Biochem Ch 8 [10-24
advertisement

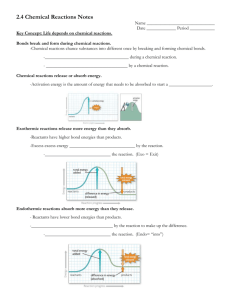
Biochemistry Enzymes as Catalysts Chapter 8 Biochem Ch 8 S= substrate E= enzyme P= product I. Enzyme-catalyzed rxn Enzymes are usually proteins that act to speed the rate of a rxn; they do not “invent” new rxns Catalytic power: rate w. enzyme / rate w.o. enzyme = usually about 106-1014 Enzyme fxn steps: 1) brings substrate into the right orientation 2) Makes/breaks bonds 3) releases products 4) returns to original state 1) binding of substrate: E+S ES 2)conversion of bound substrate to bound products: ESEP 3) release of product: EP E+P Specificity- the ability of an Enzyme to select just one substrate and distinguish this substrate from a group of very similar compounds A. The active site -usually a crevice or cleft in the enzyme -cofactors and fxn’l goups from the chain help to transform the SP Substrate Recognition site- where substrate binds to enzyme initially Functional group- part of the enzyme that interacts with and does all the work to the S; can be directly from the chain, or from a bound coenzyme. Coenzyme- metals or complex organic molecules B. Subsrate-Binding Sites 1. Lock and Key model for S binding -surface that “recognizes” the S and binds with multiple hydrophobic interactions, electrostatic interactions or hydrogen bonds -binding of compounds with a structure that differs from the S even to a small degree may be prevented by steric hindrance and charge repulsion 2. Induced fit model for S binding -once the S binds, the enzyme undergoes conformational changes that repositions the side chains of the a.a in the active site and increases the number of binding interactions -flexible, dynamic surface that warps and changes to best align S to fxn’l groups promotes rxn C. The Transition –State complex Transition-state complex- an unstable, high-energy complex with a strained electronic config. that is intermediate btwn S and P; bonds with in the enzyme make the transition more stable and possible. -Usually at the highest energy level- most unstable S config, but most tightly bound to the fxn’ groups -Activation Energy (EA)- difference in E btwn the S and the transition-state complex -Enzymes work by lowering the EA -From here, collapses backwards into Substrates, or decomps into products -enzyme usually returns to it original form, binds to another S, rinse/repeat Importance to Pharm: Transition-State of substrate binds more tightly to the enzyme than does the Substrate Transition-state analogs are more potent inhibitors of an enzyme than S analogs (they look like the substrate does in the T-S complex) -a drug developed as a transition-state analog would be highly specific for the enzyme it is designed to inhibit, but are highly unstable and unlikely to reach the site of action -so drugs being designed as Biochemistry Enzymes as Catalysts Chapter 8 1) almost T-S analogs but are stable 2) “prodrug” that gets mod’d in the body into the T-S analog at the site of action 3)using the T-S analog to design a complementary antibody abzymes- catalytic antibodies. Have a similar a.a. arrangement to the active site of the enzyme in the T-S -they act like artificial enzymes -abzymes have been created against analogs of the T-S compex of cocaine esterase, the enzye that degrades cocaine in the body. These abzymes have esterase activity. Monthly injections of the drug can destroy cocaine in the blood, decreasing the dependence of addicted folks. II. Catalytic mechanism of chymotrypsin -digestive enzyme released into intestines that catalyzes the hydolysis of specific peptide bonds in denatured proteins -a serine protease; uses serine in the active site to form a covalet intermediate during proteolysis Scissile bond- the bond that is cleaved in lysis Hydrolysis- use of water to break a bond; -OH added to the carbonyl carbon, -H added to N Proteolysis- the hydrolysis of a peptide bond in a protein A. The rxn without the enzyme -the oxygen would carry a full (-) charge in its T-S - the rate of the rxn would be very slow bc there are too few OH- molecules in H2O with enough energy to form the T-S complex and too few OH- molecule colliding with the S at the right orientation B. Catalytic processes in the rxn cat’d by Chymotrypsin Two stage rxn 1) cleavage of the peptide bond in the denatured substrate protein and the formation of a covalent acyl-enzyme intermed. 2) hydrolysis of the acyl-enzyme intermed. to release the remaining portion of the subtrate protein -rate of rxn increased bc fxn’l groups in the enzyme: - active site activate the attacking hydroxyl group -stabilize the oxyanion TS complexes -form covalent intermeds - destabilize the leaving group 1) Specificity of binding to chymotrypsin -hydrolyzes the peptide bond on the carbonyl side of Phe, Tyr, and Trp in a denatured protein -recog site has hydrophobic pocket; holds onto the hydrophobic a.a at the carbonyl group of the scissile bond -Glycines hold the substrate in place -proximity and orientation- specificity is improved by moving the attacking group into position 2. Formation of the acyl-enzyme intermeditate in chymotrypsin nucleophilic catalysis- a nucleophile is a chemical group that is attracted to the (+) charged nucleus 1st stage of rxn- Ser195 hydroxyl group attacks the carbonyl carbon of the peptide bond of the denatured protein S…cleaving it -Aspartate and His coop in coverting the hydroxyl group into a better nucleophilic attacking group by giving it a more (-) charge. Acid-base catalysis- (the method by which) His acts as a base and pulls a proton from the serine hydroxyl; the protonated his is stabilized by the (-) of nearby aspartate -needed because the pK of serine hydroxyl group is very high; needs stabilizing Catalytic triad- the His-Ser-Asp team work combo is referred to as.. -this same combo is found in many proteases in the digestion and clotting system 2nd stage: an oxyanion tetrahedral transition-state complex formed that is stabilized by H bonds with NH groups in the backbone -described as showing “electronic strain” – an electrostatic surface that would be highly improbable if it were not stabilized by the bonds with fxn’l groups on the enzyme. Biochemistry Enzymes as Catalysts Chapter 8 Stabilization of the T-S complex- lowers the energy lvl and increases the number of molecules that reach this energy lvl Covalent catalysis-forming a full covalent bond with the carbon of the carbonyl group as the peptide bond is cleaved to create a stable intermediate (Serine does this in the next step) -very commonly done in proteins (usually by serine or cysteine) -Is hydrolyzed by acid-base catalysis later destabilization of developing product- subjected to some degree of charge repulsion 3. Hydrolysis of the acyl-chymotrpsin intermediate -hydrolyzation to release the bound carbonyl-side peptide -histidine activates water to form an OH- for nucleophilic attack second oxyanion T-S complex -His adds a proton back to sering, rxn is completeproducts dissociate C. Energy Diagram (image) III. Fxn’l groups in catalysis -all enzymes use proximity and orientation, stabilization of Transition state by electrostatic interactions, not all form covalent intermediates cofactors- nonprotein compounds that participate in the catalytic process coenzymesmetal ions- (Fe2+, Mg2+, Zn2+) metallocoenzymes- (like the Fe2+-heme in Hb) A. Functional groups on a.a. side chains -almost all of the polar a.as take part in catalysis in one or more enzymes -Ser, Cys, Lys, His- covalent catalysis -His- it’s pKa allows it to donate and accept protons at neutral pH- often in acid-base catalysis -most polar a.a. side chains are nucleophilic and partake in nucleophilic catalysis by stabilizing more (+) groups that develop during rxn B. Coenzymes in Catalysis-usually made from vitamins -have very little specificity and very little activity without an enzyme -each coenzyme does specific types of rxns: 2 classes: 1. activation-transfer coenzymes -form a covalent bond with a portion of the substrate -activates the substrate for transfer, addition of water, etc. -fxn’l group- part that binds to the substrate -separate part binds to the enzyme -Thiamine (B1) pyrophosphate- made in human cells by B1 added to pyrophosphate. -pyrophosphate= chelates Mg2+, which binds to the enzyme -fxn’l group extends into the active site is a reactive C with a removable proton -in every enzyme-substrate rxn that it takes part in, it always does the same thing: The reactive C forms a covalent bond with a substrate keto group, and cleaves the nearby C-C bond -CoA, biotin, pyridoxal phosphate= are all activation-transfer coenzymes made from vitamins -CoA (CoASH) made from B5 (pantothenate)- binds to enzyme with an ADP bond; fxnl group is a sulffydryl group on other end, is a nucleophile that always attacks carbonyl groups and forms acyl thioesters -most coenzymes are regen’d during the rxn. -CoASH and a few redox coenz are transformed during the rxn into products but dissociate from the enzyme at the end of the rxn -the original form is usually re-made in other rxns -the amt of them in the cell is usually constant -Biotin- covalently bonded to a lysine in carboxylases -fxnl group= N atom that covalently binds a CO2 (requires energy); activates it for add’n of another molecule -only functions in carboxylation reactions in humans Biochemistry Enzymes as Catalysts Chapter 8 -pyridoxal (B6) phosphate-has reactive aldehyde forming a covalent bond with amino groups -a ring nitrogen pulls e-‘s out of a bond, until it breaks 3 things in common with all of the above: 1) specific group involved in binding the enzyme 2) separate and diff fxnl or reactive group that participates directly in the catalysis of one type of rxn by forming a covalent bond with the substrate 3) dependence on the enzyme for additional specificity of substrate and add’l catalytic power 2. oxidation-reduction coenzymes -aid enzymes called oxidoreductases -oxidized=losing e-‘s; so the oxdized C has fewer Hatoms or gains an O atom -reduced=gaining e-’s; shows as the gain of H or the loss of O NAD+ and FAD=can transfer e-‘s together with Hydrogen Vit C and E- act as antioxidants and protect against free radical injury -dehydrogenases- (enzyme subclass)-they transfer H (in form of H atoms of hydride ions) from the substrate to an eacceping coenzyme such as NAD+ -Oxidation-Reduction coenzymes- follow all the same rules as activation-transfer coenzymes, except that they do not make covalent bonds -Enzyme- Lactate dehydrogenase catalyzes transer of e- from lactate to NAD+ -Coenzyme- NAD+= synth’d from niacin (forms the nicotinamide ring) and ATP -has ADP portion that binds to the enzyme; causes conf. change -fxn’l group= the carbon on the ring opposite the (+) N; this carbon accepts the hydride ion transferred from a specific carbon atom on the substrate. This forms a keto group (C=O) when the H+ from the substrate –OH group dissociates. **Pst! Hydride= H atom with 2 e-‘s -enzyme is to contribute a histidine N, that can bind the dissociable proton on lactate, thereby making it easier for NAD+ to pull off the other Hydrogen with both e-’s. -NADH dissociates C. Metal ions in Catalysis -Metal ion, which have a positive charge, act as electrophiles -assisit in binding to the substrate -stabilize developing anions in the rxn -can accept and donate e-‘s in redox rxns -capable of bindings multiple ligands in their coordination sphere -help to bind substrates to enzymes or coenzymes -ex: Phosphate groups of ATP are usually bound to enzymes through Mg2+ chelation -can bind to anionic substrates or intermediates of the reaction to alter their charge distriution; adds to catalytic power -ADH (alcohol dehydrogenase) transfer e-‘s from EtOH to NAD+ to make acetaldehyde and NADH -In active site- serine pulls a proton off the ethanol –OH group makes a (-) charge on the Oxygen stabilized by zinc; this electronic config allows the trasnfer of a hydride ion to NAD + D. Noncatalytic roles of cofactors-can bind portions of an enzyme together to help make the 3* structure -can serve as substrates that are cleaved during the rxn IV. Optimal pH and temp -enzyme fxn is best in physiologic pH; any higher or lower and fxn is diminished -Most human enzymes fxn best at about 37* C (lower, and the vibrational energy is not high enough) (higher and the proteins get denatured) V. Mechanism- based inhibitor Biochemistry Enzymes as Catalysts Chapter 8 Inhibitors- are compounds that decrease the rate of an enzymatic rxn; mimic or participate in an intermed step of the catalytic rxn; includes transition state analogs and compounds that can irreversibly bind with fxn’l groups in the active state. A. Covalent inhibitors -form covalent bond in the active site -ex: diisopropylphosphofluoridate (DFP)- an organophosphorus compoud; prototype for nerve gas Sarin and other toxins like insecticides -works by forming a irreversible bond with acetylcholine esterase preventing acetylcholine from being removed from the nerve synapses -also inhibits serine proteases, but the effects of those are not lethal -ex- aspirin- works through covalent acetylation of an active site serine in the prostaglandin endoperoxide synthase (cyclooxygenase) -aspirin resembles a portion of the prostaglandin precursor that is a substrate for the enzyme B. Transition-State Analogs and Compounds that Resemble Intermediate Stages of the Reaction -discussed earlier 1) Penicillin -binds tightly to glycopeptidyl transferase -this enzyme is required for synthesis of the bacterial cell wall -it catalyzes a partial rxn with penicillin that covalently binds penicillin to its own active-site serine -suicide inhibitor- active site inhibitors that undergo partial rxn to form irreversible inhibitors in the active site (like penicillin) 2) Allopurinol- drug used to treat gout. Decreases urate production by inhibiting xanthine oxidase -the enzyme commits suicide by converting a drug to a transition-state analog -enzyme contains a molybdenum-sulfide (Mo-S) complex that binds the substrates and x-fers the e-‘s required for the oxidation reactions it normally does. -Xanthine oxidase oxidizes allopurinol to oxypurinol which binds tightly to the Mo-S complex in the active site -Enzyme commits suicide cannot do its usual job of degrading purines into Uric Acid Rxn: GMPGuanineXanthine-----xanthine oxidaseUrate AMPhypoxanthine-----xanthine oxidaseXanthineUrate C. Heavy metals -toxicity caused by tight binding of metals -Hg, Pb, Al, or Fe to a fxn’l group in an enzyme -are mostly unspecific in the enzymes that they inhibit -Hg- binds so many enzymes (mostly at sulfhydryl groups) that they don’t know which of the inhibited enzymes is responsible for toxicity -Pb-inhibits through replacing the normal fxn’l metal in an enzyme (Ca, Fe, Zn); neurological toxicity results from displaced Ca in regulatory proteins that are important for CNS and other tissues Biochemistry Enzymes as Catalysts Chapter 8 Clinical Dennis Veere Presented to ER after ingesting insecticide malathion- used for killing flies and other insects Sx: vomited repeatedly; c/o abd cramps, salivating and uncontrollable defecation, involuntary twitching of extremity muscles TX: nasogastic tube for stomch lavage, IV fluids, recorded vitals: p: 48, bp: 78/48 -sx caused by inhibition of acetylcholine esterase. Malathion is metabolized in the liver to a toxic derivative (malaoxon) that binds to the active-site serine in acetylcholine esterase and other enzymes. ACh builds up in the synapse and overstimulates the ANS…explaining his vomiting, abd cramping, salivation and sweating. ACh is also a NT for the somatic motor nervous system, where its buildup caused the involuntary motor twitch. Lotta Topaigne: Dx: acute gouty artritis in R great toe Caused by insoluble urate crystals Tx: allopurinol 150mg BID Al Martini 44 y.o old alcoholic for the past 5 years Sx: diminished appetite; sudden irritability, confusion after 2 fifths of scotch and eating very little, poorly oriented to person, time, place P: 104; BP slightly low, early congestive heart failure Most tissues in the body are affected by chronic ingestion of excessive EtOH (liver, pancreas, heart, reproductive organs, CNS, and fetus mostly) -things directly caused by EtOH: psychtropic effects on the brain, inhibition of vit transport, -most other issues brought on by EtOH metabolism -Functional deficiency: -drugs and toxins that inhibit protein required for conenzyme synth can cause symptoms of vit defici. Dietary Deficiency- inadequate intake Deficiencies of any sort causes: -Apoenzyme- enzyme without its cofactor Ethanol is antivitamin- decreased the cellular content of almost every coenzyme -ex: decreases thiamine absorption through intestinal mucosa; Thiamine is converted to Thiamine pyrophosphate (TPP); is a coenzyme in decarboxylation of alpha-keto acids (think krebs); dysfunction occurs in the CNS and PNS and cardiovascular system and other organs. -ex: acetaldehyde (produced from EtOH oxidation) displaces pyridoxal phosphate from its protein-binding sites, thereby accelerating its degradation -is highly reactive, toxic and immunogenic, is the cause of liver injury in alcoholism -ADH is active as a dimer with an active site containing zinc in each subunit -at low level EtOH intake, much of the acetaldehyde is safely oxidized to acetate in the liver by acetaldehyde dehydrogenases. Biochemical Comments: OxidoreductasesRxns in which at least one substrate gains e-‘s/becomes reduced and another loses e-‘s/becomes oxidized -dehydrogenases-accepts/donates e-‘s in the form oh hydride or H atoms - usually works with coenzyme NAD/NADH - hydroxylases or oxidases-when one O atom goes into a substrate and the other O into water, or when both atoms go to water -require metal ions (like Fe2+ for e- transfer) -Oxygenases- when both atoms of O are incorporated into the acceptor TransferasesCatalyze group transfer rxns (the transfer of a fxn’l group from one molecule to another) -Kinases- transfer high-energy phosphate groups -Glycosyltransferase- transfer carbohydrate residues Biochemistry Enzymes as Catalysts Chapter 8 -Acyltransferase- transfer fatty acyl groups For those 3, the group being transferred exists as a good leaving group -Transaminases or aminotransferases: do Transamination=Nitrogen group from an a.a is donated to an alpha-keto acid -forms either a new a.a + the alpha-keto acid corresponding to the donor a.a. -pyridoxal phosphate required for all transamination rxns -synthases- transferases called this when the important aspect of the rxn is the compound made ex:glycogen synthase- transfers a glucosyl residue from UDP-glucose to the end of a glycogen molecule Hydrolases-cleavage of C-O ,C-N, C-S by addn of water (adding –OH to on part of thte bond and -H to the other Enzyme class name based on the group being cleaved (proteases= hydrolase that cleaves protein) Lyases- cleavage of C-C, C-O ,C-N bonds by means other than hydrolysis or oxidation. -Aldolases- on C-C bonds; -decarboxylases- on C-C bonds; when carbon dioxide is released from a substrate -thiolases- on C-C bonds- when the sulfer containing nucleophile of cysteine or CoASH is used to break a C-C bond - need an e- sink to stabilize the carbanion formed during breakage -dehydratases-remove the elements of water from 2 C-C bonds C=C -synthases- also fall into this broad category Isomerases- rearrangement of the existing atoms of a molecule; make isomers of the starting material -isomerases- rearrange the bond structure of a compound -mutases- catalyzes movement of a phosphate from one atom to another Ligases- synthesis C-C, C-S, C-O, C-N bonds in rxns coupled to the cleavage of ATP or another nucleotide. -Carboxylases- add CO2 to another compound; require ATP for Energy; usually also requires biotin -Sythetases- (not sythases)- these derive energy for the new bond from ATP