Cellular Respiration: From the Department of Redundancy Department.
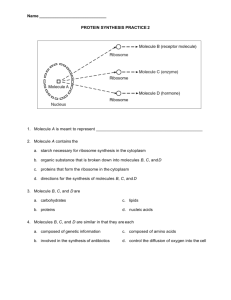
It seems natural for us to think about cellular respiration in the context of our own mammalian
bodies, but to understand respiration in a larger context, it is necessary to step back and consider
plants, fungi, and cellular microbes. In bacteria and archaea, there are no mitochondria - these
organisms have a more streamlined system. However, we find the same molecules involved in cellular
respiration in ALL forms of cellular life, regardless of the particular structural features of the cell. The
commonality and universal nature of the molecules involved is much more significant - really - than
the way the cell is structurally put together.
Anaerobic Respiration: Glycolysis is the anaerobic (without oxygen) process that generates 2 ATP
molecules per glucose molecule. It takes place in the cytoplasm (outside mitochondria). Glycolysis in
your cells and glycolysis in bacterial cells is almost identical, strongly inferring that glycolysis is a
metabolic pathway that was present in the nebulous common ancestor of all life. There are many
bacteria and archaea that never evolved the aerobic (oxygen-requiring) pathways of the Kreb Cycle
and Electron Transport (with ATP Synthase). (Or did they have it at one time then get rid of it?) We
tend to find these anaerobic organisms in extreme environments that might represent environments
similar those of the early earth (before free oxygen was present). For example, in deep ocean
hydrothermal vents we find heat-tolerant microbes that are killed by oxygen. We also find glycolysis
in these same organisms…. the same glycolysis that is happening in almost all of your cells right now.
Gycolysis alone is capable of phosphorylating 2 ATPs per glucose molecule, and these 2 ATPs are
produced really fast. Since the phosphorylation does not involve electron transport, proton gradients,
or ATP synthase, The phosphate transferred to ADP in glycolysis is transferred from another
molecule. Phosphorylation via acquisition of phosphate from an intermediate (substrate) molecule is
called SUBSTRATE-LEVEL PHOSPHORYLATION.
Aerobic Respiration: In aerobic organisms, the most important products of glycolysis are the reduced
electron carrier molecules, NADH. More energy is contained in the NADH molecules reduced in
glycolysis than is contained in the 2 ATPs. The 2 NADH molecules transfer their electrons to the
electron transport chain. Meanwhile….
Preparing pyruvate for the Kreb Cycle: The 2 pyruvate (pyruvic acid) molecules from glycolysis
contain the lion's share of energy from the original glucose molecule. Pyruvate does not enter the Kreb
Cycle directly. It is partially oxidized, releasing a CO2 molecule and generating one more NADH (per
pyruvate… 2 more per glucose) for the ETC. What then remains of the pyruvate is the 2 carbon
molecule, acetate. At this point (we're in the mitochondria in eukaryotes) coenzyme A (CoA) binds to
acetate, forming a complex molecule of acetyl CoA.
Acetyl CoA is the entry point into the Kreb cycle for carbohydrates, fatty acids, and some amino acids.
(Other amino acids enter the Kreb cycle at other points). When the 2-carbon acetyl CoA enters the
Kreb cycle, it first binds to a 4-carbon molecule, oxaloacetate, and the resulting 6-carbon molecule is
citric acid (citrate). Thus, we often refer to the Kreb cycle as the CITRIC ACID CYCLE.
Citric Acid Cycle (CAC): This takes place in the interior of mitochondria, the matrix (or in the deep
interior of bacterial cells). What is left of the food molecules (carbos, proteins, fats) is oxidized all the
way down to CO2, and the energy is transferred to NAD+ and FAD++ (reducing them to NADH and
FADH2). These reduced electron carriers are the most significant molecules produced in the CAC/KC,
but you also get a few ATPs via substrate-level phosphorylation. Oxygen is not directly needed in
the citric acid/Kreb cycle, but if oxygen is absent, the reduced electron carriers will be unable to
unload their electrons onto the ETC, and thus in anaerobic conditions, the CAC will shut down like
one gigantic molecular traffic jam. Total gridlock. Technically, the CAC is aerobic. Note: Oxygen is
the final electron acceptor in the ETC, and without it there to pull off the electrons, the ETC shuts
down, leaving the loaded electron carriers (NADH and FADH2) with no place to dump their electrons.
Thus, the CAC is shut down also.
Electron Transport/Chemiosmosis: Electrons are delivered to the ETC (electron transport chain) by
NADH and FADH2. Recall that we get (from one glucose) 2 NADH molecules in glycolysis, 2 more
from the preparatory reactions, and 6 more from the CAC. So in this scenario, we have 10 NADH
molecules from one glucose molecule. The CAC also reduces 2 molecules of FADH2 (per glucose).
At this point, we have 4 ATPs (2 from glycolysis, 2 from the CAC… again, this is the yield per
glucose molecule), but most importantly we have 10 NADH molecules and 2 FADH2 molecules. The
latter two supply the electrons for electron transport and at this point they are carrying the vast
majority of chemical energy from the original food molecule.
By now, you ought to know that bacteria do not have mitochondria, and yet they go through the CAC
and the ETC (the aerobic bacteria do, anyway). These microbes are all around us and inside us. They
are everywhere. They collectively contain the majority of the planet's genetic diversity. Nevertheless,
we will be good students (and teacher) and proceed with this story of respiration as if it always
happens in mitochondria, because that's the presentation you see in your text. So the story continues
with mitochondria as the setting.
NADH and FADH2 deliver their high energy electrons to the electron transport chain located on the
inner mitochondrial membrane. The CAC reduces these molecules in the mitochondrial matrix, so
they are right where they need to be. The electrons are transferred to ultra-complex, membraneembedded proteins that contain iron components to facilitate the transfer of electrons. Chief among
these molecules are the CYTOCHROMES. Cytochromes are similar in many respects to the
photosystems in the light reactions of photosynthesis. As the electrons are moving through the ETC
from one cytochrome complex to the next, protons are being pumped from the matrix to the intermembrane space. Oxygen is waiting at the end of the ETC and as you know, oxygen is very attractive
to electrons and exerts the most significant pull on electrons. Thus, chemiosmosis in this situation is
called OXIDATIVE PHOSPHORYLATION. At the end of the ETC, you find that beautiful ATP
synthase complex cranking out ATPs from ADP + Pi. This is where most of our ATP comes from.
To summarize…. (all yields are based on one glucose molecule)
Glycolysis - glucose split to 2 pyruvates, 2 ATPs via substrate-level phosphorylation, and 2
NADHs for the ETC.
Preparation of pyruvate: 2 more NADH for the ETC, 2 CO2s released.
CAC - food molecules completely oxidized to CO2, 2 more ATP via substrate-level
phosphorylation, and (per glucose) 6 NADH and 2 FADH2.
ETC - Energy from 10 NADH and 2 FADH2 is used to phosphorylate 32 ATPs!!!!! Then the
oxidized (energy depleted) forms of the molecules (NAD+, and FAD++) go back to the matrix or
cytoplasm to get reduced all over again in the CAC (mostly) or glycolyis.
FERMENTATION: This is another name for anaerobic respiration. ATPs are generated via
glycolysis, and pyruvate is converted to either lactic acid (lactate) or ethyl alcohol (ethanol). Most
significantly, the NADH is oxidized back to NAD+ so that glycolysis can continue to operate.
Fermentation has been exploited by humans in some cases, and as you will see, fermentation in muscle
cells contributes to soreness.
When oxygen is not available, organisms and cells can produce ATP via glycolysis alone at a rate of 2
ATPs per glucose molecule. The problem comes when all of the NADH is in the reduced form.
Fermentation oxidizes the NADH back to NAD+ so glycolysis can continue. In some cells, the
pyruvate is converted to lactic acid (lactate). In other cells (some yeast and bacteria), the pyruvate is
converted to ethyl alcohol (ethanol).
Lactic acid fermentation: Bacteria and muscle cells come to mind. Lactic acid is a preservative in
foods like sour kraut, kim chi, or olives. However, in muscle cells that are called upon to work harder
than they are conditioned to work, lactic acid can build up and contribute to the muscle soreness
associated with strenuous physical activity. Small tears in muscles also contribute to the
aforementioned soreness.
Alcohol fermentation: Some bacteria and yeast. Yeast are unicellular fungi, and prefer oxygen
environments, but will produce ethanol (ethyl alcohol) as a toxic by-product. As you may know, this is
a natural process, and humans have been manipulating nature for millennia to get more alcohol from
wine juice, apple juice, or grainy soup. Wine and other alcoholic beverages go back to the earliest
recorded history.
Nowadays, scientists are manipulating the process more than ever before because ethanol is a viable
fuel for cars and trucks. Ethyl alcohol is toxic to cells. It is the active ingredient in hand sanitizers
because it dissolves cell membranes. Naturally, yeast cells are deactivated with the ethanol level
reaches about 5%. Humans have been selecting for strains of yeast that have higher tolerance for
alcohol. So far, we've been able to select for strains that can tolerate ethanol levels up to 20% or more.
As you might imagine, the biofuel industry prefers higher ethanol content. Most gasoline these days
has 10% ethanol added to it. Pure ethanol has to be distilled from the fermented carbohydrates. This
is relatively simple since it boils at 80 C (compared to 100 C for water).
 0
0