobe (i. e., in most humans, the left one), that so is involved in reductionist and analytical processes, while the non dominant parietal lobe is involved in holistic and synthetic ones. The corpus callosum allows connection between the two parietal lobes, permitting the mind to combine the holistic and the reductionistic approaches. 21) The causal operator enables the mind to interpret all of reality as a sequence of specific causes and effects. It is believed to result from the connections between the left frontal lobe and the left orientation association area; it spontaneously and actively tends to impart a sense of causality on all the events. This function of the mind is genetically hardwired in all humans, and may be called as the causal imperative, or, according to Levy-Bruhl, the need of explanation. 22) The abstractive operator permits the organism to form general concepts. These abstract concepts are constructed from the perception of individual facts and various individual objects, derived from inductive functions. These operators likely reside in the inferior portion of the parietal lobe. V. Ramachandran has shown experimental evidence that the angular gyrus, belonging to the inferior parietal lobule, plays a major role in metaphoric understanding. 23) The binary operator organizes reality by reducing the most complicated relationships to simple pairs of opposites. According to d'Aquili and Newberg it resides in the inferior parietal lobe on the dominant side. However, as its activity appears as a stereotyped one, this operator may have its unconscious origin in the R-complex, likely in the caudate nucleus, which has numerous links with neocortical association areas, so it may become conscious in the parietal lobe. 24) The quantitative operator permits the abstraction of quantity from the perception of various elements, and allows mathematical operations but also the estimation of time and distance, the awareness of amounts of things as food or enemies approaching, and the need to order objects or sequences of events by some numerical system. It likely resides in the general area of the inferior parietal lobe. 25) The linguistic operator has two neocortical components: the Broca’s area in the frontal lobe and the Wernicke’s area in the superior temporal gyrus, near the inferior parietal lobule. The former area permits to speak correctly, the latter allows understanding of the meanings of words or sentences that have been heard. We may note that this area is contiguous with abstractive (and metaphoric) area. The linguistic operator may be considered the cortical correspondent and extension of the reptilian semiotic operator. 26) The insight operator let us to succeed in problem solving in a sudden way, after and by an unconscious elaboration and not by attempts and errors: the solution reach the consciousness suddenly, often while the subject is thinking or doing other matters. Remembering the exclamation shouted by Archimedes when the solution of a problem suddenly rose to his consciousness while he was in the bath, this operator may be termed “eureka” (I found!) operator. 27) The metacognition (or empathy) operator permits the awareness of one’s own mental states and relates behaviors to social or environmental circumstances; it allows the development of a “theory of mind”, i. e. an awareness of other people’s minds’ content. It likely resides in frontal lobes, mostly in the right hemisphere (Stuss et al. 2001), and “mirror neurons” (Gallese 2005) may contribute to this operator. “Theory of mind”, “metarepresentation”, “metacognition”, “mind reading” and “mental state attribution” are terms that refer to the “awareness of one’s own mental states, beliefs, attitude and experiences, the relationship between these and external events, and also of the mental states of others and the implications for their motives and intentions”, as summarized by Stuss et al. (2001, p. 279). They note that only humans and a few species of great apes are capable of attributing mental states to others, making us, as Kirkpatrick noted, “all amateur psychologists” (2005, p. 274). We may assume that some of these cognitive operators are functioning in Mammals based on both to neocortical development and to complexity of connections among cortical centers. The holistic, reductionist, abstractive, quantitative, insight and ethical operators may be assumed working in a variable way in the different Orders of Mammalia, presumably more in Proboscidea (elephants), Cetacea (dolphins) and Primates (apes). The binary operator seems to be typically human, as well as the quantitative operator and its connections with other operators (as the linguistic and abstractive ones) to allow symbolic mathematics rising (but see Hauser et al. 2002 with regard to animal cognitive abilities and quantitative and linguistic functions). The causal operator, as it is endowed with an high abstractive ability, is produced thanks to human neocortical complexity: on the grounds of our knowledge, it seems to result from the inclusion and development of a mammalian neocortical operator (which has more or less abilities according to the various species), and that may be termed as “agency detection operator”: it attributes an intentionality as the concrete causes of phenomena. A first description of an animal behavior that we may refer to this operator has been reported by Darwin (1871, p. 67): “my dog … was lying on the lawn during a hot and still day; but at a little distance a slight breeze occasionally moved an open parasol … As it was, every time … the dog growled fiercely and barked. He must, I think, have reasoned to himself in a rapid and unconscious manner that movement without any apparent cause indicate the presence of some strange living agent, and no stranger had a right to be in his territory”. Agency operator evolved in Mammals, by means of natural selection, not only for intruder detection, but also for predator detection: Barrett (2000) has termed this brain function as agent detection device. As exposed by Boyer for humans (2001, p. 145), “Our evolutionary heritage is that of organisms that must deal with both predators and prey. In either situation, it is far more advantageous to overdetect agency than to underdetect it. The expense of false positive (seeing agents where there are none) is minimal, if we can abandon these misguided intuitions quickly. In contrast, the cost of not detecting agents when they are actually around (either predator or prey) could be very high.” Thanks to this over-developed neocortex, Homo sapiens can be said to possess a developed agency detection operator, termed by J. Barrett (2000) as Hyperactive Agent-Detection Device (HADD), extended also in a more abstract form, in the causal operator. In Reptiles and Birds, R-complex has the ability of recording temporal sequences of events. Such sequences in order of time do the function of sequences in order of cause: this leads to routine activities, in conformity with precedent situations, ritualistic behaviors or associations between events and behaviors that may involve both Central and Peripheral Nervous Systems. Pavlov's conditioning experiments have shown that an animal may unconsciously and automatically put in connection stimuli and answers that normally have no link. Natural selection has favored survival and reproduction of those animals that, provided with a rudimentary forebrain, succeeded in linking and recording events that happened in a temporal sequence. Individual provided with such an ability had more fitness because they could anticipate favorable events, repeat positive and avoid negative experiences. Indeed, if it is true that not all the events that happen in a temporal order are also linked by a casual order, it also true that all the events that have a causal connection have also a temporal sequence: temporal relations are redundant in respect to causal ones, but for animal surviving it is better overloading than lacking. This “pseudo causal” association made by the R-complex is the fundamental, and almost the only, link among events Reptiles and Birds may “think”. It is basic also for Mammals, but they can elaborate sensorial or recorded data by means of the Neocortex. A mammal, in comparison with a reptile, more easily forgets a connection if it does not happen afterwards: this ability is linked to brain complexity. In Mammals, the R-complex is connected with the Neocortex in a more complicated way than in Reptiles and Birds. In Primates, as shown by Parent et al. (1995, p. 135), inputs from the frontal lobe (motor cortex) terminate mostly in the lateral putamen to constitute the sensorimotor striatal region (SMSR in fig. III. 1). Inputs from the hippocampal and parahippocampal gyri, the cingulated cortex and the amygdala, terminate in the nucleus accumbens and part of the olfactory tubercle to constitute the limbic striatal region (LSR). Inputs from the prefrontal cortex as well as from associative areas of the parietal and temporal lobes terminate in the head of the caudate nucleus and the rostral part of the putamen to constitute the associative striatal region (ASR). So in the mammalian R-complex we may locate at least four components: 1) a behavioral one, made up by globus pallidus and nucleus accumbens, that is the main seat of behavioral operators as the specific, sexual, territorial, isopraxic, and hierarchic ones; 2) a senso-motor component (which shows its weight in the Parkinson’s disease); 3) a limbic component and, 4), an associative one, as above described. In Mammals the R-complex maintains the ability of fixing pseudo causal connections among phenomena (post hoc, propter hoc). In the associative component of the mammal R-complex may occur both the link among real events and the association, mostly unconsciously, of ideas, memories or emotions originated in other brain areas such as the Limbic System or the Neocortex. Particularly in humans, the associative striatum may link not only real events, but also events “imagined” by Neocortex, Moreover, the pseudo causal sequence may be reversed: not only from the “cause” to the “effect”, but “given an event” then “connect it with a cause”. These “free associations”, unconscious and reasonless, made by humans in ways that are structural similarly, but whose contents are various in each being, had been discovered by psychoanalysis long time before their cerebral origin had been located. The thought processed by the reverberating circuit Neocortex – R-complex – Neocortex – etc., is called “magical thought” and it may be expressed by language. Magical thoughts, as “that events which have been observed to occur simultaneously or to follow a particular sequence will continue to follow the same pattern” or “that like produces like” (Vernon 1962, p. 65), may lead the development of ritualistic actions, also up to obsessive and compulsive behaviors. The Limbic System, particularly its serotonergic component, aims to inhibit the magic and ritualized activity the associative striatum tends spontaneously to generate. But the Limbic System, if it fails in doing so, then adds an emotive component to ritualistic or compulsive behaviors, producing a released condition when they are completed, and an anxious condition when they are brusquely interrupted or changed. The Neocortex, as we have seen, has “induction” and “causal” operators, so that from the particular observations that different phenomena are effects of certain causes, it infers that “every phenomenon is the effect of a cause”. The abstractive operator lies in the inferior portion of the parietal lobe, and the causal operator results from the connection of the left association area /in the parietal lobe) with the frontal lobe: the parietal part of the causal operator is close to the abstractive operator, and it seems that, for its activity, the causal operator utilizes previous or almost contemporary investigations done by the abstractive operator. Therefore, the Neocortex, by means of its cognitive operators, allows humans some rational way of interpreting, or explaining, the reality: it is able to do this more or less well, according to its knowledge. Summarizing, the link among phenomena elaborated by the whole brain may be : a) a rational and verifiable one, if the Neocortex can find the cause-effect nexus; b)a magic one, that is not verifiable, that is characterized by more or less high levels of fancy, and that is held by faith. Magical thought has its unconscious origin in the R-complex, and is made conscious by the Neocortex, which tries to give it an appearance of rationality. § IV. Dominance-Submission Behavior in Religion, and the Principles of Uniformatism and of Correlation. In 1821 F. Schleiermacher wrote that “religion is the feeling of sheer dependence on God” (quoted by Burkert, 1996, p. 80), who afterwards adds: “religion is generally accepted as a system of rank, implying dependence, subordination and submission to unseen superiors. The awareness of rank and dependence in religion is particularly clear in all the ancient religions. God means power, rule, and honors due.” (p. 81). As noted by Morris (1967; 1994 p. 121) “... in a behavioral sense, religious activities consist of the coming together of large groups of people to perform repeated and prolonged submissive displays to appease a dominant individual. The dominant individual concerned takes many forms in different cultures, but always has the common factor of immense power.” This notion of more powerful beings who determine the fate of humans is a widespread tendency, if not the nucleus of religion. Utilizing data from sciences as different as Neurobiology, Paleontology, and Compared Ethology, we apply the Principle of Uniformitarianism, that, in its broader form, affirms natural laws we observe in the present world are the same that operated in the past and the same will operate in the future. In general terms, natural laws are invariable both in space and in time. We seek to therefore understand the source of our “Supreme Power Being” in the evolutionary pre-history of dominance and submission relations in the animal kingdom. The Principle of Correlation proceeds from the principle of uniformitarianism: if particular phenomena, or peculiar features, of living beings regards their environment, or regards themselves, are in correlation in the present world, we may infer that they were in correlation also in the past. It implies that the knowledge of some documented by fossils features allows us to infer the presence, in those beings of the past, of other non fossil, but with the previous ones correlated, features. However, the correlation principle must be used with caution, as two phenomena or two features may show themselves together by chance. So we can correctly utilize the correlation principle either when we know the causal relation which determines the correlation, or when the statistical analysis allows us to exclude a more chance correlation. The hypothesis that religious behavior, religious thought and the origin of the “Powerful Being” idea are natural may be substantiated by evidence of the roots of such behavior among Hominids and non-human Primates. For confirmation of the validity of this hypothesis we find the existence of a dominant “chief figure,” potential or in action, to whom others show submission behavior. § V. Dominance-Submission Behavior in Primates In Primates strong links have been established between sexual organ or behaviors and territoriality and dominance-submission behaviors, so that male genitals and sexual behavior signify dominance on a territory or on a social group, while female genitals and behavior become symbols of subordination. Wickler (1967) has described the so called sentinel behavior in baboons: one, or more, male dominant individuals sit at lookout sites thighs spread and a display of partial erection. This display is regarded as an optical marker of boundaries, warning other monkeys not to intrude (fig. V. 1 a). The male squirrel monkey, seeing another male or its own image on a mirror, “flexes the head to one side, retracts the corners of the mouth while making high-pitched, peeping vocalizations, spread one or both thighs, and makes thrusting movements with the fully erect phallus” (MacLean 1990, p. 171) (fig. V. 1 b). The rostral two-thirds of the medial segment of the globus pallidus seems to be the main site involved in mirror display and therefore in aggressive intraspecific behavior, or, in other words, this part of the pallidum seems to be the main site of the hierarchic operator. Such valences of sexual behavior and genitals are also evident in Homo sapiens. MacLean (1990, p. 233) refers that Gajdusek (1970) “has called attention to the parallel between the display behavior of squirrel monkeys and certain rituals of Melanesian tribes”, or that some New Guinea groups when frightened, excited or surprised meet the event by a penile display dance. This dance is similarly used to express both dominance and aggression. At the present time the rugby team All Blacks displays the Maori haka dance before any game. In many cultures in the world, house guards (stone or wooden monuments showing an erect phallus) have been used to mark territorial boundaries (fig. V. 1 c). In ancient Athens stylized statues of Hermes, endowed with an erect phallus, were posed at street corners. (Fig. V. 1 d). The erect phallus, symbolized by some means as, for example, the middle finger, is an human challenge display utilized by male or female individuals (fig. V. 1 f). For more than two centuries, Occidental men have found a discrete form of phallic display: the neck-tie. Perhaps it is superfluous as a masculine signal, since bearded men more easily give up tie in habitual dressing. Returning to Primates, the mounting threat is a form of rank demonstration, and it can be performed both by a male towards another male (fig. V. 2 b) and by a female towards a lower rank female (fig V. 2 c). Female sexual presentation can be used by low rank individuals in order to acquire group chief’s support (fig V. 2 d). Primate female sexual presentation may be observed in some human profane (fig. V. 2 e) or religious (fig. V. 2 f) submissive gestures. Sometimes female sexual presentations are utilized as challenge displays: doing so, the displayer means that he/she considers his/her opponent as impotent. § VI Primate social behaviors and sexual dimorphism. Primate societies may be grouped according to breeding systems as follows: a) Monogamous breeding systems (Mo); b) Single male breeding systems (SM); c) Multi-male breeding systems (MM). Several scholars (as Clutton-Brock and Harvey 1977; Leutenegger and Kelly 1977; Harvey et al. 1978; Pickford and Chiarelli 1986; Hinde 1987) have attempted to correlate these systems with male/female sexual dimorphism in body size and in canine tooth size. These studies have shown that, at first view, monogamous species show a very low dimorphism, while single-male and multi-male species show a variable degree: as Buss (1999, p. 286) has summarized these studies “the greater the effective polygyny the more pronounced the sexual dimorphism”. However, results from in-dept studies have shown less linearity: after a long series of studies (as Plavcan et al 1992, 1997), Plavcan (2001) has made clear that sexual dimorphism is more related to malemale competition levels than to breeding systems. Plavcan and van Schaik (1992) classified primate species into four “competition levels”, labeled from 1 to 4, on the basis of the “intensity” and “potential frequency” of male-male competition. Competition level 1 contains low-intensity, low-frequency species. Competition level 2 contains low-intensity, high-frequency species. Competition level 3 contains high-intensity, low-frequency species. Competition level 4 contains high-intensity, high-frequency species. Low-frequency competition is typical of breeding groups that contain only a single, adult male: so monogamous species belong to level 1, while single-male species belong to level 3. But species in which single male breeding groups come together to form large groups daily, such as Papio hamadryas, Theropithecus gelada, and Nasalis larvatus, belong to level 4 because they are high frequency species. Multi-male species mainly belong to competition levels 2 and 4. Primates’ breeding and social systems, some representative species and their competition levels, as estimated by Plavcan and van Schaik (1997, p. 347-348), are shown in table VI. 1. § VII. Dominance-Submission Behavior in Hominin Phylogeny. Homo sapiens is an African ape, belonging to the order of Primates, infraorder Catarrhinae, that is now formed by two superfamiliae, Cercopithecoidea and Hominoidea. Evolution of Catarrhinae occurred in Africa and Eurasia. The first family of catarrhins was Propliothecidae, whose main genera were Propliopithecus and Aegyptopithecus. This latter one, a cat size monkey, tail having and showing an evident canine size sexual dimorphism, is considered as the ancestor of both Cercopithecoidea and Hominoidea, the first ones having a tail and quadrupedal walk, the second ones tail absence and the possibility of braching gait. Proconsul, Dryopithecus, Oreopithecus, and Ouranopithecus are the main genera of Miocene fossil Hominoidea. The 8 Myr ago common ancestor of humans and chimpanzees “was chimplike forest dwelling and predominantly arboreal and fruit-eating” (Wood & Brooks 1999, p. 219). The tectonic fracture series forming the African Rift Valley system caused, about 86 Million years ago, a geographic barrier that divided Hominoids apes in two distinct populations, and the differentiation of the eastern environment respect the western one. This latter remained, and still does, an equatorial forest, while in the east side, owing to a growing dry environment, as a result of disturbances in atmospheric circulations caused by the Rift Valley, disperse woods succeeded in the first millennia, and then savannah developed. The eastern (future Hominini) and the western (future Panini) populations of Hominids were so each other isolated and became subject to increasingly different selective pressure, as Hominins faced at first disperse woods and subsequently the savanna, while Panins remained in equatorial forest, where they differed into two species, Pan throglodytes and P. paniscus. The most ancient Hominini so far discovered, and dated back to 5.8 – 5.2 Mya, have been assigned to the species Ardipithecus kadabba (Haile-Selassie et al. 2004). Fossils happen chimplike and the environment in which they lived was still wood. The most likely competition level attributable to this hominin species is the second: A. kadabba most probably lived in multi male social and breeding groups, as present Pan species do. In subsequent Hominini, such as Ardipithecus ramidus (4.5 Mya), Australopithecus anamensis (4.2 – 4 Mya), and Australopithecus afarensis (4 – 2.5 Mya), we may note improvements in bipedalism and increase in sexual dimorphism. A. afarensis lived mostly in savanna. Primate models of ancient hominin behaviors. Reconstructing hominin evolution and social behavior, some scholars (as De Waal 2000), privileging genetic proximity, have chosen chimps as a species model; others, giving more consideration to environmental influences, have chosen baboon species. Both of these approaches have their strengths, but neither alone is a complete model for Hominins, who must be studied as Primates that, genetically and morphologically chimp-like, underwent an environmental selective pressure that might give them behavioral baboon-like traits, as we know that similar ecological pressures produce similar adaptations even in genetically different animals (convergent evolution) (e.g., see C. Jolly 2001). As summarized by McFarland (1985, pp. 159-60), comparative studies have shown that in harsh environments baboon troops are dominated by a single male, whereas in rich environments the troops are multi-male. Papio hamadryas and Theropithecus gelada are typically SM, while Papio cynocephalus lives in rich environments and has typical multi male troops. Papio anubis and P. ursinus (Chackma baboons) live in intermediate environments. They have multi-male troops and dominant males have privileged access to females. Often some dominant males form a “central hierarchy” and cooperate in maintaining their social position against rivals, in protecting mothers and infants and in defense against predators. Male anubis baboons have a smaller but evident cape, and show a marked sexual dimorphism in body and canine teeth sizes. Each male hamadryas baboon that possess an harem of females does not try to mate with a female of another harem, while male anubis baboons try. From this and other behavioral differences, it follows that “hamadryas males exhibit affiliative and mutually supportive behaviors in a variety of social contexts in which anubis males tend to be indifferent or hostile to each other” (Kaplan et al. 1999, p. 518). The neurobiological basis of these different behaviors mostly lie in the serotonergic system’s activity, that is lower in anubis than in hamadryas males, as shown by observations carried out in anubis and anubis-hamadryas hybrid males (Kaplan et al. 1999). It follows that, within the fourth level, hamadryas baboons are less competitive than anubis baboons. As stated by Plavcan and van Schalk, “increasing sexual dimorphism in both canine tooth size and body weight is strongly correlated with increasing competition level” (1997, p. 353). Such correlation allows us to assess competition level from sexual dimorphism, and vice-versa: we may control this statement in extant species and infer it for fossil species. As it lived in harsh environment, we may reasonable suppose that A. afarensis, which showed a marked body size sexual dimorphism, underwent a selective pressure that gave baboon-like its behavior and social organization. According to the correlation principle, we may chose Papio hamadryas or Theropithecus gelada as species that better may give us a model of A. afarensis male behavior (high male – male competition) and social/breeding organization (SM breeding groups that daily come together to form large groups). So the most likely competition level attributable to A. afarensis, and to the precedent species A. anamensis, which appears to have been very dimorphic too (Ward et a. 2001), is the fourth one. As noticed by Flinn et al. (2005, p. 26), “Australopithecus body mass dimorphism suggests that these early hominins were polygynous, as significant mass dimorphism is not associated with monogamy in any extant primate (Plavcan 2001)”. So, we may likely suppose that A. afarensis had an SM breeding system and an MM social system. Sexual selection in hamadryas and gelada baboons favors males with large hair capes: in harsh environment, males that seem to be bigger are more fit than males actually larger (that are more expensive in nutrition and easilier victims for predators). According to an hypothesis that has had Lovejoy among its first author (1980), it might have been that, in a hominin SM breeding system group (likely belonging to A. afarensis), it were born by chance a female having a concealed ovulation (i. e. without estrus or sexual swellings and perhaps with a little masculine appearance, having a little hair cape). In this case, the dominant male might have no close watch on her, and she could more easily mate with a subordinate, and so less dimorphic, male. From such mates it could be born both sons having little dimorphism and daughters having concealed ovulation and probably a continuous sexual receptiveness. These modifications very probably gave origin to a new species: Australopithecus garhi is the fossil hominin most likely depict it. Male individuals of such a new species, being less dimorphic and less competitive each other, could easily co-operate in defense against predators and for food gaining. Moreover, if A. afarensis males behaved as Papio hamadryas males do at present, they might exhibit supportive behaviors in various social contexts: this condition might be a pre-adaptive feature for more supportive behaviors in A. garhi. It seems that A. garhi was gatherer and scavenger, rather than hunter, because its body was small and its legs were short. Fossils of the subsequent genus Homo are characterized by low body size dimorphism (competition levels 1 or 2), and absence of canine tooth size dimorphism, which may imply breeding and social systems unique among primates (Ruff 2002). Concealed ovulation and female continuous sexual receptivity formed two behavioral selective factors because of males that insisted in mating with the same female gained more reproductive success compared with males that tried to mate with all the females, risking impregnating no one. Such a situation lead to form some adaptations supporting long-term mating relationships and pair bonds, towards the success of monogamy in a primate species in which, owing to its basic genetic inheritance, both males and females were polygamous. Because of this complex interactions between “towards monogamy adaptations” and “basic polygamous inheritance”, we have chosen to term “pseudoMo” the human breeding system from early Homo to hunter-gatherer Homo sapiens. Subsequently, depending on environmental and cultural circumstances, Homo sapiens has produced various kind of marital systems: in some there has been a reappearance, at least in part, of an SM breeding system, with lawful or tolerated polygyny. As regards environmental adaptations, according to Bramble and Lieberman (2004, p. 345), “endurance running, defined as running many kilometers over extended time periods using aerobic metabolism ... is unique to humans among primates”. Fossil data suggest that this capability evolved in Homo habilis/rudolfensis for scavenging, and subsequently in Homo ergaster/erectus for hunting. Endurance running, as remarked by Bramble and Lieberman (2004, p.351), “may have made possible a diet rich in fats and proteins thought to account for the unique human combination of large bodies, small guts, big brains and small teeth” (see also Fialkowski 1986, 1987; Aiello & Wheleer 1995). Fig VII. 1 shows fossil species, which are thought to be inside the human phylogenetic tree, disposed according to their estimates of male-male competition levels. Scheme VII. 1 correlates the human evolution with breeding systems. As regards social organization, combining data from body dimorphism, ecological environment and stone tools, there is a general consensus that humans lived as hunter gatherers until a few thousands of years ago (that is, until the rising of agriculture, in some regions before, in other after, in other never), and that these social groups were egalitarian as regards the distribution of resources and reproduction, that is they had MM social system and pseudo-Mo breeding system. § VIII. The Immense Power Being as a projection of primate’s group chief: Freud’s and Morris’ hypotheses and criticism. Since religious displays are characterized by submission to a dominant individual, they may be the expression of an SM social system which has its neurological background in the hierarchy operator of the R-complex, as described in § III. On the other hand, according to evolutionary psychology principles, a dominant individual may not be conceptualized by a brain which is not prepared to conceive individual relations as hierarchic relations. Therefore, the “Immense Power Being” concept must have been the result of the projection of the image of a group’s dominant individual in a superhuman world: an example of such a model concerning the origin of religion is found in Freud’s Totem und Taboo (1912/13). In this work, which aims to explain the origins of totemism and exogamy, rather than religion, Freud utilized the concept of the “primitive horde” hypothesized by Darwin in The Descent of Man (1871), and the concept of the sacramental sacrifice of the totemic animal described by W. R. Smith in Lectures on the Religion of Semites (1889). According to Freud, in the “primitive horde” the father forbade his sons to mate with their sisters, that he kept to himself: one day, the excluded brothers came together, and killed and devoured their father, putting an end to the paternal horde. Eating him, they fulfilled a sort of identification with him, and each of them took possession of a part of his power. But afterward, as Freud hypothesized inferring by means of psychoanalysis, they felt remorse for their action, and, as dead, the father became more powerful than he was as live: his sons, owing to the “posthumous obedience”, inferred from psychoanalysis, prohibited the killing of father’s substitute, the totem, and mating with their sisters (from which the origin of exogamy). In order to explain the persistence of totemism and exogamy in subsequent human societies, Freud assumed memories, that are acquired and individual characters, as genetically hereditary characters, while geneticists were showing acquired characters were not. For that reason, now we may consider Freud’s tale only as a myth regarding the passage from SM hominin societies to pseudo-Mo ones. In The Naked Ape (1967; 1994, p. 121-122), D. Morris has proposed a sort of biological updating of the Freudian hypothesis. He has wrote that before human ancestors evolved into cooperative hunters they lived in social groups dominated by a single male. But “with the growth of the cooperative spirit so vital for successful group hunting, the application of the dominant individual’s authority had to be severely limited if he was to retain the active, as opposed to passive, loyalty of the other group members. They had to want to help him instead of simply fear him. He had to become more “one of them”. The old-style monkey tyrant had to go, and in his place there arose a more tolerant, more cooperative naked ape leader. … This change … nevertheless left a gap. From our ancient background there remained a need for an all-powerful figure who could keep the group under control, and the vacancy was filled by the invention of a god. The influence of the invented god-figure could then operate as a force additional to the now more restricted influence of the group leader”. We are in agreement with Morris about the basic point that the “all-powerful being” is a projection of the ancient “old-style monkey tyrant”, but Morris’ fast reconstruction presents some points that need explanations, so that it must be regarded as a further research stimulating model rather than a complete theory. Let us discuss two of this points: 1) Morris does not state precisely the period of the human evolution in which the “old-style tyrant” was eliminated and the god-figure was invented: in his reconstruction these two events are regarded as or almost contemporary. But: a) we may reasonably assert that the ability of thinking abstract, superhuman and invisible beings was achieved only by hominins endowed with a large brain, presumably after Homo sapiens speciation. b) We know, from the reconstruction of hominin social behavior based on fossil data dimorphism, and above reported, that the “old-style tyrant” was replaced by more cooperative leaders about two million of years ago. Therefore, if Morris’ postulated “invention of god” happened after Homo sapiens speciation, his reconstruction does not hold in due consideration the very long time passed from the “old-style tyrant” cessation; vice versa, if it happened immediately after this cessation, it cannot be considered reasonable because the brains of two millions years ago hominins could not think superhuman and invisible beings. 2) A “gap” is not a stimulus, but the absence of a stimulus. It is necessary a stimulus that, as it is inexplicable, suggests a “gap” of power that must be filled. Returning to our reconstruction, from the evolution of social behavior we have depicted, it logically descents that, in groups of lacking dimorphism Hominini (especially from early Homo), there were no individual who, as dominant male in SM social groups, might act as a concrete example of an “Immense Power Being”. In other words we can reject the possibility that the divine being idea might have been originated by a projection of a dominant male actually living in those groups. How did it happen, therefore, that among human beings with a documented low male-male competition social system we can find a projection in the super-human world of a being with immense power, that should have to be associated with a high malemale competition social system, supported by a strong hierarchy operator’s activity? Although inhibited by the Limbic System, the human brain R-complex preserved (and still preserves) structures and hierarchy forming functions which give rise to conceive powerful leaders. The neocortical elaboration of the concept of an “Immense Powerful Being” can only be created, according to the triune brain model, after a proposal of the R-complex to the Neocortex. Consequently we can deduce that at some time in the course of human evolution such a nervous structure was set free by the inhibitory action of the Limbic System, presumably as a consequence of a strong external stimulus which, causing a psychic trauma, weakened the inhibitory action of the Limbic System. § IX. Towards a Neurobiological and Evolutionary Psychology Explanation: a) The Traumatic and Triggering Stimulus. We could presume that this traumatic stimulus consisted in the acquisition of the awareness of own mortality. Indeed, as the human brain, presumably in Homo sapiens, achieved the complexity that allowed it to think over it happened in surrounding environment, human beings realized that whereas animals died because another animal killed them, human beings could die even if no animal killed them: For example, at a certain point of his life, with no apparent cause, an human being felt ill, also pain, and died. This event, in various ways, occurred always: Every human being had not a predator’s (or accident’s) victim, invariably died anyway. The abstractive operator, from every single death event, inferred that every human being had to die, i. e. it allowed human beings to be aware of their own mortality. Thanks to the linguistic operator, this awareness of death became collective: Even human beings who had not arrived alone at the concept of the universality of death came to know of it by other humans. We can presume this awareness was very shocking. Most scholars agree that human are, and always were, troubled by their own mortality, and some of them had explained many cultural human features as derived from this awareness (Terror Management Theory: See, for example, Greenberg et al. 1997, or Salzman 2001). Moreover, as Hinde has noticed (1999, p. 59), among presentday humans, even if “most people profess to accept the prospect of death, indirect methods … indicate that unconscious fear is widespread (Beit- Hallahmi and Argyle 1997; Hood et al. 1996)”. Ethnological research of the past two centuries has shown that preliterate people do not think death as a natural datum: “On the contrary, all the narratives on the origin and cause of death have shown in evidence, without exceptions, the unnaturalness of death” (Widengren 1969). So we may infer that Homo sapiens is the only animal who knows he is mortal, and that he does not immediately recognize this fact as a natural datum: he may think so by means of a rational effort. Homo sapiens, as historical and ethnographic data show, rather and immediately feels that death is a violence he has to suffer: all the more reason it could have been considered so by the first Homo sapiens who were conscious of death. We cannot know when this awareness took place. McBrearty & Brooks, in their very expansive review of the origin of modern human behavior (2000), refer (p. 519) that “earliest evidence for burial among H. sapiens is found in the Levant at the site of Qafzeh”, that these are dated to ca. 120, - 90,000 years ago and that at least one may be associated with grave goods. b) A scheme of the Triggered Psychic/Neurobiological Mechanism. On the basis of our neurobiological knowledge, we can try to reconstruct the psychic mechanism triggered by first death awareness in ancient humans, roughly locating the involved brain structures using the triune model, as follows and as it is depicted in fig. IX. 1. 1) Homo sapiens perceived death. 2) The Neocortex realized the awareness that all human beings had to die, and in the same time death was felt as a violence. 3) Neocortical causal operator, as it spontaneously tends to impart a sense of causality on all the events (the causal imperative or need of explanation), tried to find the cause of the mortal violence exerted over humans. But it obviously failed in finding an empirical cause. 4) The Limbic System received the awareness of death and the anxiety derived from the lack of any empirical explanation, and, as it was shocked by them, turned them into a distressing stimulus affecting the Neocortex and R-complex; at the same time its inhibitory action on the R-complex decreased. 5) According to R-complex way of thinking (due to its behavioral operators), violence must be performed by a dominant individual. To a similar conclusion ancient Homo sapiens’ R-complex had unconsciously to come owing to its internal neuronal circuits linking cognitive operators of the caudate nucleus (the “thinking” part of the R-complex and main site of the pseudo-causal operator) with behavioral operators as the hierarchic one lying in the pallidum. 6) The result of such a reptilian elaboration was projected to Neocortex, where it became conscious. The Neocortical causal operator, as no other active and empirical cause he might find, had to recognize in the reptilian concept of a dominant individual the agent of the mortal violence inflicted on humanity. Such a recognition was not difficult to causal operator, because, as we have said above (§ III), it includes an agent-detection device. Therefore: 7) The Neocortex formulated the “Immense Powerful Being” concept. 8) However it was still an indefinite concept. The neocortical association areas, in relation to the ecological, cultural and social environments (9 a), and with more or less relevant contributions by the caudate nucleus (thank to the reverberating circuit Neocortex – Associative Striatal Regions – Neocortex – etc., as we have described in § III), developed various systems of myths (9 b) in order to explain mankind-god relations (first the reason of mortal violence inflicted to humans) and systems of rites allowing the establishment of some kind of communication between humans and the Powerful Being(s). Inside the neocortical association areas that tried to explain the reason of mortal violence inflicted by the “Immense Powerful Being” to the humans, an important role might be performed by the “metacognition” or “theory of mind” operator. Finally, but it is fundamental, it must be emphasized that every system of belief may be felt as true only if the Limbic System’s existential operator (9c) gives it a sense of reality. In other words, in the presence of equivalent fantastic tales (regards neocortical judgment) the existential operator chooses which are fancy products and which are sacred, or revealed (and so absolutely true), tales. Thanks to the activities of above mentioned operators, we may agree with Geertz characterization of religion, as well as a totalitarian ideology, as a system of symbols which acts to establish powerful, pervasive, and long-lasting moods and motivations in humans by formulating conceptions of a general order of existence and clothing these conceptions with such an aura of factuality that the moods and motivations seem uniquely realistic (Geertz 1966). However, in Geertz’s view, religion, or a totalitarian ideology, is a Cultural System, in our view it is a Bio-cultural System based on neurognostic development, driven by the logical emergent properties of our innate operators. Cognitive or behavioral operators, such as the hierarchic one, may correspond to Jung’s archetypes. As remarked by MacLennan (2002), archetypes are not innate images but dynamical structures of perception and behavior. They reside in the collective unconscious, for the archetypes are unconscious until they are activated, and they are collective in that they are common to all humans. They become images after they are activated by a sign stimulus. In the present reconstruction of the “Powerful Being Concept” rising, hierarchic and agent detection operators are the most important archetypes, and the awareness of death is the sign stimulus. c) The “Soul” Concept As the invisible “Powerful Being” is the “chief” of human beings, he is like them, that it is to say human beings are like him, and so also humans have an invisible part, the “soul”. This more or less, might have been the line of reasoning that allowed the first humans that produced the “Powerful Being” concept to produce also the “soul” concept. This reconstruction of the relation between “Spiritual Beings” and “soul” is symmetrical, or anti-parallel, in comparison with that one of Tylor, according to humans first postulated souls in all of nature and then proceeded to postulate “spiritual beings” or gods. We think that the triggering stimulus we have proposed to elicit the god concept as first is more fit than that one proposed by Tylor to elicit the soul concept as first. The typical human capacity of thinking insensate beings has probably been favored by oneiric activity. The “World of Dreams”, in fact, could have been explained by the first human beings as a reality - apart from, different from, the sensate one, but not less real. In support of this idea, we note that in religious thought dreams have often been seen as a form of communication with the divine § X. Reptilian, Limbic, and Neocortical Divine Beings As Divine Beings result from the projection of a brain activity, their features must be result from the mixing in various parts of projections arising from the R-complex, the Limbic System and the Neocortex. We subsequently examine each of the main features these three brains that contribute to the emergence of the “Immense Power Being” concept. Reptilian Divine Beings In the “Immense Power Being” projection of the group-chief, independent of cultural variations, we note the manifestation of a more or less complete SM groupchief prerogative: it exercises its power on a territory, dominates other males, which behave submissively, and has priority in access to food and to females. In the same manner, its supernatural projection exercises: a) Territorial domination. This may take the form of control over a single people or be more wide, to include all of the world. b) Domination over males. This may be shown in various ways in which males: - assume positions that denote submission and originate from the female sexual presentation; genuflection (from the Latin words genu, knee, and flectere, to bend) is an euphemistic word or action rendering an unconscious primate female sexual presentation to a dominant male (fig V 8 e). - submit themselves to sexual mutilations (such as circumcision) that symbolizes castration or a reduction of masculinity, so that full masculinity remains an exclusive attribute of the “Powerful Being,” and others are required to abstain from sexual relations c) Priority in food access: ritual offers, especially of early fruits. d) Ownership of females: matrimonial rites, consecration of virginal maidens to the service of the god, or, on the contrary, sacred prostitution, by which the god shows his magnanimity towards submissive males. In some cases the priority in female possession is delegated by the god to his representatives on earth (“ius primae noctis”). It is not surprising that if males must assume a submissive attitude toward the “Powerful Being”, females must do likewise. In summary, a Reptilian Divine Being is obsessed with sex and the control of his subordinates’ sexual activities, as an SM Primate group-chief does. We may tell that a Reptilian Divine Being is not only “anthropomorphic”, but, more properly, “pithecomorphic” (pithec- being the Greek root for “monkey”). Limbic Gods. Limbic Divine Beings show features arising from limbic operators, mainly the “falling in love” and attachment operators but also nursing and call ones: these operators constitute an attachment system (see Kirkpatrick 2005). The mammalian attachment system was selected to make the organism acutely aware of the whereabouts of caregivers who can provide protection. Research on wide range of mammalian species illustrates the desire of the infant to remain in close proximity, even in contact with the mother, illustrating a special kind of psychological bond . Virtually any threat to the infant activates this attachment system, leading the infant to seek physical contact with the caregiver, or vocalizing distress to attract the caregiver. The attachment system is an evolved mechanism, an adaptation for maintaining close proximity of a dependent infant with the caregiving parent. This contact reduces fear and the anxiety, and induces a feeling of security, a basis from which a sense of mastery of the environment emerges. This bonding with the care giver is so fundamental to mammalian survival that the failure to bond may produce a withering away syndrome leading to death. This need for protection is not limited to the infant phase, but persist as an underlying need during adulthood. This mamalian attachment bond between infant and mother maybe generalized to a variety of other caretakers, as well as peers and mates. These attachment processes are lifespan issues, being significant factors in emotional adjustment from birth through end of life. Romantic love, for example, has been characterized as the integration of the attachment system with the caregiving and reproductive systems. According to Kirkpatrick (2005), the attachment system constitutes the basis for the relationships with the most important significant other--God. In many religions, God is often portrayed as an ideal attachment figure, as well as a model for one's own caregiving and attachment behaviors. Kirkpatrick provides a detailed analysis to illustrate how Christian relationships with God constitute attachment relationships that provide a sense of safety and security. He reviews a range of psychological studies that illustrate that the attachment relationship with God is “real”, and functions similarly to parent child relations during childhood. Kirkpatrick reviews a variety of studies to illustrate the major ways in which God functions as a substitute attachment figure. For those with deep religious commitment, their perceived relationships with God parallel the expressions of emotional love that are found in attachment relationships with parents. “In particular, beliefs about what God or gods are like, and the ability to have personal relationship with God, appear to be consistent with one's experience in human relationships with attachment figures” (Kirkpatrick pp.125-26). This is one of correspondence, where and God and human attachment relationships are similarly structured. Another is an opposite pattern of compensation, and where people who lack secure attachment relationships find in their relationships with God the interpersonal relationships that they lacked and interaction with others. This pattern of attachment relationship with God is often characterized as “falling in love”, and often manifested as a dramatic religious conversion. This latter pattern has generated many theories of the origins of religion, suggesting that the basic dynamics of beliefs and a God reflects a substitute relations for beliefs and the caring power of a father or other caregivers. A variety of empirical studies indicate that God relationships not only function psychologically as attachment relationships, but that in doing so they provide a range of psychological benefits. The extent of one’s religious commitment contributes directly to an internal locus of control, a sense that one is personally competent and able to address the problems that one confronts. This sense of religiosity inhibits feelings of anxiety and contributes to a sense of optimism about the future. People who experience a sense of the presence of God during prayer or other ritual experiences are likely to have higher levels of well-being. This sense of intrinsic religious orientation, a genuine commitment to one's religious beliefs, is popularly associated with a variety of measures of health including inducing a sense of personal competence and control and freedom from worry. Summarizing, Limbic Divine Beings show feature of parents (father or mother like), or brother/sister. These features do not exclude feelings of fear, but sometimes they permit some licences: after all a swearer is like a child strongly protesting against his parents. Neocortical Absolute Beings. The human being has subsequently elaborated the “Perfect Being” concept. Such a Being is the fruit of philosophical elaboration in the Neocortex. Theological and philosophical speculations have tried to conciliate the “Powerful” with the “Perfect” Being, mainly trying to rationalize and justify actions associated with belief in the Powerful Being which seemed incompatible. with human moral standards. So the human being has reached various conceptions of the Perfect Being, such as Spinoza's “Deus sive Natura,” the deism of most of followers of the Enlightenment, the Idealistic Absolute Spirit, the Masonic Great Architect of the Universe. We can make no choice in this regard, even if we think much more likely the existence of a “Perfect Being” which is his neocortical image rather than one of reptilian origin, such as the “Powerful Being”. Reptilian, Limbic, and Neocortical Gods in human history. Returning to our § IX b scheme, the first Powerful being that emerged in human mind presumably was primarily endowed with Reptilian features. However, the subsequent development of the concept of a soul that, being similar to the Supernatural Being, was believed to be immortal, attenuated the fear of death. Also human beings had supernatural features, and other spiritual beings, more human-like, i. e. more or less good, more or less bad, might be conceived to exist. As ancient hunter-gatherers most likely lived in egalitarian societies, the conception of a Reptilian God was in some aspect antithetical regards their social organization: We may presume that rapidly the Supreme Being assumed Neocortical and Limbic features, the first or the second ones prevailing according to other factors influencing the cultures and ideas development, whereas Reptilian features became more properly typical of less important spiritual beings, ghosts, devils and similar. We may think these conceptions broadly spread and persisted in hunter-gatherers till now: This form of religion is called shamanism, a topic on which we refer to Winkelman’s studies. In the regions of the world in which the demographic growth was faced by agriculture (Harris 1977), more complex social entities such as chiefdoms began to appear, followed by kingdoms and empires, as summarized by Summers (2005), who adds that the development of these societies was associated with increasing despotism, in term of control of resources and differential reproduction. As the history of ancient civilizations (Egypt, Mesopotamia, China, India, Greece and Mesoamerica) shows, Divine Beings that we may consider “reptilian” became very important, and often the more expensive rituals and sacrifices were addressed to them. In most agricultural chiefdoms, patrilinear lineages were subsequent to matrilinear lineages (as females first practiced agriculture and had the ownership of the fields), and in many civilizations this sociality lasted till our historical period. Pettazzoni (1956) illustrated that the Supreme Being Concept assumed various features according to the civilization he was belonging to: So in a patriarchal civilization the Supreme Being had father’s features, in agricultural matrilinear societies had the Mother Earth female features. Nevertheless, whereas the male Supreme Being was omniscient and all-seeing, the female Mother Earth never was so: The power of the two kinds of Supreme beings was different, and Pettazzoni failed to find the cause. On the basis of our knowledge of Primate societies, we may now suggest an explanation: As in Primates the male sex means dominance and the female sex means submission: the Mother Goddess, although she might rule most natural events, could not completely assume Immense Power Being’s features. In other words the conception that reptilian deities were the first and limbic and neocortical ones were subsequent in a progressive sequence is conceptually wrong, as it was been disproved by ethnological studies about early cultural evolutionary theories, such as Tylor’s or Frazer’s ones. In summary, in a Neo-Darwinian perspective we may study the cultural history of a population and may distinguish in it some stages, but they are not in a pre-defined sequence, nor they are necessarily in a progression of increasing values. In most hunter-gatherer human group is (and was) present the believe in a Supreme Being that is thought as not involved in human affairs, as a “Neocortical” Divine Being can do, whereas in most agricultural groups (necessarily originated from hunter-gatherer groups) are present one or more Divine Beings that are involved in human affairs, claim sacrifices and often are obsessed with the control of human sexual life as a primate group-chief might do, so that we may call them “reptilian” deities. § XI. Divinized human chiefs and charismatic leaders. In the course of history, human beings that, by birth or historical events, achieved absolute or complete power on other humans did not usually (or, as we know, ever) show their power as having human nature, but as endowed with divine nature, or as invested by Gods. For example, ancient Egypt’s kings, Inca, Chinese and Japanese emperors were gods or gods' sons. Also Roman emperors, clashing with the traditional laicism of the previous consular power, claimed to be regarded as gods. In ancient Israel, kings as Saul or David were such since they were anointed by a prophet who acted on divine behalf. In the Holy Roman Empire, emperors were crowned by Catholic Popes. In the European feudal system, kings, feudatories and vassals claimed their authority over common people was based on divine behalf, as sanctioned by the Holy Bible, such as in Pauline Letters (Rom 13, 1-2: “1Let every person be subject to the governing authorities. For there is no authority except from God, and those that exist have been instituted by God. 2Thereforehe who resists the authorities resists what God has appointed, and those who resist will incur judgment”). We may state that from studies carried out on human societies by syncronic or diacronic methods, it seems that every absolute power cannot be presented as merely “human”, but only as “divine”. This suggests that, after the transition from SM breeding and social systems to pseudo-Mo breeding and MM social systems, no human could claim to be endowed with an absolute power. When thousands of years later, an absolute powerful being was conceived as the agent of death, he was conceived as superhuman. The Immense Power Being idea proposed by the R-complex had to be projected outside the mankind: the new “group chief” was similar to human beings, but he was not human. And when, at least from the development of chiefdoms, an human being was able to achieve a sort of absolute power, he could not be considered a simple human being, but someone endowed with divine features. The deified group chief returned among humans deifying in turn a human chief. An intriguing figure of chief is formed by charismatic leader. It’s generally about a human being that is not a chief by birth but that is able to convey towards his/her person people’s expectations in a particular historical time. He/she proposes the answer to such expectations exposing an ideological system that rapidly spreads not because its rationality rather because it stimulates audience’s emotionalism. This implies that who proposes himself as leader must have especially rhetorical abilities, since rationality is less important. Emotional stimuli destabilize in audience the control action of the Limbic System: as we have said above (§ II), according to MacLean, alterations of the Limbic System’s activities can produce various experiences and feelings, as those associated with “revelation” of truths, and strong feelings attached to systems of beliefs (that, in this case, are in progress), regardless of whether they are rational or irrational. When the Limbic System, instead of inhibiting, takes a function of emotional stimulus, R-complex’s operators may be disinhibited, particularly two of them: the hierarchic and the isopraxis ones. The hierarchic operator’s action, in the followers, causes that who was able to become a charismatic leader assumes a very strong power upon followers, and as a rule he is called and rewarded as a “leader” (dux, fuhrer, helmsman, and the like) that will be able to go with his followers towards a mythological end assigned to the History. If such an end is conform to a religious faith, the charismatic leader become the person chosen both by people and by divine being, the Anointed of the Lord, the Man of the Providence, the God’s Prophet and the like, and his power become total. At the same time the isopraxis operator causes the followers to imitate leader’s movements, or to respond to his harangues marching or dancing all together for him, and singing his praises; in such a way they standardize their behaviors and their mental attitudes. They become easily indoctrinated and execute even most despicable orders without any objection. What might be considered as inhuman or dreadful if done by a single human, can become a lawful or holy action if it is done by altogether owing to leader’s incitement. § XII. Conclusive remarks on our hypothesis. The biological bases of religion, that we have supposed from § I, consist in summary in a few generalized characteristics of the human way of thinking, owing to brain structures and operators, as: 1) the inductive capacity of abstracting from single observations general rules (inductive operator); 2) the necessity of finding a cause-effect nexus among observed phenomena 3) 4) 5) (causal operator), and, more properly, that some causes indicate the presence of some living agent (agency detection operator); the capacity of projecting one’s own thought and emotions in others (theory of mind, or metacognition , operator); the capacity to accept dominant and subordinate roles in a hierarchic society, accepting rewards vand punishments by dominant individuals. These generalized structural characteristics have been produced by natural selection because they allowed Mammals (and Hominini in particular) to face better the environment. So religion, regards its origins, must be considered an emergent product of naturally selected mind functions. This perspective might be seen as in agreement with scholars such as Maser & Gallup (1990), Kirkpatrick (1999, 2005), Boyer (2003), expressed in the conclusions of Atran & Norenzayan (2004) that “religion is not an evolutionary adaptation per se, but a recurring cultural by-product of the complex evolutionary landscape that sets cognitive, emotional and material conditions for ordinary human interactions” (A. & N. 2004, p.713). We may reasonably differ from this perspective in seeing the superordinate god concept as an evolved mechanism for allowing the integration of larger groups of humans, making a “Dominant other” so distant and yet powerful that it could unite millions to a common cause. The “Immense Power Being” concept certainly has shown its ability to foster reproduction and survival in its members in the major religions of the world today. In allowing humans to extend male-male bonds, provided a major evolutionary extension of the human ability to live in groups supordinated to a common hierarchical power. While the “Immense Power Being Concept” exapts many prior dominance structures, as well as social and personal operators, it provides something more in the conceptualizations of a omniscience, omnipotence and superordinate identity. The Immense Power Being concept take the conceivable possibilities beyond human pale, symbolically representing further possibility for human modeling and development. This moves developmental goal beyond the genetically driven selfish interests of individual inclusive fitness to permit a commitment to supporting a larger non-kin social group that helps ensure intergroup competition for survival. In these sense, the Immense Power being concept provides a mechanism for integrating larger groups that intensifies intergroup selection (D.S. Wilson). Our “agency detection mechanism” evolved to respond to things “out there” that were vitally important for our ancestors to recognize; it come to be focused on other groups, humans’ most important competitors and predators. But it was extended to connect with for all sorts of phenomena, particularly those causal systems involving unknown complex agents, designs and origins. Spirit concepts involve these kinds of agents with capacities beyond ordinary human capabilities. Spirit beings possess a variety of personal and social characteristics of humans, as well “non-intuitive” properties that “contradict” both the ordinary principles of the Universe and the ways in which our brains understand the Universe. Spirits “supernatural” abilities expanded beyond our human behavioral capacities. What pre-adaptation constitutes the basis for the exaptation providing the basis for super human capabilities manifested in the supernatural? What exaptation provides the counterintuitive properties we ascribe to the supernatural? The lack of an apparent answer to these questions suggests that religion may provide an adaptation through those aspects of the supernatural premise that relate to these non-intuitive, contradictory, and “super”-human capabilities. The supernatural concept represents an extreme manifestation of the symbolic process that makes it possible for us to represent and thereby conceive of “things” we can never actually see or touch as empirical objects. This leads to another way in which religion’s supernatural assumption is adaptive. For the belief that “supernatural others” have access to our thoughts helps to inhibit deception and encourage group loyalty and sacrifice. Wildman points out that this provides a basis for a compelling interest in adhering to moral standards established by the supernatural. As the German philosopher Immanuel Kant contended, moral reasoning requires a religious framework in order for it to be rational, assuming an ultimate moral authority that establishes standards and allocates rewards and punishments. To the extent that the omniscient (all-knowing) properties of the supernatural engage possibilities beyond that of our innate mental hardware, religious thought provides an adaptation. The supernatural concept may offer an additional advantage by expanding our ability to internalize these “supernatural others” and their super human capabilities, thereby providing a basis for addressing problems of reliability in assuming the honesty of others. The super human capabilities that enable omniscience can provide a policing function and assure one’s adherence to normative expectations, even if our normal social others—the other members of our society—are not capable of detecting our behavior. However different natural or social environments may act different selective pressures on the different genes that constitute the biological foundations of brain operators. For example, in some societies a well-functioning hierarchic operator (producing submissive individuals) may be fitter that a minor one (producing anarchic individuals). So, as submissive individuals may have more reproductive success than anarchic individuals, parallel reptilian gods may have more followers than limbic or neocortical gods. Another case may be done by genetic drift, in which a particular gene, for example that favoring magic thought, or mystical behaviors, may be abundant in the group that founds a new population in a new land. In this respect, the adaptative value of religion, that some scholars (as Bulbulia, 2004, or Sosis, cited) are studying, has to be investigated. But in an other paper. Figures (Draft that has to be retouched) Fig Telencephalon Diencephalon II.1 Endbrain Mesenc- Metenc- Myelen- ephalon ephalon cephalon Mid- Hindbrain Interbrain F o r e b r a i n brain E n c e p h a l o n S p i n a l C o r d Abbreviations. BG: basal ganglia; CB: cerebellum; D CTX: dorsal cortex (neo-cortex); HY: hypothalamus; L CTX: lateral cortex (palaeo-cortex); M: medulla oblongata; M CTX: medial cortex (archi-cortex); P: pons; SP: spinal cord; T: tectum; TG: tegmentum mesencephali; TH: thalamus. Fig. II.3 Scheme of the R-Complex in Saimiri (adapted from MacLean) Fig. II:4.The three main regions of the Limbic System (adapted from MacLean) Abbreviations: A.T.: Anterior Thalami Nucleus; HYP.: Hypothalamus; M.: Mammillary body; M. F. B.: Medial Forebrain Bundle; OLF.: Olfactory; PIT.: Pituary gland, or hypophysis. Fig. III.1 (a: section at anterior commessure level; b: more rostral section). Abbreviations: AC: Anterior Commissure; Acc: Nucleus Accumbens; ASR: Associative Striatal Region; CC: Corpus Callosum; CD: Caudate Nucleus; GP: Globus Pallidus; LSR: Limbic Striatal Region; PUT: Putamen; SMSR: SensoriMotor Striatal Region. Fig. V.1 a Fig. V.1 b Fig. V.1 c Fig. V.1 d Fig. V.1 e Fig.V.1 f Fig. V.2 a : copulation in Papio Fig. V.2 b : a male mounts a Fig. V.2 c : a female mounts a hamadryas. subordinate subordinate male as rank demonstration. female as demonstration Fig. V.2 d A low rank individual acquires group chief’s support against an high rank male. Fig. V.2 e Homages to an ancient Egyptian dignitary. Fig. V.2 f Christian fundamentalists in prayer. rank Table VI 1 Breeding groups Social Groups Representative species Competition levels Mo or Pair Hylobates spp. Symphalangus spp. I° level I° level MM Lemur catta II° or IV° level Pan spp. Papio cynocephalus Papio ursinus Papio anubis II° level IV° level IV° level IV° level Gorilla gorilla III° level Pongo pygmaeus Presbytis cristatus Presbytis obscurus Presbytis entellus III° level III° level III° level IV° level Papio Hamadryas Theropithecus gelada IV° level IV° level Mo MM MM MM SM SM Multi SM Fig. VII 1 Homo sapiens 1200-1500 cm3 ] Low sexual dimorphism. ] Pseudo Mo , but also SM ] and MM breeding groups. Homo ergaster/erectus ≈ 1000 cm3 ] Low sexual dimorphism ] Probable pseudo Mo breeding Homo rudolfensis/habilis ≈ 600 cm3 Australopithecus ≤ 500 cm ] and MM social groups. ] Dimorphism reduction; probable 3 garhi ] passage from SM to pseudo Mo ] breeding groups. Australopithecus < 500 cm3 afarensis ] High sexual dimorphism; ] probable SM breeding groups. Australopithecus anamensis ] Medium (chimp-like) dimorphism; Ardipithecus kadabba < 500 cm3 4 ] probable MM breeding groups. 2 Male-male competition levels (Plavcan and van Schaik) Scheme VII 1: Figure’s VII 1 table Cranial capacity Sexual dimorphism and breeding system Fig. IX. 1 (“Fig. 10” refers to a former numeration: it has to be deleted). References Adolphs R., D. Traner & A. R. Damasio 1998 The human amygdala in social judgement. Nature 393: 470-474. Aiello L. C. & P. Wheeler 1995 The expensive-tissue hypothesis: The brain and the digestive system in human and primate evolution. Current Anthropology 36: 199-221. Alper M. 1996 The “God” Part of the Brain. New York: Rogue Press. (5th Edition: 2001). Ariëns Kappers C. U. 1909 The phylogenesis of the palaeo-cortex and archi-cortex compared with the evolution of the visual neo-cortex. Arch. Neurol. Psychiatry (London) 4: 161-173. Aron A., H. Fisher., D. Mashek, G. Strong, H. Li, L. L. Brown 2005 Reward, motivation and emotion systems associated with early-stage intense romantic love. J. Neurophysiol 94: 327-337. Atran S. & A. Norenzayan 2004 Religion’s evolutionary landscape: counterintuition, commitment, compassion, communion. Behavioral and Brain Sciences 27: 713-730. Barrett J. L. 2000 Exploring the natural foundations of religion. Trends in Cognitive Sciences 4: 29-34. Baumgarten H. G. & M. Göthert (eds.) 1997 Serotoninergic Neurons and 5-UT Receptors in the CNS. Berlin: Springer-Verlag. Beit-Hallahmi B. & M. Argyle 1997 Religious behavior, belief, and experience. London: Routledge. Boyer P. 2001 Religion Explained: Evolutionary Origins of Religious Thought. New York: Basic Books. 2003 Religious thought and behaviour as by-products of brain function. Trends in Cognitive Sciences 7 (3): 119-124 Bramble D. M. & D. E. Lieberman 2004 Endurance running and the evolution of Homo. Nature 432: 345-352. Brelich A. 1970 “Prolegomenes à une histoire des religions”. In: H. Ch. Pucch (ed.), Histoire des Religions. Paris: Librairie Gallimard. Bulbulia J. 2004 The cognitive and evolutionary psychology of religion. Biology and Philosophy 19: 655-686. Burkert W. 1972 Homo necans: Interpretationen altgriechischer Opferriten un Mythen. Berlin: Walter de Gruyter. 1996 Creation of the Sacred. Tracks of Biology in Early Religions. Cambridge: Harvard University Press. Buss D. M. 1999 Evolutionary Psychology. The New Science of the Mind. Boston: Allyn & Bacon. Buss D. M. & T. K. Shackelford 1997 Human Aggression in Evolutionary Psychological Perspective. Clinical Psychology Review 17 (6): 605-619. Chaline J., A. Durand, D. Marchand, A. Dambricourt Malassé, MJ. Deshayes 1996 Chromosomes and the origins of Apes and Australopithecines. Human Evolution 11(1): 43-60. Clutton-Brock T.H. & P.H Harvey 1977 Primate ecology and social organization. The Journal of Zoology 183: 1-39. Cory G. A. & R. Gardner (eds.) 2002 The Evolutionary Neuroethology of Paul MacLean: Convergences and Frontiers. Westport: Praeger. d’Aquili E. G. & A. B. Newberg 1999 The Mystical Mind: Probing the Biology of Religious Experience. Minneapolis: Fortress Press. Darwin C. R. 1871 The Descent of Man and Selection in Relation to Sex. London: Murray. Delgado J.M.R. 1969 Physical control of the Mind. Toward a Psychocivilized Society. New York: Harper & Row. DSM III 1985 The Diagnostic and Statistical Manual of Mental disorders, III ed. Washington: American Psychiatric Association Press. Durkheim E. 1912 Les formes élémentaires de la vie religieuse (Le système totémique en Australie). Paris: F. Alcan. Ernandes M. & S. Giammanco 1998 MacLean’s Triune Brain and the Origin of the “Immense Power Being “ Idea. The Mankind Quarterly 39 (2): 173-201. Feuerbach L. A. 1841 Das Wesen des Christenthums. Leipzig. (The Essence of Christianity, G. Eliot tr., 1989, Prometheus Books//1958, P. Smith// 1942, Harp C.). Fialkowski K. 1986 A Mechanism for the Origin of the Human Brain: A Hypothesis. Current Anthropology 27: 288290. 1987 On the Origins of the Human Brain: Preadaptation vs Adaptation. Current Anthropology 28: 540543. Firth R. 1963 Offering and sacrifice. Journal of the Royal Anthropological Institute 93: 12-24. Flinn M. V., D. C. Geary, C. V. Ward 2003 Ecological dominance, social competition, and coalitionary arms races: Why humans evolved extraordinary intelligence. Evolution and Human Behavior 26: 10-46. Freud S. 1912/13 Totem und Tabu. Leipzig: Heller. (Totem & Taboo, 1989, Buccaneer Bks//1976, Amereon Ltd//1960, Random). Gazzaniga M. S. 1985 The Social Brain. Discovering the networks of the mind. New York:Basic Books Inc. Geertz C. 1966 Religion as a cultural system. In: Banton M. (ed.) Anthropological approaches to the study of religion, p. 1-46. London: Tavistock. Haile-Selassie Y., G. Suma, T. D. White 2004 Late Miocene Teeth from Middle Awash, Ethiopia, and Early Hominid Dental Evolution. Science 303: 1503-1505. Hamer D. 2004 The God Gene. New York: Doubleday. Harris M. 1977 Cannibals and Kings. The Origins of Cultures. New York: Random House. 1979 Cultural Materialism: The Struggle for a Science of Culture. N. Y.: Random House. 1989 Our Kind. New York: Harper & Row. Harvey P.H., M. Kavanagh. & T.H. Clutton-Brock 1978 Sexual dimorphism in primate teeth. The Journal of Zoology 186: 475-485. Hauser M. C., N. Chomsky & W. T. Fitch 2002 The Faculty of Language: What Is It, Who Has It, and How Did It Evolve? Science 298: 15691579. Higley J. D., P. T. Mehlman, D. M. Taub, S. B. Higley, S. J. Suomi, M. Linnoila, J. H. Vickers 1992 Cerebrospinal Fluid Monoamine and Adrenal Correlates of Aggression in Free-Ranging Rhesus Monkeys. Archives of General Psychiatry 49: 436-441. Hinde R.A. 1987 Individuals, Relationships and Culture. Cambridge: Cambridge University Press. 1999 Why Gods Persist. London – New York: Routledge. Hubert H., Mauss M. 1898 Essai sur la nature et la fonction du sacrifica. L’Année sociologique 2: 29-138. Insel T. R. & L. J. Young 2000 Neuropeptides and the evolution of behavior. Current Opinion in Neurobiology 10: 784-789. Isaacson R. L. 1982 The Limbic System. Second edition. New York: Plenum Press. Jacobs B. L & F- C. Azmitia 1992 Structure and Function of the Brain Serotonin System. Physiological Reviews 72 (1): 165-229. Jolly C. 2001 A proper study for mankind:Analogies from papionin monkeys and their implications for human evolution. Yearbook of Physical Anthropology 44: 177-204. Kandel E. R., J. H. Schwartz & T.M. Jessel 1996 Principles of Neural Sciences (3rd edition). Appleton & Lange. Kaplan J. R., J. Phillips-Conroy, M. B. Fontenot, C. J. Jolly, L. A. Fairbanks, & J. J. Mann 1999 Cerebrospinal Fluid Monoaminergic Metabolite Differ in Wild Anubis and Hybrid (anubis hamadryas) Baboons: Possible Relationships to Life History and Behavior. Neuropsychopharmacology 20 (6): 517-524. Kant I. 1781 Kritik der reinen Vernunft. Riga: J. F. Hartknock (Critique of Pure Reason, Politis V. ed., 1993, C. E. Tuttlel// Meiklejohn J. M. tr., 1990. Prometheus Books// Smith N. Y. ed., 1969, St Martin). Kirkpatrick L. A. 2005 Attachment, Evolution, and the Psychology of Religion. New York: The Guilford Press. Koestler A. 1978 The ghost in the machine. London: Hutchinson and Co. Leutenegger W. & J. T. Kelly 1977 Relationship of Sexual Dimorphism in Canine Size and Body Size to Social, Behavioral, and Ecological Correlates in Anthr. primates. Primates 18(1): 117-136. Lorenz K. 1973 Die Rückseite des Spiegels. Versuch einer Naturgeschichte menschlichen Erkennens. München: R. Piper & Co. Verlag. (Behind the Mirror: A Search for a Naturai History of Human Knowledge, Taylor R. tr., 1978, Harcourt Brace, P. Smith) MacLean P.D. 1970 The triune brain, emotion, and scientific bias. In F.O. Schmitt (ed.), The neurosciences: Second study program, pp. 336-349. New York: Rockefeller University Press. 1973a A triune concept of the brain and behavior. In T. J. Boag & D. Campbell (eds.), The Clarence M. Hincks Memorial Lectures, 1969, pp. 1-66. Toronto: University of Toronto Press. 1973b The brain’s generation gap: some human implication. Zygon, Journal of Religion and Science 8: 113127. 1973c Effects of pallidal lesions on species-typical display behavior of squirrel monkey. Fed. Proceedings 32. 384 ... 1978 A Mind of Three Minds: Educating the Triune Brain. Chicago: University of Chicago Press. 1985a Evolutionary psychiatry and the triune brain. Psychological Medicine 15: 219-221. 1985b Brain Evolution Relating to Family, Play, and the Separation Call. Arch. of General Psychiatry 42: 405-417. 1990 The Triune Brain in Evolution. Role in Paleocerebral Functions. New York: Plenum Publ. Co. MacLennan 2002 Evolutionary Neurotheology and the Varieties of Religious Experience. In: R. Joseph (ed.), Neurotheology: Brain, Science, Spirituality, Religious Experience, pp. 305-314, University Press, California Mandell A. J. 1980 Toward a Psychobiology of Transcendence. God in the Brain. In: Davidson & Davidson (eds.), The Psychobiology of Consciousness. New York: Plenum Press. Marazziti D., H. S. Akiskal, A. Rossi, G. B. Cassano 1999 Alteration of the platelet serotonin transporter in romantic love. Psychol. Med. 29: 741-745. Maser J. D. & G. G. Gallup 1990 Theism as a By-Product of Natural Selection. The Journal of Religion 70 (4): 515-532. McBrearty S. & A. S. Brooks 2000 The revolution that wasn’t: a new interpretation of the origin of modern human bahavior. Journal of Human Evolution 39: 453-563. McFarland D. 1985 Animal Behaviour. Harlow (UK): Longman Scientific & Technical. McNamara P. 2002 The motivational origins of religious practices. Zygon. J of Rel. & Sci. 37 (1):143-160. Mehlman P. T., J. D. Higley, I. Faucher, A. A. Lilly, D. M. Taub, J. Vickers, SJ. Suomi, M. Linnoila 1992 Low CSF 5-HIAA Concentrations and Severe Aggression and Impaired Impulse Control in Nonhuman Primates. American Journal of Psychiatry 151 (10): 1485-91. Morris D. 1967 The Naked Ape. London: Jonathan Cape. (1994, London: Vintage Books/Random House). Morris J. S., A. Öhman & R. J. Dolan 1998 Conscious and unconscious emotional learning in the human amygdala. Nature 393: 467-470. Newberg A. B. & E. G. d’Aquili 2001 Why God won’t go away. Brain Science and the Biology of Belief. New York: Ballantine Books. Niewenhuys R., H. J. ten Donkelaar & C. Nicholson (eds.) 1998 The Central Nervous System of Vertebrates. Berlin: Springer-Verlag Oatley K. 2004 Emotion:Brief History. Blackwell Publishing. Pals 1996 Seven Theories of Religion. New York: Oxford University Press. Parent A., Hazrati L.-N. 1995a Functional anatomy of the basal ganglia. I. The cortico-basal ganglia-thalamo-cortical loop. Brain Research Reviews 20: 91-127. 1995b Functional anatomy of the basal ganglia. II: The place of sub-thalamic nucleus and external pallidum in basal ganglia circuitry. Brain Research Reviews 20 (1): 128-154. Pettazzoni R. 1956 The All-Knowing God. London: Methuen. (Or.: L’onniscienza di Dio. Torino, 1955). Pickford M. & B. Chiarelli (eds.) 1986 Sexual dimorfism in living and fossil primates. Florence:. Il Sedicesimo. Pinker S. 1997 How the mind works. New York: W. W. Norton & Co. Plavcan J. M. 2002 Sexual dimorphism in primate evolution. Yearbook of Physical Anthropology 44: 25-53. Plavcan J. M. & C. P. van Schaik 1992 Intrasexual competition and canine dimorphism in anthropoid primates. American Journal of Physical Anthropology 87:461-477. 1994 Canine Dimorphism. Evolutionary Anthropology 2: 208-214. 1997 Interpreting hominid behavior on the basis of sexual dimorphism. Journal of Human Evolution 32: 345-374. Porges S. W. 2001 The polyvagal theory: phylogenetic substrates of a social nervous system. Int. J. of Psychophysiology 42: 123-146. Povinelli D. J. & J. Vonk 2003 Chimpanzee minds: suspiciously Human? TRENDS in Cognitive Sciences 7(4): 157-160. Raleigh M.J., G. Brammer, A. Yuwiler, J. W. Flammery, M. T. McGuire, E. Geller 1980 Serotonergic influences on the social behavior of vervet monkeys (Cercopithecus aethiops sabaeus). Experimental Neurology 68 (2): 322-34. Raleigh M.J., M. T. McGuire, G. L. Brammer, D. B. Pollack and A. Yuwiler 1991 Serotonergic mechanisms promote dominance acquisition in adult male vervet monkeys. Brain Research 599: 181-190. Rapoport J. L 1989 The Biology of Obsessions and Compulsions. Scientific American 260 (3): 62-69. Rappaport R. A. 1970 The sacred in human evolution. Annual Review of Ecology and Systematics 2: 23-44. Reiner A., L. Medina & C. L. Veenman 1998 Structural and functional evolution of the basal ganglia in vertebrates. Brain Research Reviews 28 (3): 235-285. Ruff C. 2002 Variation in Human Body Size and Shape. Annual Review of Anthropology 31:211-232. Smith W. R. 1889 Lectures on the Religion of the Semites. Edinburgh: Blackwell. Sosis R. & C. Alcorta 2003 Signaling, Solidarity, and the Sacred: The Evolution of Religious Behavior: Evolutionary Anthropology 12: 264-274. Sosis R. & E. R. Bressler 2003 Cooperation and Commune Longevity: A Test of the Costly Signaling Theory of Religion. CrossCultural Research 37 (2): 211-239. Spinoza B. (posthumous; written: ante 1677). Epistularum liber, XIX, (Correspondence, Wolf A. ed., F. Cass). Stuss D. T., G. G. jr Gallup, & M. P. Alexander 2000 The frontal lobes are necessary for “theory of mind”. Brain 124: 279-286. Swanson L. W. 2000 What is the brain? Trends in NeuroScience 23 (11): 519-527. ten Donkelaar H. J. 1998 Reptiles. In: R.. Niewenhuys et al. (eds.), pp. 1315-1524. Tylor E. B. 1871 Primitive Culture. London: J. Murray. van der Leeuw G. 1933 Phänomenologie der Religion. Tübingen Vernon G. M. 1962 Sociology of religion. New York: McGraw-Hill. Vico G. 1744 Scienza nuova terza (3rd ed.; 1st: 1725), Napoli. En. trans.: New Science, 3rd ed. London: Penguin, 1999. Voogd J., R. Niewenhuys. P. A. M. van Dongen and H. J. ten Donkelaar 1998 Mammals. In- R. Niewenhuys et al. (eds.), pp. 1636-2097. Ward C. V., M. G. Leakey, A. Walker 2001 Morphology of Australopithecus anamensis from Kanapoi and Allia Bay, Kenya. Journal of Human Evolution 41:255-368. Weber M. 1904/5 Die protestantische Ethik und der Geist des Kapitalismus. Archiv für Sozialwissenschaft und Sozialpolitik 20/21: ...; Book edition: 1922. Tubingen: Mohr. Weiger W. A. 1998 Serotonergic modulation of behaviour: a phylogenetic overview. Biological reviews of the Cambridge Philosophical Society 72: 61-95. Whybrow P. C. 1999 A Mood Apart. The Thinker’s Guide to Emotion and Its Disorders. New York: HarperPerennial. 2005 American Mania: When More is not Enough. New York: W. W. Norton & Co. Wickler W. 1967 Socio-sexual Signals and their intra-specific Imitation among Primates. In : D. Morris (ed.) Primate Ethology, pp. 69-147. London: Weidenfeld & Nicolson. 1969 Sind wir Sunder? München: D. V. Th. Knaur Nachf. Winkelman M. 1998 Aztec human sacrifice: Cross cultural assessments of the ecological hypothesis. Ethnology 37: 28598. 2000 Shamanism: The Neural Ecology of Consciousness and Healing. Westport: Greenwood. Winkelman M. & J. Baker 2008 Supernatural as Natural. A Biocultural Theory of Religion. Prentice Hall. Widengren G. 1969 Religionsphänomenologie. Berlin: Walter de Gruiter. Wolters J. O., H. J. ten Donkelaar, H. W. M. Steinbusch, A. A. J. Verhofstad 1983 Distribution of serotonin in the brain stem and spinal cord of the lizard Varanus exanthematicus: an immunohistochemical study. Neuroscience 14: 169-193. Wood B. 1994 The oldest hominid yet. Nature 371: 280-1. Woob B. & A. Brooks 1999 Human evolution: We are what we ate. Nature 400: 219. Sources of Figures: they have to be added