Supplementary Figure - Word file
advertisement

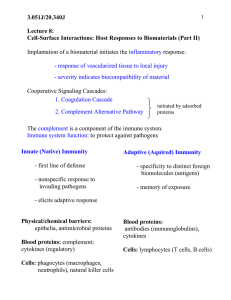
Supplementary information Structure of C3b reveals conformational changes underlying complement activity Bert J.C. Janssen1, Agni Christodoulidou 2, Andrew McCarthy3, John D. Lambris2 and Piet Gros1,# 1 Crystal and Structural Chemistry, Bijvoet Center for Biomolecular Research, Faculty of Sciences, Utrecht University, Padualaan 8, 3584 CH Utrecht, The Netherlands 2 Dept. of Pathology and Laboratory Medicine, School of Medicine, University of Pennsylvania, Philadelphia, USA 3 European Molecular Biology Laboratory, Grenoble outstation, 38042 Grenoble Cedex 9, France # to whom correspondence should be addressed: p.gros@chem.uu.nl Supplemental Table 1, Crystallographic data a, Data collection and refinement statistics Data for the outermost shell are given in parenthesis. b, Molecular replacement scores from Phaser1 Supplemental Table 2, Domain rotations and translations between C3, C3b and C3c C3b was superposed onto C32 (pdb code 2A73) and C3c2 (pdb code 2A74) on the basis of MG1, MG2, MG4-MG6 of the ring. Using this superposition the rotation and center-of-mass translation of the domains were calculated with SUPERPOSE in the CCP4 package3. Supplemental Figure 1, Electron density for each domain Electron density (using coefficients 2mFobs-DFcalc, φcalc) of C3b contoured at 1σ (blue wire frame) for each domain (ribbon representation; coloured according to Figure 1). a, MG1; b, MG2; c, MG3; d, MG4; e, MG5; f, MG6 ; g, LNK; h, MG7; i, CUB; j, TED; k, MG8; m, anchor; n, C345C. Supplemental Figure 2, Cleavage sites for factor I Inactivation of C3b to iC3b by factor I cleavages occurs in the CUB domain. First factor I cleaves between the fully exposed residues Arg1281 and Ser1282 in loop β6β7 of CUB which generates iC3b1. A second cleavage occurs between Arg1298 and Ser1299 which are in strand β8 of CUB generating C3f (18 residues) and iC3b24, 5. These residues are partly occluded in C3b in the interface of CUB with MG2; this also indicates that the structure and orientation of CUB with respect to the rest of C3b changes in the conversion to iC3b liberating this cleavage site. Third, factor I cleaves between Arg932 and Ser933 in loop β3’-β4 of CUB, resulting in the formation of C3dg (40 kDa) and C3c (135 kDa)6 (Figure 4). This indicates that factor I might bind in various ways to the CUB domain assisted by one of the cofactors (e.g. factor H, CR1 or MCP) which might be involved in the unraveling of the CUB domain enabling factor I to cleave. Supplemental Figure 3, Modeling of 2-macroglobulin (2M) based on 25-Å resolution EM reconstruction maps a, Model and EM-reconstruction map of native 2M b, Model and EM-reconstruction map of methyl-amine treated 2M Shown are the four monomers in different colors with the same view of the two 2M forms side-by-side. Electron reconstruction maps7, 8 were kindly provided by S.J. Kolodziej and J.K. Stoops. Monomers of native 2M were constructed by removing the LNK, ANA and C345C domains from the structure of native C3; C345C is absent and LNK-ANA are expected to have a different fold in 2M. Monomers of methylamine-treated 2M were constructed from C3b by removing LNK and C345C. Subsequently, dimers of the two different forms were made based on the two disulfide bonds that cross link the monomers (disulphide bonds 256-410 and 410-256; C3 numbering used). For each of the two forms we constructed tetramers, dimer of dimers, based on the two EMreconstruction maps applying strict 222 symmetry. The C3-based model of native 2M fits reasonably well into the EM-map. The differences indicate that possibly MG7 will be displaced (as indicated by an arrow); in addition, the bottom part of the ring (domains MG1-4-5) are possibly in contact with its symmetry-related part of another molecule in the tetramer (as indicated by an arrow marked “ring”). The C3b-based model of methylamine treated 2M fits well into its EM-map. The TED domain, however, should likely be pointing more into the central cavity (indicated by an arrow) with its thioester moiety pointing inwards. The overall fit of these two models indicate that the conformational changes observed for C3 and C3b likely occur in the conversion of tetrameric, native 2M into activated, methylamine-treated 2M. The 2M models are in good agreement with functional data9. The receptor binding domain (RBD) changes from hidden in native 2M to exposed in methylaminetreated 2M. The bait region (indicated by an arrow) is exposed, assuming that it adopts the same position as the ANA domain does in C3. Supplemental Movie, Transition of C3 into C3b The movie was prepared using CNS10, 11 and is only based on the structures of C32 (first frame) and C3b (last frame). Colours used according to Figure 1A. 1. 2. 3. 4. 5. 6. 7. 8. 9. 10. 11. Storoni, L. C., McCoy, A. J. & Read, R. J. Likelihood-enhanced fast rotation functions. Acta Crystallogr D Biol Crystallogr 60, 432-8 (2004). Janssen, B. J. et al. Structures of complement component C3 provide insights into the function and evolution of immunity. Nature 437, 505-11 (2005). The CCP4 suite: programs for protein crystallography. Acta Crystallogr D Biol Crystallogr 50, 760-3 (1994). Harrison, R. A. & Lachmann, P. J. Novel cleavage products of the third component of human complement. Mol Immunol 17, 219-28 (1980). Davis, A. E., 3rd & Harrison, R. A. Structural characterization of factor I mediated cleavage of the third component of complement. Biochemistry 21, 5745-9 (1982). Lachmann, P. J., Pangburn, M. K. & Oldroyd, R. G. Breakdown of C3 after complement activation. Identification of a new fragment C3g, using monoclonal antibodies. J Exp Med 156, 205-16 (1982). Qazi, U., Kolodziej, S. J., Gettins, P. G. & Stoops, J. K. The structure of the C949S mutant human alpha(2)-macroglobulin demonstrates the critical role of the internal thiol esters in its proteinase-entrapping structural transformation. J Struct Biol 131, 19-26 (2000). Qazi, U., Gettins, P. G., Strickland, D. K. & Stoops, J. K. Structural details of proteinase entrapment by human alpha2-macroglobulin emerge from threedimensional reconstructions of Fab labeled native, half-transformed, and transformed molecules. J Biol Chem 274, 8137-42 (1999). Sottrup-Jensen, L. Alpha-macroglobulins: structure, shape, and mechanism of proteinase complex formation. J Biol Chem 264, 11539-42 (1989). Brunger, A. T. et al. Crystallography & NMR system: A new software suite for macromolecular structure determination. Acta Crystallogr D Biol Crystallogr 54 (Pt 5), 905-21 (1998). Echols, N., Milburn, D. & Gerstein, M. MolMovDB: analysis and visualization of conformational change and structural flexibility. Nucleic Acids Res 31, 478-82 (2003).