Conservation Genetics Resources Online Resource 1 Development
advertisement

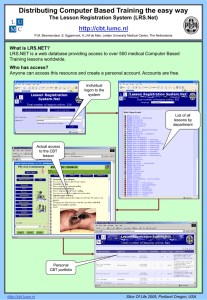
Conservation Genetics Resources Online Resource 1 Development of 20 TaqMan assays differentiating the endangered shortnose and Lost River suckers M. S. Hoy and C. O. Ostberg U.S. Geological Survey, Western Fisheries Research Center, 6505 NE 65th Street, Seattle, WA 98115 Corresponding author: Carl O. Ostberg, costberg@usgs.gov Methods Samples Shortnose sucker (SNS) (Chasmistes brevirostris) and Lost River sucker (LRS) (Deltistes luxatus) fin tissues were collected from adults (2004 – 2006, 2009, and 2011) in Upper Klamath Lake, southern Oregon, and Clear Lake, northern California, and whole young-of-the-year (YOY) LRS and SNS collected from Upper Klamath Lake (2004 – 2011). Adults were identified to species by snout and lip morphology. Young-ofthe-year were identified to species by vertebral counts, with the exception that gill raker counts were incorporated for individuals having 44 vertebrae or less. Young-of-the-year SNS and LRS are primarily differentiated by their number of vertebrae; SNS have 41 – 44 and LRS have 44 – 48 (Markle et al. 2005). Gill raker number is used to assist species identification for YOY having 44 vertebrae, although gill raker number overlaps between LRS and SNS. In addition, SNS and Klamath largescale sucker (Catostomus snyderi) have similar numbers of vertebrae and their differentiation relies on gill rakers and other meristic characters. Species identification was performed by field biologists at U.S. Geological Survey (Western Fisheries Research Center, Klamath Falls Field Station) and Bureau of Reclamation (Klamath Basin Area Office). Herein, we designate the species identifications by field biologists as the a priori assignment. Genomic DNA was extracted from fin tissues using the UltraClean Tissue and Cells DNA Isolation Kit (MO BIO Laboratories). Marker discovery using locus-specific sequencing Two marker discovery approaches were applied to identify species-diagnostic candidate SNPs; locusspecific sequencing and restriction-site-associated DNA (RAD) sequencing (Baird et al. 2008). For locusspecific sequencing, PCR primers were generated from SNS nuclear DNA (nDNA) sequence data available in Genbank and were used to amplify and sequence 15 YOY LRS and SNS each. Sequences were edited and aligned using SEQUENCHER v 4.10.1 (Gene Codes Corporation). One PCR primer set designed from accession number AY351369.1 (AL176; F:5’-TCATAATAACCATCACATCAC-3’; R:5’AGTCCCATTGGCGAGAG-3’) amplified a 435 base pair product that contained a species-diagnostic candidate SNP. PCR chemistry for AL176 amplification consisted of 10 ng genomic DNA, 10x Reaction Buffer (Bulldog Bio), 500 µM dNTP Mix (Bioline), 0.5 µM of each primer, and 1.0 units BioReady rTaq (Bulldog Bio). Cycling conditions consisted of initial denature at 94oC for 5 minutes, followed by 35 cycles at 94oC for 30 seconds, anneal at 52oC for 15 seconds, and extension at 72oC for 25 seconds, followed by a final extension at 72oC for 5 minutes. A restriction enzyme assay was designed for the AL176 candidate SNP using TasI (MBI Fermentas) and was applied to 66 adults from each species. The TasI-digested AL176 amplicon produced two restriction fragment length polymorphism patterns: one common to SNS (21, 26, and 388 base pairs) and a second common to LRS (21, 26, 58, and 330 base pairs). We also amplified and sequenced 771 base pair of the mitochondrial DNA (mtDNA) Cytochrome B (CytB) gene in 15 YOY LRS and SNS each, using primers L14725 and H15573 (Meyer 1993). PCR chemistry consisted of 10 ng genomic DNA, 10x Reaction Buffer (Bulldog Bio), 500 µM dNTP Mix (Bioline), 0.5 µM of each primer, and 1.0 units BioReady rTaq (Bulldog Bio). Cycling conditions consisted of initial denature at 94oC for 5 minutes, followed by 35 cycles at 94oC for 30 seconds, anneal at 56oC for 1 minute, and extension at 72oC for 1 minute, followed by a final extension at 72oC for 5 minutes. The CytB amplicons were sequenced, edited and aligned using SEQUENCHER v 4.10.1 (Gene Codes Corporation), and yielded two haplotypes: one common to SNS and an alternate common to LRS. A restriction enzyme assay that differentiated the two CytB haplotypes was designed using BamHI (MBI Fermentas), and this restriction enzyme assay was applied to 66 adults from each species. The BamHIdigested CytB amplicon produced two restriction fragment length polymorphism patterns: one common to SNS (102,159, and 510 base pairs) and a second common to LRS (159 and 612 base pairs). Taqman assays were designed for the AL176 SNP (designated as Dlux176) and a SNP in CytB (designated as Dlux434) using Primer Express v3.0 (Applied Biosystems) (See Table 1 in main article). For each assay, we designated the allele most common to a priori identified SNS and LRS as ‘A’ and ‘B’, respectively. Marker discovery using RAD sequencing For RAD sequencing, we selected 24 SNS, consisting of 12 adults from Upper Klamath Lake and 12 from Clear Lake, and 24 LRS, consisting of 13 adults and 4 YOY from Upper Klamath Lake and 7 adults from Clear Lake. A RAD library was generated for each individual using 500 ng of SbfI digested genomic DNA following the methods described by Etter et al. (2011). Two pooled libraries of bar-coded fragments, each containing 12 LRS and 12 SNS, were sequenced by University of Oregon on an Illumina Hi Seq 2000 instrument. We analyzed single-reads and applied several modules from STACKS v. 0.99992 (Catchen et al. 2011) to sort, filter, and align sequence reads into RAD loci. The application process-radtags was used to filter read quality and remove sequences without intact barcodes. Reads with quality score < 33 were removed. Next, we applied ustacks to build stacks of RAD loci for each individual, using a minimum coverage depth of 5 total reads per individual, and enabled the removal and deleveraging algorithms to filter out highly repetitive loci. The application cstacks was used to form a catalog of RAD loci common to all members by merging loci across individuals. We allowed three nucleotide mismatches between loci for the between-individual distance parameter. Last, sstacks was used to match RAD loci from individuals against the catalog formed in cstacks. Species-diagnostic candidate SNPs were identified within the catalogue by first applying scripts that removed monomorphic loci and loci containing two or more nonconsecutive SNPs (https://github.com/marshalhoy/RAD-suckers). We then removed loci having > 20 % heterozygosity across all individuals. Next, individuals having > 10 % heterozygosity over all loci were removed from the catalogue. This step was performed to remove potential SNS-LRS hybrids from the catalogue. Finally, we applied a script to retain loci fixed for alternate alleles between SNS and LRS. The average read coverage for species-diagnostic candidates was 64 reads/SNP for SNS and 56 reads/SNP for LRS. We designed 28 TaqMan assays using the first 75 bases of each RAD locus to avoid sequencing errors that may be incorporated toward the 3’ terminus of single reads. TaqMan sssays were designed using Primer Express v3.0 (Applied Biosystems) and assays were run on a viiA7 Real-Time PCR System (Applied Biosystems). Eighteen of the 28 Taqman assays produced high quality amplification products (See Table 1 in main article). Validation of these 18 assays against the individuals included in the final catalogue indicated that 99.2 % of genotypes were congruent between RAD sequences and TaqMan assays. For each assay, we designated the allele most common to a priori identified SNS and LRS as ‘A’ and ‘B’, respectively. Evaluation of TaqMan assays TaqMan assays were evaluated in 160 YOY identified a priori as LRS (N = 79) and SNS (N = 81) from Upper Klamath Lake. These YOY were not used in the RAD library construction and consisted of individuals pooled across years (2004 – 2010). Allele frequencies and observed and expected heterozygosity were calculated for the a priori identified species, and multilocus FST was estimated between SNS and LRS using GENEPOP 4.2 (Raymond and Rousset 1995). We tested each locus for conformation to Hardy-Weinberg equilibrium and tested all permutations of nuclear locus pairs for genotypic linkage disequilibrium using GENEPOP 4.2. Significance levels were adjusted using the false discovery rate (FDR) procedure (Benjamini and Yekutieli 2001). Test results were considered significant when P exceeded the FDR adjusted critical value (α) for each series of simultaneous tests. References Baird NA, Etter PD, Atwood TS, et al (2008) Rapid SNP discovery and genetic mapping using sequenced RAD markers. Plos One 3 e3376 Benjamini Y, Yekuteili D (2001) The control of false discovery under dependency. Ann Stat 29:1165-1188 Catchen JM, Amores A, Hohenlohe P, Cresko W, Postlethwait JH (2011) Stacks: building and genotyping loci de novo from short-read sequences. G3-Genes Genom Genet 1:171-182 Etter PD, Bassham S, Hohenlohe PA, Johnson EA, Cresko WA (2011) SNP discovery and genotyping for evolutionary genetics using RAD sequencing. Methods Mol Boil 772:157-178 Markle DF, Cavalluzzi MR, Simon DC (2005) Morphology and taxonomy of Klamath basin suckers (Catostomidae). West N Am Naturalist 65:473-489 Meyer A (1993) Evolution of mitochondrial DNA in fishes. In: Hochanchka PW, Mommsen TP (eds) Biochemistry and molecular biology of fishes. Elsevier, New York, pp 1-38 Raymond M, Rousset F (1995) Genepop (version1.2): population genetics software for exact tests and ecumenicism. J Hered 86:248-249