COX-2 overexpression increases bone metastasis of MDA
advertisement

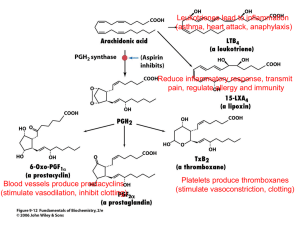
Revised MS#ONC-2006-00864 (marked copy) Original Article COX-2 involvement in breast cancer metastasis to bone Balraj Singh1, Jacob A. Berry1, Angela Shoher2,3, Gregory D. Ayers4,5, Caimiao Wei4, and Anthony Lucci*,1 1 Department of Surgical Oncology and 4Department of Biostatistics & Applied Math, The University of Texas M.D. Anderson Cancer Center, Houston, TX 77030, USA; 2Baylor College of Medicine, Houston, TX 77030, USA. *Correspondence: A Lucci, Department of Surgical Oncology, Unit 444, The University of Texas M.D. Anderson Cancer Center, 1515 Holcombe Boulevard, Houston, TX 77030, USA. Phone: 713-563-1871; Fax: 713-792-4689; E-mail: alucci@mdanderson.org 3 Current address: Johns Hopkins University School of Medicine, Baltimore, MD, USA. 5 Current address: Department of Biostatistics, School of Medicine, Vanderbilt University, Nashville, TN, USA. Running title: COX-2 in breast cancer metastasis to bone B Singh et al Abstract Cyclooxygenase-2 (COX-2) is expressed in 40% of human invasive breast cancers. Bone is the predominant site of metastasis in case of breast cancer. We investigated the role of COX-2 in a suitable mouse model of breast cancer metastasis to bone using the whole-body luciferase imaging of cancer cells. We provide several lines of evidence that COX-2 produced in breast cancer cells is important for bone metastasis in this model including 1) COX-2 transfection enhanced the bone metastasis of MDA-435S cells, and 2) breast cancer cells isolated and cultured from the bone metastases produced significantly more prostaglandin E2 (an important mediator of COX-2) than the parental injected cell populations of breast cancer cells. Next, we found that a COX-2 inhibitor MF-tricyclic inhibited bone metastasis caused by a bone-seeking clone both in prevention regimen (in which case mice started receiving MF-tricyclic one week prior to the injection of cancer cells) and in treatment regimen (in which case mice received MFtricyclic after the development of bone metastasis). These studies indicate that COX-2 produced in breast cancer cells may be vital to the development of osteolytic bone metastases in patients with breast cancer, and that COX-2 inhibitors may be useful in halting this process. Keywords: breast cancer, COX-2, mouse model, MF-tricyclic, bone metastasis, cancer prevention. 2 B Singh et al Introduction Cyclooxygenase (COX) enzymes, COX-1 and COX-2, mediate the production of prostaglandins and thromboxanes from arachidonic acid. COX-1 is expressed constitutively, whereas COX-2 is induced in inflammation and cancer. The extracellular stimuli that induce COX-2 include growth factors, cytokines, tumor promoters, hypoxia, ionizing radiation and carcinogens (reviewed by Singh and Lucci, 2002; Howe and Dannenberg, 2003; Wang and DuBois, 2004). COX-2 is overexpressed in most human epithelial cancers and has been linked to various aspects of cancer progression both early and late in the process. Epidemiological studies have reported a significant reduction in the incidence of human gastro-intestinal cancers with COX inhibition by NSAIDS (Shaheen et al., 2002). Although the evidence for breast cancer prevention is less strong, a recent meta-analysis of 14 studies (six cohort, eight case-controlled) gave a combined estimate of a reduced relative risk of 0.82 (95% confidence interval 0.75-0.89) with regular NSAID use (Khuder and Mutgi, 2001). There is evidence to implicate COX-2 in breast cancer tumorigenesis. One study showed that 56% of infiltrating mammary carcinomas and intraductal carcinomas expressed significant levels of COX-2, while benign breast tissue at least 1 cm from a malignant lesion did not express COX-2 (Soslow et al., 2000). In a murine model of metastatic breast cancer, PGE2 levels are positively correlated with increased tumorigenic and metastatic potential (Kundu et al., 2001). Perhaps the most convincing evidence that COX-2 causes breast cancer in animals comes from transgenic mice in which COX-2 was overexpressed in mammary tissue by using the mouse mammary tumor virus (MMTV) long-terminal repeat promoter. More than 85% of these mice developed tumors, indicating that COX-2 overexpression alone is sufficient to cause breast tumors (Liu et al., 2001). 3 B Singh et al Ristimaki et al. (2002) analyzed the expression of COX-2 protein by immunohistochemistry in tissue array specimens of 1576 invasive breast cancers. Moderate to strong expression of COX-2 protein was observed in 37.4% of the tumors, and it was associated with unfavorable distant disease-free survival. Elevated COX-2 expression was associated with a large tumor size, a high histological grade, a negative hormone receptor status, a high proliferation rate (identified by Ki-67), high p53 expression, and the presence of HER-2 oncogene amplification, along with axillary node metastases and a ductal type of histology. These results indicate that elevated COX-2 expression is more common in breast cancers with poor prognostic characteristics and is associated with an unfavorable outcome. We are investigating the mechanisms and biochemical pathways through which COX-2 contributes to various aspects of breast cancer progression, including metastasis. In this study, using a suitable mouse model of breast cancer metastasis, we show that COX-2 overexpression in breast cancer cells correlates with increased metastasis to bone. Starting with the heterogeneous populations of breast cancer cells, cells overexpressing PGE2 were favored at the sites of bone metastastases in the mouse model. Thus our studies provide further evidence for the role of COX-2 in breast cancer metastasis to bone. We also provide evidence that a COX-2 specific inhibitor, MF-tricyclic, halts breast cancer metastasis to bone in this pre-clinical mouse model. Results COX-2 overexpression increases bone metastasis of MDA-435S cells To determine whether COX-2 overexpression in breast cancer cells would enhance breast cancer metastasis to bone, we utilized a nude mouse model in which cancer cells are inoculated into the 4 B Singh et al left ventricle of female mice. Others have shown previously that MDA-435 cells yield efficient bone metastasis in this model, affecting 100% of the injected mice (Harms and Welch, 2003). These cells express a low level of COX-2 (Gilhooly and Rose, 1999). Therefore, to ask whether COX-2 overexpression would enhance bone metastasis (either kinetically or overall burden), we co-transfected MDA-435S cells with a COX-2 expression vector, pSG5-COX2 (encoding human COX-2 protein), and pEF1a-Luc-IRES-Neo vector (encoding luciferase protein and neomycin resistance). After selecting the stably transfected cells in 0.4 mg/mL geneticin, we analyzed them for luciferase activity and COX-2 expression. The pEF1a-Luc-IRES-Neo vector allows a longlasting and relatively high-level stable luciferase expression in the absence of geneticin and after injecting into mice. As shown in a western blot in Figure 1A, COX-2 protein level was significantly increased in COX-2 transfected cells as compared to luciferase-transfected MDA435S cells (compare lanes 1 and 2). The COX-2 band is not visible in control MDA-435S cells at this short exposure of the X-ray film. To obtain a better estimate of the COX-2 protein and its enzyme activity, we measured the production of a one of the prostaglandins, PGE2, in transfected cells. PGE2 is the major mediator of COX-2 in breast cancer cells. Both luciferase-transfected and COX-2 transfected cells were exposed to 10 M arachidonate for 30 min, and PGE2 was measured in the medium with an immunoassay. Of importance, the COX-2 transfected MDA435S cells produced significantly higher level of PGE2 (approximately 22-fold) as compared to the control cells transfected with the luciferase vector (Figure 1A, right panel). We conclude that the recombinant COX-2 protein is functional in the transfected cells. To determine whether COX-2 overexpression in MDA-435S cells would increase their metastasis in the mouse model of bone metastasis, we injected the left cardiac ventricle of two groups of 5 female nude mice with 0.1 million cells of either MDA-435S cells stably transfected 5 B Singh et al with luciferase alone (Figures 1B and 1D, left panels) or with luciferase plus COX-2 (Figures 1B and 1D, right panels). Starting at day 28, we quantitated the metastasis on a weekly basis until day 56 by whole-body luciferase imaging with the IVIS imaging system 100 (Xenogen Corporation, Alameda, CA). We measured the relative rate of bone metastasis in mice as the relative amount of light produced from the luciferase activity in breast cancer cells as radiance flux per second using Xenogen Living Image software (Figure 1B). Average radiance flux per mouse in mice injected with control and COX-2 transfected cells (data derived from Figure 1B) is graphed in Figure 1C for comparing the groups. The P-values of F-test of the fixed effects were: treatment 0.635, day <0.0001, day*treatment 0.284. The P-values for comparing treated vs. control on day 28, 35, 42, 49, 56 were 0.628, 0.265, 0.089, 0.064, and 0.078. Based on the observed data, on day 56, the mean and standard deviation of log-transformed radiance flux for the MDA-435S group were 2.79 and 0.34; the mean and SD for the MDA-435S COX-2 group were 3.91 and 0.43. With 5 samples for MDA-435S and 3 samples for MDA-435S COX-2, we had 74% of power to detect a difference of 1.12 in the mean of log-transformed radiance flux at a significance level of 0.05 using a two sample Satterwaite t-test. Furthermore, power is relevant post-study only for statistically non-significant results. For significant p-values, power is not relevant. At the end of the experiment, we sacrificed mice to confirm pathologically that the metastasis observed by luciferase imaging was indeed localized to bone. The representative luciferase image of the mice injected with control cells (Figure 1D, left panel) and COX-2 transfected cells (Figure 1D, right panel) at day 35 are shown in the ventral position. All the data in Figure 1B-1D is from imaging mice in ventral position. We also collected the data from imaging the same mice in dorsal position and obtained similar results with the exception that 6 B Singh et al signal intensities were lower in dorsal position than in ventral position, presumably due to differences in the depth of metastatic loci from the skin. These data showed clearly that COX-2 overexpression significantly increased metastasis in this mouse model of breast cancer metastasis to bone. Increased PGE2 production in bone-seeking clones of breast cancer cells We hypothesized that if COX-2 protein expressed in breast cancer cells confers a functional advantage on cells in causing bone metastasis, one might expect that starting from a heterogeneous population of cells in culture, cells with a higher level of COX-2 activity would be selected at the site of bone metastases. To test this, we injected breast cancer cells into left cardiac ventricle of female nude mice, and upon the development of bone metastasis, isolated and cultured cells from the metastasis sites at tibia/femur. In this manner, we isolated such boneseeking clones from breast cancer cell populations representing a wide range of PGE2 production, including both untransfected and COX-2 transfected MDA-231 and MDA-435S cells (Figure 2). Figure 2A shows a luciferase image (left panel) and an X-ray (right panel) of mouse number 12 (showing osteolytic bone metastasis) injected with luciferase-transfected MDA-231 cells. We used the bone-seeking clone BSC-12 isolated from mouse number 12 for evaluating the effect of a COX-2 inhibitor in the mouse model (see below). Luciferase images of several other mice from whom the bone-seeking clones were isolated, are shown in Figure 2B. We analyzed the bone-seeking clones generated from several mice (shown in Figure 2A and 2B) in this manner for the COX-2 activity (by measuring PGE2 production) and compared these values to those from the parental cells. This data shows that the bone-seeking clones produce an increased level of PGE2 as compared to the heterogeneous mixed population of cells that were 7 B Singh et al injected into mice (Figure 2C and 2D). This was true in all cases irrespective of whether we used untransfected or COX-2 transfected MDA-231 or MDA-435S cells. These results support our hypothesis and suggest that PGE2 production in breast cancer cells must confer some functional advantage on them so that they complete a cascade of events leading to successful bone metastasis. Inhibition of bone metastasis with a COX-2 inhibitor Next, we evaluated the role of COX-2 in causing bone metastasis by an alternative experimental approach involving the use of a COX-2 specific inhibitor, MF-tricyclic. These experiments had two objectives: 1) to confirm the observations described above that COX-2 in breast cancer cells is important for bone metastasis by showing that a COX-2 inhibitor would inhibit bone metastasis, and 2) to demonstrate in the pre-clinical mouse model that a COX-2 inhibitor would be effective in preventing and treating breast cancer metastasis to bone. We chose to use MFtricyclic, a highly selective COX-2 inhibitor, in diet at a concentration (67 mg per kg diet; approximately 13 mg/kg of mouse weight/day) that has been shown to be effective in inhibiting colon cancer in a mouse model (Lal et al., 2001). First to evaluate the effect of MF-tricyclic in a prevention mode, the treatment group of 5 mice was fed on the MF-tricyclic diet for 7 days before injecting them with the bone-seeking cells from the BSC-12 clone. Then the mice were maintained on the MF-tricyclic diet for the duration of the experiment. The control group of 5 mice was fed on the diet without MF-tricyclic at all times and injected with the BSC-12 cells similar to the treatment group. Relative metastasis burden in individual mice, as measured by whole body luciferase imaging of mice at weekly intervals from day 20 to 34, is shown in Figure 3A; average metastasis burden per mouse in 8 B Singh et al control and treatment groups is shown in Figure 3B. A luciferase image of mice in both control and MF-tricyclic groups on day 27 is shown in Figure 3C. The P-values of F-test of the fixed effects were: treatment 0.062, day <0.0001, and day*treatment 0.013. The P-values for comparing treated vs. control on day 20, 27, 34 were 0.629, 0.151, and 0.0205. These results showed that, as compared to the control group, MF-tricyclic caused a significant inhibition of bone metastasis at day 34 (Figure 3). In a clinical setting, it may not be possible to always use COX-2 inhibitors in prevention regimen. Therefore, from the translational standpoint, it is important to learn whether a COX-2 inhibitor would be effective in a treatment regimen in our pre-clinical model. To address this question, we started by feeding a group of 10 mice on the normal control diet. Fourteen days after injecting BSC-12 cells, mice were luciferase-imaged and then divided into two groups, each group having approximately equal metastases in terms of volume and distribution, as measured in radiance flux. One of these groups continued on the control diet while the other group was switched to the MF-tricyclic diet for the remainder of the period. The metastasis burden, measured by luciferase-imaging on a weekly basis, is shown in Figure 4A; average metastasis burden per mouse in control and MF-tricyclic groups is shown in Figure 4B. A luciferase image of mice in both control and MF-tricyclic groups on day 28 is shown in Figure 4C. The P-values of F-test of the fixed effects were: treatment 0.263, day 0.178, and day*treatment 0.045. The Pvalues for comparing treated vs. control on day 14, 21, 28 were 0.818, 0.091, and 0.034. These results showed that MF-tricyclic caused a significant inhibition of bone metastasis as compared to the control group at day 28 (Figure 4). The inhibitory effect of MF-tricyclic was evident in two ways, 1) reduced rate of growth of metastasis in the initial period of 35 days (21 days of MF- 9 B Singh et al tricyclic treatment), and 2) longer lag period before the exponential growth of metastasis (compare Figure 4A and 4B). Discussion Our results indicate that COX-2 mediated production of PGE2 in breast cancer cells enhances metastasis to bone, a major site of breast cancer metastasis in humans. Main features of the preclinical mouse model used in this study included: 1) route of introduction of cancer cells through left ventricle thus allowing efficient metastasis to bone, 2) stable long-term expression of luciferase in breast cancer cells without any selection pressure in the mouse background, and 3) ability to image and quantitate metastasis in living mice over time with a high degree of sensitivity. One can introduce the desired genetic changes in cancer cells and evaluate their effect on bone metastasis. We chose to investigate the effect of COX-2 in breast cancer metastasis to bone because 1) COX-2 is expressed in poor-prognosis human breast cancers including those overexpressing HER2, and 2) studies with cell lines and mouse models indicate that COX-2 overexpression in breast cancer cells inhibits apoptosis of breast cancer cells and promotes metastasis (Liu et al., 2001, Basu et al., 2004, Singh et al., 2005). Other in vitro studies have shown that breast cancer cells can stimulate osteoblasts and stromal cells to produce PGE2, which in turn induces osteoclastogenesis thus causing osteolytic bone metastasis (Ono et al., 2002; Morgan et al., 2004). Our cumulative evidence for the role of COX-2 in bone metastasis is based upon: 1) increased bone metastasis upon COX-2 transfection of breast cancer cells (Figure 1), 2) inhibition of bone metastasis (in both prevention and treatment regimens) by MF-tricyclic (Figures 3 and 4), and 3) demonstration that bone-seeking clones of breast cancer cells produce 10 B Singh et al higher levels of PGE2 than the parental cell population (Figure 2), perhaps reflecting that PGE2 confers an advantage on breast cancer cells for bone metastasis. Although we have not investigated the mechanism of COX-2 mediated bone metastasis here, there is evidence that PGE2 induces bone resorption both by stimulating RANKL (receptor activator of NF- B ligand) production in osteoblasts (Li et al., 2002) and by directly acting on osteoclasts (Fujita et al., 2003). We also have evidence that COX-2 mediated bone metastasis involves induction of interleukin-8 and interleukin-11 cytokines in breast cancer cells (Singh et al., 2006a, 2006b), both of which have well established roles in bone metastasis (Girasole et al., 1994, Kang et al., 2003, Bendre et al., 2003). Our criteria to demonstrate that COX-2 produced in breast cancer cells is important for bone metastasis are similar to those employed by others in case of other genes involved in bone metastasis (e.g., see Kang et al., 2003). The results showing a clonal selection of PGE2 overproducing breast cancer cells at the site of bone metastasis provide a strong evidence for the role of PGE2 in this process. In another study, BALB/c-MC cells producing a low level of COX2 formed osteolytic bone metastasis upon injection into the heart of Balb/c mice (Ono et al., 2002). Although these authors did not analyze metastatic clones, they found that bone marrow stromal cells adjacent to tumor cells expressed COX-2 protein. Results of our study and those of Ono et al. (2002) indicate that production of PGE2 in the tumor and/or its microenvironment is important for bone metastasis. While PGE2 produced by breast cancer cells has a specific role in bone metastasis, our study does not rule out additional roles of COX-2 in cancer progression, which are not specific to bone metastasis. Role of COX-2 in breast cancer has been demonstrated using several different rodent models, including 7,12-dimethyl-benz(a)anthracene induced mammary carcinogenesis in rat (Harris et al., 2000), MMTV-COX-2 and MMTV-HER2 11 B Singh et al transgenic mouse models (Liu et al., 2001; Howe et al., 2002; Chang et al., 2004), MMTVHER2 transgenic/COX-2 knock-out mouse model (Howe et al., 2005), and xenograft mouse models involving inoculation of cancer cells into fat pad (Connolly et al., 2002; Roche-Nagle et al., 2004) . These studies indicate an important role of COX-2 in primary tumor growth, angiogenesis and metastasis. With the use of other pre-clinical models, others have shown that a COX-2 inhibitor celecoxib inhibits angiogenesis and induces apoptosis in breast cancer cells in a mouse model of spontaneous metastasis (Basu et al., 2004). In another model involving MDA435-FL-GFP human breast cancer xenografts in nude mice, Zhang et al. (2004) showed that celecoxib inhibits overall tumor burden and lung metastasis. COX-2 could have yet additional roles including a role in the pre-malignant phenotype (Crawford et al., 2004) and a role in suppressing tumor-specific host immune responses (Kundu et al., 2005; Basu et al., 2006). Our ongoing unpublished studies on the analysis of RNA and protein profiles in COX-2 transfected breast epithelial cells are consistent with the multiple roles of COX-2 in cancer progression and metastasis. The outcome of tumor metastasis may be influenced at multiple steps in the mouse model. We chose an appropriate model, widely used by others investigating the mechanisms of bone metastasis, to investigate whether COX-2 is important for bone metastasis. Although the model has several features making it suitable for our studies, measurement of metastasis by whole-body luciferase imaging does not allow us to distinguish between various steps at which COX-2 could contribute to the final outcome of metastasis. The major contribution of COX-2 could be to support growth at the secondary site but its other roles cannot be ruled out (e.g., inhibition of apoptosis in the bone microenvironment) that could be important in establishing 12 B Singh et al metastasis. Particularly the initial steps of the process (before the appearance of measurable signal) are least understood. There may be several mechanisms to account for the increased COX-2 activity at the metastasis sites. On the one hand alterations in cancer cells themselves, e.g, overexpression of HER-2, may lead to COX-2 activation (Subbaramaiah et al., 2002). On the other hand alterations in the tumor microenvironment, e.g., release of transforming growth factor-beta from the bone, could induce COX-2 in cancer cells (Hiraga et al., 2006). To determine whether COX-2 expression in primary breast cancer would be predictive of bone metastasis, we are currently analyzing bone marrow micrometastasis (BMM) in breast cancer patients (A Lucci, S Krishnamurthy, B Singh, manuscript in preparation). The results of this clinical study support our hypothesis regarding the role of COX-2 in bone metastasis. Our results of this study in the mouse model, particularly those obtained with the COX-2 inhibitor, provide support for the clinical trials with the COX-2 inhibitors to halt breast cancer metastasis to bone. Materials and methods Cells and DNA transfection The human breast cancer cell lines (MDA-231 and MDA-435S) were obtained from American Type Culture Collection (Manassas, VA) and maintained in RPMI 1640 medium, supplemented with 10% fetal bovine serum, 100 units/ml penicillin, and 100 g/ml streptomycin, in a humidified 5% CO2 atmosphere. COX-2 overproducing cells were generated by co-transfecting breast cancer cell lines with a human COX-2 expression vector pSG5-COX2 (Guo et al., 1996) (obtained from Richard J. Kulmacz, University of Texas Health Science Center, Houston, TX) and pEF1a-Luc-IRES-Neo vector (obtained from David Spencer, Department of Immunology, 13 B Singh et al Baylor College of Medicine, Houston, TX) at a 20:1 ratio by the Lipofectamine protocol (Invitrogen, Carlsbad CA), and by selecting stably transfected cells in the presence of geneticin (0.4 mg/mL, Invitrogen). The control cells were transfected with the pEF1a-Luc-IRES-Neo vector alone. After the initial selection, stably transfected cells were maintained in the presence of 0.1 mg/mL geneticin. The experiments with the stably transfected cells were carried out within 10 passages. Detection of COX-2 by western immunoblotting Cell lysates were prepared by scraping the cells in a sample buffer for SDS-PAGE containing 62.5 mM Tris-HCl pH 6.8, 2% SDS, 10% 2-mercaptoethanol, 10% glycerol, and 0.025% bromophenol blue, and boiling immediately for 5 min followed by centrifugation for 15 min at 14,000 rpm in a microcentrifuge at room temperature. Proteins were resolved by SDS-PAGE on an 8% gel and then electroblotted onto nitrocellulose membrane. A lane containing pre-stained molecular weight markers (Diversified Biotech, Boston, MA) was always included during SDSPAGE. The nitrocellulose membrane was pre-incubated with the blocking agent to reduce nonspecific protein binding and then incubated with an anti-COX-2 monoclonal antibody (Cayman Chemical, Ann Arbor, MI). After incubation with the sheep anti-mouse IgG conjugated to horseradish peroxidase, the blots were processed with the ECL Advance Western blot detection reagents (Amersham Biosciences, Piscataway, NJ) according to the manufacturer’s instructions. The filters were stripped and re-probed with a monoclonal β-actin antibody (Sigma Chemical Company, St Louis, MO), to serve as gel loading controls. 14 B Singh et al PGE2 assay The level of PGE2 released by breast cancer cells was measured by a colorimetric enzyme immunoassay using a kit (Cayman Chemical Company, Ann Arbor, MI) as described previously by others (Subbaramaiah et al., 2002). The growth medium of the cells was replaced with fresh medium containing 10 uM sodium arachidonate 30 minutes prior to collecting the sample for the assay. The samples were stored frozen at –80C prior to the assay. The assay utilizes 96-well plates coated with goat anti-mouse IgG that is incubated with limited amounts of anti-PGE2 monoclonal antibody and a mixture of fixed amount of acetylcholine esterase-coupled PGE2 and the growth medium containing PGE2. After washing the wells, the bound acetylcholine esterase is assayed by a colorimetric reaction. The assay is based on competition between free PGE2 and acetylcholine esterase-coupled PGE2 for binding to a fixed number of antibody molecules. Thus the amount of color produced is inversely proportional to the amount of PGE2 present in the medium. These assays were carried out on two parallel cell cultures assayed in duplicate. We normalized the PGE2 production on an equal protein basis by measuring protein concentration in cell lysates from two parallel cell cultures assayed in triplicate using a protein assay kit (BioRad, Hercules, CA). Whole-body luciferase imaging of mice Breast cancer cell lines (MDA-435S and MDA-231) were stably transfected with luciferase alone or luciferase plus COX-2. After confirming that these cells produce luciferase and COX-2 (by western blot and PGE2 assay), we injected 100,000 cells (in 0.1 mL RPMI 1640 medium without serum and phenol red) into left cardiac ventricle of 4-week old female athymic nude mice (Harlan, Indianapolis, IN) with a 27-gauge syringe as described by others (Arguello et al., 15 B Singh et al 1988). Injections were carried out under deep anesthesia with pentobarbital. Tumors in mice were visualized by whole-body luciferase imaging with IVIS imaging system 100 (Xenogen Corporation, Alameda, CA). Briefly, 1 mg potassium salt of D-luciferin in 0.1 mL phosphate buffered saline was injected intraperitonially with a 25-gauge syringe. Then mice were kept under anesthesia with isofluorane. Starting at ten minutes after the luciferin injection, images were collected for 5 minutes each in ventral and dorsal position, respectively. Relative tumor metastasis burden in mice was calculated with the Living Image software version 2.50 (Xenogen). X-rays To confirm bone metastasis by X-rays, mice were deeply anesthetized with an intraperitonial injection with pentobarbital and X-rayed in prone and lateral positions at 35 kV for 6 sec with a Faxirton imaging system (Faxitron X-ray Corporation, Wheeling, IL). Isolation of bone-seeking clones For the isolation of bone-seeking cells, after detecting metastases in hind limbs by luciferase imaging, mice were sacrificed. Tumor cells were flushed out of bone with a syringe, washed with phosphate buffered saline, and cultured in normal complete RPMI 1640 medium as described by others (Kang et al., 2003). Statistical analysis Statistical analysis was performed using two-sample t-test (Figure 2), and a linear mixed model with repeated measures (Figures 1, 3, and 4). The linear mixed model was fitted to analyze 16 B Singh et al radiance flux using SAS (version 9.0). Radiance flux was first transformed using the natural log scale to better fit the assumptions of linear mixed model. The linear models include fixed effects of treatment, day effect as continuous variable, and the interaction between day and treatment (treated vs. control), with repeated measures over time. We used the first-order autoregressive covariance structure for the MDA-435S COX-2 and the treatment with MF-tricyclic data sets, and used the Compound Symmetry covariance structure for the prevention with MF-tricyclic data set. Due to missing observations at the later time points, the time points included in the linear mixed model for MDA-435S COX-2, prevention with MF-tricyclic, and treatment with MF-tricyclic were 5, 3, and 3, respectively. Comparisons between the treated and control group at each time were made within the linear mixed models. To analyze our data, we tried difference covariance structures including 1) Autoregressive (AR(1) ), 2) Heterogeneous Autoregressive (ARH(1) ), 3) Compound Symmetry(CS), and 4) Heterogeneous Compound Symmetry (CSH). ARH(1) and CSH take into account the heterogeneity between animals. In the final models, we chose the covariance structure based on the Akaike’s information criterion, Bayesian information criterion, and simplicity. P values of 0.05 or less were considered significant. Acknowledgments These studies were initiated when the authors were at Baylor College of Medicine, Houston, TX. We thank Richard J. Kulmacz (University of Texas Health Science Center, Houston, TX) for kindly providing pSG5-COX2 vector, Timothy Hla (University of Connecticut health Center, Pharmington, CT) for pOSML-COX2GFP vector, and David Spencer (Baylor College of Medicine, Houston, TX) for providing the pEF1a-Luc-IRES-Neo vector. We also thank David 17 B Singh et al Spencer for and help with the whole-body luciferase imaging of mice. These studies were supported in part by the grants from the Wendy Will Case Cancer Fund, Inc., Methodist Hospital Foundation and by the grants R21 DK067682 from the National Institutes of Health and DAMD17-03-1-0669 from the United States Army Medical Research and Material Command. 18 B Singh et al References Arguello F, Baggs RB and Frantz CN. (1988). Cancer Res. 48, 6876-6881. Basu GD, Pathangey LB, Tinder TL, Lagioia M, Gendler SJ and Mukherjee P (2004). Mol. Cancer Res. 2, 632-642. Basu GD, Tinder TL, Bradley JM, Tu T, Hattrup CL, Pockaj BA and Mukherjee P. (2006). J. Immunol. 177, 2391-2402. Bendre MS, Montague DC, Peery T, Akel NS, Gaddy D, and Suva LJ (2003) Bone 33, 28-37. Chang SH, Liu CH, Conway R, Han DK, Nithipatikom K, Trifan OC, Lane TF and Hla T. Proc Natl Acad Sci U S A. (2004) 101, 591-596. Connolly EM, Harmey JH, O'Grady T, Foley D, Roche-Nagle G, Kay E and Bouchier-Hayes DJ. (2002). Br. J. Cancer. 87, 231-237. Crawford YG, Gauthier ML, Joubel A, Mantei K, Kozakiewicz K, Afshari CA and Tlsty TD. (2004). Cancer Cell 5, 263–573. Fujita D, Yamashita N, Iita S, Amano H, Yamada S and Sakamoto K. (2003). Prostaglandins Leukot. Essent. Fatty Acids 68, 351-358. Gilhooly EM and Rose DP. (1999). Int. J. Oncol. 15, 267-270. Girasole G, Passeri G, Jilka RL and Manolagas SC. (1994). J. Clin. Invest. 93, 1516-1524. Guo Q, Wang LH, Ruan KH and Kulmacz RJ. (1996). J. Biol. Chem. 271, 19134-19139. Han C, Demetris AJ, Liu Y, Shelhamer JH and Wu T. (2004). J. Biol. Chem. 279, 44344-44354. Harris RE, Alshafie GA, Abou-Issa H and Seibert K. (2000). Cancer Res. 60, 2101-2103. Hiraga T, Myoui A, Choi ME, Yoshikawa H and Yoneda T. (2006). Cancer Res. 66, 2067-2073. 19 B Singh et al Howe LR, Chang SH, Tolle KC, Dillon R, Young LJ, Cardiff RD, Newman RA, Yang P, Thaler HT, Muller WJ, Hudis C, Brown AM, Hla T, Subbaramaiah K and Dannenberg AJ. (2005). Cancer Res. 65, 10113-10119. Howe LR and Dannenberg AJ. (2003). J Mammary Gland Biol Neoplasia 8, 31-43. Howe LR, Subbaramaiah K, Patel J, Masferrer JL, Deora A, Hudis C, Thaler HT, Muller WJ, Du B, Brown AM and Dannenberg AJ. (2002). Cancer Res. 62, 5405–5407. Kang Y, Siegel PM, Shu W, Drobnjak M, Kakonen SM, Cordon-Cardo C, Guise TA and Massague J. (2003). Cancer Cell 3, 537-549. Khuder SA and Mutgi AB. (2001). Br. J. Cancer. 84, 1188-1192. Kundu N, Walser TC, Ma X and Fulton AM. (2005). Cancer Immunol. Immunother. 54, 981987. Kundu N, Yang Q, Dorsey R and Fulton AM. (2001). Int. J. Cancer 93, 681-686. Lal G, Ash C, Hay K, Redston M, Kwong E, Hancock B, Mak T, Kargman S, Evans JF, Gallinger S. (2001). Cancer Res. 61, 6131-6136. Li X, Pilbeam CC, Pan L, Breyer RM and Raisz LG. (2002). Bone 30, 567-573. Liu CH, Chang S-H, Narko K, Trifan OC, Wu M-T, Smith E, Haudenschild C, Lane TF and Hla T. (2001). J. Biol. Chem. 276, 18563-18569. Morgan H, Tumber A and Hill PA. (2004). Int. J. Cancer 109, 653-560. Ono K, Akatsu T, Murakami T, Kitamura R, Yamamoto M, Shinomiya N, Rokutanda M, Sasaki T, Amizuka N, Ozawa H, Nagata N and Kugai N. (2002) J Bone Miner Res. 17, 774-781. Ristimaki A, Sivula A, Lundin J, Lundin M, Salminen T, Haglund C, Joensuu H and Isola J. (2002). Cancer Res. 62, 632-635. 20 B Singh et al Roche-Nagle G, Connolly EM, Eng M, Bouchier-Hayes DJ, and Harmey JH. (2004). Br. J. Cancer. 91, 359-365. Shaheen NJ, Straus WL and Sandler RS. (2002). Cancer 94, 950-963. Singh B and Lucci A. (2002). J. Surg. Res. 108, 173-179. Singh B, Berry JA, Shoher A, Ramakrishnan V and Lucci A. (2005). Int J Oncol. 26, 1393-1399. Singh B, Berry JA, Shoher A and Lucci A. J Surg Res. (2006a). 131, 267-275. Singh B, Berry JA, Vincent LE and Lucci A. J Surg Res. (2006b). 134, 44-51. (Ref completed) Soslow RA, Dannenberg AJ, Rush D, Woerner BM, Khan KN, Masferrer J and Koki AT. (2000). Cancer 89, 2637-2645. Subbaramaiah K, Norton L, Gerald W and Dannenberg AJ. (2002). J. Biol. Chem. 277, 1864918657. Wang D and DuBois RN. (2004). Semin. Oncol. 31, 64-73. Zhao Y, Agarwal VR, Mendelson CR and Simpson ER. (1997). J. Steroid Biochem. Mol. Biol. 61, 203-210. Zhang S, Lawson KA, Simmons-Menchaca M, Sun L, Sanders BG, Kline K. (2004). Breast Cancer Res Treat. 87, 111-121. 21 B Singh et al Titles and legends to figures Figure 1 COX-2 transfection increases bone metastasis in MDA-435S breast cancer cells A: Upper left panel: Cell lysates from the luciferase transfected MDA-435S (lane 1) and COX-2 transfected MDA-435S cells (lane 2) were subjected to western blotting with a COX-2 monoclonal antibody as described in Materials and methods. The abundance of COX-2 protein band, detected in 72-kDa size range, increased dramatically in COX-2 transfected cells. Lower left panel: Nitrocellulose membrane shown in upper panel was stripped and re-probed with a βactin antibody. Right panel: PGE2 produced in the medium from luciferase-transfected control and COX-2 transfected MDA-435S cells was measured as described in Materials and methods. B: Measurement of bone metastasis by whole-body luciferase imaging. Mice in the groups of five were injected with either luciferase-transfected control (left panel; mice M26-M30) or COX2 transfected MDA-435S cells (right panel; mice M31-M35). Mice were imaged weekly starting at day 28, and relative luciferase signal (shown as radiance flux in photons per second) over the body of the mice quantitated as described in Materials and methods. Mouse M32 died as a result of cardiac injection. Mouse M31 was sacrificed on day 42 for the isolation of a bone-seeking clone from the tibia/femur. P = 0.05 at day 56 using a two sample Satterwaite t-test. C: Average metastasis burden per mouse for each group (mice injected with control or COX-2 transfected MDA-435S cells) was determined by averaging the metastases for all the mice at the given time points, for an easy comparison of the groups. 22 B Singh et al D: The images shown in this photograph were taken in ventral position 35 days after injecting cells. A bar on the right shows the pseudocolor representation of the relative intensities of luciferase signal. Average flux per mouse: MDA-435S injected mice, 0.52 million photons/sec; MDA-435S-COX-2 injected mice, 0.82 million photons/sec. Figure 2 Increased production of PGE2 in the bone-seeking clones of breast cancer cells A: The left panel shows a luciferase image of mouse number 12 in ventral position, from whom bone-seeking clone BSC-12 was isolated. The right panel shows an X-ray of the right leg of the mouse number 12, showing an extensive bone osteolysis that corresponds to the location of the luciferase signal on the mouse. B: Luciferase images of mice that were used for isolating the bone-seeking clones. The arrows indicate the metastases from where BSCs were isolated. The numbers below the images refer to the mouse number and the clone number (e.g., BSC-95 was isolated from the mouse 95). C and D: Parental cells and bone seeking clones isolated from mice injected with MDA-231 (C left panel), MDA-435S (C right panel), COX-2 transfected MDA-231 (D left panel), and COX-2 transfected MDA-435S cells (D right panel) were cultured as described in Materials and methods. The cells were exposed to fresh medium containing 10 uM sodium arachidonate for 30 minutes prior to collecting the medium for the PGE2 immunoassay with a kit (Cayman Chemical Company, Ann Arbor, MI). P < 0.02 in all comparisons involving parental cells and boneseeking clones. 23 B Singh et al Figure 3 Prevention of the bone metastasis with MF-tricyclic A: Mice were fed on a normal diet (left panel) or the diet pre-mixed with 0.0067% MF tricyclic (right panel) beginning at 7 days prior to injecting the breast cancer cells. 0.1 million cells of a bone-seeking clone BSC-12, that is derived from MDA-231 cells, were injected into the left ventricle of female athymic mice (n = 5). Mice were imaged weekly starting at day 20, and relative luciferase signal (shown as radiance flux in photons per second) over the body of the mice quantitated as described in Materials and methods. One mouse in the control diet group, M49, died as a result of heart injection. One mouse in the MF-tricyclic diet group, M43, died on day 34 presumably from metastasis. (P = 0.02 at day 34). B: Average metastasis burden per mouse for each group (mice fed on control or MF-tricyclic diet) was determined by averaging the metastases for all the mice at the given time points, for an easy comparison of the groups. C: The images shown in this photograph were taken in ventral position 27 days after injecting the BSC-12 cells. A bar on the right shows the pseudocolor representation of the relative intensities of luciferase signal. Average flux per mouse: control diet group, 5.47 million photons/sec; MF tricyclic diet group, 2.64 million photons/sec. 24 B Singh et al Figure 4 Treatment of the bone metastasis with MF-tricyclic A: We started by feeding a group of 10 mice on the normal control diet. BSC-12 cells were successfully injected into the left ventricle of 6 out of 10 mice. Fourteen days after injecting BSC-12 cells, mice were luciferase-imaged and then divided into two groups, each group having approximately equal metastases in terms of volume and distribution. One of these groups continued on the control diet while the other group was switched to the MF-tricyclic diet for the remainder of the period. Mice were imaged weekly starting at day 20, and relative luciferase signal (shown as radiance flux in photons per second) over the body of the mice quantitated as described in Experimental procedures. Mouse M9801 was sacrificed on day 42 for the isolation of a bone-seeking clone from a large bone metastasis in a leg. All other mice died of metastasis: M9803 on day 35, M9804 on day 49, M9802 on day 42, and both M9805 and M9606 on day 62. (P = 0.03 at day 28). B: Average metastasis burden per mouse for each group (control or MF-tricyclic treated mice) was determined by averaging the metastases for all the mice at the given time points, for an easy comparison of the groups. C: The images shown in this photograph were taken in ventral position 28 days after injecting the BSC-12 cells, 14 days after switching to MF-tricyclic diet in the case of treatment group (right panel). A bar on the right shows the pseudocolor representation of the relative intensities of luciferase signal. 25