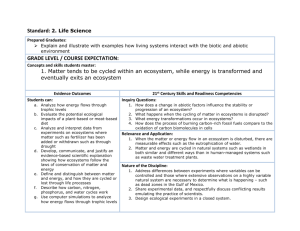
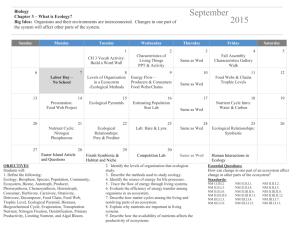
jec12025-sup-0001-AppendixS1
advertisement

Supporting material Appendix S1. A list of the 754 submitted questions. Those who were consulted and provided questions include Steve Marcelo Aizen, Tiffany Aslam, Mark Auliya, Sandro Azaele, Avi Bar-Massada, Patty Balvanera, Lior Blank, Ute Bradter, Judie Bronstein, Rob Brooker, Yvonne Buckley, Harald Bugman, Tom Cameron, Scott Chamberlain, Jo Clark, Erika Crispo, Chris Dickman, Carsten Dormann, Alison Dunn, Liz Elliot, Rampal Etienne, Dan Flynn, Enric Frago, Gregoire Freschet, Stephan Gale, Michael Gundale, Richard Gunton, Michael Gerisch, Richard German, Sofia Gripenberg, Camille Guilbaud, Kyle Harms, Laura Harrison, John Healey, Mark Hebblewhite, Klaus Henle, Joe Hicks, Steven Higgins, Joaquin Hortal, Etienne Laliberte, Jan Leps, Jonathan Levine, Tom de Luca, Chris Lortie, Neal Hockley, Mike Hutchings, Hans Jacquemyn, Karin Johst, Laurence Jones, Kat Jones, Micael Jonsson, Hans Kasperidus, Reinhard Klenke, Roger Kouyos, Robert May, Jason Matthiopoulos, Jessica Metcalf, Sean McMahon, Shai Meiri, Katrin Meyer, Rebecca Morris, Rosmarie Müller, Marie-Charlotte Nilsson, Tim Paine, Guy Pe’er, Will Pearse, Nathalie Pettorelli, Avi Perevolotzky, Alex Pigot, Marco Plebani, Janielle Porter, Trevor Price, Simon Rowe, Rob Salguero-Gomez, Santiago Saura,Martin Schädler, Boris Schröder, Alexander Singer, Eleanor Slade, Phil Stephens, Maja Sundqvist, Jeremy Thomas, Rebecca TierLange, Johan du Toit, Jason Tylianakis, Jun-Ichirou Suzuki, Marie Vanderwalle, Ciska Veen, Rebeca Velazquez, Steve Votier, Mark Whittingham, Kerstin Wiegand, Kathy Willis and Tobias Zust. 1. There’s a lot of talk about eco-evolutionary feedbacks now. But how fast can they happen? 2. What evolves first, a specialist or a generalist? 3. Will the breakdown of biogeographical barriers slow or accelerate the evolution of new species? 4. How does phenotypic plasticity interact with environmental pressures to influence evolution? 5. Does indeed nothing make sense in evolution except in the light of ecology? Or vice versa? 1 6. Functional traits: can one predict evolutionary pressures based on puntual measurements of functional traits? 7. Is there any signal of microevolutionary change at the level of communities or ecosystems? 8. Is evolutionary suicide more than interesting theory? 9. Under what circumstances will evolutionary change keep pace with environmental change? 10. Is the evolution of competitive dominance actually rather common? (thinking of monodominants like giant sequoias, humans, tree climax species in general... that only seem to be knocked back by major climatic disturbances or pathogens?) 11. To what extent are insect species adapted to their local biotic or abiotic environments at regional or landscape scales? 12. Is there a general theory of scaling? And how can we find it? 13. How local is adaptation? 14. Studies on scaling laws: they seem to ignore history and assume that species are interchangeable. Is this the best we can do? 15. As genetic studies are increasing with ever more sophisticated tools, it is likely that the uncovering of cryptic/new diversity and even cryptic/new species will be increasing - how do these new discoveries impact on our understanding and evaluation of biodiversity? 16. Senescence: how universal is senescence? Under which habitat do senescence rates slow down (or even go negative?). Is there phylogenetic inertia for senescence patterns? Does the internal organization of modular organism predict which species undergo/escape senescence? 17. How important are genetic and epigenetic diversity for adaptation to environmental variation? 18. What is the ecological significance of epigenetic variation? 19. What are the relative influences of genetics, epigenetics and non-genetic processes on phenotypes? 20. What is the ecological importance of epigenetic variation? 21. What aspects of the biotic and abiotic environment lead to epigenetic variation? 22. Which phenotypic traits are subject to epigenetic variation? 23. What maintains consistent individual differences (personality) in animal behaviour?/Is variability in behaviour a strategy? 2 24. What are the magnitudes and causes of inbreeding preferences in nature? 25. What is the physiological basis of life-history trade-offs? 26. Why do closely related species often exhibit very different amounts of intraspecific variability? 27. Why can't we find evidence of the cost of phenotypic plasticity? 28. Why do most species have stage-structured life cycles? 29. Why are there no genes for stochasticity (plasticity) in phenotype? 30. Does genomics have a role in ecology? 31. What is the value of genetic capital? 32. Is there red-queen hypothesis for environmental change and evolution – running to stand still? 33. How fast can evolution proceed in a changing and variable environment? 34. Does molecular detail alter or overturn predictions from ecological and life history theory; or is it just more natural history for the ecologist? 35. Does molecular insight into the mode and rate of trait evolution change ecological theory? 36. What ecological and genetic factors constrain evolution in a changing and variable environment? 37. How does phylogeny alter our interpretation of morphology, physiology and ecology? 38. What cues do organisms use to determine their phenology? 39. When does local adaptation lead to speciation? 40. Do species concepts matter? 41. How many of the described morphospecies are functionally distinct? 42. How do you define a species? 43. Do we need an ecological understanding of molecular systems? 44. What is the role of the epigenome in phenotypic plasticity? Can the capacity to utilise epigenetic mechanisms make some species more “adaptable” than others? 45. How has the Evolution of sex occurred? 46. How do animals compete for resources? This is important as it is the process that drives the unequal distribution of resources and thus natural selection, however, the current explanations in all behavioural ecology texts is fundamentally flawed. 3 47. In a world of increasing international interconnectedness, will eukaryotes increasingly follow the microbial rule: everything is everywhere but the environment selects? 48. Do southern hemisphere plants have more conservative traits than those in the northern hemisphere? 49. Does the rate of succession and the spatial distribution of vegetation differ between climatically similar areas because of the available biota (e.g. animal dispersers)? 50. Can fields of farmed saltmarsh, mangrove, seagrass replicate, equal or increase the ecological productivity on the same area of natural coastal complex? 51. What geographical and environmental features shape connectivity among riverine networks? 52. What are the consequences of the Bering strait opening up? 53. What will be the impacts of the more permanent opening of the north passage? 54. Are corridors always important for the distribution and abundance of animals and plants, or does the importance of corridors vary widely in different landscapes? 55. To what extent do the low dispersal rates found in many European insects and plants reflect historical adaptations to the availability and dynamics of species’ habitat-patches under defunct anthropogenic forms of land management? 56. Will the breakdown of biogeographical barriers lead to sustained increases in local diversity? 57. To what extent do dispersal traits currently limit the connectivity of fragmented habitats? 58. What is the impact of landscape structure on movement (dispersal), connectivity, and population dynamics? (This is related to the older question about the relative impact of habitat loss vs. fragmentation which is still unsettled, but revitalized in the current context of range shifts under climate change) 59. What are the costs of dispersal? 60. How important is habitat connectivity for successful dispersal and population persistence? 61. What is the heritability/genetic basis of dispersal and movement behaviour? 62. What is the degree of within-individual and among-individual consistency and variation in seasonal movements and locations? 63. What rules shape the development of locations and movements? 4 64. What type of information do real animals use to make dispersal and foraging decisions and how do they process it in relation to its value? 65. How do environmental conditions (meteorological or oceanographic) impact the costs of animal movement? 66. To what extent does morphology constrain animal distributions? 67. Where do wide-ranging large marine animals (e.g. cetaceans, turtles, etc.) come from and where are they going? How do we describe the spatial connectivity for organisms that have the potential for wide dispersal? 68. What cues drive migration and will those cues change in response to differential rates of change on species' breeding and non-breeding grounds? 69. Is Levy flight the most suitable model to underpin analysis of animal movement trajectories ? 70. Seeds that disperse very long distances or survive in the soil for a very long time represent the tail of the distribution. Is this tail dominated by seeds with particular traits, or simply by seeds that happen to be the most abundant (and thus inevitably the smallest)? 71. Is the persistence of species assemblages in fragmented landscapes driven by dynamical extinction-recolonisation equilibria? 72. How do climatic conditions (past and present) affect the ability of animals to fly? 73. Is dispersal the key to persistence under intense habitat fragmentation and shifting climatic conditions? If so, how do we model/study dispersal in organisms that range in size from pollen to top predators? 74. What are the effects of fragmentation on the evolution of morphology and demography of species? 75. How do anthropogenic changes to land cover affect dispersal and migration patterns? 76. We argue over what is an appropriate ecological unit for a particular study, yet we all agree that such things don't exist. Under what framework can we link processes at a smaller scale (e.g. dispersal limitation) into a wider scale (e.g. range size)? 77. Is it possible to infer the dispersal range from traits? 78. What is dispersal? 79. While theory predicts that species should go extinct under certain fragmentation and isolation thresholds, this does not seem to be the case. What allows them not to go extinct then? 5 80. What are the contributions of biogeographical factors and historical contingencies in determining present day ecological processes? 81. How can models of species distribution (e.g. resource selection functions) be successful at predicting distribution under changing environmental conditions? 82. How can we find a general mechanistic understanding of distribution patterns and range dynamics? 83. After accounting for all confounding variables, do invasive alien species have different traits from those of native and non-invasive alien species? 84. Are there general traits or variables to predict why some alien species are invasive? 85. How long does it take for an invasive species to become a native species? 86. When does biological invasion matter? 87. Which traits promote plant invasiveness 88. Are there thresholds of occupancy beyond which efforts to control invasive species (e.g. weeds) are ineffective, and, if so, what are they? 89. Can we identify characteristics of likely problematic invasive species? 90. Can we predict whether a species will become invasive in new habitats/environmental change by evaluating its capacity for adaptive evolution? 91. If a species is trying to invade and the area already presents congenerics, does it make invasion more or less likely? 92. How do individual dispersal traits affect invasions? 93. Will invasive species eventually become naturalized or integrated into local communities with reduced impacts? 94. Are patterns of biogeography and diversity similar across taxonomic groups? 95. Where do we draw the line between naturalized vs invasive species? At what point does a former invasive become integral to the functioning of the ecosystem? 96. Have we been studying uninformative patterns in ecology? For instance, speciesabundance distributions, species-area curves, body size patterns. 97. If habitat diversity drives the Species-Area relationship in many cases, why does habitat diversity itself scale so consistently with area? 98. What controls spatial variation in speciation and extinction rates? 99. What is the feedback between diversity and diversification? 100. How do range margins evolve and what determines them? 101. How does climate/latitude/elevation influence range margins? 6 102. Is there a synoptic model of global patterns of biodiversity? (Maintained at the Institute for Global Biodiversity Simulation, of course). 103. What observable variables can be used to assess the distance of a population from a “tipping-point” beyond which it is in jeopardy of extinction? 104. Can we accurately predict the “extinction debt” from the loss and fragmentation of natural habitats, and can we find ways to forestall paying it off? 105. Which native woody species should be planted, and how, to satisfactorily restore degraded land and reintegrate forest fragments in the Asian tropics and subtropics? 106. How do we establish if jellyfish are taking over perturbed marine systems ? 107. Can we infer process from pattern? One of the fundamental issues in ecology is that the dynamics generally operate over time-scale that are much longer than grant income, or even academic careers. Therefore the types of long time-series necessary are often difficult to obtain -- however, in principle the current state of the population and its spatial pattern may tell us something about the underlying processes. 108. Can we resolve the relationship between dispersal and the distribution of adaptive variation and its impact on meta-communities? 109. Did most rare and narrow-niche species reach their currently few remote and isolated locations via extremely rare and improbable dispersal events, or are they typically remnants of once-relatively-continuous distributions? 110. What is the effect of fragmentation on taxa that operate at specific scales? 111. What is the effect of fragmentation on biodiversity? 112. How important is (habitat) heterogeneity for understanding the distribution of species at local and larger scales? 113. What determines natural treelines? 114. How, and how strongly, does long-distance migration link otherwise separate ecosystems? 115. How long does it take species geographic distributions to equilibrate with climate? 116. What maintains species range margins? 117. What are the typical spatial scales over which behavioural and population processes operate in insects, and do they vary systematically for different guilds and trophic levels? 118. How much information is buried in spatial patterns with respect to the underlying processes and how can we extract this information? 7 119. How important are offspring sex ratios as drivers of population dynamics? 120. Vole cycles – why did they disappear and now seem to reappear? 121. Multiple stable states and irreversibility are understudies. Specifically, does biodiversity protect against flips to the less desirable state? 122. What determines the relative importance of direct and indirect effects of environmental change on population dynamics? 123. How coarsely can we consider density dependence while still predicting future population dynamics accurately? 124. Demography: how does belowground population dynamics affect aboveground population dynamics? 125. How can we disentangle the effects of the environment, internal processes of population dynamics, and stochasticity on observed species distribution patterns? 126. Do observed population trajectories reflect equilibria or transient dynamics? Do the transient population dynamics observed in mathematical models occur rarely, often, or ubiquitously in real populations? 127. Is the tempo of natural selection on genes affecting fitness and demography sufficient to cause instability in population dynamics (Chitty 1954 revisited!)? 128. To what degree does genetic variation influence population dynamics and persistence and over what timescales? 129. Do different demographic rates vary over different spatial scales, and how do they then combine to influence spatio-temporal population dynamics? 130. How do disturbances influence population dynamics of species with different lifehistory strategies? 131. How does isolation affect population dynamics and persistence? 132. How are population dynamics and evolutionary dynamics linked? 133. Do most organisms live near their physiological limits? 134. Can we predict species interactions based on phylogenetic relatedness, and do how do similarities in species traits correlate with phylogenetic relatedness? 135. What is the ecological significance of adaptive genetic diversity? 136. How do species interactions and abiotic constraints interact to determine the distribution of species? 137. What is the relative role of biotic and abiotic processes in determining presence/absence, abundance and composition of populations? 8 138. What are the relationships between the quality and quantity of plant resource investment toward their different tissues? 139. In long lived species (e.g. turtles) that produce many offspring in a given year across multiple years, is the offspring contribution to the adult population regular across years or are there always bumper “year classes” 140. How does the 'personality' of individual animals affect the performance (e.g. reproduction, dispersal) of the populations to which they belong? 141. Given the unmanageable multitude of hypotheses about population declines (and of combinations thereof), when a population starts to decline, how can we organize research effort in a way that can isolate the cause(s) and possible mitigation mechanisms? 142. Is genetic variation sufficient for populations to adapt to environmental stressors? 143. What determines the lower threshold of population abundance? 144. Can several reciprocal feedback processes simultaneously shape the dynamics of interacting populations? 145. What are the population dynamic consequences of delayed life-history effects in space and time? 146. What causes massive variability in recruitment? 147. What are the magnitudes of direct and indirect selection on reproductive strategies in the wild? 148. How does environmental stochasticity shape functional traits of species? 149. Does global change disrupt aboveground-belowground interactions due to differences in the spatial and temporal scales of processes above- and belowground? 150. What affects the predictability of population size? 151. What is the relationship of species traits to their ecological behaviour? This is again meant in very wide sense – can the physiological properties of a species determine its habitat preferences? But also, how much can the species’ “soft traits”, i.e. easily measurable (say morphological) properties, predict physiological behaviour, and, in consequence, its habitat preferences. 152. Why do some non-native species require disturbance events to colonise whereas others do not? 153. To what extent are the effects of abiotic conditions on individual fitness direct versus indirect (e.g., by changing ecological interactions)? 9 154. What explains the persistence of small populations over long time periods? Are they just the lucky ones, have they evolved adaptations to being small, or do they simply live in a very predictable environment? 155. Why do only some trees lose their leaves in winter? 156. To what extent do population processes reflect the effects of stochasticity? 157. Should most ecological models be age-structured? and if so how? It is clear that many marine organisms have huge variation in size between larvae and adult; in addition for most complex organisms survival, reproduction and predation are all linked to age. Should we be using coupled PDEs (Partial Differential Equations) that can capture age-structure in all of our models? 158. To what extent does adaptation drive ecological dynamics at different scales? 159. How does environmental stochasticity interact with the intrinsic dynamics of population systems, and how will changes in environmental signals filter through to affect dynamics? 160. How should we model ecological systems in order to minimise forecasting error? 161. How does density-dependence function in non-equilibrium systems? 162. Is hysteresis ecologically relevant, or is it only Scheffer pestering us with cusps? 163. Should we discard deterministic models, and switch to stochastic ones? 164. Should we discard frequentist statistical methods and go Bayesian? 165. Should we move from hypothesis testing to a more systems based analytical approach? 166. What alternatives to long-term time-series are there for detecting change in natural environments and linking this to specific drivers? 167. How important are transgenerational effects to population dynamics? 168. How important are plastic responses to population dynamics? 169. Does diet choice, habitat choice or mate choice govern lifetime reproductive success (or Ro; or population growth in general) 170. How can behaviour of individuals shape population behaviour in response to disturbance? 171. What determines population numbers in a given environment? 172. What proportion of species are helpfully viewed as metapopulations? 173. What are the causes of under crowding? 174. What are the levels that under crowding occurs? 175. What determine under crowding? 10 176. How does the ecology of a species change through time? 177. Can remote sensing and DNA barcoding be combined to allow real time population and community monitoring on multiple scales? 178. Can the shape of a functional response be predicted? Can you use species traits and system characteristics to predict the form of a functional response? 179. How important are animal population dynamics to plant population dynamics in different biomes? 180. How important are individual differences in determining population dynamics? 181. How much are typical population sizes determined by extreme events? 182. To what extent does food-meditated spring body condition determine subsequent breeding success? 183. What is the long-term implication of nutrient removal from rural landscapes and concentration in urban centres? 184. Is P limitation a factor that will be of overwhelming importance in the long run also in temperate forests? 185. Is competition for light inherently more asymmetric than for nutrients? 186. Which nutrient(s) control productivity of terrestrial biomes, from the poles to the tropics? 187. Among plants, are the best competitors those that are least sensitive to a shortage of limiting resources? 188. Does seasonality (the alternation of periods with different resource abundance and renewal regimes) contribute to instability of trophic interactions? 189. What role do endosymbionts play in regulating the dietary range of insects? 190. What is the relative importance of biotic versus abiotic feedbacks between the plant and soil environment? 191. What is the relative importance of biotic interactions (e.g. predation, competition) versus abiotic interactions (e.g. fire, weather) in structuring animal and plant communities in arid environments? 192. Are communities structured more by biotic interactions between species, or abiotic interactions between each species individually and the environment? 193. Why would nutrient resorption (THE major source of resource conservation) be decoupled with the plant economics spectrum (that is, other “conservative” traits)? 194. What is the most important carbon source for below-ground animals and fungi: roots or litter? 11 195. How do aboveground-belowground interactions determine why species are and the assemblage of ecological communities? 196. How important is allelopathy in natural plant communities? 197. What excludes fast-growing plants from infertile soils? 198. How do organisms void elemental excesses when dealing with non-ideal quantities and qualities of resources? 199. How do resource pulses in arid environments affect resource use and interactions between organisms? 200. Which ecological processes are similar between micro- and macroorganisms? 201. How does ecological disturbance govern C, N and P dynamics and capital in terrestrial ecosystems? 202. What is the role of species interactions in driving ecosystem processes, including fluxes in the ecosystem of C, N and P? 203. In highly nutrient limited ecosystems, such as arctic tundra, what will be the key limiting factors for those plants that will dominate as a result of a future climate? 204. What factors modify the reciprocal flow of nutrients between aquatic and terrestrial systems and what are the ecological consequences of such modifications? 205. How do major macronutrient cycles (N, P) interact to modify carbon stocks and flows? 206. What will be the overall magnitude of the biogeophysical (e.g., albedo) and biogeochemical (e.g., cycles of C and H2O) feedback from the global forest to the atmosphere under a changing climate? 207. How will greenhouse gas emissions from soils vary as peat lands dry out and tundra warms? 208. How can we measure plant biomass without killing them? 209. How does the summation/ integration of individual plant and plant unit allow responses to local environmental conditions? 210. How do plants adapt to changes in CO2? 211. C4/C3 photosynthesis – are the C4 ones getting more common in nature? 212. What determines the strength of trophic interactions? 213. Do trophic interactions just reflect metabolism? 214. How do species traits influence the strength of trophic interactions? 215. Do interaction strengths determine food web structure? 12 216. What are the interelationships between body mass, abundance and food web structure? 217. How does spatial scale affect the structure of food webs? 218. How does warming affect the strength of trophic interactions? 219. Are alpine plants pollen limited? 220. What is the relative importance of direct (consumption, competition) vs indirect (induced behavioural change) in determining the effect of one species on others? 221. How do the allometric scalings of attack rates and handling times differ among biomes, and between the land and ocean? 222. What controls relative strength of competitive vs facultative interactions? 223. What limits size of top predators? 224. How affect interspecific interactions (e.g. trophic) distributions of interacting species at different spatial scales? 225. Is competition between plants more intense under productive or unproductive conditions? 226. How important are ecological processes (i.e. competition/predation/mutualisms) in explaining the diversification of life? 227. Phylogenetic community structure (PCS): How do mutualisms/predation contribute to PCS? We have not yet looked at interactions other than competition for driving PCS, despite the fact that we know mutualism/predation are important in general. 228. How important is intra-guild predation in structuring communities? 229. How is vulnerability to predation affected by rarity? 230. Is competition between plants predictably replaced by facilitation in harsh environments? 231. Is there a competition-colonisation trade-off in plants? 232. What is the relative incidence of competition vs. facilitation in plant communities? 233. What are the effects, and the relative contribution of competition and facilitation to determine local and regional patterns of plant diversity? 234. Are bipartite mutualistic networks really just driven by competition within each partite (e.g., plants in a plant-pollinator network) for access to partners within the other partite (e.g., pollinators within the same plant-pollinator network). 235. How is the process of competition, the actual fight for resources between two neighbouring individuals, regulated (by time, by external environmental drivers) 13 and how does it then translate into the final well-known measures of plant success, e.g. biomass, seed production? 236. How widespread and important are enemy-mediated indirect interactions (apparent competition, apparent mutualism) in ecological communities? 237. Does the balance among species interactions (such as competition and facilitation) shift in a predictive way along environmental gradients? (this is basically the stressgradient hypothesis which is still unresolved) 238. How does the relationship between ecological processes such as competition or facilitation and spatial patterns change across scales 239. Do prey-predator size ratios drive dynamics, or does selective extinction drive observed size ratios? 240. What processes or factors lead to the persistence of mutualisms? 241. What is the relative importance of trophic and non-trophic interactions? (How does trophic connectance compare with non-trophic connectance, and what determines variation in this balance?) 242. How opportunistic are ecological interactions i.e. do mutualisms etc arise rapidly or do they require long periods of time to evolve? 243. Are phylogenies a good surrogate for the differences in ecological role among species? 244. How might temporal- or condition-dependent assimilation efficiency affect our understanding of food webs? 245. How do pervasive structural features of interaction webs (e.g., nestedness) originate? 246. Under what ecological conditions do the benefits of cooperating with other species outweigh the costs of doing so? (this is the most basic question one can ask about facilitation and mutualism, and one for which we still don't have a clear answer.) 247. What aspects of ecological networks affect their stability? 248. How do food webs respond to temperature change and variation? 249. What is the ecological role of physical processes (internal waves, atmospheric turbulence) in shaping trophic interactions from micro to macro spatial scales? 250. What are the impacts of introduced species on native food webs? 251. How does the composition of a parasite/commensal/symbiotic community impact on host fitness? 252. To what extent do plant mediate interactions between herbivores and microbes? 14 253. Does artificial plant breeding disrupt co-evolved plant-insect interactions and plant defence mechanisms? 254. How important are symbiotic/commensal microorganisms for host evolution? 255. Are there really ubiquitous (or close to it) distributions of micro-organisms that contrast clearly with limited distributions of macro-organisms. If so, where is the discontinuity and why? 256. How important is consideration of coinfection in understanding disease dynamics? 257. How does ecology influence the emergence and dynamics of disease? 258. How will climate change affect disease emergence and dynamics? 259. What is the importance of plasmids in bacteria/symbionts in insects/other laterally transferred parts of genome in altering ecological/evolutionary dynamics? 260. Can ecological understanding help in the management and treatment of infection in individuals? 261. What part is played by parasites and disease in food webs? 262. How can we understand (and predict) the evolution of diseases? Why are diseases virulent when this causes morbidity or mortality in the host and therefore reduces the chances of transmission? Why are some infections short-lived while others are persistent? What key features do we need to understand these different adaptations to the pathogen life cycle. Do we need a full understanding of the pathogen's genetics or can we safely assume that disease characteristics evolve in a continuum? 263. Is the mode of transmission of pathogens a good predictor of disease dynamics? 264. How are directly transmitted parasites maintained in fragmented host populations? 265. What effect does host dispersal ability have on the spatial spread of parasites with different life history strategies? 266. What is the role of population structure in the transmission of infection? Clearly human (and animal) populations are segregated at a variety of scales: households, social groups, work-places, communities, nations etc etc -- how much of this structure is needed and can any of it be approximated? 267. To what extent do plant-associated microbes such as plant pathogens and foliar endophytes drive patterns of insect herbivory? 268. How do plant-mediated interactions between beneficial microbes and insects affect plant fitness and productivity? 269. How do plant-associated microbes affect plant-induced defenses against insect herbivores? 15 270. How do belowground symbioses between plants and microorganisms contribute to the ecology and evolution of aboveground plant-herbivore interactions? 271. To what extent is plant community composition and diversity determined by interactions with above-ground (e.g. insects) and below-ground (e.g. soil pathogens) natural enemies? 272. Does ecological theory developed for plants and vertebrates apply equally to microorganisms? 273. Is the resource allocation model that underpins most ecological thinking correct? Are animals really limited by food/energy supply that they must allocate between alternative functions, or do other physiological constraints act first? 274. Does microbial diversity directly reflect stability and productivity in the soil environment? 275. What is the extent of microbial diversity and what is its relationship to functional diversity? 276. Can the well established body size abundance/diversity relationships be extended to microbial organisms? 277. Do macro-ecological theories such as the island biogeography theory apply to microbial ecosystems? 278. Does microorganism diversity (functional type, species, genotype) matter for ecosystem processes? 279. Is successful forest restoration on a workable timescale dependent on mycorrhizal inoculation? 280. How do we measure wildlife disease intensity? 281. How much detail of the parasite community is needed in order to predict the impact of disease on host populations? 282. What is the role of diseases imported with invasive organisms? 283. What are the effects of heterogeneity in host-parasite infectivity to the epidemiology of the disease? 284. Would thinking of the immune system as an ecosystem (sequentially assembled to a given level of diversity in the first years of life) designed to cope with “invasive” infections help in better design of vaccines? 285. When can heterogeneity in host response to parasites be safely ignored? 286. How do microparasites influence communities and food web theory? 287. What is the role of parasites in maintaining host species diversity? 16 288. Can we understand the ecology of microbial communities inside other organisms (e.g. in the gut)? 289. When does one or a few infectious organisms drive host-parasite population dynamics and when does the community matter? 290. What are the role of diseases in shaping the community? 291. Are there endemic, endangered or even extinct microbes? 292. How different would communities be if there were no diseases? 293. What is the level of functional redundancy in prokaryotic communities in terrestrial and aquatic environments? (ie does prokaryotic diversity matter?) 294. How are parasite communities assembled? 295. How important are parasites from an ecological perspective? [are parasites or predators the dominant factor controlling population sizes?] 296. To what extent are community dynamics pre-determined by macroevolutionary patterns? 297. Is functional redundancy common? Can we remove species without ill effects on the others? 298. Which community properties, if any, make some ecosystems more resilient than others to environmental change? 299. What (if any) kind of relationship is there between genetic diversity, functional diversity, and functional redundancy? 300. How important are functionally redundant species for the resistance of biodiversity to disturbances? 301. Do disturbances impact different trophic levels equally? 302. How do behavioural interactions scale across food webs? 303. Are aquatic communities really more size structured than terrestrial ones? 304. Is the stability of ecological communities predictable? 305. With which traits can we predict the functioning of extinct communities? 306. Will coral reef communities be able to adapt or 'migrate' in response to ocean change (temperature/depth/pH etc.) 307. Are diverse communities more robust to disturbance than depauperate ones? 308. Are diverse communities more robust to environmental change than depauperate ones? 309. To what extent are communities discrete entities that can be studied in isolation? 17 310. Given the global declines of pollinators, what levels of resilience can we expect in insect and plant communities to continue effective propagation of species namely crops and foods? 311. What determines the resilience of communities under environmental change? 312. In which types of natural communities do equilibrial co-existence models apply and in which are conditions sufficiently non-equilibrial that equilibiral co-existence models do not apply? 313. Improved understanding of which fundamental ecological issues by a broad crosssection of society would significantly improve the conservation status of biological species and communities? 314. Which is the role of plant-emitted volatile organic compounds in the structure and function of insect communities? 315. What role does the "community context" have for constraining or facilitating the responses of individual species to environmental change? 316. How do different plant community types respond to changes in the same environmental factors (i.e. temperature, precipitation, nutrient availability)? 317. How precise and accurate must model structure and parameter estimates be, in order to predict community responses to environmental change with adequate precision and accuracy? 318. What determines the predictability of species extinctions and community collapse? 319. What is the importance of intraspecific variability for community stability? 320. Do marine invasives perform additional functional roles to those existing in the native community subject to invasion? 321. How well can community stability and function be predicted from body size distributions, and how effectively could size-structure serve as a measure of community “health”? 322. Which structural properties of interaction webs actually increase community stability and biodiversity maintenance? 323. How do predator functional responses change with community dynamics? 324. Which rules of population biology might be easily generalized to community ecology, e.g. density-yield relationships, diversity "strategies" (and whether they exist and can evolve within clones, populations or even communities), self-thinning and allometric relationships, "community genetics and evolution", resource vs. pathogen vs. mutualist niches structuring communities? 18 325. Do observed changes in kelp forest composition affect associated biodiversity? 326. Do webs of biotic interactions “collapse”, and if so what variables can predict this? 327. Under what circumstances are different types of disturbance interchangeable (with respect to consequences for species composition, diversity, etc.)? 328. Are some species functionally redundant, even in the context of stochastic and deterministic environmental changes? 329. To what extent are the processes underlying increased functioning also the processes that give coexistence? 330. How much of response to changing environments will be intraspecific and how much interspecific turnover? 331. What is the relative importance of phenotypic plasticity, genotypic variation, or inter-specific variation in determining how communities respond to environmental change, and how do this vary across different trophic levels or across different ecosystem types? 332. Which mechanisms allow the long-term coexistence of grasses and woody plants over such a wide range of systems? (This is basically the “savanna question”) 333. How does random arrival of species control the succession of ecological systems after disturbances? 334. Are there fundamental mechanistic differences between aquatic and terrestrial systems? 335. How do long-term structure of networks change with global warming? There are a lot of studies now describing structure of ecological networks, but we don't know what will, or is expected to, happen with increasing warming of our earth. 336. What are the consequences of specifying models and theory in terms of size, biomass or numbers in population and community ecology? 337. When is it appropriate to replace species identity with size in population and community ecology? 338. Are the trade-offs and traits that structure terrestrial communities the same as those that structure aquatic communities? 339. At what level, and to what extent are recognised ecosystems structured on dominant keystone species? 340. Why does community composition change in space - is it more than the sum of the parts? 341. Are redundancy and resilience linked at the community level? 19 342. Do community traits confer the ability to predict responses? 343. What are the relative roles of factors affecting species composition at fine sptial and temporal scales (namely, chance events, dispersal limitations, environmental factors and biological interactions) -- and how all this varies between biomes and eons? 344. How does flexibility of traits affect species coexistence? 345. Can you predict the consequences of species extinctions? 346. Do all species interact? Do all species interact even if weakly? If some interactions are so weak do we simply not observe them? 347. How can we integrate multispecies dynamics across spatial scales? How do we scale up from small to large processes? 348. How do weak interactions affect multiple forms of stability? 349. Climate dependence in which aspects of demography and / or species interactions, explain species-climate correlations? 350. How saturated or close to equilibrium are communities? 351. What factors govern the composition of ecological communities? 352. Are local communities only legacy, i.e. is there no convergence of community structure for sites which start with different species sets? (In other words: Will the founder-effect dominate communities for ever?) 353. Can we predict the performance of species in new environments/communities based on species’ traits or trait combinations? 354. What role does facilitation play in the development and structure of communities? 355. How does community composition and structure arise from small-scale competitive interactions? 356. To what extent do local competitive interactions scale-up to produce a decisive influence on community level patterns and processes? 357. How connected are communities? 358. The role of taxonomy in ecosystem niche overlap / stability. Is a speciose area A more stable than area B if in B the species aren’t closely related? 359. How are ecosystems altered through the simultaneous gain of species (through invasion) and loss of species (through extinction)? 360. Does the number of species have any effect on the properties of natural ecosystems, beyond those predictable from traits of individual species? 361. Are there any practically usable proxys for measuring the 'health' of an ecosystem? 20 362. How do structural features of ecological communities relate to the dynamics of those communities, e.g., their stability to outside perturbation, and how do both relate to ecosystem services provided? 363. What practical methods can we use to tell when an ecosystem is near a “tipping point,” after which dramatic and difficult-to-reverse changes will occur? 364. What confers robustness to outside perturbations on an ecosystem, and how is this robustness affected by climate change and other human impacts? 365. Can we predict the future state of ecological systems? At what resolution of measurement, and level of ecological organization and for what system types is this possible? 366. What are the ecosystem impacts of worldwide predator declines? 367. Desert ecology: how do punctual and intense climatic events (droughts, rains) affect the ecology of populations and communities on the long-run? how will changes in community structure in deserts affect other ecosystems? 368. Whether and to what extent species extinction affects ecosystem properties. Does it matter to the ecosystem to lose one or a few species, or can we loose easily half of the species without the ecosystem being affected? 369. What is the importance of functional-group diversity to sustain ecosystem productivity across biomes? 370. What is the contribution of biodiversity to sustain long-term productivity in manmade ecosystem? 371. How resilient are species interactions within variable environments? To conserve interactions among species within such variable environments - interactions that may be essential for healthy ecosystem functioning - do we need new concepts, approaches and theories; or alternatively, is a side effect of conserving enough species that we'll end up preserving critical interactions as well? 372. What are the ecological impacts of the radioactive materials released from the Fukushima nuclear power plant? It is obvious that materials with ‘strong’ radiation have substantial effects on an ecosystem but we still do not know what effects materials with ‘weak’ radiation have. 373. How predictable are the impacts of climate/global change on ecosystems and their functioning in terms of transient dynamics, interspecific interactions types, initial conditions, time horizon and phenotypic plasticity? 21 374. Are humans part of ecosystems and if so, what is their ecological role? Are they an integrated part of it? 375. Does the complexity of ecosystem processes increases or decreases across ecological and spatial scales, and is the complexity of predicting them increase alongside? 376. What are the effects of overpopulation, degradation of global ecosystems towards speciation processes and species’ adaptive responses? 377. How can we model feedbacks and interactions between abiotic and biotic ecosystem/landscape components (under non-stationary conditions)? 378. Which factors and mechanisms determine the resilience of ecosystems/landscapes? And how can we better manage them in order to reliably provide ecosystem/landscape services? 379. How do tightly-linked coevolving organisms (mutualisms, pathogens) shape communities and ecosystem processes? 380. How deterministic are ecosystem dynamics? 381. What are the relative importances of abiotic stochastic factors versus deterministic ecological interactions in shaping ecosystem dynamics? 382. How do we consider spatial relationships in ecosystems? 383. How can ecosystem functions be maintained as communities change due to altered abiotic conditions and biotic interactions? 384. How much the enhancenment of connectivity between habitat areas can compensate for habitat loss over different spatial and temporal scales and for different types of ecological processes and ecosystem services? 385. How important are the dynamics of individual traits and life histories in understanding population and ecosystem change in the wild? 386. How can we predict the ecological / ecosystem effect of eliminating a species? 387. What role does biodiversity play in ecosystem resilience? 388. How vulnerable/stable are ecosystems in the face of environmental change? Can tipping points be identified in ecosystems before they are passed? 389. Are ecosystems subject to real “tipping points” the exception or the rule? How real is irreversibility in ecosystem change? Under what circumstances does it occur? 390. Are more or less ‘natural’ ecosystems more resilient to change? 391. Will the complexity of interactions that occur in ecosystems always prevent us from making predictions that are reliable beyond the short term? 22 392. What role do transients play in the dynamics of ecological systems? 393. Ecologists focus on differences between systems, and how those processes change over time. What kind of processes are consistent within a particular habitat across different datasets (e.g. a woodland), and yet do not exist in other systems (e.g. a grassland). 394. To which extent might tipping points in the earth system be influenced by terrestrial vegetation? 395. What are the key biosphere-atmosphere interactions that affect climate and air quality at multiple spatial scales? 396. How dependent on hydrological regimes is the ecology of seasonal wetlands (especially poorly known are dune wetlands & mountain flushes) and what are the implications of climate change? 397. At what scale does geology replace ecology in explanations of biodiversity and ecosystem function? 398. How much do you need to know to predict how ecosystems function across spatiotemporal scales? 399. Are ecosystems significantly impacted by increasing environmental variability? 400. How can interactions besides feeding and pollination be included in ecological networks, e.g. transmission, dispersal, immunity? 401. Are there common themes in resilience? 402. What is a resilient ecosystem? 403. What are good metrics for resilience? 404. What is the relation between the structure of interconnections and the way they behave – within an environment? 405. Are there general patterns in ecosystem perturbations? 406. Can we acurately describe an ecosystem? Empirical description at all spatial and temporal scales. 407. Are there real differences between biomes? Are there regularities in ecological systems between marine, freshwater and terrestrial systems. 408. How do you measure the complexity of an ecosystem? 409. How will ecosystem services be affected by global environmental change? 410. How is ecosystem function altered when biodiversity is lost through realistic extinction scenarios? 23 411. Will ecosystems that have lost species or those with a high proportion of introduced species respond differently (from those that haven't) to environmental change? 412. How does biodiversity affect the functioning and stability of natural ecosystems 413. How does spatial scale affect the relationship between biodiversity and ecosystem processes? 414. How much diversity is enough? How much diversity is needed at landscape scales to maintain ecosystem processes at local scales. 415. What is the relative contribution of biodiversity (species richness, species identity, functional identity, functional diversity) to ecosystem functioning, relative to that of conditions (climatic or microclimatic) and resources (nutrients, light)? How do these different contributions operate? 416. How does the form of the biodiversity-ecosystem functioning relationship change in different contexts (e.g. environmental change scenarios)? 417. What is the effect of species richness and diversity on ecosystem functioning? 418. Question of importance of diversity for functioning (and stability) – again, can be considered at various temporal and spatial scales. Should include genetic diversity of a population, but also (species, phylogenetic, functional) diversity of a community. 419. What is the shape of relationship between biodiversity and ecosystem functioning? 420. How does the relationship between biodiversity and ecosystem function vary with different functions? 421. What measures of biodiversity best characterise relationships to ecosystem function? 422. Have we focused research too much at the species level? Does it matter how many species there are for ecosystem function? 423. How substitutable are species (in terms of ecosystem function/services/cascades) and can we predict this in advance (i.e. can we predict which ones we can afford to lose/can we predict the effects of losing particular species)? 424. What is the differential sensitivity of different ecosystem functions to the loss of different taxa or different functional groups for different types of ecosystems? What mechanims explain such differences (or similarities)? 425. What are the effects of biodiversity change (species loss, change in species taxonomic or functional composition) on ecosystem functioning and the ability to deliver ecosystem services at different spatial scales (ranging from laboratory 24 experiments to plots, landscapes, municipalities, states or countries)? What key processes operate at these different spatial scales in relation to such effects? 426. How does the structure of ecological networks affect ecosystem functioning? 427. What ecosystem services are provided by poorly known ecosystems? (E.g. deep oceans, groundwater) 428. How much does genetic variability contribute to ecosystem functioning? In what ways does this variability play a role in what we perceive as “ecology” 429. What is the relationship between genetic diversity and ecosystem functioning? 430. What is the role of biodiversity in determining the resilience of ecosystem function in rapidly changing environments? 431. How reliable are financial valuations of ecosystem processes and ecosystems? 432. How importance(t) is species rarity for ecosystem functioning? 433. In ecosystems that are effectively novel, due to recent anthropogenic extinctions of some species and/or invasions of others, what are the essential functional units of the food web that have to be filled (with exotic species if necessary) to restore ecosystem services? 434. What is the best way to assist migration of species to maintain ecosystem function? 435. Is tree species identity important regarding the provision of a wide array of ecosystem services from forests (e.g., C storage, timber production, biodiversity for biota other than trees, protection from avalanches and rockfall, etc.)? 436. How much does biodiversity contribute to ecosystem services? 437. How much does an individual species or species group contribute to ecosystem function? 438. What are the critical levels of ecosystem stocks necessary to maintain ecosystem services? 439. What is the relative importance of top down versus bottom up processes in regulating ecosystem function? 440. Do rare species matter to ecosystem functioning? 441. To what extent are rare species important in ecosystems? 442. Do species matter (more than any other component of biodiversity) for ecosystem functioning? 443. Can technology replace most (or all) ecosystem services? 444. When is biodiversity unimportant and what are the minimum levels we can tolerate with reasonable function maintained? 25 445. How do we integrate between spatial scales and understand how spatial structure influences ecosystem function? 446. What do we need to measure to know what the function of a component organism of an ecosystem does? 447. To what degree is it possible to describe ecosystem functions (in space or time) from partial knowledge? 448. Are the relationships between ecosystem functions the same in different environments? 449. What are important ecosystem functions in coastal systems? 450. How do we identify the thresholds limits in number of species, number of particular functions? 451. How can human knowledge obtained from domesticating natural ecosystems, and managing ecosystems, best be used to improve ecological theory? 452. How important are rare species in the structure and functioning of ecological communities? 453. How can we predict the response of complex systems to perturbations when we have incomplete knowledge? 454. Are productivity and network complexity related? 455. Can we predict or understand when do tipping points exist? Or: Can tipping points be predicted and if so how? 456. Is it important that models of complex ecosystem can be stable? What are the relationships among the many existing mechanisms (phenomena) that can stabilise complex ecosystems? What is the relationships between complexity and stability? 457. Can we predict the responses of ecosystems to land use change based on the traits of species? 458. How does biodiversity influence ecosystem stability? 459. What are the effects on ecosystem function of large-scale extinctions? 460. How can we reliably predict critical thresholds/tipping points in ecosystems? (e.g. with respect to the effect of biodiversity loss on ecosystem functioning) 461. When does if matter about the individual species for ecosystem function? NB taxa vs. function? 462. How can ecological complexities, such as the resilience of natural capital stocks, be incorporated into the evaluation of natural capital and ecosystem services? 26 463. Does the phenomenon of critical slowing down as an indicator of the proximity of tipping points translate from microcosm studies to field ecosystems? 464. How does soil biota modify the physical environment, and so impact the flow of matter and energy and the delivery of ecosystem goods and services? 465. If a priority is given to restoring rivers to reference conditions as defined under the Water Framework Directive, what would be the trade-offs involved between different ecosystem services 466. How do the stressors of flow alteration, nutrients, sediments and channel morphology combine and interact to define the conditions for aquatic and wetland species? 467. When is the identification of cryptic species crucial for our understanding of the functioning of ecosystems? 468. What are the implications of climate change induced reductions in mean fish size for their roles in lake ecosystems? 469. How do we measure ecosystem services and their link to ecosystem structure and function? 470. What are the specific components of ecosystem resilience under greatest threat from agriculture and what are the consequences of loosing this resilience in terms of ecosystem services? [Bruce Howard] 471. Is there anything inherently different about the biogeochemical cycles of the northern and southern hemispheres? 472. Do ecosystem functions evolve through time? 473. Does ecosystem disassembly follow the same rules as assembly? 474. What traits of organisms, are important in determining different ecosystem properties? 475. What are the relationships between multiple measures of stability? 476. What makes a species important? Can you predict the importance of species for stability or functioning of ecosystems based on its traits? E.g. position in a food web, phylogeny, body size? 477. Can we predict ecosystem dynamics? Can we model the dynamics of real ecosystems with all their component parts? 478. Acidification vs warmer temperatures in coral and in primary producers in general: which one is the worse problem, and do they interact? 27 479. How important is adaptation by natural selection in mediating species responses to climate change? 480. How will whole ecological communities respond to climate warming and other aspects of global change? Can information about species responses be scaled up to understand whole-community responses? 481. How do we develop a robust mechanistic understanding of empirical links between climate change and long term changes in species distribution and abundance ? 482. Will species with TSD go extinct in the face of projected climate warming ? 483. Will climate change cause mass extinction? 484. Will climate change uncouple trophic links due to phenological change? 485. What impact is climate change/ocean acidification having on benthic primary production? 486. How are responses of primary consumers to climate change/ocean acidification altered/mediated by changes in composition/biomass of primary productivity? 487. What potential maladaptation issues can arise from climate change adaptation measures (e.g. artificial coastal defences)? 488. Is attribution necessary in climate change impacts science? 489. How can we link local responses of individuals, populations and species to global climate change? 490. Given current and imminent climate change and our ongoing history of intervention, management, translocation and introduction of non-native species and genotypes, how necessary is it to ensure the conservation of observed patterns of genetic variation? 491. How does the role of plant-plant (or other biotic interactions) change in relation to prevailing environmental conditions such as climate? Is this predictable? 492. How do natural communities cope with or even adapt to increased frequencies of extreme weather events predicted under global climate change? 493. What is more important for biodiversity loss: land use or climate change? 494. How will changes in climate affect primary productivity dynamics around the globe? (bearing in mind that climate change is linked to changes in averages, seasonal patterns and occurrence/severity of extreme climatic events; and that changes in primary productivity dynamics could involve changes in average biomass production, changes in seasonal patterns in primary productivity and/or changes in level of inter-annual variability in primary production). 28 495. How will the interactions between land cover modifications, climate change and socioeconomic transformations drive the patterns of occurrence and intensity of wildfires and their ecological impacts in the future? 496. The premise that climate change will uniformly affect species distributions is known to be false because of the apparent unpredictability of climate-mediated changes to species interactions in food webs - so , how can applied ecologists predict how climate change will affect species interactions in food web dynamics? 497. How & how fast will species distributions change with climate change? 498. How does climate change shift species' distributions? 499. Can we predict future distribution patterns based on climate change projections? 500. Which aspects of climate change are likely to lead to widespread tree mortality, and what are the underlying mechanisms? 501. How will climate change influence primary production (the underpinning of all ecology)? 502. How will change in land-cover feedback on the climate (e.g. through combined effects of changes in carbon sequestered and changes in albedo)? 503. With reference to climate change is the term “adaptation” a source of serious and even dangerous misunderstanding? 504. What is the role of interspecific interactions in buffering species responses to global change? 505. Comparison of recent global changes to those during the last glacial period. I am not sure how to formulate this as a question that would not sound too trivial. Nevertheless, I believe that we can get a perspective for what is going on from the changes that happened from the last glacial maximum (or perhaps, from the whole quarternary). 506. How sensitive are organisms to changes in ocean pH/ocean chemistry? 507. Do soil microorganisms represent a significant sink for atmospheric CO2? 508. How much biodiversity are we loosing and how much will we lose (assuming rate of loss will slow as vulnerable species/habitats are converted)? 509. How many species do we really have, considering those that were already lost and those that might already start evolving (as a dynamic process)? How many species did we loose? 510. At what rate are we discovering new species compared to the number of species under threat/or near extinction and how has this changed over the past decade? 29 511. In the face of rapid environmental change, what determines whether species adapt, shift their ranges and/or go extinct? 512. What are the long-term impacts of invasive species worldwide? 513. The modern world presents animals with challenges such as windows, light pollution, cars ... we often study adaptation to pollution etc., but do things ever adapt to these urban sources of trouble? 514. What are the ecosystem level effects of ocean acidification? 515. How successful are protected areas when surrounded by human settlement? 516. How can the impacts of human activities be separated from environmental impacts for commercial species? 517. Given current/projected per capita rates of consumption, at what critical regional or global human biomass do efforts in conservation become no longer viable? 518. Is human population optimally distributed? 519. Does the well being of a nation increase or decrease the probability of humanwildlife conflicts? 520. Which silvicultural treatments and other management practices in human-made or seminatural forests (and other woodland habitats) can promote the adaptation of plant and animal communities to changing environmental conditions and increasing societal pressures? 521. How does biodiversity change impact on human health and well-being? 522. How much biodiversity would be left in a world where the whole human population were adequately fed? 523. Does harvesting result in human-induced evolution of harvested species? 524. How do the destruction and fragmentation of habitat affect biodiversity? 525. Is “fishing down the food chain” fact or fiction ? 526. What is the level of bycatch in fishing gear for endangered marine animals and how can it be mitigated ? 527. What effect will loss of land races of agricultural crops have long term? 528. Can we restore biodiversity? (non-existing species cannot be brought back) 529. How do genetically modified organisms affect biodiversity? 530. Which of genetic, life history or food web constraints are more relevant to predicting and detecting responses to environmental change? 531. How do we understand ecosystems of which humans are a part? 532. Does ecology exist in urban environments? 30 533. Does ecology only exist in pristine areas? 534. What is the effect of history on current and future biodiversity trends? 535. What caused the discontinuation of lemming cycles in northern Europe? 536. How do we frame ecological conservation objectives for fragile ecosystems, such as the arctic, which are under stress from development and climate change pressures? 537. Will high atmospheric CO2 lead to a surge in ecosystem productivity and/or a reduction in water stress in natural and disturbed ecosystems? 538. How will ecological communities respond to high atmospheric CO2? 539. How will tropical forests respond to warmer temperatures? What is the thermal niche of tropical forests? 540. Will climate change ultimately reduce or increase species richness? 541. How does warming alter the structure and stability of communities? 542. How will warming affect species loss? 543. How will species demographic rates be affected by global warming? Warm versus cold blooded species. 544. How influential will the genome, epigenome and second genome (i.e. symbiogenome) be in controlling local adaptation to climate change? 545. What will be the magnitude of the terrestrial carbon sink in 50 and 100 years time? 546. What is the legacy of human disturbance on the structure and composition of tropical ecosystems? 547. How will climate change alter disturbance regimes? 548. What geographic and biodiversity gaps remain in the global terrestrial protected area system? 549. Can we find a way to support long-term ecological research (individual-based field studies) in the tropics, given the high levels of tropical diversity and disproportionate threats. Funding should be allocated outside of the typical 3-5 year grant schemes, allowing consistent data collation. 550. Do non-agricultural and semi-natural habitats and communities contain a genetic capital that could be used for crop/ livestock improvement? 551. What is the most important social change within the ecological community to affect positive decisions in society? 552. Can we pretend that human activity is just like that of any other member of an ecosystem and analyse "us" in the same way as other keystone species? 553. Why is ecology (and ecosystems) confusing to law makers? 31 554. Is the change from a legal to an economic epistemology likely to reduce extinction rates? (or) Is ecosystem services thinking dangerous? 555. What can we learn from experimental systems? Do we need the ecological equivalent of the geneticists C.Elegans? Should we be striving for a single simple system against which we can test our ideas and understanding? 556. What is the most appropriate baseline for determining future change in status of marine systems (for policy directives such as the MSFD Good Environmental Status, MPA assessments etc.)? 557. What are the key species required to maintain human persistence? 558. How can be used sound and well-tested ecological theory to decrease poverty and improve human welfare? 559. Overpopulation has disrupted ecological stability, short-term and long-term negative impacts (will) affect humans, thus the costs of loosing biodiversity are high? So, what are the ultimate sustainable strategies to shift to a stable culture for humans and the biosphere…?? 560. When is it appropriate for human interactions with ecological systems to be integrated into ecological models rather than treated as an external driver? 561. Which ecological theories or generalisations have actually proved useful when applied to human management of the environment? 562. Should we draw limits to the scope of the discipline of ecology and if so where (e.g. with respect to the totality of ways in which human beings interact with their environment, or with respect to physiological and other processes occurring within organisms, geochemical cycles in atmosphere, earth and ocean, hydrology etc.)? 563. To what degree should methods drive progress in ecology? 564. To what degree is progress in ecology constrained by the focus on experimental approaches? 565. What more can ecology learn from economics? 566. Is it time for the peer-review model of science and academia in general to evolve and possible increase transparency particularly to the public? 567. What ecological concepts and terms have proved most useful in knowledge exchange with people who have not had a formal education in ecology? 568. How important is it for ecological research to seek to bust commonly held “myths” about ecology? 32 569. Which ecological myths should we try hardest to remove from formal educational curricula? 570. What kind of ecology do we want? 571. Is there such a thing as pure ecology? 572. Can we afford to keep biodiversity? 573. What forms of response do species take to drivers of change? 574. Do ecologists need to be natural historians? 575. Does ecology matter to conservation when the problem is human behaviour? 576. How is our understanding of ecology influenced by publication bias? 577. Are there landscapes/communities/taxonomic groups that we have not focused on? 578. Are there theories/concepts in ecology that we should abandon? 579. Do we have the tools and education programme to train the current and next generation of ecologists in quantitative (maths and statistics) methods that can answer major questions? 580. Would improved ecological education of the population improve environmental decision making by our leaders? 581. Is taxonomic bias (everyone loves big mammals) useful or dangerous to engaging the public in ecological issues? 582. How can ecologists get long term funding for long term projects? 583. Under what conditions is ecology a predictive science? 584. Are there generalisations (general laws) in ecology? 585. Can simple models really be informative for specific ecological problems? 586. What advances can ecology make by harnessing the power of modern technology (sensing, informatics, computing)? 587. Is the generalised shape of stressor-response relationships appropriate to all situations? 588. Can the distribution and flow of information help integrate ecology with biology and physics? 589. Why is it that so many people (including “ecologists”) cannot discriminate between ecology as a science and simple surveying / counting? 590. Plant ecology still largely lacks any significant predictive powers. I think this is true at a range of scales from the individual to, populations to communities. The subject is still largely descriptive, all be it in a much more quantitative way than 30 years ago. You could argue, as Grime C-R-S does, that his concept of plant strategies/ 33 functional types was capable of providing an element of predictability, but I don’t believe this approach has been successful. 591. What laboratory systems can be developed and usefully canonized as broadly useful model systems for population and community ecology, in the same way drosophila, e. coli, and yeast, for instance, have been canonized as broadly useful systems in other fields of biology? 592. How do we begin to model the complexity of the natural world? What types of model / interactions should be assumed? Is a reductionist approach where we only consider 1,2 or 3 interacting species ever going to be successful? 593. Are ecologists’ efforts to construct a pseudo-physical unified theory productive – or are they bound to fail since biological systems are too complex to be reduced to a handful of physical laws? 594. Is there a unifying theory of ecology? 595. Body size of an organism is strongly correlated to many physiological and ecological properties, and can be used to help understand population-, community-, ecosystem-, and global-scale phenomena. What additional organism traits must be incorporated into models to more fully understand phenomena at higher levels of organization? 596. Are there universal ecological models, or does each ecosystem and species require its own formulation? Are there basic well-formed building blocks that will allow us to create functioning models. 597. Can we afford to degrade biodiversity further? 598. Can we replace palm oils? 599. Is our system of democracy capable of delivering good environmental outcomes? 600. Can we identify commonalities of where predictive ecological models have worked? 601. Comparative demography: is there a population economics spectrum (very much along the ways of the leaf economic spectrum idea)? 602. Considering population ecology relates to species across social and catchment boundaries, why do we continue to have many scientists working on the same or similar topics in departmental silos across states and countries? 603. How can the proportion of students studying ecological disciplines at university be increased? 34 604. How can market funds be channeled into on-the-ground protected area monitoring and protection? 605. What political, economic and social instruments are required to curtail the illegal wildlife trade? 606. What is the effect of changing food availability for consumers and how can we make quantitative predictions of this? 607. What is the most effective way of combining observations and experiments in improving ecological knowledge? 608. Can ecology be a truly predictive science? (i.e. useful ability to generalise from feasible study effort to novel situations) 609. How effective has improved knowledge of ecological mechanisms been in our capacity to predict future change? 610. How reliable or accurate have past ecological predictions been and on what basis have these predictions proven true (i.e. from lab or field experiments, modelling, theory or collective research)? 611. How can the culture of ecology as an academic discipline be modified to make progress more rapid and to increase the usefulness of ecology for society? 612. How can the review process be modified to increase the quality and usefulness of outputs? 613. What is the error rate of published ecological literature, what is the nature of published errors, and how do errors affect academic understanding and policy? 614. What can policy ever learn from realistic ecological models, since they do not contain the "unknown unknowns"? 615. How can we measure the ‘importance’ of an ecological process? 616. Is an environmental Kuznet’s curve at the global scale possible? 617. How does biodiversity contribute to human mental health and well-being? 618. In view of frequent gaps between optimization models and experimental data, should we re-think the concept of optimization, currently presumed an axiom in behavioural ecology? 619. Do ecological laws apply to other domains (banking, internet, memes, exobiology)? 620. Are undergraduate ecology curricula appropriate? 621. Which ecological generalities apply to humans? 622. How important is improved ecological understanding of the natural mechanisms enabling species co-existence to improvement of our conservation of biodiversity? 35 623. Ectotherms appear generally to be getting smaller as a consequence of climate change in aquatic systems. What are the causes of this, how universal is it, and what are the consequences for biodiversity, community structure and ecosystem function. 624. Can we understand the ecology of organisms on other planets etc.? 625. What are the statistical consequences of model misspecification, and how do they affect ecological theory and understanding, predictions about the future states of ecosystems, and policy development? 626. Are mixed effects models the panacea people think they are for ecology. 627. How can we best produce predictive models that encapsulate different aspects of ecological understanding, so that they can be rapidly combined into accessible tools for exploring multifaceted scenarios? 628. Does modelling the observation process inherent to any ecological investigation improve our ability to infer population processes? 629. Can ecological models ever be truly predictive? Consider the SORTIE model of Pacala and co-workers, or the Florida panther model developed by Gross and colleagues; even these highly sophisticated models fail to capture the full dynamics and are far from being predictive. This obviously links to questions on how ecological modelling can be used in practical situations. 630. How do we begin to parameterise ecological models? This is made much more complex by the number of interacting species, the range of different model choices and the fact that we generally only observe populations close to equilibrium (such that non-linear effects are difficult to measure). 631. Can we develop a robust, scale-explicit and non-ambiguous measure for comparing species diversity between locations and systems – in order to replace the scalesensitive, ambiguous and misleading β-diversity? 632. Can we employ technology to count species and individuals. 633. What are the relative prospects of success of stochastic versus mechanistic predictive ecological models? 634. What are the roles of sampling methods in understanding ecology? 635. Is hysteresis the exception or the norm in ecological systems? 636. What do species do? My reasoning is that for most species, irrespective of how common they are, we simply don't know. And when we do, its for one or two processes only. Knowing, I would argue, is fundamental to everything else. 36 637. Some fish stocks are now bouncing back after overexploitation. Are the prospects of this happening impacted by evolution? 638. Evolutionary theory predicts that conspecifics often benefit, evolutionarily speaking, by developing traits that do not benefit the group (Rankin et al. 2007). But does this really show: How often does intraspecific conflict have a significant impact on population growth? 639. Does microevolution play an important role for the success of a species on an ecological time scale? 640. How, and at which spatial and temporal scales, do we integrate the ecology of species interactions with the geographic mosaic theory of coevolution? 641. How do we best include rapid contemporary/ongoing evolution and coevolution in ecological studies? 642. How do evolutionary and ecological dynamics interact? 643. To what extent can we ignore evolution in addressing a specific ecological question? 644. Does evolution/coevolution increase (or reduce) the complementarity and productivity of communities? 645. What are the relative potentials of plasticity and evolution to enable life history adaptation, and how do those vary among traits and species? 646. Does ecological and evolutionary change always occur simultaneously? 647. How important is short-term evolutionary change in determining the response of ecosystems to environmental change? 648. Will evolutionary rescue play an important role in natural populations? 649. Do extinction times have an activation energy? 650. What features of animals allow them to deal with warmer temperatures? 651. Is it true that negative environmental impacts on bigger species (those of larger body size) are, all else equal, more damaging due to life history traits that associate with large bodies — except maybe in fish?? 652. How do fecundity, body size and generation time relate across taxa? 653. What is the relative contribution of different levels of selection (individual, kin, group, lineage, etc) to life-history evolution and the resulting population dynamics? 654. What drives mate choice, and how can this impact on conservation? 655. How significant is the fact that early life conditions influence adult phenotype? Do epigenetic effects play any role at the ecological level? 37 656. Does genetic variation within a species matter to ecologists? 657. Demo-genetics: are assumptions currently being made on population models regarding the contribution of "mothers" at time t to "babies" at t+1 correct from a genetic standpoint? 658. How does epigenetic variation impact a population's resistance to disturbance and environmental change? 659. What are the magnitudes and durations of cross-season carry-over effects on lifehistories? 660. What are the intrinsic covariances/trade-offs among major life-history traits (agespecific reproduction and survival) and to what degree do they constrain elasticities, sensitivities and population growth rate? 661. How do operational sex ratios compare to offspring sex ratios in species with temperature dependent sex determination ? 662. Are behavioural syndromes important? 663. How can we measure fitness in fungi? 664. Is there an integrative long-term measure of fitness? Would one make sense in the context of temporally varying selective environments? 665. How accurate are estimates of baseline population (i.e. pre-exploitation) levels? 666. How does genotypic variation determine ecological performance? 667. What are the limits of phenotypic plasticity for adaptation to environmental change (common gardern experiments, transplant experiments)? 668. Should we worry about the consequences of conservation actions that mix different (locally adapted) gene pools of a single species? Do these management options represent best practice in conservation? 669. Will species specialize, adapt and evolve to urban environments? If so how and when? 670. Is adaptation to urban environments an evolutionary process? Are abundance & body size relationships constraining that: large-bodied individuals have small population? 671. Is there a selective pressure to be cute, given how much conservation relies on the values people hold? 672. Are co-evolutionary processes important for population responses to environmental perturbation? 38 673. When is trait variation more important than rapid evolution in response to environmental change? 674. What are the traits of migrant insects vs. residents and birds? 675. What ecological questions are best suited to molecular data? 676. What molecular methods are best suited to ecological questions? 677. Do subpopulations within meta-populations share adaptive variation evenly, or do they represent adaptive mosaics? 678. What is more important, climate or ecology? When climate drives recruitment, can processes such as predation or competition overwhelm the climate signature. 679. What explains life history differences in autotrophs globally? 680. How do evolutionary changes affect structure of density-dependence? 681. What are the magnitudes and mechanisms of sex-specific genetic variation in fitness? 682. What is the genetic basis of variation in fitness? 683. How important is individual variation for the outcome of ecological interactions? 684. How do species traits and the environment contribute to determine ecological network structure? 685. Why so many species? (Or, why not more species?) 686. Why THIS level of species richness at a given site? 687. Is there a way to predict, from first principles, how many species can coexist in a given area? 688. Why are there so many species? 689. To what extent does competition influence species distributions? Do assembly rules exist? 690. How relevant are assembly rules in a world of biological invasion? 691. Are there community assembly rules? 692. What is / is there a saturation level for the number of species within trophic levels? 693. What is the amount of vacant ecological niche space? 694. Why are there so many vacant niches? 695. Does niche partitioning exist and if so does it contribute to species coexistence? 696. Are most plant communities unsaturated with species? 697. Does coexistence of species in plant communities depend on forces (niche partitioning, limiting similarity or complementarity) that prevent coexisting species from being too similar? If so, what are the key traits? 39 698. The humped-back model of herbaceous plant diversity (a unimodal relationship between annual production (standing biomass and litter) and maximum species richness) is the most frequent relationship in natural plant communities. Are there particular circumstances under which such a relationship is not found? 699. Why do abundance distributions in natural communities conform so closely to the predictions of neutral theory when so often a clear competitive “winner” emerges when species encounter each other in pair-wise fashion? 700. What are the relative contributions of stochasticity, species interactions and environment in governing the composition and dynamics of plant communities and how does this depend on species richness? 701. Which mechanisms stabilize species rich communities? 702. To which extent is species composition (and in consequence also the diversity) controlled by available species pool (or dispersal limitation), and to which extend by interaction within community. It is clear that the answer will depend on spatial scales of comparison (i.e. on the grain, i.e. size of community, and on the extent, i.e. size of area within which we compare individual “communities”. There is a similar question (because the species pool is to a large extent determined by historical phenomena, to which extent are recent communities shaped by recent ecological interactions, and to which extent by historical phenomena (which can include both, the long-term evolutionary history, but also the postglacial migration). 703. Can trait-based approaches help us to predict shifts in community structure (composition, abundance) following environmental change? 704. What allows species to co-exist, and a secondary but related question, are their emergent functional properties that occur when certain collections of species coexist? I think this remains largely un-resolved. 705. To what extent do stochasticity and environmental noise break deterministic plant interactions, and how does this depend on species richness? (If the biotic neighbourhoods and environmental conditions experienced by individuals are is highly variable species may converge on similar generalist life history strategies). 706. Does stochastic niche theory explain biodiversity patterns better than niche or neutral theory of biodiversity? 707. Are simple tradeoffs the answer to multispecies coexistence? Are more complex tradeoffs required? 40 708. Can we determine the biodiversity properties (e.g. species-area relationship) of a large area from spatially structured “point” surveys? 709. What is the best way to configure a given amount of total habitat if we are to maximise the number of habitat-dependent species that we can conserve – both in the short term and in the long run? 710. Under what circumstances (and at what scales) do temporal and spatial environmental heterogeneity most increase (or decrease) diversity? 711. Why is there usually a positive correlation between range and local abundance in plants? 712. Does coexistence require space? How much does space heterogeneity affect coexistence, and how does this vary across spatial scales? 713. How do species coexist? 714. What is the relative importance of stochastic versus deterministic processes in controlling diversity and composition of communities, and how does this vary across ecosystem types or across trophic levels? 715. Does an ecosystem have any ‘vacant’ niches? If it has, how many and why does it have them? 716. How do we explain species co-existence across scales? (current theories, including niche and neutral theory, clearly do not provide the answer) 717. Why do some cities maintain so much more life than other cities? 718. How many undescribed species are there out there? 719. Are species numbers increasing or decreasing? 720. How does soil biodiversity influence plant diversity? 721. How far does counting species take us? 722. How do the relative importance of niche and neutral processes varies among communities? 723. At what stage(s) in forest succession does the species density peak for arthropods, nematodes, fungi and microbes? 724. What traits govern coexistence? 725. Are the traits that govern coexistence in soil, plants and animals similar? 726. How do major life history trade-offs, common to most organisms result in such immense diversity? 727. Does diet choice, habitat choice or mate choice govern the structure of ecological communities? 41 728. What aspects of community composition are predictable in space and time? 729. What stabilises consumer resources antagonistic interactions? 730. How can many species evolve and coexist in an seemingly unstructured (marine) environment? 731. What is the role of community traits in shaping the environment? 732. How many species does it take to form a community? 733. How do ecological networks arise and how do we best describe them? 734. Why is species diversity often apparently higher than niche diversity and why are hyper-diverse regions often phylogenetically overdispersed? 735. How can earth observation and field monitoring data be used to monitor biodiversity at multiple geographical scales? 736. The CBD 2010 targets were for reductions in the rate of biodiversity loss, a good ecological indicator yet we could not measure this with any confidence. Can we develop more consistent sampling of global biodiversity and derive more precise and measurable targets that could facilitate delivery of ecological aims? 737. Why are the tropics so diverse? 738. Which theory best explains the latitudinal gradient? If more than one theory is plausible – can we assign weights to the role of each explanation? 739. Is ecological diversity a self sustaining phenomena? 740. Are southern temperate forests more species rich than northern temperate forests? 741. What happens to a species' niche once the species goes extinct? 742. How many species could the world theoretically support? I.e., how saturated is the world? 743. Is plant diversity the main driver of wider community diversity in tropical forests? 744. What are the ecological mechanisms contributing to high biodiversity in the tropics? 745. Do latitudinal diversity gradients result primarily from local-scale ecological processes (e.g., strength of density-dependent competition or predation) or largerscale evolutionary / biogeographic processes (e.g., differential rates of speciation, colonization or extinction)? 746. How can so many species co-exist? This seems to be an eternal, but still not fully solved question. I expect that the effect of pathogens (and, albeit to a lesser extend also predators s.l.) will be found as most important, but there will be undoubtedly other mechanisms of “increase when rare”. I believe that we need some progress in ecological theory here, but mainly, the mechanisms suggested by the theory should 42 be demonstrated experimentally. The question might be formulated also as “What are the mechanisms maintaining the species diversity?” Part of the question can also be the differentiation of traits, on consequently, the relationship between the species diversity, functional diversity, and phylogenetic diversity. 747. How do we explain latitudinal gradients in species richness? 748. Why are there more species in the tropics than in temperate climes? 749. Do latitudinal diversity gradients result primarily from local-scale ecological processes (e.g., strength of density-dependent competition or predation) or largerscale evolutionary / biogeographic processes (e.g., differential rates of speciation, colonization or extinction)? 750. What are the mechanisms behind the latitudinal gradient in biodiversity? 751. To what extent phylogenetic diversity does reflect functional-group diversity? 752. What proportion of biodiversity is deterministic versus based on random processes? 753. To what extent are communities a function of the external environment (abiotic /biotic environment) 754. At what scale does ecology become too specific to predict regional and global patterns of diversity? 43