Sordaria Lab
advertisement

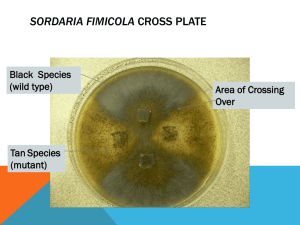
Sordaria fimicola Genetics Sordaria fimicola, a common species of dung ascomycete, produces eight ascospores in a linear arrangement within an ascus, a bag-shaped, spore-containing structure. The ascospores are products of meiosis. Hybrid asci resulting from hyphal fusion between wild-type and mutant-tan (or gray) strains will contain four ascospores of each color. Their arrangement will allow determination of crossing over and to calculate the gene to centromere distance. Procedures Preparation of Stock Cultures 1. Disinfect the work surface and wash hands. 2. When ready for use, label two of the cornmeal-glucose-yeast agar dishes “wild,” two “gray”, and two “tan.” 3. Using aseptic technique, inoculate the dishes with the appropriate culture. Remove the top from the tube of the wild-type Sordaria fimicola, and flame the mouth over a Bunsen burner for a few seconds. With a flamed, cooled scalpel or spear point needle, remove a portion of the culture containing perithecia (black pepper grain appearance) and transfer to the middle of a cornmeal-glucose-yeast agar dish. Repeat this procedure to prepare another wild-type culture. 4. Using the other tubes, follow step 3 to prepare two gray and two tan stock culture dishes. 5. Incubate the dishes for 5 to 7 days out of direct sunlight at room temperature (22° - 25°C) until perithecia have formed at the periphery of the dishes. Preparing the Crosses 1. Disinfect the work surface and wash hands. 2. Label one half of the Sordaria crossing agar dishes “+/g” and the other half “+/tn” to indicate crosses between the wild-type and mutant gray (or tan) strains. 3. Divide the dishes into quarters and use a wax pencil or permanent marker, indicate the positions of wild type (+) and gray (g) or tan (tn) cultures. The different strains should be located opposite one another on the culture dish. 4. Using a flamed, cooled, scalpel or spear point needle, cut the agar in the stock culture dishes into 0.5 cm cubes. Place the cubes upside down over the indicated positions on the surface of the crossing agar. Each plate will contain two blocks of the wild-type culture and two blocks of either tan or gray culture. 5. Incubate the dishes out of direct sunlight and at room temperature. 6. From 8 days after inoculation until forcible discharge of the spores, genetic data can be obtained. Usually, the cultures should be ready for microscopic examination in 8 to 10 days, but at cooler temperatures, 14 to 15 days may be required. In order to obtain accurate data, it is essential that mature ascospores be counted. If it is difficult to distinguish microscopically between the wild-type and gray or tan spores, the ascospores are too immature to collect data. Incubate the cross dishes for another day or two and observe again. Microscopic Examination 1. Disinfect the work surface and wash hands. 2. Remove a few perithecia from the cross dishes with a flamed, cooled loop and prepare a wet mount. Note from which cross plate (“+/tn” or “+/g”) you are removing the perithecia. 3. Press the cover slip gently using the thumb or an eraser to crush the perithecia and release the rosettes of asci. If too much pressure is applied, the ascospores will be forced out of the asci, making it impossible to collect data. 4. Using low power, examine the slide and locate the rosettes of hybrid asci containing ascospores of two different colors. The wild-type ascospores appear black, while the gray and tan spores are a lighter color. 5. After locating a rosette of hybrid asci, use high power to observe the ascospores and determine is crossing-over has occurred. If crossing over has not occurred, segregation of the genes for spore color has taken place during meiosis I (MI) and the ascospores will be arranged in a 4:4 ratio. If crossing over has occurred, segregation of the genes for spore color do not segregate until meiosis II (MII) and the arrangement of ascospores will be either 2:4:2 or 2:2:2:2. 6. Each group should count 100 to 200 asci. Collate class data. 7. Chromosome maps for the two mutant genes are constructed by dividing the %MII by 2. Strains Crossed No. of MI Asci (4:4) No. of MII Asci (2:4:2 or 2:2:2:2) Total Asci %MII (No. MII/total) Map Units %MII/2 (g) x (+) (tn) x (+) Discussion Sordaria fimicola belongs to the fungal class of ascomycetes, or sac fungi. These fungi produce sexual spores called ascospores, borne in a bag-shaped structure, the ascus. The asci are contained in a flask-shaped structure, the perithecium, which has a small opening at the apex, the ostiole. The cavity of the perithecium is lined with sterile tissue, the periphyses. Sordaria is found in nature on herbivore dung. It is a coprophilous, or dung-loving, fungus. It exhibits several characteristics of these fungi, which typically pass through an herbivore’s digestive system during a portion of the life cycle: 1. The ascospores are a dark color to protect them from ultraviolet light damage. 2. The perithecia demonstrate a phototrophic response to light. The neck of the perithecium bends toward the light, and the spores are discharged toward the light. 3. Spores are explosively discharged to move them beyond the “zone of repugnance” (or dung heap), because herbivores will not graze near their own dung. 4. The spores are mucilangenous so that they adhere to the vegetation impacted. When the mucilage dries, the spores firmly attach to vegetation, where they remain until consumed by an herbivore. Sordaria reproduces sexually only. It has lost the ability to reproduce asexually. It is homothallic; it reproduces sexually with itself. A haploid ascospore germinates producing a monokaryotic (“one nucleus”) hypha. The nucleus divides by mitosis and these nuclei distribute themselves in the growing hypha. Septa (cross walls) are laid down and branching occurs. At certain points, the mycelium forms a coiled ascogonium, the female sex organ. From the same mycelium, an antheridium, the male sex organ, forms. The ascogonium forms a long, hair-like hypha, the trichogyne. The trichogyne grows until it contacts the antheridium. The wall of the antheridium dissolves, and its contents enter the ascogonium. Plasmogamy (the fusion of the cell contents except the nucleus) occurs, and an ascogenous hypha forms from the ascogonium. The nuclei from the ascogonium and antheridium divide repeatedly by mitosis, pair up, and migrate into the ascogenous hyphae. Septa are laid down so that each cell contains one nucleus from the ascogonium and one from the antheridium. The ascogenous hyphae is dikaryotic; it contains one haploid nucleus from each parent. A tip of the ascogenous hyphae curves over, forming a crozier or “shepherd’s crook” with two nuclei. Each nucleus divides by mitosis. Septa are formed to divide the crozier into three cells. The uppermost cells contain two nuclei, one from each parent. These nuclei fuse (karyogamy) to form a diploid cell. The nucleus immediately undergoes meiosis I and meiosis II, resulting in four haploid cells. Another mitosis follows, resulting in eight haploid ascospores. This process is repeated many times to form the asci contained within the perithecium. The sterile tissue from the old ascogonium forms the perithecium wall and the periphyses. The order of ascospores in the ascus reflects the order in which the chromosomes are segregated during meiosis. This can be clearly visualized if the diploid nucleus is a hybrid for strains with two different spore coat colors. The wild-type strain (+) produces a dark spore, while mutant-tan (tn) or gray (g) strains produce lighter-colored spores. If crossing-over does not occur, these genes segregate during meiosis I to produce a 4:4 sequence of ascospores. However, if crossing-over did occur, the genes do not segregate until meiosis II to produce a 2:2:2:2 or a 2:4:2 sequence of ascospores. Since the rate of crossing-over increases with the distance between two points on a chromosome, it is possible to use the rate of crossing to determine the relative distance from gene locus to centromere. In Sordaria, each meiosis II ascus that exhibits crossing-over represents a tetrad of chromatids where there has been an exchange involving only two of the four chromatids. Therefore, the percentage of meiosis II asci must be divided by two to determine gene-to-centromere distance in map units. Extensive research has shown that the gene for tan spores is about 26 map units from the centromere, while the gene for gray spores is about 60 units. However, because the incidence of double crossover increases with the distance between any two points on a chromatid, distances greater than 33.3 map units cannot directly be calculated for Sordaria with the method used here. Therefore, a value of about 33.3 map units should be expected for the gene for gray spores. Questions: 1. Can a haploid cell undergo meiosis? Can it divide by mitosis? 2. Why do you expect the diploid number of chromosomes always to be an even number and never an odd number? 3. How does crossing over contribute to genetic variability? Does this have any evolutionary significance? 4. Using a Chi-square, determine if the crossover frequency found in your Sordaria culture matching that reported in the literature. Source: Sordaria Genetics Kit 15-5848. 1999. Carolina Biological Supply Company. North Carolina, USA.