Puberty and the oestrus cycle

Puberty and the oestrus cycle:
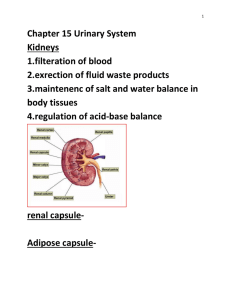
The endocrine and nervous systems play interwoven role in the cascade on events leading to the formation of mature gametes, fertilization, establishment and maintenance of pregnancy, birth and, finally, rearing of offspring. These processes begin at puberty.
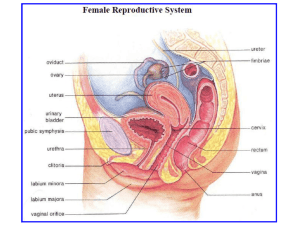
In the female, puberty is marked by the onset of regular cyclic activity in the ovary affecting behavior and the entire genital system(the oestrus cycle in domestic animals).
Factors influencing the onset of puberty include age, body weight, breed nutrition, disease and, in some species, season of the year and proximity of a male. The ages at which the common domestic species reach puberty are listed in Table 3-1 page 27.
After the onset of puberty, signals provided by certain regions in the brain, including the pineal gland, hypothalamus and the pituitary gland, allow for the fertilizable oocytes.
From the anterior pituitary gland, the gonadotropins (i.e hormones stimulating cells within the gonads) FSH (follicle-stimulating hormones) and LH (Luteinizing hormone) are released. This release is controlled by GnRHs (gonadotropin-releasing hormones) that are secreted from the hypothalamus and conveyed to the anterior pituitary gland through hypothalamo-hypophyseal portal blood circulation. Secretion of GnRHs, and thus of FSH and LH is influenced by visual, olfacrory, auditory and tactile stimuli from the environment and also by homeostatic feedback systems within the animal.
FSH released from the pituitary gland reaches the ovary by the systemic circulation. In the ovary, it stimulates a pool of growing follicles to develop. Follicular growth occurs when follicles are recruited from the primordial pool and develop into primary, secondary and tertiary follicles. The majority of growing folliclesail to complete their growth phase, most degenerate through a process referred to as atresia with only minority completing their growth to the point of ovulation.
At least in the large domestic species follicular growth is initiated during fetal life. However, none of the oocytes enclosed in the follicles resume meiosis until puberty is reached.
Up on activation of the primordial follicle, the follicular cells start to proliferate and form a cuboidal monolayer around the oocyte to establish the primaryfollicle.(Figures 4-8, 4-9, 4-10,in the text book). The follicular cells are now called granulosa cells. With this activation, a phase of oocyte growth is initiated in which the oocyte of the domestic species grows from less than 30 µ m to more
than 120 µ m in diameter. Granulosa cells proliferate to form several layers around the oocyte in what becomes known as the secondary follicle. The oocyte and the surrounding granulosa cells synthesize certain glycoproteins that are deposited between itself and the surrounding granulosa cells as the zona pellucida. This structure is traversed by numerous projections from the innermost granulosa cells which thereby maintain contact with the oocyte through gap junction.
The stromal cells surrounding the granulosa cells differentiate into an inner theca interna, a layer of steroid-producing cells, and the outer theca externa, made up of concentric layers of cells that have supportive function.
As development continues, fluid-filled spaces appear between the granulosa cells and coalesce into a single cavity, the antrum, characterizing the tertiary follicle. In parallel with the expansion of the antrum, the oocyte becomes located in a protrusion of granulosa cell, the cumulus oophorus, extending into the antrum.
As the follicle develops, so does the oocyte until it achieves it’s a characteristic structure. The process of ovulation is triggered by the preovulatory LH surge.
Follicular and oocyte maturation
The tertiary follicle continues its development and, it selected for ovulation, enters a final phase of follicular and oocyte maturation stimulated by thepreovulatory LH surge.
The period from the onset of the LH surge to ovulation is species-specific and varies from less than
12 hr to more than 40 hr. The preovulatory maturation of the oocyte has nuclear as well as cytoplasmic components. Nuclear maturation refers to the process of meiosis which is resumed from diplotene stage (resting stage) of 1 st meiosis and continues to the metaphase of 2 nd meiosis when the oocyte is ovulated. The nucleus of the primary oocyte is often referred to as the germinal vesicle, and resumption of meiosis is morphologically evidenced by breakdown of this structure.
Cytoplasmic oocyte maturation involves the structuring and modulation of many of the organelles of the oocyte.
Oestrogens which are produced in the granulosa and thecal cells lining the follicle and surrounding the oocyte, exert a positive feedback on the hypothalamic GnRH secretion. Another principal effect of the oestrogens is to induce oestrous symptoms, and so puberty is often signalled by the occurrence of the first period of oestrus or heat.(sexual receptivity).
After puberty the animal enters a phase of life characterized by repeated oestous cycles. The oestrous cycle is subdivided into prooestrus, oestrus, metoestrus, and dioestrus (Tables 3-2,3-3) pages 27, 28.
In most species, the increasing levels of oestrogens produced during prooestrus reach a certain threshold level that leads to a major surge of GnRH being secreted by the hypothalamus during oestrus. On reaching the anterior pituitary gland, the GnRH stimulates a secretory surge of mainly
LH. In the ovary, LH is needed to bring about the final maturation of the oocyte(s) and their release through the process of ovulation.
Following ovulation,the cells that lined the follicle before its rupture become transformed into a corpus luteum, under the continued influence of LH, during metoestrus. The development of the corpus luteum begins gradually a few hours before ovulation and is marked by the synthesis of progesterone, instead of oestrogens. The corpus luteum formed by these cells after ovulation is a well-defined, sometimes slightly cavitated, spherical structure, named for its yellowish colour.
During dioestrus, the production of progesterone by the corpus luteum reaches its maximum. The principal roles of progesterone are: (1)to exert a negative feedback on the hypothalamus, inhibitingGnRH release and, therefore, new recruitment of oocytes for ovulation. (2) to prepare the endometrium for pregnancy.
If insemination and conception occur, progesterone is the principal hormone responsible for the maintenance of pregnancy.
In the non pregnant animal, the life span of the corpus luteum is relatively short, in the absence of an embryo within the uterus, the endometrium releases prostaglandins-F2α leading to luteolysis
(regression of corpus luteum). The consequent decline in progesterone results in removal of the progesterone block on the hypothalamic GnRH secretion and allows for resumption of oestrus cycle.
In the pregnant uterus, endometrial prostaglandin F2α release into the blood stream is blocked, leading to persistence of the corpus luteum.
The domestic breeds of pigs and cattle are non-seasonal polycyclic animals, meaning that sows and cows experience recurring cyclic activity throughout the year, interrupted only by pregnancy, lactation or pathological conditions.
In contrast, the mare, ewe, doe and queen are seasonally polycyclic, their cyclicity is profoundly influenced by the amount and timing of light. The bitch is monocyclic and experiences long of anoestrus between single periods of oestrus. Normally, one or two (sometimes three) oestrus periods are seen per year separated by longer anoestrus periods with no obvious seasonality.
The corpus luteum produce progesterone until the end of the third month. Thereafter, the placenta takes over until term.