IB104 - Lecture 29 - Genetics
advertisement

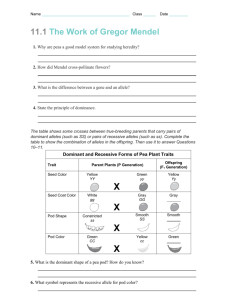
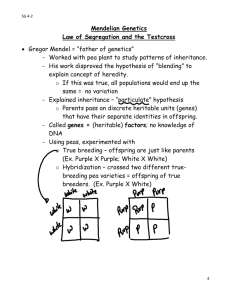
IB104 - Lecture 29 - Genetics - 11/6/13 Reading: Starr et al, Chapter 13 What is so special about Gregor Mendel? A major problem with trying to study patterns of inheritance (that is, genetics) was that comparison of overall characters of individuals, e.g. people, left one with an overwhelming array of small differences, and if these were followed through generations then it seemed that they were simply mixed and averaged in progeny. In the absence of real evidence, a variety of wild notions about mechanisms of inheritance were postulated in the 19th century, egg. "gemmules" of Charles Darwin that were supposed to carry characteristics from cells to gonads to be passed on to the next generation. This was part of the notion of acquired characters, attributed mainly to Jean Baptiste Lamarck. Another notion was that genes on chromosomes were parsed out to various tissues for their development. It is now clear that inheritance of acquired characteristics is not true and that all cells with nuclei are totipotent, at least in theory and now in practice with cloning of various animals, mammals included. Mendel instead used unit traits that characterized individuals, almost to the point of using qualitative versus quantitative traits. This is the heritage of Mendelian genetics and is the basis for all the understanding of genetics we will talk about. He also used a numerical and statistical approach to asking whether his results fit his theories, essentially the basis for all modern biology, some of which you will see in your labs. What did he do? He was deputized by the abbot of his monastery in Brno in what was until recently Czechoslovakia to work on inheritance of characteristics in peas, back in the 1860's. He chose a series of lines that had been bred for various peculiarities. These were true-breeding lines, that is, both alleles at the locus of interest were the same, so that every generation one obtained all individuals with the same peculiar characteristics. Then he crossed (mated) individuals of pairs of lines to see what would happen. Such controlled crossing is relatively easy in animals, other than humans, but in plants such as peas where flowers have both male and female parts, it needs some manipulation, i.e. removal of the male anthers from the "female" plants before fertilization. He had a series of at least seven characters to work with, egg. pea shape and color, pod color, flower color, and plant height. His first observation was that the hybrids were all of one kind. He might have quit at this point, but instead left the hybrids to fertilize themselves and monitored the "grandchildren". This is equivalent to crossing the hybrid or so-called F1 (for first filial generation) individuals with each other in animals. In each case, the "lost" character was returned, and in a 3:1 ratio, give or take a bit (that is, roughly a quarter of the F2 progeny had the lost characteristic back). What was going on? Remember that he did not know about genes or chromosomes or diploidy or meiosis, but figured out mathematically that the best way to explain this was to assume first that the characters are controlled by particulate factors we now call genes. Second, that one of these factors was dominant in expression thereby masking the presence of the other in the F1, and third that there were two copies of each factor such that the ratio in the 2nd generation of progeny was 3:1. We have several terms to talk about these situations. Homozygote refers to when an individual has two copies of the same allele, versus heterozygote where an individual has one copy each of two different alleles. Also dominant versus recessive, i.e. some alleles when present in a heterozygote mask the presence of another allele. One can commonly understand this today in terms of the genes and biochemistry in that the dominant allele is commonly the "normal" version, while recessive alleles are commonly mutant versions that destroy the gene's function. Given the built-in redundancy of diploids, the presence of a single normal copy is generally sufficient to yield a normal individual, hence that allele is dominant. And indeed it turns out that the recessive characteristics Mendel worked with were caused by mutations in particular genes that inactivated the particular gene (e.g. removed the ability to make seed or pod color, or shortened the plant overall). Finally, one needs to distinguish between genotype and phenotype, the former being the actual combination of alleles an individual has, and the latter the resultant appearance of the individual. Make sure you understand what is going on at the gamete level, because the first step in solving any genetic problem is figuring out what kinds of gametes are formed, and then randomly fusing them with each other to give progeny of various kinds. This example exemplifies the first principle of genetics, the principle of segregation, i.e. in Meiosis I the homologs segregate faithfully to separate gametes. Dihybrid crosses and independent assortment What happens if we cross a green seed and tall plant (dominant alleles) with a yellow seed and short plant (recessive alleles)? Now we are dealing with two different loci and two alleles of each in the cross. Recall the chromosomes in meiosis - if these loci are on different chromosomes then their alleles on the homologs will assort into the gametes independently of their parental origin. To work out this problem, we first figure out all the possible gametes, then we do a Punnet Square (named for an Englishman who first used this method to do genetics of chickens) to figure out all the possible genotypes in the F2. Then we convert these genotypes to phenotypes to see what Mendel got. In this example, with green seed and tall plants being dominant to yellow seed and short plants, in the F2 one obtains a classic 9:3:3:1 ratio of 9 green seed, tall plants: 3 green seed, short plants: 3 yellow seed, tall plants: 1 yellow seed, short plants. Linkage When two loci are on the same chromosome, then they are likely to be inherited together, not at random as in independent assortment. However, the possibility of crossing over resulting in recombination between different alleles from the mother and father homologs makes it more complicated. So, if two loci are very close to each other on a chromosome their alleles will almost always be inherited together, that is, they are closely linked. If they are at opposite ends of the same chromosome then crossovers will routinely occur between them and they will be only weakly linked to each other, and will be inherited almost as if by independent assortment. It turns out that of the 7 true-breeding characteristics Mendel studied,, six were on different chromosomes, and the seventh was so weakly linked to another that it behaved genetically as if on a separate chromosome. Either Mendel was incredibly lucky, or he discarded and did not work further with characteristics that did not fit his theories. In either case he discovered the basic rules of inheritance and genetics, rules that apply to all diploid eukaryotic organisms as we shall see next with humans.