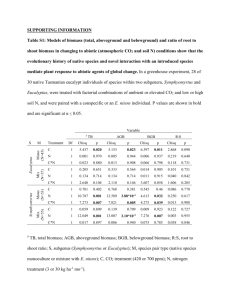
Arcano, Rick, Allometric Model Development, Biomass Allocation
advertisement

To the Graduate School: The members of the Committee approve the thesis of Rick Arcano on April 26, 2005. Daniel B. Tinker, Co-Chairman Gregory K. Brown, Co-Chairman Elise G. Pendall Peter D. Stahl APPROVED: Gregory K. Brown, Head, Department of Botany Don Roth, Dean, The Graduate School ALLOMETRIC MODEL DEVELOPMENT, BIOMASS ALLOCATION PATTERNS, AND NITROGEN USE EFFICIENCY OF LODGEPOLE PINE IN THE GREATER YELLOWSTONE ECOSYSTEM By Rick Arcano A thesis submitted to the Department of Botany and the Graduate School of the University of Wyoming in partial fulfillment of the requirements for a degree of MASTER OF SCIENCE In BOTANY Laramie, Wyoming May, 2005 Table of Contents Abstract ………………………………………………………………………………….iii Acknowledgments ………………………………………………………………………….…iv Preface ……………………………………………………………………..…………...v Chapter 1 Allometric Model Development for Mature Lodgepole Pine Forests in the Greater Yellowstone Ecosystem ……………………………………………...……….1 Chapter 2 Biomass Allocation Patterns and Nitrogen Use Efficiency for Mature Lodgepole Pine Forests in the Greater Yellowstone Ecosystem ………………………...40 Summary …………………………………………………………………………….......89 ii Arcano, Rick, Allometric Model Development, Biomass Allocation, and Nitrogen Use Efficiency of Lodgepole Pine in the Greater Yellowstone Ecosystem, M.S., Department of Botany, May, 2005. Estimation of Net Primary Production (NPP) in lodgepole pine (Pinus contorta var. latifolia (Engelm. Ex Wats.) Critchfield) forests is critical for understanding how climateinduced or anthropogenic changes in fire frequency and pattern may affect the carbon balance and biomass allocation across the Greater Yellowstone landscape. In addition, biomass allocation patterns are influenced by nitrogen use efficiency (NUE), yet little is known about their interactions. Allometric equations that quantify biomass production of lodgepole pine in the Greater Yellowstone Ecosystem (GYE) were developed in the summer of 2004. Statistical comparisons were used to determine whether allometric models differed between forests of different ages and densities, and t-tests were used to determine if allometric models and biomass allocation patterns in the GYE differed from southeastern Wyoming and in British Columbia, Canada. One-way ANOVA was used to determine whether biomass allocation patterns and NUE differed between stand densities and ages, and correlation analyses were used to determine how they were related. Allometric models differed by both density and age, but differed more by age. Biomass allocation patterns differed between forest density and age, but coarse root allocation was relatively constant across forest densities and ages. Aboveground biomass allocation patterns and allometric models developed in this study differed significantly from allometric models developed elsewhere, independent of stand density and age. NUE was highest in the oldest stand, and was correlated with site productivity and biomass allocation patterns. NPP and Net Ecosystem Production (NEP), which differ among lodgepole pine ecosystems, are impacted by forest structure, and are at least partly impacted by the interrelationships among allometric model development, biomass allocation patterns, and NUE. iii ACKNOWLEDGMENTS I would like to thank my graduate advisor, Daniel B. Tinker for providing me the opportunity to be part of the Department of Botany at the University of Wyoming and providing me all the tools necessary to succeed. I would like to thank my graduate committee -- Drs. Gregory K. Brown, Elise G. Pendall, and Peter D. Stahl for their guidance and support. I also received invaluable support from the entire botany department, including my fellow graduate students, especially Abbie Gongloff, Jennifer Andersen, Mark Andersen, Peter Ebertowski, and Erin Foley. I had five diligent and dedicated field assistants whom willingly and effectively participated in sometimes grueling and tedious field and lab work for this study. They are Lance East, Lance Farman, Christine Regester, Kellen Nelson, Terez Tepe, and Heather Lyons. I have estimated that I would have needed 6-10 more years to complete this study without them. I am very appreciative of my access to the experience and expertise of Drs. Michael G. Ryan of the Rocky Mountain Research Station of the USDA Forest Service, and Daniel M. Kashian and William H. Romme from the Department of Forest, Rangeland, and Watershed Stewardship at Colorado State University. Drs. David E. Legg from the Entomology Department at the University Wyoming and Rudy M. King from the Rocky Mountain Research Station of the USDA Forest Service provided me with statistical advice. I was permitted to conduct my research on the Ashton-Island Park Ranger District of the Caribou-Targhee National Forest and deeply appreciate it. Without the help of everyone involved, completion of this project would not have been possible. This research was supported by grants from the Plummer Scholarships for Environment and Natural Resources Conservation and Management, University of Wyoming / National Park Service Research Center, and the USDA Joint Fire Sciences Program. iv PREFACE The global climate has been subject to dramatic change over the past half century, and the climate of the intermountain west is undergoing similar alterations. Both naturally and anthropogenically-altered climatic patterns in the western U.S. are likely to cause substantial changes in natural fire regimes that could result in alterations in forest stand structure and productivity (Amiro et al. 2000; Dale et al. 2001). These changes in forest structure and productivity may cause changes in Net Ecosystem Production (NEP), which varies widely across many forested landscapes. The variability in NEP is produced by heterogeneity in stand ages and densities that are impacted by natural and anthropogenically altered disturbance regimes (Chapin et al. 2002; Turner et al. 2004). To determine how NEP is affected by altered disturbance regimes, quantification of forest structure and determination of how lodgepole pine (Pinus contorta var. latifolia (Engelm. ex Wats.) trees and stands allocate their biomass above and belowground is needed. Detailed estimates of forest structure and function requires accurate and easily obtained measurements of aboveground and belowground tree biomass, which are necessary components for determining Net Primary Production (NPP) of forested ecosystems. To accurately estimate NPP, it is important to use allometric equations that consider differences in allocation patterns of lodgepole pine biomass, which may vary with tree density and stand age (Pearson et al. 1984). In this study, we developed new allometric models for predicting above and belowground biomass in mature lodgepole pine forests of the GYE. We also harvested lodgepole pine above and belowground in three stands differing in densities and ages to determine the allocation patterns of mature lodgepole pine forests in the GYE across varying forest structures. This study was part of a larger study funded by the USDA Joint Fire Sciences program to determine the carbon budget of the Greater Yellowstone Ecosystem and how this v budget is changed by alterations in climate and fire regime. Development of allometric equations for lodgepole pine, which comprises over 80% of the overstory vegetation (Despain 1990) in Yellowstone National Park, is critical for estimating carbon pools and Net Ecosystem Production across the Greater Yellowstone landscape. Determining the use efficiency for essential nutrients such as nitrogen (N) is also important for improving our understanding of biomass accumulation of lodgepole pine. Gross Primary Production (GPP), a major component of NEP, is equal to the product of resource supply, the proportion of the resource supply that is taken up, and the efficiency of resource use (Binkley et al. 2004). In this study, we estimated NUE in three sites in the Greater Yellowstone Ecosystem that varied by stand age and tree density to determine how NUE relates to variability in forest structure and site index (see Methods), and to identify relationships between NUE and biomass allocation patterns. The specific objectives of this study were (1) to develop allometric models for predicting above and belowground biomass of mature lodgepole pine trees in the GYE; (2) to determine how allometric model development and application for lodgepole pine differs with stand density and age; (3) to compare and contrast allometric equations developed in this study to allometric equations developed in other locations to determine model variability and applicability across geographic locations independent of forest structure. (4) to determine how patterns of above and belowground biomass allocation for mature lodgepole pine forest of the GYE vary with stand age and density; (5) to quantify patterns of nitrogen use efficiency (NUE) for lodgepole pine forests among these differing forest structures and productivities; and (6) to examine the relationships between NUE and biomass allocation patterns. vi Chapter 1 will address the allometric model development of lodgepole pine in the GYE, and will also focus on potential differences in allometric models with differing stand density, stand age, and geographic location. Chapter 2 will focus on biomass allocation patterns and nitrogen use efficiency. Specifically, it will focus on how biomass allocation and nitrogen use efficiency differ with stand density, stand age, and site productivity. In addition, the potential relationships between biomass allocation patterns and nitrogen use efficiency will be examined. Next, I will summarize the findings and results of both chapters in a final summary section. vii LITERATURE CITED Amiro, B. D., J. M. Chen and J. Liu 2000. Net primary productivity following forest fire for Canadian ecoregions. Can. J. For. Res. 30: 939-947. Binkley, D., J. L. Stape and M. G. Ryan 2004. Thinking about efficiency of resource use in forests. Forest Ecology and Management 193: 5-16. Chapin, S. F. I., P. Matson and H. A. Mooney (2002). Principles of terrestrial ecosystem ecology. New York, NY, Springer-Verlag New York, Inc. Dale, V. H., L. A. Joyce, S. McNulty, R. P. Neilson, M. P. Ayres, M. D. Flannigan, P. J. Hanson, L. C. Irland, A. E. Lugo, C. J. Peterson, D. Simberloff, F. J. Swanson, B. J. Stocks and B. M. Wotton 2001. Climate change and forest disturbances. Bioscience 51: 723-734. Despain, D. G. (1990). Yellowstone vegetation: consequences and environment in a natural setting. Boulder, CO, Roberts Rinehart, Inc. Pearson, J. A., T. J. Fahey and D. H. Knight 1984. Biomass and leaf area in contrasting lodgepole pine forests. Can. J. For. Res. 14: 259-265. Turner, M. G., D. B. Tinker, W. H. Romme, D. M. Kashian and C. M. Litton 2004. Landscape patterns of sapling density, leaf area, and aboveground net primary production in postfire lodgepole pine forests, Yellowstone National Park (USA). Ecosystems 7: 751-775. viii Chapter 1 Allometric Model Development for Mature Lodgepole Pine Forests in the Greater Yellowstone Ecosystem INTRODUCTION Natural and anthropogenic effects on climatic patterns in the western U.S. are likely to cause substantial changes in natural fire regimes that could result in alterations in forest stand structure and productivity (Amiro et al. 2000; Dale et al. 2001). Quantification of forest structure and function requires accurate and easily obtained measurements of aboveground and belowground tree biomass, which are necessary components for determining Net Primary Production (NPP) of forested ecosystems. In Yellowstone National Park (YNP), lodgepole pine (Pinus contorta var. latifolia (Engelm. ex Wats.) Critchfield) comprises over 80% of the overstory vegetation (Despain 1990). Estimating the biomass of these vast forests is a critical component in determining the NPP of lodgepole forests across the entire Greater Yellowstone Ecosystem (GYE). To accurately estimate NPP, it is important to use allometric equations that consider differences in allocation patterns of lodgepole pine biomass, which may vary with tree density and stand age. Useful allometric equations for above and belowground biomass of lodgepole pine have been developed in Alberta, Canada by Johnstone (1971), in southeastern WY by Pearson et al. (1984), in southeastern British Columbia by Comeau and Kimmins (1989), and in Washington and Oregon by Gholz et al. (1979). More recently, allometric equations for predicting biomass in 10 year-old post-fire forests have also been developed in Yellowstone National Park (YNP) for aboveground biomass by Turner et al. (2004), and for belowground biomass by Litton et al. (2003). Although Litton et al. (2004) later suggested that allometric models developed by 1 Comeau and Kimmins (1989) and Pearson et al. (1984) could be appropriate for use in the GYE, the models were tested on only five mature trees in a single stand, and did not account for natural variability in stand density and age. In addition, allometric models for predicting belowground biomass were not critically evaluated. Therefore, new allometric models specific to mature forests of the Greater Yellowstone Ecosystem are needed. In this study, we developed new allometric models for predicting above and belowground biomass in mature lodgepole pine forests of the GYE. Recently, allometric models were developed and applied to predict aboveground NPP of young post-fire forests across the entire Yellowstone landscape by Turner et al. (2004). Their landscape scale extrapolation was made possible by the applicability and accuracy of their allometric models. Allometric equations developed for lodgepole pine and many other species (Ter-Mikaelian and Korzukhin 1997) have proven to be very good predictors of above and belowground biomass for individual trees, where coefficients of determination are often above 0.90 for aboveground tree components and for roots >10mm diameter. Despite the high quality of allometric equations, previous studies have suggested that they may not be universally applicable in stands of varying densities, ages, and site qualities. Johnstone (1971) identified a need to consider stand density and age when he developed allometric models for 100-year-old lodgepole pine in Alberta, Canada. Since that study, Pearson et al. (1984) suggested that differences in stand density, age, and site quality may have caused considerable variability in tree crown biomass, and both Pearson et al. (1984), and Comeau and Kimmins (1989) found that the foliage biomass: sapwood area ratio decreased with increasing stand density. Variability among sites suggests that allometrics are also likely to differ among geographic locations due to differences in substrate, topography, and climate. Keyes and Grier (1981) found that Douglas-fir 2 had proportionately more total root biomass on a low productivity site than on a high productivity site, and Comeau and Kimmins (1989) found that belowground production represented a greater proportion of total production in two xeric sites compared to two mesic sites. Therefore, changes in allocation of biomass with differing site productivities may cause changes in allometric relationships across gradients in site productivity that could be caused by differences in substrate, topography, and climate. Objectives of this study were to: (1) develop allometric models for predicting above and belowground biomass of mature lodgepole pine trees in the GYE; (2) determine how allometric model development and application for lodgepole pine differs with stand density and age; and (3) compare and contrast allometric equations developed in this study to allometric equations developed in other locations to determine model variability and applicability across geographic locations independent of forest structure. We hypothesized that (1) allometric models developed for estimating biomass of above and belowground tree components in the GYE would vary with stand density and age; and (2) that allometric models developed for lodgepole pine in the GYE will differ from models developed elsewhere due to differences in substrate, topography, and climatic conditions. METHODS Study Area The study area was located within the GYE on the Caribou-Targhee National Forest (CTNF) bordering YNP. The GYE encompasses portions of three states in the western US: Wyoming, Idaho, and Montana, and is centered around YNP and its surrounding mountains, valleys, and subalpine plateaus. Elevations in YNP range from 1,620 m near Gardiner, Montana 3 to 3,333 m in the Absaroka mountain range of Wyoming. Mean annual precipitation ranges from less than 28 cm yr-1 at lower elevations to about 180 cm yr-1 on the southwestern plateau (Knight 1994). Temperatures are cold during the winter, where high temperatures less than 0C occur for an average of 87.6 days year-1, and temperatures are rarely more than 32C during the summer (Dirks and Martner 1982). There are three main substrates for soil development on the subalpine plateaus of the ecosystem: the least fertile rhyolite; the less infertile rhyolite; and lake bottom sediments (Despain 1990). The dominant forest type of the ecosystem is lodgepole pine forest, which occurs at middle elevations. Spruce/ Fir (Picea engelmannii /Abies lasiocarpa) forests occur at higher elevations and Douglas-fir (Pseudotsuga menziesii) forests occur at lower elevations. Whitebark pine (Pinus albicaulis) dominates at the upper treeline and sagebrush (Artemesia sp.) occurs at the lower treeline. Aspen (Populus tremuloides) occurs more commonly within the ecosystem outside of YNP (Knight 1994). Field and Lab Methods Allometric equations were developed in three lodgepole pine stands on the CTNF that represented two age classes and two density classes (Table 1). In the young (64 years old) aggrading age class, two stands of different densities were examined; one dense (YD) (2,452 trees/ha) and the other sparse (YS) (725 trees ha-1). The two young stands were located within the CTNF near Island Park, ID. Because densities of lodgepole pine stands tend to converge as they get older (Kashian et al. 2005), a single sparse (674 trees ha-1) stand was sampled in the older age class (OS) (164 years old). Although sites differed in density and age, they were located on similar soils. The Koffgo soil series consists of loamy-skeletal, mixed, superactive Vitrandic Cryochrepts. Parent 4 material was local residuum, colluvium, or alluvium developed from volcanic ash, igneous rocks, and loess (Bowerman et al. 1999). Elevations ranged from 1951m to 2249m, which is within the elevation range of lodgepole pine found for the GYE. Precipitation was approximately 114.3 cm yr-1 in the oldest stand and approximately 76.2 cm yr-1 in the younger stands (Dirks and Martner 1982). All sites were located at least 50 m from the road to facilitate equipment hauling and to avoid road influences. Stand basal area was similar between the OS (16.84 m2 ha-1) and YS stands (19.71 m2 ha-1), but was quite different for the YD stand (28.32 m2 ha-1) (Table 1). All aboveground tree biomass for a total of 46 trees was harvested within the three stands, and 24 root systems were excavated to develop allometric equations where easily obtained morphological parameters, such as diameter at breast height, were tested as predictors of above and belowground tree components (foliage biomass, coarse root biomass, etc.). Fourteen trees were harvested in the YD stand and 15 trees were harvested in the YS stand; 17 were harvested in the OS stand. For belowground components, 5 root systems were excavated in both the YS and OS stands due to logistical difficulties associated with large root systems, while 14 root systems were harvested in the YD stand. Initially, we decided to harvest 15 trees in each stand. However, in the OS stand, three extra trees were harvested to ensure that all trees were within the proper age range, and one tree had to be removed because it was younger than the acceptable age range (150-165 years) for that stand. In addition, some field data for one tree in the YD stand was missing, and therefore, that tree was not included in the analyses. Five trees were harvested along each of three 25m transects in each stand. Trees were generally selected at 5m intervals along each transect, but more importantly, trees were selected based on their diameter to represent the range of tree sizes found for trees in their respective stands. Trees were not chosen if they had any of the following characteristics: unusually poor 5 tree form, such as crook or sweep of the bole; major forking of the bole; excessive mistletoe; any defect that would alter the biomass of the tree, such as heart rot or insect damage; or any tree outside of the acceptable age range (15 years of the oldest tree in the stand). Prior to harvest, DBH (diameter at breast height, 1.37m) was measured and crown width was estimated using a meter tape. After felling of the tree, total height and height to crown base were measured. Crown base was defined as the point along the bole at the bottom of roughly 90% of the crown mass, and crown length was calculated as: CL = H – HCB (1) where CL = crown length, H = total tree height, and HCB = height to the base of the live crown. Aboveground Components Tree Bole - Each bole was harvested at ground level and all branches were removed. The bole was cut into three to four sections with 1 to 2 discs cut out as subsamples to determine moisture content for dry weight. For each bole, two discs were always taken at DBH, and at 90% of crown base (the location on the bole where most of the crown began). Two discs were taken from each section if the bole forked near the top or if DBH and crown base were one meter or closer in proximity. Each bole section was weighed separately using a digital hanging scale and the disc weight of the subsample was added back into the weight for its respective section. Discs were then dried to a constant weight at 70C in the lab to determine moisture content. Dry: wet weight ratio for each disc was then applied to determine dry weight of the entire bole section. For each DBH and crown base subsample, the following measurements were taken for determining sapwood area: phloem + bark thickness, total diameter, and heartwood diameter. Two perpendicular measurements were taken and averaged for each of these parameters except for diameter at breast height, which was measured with a diameter tape prior to harvest. 6 Sapwood diameter can be determined by subtracting the diameter of phloem + bark and heartwood diameter from total diameter. Therefore, the following equation was used to determine sapwood area: SA = a – b (2) where SA = Sapwood Area, a = the basal area excluding phloem and bark, and b = heartwood area. Branches - Branches were cut flush with the tree bole and were separated from foliage at 6.4 mm in diameter. Thus, branches consisted of all shoots minus the tree bole that were greater than 6.4 mm in diameter, because biomass smaller than 6.4 mm are likely to be consumed by fire (Despain 1990), allowing for post-fire estimates of branch biomass, independent of small twigs that were likely to be consumed the fire. Branches were compiled rather than separated into three crown sections, because one subsample from all sections was deemed sufficient to determine the dry weight of the entire component. A subsample of approximately 4.0 L was taken to determine moisture content for dry weight for each tree, where it was then dried in the lab. Fine Fuels - The fine fuels component was considered to be all needles and associated twigs less than 6.4 mm in diameter. These were combined because needles and twigs smaller than 6.4 mm diameter are likely to be consumed by fire (Despain 1990). The fine fuels component was maintained in separate piles of lower, middle, and upper crown sections for each tree, since foliage moisture contents were likely to vary with crown height (Brown 1978). A random subsample approximating the size of approximately 4.0 L volume was taken from each crown section to determine moisture content. The fine fuels subsamples were then weighed to 7 obtain wet weight. If immediate weighing was not possible, subsamples were stored in a cooler for no more than 5 days in plastic bags, to prevent moisture loss prior to weighing. The current year’s growth (2004) was removed and discarded, because many of the needles were not fully expanded, and samples were collected at varying times throughout the summer of 2004. The subsample was weighed again to determine the proportion of the fine fuels component from the current year’s growth. This proportion was then subtracted from the entire foliage component. After weighing, a random sample of 10 needle fascicles was taken from each fine fuels subsample, and the length and width of each fascicle was measured to determine leaf area. A tapered, bisected cylinder was used to determine surface area represented by each fascicle (Pearson et al. 1984 ; Madgwick 1964). This sample was dried and subsequently weighed so that surface area of this subsample could be extrapolated to the entire tree’s needle biomass. This sample was also weighed while still wet for subsequent drying and weighing to determine moisture content for dry weight determination of total needle biomass. The dry weight proportion of total foliage biomass that was needles was determined from separating needles from twigs after the foliage subsamples were dried. Belowground Components For each tree, the entire coarse root system (>10mm diameter) was excavated with the encouragement of a backhoe or come-a-long. Prior to excavation, smaller roots (≈ 10-20 cm) that could potentially be damaged by the backhoe or other excavation techniques were removed by hand. After excavation, the root system was divided into four size classes: root crown (i.e. the massive structure directly beneath the tree bole), lateral roots >50mm in diameter, lateral roots 25-50mm in diameter, and lateral roots 10-25mm in diameter. Total weight of each root size class was weighed using a digital hanging scale (Salter-Brecknell). Subsamples were taken 8 and weighed to determine moisture content for dry weight of each size class. Subsamples were then dried at approximately 70°C where the dry: wet weight ratio was applied to its corresponding size class for determination of total coarse root system dry weight. Statistical Analyses Model Development All allometric models were developed with SPSS 13.0 (SPSS Inc. 2005). Models for each tree component (Table 2) were developed for the three individual stands, for all sites combined, and were also pooled by density and age. The following criteria were used to develop the best models for predicting biomass of lodgepole pine: A.) The model must be biologically reasonable (Hilborn and Mangel 1997). For instance, an exponential model may yield the highest coefficient of determination (R2) for predicting total coarse root biomass with tree basal area, but an exponential increase in root biomass with basal area is not biologically reasonable. B.) The model must be of a form (linear or non-linear) that fits the data, although a model that does not produce a proper fit may achieve a higher R2. The proper model form was chosen using the following methods: 1. All potential predictors were plotted against each biomass component. 2. For any given plot, if a relationship was observed, data were examined to determine its linearity of curvilinearity. 3. To detect more subtle patterns of linearity or curvilinearity of data, the predictor was plotted against its residuals. If the relationship was linear ( y = ax+b ), a shotgun pattern around the horizontal axis through the origin was apparent. If the pattern appeared to be curvilinear, a curvilinear analysis was chosen. A nonlinear 9 power function (y = axb, where y was the dependent variable, x was the independent variable, and a and b were constants) best fit much of our data. 4. Where more than one predictor exhibited a non-linear relationship, coefficients of determination (R2), mean square errors, and plots of residuals were used to determine the best model. In addition, multiple predictors were combined into a multiplicative power model ( y = a xb x2c ) and compared to models with one predictor. C. If two predictors were deemed to be relatively equal based on their biological plausibility, coefficient of determination (r2/R2), standard error / mean square error, and plot of the residuals, the most easily measured independent variable was chosen. Model Comparison Density and Age To determine whether allometric models differed between stands of varying densities and ages, the sum of squares of the residuals for each model, pooled by density and age, were compared using the extra sum of squares analysis for nested models (Bates and Watts 1988). The null hypothesis was that slopes and intercepts of the models were the same. If either slopes or intercepts differed significantly ( < 0.05), it was determined that the models would produce biomass estimates significantly different from one another. In addition, equations for total aboveground biomass and coarse root biomass were used to predict biomass in a different stand from where they were developed. For instance, pooled equations from the two young stands were used to predict above and belowground biomass in the older stand, to determine the degree of error produced from using an inappropriate model. 10 Geographic Location Comparisons Models for mature lodgepole pine forests in this study were compared to models developed by Pearson et al. (1984) in the Medicine Bow Mountains of southeastern Wyoming and by Comeau and Kimmins (1989) in British Columbia. These models were used to estimate biomass using the same measured parameters used to develop allometric models for this study. Selected models were similar in stand density and age to allometric models developed for this study. Comparison was further facilitated by selecting sites from this study that were similar to stands used to develop the models for Pearson et al. (1984) and Comeau and Kimmins (1989). Paired, two-tailed t-tests were used to assess whether biomass estimates from this study differed statistically from biomass estimates produced from other models (Comeau and Kimmins 1989); (Pearson et al. 1984) ( = 0.05). In addition, actual biomass values from this study were compared against values estimated from the application of our allometric models to determine whether our estimates biomass values were more similar to the actual values than estimated values calculated from the application of models developed by Pearson et al. (1984) and Comeau and Kimmins (1989). RESULTS Model summary Forty-eight allometric models for all measured tree components are shown in Table 2. R2 values ranged from 0.54 to 0.99, and 37 out of 48 of the models had R2 values greater than 0.80. Generally, for any given component, model strength decreased with increasing tree size, indicating greater variability in the biomass of larger trees than smaller trees. Volume (basal area (cm2) * total height (cm)) was the best predictor of total aboveground and bole biomass 11 across all stand densities and ages. Basal area (cm2) was the best predictor of coarse root biomass, root crown biomass, and lateral root biomass. Best predictors of branch biomass were basal area and crown length, while basal area and sapwood area best predicted foliage and needle biomass (Table 2). Allometric models for total aboveground biomass, bole biomass, total coarse root biomass, and lateral root biomass were the most robust models, because only 1 of those 24 models had R2 values below 0.89. In contrast, the other 24 models, which were for root crown, branches, fine fuels, and needles were less robust, because R2 values were below 0.80 for 9 of the 24 models. For total aboveground biomass, the best model was for all sites combined (R2 = 0.95). Allometric models indicated that pooling the stands by age (R2(old) = 0.94, R2(young) = 0.95) produced better models than pooling stands by density (R2(dense) = 0.84, R2(sparse) = 0.94). The same pattern was observed for bole biomass, branch biomass, and root biomass components (Table 2). An exception for this is lateral root biomass where 94% of the variation in lateral root biomass was explained by basal area after developing separate models stratified by age, compared to stratification by stand density where 89% and 91% of the variation in lateral root biomass was explained by basal area for the two sparse stands and the single dense stand, respectively. Model Comparisons Between Densities and Ages Where statistical model comparisons were possible, most allometric models developed differed significantly between densities and ages (p < 0.05, Table 3.). Although formal statistical comparisons were not possible if either the model form or the predictor variables between two models were considered to be different, those models were nevertheless considered to be qualitatively different, because the allometric relationships were different. However, models for the tree bole (extra sum of squares, p = 0.622), root crown (extra sum of squares, p = 0.396), and 12 lateral roots (extra sum of squares, p = 0.608) were not significantly different when stands were pooled by density, and models for root crowns (extra sum of squares, p = 0.036) and lateral roots (extra sum of squares, p = 0.020) were only modestly different. In addition, equations for total aboveground biomass were less different when pooled by density (extra sum of squares, p = .037) than by age (extra sum of squares, p < 0.001). Model Comparison Between Geographic Locations Generally, predicted biomass values from equations developed in this study were closer to actual biomass of lodgepole pine in the Greater Yellowstone Ecosystem than predicted values from equations developed by Comeau and Kimmins (1989) and Pearson et al. (1984) (paired ttest, p<0.05, Table 4, Figure 2 and 3). Predicted values applying our study’s equations typically produced biomass values that were not significantly different (paired t-test, p > 0.05) from actual biomass values. However, estimates of branch biomass applying Pearson et al.’s equations were slightly more similar to actual values (paired t-test, p = 0.005) than were estimates values applying our equations (paired t, test, p = 0.003). Although Pearson et al.’s models appeared to be better at predicting actual branch biomass in the GYE according to paired t-tests, more indepth examination of Table 4 revealed a different pattern, because the mean value for branch biomass was closer to the actual value when applying this study’s equations. In addition, Figure 3 identified a clear departure of Pearson et al.’s values from the actual values with increasing tree diameter, while this study’s predicted values followed the actual values more closely with increasing diameter. An additional caveat is for the comparison between bole biomass for this study and bole biomass for Pearson et al.’s study, because Pearson et al’s predicted values for bole biomass were statistically similar (paired t-test, p = 0.063) to actual bole biomass from this study. However, predicting bole biomass with our study’s equations produced predicted bole 13 biomass values that were more similar to actual bole biomass values from this study (p = 0.514). The greatest difference in observed versus predicted biomass between studies was for larger trees (Table 4, Figure 2 and 3), supporting the observation of increased variation in allometric models with increasing tree size. Biomass estimates produced by models from studies northwest (Comeau and Kimmins 1989) and southeast (Pearson et al. 1984) of the GYE produced significantly lower estimates of root biomass, higher estimates of needle and branch biomass, and relatively comparable estimates of bole biomass (Table 4, Figure 2, 3). The root biomass estimates generated from models developed in the Medicine Bow Mountains produced slightly lower biomass estimates than from this study (Table 4, Figure 3). However, a portion of this difference is probably attributed to differences in the definition of a root crown between studies. For this study, a root crown was the portion of the coarse root system that is directly below the basal swell of the tree bole and within the lateral extent of the diameter of the tree at ground level. In contrast, Pearson et al. (1984) defined the root crown as the structure directly below the basal swell of the tree bole extending laterally to a distance of 1.5 x DBH. DISCUSSION Variation in stand density and age affected which of several measured tree morphometric parameters would best predict biomass of lodgepole pine tree components. The product of tree basal area and total height (volume) was the best predictor of total aboveground and bole biomass (Table 2). Notably, a tree’s ability to produce stemwood biomass, a major component of tree bole and total aboveground biomass, is strongly influenced by site productivity (Barnes et al. 1980). Therefore, a model including tree height, a variable relating more strongly to site productivity than any other measured parameter (Barnes et al. 1980), can explain a large amount 14 of the variability in bole and total aboveground biomass. In contrast, tree height was not found to be a useful variable for predicting the biomass of other tree components (Table 2). In addition, the decrease in variability of the dependent variable that can be explained by its respective predictor with increasing tree size may be due to greater variability in biomass allocation patterns with increasing tree size. Total coarse root biomass (>10 mm), root crown biomass, and lateral root biomass are best explained by tree basal area (Table 2), where tree diameter and basal area are measured parameters most closely correlating with spacing effects related to tree density (Barnes et al. 1980), suggesting coarse root biomass is at least partly dependent on tree spacing. Further studies incorporating nearest neighbor methods may be needed to determine effects of spacing on the coarse root biomass of individual trees, such as the approach by (Mainwaring and Maguire 2004) where they modeled the relationship between spacing among individual trees and tree diameter growth for lodgepole and ponderosa pine in central Oregon. They found that spatially explicit models including distance-dependent measures of competition among trees explained more of the variability in tree diameter growth than did implicit measures of competition based on stand-level averages. Their results suggested that allometric models have the potential to be improved further by including explicit measures of competition. Most of the variability in needle and foliage biomass is explained by basal area and sapwood area (Table 2). Sapwood area has been found to be a good predictor of foliage biomass in other studies (Snell 1978; Grier and Waring 1974; Pearson et al. 1984; Comeau and Kimmins 1989), because it is a good measure of a tree’s ability to conduct water and nutrients (Taiz and Zeiger 2002). Needle and foliage biomass in the older stand was best explained either by sapwood area or a combination of sapwood and basal area (Table 2). This indicates foliage 15 biomass may be better explained by a tree’s ability to conduct water and nutrients in older forests where water and nutrients are more limiting (Smith and Long 2001; Berger et al. 2004; Ryan et al. 2004). In contrast, basal area was the single best predictor of foliage biomass in younger stands (Table 2), indicating that foliage biomass in younger forests is better explained by tree spacing rather than by the tree’s ability to conduct water and nutrients. Differences between densities and ages Allometric models were expected to differ between densities and ages due to differences in biomass allocation patterns. For the most part, this was supported by the data (Table 3). However, models for tree bole, root crown, and lateral roots did not differ when pooled by density, and models for aboveground biomass were less different when pooled by density than by age. One plausible explanation for this pattern is that stand densities may not have been different enough for some of the models to differ according to stand density or to be less different than for models pooled by age. Although 2,452 trees ha-1 was considered in this study to be a dense stand, “dog-hair” stands of lodgepole pine that are of comparable age can sometimes reach densities higher than 10,000 trees ha-1 (Koch 1996). Therefore, the relatively small difference in stand densities in this study may be a factor leading to the relative similarity of models pooled by density compared with models pooled by age. Although allometric equations appear to be more similar across densities, application of models significantly different in density from where they were developed should be approached with caution. Furthermore, inappropriate model use can cause significant errors when extrapolating allometric equations to the landscape scale. The estimated errors produced by inappropriate model use appeared to be related to differences in aboveground: belowground biomass allocation, where an overestimation of 16 aboveground biomass was coupled with an underestimation of belowground biomass. Predicting biomass in the OS stand using the pooled model from the younger stands (YS and YD) produced tremendous error, where total aboveground biomass was 127.0 Mg/ha with the proper model for that stand, while it was nearly double (215.4 Mg ha-1) using the improper model. For total coarse root biomass, the error was less pronounced, but still evident, where it was 12.1 Mg ha-1 for the appropriate model and 10.2 Mg ha-1 for the inappropriate model. Predicting biomass in the sparse stand using the model developed in the dense stand underestimated total aboveground biomass significantly, where it was 85.0 Mg ha-1 for the correct model, but was 61.5 Mg ha-1 for the incorrect model. The inappropriate model overestimated for total coarse root biomass, it was 10.5 Mg ha-1 for the appropriate model and 13.6 Mg ha-1 for the inappropriate model. Differences between Geographic Locations Differences in allometric equations among geographic locations are likely attributed to differences in soil, topography, and climate. The soils on which Pearson et al. (1984) developed allometrics in the Medicine Bow Mountains were generally less infertile and were derived primarily from glacial till and fluvial conglomerates, while soils in British Columbia were Orthic Eutric Brunisols for xeric sites and Brunisolic Gray Luvisols for mesic sites (Comeau and Kimmins 1989). In contrast, soils of this study were more infertile and were developed on volcanic substrates (Bowerman et al. 1999). Sites where allometrics were developed by Comeau and Kimmins (1989) were more productive than sites in this study, where higher site index (SI), which is associated with higher productivity, ranged from 14.3 to 20.5m at 100 years for Comeau and Kimmins (1989) compared to our study where SI ranged from 14.3 to 16.0 at 100 years. 17 Bole biomass was the only component that did not differ between the GYE and the Medicine Bow Mountains (Pearson et al. 1984), and was the component most similar between the GYE and British Columbia (Comeau and Kimmins 1989), indicating that relationships between bole biomass and its predictor(s) (basal area and total height) are relatively constant across gradients of elevation, topography, climate, and site productivity. This further indicates that a tree will allocate a relatively constant proportion of available resources throughout its lifespan to wood production, indicating a tree may compensate for reductions in root and foliar biomass through increases in light, nutrient, and overall photosynthetic efficiency. However, increases in light use efficiency typically coincide with increases in leaf area (Smethhurst et al. 2003). Therefore, the ability of a tree to produce wood across sites may be controlled by interactions between nutrient and light use efficiency, where a reduction in light use efficiency is coupled with an increase in nutrient use efficiency (Binkley et al. 2004). This finding is consistent with Tilman's (1982) resource ratio model of plant competition that implies stability among competing plants at different ratios of important resources, such as nitrogen and light (Gurevitch et al. 2002). Despite the relatively constant relationships between bole biomass and its predictors across geographic locations, predicted bole biomass values when applying our equations as opposed to those developed by Pearson et al. (1984) were closer to actual bole biomass (Table 4, Figure 3), indicating the need for using locally developed allometric models. Needle biomass estimates were lower for less nutrient poor sites (Pearson et al. 1984; Comeau and Kimmins 1989) than for actual and predicted biomass from sites in the GYE (Table 4, Figure 3), indicating foliage biomass was higher increasing photosynthate to compensate for more nutrient limiting conditions that might otherwise decrease wood production. This potential 18 balance between light use and nutrient availability may account for some of the homogeneity in bole biomass across geographic locations. Root biomass in the GYE was over predicted by models developed in British Columbia and southeastern Wyoming, indicating root crown and coarse root biomass is dependent upon a site’s ability to support production of wood biomass. This suggests that coarse root biomass has little impact on the trees ability to uptake water and nutrients, because less coarse root biomass should have been needed to uptake water and nutrients in British Columbia and southeastern Wyoming, which are less productive sites than the GYE. The greatest difference in biomass of tree components across geographic locations was for larger trees (Figure 2, 3), indicating site conditions become increasingly important as forests become older and trees become taller. This is attributed to an increase in nutrient limitation (Berger et al. 2004; Ryan et al. 1997; Smith and Resh 1999; Smith and Long 2001), hydraulic limitation (Ryan and Yoder 1997), and maintenance respiration (Ryan et al. 1995) with increases in stand age and tree height. SUMMARY AND CONCLUSIONS Allometric models developed for mature lodgepole pine in the Greater Yellowstone Ecosystem were very robust for predicting tree biomass and showed promise for estimating landscape level biomass of lodgepole pine forests, because R2 values were often above 0.80. Allometric equations were found to differ between stand densities and ages according to statistical tests and qualitative observations, and they were also identified to be different among geographic locations independent of stand density and age. In addition, applications of inappropriate models sometimes produced tremendous errors and have the potential to do so in 19 further applications. Therefore, wise use of these models is critical, as application of the appropriate model to the appropriate stand is vital due to differences identified by this study among forest structures and geographic locations. We strongly suggest that models not be applied to stands differing significantly in density and age from the stands where the models were developed, and that models not be applied to geographic locations of considerable distance from where the models were developed. In addition, allometric models developed in this study have many potential uses from determination of NPP for carbon studies using multi-year measurements, fire modeling, and for determination of wood biomass by foresters. 20 Table 1. Sites for the development of allometric models in the Greater Yellowstone Ecosystem. Soils for all three sites were in the Koffgo series (see Methods). NAD 83, UTM Zone Stand Density 12 Stand (trees > 5cm Northing Easting Site Elevation Age DBH per Stand basal area (m) (m) Name (m) (years) hectare) (m2 per hectare) Grassy Lake 2249 4886015 511735 165 674 175 16.84 14.3 Coffee Pot 1951 4926541 472232 64 725 72 19.71 11.4 US 20 1951 4925932 472657 64 2452 929 28.32 30.4 21 Table 2. Allometric models for predicting biomass of eight different above and belowground components of P. contorta in the Greater Yellowstone Ecosystem. Non-linear power functions are listed first, and linear equations are at the end of the table. MSE is the mean square error for non-linear model and SE is the standard error of the estimate for linear the model. X is the morphometric predictor variable and Y represents the response variable (biomass (Kg)). The subscript 2 is for predictors that are associated with the coefficient c in non-linear models. Non Linear Power Functions (y = axb, or y = axbx2c) (Y) Total Aboveground Biomass Tree Bole Total Coarse Root Biomass (>10mm) Root Crown Biomass Lateral Root Biomass (>10mm) Branches (X) Volume Volume Basal Area Basal Area Basal Area Basal Area, Crown Length2 Basal Area - Site(s) All Sites Old, Sparse 2 Young Stands 2 Sparse Stands Young, Dense Young, Sparse Old, Sparse All Sites Old, Sparse 2 Young Stands 2 Sparse Stands Young, Dense Young, Sparse All Sites Old, Sparse 2 Young Stands 2 Sparse Stands Young, Dense Young, Sparse All Sites Old, Sparse 2 Young Stands 2 Sparse Stands Young, Dense Young, Sparse All Sites Old, Sparse 2 Young Stands 2 Sparse Stands Young, Dense Young, Sparse DBH range (cm) 5.4 - 33.3 11.3 - 33.3 5.4 - 25.0 11.3 - 33.3 5.4 - 15.6 11.7 - 25.0 11.3 - 33.3 5.4 - 32.0 11.3 - 32.0 5.4 - 24.3 11.3 - 32.0 5.4 - 15.6 12.7 - 24.3 5.4 - 32.0 11.3 - 32.0 5.4 - 24.3 11.3 - 32.0 5.4 - 15.6 12.7 - 24.3 5.4 - 32.0 11.3 - 32.0 5.4 - 24.3 11.3 - 32.0 5.4 - 15.6 12.7 - 24.3 5.4 - 33.3 11.3 - 33.3 5.4 - 25.0 11.3 - 33.3 5.4 - 15.6 11.7 - 25.0 22 n 46 17 29 32 14 15 17 24 5 19 10 14 5 24 5 19 10 14 5 24 5 19 10 14 5 46 17 29 32 14 15 a 0.005 0.003 0.0001 0.010 0.00004 0.0004 0.002 0.028 0.188 0.006 29.939 0.005 0.002 0.020 0.224 0.006 0.050 0.007 0.007 0.008 0.010 0.001 0.011 0.0004 0.00003 0.022 0.003 0.001 0.042 0.002 0.004 b 0.793 0.817 1.088 0.741 1.155 1.001 0.822 1.139 0.845 1.406 -0.179 1.467 1.579 1.109 0.745 1.304 0.968 1.267 1.284 1.193 1.146 1.564 1.147 1.738 2.153 0.683 1.136 1.855 0.528 1.548 1.497 c 1.276 0.850 -0.291 1.288 - R2 0.95 0.94 0.95 0.94 0.84 0.92 0.93 0.95 0.93 0.97 0.99 0.98 0.95 0.84 0.71 0.95 0.73 0.93 0.89 0.94 0.94 0.94 0.89 0.91 0.94 0.69 0.86 0.88 0.54 0.91 0.76 MSE 679.6 758.4 289.4 810.0 109.9 445.2 608.5 11.8 36.4 2.0 5.8 0.3 8.3 13.6 65.5 1.1 33.8 0.2 5.3 2.1 4.8 1.0 5.2 0.2 2.6 69.3 21.6 34.6 96.2 0.4 67.9 Fine Fuels Needles Basal Area, Sapwood Area2 Basal Area Sapwood Area Basal Area Basal Area, Sapwood Area2 Basal Area Sapwood Area Basal Area - All Sites Old, Sparse 2 Young Stands 2 Sparse Stands Young, Dense Young, Sparse All Sites Old, Sparse 2 Young Stands 2 Sparse Stands Young, Dense Young, Sparse (X) Volume - Site(s) All Sites 2 Young Stands 2 Sparse Stands Young, Dense Young, Sparse 5.4 - 33.3 11.3 - 33.3 5.4 - 25.0 11.3 - 33.3 5.4 - 15.6 11.7 - 25.0 5.4 - 33.3 11.3 - 33.3 5.4 - 25.0 11.3 - 33.3 5.4 - 15.6 11.7 - 25.0 46 17 29 32 14 15 46 17 29 32 14 15 0.418 0.524 0.007 1.204 0.002 0.011 0.192 0.249 0.007 0.563 0.002 0.014 0.466 0.323 1.512 0.638 1.695 1.427 0.525 0.323 1.426 0.701 1.600 1.312 0.418 0.403 0.317 0.471 - 0.68 0.81 0.93 0.52 0.96 0.86 0.73 0.82 0.89 0.61 0.91 0.78 184.8 58.3 48.7 220.9 1.4 97.1 67.0 33.0 30.7 78.5 1.5 60.3 a 0.0002 0.0002 0.0002 0.0002 0.0002 b 3.420 5.482 1.921 -0.327 0.055 c r2 0.96 0.93 0.95 0.79 0.95 SE 17.9 8.9 20.8 8.9 7.6 ( Linear Models, y = ax + b ) (Y) Tree Bole DBH range (cm) 5.4 - 33.3 5.4 - 25.0 11.3 - 33.3 5.4 - 15.6 11.7 - 25.0 23 n 46 29 32 14 15 - Table 3. Allometric model comparison between densities and ages of mature P.contorta stands in the Greater Yellowstone Ecosystem. Results are from the extra sum of squares analysis for nested models (Bates and Watts 1988). All possible statistical comparisons are shown. Models for comparisons not in this table had different model forms. Dependent Variable Model Comparison F p-value Total Aboveground Biomass Old vs. Young 11.726 <.0001* Total Aboveground Biomass Sparse vs. Dense 3.514 0.037* Branches Old vs. Young 19.706 <.0001* Total Coarse Roots (>10mm) Old vs. Young 8.128 <.0001* Total Coarse Roots (>10mm) Sparse vs. Dense 57.919 <.0001* Root Crown Old vs. Young 3.937 0.036* Lateral Root Biomass (>10mm) Old vs. Young 4.790 0.020* Bole Sparse vs. Dense 0.481 0.622 Root Crown Sparse vs. Dense 0.971 0.396 Lateral Root Biomass Sparse vs. Dense 0.508 0.608 * p-values are significantly different ( = 0.05) Values in bold were not significantly different 24 Table 4. Comparison of actual P. contorta individual tree biomass vs. estimates from this study in the Greater Yellowstone Ecosystem and from studies in British Columbia by Comeau and Kimmins (1989) and Pearson et al. (1984). All p-values < 0.05 are in bold. The upper p-value for each component is the comparison between actual values from this study and predicted values from this study, and the lower p-value is for the comparison between actual values from this study and predicted values applying the equations from other studies. Component Bole Branches Needles Total Root Bole Branches Needles Root Crown Minimum Biomass (Kg) Maximum Biomass (Kg) Mean Biomass (Kg) Age Range Density Range This Study (Actual) 10.2 136.8 50.7 64 725 - 2452 This Study (Predicted) 3.3 135.5 51.1 - - 0.804 Comeau and Kimmins 6.7 170.9 67.9 70 - 78 1770 - 3580 <0.001 This Study (Actual) 0.3 55.9 14.0 64 725 - 2452 This Study (Predicted) 0.3 47.6 14.9 - - 0.401 Comeau and Kimmins 0.7 33.2 9.6 70 - 78 1770 - 3580 0.037 This Study (Actual) 0.2 59.3 16.2 64 725 - 2452 This Study (Predicted) 0.6 48.1 16.5 - - 0.739 Comeau and Kimmins 0.4 24.5 5.5 70 - 78 1770 - 3580 <0.001 This Study (Actual) 0.7 33.8 8.1 64 725 - 2452 This Study (Predicted) 0.4 32.5 8.3 - - 0.400 Comeau and Kimmins 5.8 32.1 15.0 70 - 78 1770 - 3580 <0.001 This Study (Actual) 26.0 329.0 137.1 64 - 165 674 - 2452 This Study (Predicted) 7.0 339.8 106.3 - - 0.514 Pearson et al. 24.7 256.6 121.8 75 - 240 <2500 0.063 This Study (Actual) 1.7 55.9 21.7 64 - 165 674 - 725 This Study (Predicted) 3.1 29.1 16.0 - - 0.003 Pearson et al. 3.2 32.9 15.6 75 - 240 <2500 0.005 This Study (Actual) 1.6 44.7 25.8 165 674 This Study (Predicted) 8.5 45.4 25.9 - - 0.928 Pearson et al. 9.3 30.3 17.1 240 420 <0.001 This Study (Actual) 0.6 35.9 7.7 74-165 674 - 2452 This Study (Predicted) 0.5 33.4 7.9 - - 0.763 Pearson et al. -0.1 54.1 13.2 75 - 240 <2500 <0.001 Site 25 p-value Figure 1. Three Stands Differing in Densities and Ages for allometric model development of P. contorta in the Greater Yellowstone Ecosystem. Individual stands are depicted by a black square. 26 Figure 1: YS YD OS 27 Figure 2. Comparison of biomass values among those directly measured by this study, those predicted by this study’s allometric models, and those predicted by Comeau and Kimmins’ allometric models. 28 Figure 2. Branches Tree Bole 60 180 This Study (Actual) This Study (predicted) Comeau and Kimmins (predicted) 160 This Study (Actual) This Study (predicted) Comeau and Kimmins (predicted) 50 140 Biomass (Kg) 120 40 100 30 80 60 20 40 10 20 0 0 0 5 10 15 20 25 30 0 5 diameter at breast height (cm) 15 20 25 30 diameter at breast height (cm) Total Coarse Roots Needles 70 40 This Study (Actual) This Study (predicted) Comeau and Kimmins (predicted) 60 This Study (Actual) This Study (predicted) Comeau and Kimmins (predicted) 50 Biomass (Kg) 10 30 40 20 30 20 10 10 0 0 0 5 10 15 20 25 30 0 diameter at breast height (cm) 5 10 15 20 diameter at breast height (cm) 29 25 30 Figure 3. Comparison of biomass values among those directly measured by this study, those predicted by this study’s allometric models, and those predicted by Pearson et al.’s allometric models. 30 Figure 3. Tree Bole 400 This Study (Actual) This Study (predicted) Pearson et al. (predicted) 300 Biomass (Kg) Branches 60 This Study (Actual) This Study (predicted) Pearson et al. (predicted) 50 40 200 30 20 100 10 0 0 0 5 10 15 20 25 30 35 0 diameter at breast height (cm) 10 20 25 30 35 Root Crown 60 This study (Actual) This study (predicted) Pearson et al. (predicted) 40 15 diameter at breast height (cm) Needles 50 Biomass (Kg) 5 This study (Actual) This study (predicted) Pearson et al. (predicted) 50 40 30 30 20 20 10 10 0 0 5 10 15 20 25 30 35 0 5 diameter at breast height (cm) 10 15 20 25 diameter at breast height (cm) 31 30 35 LITERATURE CITED Amiro, B. D., J. M. Chen and J. Liu 2000. Net primary productivity following forest fire for Canadian ecoregions. Can. J. For. Res. 30: 939-947. Barnes, B. V., D. R. Zak, S. R. Denton and S. H. Spurr (1980). Forest Ecology. New York, NY, John Wiley & Sons, Inc. Bates, D. M. and D. G. Watts (1988). Biometrics: nonlinear regression and its applications. New York, NY, John Wiley & Sons. Berger, U., H. Hildenbrandt and V. Grimm 2004. Age-related decline in forest production: modelling the effects of growth limitation, neighbourhood competition and self-thinning. Journal of Ecology 92: 846-853. Binkley, D., J. L. Stape and M. G. Ryan 2004. Thinking about efficiency of resource use in forests. Forest Ecology and Management 193: 5-16. Bowerman, T. S., J. Dorr, S. Leahy, K. Varga and J. Warrick (1999). Targhee National Forest ecological unit inventory, United States Department of Agriculture, Forest Service United States Department of Agriculture, Natural Resources Conservation Service University of Idaho, College of Agriculture. 1-2: 787. Brown, J. K. 1978. Weight and density of crowns of Rocky Mountain conifers. Res. Pap. Int197. Ogden, UT: U.S. Department of Agriculture, Forest Service, Intermountain Research Station; 56 p. Comeau, P. G. and J. P. Kimmins 1989. Above- and below-ground biomass and production of lodgepole pine on sites with differing soil moisture regimes. Can. J. For. Res. 19: 447454. 36 Dale, V. H., L. A. Joyce, S. McNulty, R. P. Neilson, M. P. Ayres, M. D. Flannigan, P. J. Hanson, L. C. Irland, A. E. Lugo, C. J. Peterson, D. Simberloff, F. J. Swanson, B. J. Stocks and B. M. Wotton 2001. Climate change and forest disturbances. Bioscience 51: 723-734. Despain, D. G. (1990). Yellowstone vegetation: consequences and environment in a natural setting. Boulder, CO, Roberts Rinehart, Inc. Dirks, R. A. and B. E. Martner 1982. The climate of Yellowstone and Grand Teton National Parks. National Park Service Occassional Paper 6. Gholz, H. L., C. C. Grier, A. G. Campbell and A. T. Brown (1979). Equations for estimating biomass and leaf area of plants in the Pacific Northwest. Research Paper, Forest Research Laboratory, Oregon State University. 41: 39pp. Gurevitch, J., S. M. Scheiner and G. A. Fox (2002). The ecology of plants. Sunderland, MA, Sinauer Associates, Inc. Hilborn, R. and M. Mangel (1997). The ecological detective: confronting models with data. Princeton, NJ, Princeton University Press. Johnstone, W. D. (1971). Total standing crop and tree component distributions in three stands of 100-year-old lodgepole pine. Forest biomass studies. H. E. Young. Orono, ME, University of Maine Press. 15th IUFRO Congress: 81-89. Kashian, D. M., M. G. Turner and W. H. Romme 2005. Variability in leaf area and stemwood increment along a 300-year lodgepole pine chronosequence. Ecosystems 8: 48-61. Knight, D. H. (1994). Mountains and plains: the ecology of Wyoming landscapes. New Haven, CT, Yale University Press. Koch, P. (1996). Lodgepole Pine in North America. Madison, WI, Forest Products Society. 37 Litton, C. M., M. G. Ryan, D. B. Tinker and D. H. Knight 2003. Belowground and aboveground biomass in young postfire lodgepole pine forests of contrasting tree density. Can. J. For. Res. 33: 351-363. Madgwick, H. A. I. 1964. Estimation of surface area of pine needles with special reference to Pinus resinosa. J. For. 61: 636. Mainwaring, D. B. and D. A. Maguire 2004. The effect of local stand structure on growth and growth efficiency in heterogeneous stands of ponderosa pine and lodgepole pine in central Oregon. Canadian Journal of Forest Research-Revue Canadienne De Recherche Forestiere 34: 2217-2229. Pearson, J. A., T. J. Fahey and D. H. Knight 1984. Biomass and leaf area in contrasting lodgepole pine forests. Can. J. For. Res. 14: 259-265. Ryan, M. G., D. Binkley and J. H. Fownes 1997. Age-related decline in forest productivity: pattern and process. Advances in Ecological Research 27: 213-262. Ryan, M. G., D. Binkley, J. H. Fownes, C. P. Giardina and R. S. Senock 2004. An experimental test of the causes of forest growth decline with stand age. Ecological Monographs 74: 393-414. Ryan, M. G., S. T. Gower, R. M. Hubbard, R. H. Waring, H. L. Gholz, W. P. J. Cropper and S. W. Running 1995. Woody tissue maintenance repiration of four conifers in contrasting climates. Oecologia 101: 133-140. Ryan, M. G. and B. J. Yoder 1997. Hydraulic limits to tree height and tree growth. Bioscience 47: 235-241. Smethhurst, P., C. Baillie, M. Cherry and G. Holz 2003. Fertilizer effects on LAI and growth of four Eucalyptus nitens plantations. Forest Ecology and Management 176: 531-542. 38 Smith, F. W. and J. N. Long 2001. Age-related decline in forest growth: an emergent property. Forest Ecology and Management 144: 175-181. Smith, F. W. and S. C. Resh 1999. Age-related changes in production and below-ground carbon allocation in Pinus contorta forests. Forest Science 45: 331-341. Snell, J. A. K. 1978. Comparison of tree biomass estimators - DBH and sapwood area. Forest Science 24: 455-457. Taiz, L. and E. Zeiger (2002). Plant Physiology. Sunderland, MA, Sinauer Associates, Inc. Ter-Mikaelian, M. T. and M. D. Korzukhin 1997. Biomass equations of sixty-five North American tree species. Forest Ecology and Management 97: 1-24. Tilman, D. (1982). Resource competition and community structure. Princeton, NJ, Princeton University Press. Turner, M. G., D. B. Tinker, W. H. Romme, D. M. Kashian and C. M. Litton 2004. Landscape patterns of sapling density, leaf area, and aboveground net primary production in postfire lodgepole pine forests, Yellowstone National Park (USA). Ecosystems 7: 751-775. 39 Chapter 2 Biomass Allocation Patterns and Nitrogen Use Efficiency for Mature Lodgepole Pine Forests in the Greater Yellowstone Ecosystem INTRODUCTION Net Ecosystem Production (NEP) varies widely across many forested landscapes due to variability produced by heterogeneity in stand ages and densities as well as natural and anthropogenically altered disturbance regimes (Chapin et al. 2002; Turner et al. 2004). For example, in the Greater Yellowstone Ecosystem (GYE), recent and historic fires have created a complex mosaic of varying forest stand structures (Turner et al. 1997). Understanding the variability in NEP is, at least in part, dependent upon knowledge of how individual trees allocate biomass in response to such differences in forest structure. Lodgepole pine (Pinus contorta var. latifolia (Engelm. ex Wats.) Critchfield) dominates the forested plateaus of the GYE (Tinker et al. 1994), and therefore contributes greatly to the NEP of the ecosystem. The successional pathways of stand development for lodgepole pine vary widely after disturbances such as fire, where initial seedling densities may range from < 100 to > 500,000 seedlings/ha (Turner et al. 2004). As the forests mature, this heterogeneous landscape is somewhat homogenized by a convergence in stand density (Kashian et al. 2005), resulting in considerable variability in spatial and temporal patterns of biomass accumulation and net primary production (NPP) across the landscape (Litton et al. 2004) (Turner et al. 2004). Although previous studies have identified differences in biomass allocation patterns for forests differing in density (Pearson et al. 1984; Comeau and Kimmins 1989; Litton et al. 2003; Litton et al. 2004) and age (Smith and Resh 1999; Ryan et al. 1997; Berger et al. 2004), further understanding of the relationships between stand structural characteristics and allocation patterns 40 of mature lodgepole pine forests of the GYE is critical for our understanding of how natural and anthropogenic changes in forest structure will alter NEP. In this study we harvested lodgepole pine above and belowground in three stands differing in densities and ages to determine the allocation patterns of mature lodgepole pine forests in the GYE across varying forest structures. Allocation patterns for mature lodgepole pine forests have been described in several locations throughout the northern and central Rocky Mountains (Pearson et al. 1984; Comeau and Kimmins 1989; Litton et al. 2004). In British Columbia, Comeau and Kimmins (1989) observed that allocation to total root biomass was significantly higher in more xeric stands, indicating that allocation to roots was higher under more resource limited conditions, such as those present where stand density is high and stands are old. In the Medicine Bow Mountains of southeastern Wyoming, Pearson et al. (1984) also found that allocation to roots increased with increasing stand density; in contrast, Smith and Resh (1999) found no significant shift to belowground allocation with increasing stand age. In addition, Comeau and Kimmins (1989) observed that allocation to coarse roots >5mm diameter was greater in more mesic stands, indicating a need for further study of biomass allocation patterns. The GYE differs appreciably in substrate, topography, and climate from the locations of other studies, which may cause biomass allocation patterns to differ. Although Litton et al. (2004) found that the ratio of total belowground carbon allocation as a proportion of total tree biomass was remarkably constant across extreme gradients of stand density and age in the GYE, their estimates of allocation were based on 15 year old lodgepole pine forests, and considered only five trees in a mature 110-year-old lodgepole pine forest. Therefore, more extensive data for allocation patterns of mature lodgepole pine forests in the GYE are needed. Also, studies of the biomass allocation patterns of lodgepole pine have focused largely on stand level allocation 41 patterns (Pearson et al. 1984; Comeau and Kimmins 1989; Litton et al. 2004, Smith and Resh 1999); therefore, more in depth analyses of the allocation of biomass in lodgepole pine for individual trees are needed. Determining the use efficiency for essential nutrients such as nitrogen (N) is also important for improving our understanding of biomass accumulation of lodgepole pine. Gross Primary Production (GPP), a major component of NEP, is equal to the product of resource supply, the proportion of the resource supply that is taken up, and the efficiency of resource use (Binkley et al. 2004). Also, nitrogen is an essential part of plant proteins and amino acids, including the enzyme rubisco, which is critical to the Calvin cycle of photosynthesis (Taiz and Zeiger 2002). Some important factors affecting the N availability in lodgepole pine forests include the abundance of N-fixers, such as Lupinus argenteus and Shepherdia canadensis (Fahey et al. 1985; Hendrickson and Burgess 1989), litter quality (Fahey 1983; Stump and Binckley 1993; Thomas and Prescott 2000), the abundance of coarse dead wood (Fahey et al. 1985), microbial biomass (Litton et al. 2003), inputs from precipitation (Fahey et al. 1985), and leaching from the soil (Fahey et al. 1985). In lodgepole pine forests, nitrogen-fixers, such as Lupinus argenteus and Shepherdia canadensis have been shown to be important sources of nitrogen for lodgepole pine forests (Fahey et al. 1985; Hendrickson and Burgess 1989). Lignin: N ratios for lodgepole pine litter were found to be higher than paper birch and douglas-fir, which coincided with lower net mineralization of N (Thomas and Prescott 2000), indicating that the poor litter quality typical of lodgepole pine is associated with lower N availability. In southeastern Wyoming, Fahey et al. (1985) found high N storage in dead lodgepole pine boles lying on the forest floor. In YNP, Litton et al. (2003) found that microbial biomass was significantly higher in a 110 year old 42 lodgepole stand than younger < 15 year old stands, indicating that both immobilization and net mineralization of N could be higher in older lodgepole pine stands. Inputs of N in lodgepole pine forests of the Greater Yellowstone Ecosystem are probably limited, because in lodgepole pine forests of southeastern Wyoming, Fahey et al. (1985) found that inputs of N were nearly twice as high in rainfall compared to snowfall. Therefore, in the GYE and other lodgepole pine forests of the intermountain west, where the majority of annual precipitation comes as snow (Dirks and Martner 1982; Knight 1994), inputs of N through precipitation are likely to be low. In contrast, extremely low inputs of N to the groundwater in lodgepole pine forests as found by Fahey et al. (1985), indicate efficient uptake of N by lodgepole pine. In addition, production of older lodgepole pine forests in southeastern Wyoming have been shown by (Binkley et al. 1995) to respond to fertilization, indicating that they are N limited. Therefore, NUE in lodgepole pine forests is likely to be an important factor affecting biomass production. Nitrogen use efficiency (NUE) of lodgepole pine is the amount of biomass or C fixed per unit of nitrogen that is taken up (Vitousek 1982; Aber and Melillo 2001; Chapin et al. 2002). Vitousek (1982) found that across biomes, coniferous forests were more efficient at fixing biomass per unit of nutrient uptaken than more productive temperate deciduous forests, indicating that nitrogen use efficiency (NUE) should be higher where nutrients are more limiting (Ryan et al. 1997), such as dense and older forests. In addition, (Gower et al. 1989) suggested that high NUE in lodgepole pine and Larix occidentalis could be attributed to low nutrient availability of the volcanic soils in the Washington Cascades. In contrast, Olsson et al. (1998) found an inverse relationship between NUE and stand age in southeastern Wyoming that they attributed to a decrease in aboveground productivity caused by increased allocation to roots and mycorrhizal development. Therefore, it would appear that patterns of variability in the 43 efficiency of nutrient use with forest structure are poorly understood, and this may be especially true for lodgepole pine forests of the Intermountain West. We estimated NUE in three sites in the Greater Yellowstone Ecosystem that varied by stand age and tree density to determine how NUE relates to variability in forest structure and site index (see Methods), and to identify relationships between NUE and biomass allocation patterns. The specific objectives of this study were: (1) to determine how patterns of above and belowground biomass allocation for mature lodgepole pine forest of the GYE vary with stand age and density; (2) to estimate the carbon storage in three mature lodgepole pine stands differing in density and age of the Greater Yellowstone Ecosystem; (3) to quantify patterns of nitrogen use efficiency (NUE) for lodgepole pine forests among these differing forest structures and productivities; and (4) to examine the relationships between NUE and biomass allocation patterns. We hypothesized that patterns in biomass allocation and NUE of lodgepole pine forests in the GYE would vary with forest structure and geographic location where factors affecting nutrient availability are different, and we expected that biomass allocation would differ at multiple scales (i.e. stand level vs. individual tree level). Specifically, lodgepole pine should allocate a greater proportion of total biomass to coarse roots in more dense stands and older stands, and is likely to allocate a greater proportion of aboveground biomass to the tree bole rather than foliage with increasing stand age due to increased competition for water, light, and nutrients (Ryan et al. 1997). We expected that the efficiency of nitrogen use would increase with increasing stand density and age, because of increased competition for resources in older and denser stands. (Binkley et al. 1995) found that tree basal area in older lodgepole pine stands in 44 southeastern WY responded to N fertilization, but not fertilization by P and K, indicating that lodgepole pine forests are N limited. We also expected that nitrogen use efficiency would be higher where biomass allocation to roots was higher as found by Vitousek (1982), because NUE has been found to be higher where nutrients are more limiting (Vitousek 1982; Cote et al. 2002), such as the nutrient poor sites where allocation to roots has been found to be greater for douglas-fir in the Cascade Mountains of Washington, US (Keyes and Grier 1981). METHODS Study Area The study area was situated within the GYE on the Caribou-Targhee National Forest (CTNF) adjacent to YNP. The GYE includes portions of three states in the western US: Wyoming, Montana and Idaho, and is signified by Yellowstone National Park and its surrounding subalpine plateaus, mountains, and valleys. There are three main substrates for soil development on the subalpine plateaus of the ecosystem: the least fertile rhyolite; the less infertile rhyolite; and the less infertile lake-bottom sediments (Despain 1990). The dominant forest type of the ecosystem is lodgepole pine forest, which occurs at middle elevations. Aspen (Populus tremuloides) occurs more commonly within the ecosystem outside of YNP (Knight 1994). Douglas-fir (Pseudotsuga menziesii) forests occur at lower elevations, and Spruce/ Fir (Picea engelmannii /Abies lasiocarpa) forests occur at higher elevations. Whitebark pine (Pinus albicaulis) dominates at the upper treeline and sagebrush (Artemesia spp.) occurs at the lower treeline. 45 Elevations in YNP range from 1,620 m near Gardiner, Montana to 3,333 m in the Absaroka mountain range of Wyoming. Mean annual precipitation ranges from less than 28 cm/yr at lower elevations to about 180 cm yr-1 on the southwestern plateau (Knight 1994). Temperatures are cold during the winter, where high temperatures less than 0C occur for an average of 87.6 days each year, and are rarely more than 32C during the summer (Dirks and Martner 1982). Biomass Allocation Patterns Field and Lab Methods Research sites were situated within the Greater Yellowstone Ecosystem (GYE) on the Caribou-Targhee National Forest (CTNF) adjacent to Yellowstone National Park (YNP), where the topography, vegetative cover, and substrate were comparable. Allocation patterns were determined in three stands on the CTNF using two age classes and two density classes. Actual biomass estimates were used to determine allocation patterns of individual lodgepole pine, and allometric models (Arcano and Tinker Manuscript in preparation) were used to estimate biomass at the stand level. Because densities of lodgepole pine forests tend to converge as they get older (Kashian et al. 2005), a single sparse (674 trees ha-1) stand (OS) was sampled in the older age class (164 years old) (Figure 1). In the young (64 years old) aggrading age class, two stands were studied; one dense (YG) (2,452 trees ha-1) and the other sparse (YS) (725 trees ha-1). The two young stands were located within the CTNF proximal to Island Park, ID (Figure 1). Although sites differed in density and age, they had comparable soils (Table.1). The Koffgo soil series is composed of loamy-skeletal, mixed, superactive Vitrandic Cryochrepts, which is due to the super volcano located beneath YNP (Knight 1994). The parent material consisted of local residuum, 46 colluvium, or alluvium developed on volcanic ash, igneous rocks, and loess (Bowerman et al. 1999). Mean annual precipitation was approximately 76.2 cm yr-1 at the younger stands and 114.3 cm yr-1 at the oldest stand. Elevations ranged from 1951m to 2249m, which is within the range of elevations found for the GYE. A total of 46 lodgepole pines were harvested aboveground and 24 were excavated belowground to determine their biomass allocation patterns. All sites were located at least 50m from the road to enable equipment hauling and to avoid road influences. Fourteen trees were harvested in the YD stand and 15 trees were harvested in the YS stand; 17 were harvested in the OS stand. For belowground components, 5 root systems were excavated in both the YS and OS stands due to logistical difficulties associated with large root systems, while 14 root systems were excavated in the dense stand for a total of 24 root systems. Tree Level Biomass Estimations Biomass of individual trees was measured for above and belowground components in each stand as a part of a separate study developing lodgepole pine allometrics for the Greater Yellowstone Ecosystem (Arcano and Tinker Manuscript in preparation). Biomass of the following tree components was determined: total coarse root (>10mm diameter), total root crown, total lateral root, total aboveground, tree bole, branches, foliage, and needles. Aboveground Biomass Bole - Each bole was harvested at ground level and all branches were removed. The bole was cut into three to four sections with 1 to 2 discs cut out as subsamples to determine moisture content for dry weight. For each bole, two discs were always removed for subsampling at DBH (diameter at breast height, 1.37m), and at 90% of crown base, (the location on the bole where most of the live crown began). Two discs were cut from each section if the bole forked near the 47 top or if DBH and crown base were one meter or closer to each other. Each of the bole sections were weighed separately using a digital hanging scale (Salter-Brecknell) and the disc was summed back into the weight for its respective section. Discs were then dried in the lab to determine moisture loss where the dry: wet weight ratio for each disc was used to determine the dry weight of its respective bole section. All subsamples were dried to a constant weight at 70C. Fine fuels - The fine fuels component was considered to be all needles, associated cones, and associated twigs 6.4 mm diameter, because those aboveground components are likely to be combusted in a fire, and post-fire estimates of branch biomass can then be obtained independent of combustible materials. The fine fuels component was maintained in separate piles of lower, middle, and upper crown portions for each tree, since foliage moisture contents were likely to vary with crown height (Brown 1978). A random subsample approximating the size of 4.0 L in volume was removed from each crown section to determine moisture content. The fine fuels subsamples were then weighed to obtain wet weight. If immediate weighing was not feasible, subsamples were stored in a cooler for no more than 5 days in plastic bags, to prevent moisture loss prior to weighing. The current year’s growth (2004) was removed and discarded, because many of the needles were not yet fully expanded, and samples were collected at varying times throughout the summer of 2004. The subsample was reweighed to determine the proportion of the foliage component that was from the current year’s growth. This proportion was then subtracted from the entire foliage component. After weighing, a random sample of 10 fascicles was collected from each fine fuels subsample, and the length and width of each fascicle was measured to determine leaf area. Leaf area index (LAI) could then be determined from the product of leaf 48 area determination for individual trees and stand density (see below). A tapered-bisected cylinder was used to determine the surface area represented by each fascicle (Pearson et al. 1984; Madgwick 1964). This sample was then dried and subsequently weighed so that the surface area of this subsample could be extrapolated to the entire tree’s needle biomass. This sample was also weighed while still wet for subsequent drying and weighing to determine moisture content for dry weight determination of total needle biomass. The dry weight proportion of total foliage biomass that was needles was determined from separating needles from twigs after the foliage subsamples were dried to a constant weight at 70C. Branches - Branches were cut flush to the tree bole and were separated from foliage at 6.4 mm in diameter. Thus, branches consisted of all shoots excluding the tree bole that were greater than 6.4 mm in diameter. A subsample of approximately 4.0 L in volume was taken to determine moisture content for dry weight for the branches. All subsamples were dried to a constant weight at 70C in the lab. Branches were compiled rather than separated into three crown sections, because one subsample from all sections was deemed sufficient to provide an adequate estimate of dry weight for the component. Belowground Biomass For each tree, the entire coarse root system (>10mm diameter) was excavated with the aid of a backhoe or come-a-long. Prior to excavation, smaller roots that could be damaged by burly excavation techniques were removed by hand. After excavation, the root system was separated into four size classes: root crown (i.e. the massive structure directly beneath the tree bole and within the lateral extent of the diameter of the tree at stump height); lateral roots >50mm in diameter; lateral roots from 25-50mm in diameter; and lateral roots from 10-25mm in diameter. Each size class was then weighed wet, before any moisture was lost. Subsamples of 49 approximately 4.0 L in volume were collected to determine the moisture content for dry weight of each size class, and were posthumously dried to a constant weight at 70°C where the dry: wet ratio was applied to its corresponding size class for determination of total dry weight for each size class. Dry weights for each size class were then summed. Stand Level Biomass Estimations Allometric models derived from three stands varying in densities and ages were used to determine biomass allocation patterns for above and belowground components of lodgepole pine in the three study sites. From variable radius point sampling (Avery and Burkhart 1994), tree diameters were measured at six points within each stand. Basal area could then be calculated. In order to estimate total height and height to the base of the live crown in each stand, regression equations between DBH and total height and between total height and crown base from a related study (Arcano and Tinker Manuscript in preparation) were used. However, if the relationship from those regressions was not significant (p>0.05), the mean of height or height to crown base for each of three diameter classes was applied to all trees in their respective diameter classes. These measured and estimated parameters were used in the application of allometric equations for various tree components from a related study in the GYE, (Arcano and Tinker Manuscript in preparation) and were then applied to the trees measured in variable radius point sampling (Avery and Burkhart 1994). In variable radius point sampling, stand density can be determined, where each “in” tree using an angle gauge represents a certain amount of basal area per unit area, depending on the size of the angle gauge that is being used. “In” trees are trees that are fully within the angle gauge when sighting through the gauge at the tree. The size of the angle gauge is referred to as its basal area factor (BAF). The equation for determining the density represented by each “in” tree is as follows: 50 Di = BAF/BAi (1) where Di = the density represented by the given tree, BAF = the basal area factor for the angle gauge in square feet per acre, and BAi = the basal area in for the given tree. For instance, if a tree is 7.1cm in diameter or 0.003959 m2 in basal area, and the angle gauge being used had a BAF of 2.296 m2/ha, then that tree would be represented by approximately 579 trees/ha. Next, if there were 10 trees of exactly the same size at a point, that point would have a density of 5,790 trees/ha. Next, the values for all points were averaged to determine a stand density. In addition, the appropriate allometric equation from a related study (Arcano and Tinker Manuscript in preparation) was applied to each tree in each stand to determine a biomass value for the respective component of each tree. Each biomass value was then multiplied by the density represented by each tree, which became the stand level biomass represented by that tree. For instance, a 7.1cm tree in the old, sparse stand was represented by 3.14 Mg/ha of total aboveground biomass. If there were 10 trees at that point of the same size, then the stand level total aboveground biomass would be 31.4 Mg/ha. The values for all 6 points in each stand were averaged to determine the average stand level biomass for all components. There were two sources of error when estimating stand level biomass: (1) the error associated with using regression equations for biomass determination of the trees measured in variable point sampling; and (2) the plot level error associated with the variable radius point sampling method itself. However, the accumulated error from error (1) is actually included in error (2). Therefore the perceived problem of error accumulation does not actually exist (Giardina and Ryan 2002), and comparisons of stand biomass between densities and ages remain valid (see below). Carbon Storage 51 Approximately 48% of the dry weight of a lodgepole pine is comprised of carbon (C) (Koch 1996); therefore, 48% of each biomass component was its carbon storage. The carbon storage for our study was compared against other lodgepole pine studies and other forested biomes to determine whether and how carbon storage for our study related to other studies. Statistical Analyses To determine whether allocation patterns at the stand and tree level differed between density and age, one-way ANOVAs were conducted, followed by Tukey’s HSD post-hoc analyses. The following variables were used for comparisons: total coarse root biomass: total aboveground biomass; root crown: lateral root biomass; and percent of total biomass for bole, branches, and foliage. (Note: The total coarse root biomass: total aboveground biomass ratio will be referred to in the following text as the below: aboveground biomass ratio.) Nitrogen Use Efficiency The NUE of a mature forest can be defined as the grams of biomass accumulated per gram of nutrient taken up (Vitousek 1982; Aber and Melillo 2001). Furthermore, the actual nutrient requirement for a tissue is not its concentration while alive, but its concentration upon senescence (Vitousek 1982; Aber and Melillo 2001). Therefore, litterfall and wood increment are ideal candidates for determining NUE. In a mature forest, however, litterfall is more easily measured due to the slow accumulation of wood increment in a mature forest. When measuring NUE with litterfall, NUE is dependent on the ability of the needles to translocate their nutrients back into living tissues prior to senescence; therefore, the higher the ratio of carbon: nitrogen, the higher the NUE. Although measuring NUE in litterfall provides a good estimate of NUE, measuring the entire N taken up for lodgepole pine would provide a more comprehensive estimate of NUE. 52 Eight litter trays were placed at fixed equally spaced intervals within a 50m x 50m plot on the first week of July 2004 in the three sites differing in densities and ages where lodgepole pine allometrics were developed. Litterfall was then collected on August 25th and placed in plastic bags for storage and transportation to the laboratory. It was then sorted by species and P.contorta needles were isolated. These needles were then dried to a constant weight at 70C. We then analyzed for total carbon (C) and nitrogen (N) through a vario Macro C:H:N analyzer (Elementar Americas, Inc.). NUE can be defined as units of biomass or C fixed per unit of N uptaken in litterfall (Vitousek 1982), therefore, the total C:N ratio in litterfall was considered to be the NUE. A one-way ANOVA followed by Tukey’s HSD post-hoc analysis was chosen to determine whether NUE differed among densities, ages, and site productivities. In order to determine relationships between site productivity and NUE, site index (SI) was calculated for each stand using SI curves for lodgepole pine in the Rocky Mountain, Intermountain, and Pacific Northwest regions of the US (Alexander 1966; Koch 1996). SI is only one index of site productivity, and we recognize that there are other indicators of site productivity, such as soil organic matter, which greatly affects nutrient availability (Stevenson and Cole 1999). However, SI has been shown to be a better index of site productivity than any other easily obtained parameter, because tree height is less influenced by stand density than other indices, where stands of higher productivity have the ability to produce taller trees than stands of lower productivity (Barnes et al. 1980). In addition, correlation analyses were conducted between NUE and Site Index, below: aboveground biomass, root crown: lateral root biomass, and allocation to the bole, branches, foliage, and needles. 53 RESULTS Individual Tree Level Allocation Patterns Allocation to bole biomass was highest for all three stands, but there was considerable variability for allocation to other components. The hierarchy of biomass allocation for tree level biomass allocation patterns was as follows: for the OS and YD stands it was bole > foliage > total coarse root biomass > branches, and for the YS stand it was bole > foliage > branches > total coarse root biomass (Table 2, Figure 2). Aboveground biomass was lowest for the YD and highest for the OS stand (Table 2). A striking result of the ANOVA for biomass allocation within trees was that the most structurally dissimilar stands (OS and YD) had the most similar biomass allocation patterns (p > 0.05, Table 2, Figure 2), except for the below: aboveground biomass ratio, where the OS and YD stands were significantly different from one another (Figure 2). In contrast, neither the OS and YS stands, nor the YS and YD stands were significantly different from one another (Table 3). Although differences were not significant according to the one-way ANOVA (p > 0.05) (Table 3), the root crown: lateral root ratio was lowest for the YS stand and highest for the YD stand (Table 2). The tree bole as a proportion of total tree biomass was highest for the YD stand and lowest for the YS stand (Table 2, Figure 2) while in contrast, branches, foliage, and needles as a proportion of total tree biomass were highest in the YS stand. Stand Level Allocation Patterns The hierarchy of stand level biomass allocation for lodgepole pine in the GYE was similar to the pattern observed for the individual tree level, and was as follows: for both the OS stand and YD stands it was bole > foliage > total coarse root biomass > branches; and for the YS stand it was bole > foliage > branches > total coarse root biomass (Table 4, Figure 3). This indicated that bole biomass was the most dominant component of total tree biomass in all three 54 stands. Similar to allocation patterns for individual trees, stand level biomass allocation was most similar for the most structurally dissimilar stands (OS and YD) (Table 4, Figure 3). The results of the one-way ANOVA revealed that stand density and age did not affect how stands allocated their biomass with respect to below: aboveground biomass (p = 0.07) (Table 3), although not statistically different, the two young stands did have slightly higher below: aboveground biomass ratios (0.12) than the OS stand (0.10) (Table 4). For the ratio of root crown: lateral root biomass, post-hoc analyses (Table 3) showed that the ratio of root crown: lateral root biomass was more affected by stand age than by stand density; there was no difference between the YS and YD stand (p = 0.719), but there was a significant difference when comparing the OS stand to both the YS (p = 0.010), and YD (p = 0.001) stands (Table 3). In addition, the root crown: lateral root ratio for each stand revealed a similar pattern, where it was 2.2 for the OS stand and 1.4 and 1.5 for the YS and YD stands respectively (Table 4), indicating a stronger effect of stand age than density on belowground allocation. The percentage of total biomass allocated to the tree bole was strongly affected by stand density and age, and all allocation comparisons between stands were significantly different (p < 0.05) (Figure 3). Allocation to the tree bole differed for all three stands, and it was higher for the OS and YD stands than the YS stand (Table 4, Figure 3). In contrast, allocation to branches, foliage, and needles was highest in the YS stand, and lower in the OS and YD stands (Table 4, Figure 3). Leaf Area Index (LAI) was approximately twice as high in the sparse stands compared to the YD stand, but was quite similar between the OS and YS stands (Table 4). 55 Comparing biomass allocation patterns across geographic locations independent of stand density and age When comparing stands from this study to stands in other studies that had similar densities and ages, the proportion of total biomass allocated to the tree bole was greater in this study when compared to Pearson et al.’s (1984) older stand, but was smaller for the younger stands when compared to both Pearson et al. and Comeau and Kimmins (Table 5, Figure 4). The opposite pattern was shown for branches, although there are slight differences in the definition of branches among studies, because we excluded twigs < ¼” from our branches component (see Methods) (Table 5, Figure 4). The percentage of total biomass allocated to branches and needles was greater for our study than studies developed in southeastern WY and British Columbia for all stands except the OS stand (Table 5, Figure 4). Carbon Storage in the Greater Yellowstone Ecosystem compared to other forests The patterns found for biomass allocation were identical for carbon storage (Table 5), because biomass of lodgepole pine is approximately 48% C (Koch 1996). Carbon storage for total aboveground tree biomass was 70.0 Mg ha-1 for the OS stand, 40.8 Mg ha-1 for the YS stand, and 46.7 Mg ha-1 for the YD stand. For a 110-year-old lodgepole pine forest in another study from the GYE, total aboveground tree carbon storage was 86.9 Mg ha-1 (Litton et al. 2004). For other forest types total aboveground tree biomass was 107.9 Mg ha-1 for a 250-year-old Pinus ponderosa forest in central Oregon by (Law et al. 2001), 93.4 Mg ha-1 for a 50-120-yearold temperate deciduous forest in eastern Tennessee (Curtis et al. 2002), 120.9-153.6 Mg ha-1 for tropical forests of the southwestern Brazilian Amazon (Cummings et al. 2002), and 49.9 Mg ha-1 and 39.3 Mg ha-1 for 139-year-old and 66-year-old Pinus sylvestris forests respectively in boreal Euro-Siberia (Schulze et al. 1999). 56 Nitrogen Use Efficiency and its relation to Stand Productivity and Biomass Allocation Patterns across stand densities and ages NUE, estimated as the C:N ratio of needle litterfall (see Methods), was more affected by stand age than by stand density (Figure 5). The C:N ratio in litterfall / NUE of the YS (104:1) and YD (100:1) stands were remarkably similar (p = 0.826)(Figure 5), but significantly lower than the OS stand (130:1; p < 0.05). Although correlations between NUE and site index, and between site index and biomass allocation patterns were not statistically significant (p < 0.05), several patterns emerged. NUE was inversely correlated to site index (SI) (r = -0.94), the below: aboveground biomass ratio (r = -0.99), and allocation to branches (r = -0.66), foliage (r = -0.75), and needles (r = -0.80), but was positively correlated with the root crown: lateral root biomass (r = 0.97) and allocation to the tree bole (r = 0.71) (Table 6). Correlations were strongest for the below: aboveground biomass ratio, where the correlation was nearly significant (p = 0.08), followed by root crown: lateral root biomass (p = 0.16) and site index (p = 0.23) (Table 6). DISCUSSION The results indicated (1) biomass allocation patterns differed between stand densities and ages; (2) biomass allocation patterns were different at the individual tree level than for the stand level; (3) however, allocation to coarse roots was relatively constant across stand densities and ages; (4) biomass allocation patterns were different in the GYE compared to southeastern Wyoming and British Columbia; (5) carbon storage of mature lodgepole in the GYE was comparable to, but at the lower end of carbon storage published in other forest types and biomes; (6) NUE was higher in the old stand, which was likely more nutrient limited (Binkley et al. 1995); and (7) NUE was likely related to site productivity and biomass allocation patterns. 57 Aboveground Biomass Within the aboveground portion of individual lodgepole pine and lodgepole pine stands, biomass of branches, needles, and foliage were highest in the YS stand (Table 2,4, Figure 2,3). At the stand level, branches, needles, and foliage as a proportion of total tree biomass was lowest in the OS stand, indicating that there was ample room for development of a large tree crown in the YS stand, in contrast to the YD and OS stands where there was more competition. The OS stand had more crown competition than the YD stand, because tree spacing was more homogeneous in the OS stand; in contrast, the YS stand had heterogeneous tree spacing resulting in an assemblage of open grown and clumped trees (Table 1). In addition, the OS stand may have had the lowest branch, foliage, and needle biomass due to greater self-pruning of the lower branches in the older stand, a common characteristic of lodgepole pine (Koch 1996). Although not statistically significant, the YD stand had the lowest branch, needle, and foliage allocation, indicating that greater self-pruning may occur with increasing stand density, and also indicating that the impact of self-pruning on biomass allocation is controlled by density in addition to age. Similarly, Pearson et al. (1984) found that a sparser stand (2217 trees/ha) had a greater amount of total biomass allocated to branches (8.3%) than for a denser stand on the same site (14,640 trees/ha), where only 5.2% of total tree biomass was allocated to branches. Ryan et al. (1997) suggested that young trees allocate more resources to foliage rather than to woody components, and this trend becomes less pronounced with crown closure. This is supported by the results of this study, as the open-grown YS stand had 26.5% of total biomass allocated to foliage, while the OS stand with greater observed crown competition had 10.2% of total biomass allocated to foliage (Table 2, Figure 2). Pearson et al. (1984) found that allocation to foliage was greater in a young, sparse stand (75 years old), where allocation to foliage was 58 10.0% of total tree biomass than in an older stand (110 years old), where foliage was only 6.2% of total tree biomass. Allocation to tree boles was lowest in the YS stand at the individual tree and stand levels, while bole allocation to the OS and YD stands were surprisingly comparable to each other. The low proportion of total tree biomass for boles in the YS stand is likely attributed to a high proportion of branch and foliage biomass created by a combination of its young age and opengrown stand structure. The similarity in allocation to tree boles between the OS and YD stands may be related to their competitive similarity. Although not quantified, our observations suggested both of these stands possessed extensive crown competition, while the YS stand had less crown competition. Increased allocation to boles in dense and older stands with greater crown competition was consistent with the results of Nilsson and Albrekston (1993) and Kaufmann and Ryan (1986). They found that allocation to the tree bole compared to foliage was greater for competitively suppressed trees. Future studies should include more direct measurements of competition among trees, such as nearest neighbor analyses. Leaf Area Index LAI was much greater in the sparse stands than in the YD stand, suggesting that tree canopies were much more suppressed in that stand, which is contrary to the results of Pearson et al. (1984). They found that a dense stand with 14,640 trees/ha had only a slightly lower LAI (7.1 m2/ha) than a less dense stand with 2,217 trees/ha (7.3 m2/ha). Overall, my estimates of LAI were much lower than estimates reported by Pearson et al. (1984), which could be attributed to lower stand basal areas for this study (Table 1). Basal areas for this study ranged from 16.84 – 28.32 m2/ha, but Pearson et al. (1984) reported basal areas from 26 – 64 m2/ha. Although sapwood area has been shown to be a better predictor of foliage mass and leaf area than basal 59 area by Pearson et al. (1984) and by Grier and Waring (1974), basal area has also been shown to predict foliage mass and leaf area well (Pearson et al. 1984). Therefore, leaf area index should also be affected by basal area. Belowground Biomass For allocation to belowground biomass components at the stand level, the root crown: lateral root biomass ratio was found to be significantly less for younger stands than for the single old stand, indicating that the root crown becomes a more dominant part of root systems as stand age increases, suggesting a greater need for structural support as trees become older and larger. Similarly, the root crown: lateral root biomass ratio for individual trees revealed that allocation to root crowns was lowest for the YS stand, but similarly higher for the two stands differing most in density and age (OS and YD). The YS stand was the most open-grown of the three stands and likely had the least amount of competition among trees, indicating that the root crown becomes a greater portion of the root system as belowground competition decreases. Interestingly, greater differences in stand age for the root crown: lateral root biomass ratio were evident when comparing sites at the stand level, while greater differences in stand density were revealed when examining the data at the individual tree level, indicating the importance of sampling at multiple scales for understanding biomass allocation patterns of lodgepole pine. A possible explanation for variability in allocation at differing scales is that individual tree allocation patterns are likely a function of individual tree age and tree spacing around individual trees, while in contrast, stand level allocation patterns are likely a function of the stand age and density. Below: Aboveground Biomass Ratio At the stand level, stand density and age had surprisingly little effect on the shifting of resources to above and belowground biomass, in contrast to a study by Pearson et al. (1984) for 60 lodgepole pine in southeastern Wyoming. Differences in below: aboveground biomass partitioning among densities ranging from 420 to 14640 trees ha-1 found by Pearson et al. may be at least partly attributed to the inclusion of roots smaller than 10mm diameter in their study. For this study, below: aboveground biomass was found to be constant across stand ages, which is comparable to estimates of total root biomass including fine roots across stand ages for other studies (Litton et al. 2004; Pearson et al. 1984). Despite the constant relative biomass partitioning of resources above and belowground at the stand level, below: aboveground biomass was considerably more variable at the scale of individual trees (Table 2, 4), because below: aboveground allocation was lowest in the YD stand, the site that likely had the highest belowground allocation, indicating that both stand density and age had an important effect on allocation to coarse root biomass. Although there are some similarities in the ratio of below: aboveground biomass among sites, especially at the stand level, biomass partitioning within above and belowground components was found to be highly variable with stand density and age. Biomass Allocation Comparisons Among Geographic Locations Although fine roots (<10mm diameter) were not considered in this study, direct comparisons for aboveground biomass components in different geographic regions could be made. The proportion of aboveground biomass allocated to the tree bole was significantly less for this study except for the comparison between older stands, and total bole biomass was less in this study for all stands (Figure 3). This suggests that allocation to the tree bole is in a large part driven by site productivity, because lodgepole pine forests in the GYE are less productive than lodgepole pine forests in southeastern Wyoming and in British Columbia due to a combination of poorer soils developed on volcanic rhyolite, more extreme climatic conditions, and differences in topography. In addition, site indices for this study ranged from 14.3 – 16.0m at 100 years, and 61 site indices for Comeau and Kimmins (1989) ranged from 14.3 – 20.5 m at 100 years. Furthermore, estimates of total root biomass were much lower for this study, although fine root biomass was excluded from this study (Figure 3), suggesting that total coarse root biomass is not driven by site productivity. Bole biomass allocation was found to be more constant across geographic locations for comparisons between older stands (Figure 3a), suggesting that during its lifespan, lodgepole pine may gradually compensate for lower site productivities through increases in resource use efficiency in order to attain relatively constant wood production despite differences in site productivity (Binkley et al. 2004; Aber and Melillo 2001; Vitousek 1982). Interestingly, allocation to branches and needles was higher in the GYE than in British Columbia and southeastern Wyoming, suggesting that lodgepole pine in the GYE are compensating for poor site conditions by allocating more biomass to photosynthetic tissues and to branches for support of photosynthetic tissues, although nitrogen and other soil nutrients would be needed to produce these tissues. Therefore, lodgepole pine in the GYE may have greater nutrient use efficiency than lodgepole pine in British Columbia and southeastern Wyoming, although there are no data to support this, and future research is needed on the variability of nutrient use efficiency of lodgepole pine across a gradient of site productivities in the intermountain west. Carbon Storage in the Greater Yellowstone Ecosystem compared to other forests Carbon storage for our study was comparable to another study in the GYE, and it was also comparable to carbon storage for other forest types and biomes. However, carbon storage in mature lodgepole pine trees was at the lower end of values reported for other forest types and biomes, and the results suggest that only boreal forests have lower carbon stored in aboveground tree biomass than subalpine lodgepole pine forests (Schulze et al. 1999). 62 Nitrogen Use Efficiency and its relation to Stand Productivity and Biomass Allocation Patterns across stand densities and ages NUE was significantly higher in the old, sparse stand than in the two younger stands and was more affected by stand age than by stand density, suggesting that older forests are more nutrient limited than younger forests as found by Ryan et al. (1997) and (Binkley et al. 1995). This also suggested that there was little difference in site productivity among densities. This is supported by the result that SI was lowest for the OS stand where NUE was highest, indicating that NUE was highest where site productivity was lowest, because SI is a better index of site productivity than any other easily obtained index (Barnes et al. 1980). This is further supported by the research of (Binkley et al. 1995) in southeastern Wyoming, where their results from a fertilization experiment suggested nitrogen to be more limiting in older forests. An inverse relationship between NUE and site productivity is consistent with the findings of Vitousek (1982) and Miller et al. (1979), but has recently been disputed by Binkley et al. (2004), where they suggested that for NUE to be equivalent to the grams of carbon fixed per unit of nutrient in litterfall, there would have to be a constant relationship between litter mass and ANPP. Unfortunately, the time span of this study did not allow for determination of litter mass or ANPP, and this should be pursued in the future. NUE was inversely related to the ratio of below: aboveground biomass, because allocation to coarse roots was greater on more productive sites, which had lower NUE. These results suggested that allocation to coarse roots >10mm in diameter were more similar to other woody components such as the tree bole and branches than to fine roots, whereby coarse roots are present mostly for structural support and are dependent, like the tree bole, on the ability of fine roots to absorb water and nutrients from the soil. 63 NUE was positively related to the ratio of root crown: lateral root biomass, indicating that the root crown is more similar in function to the tree bole than to lateral roots, because the root crown: lateral root biomass ratio and allocation to the tree bole are similarly correlated with NUE. If the opposite were true, the correlation between NUE and the root crown: lateral root ratio would be negative. NUE is higher where allocation to the tree bole is higher, suggesting that NUE is increasing the ability of lodgepole pine to produce woody biomass. In contrast, NUE was lower where allocation to branch, foliage, and needle biomass was higher. This indicated that increasing NUE may not be necessary to increase production of woody biomass, where there was more biomass allocated to photosynthetic tissues and branches tissues in support of those tissues. Future studies should include estimates of photosynthetic efficiency to test this hypothesis. SUMMARY AND CONCLUSIONS The allocation of resources to above and belowground lodgepole pine biomass was relatively similar between stand densities and ages as shown by the relatively constant ratios of total coarse root: total aboveground biomass. Therefore, allocation to coarse root biomass is relatively constant across forest structures. However, there was considerable variability in aboveground allocation and belowground allocation when considered independently, because the ratio of root crown: lateral root biomass differed significantly with stand density and age, and allocation to the tree bole, branches, fine fuels, and needles differed with stand density and age. In addition above and belowground allocation patterns were affected differently and the individual tree and stand levels. 64 There were considerable differences in the biomass allocation patterns of lodgepole pine among geographic locations. Despite some similarities, generalizing patterns of biomass allocation across forest structures and among geographic locations should proceed with caution, and further research into the biomass allocation patterns of lodgepole pine in the Greater Yellowstone Ecosystem and elsewhere in the Intermountain West is needed. However, if further research indicates that the ratio of coarse root: total aboveground biomass for lodgepole pine stands is approximately 0.10 to 0.12 as identified by this study, determination of belowground lodgepole pine biomass could become considerably easier, because only aboveground measurements would be needed and the appropriate ratio could be applied to determine coarse root biomass. Additionally, NUE appears to be related to site productivity and biomass allocation patterns, although further study of these relationships is needed, and additional study is needed to determine the relationships of NUE across the intermountain west. 65 Table 1. Sites for the determination of biomass allocation patterns of P. contorta in the Greater Yellowstone Ecosystem. Soils for all three sites were in the Koffgo series (see Methods). NAD 83, UTM Stand Density Zone 12 Stand (trees > 5cm Northing Easting Site Elevation Age DBH per Stand basal area (m) (m) Name (m) (years) hectare) (m2 per hectare) Grassy Lake 2249 4886015 511735 165 674 175 16.84 14.3 Coffee Pot 1951 4926541 472232 64 725 72 19.71 11.4 US 20 1951 4925932 472657 64 2452 929 28.32 30.4 Table 2. Mean individual tree level biomass, total coarse root: total aboveground biomass and root crown: lateral root biomass ratios of three Pinus contorta stands in the GYE. Mass is in Mg/ha and percent of total tree biomass for each component is in brackets. STANDS Component Young, Old, Sparse Sparse Young, Dense Biomass Mass (Kg) [%] Mass (Kg) [%] Mass (Kg) [%] 187.9 [86.3] 128.9 [87.6] 41.0 [90.1] 141.2 [64.8] 59.7 [40.6] 31.2 [68.7] Branches 14.9 [6.9] 24.3 [16.5] 2.7 [5.9] Fine Fuels 31.8 [14.6] 44.9 [30.6] 7.1 [15.6] Needles 25.8 [11.8] 27.0 [18.4] 4.6 [10.1] 30.0 [13.8] 18.2 [12.4] 4.5 [9.9] Root Crown 19.0 [8.8] 10.4 [7.1] 2.6 [5.7] Lateral Roots (>10mm) 11.0 [5.0] 7.8 [5.3] 1.9 [4.2] Total Tree Biomass 217.9 147.1 45.5 Total Aboveground Bole Coarse Roots (>10mm) - Ratios Total Coarse Root: Total Aboveground 0.16 0.14 0.11 Root Crown: Lateral Root 1.8 1.4 2.2 Table 3. Tukey’s post-hoc analysis for the total coarse root: total aboveground and root crown: lateral root biomass ratios at the stand and individual tree levels for three P. contorta stands in the Greater Yellowstone Ecosystem. Asterisks (*) signify that the one way ANOVA was not significant (p>0.05) and therefore, no post-hoc analyses were necessary. P-values in bold are significant at an of 0.05. - Tree Level Site Comparisons p-values total coarse root: total Root crown: lateral (1) (2) aboveground root Old, Sparse Young, Sparse 0.234 * Old, Sparse Young, Dense * 0.004 Young, Sparse Young, Dense Site Comparisons (1) Old, Sparse (2) Young, Sparse Old, Sparse Young, Sparse 0.282 * - Stand Level p-values total coarse root: total Root crown: lateral aboveground root * 0.010 Young, Dense * 0.001 Young, Dense * 0.719 Table 4. Stand level biomass, total coarse root: total aboveground biomass ratio; root crown: lateral root biomass ratio, and leaf area index of three Pinus contorta stands in the GYE. Mass is in Mg/ha and percent of total tree biomass for each component is in brackets. Ratios were based on values derived from individual models for the components involved in the ratios. Percentages for bole, branches, foliage, needles, root crown, and lateral roots were calculated from the sum of the bole, branches, foliage, and total roots. Percentages for total aboveground and total root biomass were calculated from total tree biomass as shown below. All components except for total tree biomass were derived from individual allometric models by Arcano and Tinker (Manuscript In Preparation); therefore discrepancies exist between total aboveground biomass and its components, as well as between total root biomass and its components. STANDS Old, Young, Young, Sparse Sparse Dense Component Mass (Mg/ha) [%] Mass (Mg/ha) [%] Mass (Mg/ha) [%] Total Aboveground 127.0 [91.3] 84.9 [89.0] 97.3 [89.5] 91.6 [75.1] 42.2 [47.3] 70.7 [64.3] Branches 5.9 [4.8] 12.9 [14.4] 8.7 [7.9] Fine Fuels 12.4 [10.2] 23.7 [26.5] 19.0 [17.3] 8.3 [6.8] 15.4 [17.2] 11.5 [10.5] 12.1 [8.7] 10.5 [11.0] 11.5 [10.5] Root Crown 8.2 [6.7] 6.6 [7.4] 7.1 [6.5] Lateral Roots (>10mm) 3.7 [3.0] 4.5 [5.0] 4.8 [4.4] Total Tree Biomass 139.1 95.4 108.8 Total Coarse Root: Total Aboveground 0.10 0.12 0.12 Root Crown: Lateral Root 2.2 1.4 1.5 2.5 - Leaf Area Index 2.1 1.2 Bole Needles Coarse Roots (>10mm) Table 5. Carbon (C) allocation in three stands differing in density and age in the Greater Yellowstone Ecosystem. Old, Sparse STANDS Young, Sparse Young, Dense C (Mg/ha) C (Mg/ha) C (Mg/ha) Total Aboveground 70.0 40.8 46.7 Bole 44.0 20.3 33.9 Branches 2.8 6.2 4.2 Fine Fuels 6.0 11.4 9.1 Needles 4.0 7.4 5.5 Coarse Roots (>10mm) 5.8 5.0 5.5 Root Crown 4.0 3.2 3.4 Lateral Roots (>10mm) 1.8 2.2 2.3 Total Tree Biomass 66.8 45.8 52.2 Biomass Component 70 Table 5: Comparison of biomass and percent of total biomass for three Pinus contorta stands in the GYE to stands in British Columbia by Comeau and Kimmins (1989) and in southeast Wyoming by Pearson et al. (1989). Percentages are in brackets. Comparisons Study This Study Biomass (Mg/ha) [%] Age Density Total Aboveground 64 2452 97.3 [89.5] 70.7 [64.3] 8.7 [8.0] 11.5 [8.4] Bole Branches Needles Comeau and Kimmins This Study 70 1900 119.4 [76.5] 107.4 [68.8] 7.1 [4.6] 4.9 [3.1] 165 674 127.0 [91.3] 91.6 [75.1] 5.9 [4.8] 8.3 [5.4] Pearson et al. 240 420 131.9 [77.5] 110.8 [65.1] 14.2 [8.3] 6.9 [4.1] This Study 64 725 84.9 [89.0] 42.2 [47.3] 12.9 [14.5] 15.4 [11.3] Pearson et al. 75 1280 96.3 [78.4] 74.4 [60.6] 9.6 [7.8] 12.3 [10.0] 71 Table 6. Correlation coefficients (r) and p-values between NUE and site index (SI), biomass allocation ratios, and proportions of total lodgepole pine biomass for three stands differing in densities and ages in the Greater Yellowstone Ecosystem Total Coarse Root: Total Root crown: Fine SI aboveground lateral root Bole Branches Fuels Needles r -0.94 -0.99 0.97 0.71 -0.66 -0.75 -0.80 p 0.230 0.082 0.155 0.497 0.593 0.463 0.409 Figure 1: Three Stands Differing in Densities and Ages for the determination of biomass allocation patterns of P. contorta in the Greater Yellowstone Ecosystem. Individual stands are depicted by a black square. 73 Figure 1: YS YD OS Figure 2: Biomass allocation patterns at the individual tree level for three P. contorta stands differing in densities and ages in the Greater Yellowstone Ecosystem. Different letters denote significance according to Tukey’s HSD post-hoc analyses. Figure 2: Proportion of Total Tree Biomass Tree Level 1.0 Old, Sparse Young, Sparse Young, Dense 0.8 0.6 0.4 0.2 0.0 B ee Tr s ole Br c an he ls ue F e Fin Component 76 s le ed e N Figure 3: Biomass allocation patterns at stand level for three P. contorta stands differing in densities and ages in the Greater Yellowstone Ecosystem. Different letters denote significance according to Tukey’s HSD post-hoc analyses. 77 Figure 3: Stand Level Proportion of Total Tree Biomass 1.0 old, sparse young, sparse young, dense 0.8 0.6 0.4 0.2 0.0 s le bo e ch an br Fin u eF els Component 78 n d ee les Figure 4: Biomass allocation patterns at the stand level for P. contorta in the Greater Yellowstone Ecosystem compared to studies in British Columbia and southeastern Wyoming. Comparisons are made using data other studies from stands of comparable density and age to stands from this study. 79 Figure 4: Young, Dense Stand Young, Sparse Stand Old, Sparse Stand 100 % of Total Tree Biomass This Study Pearson et al. This Study Pearson et al. This Study Comeau and Kimmins 80 60 40 20 0 al A Tot und gro e v bo le Bo s es dle nch nee bra v bo T d un ro g e le Bo s he nc a br lA ota les ed e n ou b ta To Component lA r eg ov nd le Bo an br ch es e ne d le s Figure 5: Carbon: Nitrogen ratio as an estimate of nitrogen use efficiency in three stands differing in densities and ages in the Greater Yellowstone Ecosystem. Letters denote significance according to Tukey’s HSD post-hoc analysis. Figure 5: Nitrogen Use Efficiency 160 140 a 120 b C:N ratio b 100 80 60 40 20 0 Old, Sparse Young, Sparse Young, Dense STAND 82 LITERATURE CITED Aber, J. D. and J. M. Melillo (2001). Terrestrial Ecosystems. San Diego, CA, Harcourt Academic Press. Alexander, R. R. 1966. Site indexes for lodgepole pine, with corrections for stand density: instructions for field use. Res. Pap. RM-24. Fort Collins, CO: U.S. Department of Agriculture, Forest Service, Rocky Mountain Forest and Range Experiment Station; 7 p. Arcano, R. and D. B. Tinker Manuscript in preparation. Allometric model development for mature lodgepole pine forests in the Greater Yellowstone Ecosystem. Avery, T. E. and H. E. Burkhart (1994). Forest Measurements. Boston, MA, McGraw-Hill Inc. Barnes, B. V., D. R. Zak, S. R. Denton and S. H. Spurr (1980). Forest Ecology. New York, NY, John Wiley & Sons, Inc. Berger, U., H. Hildenbrandt and V. Grimm 2004. Age-related decline in forest production: modelling the effects of growth limitation, neighbourhood competition and self-thinning. Journal of Ecology 92: 846-853. Binkley, D., F. W. Smith and Y. Son 1995. Nutrient Supply and Declines in Leaf-Area and Production in Lodgepole Pine. Canadian Journal of Forest Research-Revue Canadienne De Recherche Forestiere 25: 621-628. Binkley, D., J. L. Stape and M. G. Ryan 2004. Thinking about efficiency of resource use in forests. Forest Ecology and Management 193: 5-16. Bowerman, T. S., J. Dorr, S. Leahy, K. Varga and J. Warrick (1999). Targhee National Forest ecological unit inventory, United States Department of Agriculture, Forest Service United States Department of Agriculture, Natural Resources Conservation Service University of Idaho, College of Agriculture. 1-2: 787. 83 Brown, J. K. 1978. Weight and density of crowns of Rocky Mountain conifers. Res. Pap. Int197. Ogden, UT: U.S. Department of Agriculture, Forest Service, Intermountain Research Station; 56 p. Chapin, S. F. I., P. Matson and H. A. Mooney (2002). Principles of terrestrial ecosystem ecology. New York, NY, Springer-Verlag New York, Inc. Cote, B., J. W. Fyles and H. Djalilvand 2002. Increasing N and P resorption efficiency and proficiency in northern deciduous hardwoods with decreasing foliar N and P concentrations. Annals of Forest Science 59: 275-281. Cummings, D. L., J. B. Kauffman, D. A. Perry and R. F. Hughes 2002. Aboveground biomass and structure of rainforests in the southwestern Brazilian Amazon. Forest Ecology and Management 163: 293-307. Curtis, P. S., P. J. Hanson, P. Bolstad, C. Barford, J. C. Randolph, H. P. Schmid and K. B. Wilson 2002. Biometric and eddy-covariance based estimates of annual carbon storage in five eastern North American deciduous forests. Agricultural and Forest Meteorology 113: 3-19. Despain, D. G. (1990). Yellowstone vegetation: consequences and environment in a natural setting. Boulder, CO, Roberts Rinehart, Inc. Dirks, R. A. and B. E. Martner 1982. The climate of Yellowstone and Grand Teton National Parks. National Park Service Occassional Paper 6. Fahey, T. J. 1983. Nutrient dynamics of aboveground detritus in lodgepole pine (Pinus contorta ssp. latifolia) ecosystems, southeastern Wyoming. Ecological Monographs 53: 51-72. Fahey, T. J., J. B. Yavitt, J. A. Pearson and D. H. Knight 1985. The nitrogen cycle in lodgepole pine forest, southeastern Wyoming. Biogeochemistry 1: 257-275. 84 Giardina, C. P. and M. G. Ryan 2002. Total belowground carbon allocation in a fast-growing Eucalyptus plantation estimated using a carbon balance approach. Ecosystems 5: 487499. Gower, S. T., C. C. Grier and K. A. Vogt 1989. Aboveground production and N and P use by Larix occidentalis and Pinus contorta in the Washington Cascades, USA. Tree Physiology 5: 1-11. Grier, C. C. and R. H. Waring 1974. Conifer foliage mass related to sapwood area. Forest Science 20: 205-206. Hendrickson, O. Q. and D. Burgess 1989. Nitrogen-fixing plants in a cut-over lodgepole pine stand of southern British Columbia. Can. J. For. Res. 19: 936-939. Kashian, D. M., M. G. Turner and W. H. Romme 2005. Variability in leaf area and stemwood increment along a 300-year lodgepole pine chronosequence. Ecosystems 8: 48-61. Kashian, D. M., M. G. Turner, W. H. Romme and C. G. Lorimer 2005. Variability and convergence in stand structural development on a fire-dominated subalpine landscape. Ecology 86: 643-654. Kaufmann, M. R. and M. G. Ryan 1986. Physiographic, stand, and environmental effects on individual tree growth and growth efficiency in subalpine forests. Tree Physiology 2: 4759. Keyes, M. R. and C. C. Grier 1981. Above- and below-ground net production in 40-year-old douglas-fir stands on low and high productivity sites. Can. J. For. Res. 11: 599-605. Knight, D. H. (1994). Mountains and plains: the ecology of Wyoming landscapes. New Haven, CT, Yale University Press. Koch, P. (1996). Lodgepole Pine in North America. Madison, WI, Forest Products Society. 85 Law, B. E., P. E. Thornton, J. Irvine, P. M. Anthoni and S. V. Tuyl 2001. Carbon storage and fluxes in ponderosa pine forests at different developmental stages. Global Change Biology 7: 755-777. Litton, C. M., M. G. Ryan and D. H. Knight 2004. Effects of tree density and stand age on carbon allocation patterns in postfire lodgepole pine. Ecological Applications 14: 460475. Litton, C. M., M. G. Ryan, D. H. Knight and P. D. Stahl 2003. Soil-surface carbon dioxide efflux and microbial biomass in relation to tree density 13 years after a stand replacing fire in a lodgepole pine ecosystem. Global Change Biology 9: 680-696. Madgwick, H. A. I. 1964. Estimation of surface area of pine needles with special reference to Pinus resinosa. J. For. 61: 636. Miller, H. G., J. M. Cooper, J. D. Miller and O. J. L. Pauline 1979. Nutrient cycles in pine and their adaptation to poor soils. Can. J. For. Res. 9: 19-26. Nilsson, U. and A. Albrekston 1993. Productivity of needles and allocation of growth in young Scots pine trees of different competitive status. Forest Ecology and Management 62: 173187. Olsson, U., D. Binkley and F. W. Smith 1998. Nitrogen supply, nitrogen use, and production in an age sequence of lodgepole pine. Forest Science 44: 454-457. Pearson, J. A., T. J. Fahey and D. H. Knight 1984. Biomass and leaf area in contrasting lodgepole pine forests. Can. J. For. Res. 14: 259-265. Ryan, M. G., D. Binkley and J. H. Fownes 1997. Age-related decline in forest productivity: pattern and process. Advances in Ecological Research 27: 213-262. 86 Schulze, E. D., J. Lloyd, F. M. Kelliher, C. Wirth, C. Rebmann, B. Luhker, M. Mund, A. Knohl, M. Milyukova, W. Schulze, W. Ziegler, A. B. Varlagin, A. F. Sogachev, R. Valentini, S. Dore, S. Grigoriev, O. Kolle, M. I. Panfyorov, N. Tchebakova and N. N. Vygodskaya 1999. Productivity of forests in the Eurosiberian boreal region and their potential to act as a carbon sink - a synthesis. Global Change Biology 5: 703-722. Smith, F. W. and J. N. Long 2001. Age-related decline in forest growth: an emergent property. Forest Ecology and Management 144: 175-181. Smith, F. W. and S. C. Resh 1999. Age-related changes in production and below-ground carbon allocation in Pinus contorta forests. Forest Science 45: 331-341. Stevenson, F. J. and M. A. Cole (1999). Cycles of Soil: Carbon, Nitrogen, Phosphorous, Sulfur, Micronutrients. New York, NY, John Wiley and Sons, Inc. Stump, L. M. and D. Binckley 1993. Relationships between Litter Quality and Nitrogen Availability in Rocky-Mountain Forests. Canadian Journal of Forest Research-Revue Canadienne De Recherche Forestiere 23: 492-502. Taiz, L. and E. Zeiger (2002). Plant Physiology. Sunderland, MA, Sinauer Associates, Inc. Thomas, K. D. and C. E. Prescott 2000. Nitrogen availability in forest floors of three tree species on the same site: the role of litter quality. Canadian Journal of Forest Research-Revue Canadienne De Recherche Forestiere 30: 1698-1706. Tinker, D. B., W. H. Romme, W. W. Hargrove, R. H. Gardner and M. G. Turner 1994. Landscape-scale heterogeneity in lodgepole pine serotiny. Can. J. For. Res. 24: 897-903. Turner, M. G., W. H. Romme, R. H. Gardner and W. W. Hargrove 1997. Effects of fire size and pattern on early succession in yellowstone national park. Ecological Monographs 67: 411-433. 87 Turner, M. G., D. B. Tinker, W. H. Romme, D. M. Kashian and C. M. Litton 2004. Landscape patterns of sapling density, leaf area, and aboveground net primary production in postfire lodgepole pine forests, Yellowstone National Park (USA). Ecosystems 7: 751-775. Vitousek, P. 1982. Nutrient cycling and nutrient use efficiency. The American Naturalist 119: 553-572. 88 SUMMARY Biomass was estimated in three lodgepole pine forests differing in stand densities and ages in the Greater Yellowstone Ecosystem. Forty-six trees were harvested, and 24 root systems were excavated. The number of trees harvested was relatively equal among stands, but only five root systems were excavated in sparse stands due to logistical constraints, while all 14 of the harvested trees in the dense stand were excavated. Allometric equations for above and belowground tree components were developed to estimate biomass in all three stands. Additional pooled equations were developed to compare allometric relationships between differing stand densities and ages. Actual biomass estimates from this study’s allometric models were compared against estimated biomass values from this study’s equations and equations developed in southeastern Wyoming and British Columbia. Biomass allocation patterns were determined from this study’s actual biomass estimates, and biomass allocation patterns were compared across differing stand densities and ages and among geographic locations independent of stand densities and ages. Nitrogen use efficiency (NUE) was determined from the C:N ratio in litterfall in each of the three stands and was compared across stands with different densities and ages. To determine if NUE was related to site productivity and biomass allocation patterns, correlation analyses were conducted. Variation in stand density and age affected measured tree morphometric parameters that best predicted biomass of lodgepole pine tree components. Allometric models developed for mature lodgepole pine in the Greater Yellowstone Ecosystem were very robust for predicting various tree biomass components, and showed potential for estimating landscape level biomass of lodgepole pine forests, because R2 values were often above 0.90. Allometric equations were found to differ between stand densities and ages according to qualitative observations and 89 statistical tests, and they were also found to be different among geographic locations independent of stand density and age. In addition, applications of inappropriate models sometimes produced tremendous errors and have the potential to do so in future applications. The allocation of resources to above and belowground lodgepole pine biomass was relatively similar between stand densities and ages as shown by the relatively constant ratios of total coarse root: total aboveground biomass, where the total coarse root: total aboveground biomass ratio at the stand level ranged from 0.10 to 0.12. Therefore, allocation to coarse root biomass is relatively constant across forest structures. However, allocation within above and belowground sections of the tree was highly variable across stand densities and ages, and appeared to be greatly affected by stand level and individual tree level competition. Although total aboveground biomass was relatively constant across densities and ages, bole biomass was lowest in the young, sparse stand, but branch, foliage, and needle biomass was highest in the young, sparse stand. Also, biomass allocation patterns were sometimes different at the stand and individual tree levels, indicating the need for additional study at multiple scales. In addition, there were considerable differences in the biomass allocation patterns of lodgepole pine among geographic locations. Allocation to bole biomass was lower in the Greater Yellowstone Ecosystem, but allocation to branch and needle biomass was higher, which is likely a product of the lower site productivity in the GYE than in southeastern Wyoming and British Columbia. In addition, estimates of lodgepole pine biomass fit well within the residuals of global allometric models developed by Enquist et al. (1998). NUE was higher with increasing stand age, indicating higher NUE with increasing nutrient limitation. Additionally, NUE was negatively correlated with site productivity (site index), indicating higher NUE with decreasing site productivity. NUE was also negatively 90 correlated with the ratio of total coarse root: total aboveground biomass and allocation to branch, foliage, and needle biomass. It was determined that NUE is higher when allocation to coarse root, branch, needle, and foliage biomass is lower. However, NUE was positively correlated with the ratio of root crown: lateral root biomass and the tree bole, indicating that higher NUE may increase root crown and bole production. Differences in allometric equations with different forest structures and geographic locations indicates the need for careful application of allometric models, supported by the result that inappropriate model use often produced tremendous errors. Errors for individual trees produced by inappropriate model application have the potential to propagate when extrapolated to the stand and ultimately the landscape level. In addition the allometric models developed in this study showed great promise for a variety of applications including ecosystem carbon studies, fire modeling, and determination of wood biomass by foresters. Biomass allocation patterns differed among forest structures and geographic locations, indicating that understanding these differences is vital to understanding differences in NPP and NEP. In addition, NUE was shown to differ across forest structures, it was related to site productivity and biomass allocation patterns, indicating that increasing our understanding of NUE can further our understanding of how NPP and NEP vary across the landscape. 91 LITERATURE CITED Enquist, B. J., J. H. Brown and G. B. West 1998. Allometric scaling of plant energetics and population density. Nature 395: 163-165. 92