Polar bears (Ursus maritimus) are charismatic megafauna and the
advertisement
 are charismatic megafauna and the")
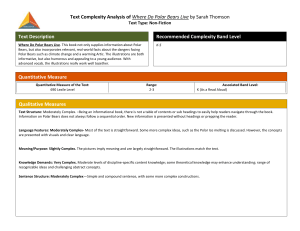
Polar bears and climate change? M. G. Dyck 1, 2 and W. Soon3 1 Natural Resources Institute, University of Manitoba, Winnipeg, Manitoba R3T 2N2 Canada 2 Correspondence address: Department of Sustainable Development Wildlife Division Box 1000, Station 1170 Iqaluit, Nunavut X0A 0A0 Canada Ph: (867) 975-5943 Fx: (867) 975-5981 Email: MDYCK@GOV.NU.CA 3 Harvard-Smithsonian Center for Astrophysics, Cambridge, Massachusetts, 02138, USA 1 Polar bears (Ursus maritimus) are charismatic megafauna and the symbol of the Arctic. For Canadian Inuit people, polar bears play an important cultural and traditional role in their lives through hunting activities and the sharing of meat. Additionally, Inuit-guided sport hunts provide crucial revenue for the otherwise economically-challenged communities (Lee and Taylor 1994). However, if one believes latest research findings, this multi-purpose natural resource faces threats by climatic change (Stirling and Derocher 1993; Stirling et al. 1999; World Wide Fund for Nature 2002). Long-term ecological studies on polar bears of western Hudson Bay (WH) indicated that this population has been exhibiting a decrease in female body mass, litter sizes, and cub survival (Derocher and Stirling 1992; 1995). It was suggested that the proximate factor for the decline in these physical and reproductive parameters between 1981 through 1998 was a change in freeze and thaw cycles of Hudson Bay causing bears to reach the shore in progressively poorer condition. A long-term warming trend of spring atmospheric temperatures was proposed, though not shown directly1, to be the ultimate factor (Stirling et al. 1999). The WH results were used and applied to generate general predictions for polar bears and their habitat (Stirling and Derocher 1993; World Wide Fund for Nature 2002). Figure 1a shows the surface air temperature records of Churchill, Manitoba2 (assumed here to be representative of WH) for four climatological seasons from 1932-2002. The large interannual variability of the seasonal temperatures suggests that confirming a long-term trend in any of these relatively short records will be difficult and the trend determination will be sensitively dependent on the time interval considered. Figure 1b attests that no statistically significant temperature trend can be confirmed for either the spring (defined specifically here as 1 Stirling et al. (1999) inferred from the mean air temperature results of Skinner et al. (1998). 2 averages of April, May and June to follow discussion in Stirling et al. 1999) or fall seasons when the full records from 1932 through 2002 or 1932 through 1992 were considered. A clear tendency towards spring warming did occurred from 1981 to about 1999 (marked by the dashed trend curve in Figure 1b) but that tendency did not persist nor any systematics observed with data updated through 2002 (yet we make no further confident inference about any concurrent ecological responses). Thus, although our independent results for temperature change and variability over the WH do not contradict Stirling et al. (1999), but the full range of temperature variability we observed argues for a less dominating role of spring temperature as the explanatory variable of the population ecology of polar bears. Figure 2 shows the records of winter and spring temperatures at Frobisher Bay (Iqaluit, Nunavut) and their respective winter and spring Arctic Oscillation (AO) circulation indices3 with two important points. First note the rather strong cooling trend (at a rate of about 0.4ºC per decade since the 1950s) for the winter and spring temperatures of Frobisher Bay by the Hudson Strait. So regional differences in the pattern of the temperature variability, especially on multidecadal time-scale, are large. This pattern of large temperature gradient between the southwestern and northeastern corners of the Hudson Bay oceanic basin has been previously noted by Skinner et al. (1998)―whom had also provided a brief but comprehensive discussion on 2 Our data source is based on the quality-controlled version of records from the NASA Goddard Institute for Space Studies web site: http://www.giss.nasa.gov/data/update/gistemp/station_data/. 3 Arctic Oscillation (AO) is a natural, planetary-scale pattern or mode of atmospheric circulation variability that is characterized by a seasaw of air mass anomaly between the Arctic basin and the midlatitude zonal ring centered at about 45º N. High (positive) AO value is defined as lower-than-normal atmospheric pressure over the Arctic and colder stratosphere which are associated with strong subpolar westerlies while the low (negative) AO value represents higher-than-normal Arctic atmospheric pressure, less cold polar stratosphere and weak subpolar westerlies. The AO index is available from http://horizon.atmos.colostate.edu/ao/Data/index.html. Although the AO is mainly a winter phenomenon because of the relatively larger variability and stronger coupling of stratospheric and tropospheric air circulation during the cold season, the relevance of AO has been demonstrated in other seasons as well (Gong and Ho 2003; Kryjov 2002; Overland et al. 2002). Please see Wallace (2000), Baldwin (2001) and Thompson and Wallace (2001) for the complete tutorials. Although there have been several attempts to suggest that the post-1969 or post-1989 tendency of the AO index to remain in the high positive AO phase is a result of the 3 climate regimes around the western Hudson Bay. Therefore the proposal of a warming spring temperatures, regardless if the fact may holds for any particular time period, for the polar bear population ecology cannot be universally applied elsewhere at other locations. The second point of Figure 2 is that the temperature and climatic conditions around the Hudson Strait and Hudson Bay areas have close association with the AO circulation index. The correlations shown in Figure 2 are highly significant with the AO variability explaining up to 20 to 50% of the temperature variances at Frobisher Bay. Why the labor of introducing and discussing the AO circulation index? Two reasons. First, from the examination of the statistics of sea level pressure and sea ice motion from the 1979-1998 data collected by the International Arctic Buoy Programme, Rigor et al. (2002) confirmed that AO circulation pattern can explain at least part of the thinning sea ice trend observed over the Arctic Ocean. But Rigor et al. (2002) clarified that instead of assuming the warming trend in the surface air temperature to cause the thinning of sea ice, it is the AO circulation pattern that produces the sea ice thinning tendency. In turn, it was the thinning of sea ice that caused the air temperature to warm because of increasing heat flux from the interface of ice-free ocean. Thus this physical picture is indeed different from Stirling et al. (1999)’s proposal in which warm spring air temperature is considered to be the ultimate root cause for the earlier spring sea ice breakup and poorer polar bear conditions. The second reason for our discussion of the AO circulation index is related to a recent finding of the AO index-climatic change effects being propagated through two trophic levels within a high-arctic ecosystem (Aanes et al. 2002). From the statistical analyses of the1987-1998 growth series of Cassiope tetragona (Lapland Cassiope) and the 1978-1998 abundance series of an introduced Svalbard reindeer population forcing by anthropogenic carbon dioxide, but the current generation of climate models and modelling efforts are simply too immature to confirm or refute such a proposal (Soon et al. 2001; Soon and Baliunas 2003). 4 near Broggerhalvoya, NW coast of Svalbard, Aanes et al. (2002) found that high positive values of AO index are associated with decreased plant growth and reindeer population growth rate. Thus, the reindeer population at Svalbard, through the mediation of the climate modulated effects on plant growth, is plausibly connected to climate through a bottom-up sequence. But Aanes et al. (2002) noted that the bottom-up scenario may be density-dependent in that at higher reindeer densities, a top-down sequence of trophic interaction is becoming more important in which grazing has a dominating influence on the forage species and plant communities. The AO index is thus promising as a robust and reliable climatic variable for further examination of the dynamic of trophic interactions under various settings of the arctic ecosystem. Science evolves through new research and critical thinking. In this paper, we examine peer-reviewed literature related to polar bears and the climate change conclusion. We believe that some published results provide room for alternative explanations why bears appear to be in poorer physical condition. Perhaps the effects of climate change, reducing sea-ice cover, and changes in freeze and thaw cycles on polar bears at higher latitudes might not be as detrimental as predicted. These changes might create more polynyas, which are productive oases in the ice (Stirling 1997), or increase marine productivity overall. Bears do not feed year-round, but during late spring when seal pups are abundant. More fat deposits might be accumulated during this time, and a “true hibernation state” like black (U. americanus) and brown bears (U. arctos) could become an evolutionary strategy for the remainder of the year, as polar bears evolved from brown bears (Kurtén 1964). Alternatively, a supplementary feeding strategy could evolve where berries and vegetation are consumed during the ice-free period, as has been observed for bears of Hudson Bay (Russell 1975; Derocher et al. 1993). 5 From 1981 through 1998, the WH population estimates were considered relatively constant at about 900 – 1200 individuals (Lunn et al. 1997a; Stirling et al. 1999). Various studies conducted on population dynamics, growth, and reproduction during this period found declining female reproductive parameters (e.g., Derocher and Stirling 1992, 1995; Stirling and Derocher 1993; Stirling et al. 1999), as well as decreasing female body length (Atkinson et al. 1996; Derocher and Stirling 1998). Whenever female body condition is poor, cub survival and litter size are negatively affected as these parameters are closely related (Derocher and Stirling 1996; Atkinson and Ramsay 1996). These declining trends were consistent with density-dependent responses, where lowered food abundance and increased intraspecific competition were suggested as causes (see Derocher and Stirling 1992 for details). Perhaps what is being observed for WH polar bears is caused by density-dependent factors (but see Derocher and Taylor 1994) rather than by climate change. How population dynamics of ringed (Phoca hispida) and bearded seals (Erignathus barbatus), polar bears’ main prey items (Stirling and Archibald 1977; Smith 1980), have changed during this period is unclear, as data are very limited (Lunn et al. 1997b). We caution the interpretation of results whenever body mass of polar bears was estimated by chest girth using a general equation (i.e., Kolenosky et al. 1989). Polar bears span a range of body mass over two orders in magnitude, depending on season, sex, age, and reproductive class (Stirling et al. 1977; Watts and Hansen 1987; Kolenosky et al. 1989). They also cycle between periods of hyperphagia and fasting, where body fat constitutes as much as 50% or as little as 10% of their total body mass, respectively (Cattet 1990; Pond and Ramsay 1992; Pond et al. 1992; Atkinson et al. 1996). This indicates that polar bears are highly variable in body mass, and that morphometry is neither accurate nor precise to estimate body mass. Not general, but rather 6 population specific equations for adult males, females, and subadults should be employed (Cattet et al. 1997). Given that the uncertain state of our understanding on the complex interaction among sea ice, atmospheric and oceanic circulations, and air and sea temperatures, we suggest that large interannual variability, which is largely stochastic in nature, dominates the climatic changes that will ultimately bring about particular ecosystem responses (perhaps yet unintelligible to us) but spring temperature is not necessarily a decisive predictor. Given the alternative interpretations regarding the decline of reproductive parameters of female polar bears of WH, we believe it is premature to make further-reaching predictions about how climate change may affect polar bears in general. It might be beneficial to re-examine existing data on temporal variation of growth for body length of females of other polar bear populations for comparison. In addition, specific population equations could be employed for body masses of WH bears. If what is being observed in WH is truly a density-dependent response, then long-term ecological studies for the Svalbard polar bear population should continue. The Svalbard population is currently the only studied population with no ongoing harvest, and is thought to have recovered from over-harvesting (Prestrud and Stirling 1994; Wiig 1998). Possible density-dependent effects could be assessed by studying and comparing these two populations. In addition, we encourage research, conducted simultaneously with polar bear studies, that examines lower trophic levels to aid in providing more insight into the proximate effects of climate change on Arctic species. We further suggest that the AO circulation index may be useful in tracking the propagation of any climatic signals through the coupled ecosystems of the Arctic land and sea that promises only the undeniable reality of multi-trophic level interactions (Steinke et al. 2002). 7 References Aanes, R., Sæther, B.-E., Smith, F.M., Cooper, E.J., Wookey, P.A. and Øritsland, N.A. 2002. The Arctic Oscillation predicts effects of climate change in two trophic levels in a high-arctic ecosystem. – Ecology Let. 5: 445 – 453. Atkinson, S.N. and Ramsay, M.A. 1996. The effects of prolonged fasting of the body composition and reproductive success of female polar bears (Ursus maritimus). – Functional Ecology 9: 559 – 567. Atkinson, S.N., Stirling, I., and Ramsay, M.A. 1996. Growth in early life and relative body size among adult polar bears (Ursus maritimus). – J. Zool. (Lond.). 239: 225 – 234.. Baldwin, M.P. 2001. Annular modes in global daily surface pressure. – Geophys. Res. Let. 28: 4115 – 4118. Cattet, M. 1990. Predicting nutritional condition of black bears and polar bears on the basis of morphological and physiological measurements. – Can. J. Zool. 68: 32 – 39. Cattet, M. R. L., Atkinson, S. N., Polischuk, S. C., and Ramsay, M. A. 1997. Predicting body mass in polar bears: is morphometry useful? – J. Wildl. Manage. 61: 1083 –1090. Derocher, A.E. and Stirling, I. 1992. The population dynamics of polar bears in western Hudson Bay. – In: McCullough, D. R. and Barrett, R. H. (eds.), Wildlife 2001: populations. Elsevier Science Publishers, pp. 1150 – 1159. Derocher, A.E. and Stirling, I. 1995. Temporal variation in reproduction and body mass of polar bears in western Hudson Bay. – Can. J. Zool. 73: 1657 – 1665. Derocher, A.E. and Stirling, I. 1996. Aspects of survival in juvenile polar bears. – Can. J. Zool. 74: 1246 – 1252. 8 Derocher, A. E. and Stirling, I. 1998. Geographic variation in growth of polar bears (Ursus maritimus). – J. Zool. (Lond.). 245: 65 – 72. Derocher, A.E. and Taylor M. 1994. density-dependent population regulation of polar bears. In: Taylor, M. (ed.), Density-dependent population regulation of black, brown, and polar bears. Int. Conf. Bear Res. Manage. Monogr. Series No.9, pp. 25 – 30. Derocher, A.E., Andriashek, D., and Stirling, I. 1993. Terrestrial foraging by polar bears during the ice-free period in western Hudson Bay. – Arctic 46: 251 – 254. Gong, D.-Y. and Ho, C.-H. 2003. Arctic Oscillation signals in the East Asian summer monsoon. – J. Geophys. Res. 108 (D2), 4066, doi:10.1029/2002JD002193: 1–7. Kolenosky, G.B., Lunn, N.J., Greenwood, C.J., and Abraham, K.F. 1989. Estimating the weight of polar bears from body measurements. – J. Wildl. Manage. 53: 188 – 190. Kryjov, V.N. 2002. The influence of the winter Arctic Oscillation on the Northern Russian spring temperature. – Int. J. Clim. 22: 779 – 785. Kurtén, B. 1964. The evolution of the polar bear, Ursus maritimus Phipps. – Acta Zool. Fenn. 108: 1 – 30. Lee, J. and Taylor, M. 1994. Aspects of the polar bear harvest in the Northwest Territories, Canada. – Int. Conf. Bear Res. Manage. 9: 237 – 243. Lunn, N.J., Stirling, I., Andriashek, D., and Kolenosky, G.B. 1997a. Re-estimating the size of the polar bear population in western Hudson Bay. – Arctic 50: 234 – 240. Lunn, N.J., Stirling, I. and Nowicki, S.N. 1997b. Distribution and abundance of ringed (Phoca hispida) and bearded seals (Erignathus barbatus) in western Hudson Bay. – Can. J. Fish. Aquat. Sci. 54: 914 – 921. 9 Overland, J.E., Wang, M. and Bond, N.A. 2002. Recent temperature changes in the Western Arctic during spring. – J. Clim. 15: 1702 – 1716. Pond, C.M. and Ramsay, M.A. 1992. Allometry of the distribution of adipose tissue in Carnivora. – Can. J. Zool. 70: 342 – 347. Pond, C.M., Mattacks, C.A., Colby, R.H., and Ramsay, M.A. 1992. The anatomy, chemical composition, and metabolism of adipose tissue in wild polar bears (Ursus maritimus). – Can. J. Zool. 70: 326 – 341. Prestrud, P. and Stirling, I. 1994. The International Polar Bear Agreement and the current status of polar bear conservation. – Aquat. Mamm. 20: 113 – 124. Rigor, I.G., Wallace, J.M. and Colony, R.L. 2002. Response of sea ice to the Arctic Oscillation. – J. Clim. 15: 2648 – 2663. Russell, R.H. 1975. The food habits of polar bears of James Bay and southwest Hudson Bay in summer and autumn. – Arctic 28: 117 – 129. Skinner, W.R., Jefferies, R.L., Carleton, T.J., Rockwell, R.F. and Abraham, K.F. 1998. Prediction of reproductive success and failure in lesser snow geese based on early season climatic variables. – Glob. Change Bio. 4: 3 – 16. Smith, T.G. 1980. Polar bear predation of ringed and bearded seals in the land-fast ice habitat. – Can. J. Zool. 58: 2201 – 2209. Soon, W. and Baliunas, S. 2003. Annual progress report on “Global Warming.”– Prog. Phys. Georgr. 27 (3): in press. Soon, W., Baliunas, S., Idso, S.B., Kondratyev, K.Ya. and Posmentier, E.S. 2001. Modeling climatic effects of anthropogenic carbon dioxide emissions: Unknowns and uncertainties. – Clim. Res. 18: 259 – 275. 10 Steinke, M., Malin, G. and Liss, P.S. 2002. Trophic interactions in the sea: An ecological role for climate relevant volatiles? – J. Phycol. 38: 630 – 638. Stirling, I. 1997. The importance of polynyas, ice edges, and leads to marine mammals and birds. – J. Mar. Syst. 10: 9 – 21. Stirling, I. and Archibald, W.R. 1977. Aspects of predation of seals by polar bears. – J. Fish. Res. Bd Can. 34: 1126 – 1129. Stirling, I. and Derocher, A.E. 1993. Possible impacts of climatic warming on polar bears. – Arctic 46: 240 – 245. Stirling, I., Lunn, N.J., and Iacozza, J. 1999. Long-term trends in the population ecology of polar bears in western Hudson Bay in relation to climatic change. – Arctic 52: 294 – 306. Stirling, I., Jonkel, C., Smith, P., Robertson, R., and Cross, D. 1977. The ecology of the polar bear (Ursus maritimus) along the western shore of Hudson Bay. – Can. Wildl. Serv. Occas. Pap. No. 33. Thompson, D.W.J. and Wallace, J.M. 2001. Regional impacts of the Northern Hemisphere annular mode. – Science 293: 85 – 89. Wallace, J.M. 2000. North Atlantic mode/annular mode: Two paradigms– One phenomenon. – Quart. J. Roy. Meteorol. Soc. 126: 791 – 805. Watts, P.D. and Hansen, S.E. 1987. Cyclic starvation as a reproductive strategy in the polar bear. – Symp. Zool. Soc. Lond. No. 57: 305 – 318. Wiig, Ø. 1998. Survival and reproductive rates for polar bears at Svalbard. – Ursus 10: 25 – 32. World Wide Fund for Nature. 2002. Polar bears at risk. – World Wide Fund for Nature, Gland, Switzerland. 11 Figure Captions Figure 1: (a) Climatological Winter (as averages of December, January and February), Spring (as averages of March, April and May), Summer (as averages of June, July and August) and Fall (as averages of September, October and November) surface air temperatures of Churchill, Manitoba (assumed to be representative of temperatures around the western Hudson Bay) from 1932 through 2002. Four-year gaps between 1993 and 1996 are missing or incomplete data. (b) Spring (defined specifically as averages of April, May and June to follow the discussion in Stirling et al. 1999; top panel) and Fall (bottom panel) temperatures with statistically insignificant (i.e., with p > 0.05) trend lines (dotted) fitted through the 1932-2002 and 1932-1992 intervals. Dashed trend line fitted through 1981-1999 verifies the spring warming episode noted by Stirling et al. (1999). Figure 2: Statistically significant (i.e., p << 0.001) correlation between temperature (solid) at Frobisher Bay, Nunavut and the Arctic Oscillation (AO) index (dotted) for Winter (as averages of January, February and March; top panel) and Spring (as averages of March, April and May; bottom panel). The axis for AO indices has reverse scales such that high positive AO values mean colder temperatures at Frobisher Bay. 52% and 20% of the variance of the winter and spring temperatures for the 1943-2002 interval are explained the respective AO indices. Both the standard Pearson’s r and non-parametric Kendall’s τ were computed and statistical significance of the results are established based on both statistics. 12