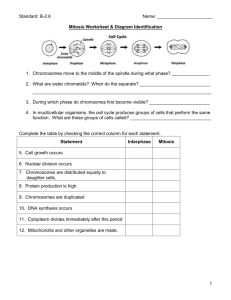
UNIT 1: INTRODUCTION TO BIOLOGY
advertisement

SAN DIEGO MESA COLLEGE SCHOOL OF NATURAL SCIENCES General Biology (BIOL 107); Instructor: Elmar Schmid, Ph.D. Chapter 8: Growth, cell division, chromosomes, cell cycle & Mitosis - Part I INTRODUCTION Growth and development are one of the hall mark features and characteristics of all forms of life on planet Earth In order for single-celled organisms, such as bacteria or protests to increase in numbers or for multi-cellular biological organism to grow in size, its cells have to multiply to increase the amount of cells, roots, stem, tissue, organ and other body materials Multi-cellular organisms, such as fungi, plants or animals grow, because their cells exponentially increase in numbers by repeating a very coordinated and synchronized cellular program, called the cell cycle, in a process called cell division In this (first) part of chapter 8 we will therefore have to have a closer look at the cellular basis of growth to fully appreciate this amazing biological process HISTORY long before man knew about the existence of genes, chromosomes and the underlaying principles of cell growth and inheritance, so-called breeding was done with life stock or certain plants to bring out a wished phenotype e.g. pigs with more meat mass or corn with larger cobs 1865: the Augustinian monk Gregor Mendel publishes his ‘Principles of Heredity’, which he discovered while conducting his famous breeding experiments with the common garden pea without knowing about the substance of genes he speaks already of ‘heritable factors’ at the end of the 19th century, the German physician Rudolf Virchov states that ‘all cells come from cells’ and that cellular growth and reproduction requires a process called cell division Mendel’s principles were not anticipated and understood before cell biologists worked out and discovered the cellular process of mitosis and meiosis on more detail Around 1910, scientists developed the chromosome theory of inheritance which states that the heritable factors (= genes in modern terminology) which are responsible for the expression of heritable traits are located on chromosomes 1 SAN DIEGO MESA COLLEGE SCHOOL OF NATURAL SCIENCES General Biology (BIOL 107); Instructor: Elmar Schmid, Ph.D. The Cell division & The cell cycle in order to for an organism to grow cell division is necessary cell division is the biological process of splitting of a unique form of cell, a so-called stem cell, into two identical daughter cells usually after stimulation of the parent cell with so-called growth factors - important growth factors are peptide molecules such as EGF, PDGF or NGF - they bind to antennae-like, so-called receptor proteins located on the surface of the cell some cells in our human body divide continuously to form new cells, such as: 1. Myelogenic stem cells in our bone marrow to form new red blood cells, platelets and immune cells 2. Keratinocytes and fibroblasts in our skin to form new protective skin layers 3. Osteoblasts in our bone marrow to build up new calcified bone material some cells start to divide only after loosing their cell-to-cell contact with neighboring cells, e.g. after tissue or cell damage e.g. liver cells after intoxication, injury or after surgical removal e.g. skin fibroblasts after injury Most cells of biological organisms, after reaching the adult stage, do not divide at all e.g. nerve cells in our brain before a cell divides it duplicates (= copies) or enlarges all of its cellular constituents, including organelles, membranes, cytoskeleton Especially the exact copying (= replication) and separation of the cell’s chromosomal genetic material in identical portions into each of the two new cells is of prime importance to assure a loss or disproportion of genetic material during growth The highly coordinated and orchestrated process of cell division assures that cells, especially cells with large amount of genetic material, such as eukaryotic cells, will produce new daughter cells with equal amounts of chromosomal material as the parental cell e.g. a dividing human cell has to copy and handle chromosomal DNA comprised of 6 billion base pairs!! Certainly not an easy task! Biological organisms with less genetic material, such as bacteria, perform a less complex cell division process called binary fission which we will look up in the last section of this chapter 2 SAN DIEGO MESA COLLEGE SCHOOL OF NATURAL SCIENCES General Biology (BIOL 107); Instructor: Elmar Schmid, Ph.D. In non-dividing eukaryotic cells, e.g. a human cell, genetic material appears as socalled chromatin fibers located within the nucleus of the cells Electron microscopic picture of chromatin chromatin fibers consist of DNA and certain proteins, called histones DNA strands are wrapped around the histone proteins these DNA/protein structures, or also called nucleosomes, can be seen through an electron microscope when a cell prepares to divide, the cellular chromatin fibers start to fold and coil to end up forming a X-shaped chromosome structure a chromosome is a cellular structure which contains the genes for genetic traits of biological organisms; it is made up of very long strands of chromatin, which (after heavy folding and coiling) during certain stages of the cell cycle (see sections ‘mitosis’ and meiosis’ below) can be seen with the help of a light microscope the total cellular DNA of a species is called the genome, which is split into a certain number of chromosomes the DNA of the complete genome is visible as a distinct number of chromosomes which is unique for each species e.g. human cells have 46 chromosomes which are visible during certain stages before and along cell division (see Graphic below) chromosomes are visible only during certain stages of the cell cycle (see sections ‘mitosis’ and meiosis’ below) the total cellular DNA of a species (= genome) is split into a certain number of chromosomes the DNA of the complete genome is visible as a distinct number of chromosomes which is unique for each species e.g. human cells have 46 chromosomes which are visible during certain stages before and along cell division 3 SAN DIEGO MESA COLLEGE SCHOOL OF NATURAL SCIENCES General Biology (BIOL 107); Instructor: Elmar Schmid, Ph.D. TThhee 4466 hhuum maann cchhrroom moossoom meess the process which is responsible for the complete duplication of the whole cellular genetic material is called DNA replication ( see Chapter 10 for more details) the whole set of DNA or the genome is duplicated shortly after DNA replication sets of X-shaped chromosomes become visible within eukaryotic cells chromosomes become only visible to us shortly before a cell enters mitosis the chromosomes can be seen through a light microscope as X-shaped bodies which show a typical banding pattern (so-called G-bands) after staining with certain dyes (see Graphic below) each chromosome is formed by two so-called sister chromatids, each sister chromatid contains genes coding for identical traits at the same spot (= locus) two sister chromatids are joined together in the so-called centromere region, which is the most narrow part of the of a metaphase chromosome the centromere plays an important role in the separation of the two sister chromatids during anaphase of mitosis and meiosis; 4 SAN DIEGO MESA COLLEGE SCHOOL OF NATURAL SCIENCES General Biology (BIOL 107); Instructor: Elmar Schmid, Ph.D. Structure and important regions of a typical metaphase chromosome Coiled euchromatin (DNA + histones) Centromere Sister Metaphasechromosome Sister Centromeric heterochromatin Telomeric end eukaryotic chromosomes are much more complex than prokaryotic chromosomes eukaryotic chromosomes contain so-called scaffolding proteins which help to organize and maintain the complex 3-dimensional chromosome structure some of these proteins are play a role in the control of gene activity (see Chapter 10) the degree of folding of coiling of chromatin fibers is not equal within a chromosome; scientists discovered areas which are made up form highly compacted chromatin, the so-called heterochromatin regions, and more loosely arranged, open chromatin regions, the so-called euchromatin regions (see Graphic above) each eukaryotic chromosome consists of one long DNA double helix which codes for thousands of genes a gene is a segment on the DNA strand of the genome which codes for a distinct protein or enzyme the long DNA double helix of each eukaryotic chromosome codes for thousands of genes the average gene is about 1000 nucleotide base pairs long almost all genes which make up an eukaryotic organism are found in the cell nucleus some genes are located on the so-called extra-chromosomal DNA which is located in mitochondria 5 SAN DIEGO MESA COLLEGE SCHOOL OF NATURAL SCIENCES General Biology (BIOL 107); Instructor: Elmar Schmid, Ph.D. eukaryotic chromosomes contain much more genes and are much more complex than prokaryotic chromosomes e.g. a human cell has about 35,000 – 40, 000 genes, while the genome of a bacterium harbors about 3000 genes eukaryotic chromosomes contain proteins which help to organize the complex 3-dimensional (X-shaped) structure some of these proteins are play a role in the control of gene activity the sequence of nucleotides (see Graphic below) or the so-called letter code which makes up a gene, determines the later shape and function of the gene product the gene product can either be a protein, which helps to build up the cell structure or an enzyme, which regulates essential part of the cell’s biochemical pathways TThhee D DN NA A sseeqquueennccee ooff aa ttyyppiiccaall ggeennee (= gene of the human enzyme superoxide dismutase) SOURCE human. ORGANISM Homo sapiens Eukaryota; Metazoa; Chordata; Craniata; Vertebrata; Euteleostomi; Mammalia; Eutheria; Primates; Catarrhini; Hominidae; Homo. REFERENCE 1 (bases 1 to 560) AUTHORS Sherman,L., Dafni,N., Lieman-Hurwitz,J. and Groner,Y. TITLE Nucleotide sequence and expression of human chromosome 21-encoded superoxide dismutase mRNA JOURNAL Proc. Natl. Acad. Sci. U.S.A. 80 (18), 5465-5469 (1983) BASE COUNT 158 a ORIGIN (human) 108 c 160 g 134 t bp1 A ATTG GGCGACGA AGGCCGTGTG CGTGCTGAAG GGCGACGGCC CAGTGCAGGCATCATCAATTTCGAGCAGA AGGAAAGTAA TGGACCAGTG AAGGTGTGGGAAGCATTAA AGGACTGACTGAAGGCCTGC ATGGATTCCTGTTCATGAG TTTGGAGATAATACGGCAGCTGTACCAGTGCAGGTCCTCACTTTAATCCTCTATCCAGAAAAC ACGGTGGGCCAAAGGATGAAGAGAGGCATGTTGGAG ACTTGGGCAA TGTGACTGCT GACAAAGATG GTGTGGCCGATGTGTCTATTGAAGATTCTG TGATCTCACTCTCAGGAGACCATTGCATCATTGGCCGCACACTGGTGGTCCATGAAAAAGCA GATGACTTGGGCAAAGGT GGAAATGAAGAAAGTACAAAGACAGGAAACGCTGGAAGTC GTTTGGCTTG TGGTGTAATT GGGATCGCCCAATAAACATT CCCTTGGATGTAGTCTGAGG CCCCTTAACT CATCTGTTAT CCTGCTAGCT GTAGAAATGT ATCCTGATAAACATTAAACA CTGTAATCTT bp561 (from: NIH/NCBI Entrez Nucleotide data base) Nucleotide abbreviation: A = Adenine T = Thymine G = Guanine C = Cytosine ATG = Start codon 6 SAN DIEGO MESA COLLEGE SCHOOL OF NATURAL SCIENCES General Biology (BIOL 107); Instructor: Elmar Schmid, Ph.D. almost all genes which make up an eukaryotic organism are found in the cell nucleus e.g. a human cell has about 100,000 genes, while a bacterium harbors about 3000 genes some genes are located on the so-called extra-chromosomal DNA which is located in mitochondria or chloroplasts Cell division of somatic cells in multi-cellular organisms not only plays a role in normal cell growth and cell repair, but also in a unique form of reproduction, so-called asexual reproduction, which leads to new daughter organisms (or organism body parts) with identical DNA molecules (= genotype) and identical appearance (= phenotype) many biological organisms, such as fungi and certain plants, rely on this form of reproduction to quickly thrive in relatively stable environments some animals, such as lizards, can replace lost body parts, e.g. a tail, regrowing a new one During each round of cell division, a cell runs through a clock work-like program, the so-called cell cycle which steers and coordinates all important cellular events while the parent cell prepares and executes its division into to equal daughter cells Today, we begin to understand the enormously intricate and complex nature and involved components of the cell cycle - the cell cycle is dependent on many proteins, so-called cyclins, and enzymes, such as the cyclin-dependent kinases - the cell cycle is a strictly controlled biological clock work which involves many proteins such as p53 and Rb (for more details on the cell cycle see separate chapter) C Ceellll ccyyccllee & &M Miittoossiiss Whenever eukaryotic cells decide, i.e. become the signal, to divide, they undergo an orderly sequence of events which is also called a cell cycle a cell cycle spans from the time a cell divides to form a daughter cell to the time point this daughter cell itself starts to divide again 7 SAN DIEGO MESA COLLEGE SCHOOL OF NATURAL SCIENCES General Biology (BIOL 107); Instructor: Elmar Schmid, Ph.D. TThhee ddiiffffeerreenntt pphhaasseess ooff tthhee cceellll ccyyccllee G0 - performance of specific cell functions e.g. protein production, metabolism 8 SAN DIEGO MESA COLLEGE SCHOOL OF NATURAL SCIENCES General Biology (BIOL 107); Instructor: Elmar Schmid, Ph.D. The cell cycle consists of two major stages: 11.. IInntteerrpphhaassee a cell spends most of the time of the cell cycle (90%) in this stage the interphase is subdivided into the three sub-phases: G0-phase - the resting, non-dividing cell which performs the “routine” metabolic or cellular activities, e.g. enzyme or hormone synthesis G G11--pphhaassee S S--pphhaassee G G22--pphhaassee the cell has a high metabolic activity to be able to perform the various, “normal” functions G G11--pphhaassee: (‘G’ refers to the gap between cell division and the occurrence of DNA synthesis) upon activation by an out-side trigger, e.g. a docking growth factor at a surface receptor, the cell increases its supply of proteins and increases the number of its organelles (mitochondria, golgi, peroxisomes, etc.) the cell grows in size S S--pphhaassee: - DNA synthesis (= DNA replication) occurs and the cell duplicates its complete set of Chromosomes (for details see separate Chapter 10-1) - the cell synthesizes most of the proteins and enzymes it needs for DNA synthesis during the S-phase, e.g. histones, DNA polymerase, Helicase, etc. - at the end of the S-phase two sister chromatids are formed by the cell - the transition from S to G2 marks the so-called mitotic checkpoint of the cell cycle - many regulator or check point proteins, e.g. the breast cancer susceptibility gene product BRCA-1 control at this point of the cell cycle G G22--pphhaassee: - the cell starts to synthesize proteins which are essential for cell division and shows a high metabolic activity again - especially proteins needed for the following M-phase, such as tubulin, actin, are produced in high amounts • By late interphase, the chromosomes have been duplicated but are loosely packed. • The centrosomes have been duplicated and begin to organize microtubules into an aster (“star”). 9 SAN DIEGO MESA COLLEGE SCHOOL OF NATURAL SCIENCES General Biology (BIOL 107); Instructor: Elmar Schmid, Ph.D. M Miittoottiicc oorr M M -- pphhaassee at the end of the G2-phase the cell installed a second checkpoint, which regulates the entering into the M-phase; so-called key-proteins of this important cellular checkpoint is the CDC25B protein and the so-called cyclins the mitotic phase is subdivided into 2 processes M Miittoossiiss Duplicated chromosomes (= sister chromatids) divide and are evenly distributed into two daughter cells C Cyyttookkiinneessiiss the cytosol is divided two; two identical daughter cells are formed Let’s have a closer look at the very dynamic cellular processes which occur during mitosis M Miittoossiiss Even though mitosis is continuous, dynamic cellular process, 5 phases are usually distinguished by biologists 10 SAN DIEGO MESA COLLEGE SCHOOL OF NATURAL SCIENCES General Biology (BIOL 107); Instructor: Elmar Schmid, Ph.D. The 5 phases of mitosis 1. Prophase Summary of changes which occur in the nucleus and in the cytosol: N Nuucclleeuuss:: the chromatin fibers become highly coiled and folded (= DNA condensation) (see Graphic below) discrete chromosomes can be seen; each duplicated chromosome appears as two identical ssiisstteerr cchhrroom maattiiddss the nuclear envelope breaks apart m miiccrroottuubbuulleess originating at the centrosomes at both poles reach the chromosomes and make contact with the kinetochore of the centrosomes chromosomes are moved to the center of the cell by forces exerted by ATPconsuming protein motors C Cyyttoossooll:: the protein-made mitotic spindle apparatus, made of hollow microtubules begins to form at the so-called MTOC which contains two centrioles microtubules grow out of from the MTOC by polymerization the 2 centrosomes move to the opposite poles 11 SAN DIEGO MESA COLLEGE SCHOOL OF NATURAL SCIENCES General Biology (BIOL 107); Instructor: Elmar Schmid, Ph.D. The way from DNA, chromatin to a metaphase chromosome (Understanding DNA condensation during prophase of mitosis) 12 SAN DIEGO MESA COLLEGE SCHOOL OF NATURAL SCIENCES General Biology (BIOL 107); Instructor: Elmar Schmid, Ph.D. 2. Pro-Metaphase • During prometaphase, the nuclear envelope fragments and microtubules from the spindle interact with the chromosomes. The microtubular spindle apparatus is beginning to form. • Microtubules from one pole attach to one of two kinetochores, special regions of the centromere, while microtubules from the other pole attach to the other kinetochore. 3. Metaphase The m miiccrroottuubbuullaarr ssppiinnddllee aappppaarraattuuss is fully formed. Chromosomes convene in line on the (virtual) metaphase plate in the center of the cell. For each chromosomes the kinetochores of the two sister chromatids face opposite poles each replicated chromosome possesses two discrete "sister" kinetochores that are positioned on the opposite sides during mitosis, sister kinetochores firmly attach to the centromere region of their associated chromosome they are important in the formation of the spindle apparatus by capturing the ends of the microtubules arriving from the MTOCs of the cell poles a kinetochore in vertebrate cells is a disk-shaped protein structure, which is composed of three distinct plate-like domains 13 SAN DIEGO MESA COLLEGE SCHOOL OF NATURAL SCIENCES General Biology (BIOL 107); Instructor: Elmar Schmid, Ph.D. 14 SAN DIEGO MESA COLLEGE SCHOOL OF NATURAL SCIENCES General Biology (BIOL 107); Instructor: Elmar Schmid, Ph.D. Mitosis & Cancer Treatment Many plant-derived molecules, such as colchicine, taxol or vinblastine, have been shown to block mitosis at this stage; they work as so-called spindle poison since they interfere with the formation or disassembly of the microtubules and arrest a dividing cell in this stage (see Graphic below) Taxol stabilizes the microtubule fibers by blocking its degradation (= depolymerization); it “freezes” a mitotic cell in metaphase as a consequence, the chromosomes cannot migrate toward the cell poles Taxol has been approved by the FDA for treatment of certain forms of cancers in humans, such as mammary carcinomas and ovarian cancer Two examples of known mitosis-blocking molecules 1. Taxol Isolated from the Pacific Yew tree 2. Laulimalide Isolated from the marine sponge Fasciospongia rimosa - mitosis blocker which kills cancer cells by blocking mitosis & triggering apoptosis - binds to polymerized tubulin and prevents the disassembly of microtubules - also binds to bcl-2 and prevents its anti-apoptotic - inhibits many different cancer cell types - is even active against cancer cells that are resistant to Taxol “Multi-drug-resistant cell types” cellular function A closer look at the microtubular spindle apparatus • The mitotic spindle, fibers composed of microtubules and associated proteins, is a major driving force in mitosis. • As the spindle assembles during prophase, the elements come from partial disassembly of the cytoskeleton. The spindle fibers elongate by incorporating more subunits of the protein tubulin. • Assembly of the spindle microtubules starts in the centrosome (or microtubuleorganizing center) which is a large protein complex which - in animals and human cells - has a pair of centrioles at the center. 15 SAN DIEGO MESA COLLEGE SCHOOL OF NATURAL SCIENCES General Biology (BIOL 107); Instructor: Elmar Schmid, Ph.D. • As mitosis starts, the two centrosomes are located near the nucleus. As the spindle fibers grow from them, the centrioles are pushed apart. • By the end of prometaphase they develop as the spindle poles at opposite ends of the cell. • During prometaphase, some spindle microtubules attach to the kinetochores located in the centromere region of each sister chromatid. A kinetochore consists of proteins and chromosomal DNA. • The kinetochores of the joined sister chromatids face in opposite directions. • When a chromosome’s kinetochore is “captured” by a “kinetochore” microtubule, the chromosome moves toward the pole from which those microtubules come from. • When microtubules attach to the other pole, this movement stops and a tug-of-war ensues. Eventually, the chromosome settles midway between the two poles of the cell, the metaphase plate. • Other microtubules (the polar microtubules) from opposite poles interact with each other, therefore elongating the cell. 16 SAN DIEGO MESA COLLEGE SCHOOL OF NATURAL SCIENCES General Biology (BIOL 107); Instructor: Elmar Schmid, Ph.D. 3. Anaphase • The ssiisstteerr cchhrroom maattiiddss are separated into full-fledged daughter chromosomes. At anaphase, the centromeres divide, separating the sister chromatids. • Each sister chromatid is now pulled toward the pole to which it is attached by spindle fibers. By the end, the two poles have equivalent collections of chromosomes. • The microtubules forming the spindle apparatus elongate or shorten the microtubules which make contact with the kinetochores of the sister chromatids (= kinetochore microtubule) start to shorten (= depolymerization) and perform a force in the direction of the MTOCs located at the cell poles the microtubules coming from the opposite poles and which make contact with each other (= polar tubules), continue to elongate; they contribute to beginning cell stretching • According to one favored hypothesis, ATP-powered motor proteins are responsible for the directed movement of the sister chromatids in anaphase. These motor proteins located at the kinetochore “walk” the attached chromosome along the microtubule toward the opposite pole. The excess microtubule sections depolymerize. 17 SAN DIEGO MESA COLLEGE SCHOOL OF NATURAL SCIENCES General Biology (BIOL 107); Instructor: Elmar Schmid, Ph.D. - The cell poles are moving farther apart and the cell begins to elongate TThhee iim mppoorrttaanntt rroollee ooff m miiccrroottuubbuulleess dduurriinngg aannaapphhaassee 18 SAN DIEGO MESA COLLEGE SCHOOL OF NATURAL SCIENCES General Biology (BIOL 107); Instructor: Elmar Schmid, Ph.D. Telophase and cytokinesis - The sister chromatids arrived at the MTOC (centrosomal region) positioned at the cell poles cell elongation continues and daughter nuclei appear at the cell poles during telophase the so-called ‘cleavage furrow’ forms with the help of the actin cytoskeleton; an actin ring forms under the cell membrane this process helps to pinch the parental cell into two daughter cells Cleavage furrow Actin ring - the ring of contracting microfilaments made up of the protein monomer actin forms a groove in the cell surface and begins to pinch the cell into two daughter cells (= cytokinesis) - at the end of cytokinesis finally two daughter cells arise Mitosis & Plant cells Due to the rather rigid cell wall which surrounds plant cells, mitosis in these cells does not show a cleavage furrow and shows different cytokinesis events. Cytokinesis in ppllaanntt cceelllss occurs by forming a so-called cell plate. Each cell plate is formed in the middle of the plant cell by fusion of many membranous vesicles which are filled with cell wall-material. As more vesicles fuse together the cell plate grows outwards and its membranes finally fuse with the plasma membrane. The cell plate content joins the parental cell wall. 19 SAN DIEGO MESA COLLEGE SCHOOL OF NATURAL SCIENCES General Biology (BIOL 107); Instructor: Elmar Schmid, Ph.D. At the end of mitosis the chromatin fibers of each sister chromatid uncoil they turn into microscopic invisible chromosomal chromatin fibers Mitosis is unique to eukaryotic organisms Mitosis is a cellular mechanism in eukaryotes which guaranties the equal distribution of identical copies of the large amounts of genetic (= DNA) material the genome is portioned into a definite number of chromosomes Mitosis is the evolutionary solution to the problem of allocating identical copies of large amounts of genetic (= DNA) material into two new cells the genome is portioned into a definite number of chromosomes, which the eukaryotic cell duplicate and separate in an extremely accurate mechanism called mitosis e.g. in yeast, errors in chromosomal distribution occur only once in 100,000 cell divisions!! 20 SAN DIEGO MESA COLLEGE SCHOOL OF NATURAL SCIENCES General Biology (BIOL 107); Instructor: Elmar Schmid, Ph.D. Cell division, cell cycle & Control cell division and the events which enable the cell to enter mitosis are tightly controlled the cell division events are controlled by installation of two so-called mitotic check points along the cell cycle two check points are positioned at: the entry into the S-phase to control DNA replication the entry into the M-phase to control chromosomal condensation and the mitosis events a protein complex, called maturation promoting factor (MPF) is involved in M-phase control Several factors influence the onset and accurate timing of cell division and cell growth 1. Growth factors (GF) amino acid-made growth factors, such as epithelial-derived growth factor (EGF) or fibroblast-derived growth factor (FGF) or platelet-derived growth factor (PDGF), are the main signals for the cell to promote cell division and are produced by specialized cells within the body there are so-called autocrine (= stimulate the same cell) and paracrine (stimulate surrounding or distant cells) growth factors known growth factors dock on so-called receptor proteins on their target cells and trigger a so-called signaling cascade (see also Chapter 10 for more details) at the end of this cascade the nucleus receives the signal of the stimulated cell the cell enters the cell cycle and cell division begins they trigger the sequential events of the cell cycle by activating a set of proteins in the cell which are part of the so-called cell cycle control system and which control the so-called check points of the cell cycle the three key check points of the cell cycle are located in the G1, G2 and Mphase; at these points the cell build in brakes that block the cell cycle from running; therefore a cell remains non-dividing or quiescent in the so-called Go-phase until it receives a go-ahead signal by a docking growth factor 2. Contact- or anchorage-dependent cell division certain cells don’t divide in suspension and need a solid surface onto which they can adhere e.g. fibroblasts need contact with so-called extracellular matrix proteins, such as collagen or fibronectin, to be able to divide many of these extracellular matrix proteins recognize structures (= receptors) on the surface of the so-called adherent cell usually cells show so-called density-dependent growth inhibition; they stop growing after getting in close contact with other cells cells growing in cell culture usually stop growing after they formed a so-called monolayer = each cell got in contact with another cell 21 SAN DIEGO MESA COLLEGE SCHOOL OF NATURAL SCIENCES General Biology (BIOL 107); Instructor: Elmar Schmid, Ph.D. under certain conditions cell growth can become dys-regulated and may lead to development of cancer cancer cells usually show a loss of cell cycle control due to mutations of components of the cell signaling cascade or of members of the cell cycle control system this leads to an uncontrolled cell division and an excessive formation of cells, or also called a tumor most tumors are benign (= friendly) and its cells remain at the same site where the tumor formation started; they can be successfully removed from the body by surgery in so-called malignant tumors, the cells leave the tumor formation and spread into other tissues or parts of the body in a process called metastasis; malignant cancers are difficult to treat and the outcome for the cancer patient is usually fatal different kind of cancers are classified dependent on the site of the body where they originated 1. carcinomas originate in the exterior or interior coverages of the body e.g. skin ( melanoma) or intestine ( colon cancer) 2. sarcomas originate in tissues which support the body e.g. bone ( osteosarcoma) muscle or connective tissue 3. leukemias and lymphomas originate in cells of the blood forming system (in the bone marrow, spleen and lymphnodes) e.g. B-lymphocytes cancer cells usually are not prone to cell-density dependent growth inhibition (= when cells get in touch with another cell) or also called contact-inhibition they therefore reach high cell densities and, when grown in cell cultures, pile up in so-called foci many cancer cells bear mutations within their genetic code and express mutated proteins or enzymes e.g. p53 protein, BCRA-1, ras protein several anti-cancer therapies are able to halt the cancer cells from spreading 1. Radiation therapy the affected tissue is irradiated with strong electromagnetic waves which mostly destroy the fast-growing cancer cells, while non-dividing, normal cells are not affected 2. Chemotherapy uses drugs that disrupt cell division by inhibiting proteins or enzymes which play an important role in cell activation or cell cycle regulation 22 SAN DIEGO MESA COLLEGE SCHOOL OF NATURAL SCIENCES General Biology (BIOL 107); Instructor: Elmar Schmid, Ph.D. most of them are so-called antimitotic drugs isolated from plants which interfere with the formation of the mitotic spindle apparatus e.g. vinblastine taxol colchicines 3. Antisense therapy short pieces of DNA or RNA (ribozymes) which base-pair with the mRNA of the mutated gene and prevents further expression of the cancer-causing protein this future anti-cancer strategy is very selective and is expected to have less side-effects for the patient 4. Gene therapy also there is no ‘magic bullet’ for fighting cancer in near sight, this future cancer therapy holds great promise and has the goal to correct the mutated and cancer-causing gene within the patients cells attenuated viruses e.g. adeno-associated virus are expected to shuttle the corrected gene into the affected cancerous tissue of the patient B Biinnaarryy FFiissssiioonn Prokaryotic cells, i.e. bacteria and most protists divide by a much faster and less comlicated biological process called binary fission the prokaryotic chromosome is a single DNA molecule after replication each copy is attached to a different part of the bacterial cell membrane; the cell begins to pull apart, and the copied and original chromosome is separated into opposite directions Binary fission of the coccoid-shaped bacterium Enterococcus faecium 23 SAN DIEGO MESA COLLEGE SCHOOL OF NATURAL SCIENCES General Biology (BIOL 107); Instructor: Elmar Schmid, Ph.D. M Miittoossiiss ccaann bbee iinnhhiibbiitteedd aanndd ddeessttrrooyyeedd bbyy m maannyy ddrruuggss aanndd ppooiissoonnss Cell division, the cell cycle and mitosis are enormously complicated cellular processes involving many steps, protein components and enzymes Many synthetic and nature-derived molecules are known to interfere with distinct steps of the cell cycle and the mitosis process Many of these molecules, such as the plant-derived colchicine, are known poisons and toxins to humans and animals Molecules which are able to interfere with the normal process of mitosis are referred to as mitotis blockers or so-called spindle poisons, All these mitosis blockers or inhibitors have in common that they prevent cells from dividing and from forming new daughter cells Due to this many of the known mitosis poisons are used today to treat forms of human cancers, which all are characterized by continuously and fast dividing socalled cancerous cells Cancerous cells have literally lost their molecular cell cycle brakes and therefore continue to remain in the cell cycle and undergo repeated cycles of mitosis important mitosis blocking molecules (= mitosis blocker) are: 1. C Coollcchhiicciinnee,, V Viinnbbllaassttiinnee - these plant-derived molecules block mitosis by preventing the formation of the mitotic spindle - they inhibit the build-up (= polymerization) of tubulin into functional microtubules 2. TTaaxxooll,, R Rhhaazziinniillaam m - stop the mitosis by stabilizing the microtubule polymer - since the degradation (= depolymerization) of tubulin is blocked, the chromosomes cannot migrate toward the poles Taxol and Vinblastine are potent anti-cancer drugs used in modern chemo-therapy Taxol was isolated 1967 from the bark of the Pacific yew tree (Taxus brevifolia) 1992: the FDA approved Taxol for refractory ovarian cancer and in 1994 for treatment of refractory breast cancer since the Pacific Yew is a slow growing tree and 2 grams of Taxol is needed for treatment of one cancer patient, scientists are currently looking for alternative sources and develop bio24 SAN DIEGO MESA COLLEGE SCHOOL OF NATURAL SCIENCES General Biology (BIOL 107); Instructor: Elmar Schmid, Ph.D. technological approaches to meet the demand in the hospitals TThhee cchheem miiccaall ssttrruuccttuurree ooff tthhee m miittoossiiss bblloocckkeerr TTaaxxooll the mitosis blocker Vinblastine (and vincristine) are so-called plant alkaloids both molecules are isolated from the Madagascar periwinkle plant (Catharanthus roseus or formerly classified as Vinca rosea) these Vinca alkaloids bind to tubulin, thus preventing the cell from forming the spindle apparatus it needs to move its chromosomes e.g. the cell poison cycloheximide blocks the activity of the DNA polymerase and prevents the duplication of the chromosomes M Miittoossiiss & &C Caanncceerr the different body parts, tissues and cells of multi-cellular organisms are the result of multiple cell divisions for the body to function normally, the organs and tissues must communicate to control the development of each of its cells and tissues if uncontrolled cell growth in a part of a tissue or body happens, due to the continuous cell divisions of one (!!) cancer cell, normal functions of the individual can be seriously impaired uncontrolled cell growth leads to tumors; two types of tumors are observable in biological organisms Malignant tumors can induce secondary tumors (= Metastases) by the release of metastatic cells, which lodge and begin to grow in other parts of the body 25 SAN DIEGO MESA COLLEGE SCHOOL OF NATURAL SCIENCES General Biology (BIOL 107); Instructor: Elmar Schmid, Ph.D. Benign tumors formed by non-aggressive cancerous cells that remain in the initial location Uncontrolled cellular growth and cancer have been shown to be the result of mutations of so-called oncogenes. Cancer cells usually show a loss of cell cycle control due to mutations of critical proteins of the cell cycle control system e.g. many women with breast and ovarian cancer carry a mutation in the so-called cancer susceptibility gene, BRCA1 e.g. a single nucleotide change in the ras oncogene, located on human chromosome 11, is frequently associated with bladder cancer e.g. many human cancers are caused by mutations of so-called Tumor suppressor genes, such as p53 or Rb Retinoblastoma a juvenile eye cancer caused by a mutation in the Rb gene located on human chromosome 13 since the gene is dominant inherited, both alleles must be mutant for the cancer to develop the Rb gene product interacts with a protein called E2F, which plays a role in the control of cellular replication during the S phase of the cell cycle p53 this tumor suppressor gene is located on the human chromosome 17 a single amino acid substitution, e.g. at amino acids 175, 248, and 273, lead to loss of function of the gene product about 50% of human cancers are associated with p53 mutations, including cancers of the bladder, breast, cervix, colon, lung, liver, prostate, and skin p53 related cancers are very aggressive and lead to a high degree of mortality in affected individuals 26