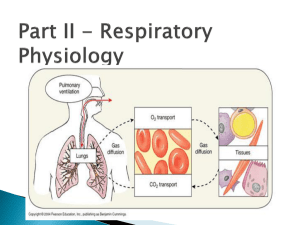
The respiratory system The cells of the body require a continuous supply of oxygen to produce energy and carry out their metabolic functions. Furthermore, these aerobic metabolic processes produce carbon dioxide, which must be continuously eliminated. The primary functions of the respiratory system include: • Obtaining oxygen from the external environment and supplying it to the body’s cells • Eliminating carbon dioxide produced by cellular metabolism from the body The process by which oxygen is taken up by the lungs and carbon dioxide is eliminated from the lungs is referred to as gas exchange. Blood–gas interface Gas exchange takes place at the blood–gas interface , which exists where the alveoli and the pulmonary capillaries come together. The alveoli are the smallest airways in the lungs; the pulmonary capillaries are found in the walls of the alveoli. Inspired oxygen moves from the alveoli into the capillaries for eventual transport to tissues. Entering the lungs by way of the pulmonary circulation, carbon dioxide moves from the capillaries into the alveoli for elimination by expiration. Oxygen and carbon dioxide move across the blood–gas interface by way of simple diffusion from an area of high concentration to an area of low concentration. The blood–gas interface consists of the alveolar epithelium, capillary endothelium, and interstitium. Taken together, only 0.5µm separates the air in the alveoli from the blood in the capillaries. The extreme thinness of the blood–gas interface further facilitates gas exchange by way of diffusion. Airways Air is carried to and from the lungs by the trachea extending toward the lungs from the larynx. The trachea divides into the right and left main bronchi; these primary bronchi each supply a lung. The primary bronchi branch and form the secondary, or lobar, bronchi, one for each lobe of lung. The lobar bronchi branch and form the tertiary, or segmental, bronchi , one for each of the functional segments within the lobes. These bronchi continue to branch and move outward toward the periphery of the lungs. The smallest airways without alveoli are the terminal bronchioles. Taken together, the airways from the trachea through and including the terminal bronchioles are referred to as conducting airways. This region contains no alveoli, so no gas exchange takes place in this area. Consequently, it is also referred to as anatomical dead space. The conducting airways carry out two major functions. The first is to lead inspired air to the more distal gas-exchanging regions of the lungs. The second is to warm and humidify the inspired air as it flows through them. The alveoli are delicate structures and may be damaged by excessive exposure to cold, dry air. Branching from the terminal bronchioles are the respiratory bronchioles. This is the first generation of airways to have alveoli in their walls. Finally, there are the alveolar ducts which are completely lined with alveolar sacs. This region, from the respiratory bronchioles through the alveoli, is referred to as the respiratory zone , which comprises most of the lungs and has a volume of about 3000 ml at the end of a normal expiration. All of the conducting airways (trachea through terminal bronchioles) are lined with pseudostratified ciliated columnar epithelium. Interspersed among these epithelial cells are mucus-secreting goblet cells. Furthermore, mucus glands are found in the larger airways. Consequently, the surface of the conducting airways consists of a mucuscovered ciliated epithelium. The cilia beat upward at frequencies between 600 and 900 beats per minute. As a result, the cilia continuously move the mucus away from the respiratory zone and up toward the pharynx. This mucociliary escalator provides an important protective mechanism that removes inhaled particles from the lungs. Mucus that reaches the pharynx is usually swallowed or expectorated. Cartilage. The trachea and primary bronchi contain C-shaped cartilage Rings in their walls; the lobar bronchi contain plates of cartilage that completely encircle the airways. The cartilage in these large airways provides structural support and prevents collapse of the airways. As the bronchi continue to branch and move out toward the lung periphery, the cartilage diminishes progressively until it disappears in airways about 1 mm in diameter. Airways with no cartilage are referred to as bronchioles. As the cartilage becomes more sparse, it is replaced by smooth muscle. Therefore, the bronchioles, which have no cartilage to support them and smooth muscle capable of vigorous constriction, are susceptible to collapse under certain conditions, such as an asthmatic attack. The pleura Each lung is enclosed in a double-walled sac referred to as the pleura. The visceral pleura is the membrane adhered to the external surface of the lungs. The parietal pleura lines the walls of the thoracic cavity. The space in between the two layers, the pleural space , is very thin and completely closed. The pleural space is filled with pleural fluid that lubricates the membranes and reduces friction between the layers as they slide past each other during breathing. This fluid also plays a role in maintaining lung inflation. The surface tension between the molecules of the pleural fluid keeps the two layers of the pleura “adhered” to each other. The water molecules pull tightly together and oppose the separation of the slides. In this way, the lungs are in contact with the thoracic wall, fill the thoracic cavity, and remain inflated. In other words, surface tension in the pleural space opposes the tendency of the lungs to collapse. Mechanics of breathing The mechanics of breathing involve volume and pressure changes occurring during ventilation that allow air to move in and out of the lungs. Thoracic volume. The volume of the thoracic cavity increases during inspiration and decreases during expiration. Inspiration. The most important muscle of inspiration is the diaphragm, a thin, dome-shaped muscle inserted into the lower ribs. A skeletal muscle ,it is supplied by the phrenic nerves. When the diaphragm contracts, it flattens and pushes downward against the contents of the abdomen. Therefore, contraction of the diaphragm causes an increase in the vertical dimension of the thoracic cavity and an increase in thoracic volume. Assisting the diaphragm with inspiration are the external intercostal muscles, which connect adjacent ribs. When the external intercostal muscles contract, the ribs are lifted upward and outward (much like a handle on a bucket). Therefore, contraction of these muscles causes an increase in the horizontal dimension of the thoracic cavity and a further increase in thoracic volume. The external intercostal muscles are supplied by the intercostal nerves. Expiration. Expiration during normal, quiet breathing is passive. In other words, no active muscle contraction is required. When the diaphragm is no longer stimulated by the phrenic nerves to contract, it passively returns to its original preinspiration position under the ribs. Relaxation of the external intercostal muscles allows the rib cage to fall inward and downward, largely due to gravity. As a result, these movements cause a decrease in thoracic volume. Lung volume. No real physical attachments exist between the lungs and the thoracic wall. Instead, the lungs literally float in the thoracic cavity, surrounded by pleural fluid. Therefore, the question arises of how the volume of the lungs increases when the volume of the thoracic cavity increases. The mechanism involves the pleural fluid and the surface tension between the molecules of this fluid. The surface tension of the pleural fluid keeps the parietal pleura lining the thoracic cavity and the visceral pleura on the external surface of the lungs “adhered” to each other. In other words, the pleural fluid keeps the lungs in contact with the chest wall. Therefore, as the muscles of inspiration cause the chest wall to expand (thus increasing the thoracic volume), the lungs are pulled open as well. As a result, lung volume also increases. Elastic behavior of lungs In a healthy individual, the lungs are very distensible; in other words, they can be inflated with minimal effort. Furthermore, during normal, quiet breathing, expiration is passive. The lungs inherently recoil to their preinspiratory position. These processes are attributed to the elastic behavior of the lungs. The elasticity of the lungs involves the following two interrelated properties: • Elastic recoil • Pulmonary compliance The elastic recoil of the lungs refers to their ability to return to their original configuration following inspiration. It may also be used to describe the tendency of the lungs to oppose inflation. Conversely, pulmonary compliance describes how easily the lungs inflate. The elastic behavior of the lungs is determined by two factors: • Elastic connective tissue in the lungs • Alveolar surface tension The elastic connective tissue in the lungs consists of elastin and collagen fibers found in the alveolar walls and around blood vessels and bronchi. When the lungs are inflated, the connective tissue fibers are stretched, or distorted. As a result, they have a tendency to return to their original shape and cause the elastic recoil of the lungs following inspiration. The alveoli are lined with fluid. At an air–water interface, the water molecules are much more strongly attracted to each other than to the air at their surface. This attraction produces a force at the surface of the fluid referred to as surface tension (ST). Alveolar surface tension exerts effects on the elastic behavior of the lungs. It decreases the compliance of the lungs. Normal lungs, however, produce a chemical substance referred to as pulmonary surfactant. Made by alveolar type II cells within the alveoli, surfactant is a complex mixture of proteins and phospholipids , the predominant constituent. By interspersing throughout the fluid lining the alveoli, surfactant disrupts the cohesive forces between the water molecules. As a result, pulmonary surfactant decreases surface tension. Ventilation Ventilation is the exchange of air between the external atmosphere and the alveoli. It is typically defined as the volume of air entering the alveoli per minute. A complete understanding of ventilation requires the consideration of lung volumes. Standard lung volumes. The size of the lungs and therefore the lung volumes depend upon an individual’s height, weight or body surface area, age, and gender. This discussion will include the typical values for a 70kg adult. The four standard lung volumes are: • Tidal volume • Residual volume • Expiratory reserve volume • Inspiratory reserve volume The tidal volume (VT) is the volume of air that enters the lungs per breath. During normal, quiet breathing, tidal volume is 500 ml per breath. This volume increases significantly during exercise. The residual volume (RV) is the volume of air remaining in the lungs following a maximal forced expiration. Residual volume is normally 1.5 l. Expiratory reserve volume (ERV) is the volume of air expelled from the lungs during a maximal forced expiration beginning at the end of a normal expiration. The ERV is normally about 1.5 l. The inspiratory reserve volume (IRV) is the volume of air inhaled into the lungs during a maximal forced inspiration beginning at the end of a normal inspiration. The IRV is normally about 2.5 l. The four standard lung capacities consist of two or more lung volumes in combination : • Functional residual capacity • Inspiratory capacity • Total lung capacity • Vital capacity The functional residual capacity (FRC) is the volume of air remaining in the lungs at the end of a normal expiration. The FRC consists of the residual volume and the expiratory reserve volume and is equal to 3 l. The inspiratory capacity (IC) is the volume of air that enters the lungs during a maximal forced inspiration beginning at the end of a normal expiration (FRC). The IC consists of the tidal volume and the inspiratory reserve volume and is equal to 3 l. The total lung capacity (TLC) is the volume of air in the lungs following a maximal forced inspiration. In other words, it is the maximum volume to which the lungs can be expanded. It is determined by the strength of contraction of the inspiratory muscles and the inward elastic recoil of the lungs. The TLC consists of all four lung volumes and is equal to about 6 l in a healthy adult male and about 5 l in a healthy adult female. The vital capacity (VC) is the volume of air expelled from the lungs during a maximal forced expiration following a maximal forced inspiration. In others words, it consists of the tidal volume as well as the inspiratory and expiratory reserve volumes. Vital capacity is approximately 4.5 l. Total ventilation. The total ventilation (minute volume) is the volume of air that enters the lungs per minute. It is determined by tidal volume and breathing frequency: Total ventilation = tidal volume × breathing frequency = 500 ml/breath × 12 breaths/min = 6000 ml/min Alveolar ventilation. Alveolar ventilation is less than the total ventilation because the last portion of each tidal volume remains in the conducting airways; therefore, that air does not participate in gas exchange. The volume of the conducting airways is referred to as anatomical dead space. The calculation of alveolar ventilation includes the tidal volume adjusted for anatomical dead space and includes only air that actually reaches the respiratory zone: Alveolar ventilation = (tidal volume – anatomical dead space) × breathing frequency = (500 ml/breath – 150 ml dead space) × 12 breaths/min = 4200 ml/min During exercise, the working muscles need to obtain more oxygen and eliminate more carbon dioxide. Alveolar ventilation is increased accordingly. Diffusion Oxygen and carbon dioxide cross the blood–gas interface by way of diffusion. The diffusion of oxygen and carbon dioxide depends on their partial pressure gradients. Oxygen diffuses from an area of high partial pressure in the alveoli to an area of low partial pressure in the pulmonary capillary blood. Conversely, carbon dioxide diffuses down its partial pressure gradient from the pulmonary capillary blood into the alveoli. The partial pressure of a gas (Pgas) is equal to its fractional concentration (% total gas) multiplied by the total pressure (Ptot) of all gases in a mixture: Pgas = % total gas × Ptot The atmosphere is a mixture of gases containing 21% oxygen and 79% nitrogen. Due to the effects of gravity, this mixture exerts a total atmospheric pressure (barometric pressure) of 760 mmHg at sea level. The PO2 of the atmosphere at sea level is 160 mmHg and the PN2 is 600 mmHg. The total pressure (760 mmHg) is equal to the sum of the partial pressures. Partial pressures of oxygen and carbon dioxide in the alveoli As explained in the previous section, the PO2 of the atmosphere is 160 mmHg. The partial pressure of carbon dioxide (PCO2) is negligible. As air is inspired, it is warmed and humidified as it flows through the conducting airways. Therefore, water vapor is added to the gas mixture. The partial pressure of the water vapor is 47 mmHg and, as a result, the PO2 is slightly decreased to 150 mmHg. The PCO2 remains at 0 mmHg. By the time the air reaches the alveoli, the PO2 has decreased to about 100 mmHg. The PCO2 of the alveoli is about 40 mmHg. Oxygen diffuses down its partial pressure gradient from the alveoli into the pulmonary capillary blood until equilibration is reached, the PO2 of this blood reaches 100 mmHg. This blood flows back to the left side of the heart and into the systemic circulation. Therefore, the PO2 of the arterial blood is 100 mmHg. Likewise, assuming that carbon dioxide diffuses down its partial pressure gradient from the pulmonary capillary blood (PCO2 = 45mmHg) into the alveoli until equilibration is reached, the PCO2 of the blood leaving these capillaries should be 40 mmHg. Therefore, the PCO2 of the arterial blood is 40 mmHg. Gas transport in blood Transport of oxygen. Oxygen is carried in the blood in two forms: • Physically dissolved • Chemically combined with hemoglobin Oxygen is poorly soluble in plasma. At a PO2 of 100 mmHg, only 3 ml of oxygen is physically dissolved in 1 l of blood. Assuming a blood volume of 5 l, a total of 15 ml of oxygen is in the dissolved form. A normal rate of oxygen consumption at rest is about 250 ml/min. During exercise, oxygen consumption may increase to 3.5 to 5.5 l/min. Therefore, the amount of dissolved oxygen is clearly insufficient to meet the needs of the tissues. Most of the oxygen in the blood (98.5%) is transported chemically pcombined with hemoglobin. A large complex molecule, hemoglobin consists of four polypeptide chains (globin portion), each of which contains a ferrous iron atom (heme portion). Each iron atom can bind reversibly with an oxygen molecule: O2 + Hb ´ HbO2 As the PO2 increases (as it does in the lungs), more will combine with hemoglobin. When the PO2 decreases, as it does in the tissues consuming it, the reaction moves to the left and the hemoglobin releases the oxygen. Each gram of hemoglobin can combine with up to 1.34 ml of oxygen. In a healthy individual, there are 15 g of hemoglobin per 100 ml of blood. Therefore, the oxygen content of the blood is 20.1 ml O2/100 ml blood. Transport of carbon dioxide. Carbon dioxide is carried in the blood in three forms: • Physically dissolved • Carbamino hemoglobin • Bicarbonate ions As with oxygen, the amount of carbon dioxide physically dissolved in the plasma is proportional to its partial pressure. However, carbon dioxide is 20 times more soluble in plasma than is oxygen. Therefore, approximately 10% of carbon dioxide in blood is transported in the dissolved form. Carbon dioxide can combine chemically with the terminal amine groups (NH2) in blood proteins. The most important of these proteins for this process is hemoglobin. The combination of carbon dioxide and hemoglobin forms carbamino hemoglobin: Hb·NH2 + CO2 ´ Hb·NH·COOH Deoxyhemoglobin can bind more carbon dioxide than oxygenated hemoglobin. Therefore, unloading of oxygen in the tissues facilitates loading of carbon dioxide for transport to the lungs. Approximately 30% of carbon dioxide in the blood is transported in this form. remaining 60% of carbon dioxide is transported in the blood in the form of The bicarbonate ions. This mechanism is made possible by the following reaction: The carbon dioxide produced during cellular metabolism diffuses out of the cells and into the plasma. It then continues to diffuse down its concentration gradient into the red blood cells. Within these cells, the enzyme carbonic anhydrase (CA) facilitates combination of carbon dioxide and water to form carbonic acid (H2CO3). The carbonic acid then dissociates into hydrogen ion (H+) and bicarbonate ion (HCO3– ). As the bicarbonate ions are formed, they diffuse down their concentration gradient out of the red blood cell and into the plasma. This process is beneficial because bicarbonate ion is far more soluble in the plasma than carbon dioxide. As the negatively charged bicarbonate ions exit the red blood cell, chloride ions, the most abundant anions in the plasma, enter the cells. This process, referred to as the chloride shift, maintains electrical neutrality. Many of the hydrogen ions bind with hemoglobin. As with carbon dioxide, deoxyhemoglobin can bind more readily with hydrogen ions than oxygenated hemoglobin. This entire reaction is reversed when the blood reaches the lungs. Because carbon dioxide is eliminated by ventilation, the reaction is pulled to the left. Bicarbonate ions diffuse back into the red blood cells. The hemoglobin releases the hydrogen ions and is now available to load up with oxygen. The bicarbonate ions combine with the hydrogen ions to form carbonic acid, which then dissociates into carbon dioxide and water. The carbon dioxide diffuses down its concentration gradient from the blood into the alveoli and is exhaled. Regulation of ventilation The rate and depth of breathing are perfectly adjusted to meet the metabolic needs of the tissues and to maintain a PO2 of 100 mmHg, a PCO2 of 40 mmHg, and a pH of 7.4 in the arterial blood. Breathing is initiated spontaneously by the central nervous system and occurs in a continuous cyclical pattern of inspiration and expiration. The three major components of the regulatory system for ventilation are: • Medullary respiratory center • Receptors and other sources of input • Effector tissues (respiratory muscles) Aggregates of cell bodies within the medulla of the brainstem form the medullary respiratory center, which has two distinct functional areas: • Dorsal respiratory group • Ventral respiratory group The aggregate of cell bodies in the dorsal region of the medulla is the dorsal respiratory group (DRG). The DRG consists primarily of inspiratory neurons. These neurons are self-excitable and repetitively generate action potentials to cause inspiration. The inspiratory neurons descend to the spinal cord where they stimulate neurons that supply the inspiratory muscles, including those of the phrenic nerves and the intercostal nerves. These nerves then stimulate the diaphragm and the external intercostal muscles to contract and cause inspiration. When the inspiratory neurons are electrically inactive, expiration takes place. Therefore, this cyclical electrical activity of the DRG is responsible for the basic rhythm of breathing. Furthermore, the DRG is likely the site of integration of the various sources of input that alter the spontaneous pattern of inspiration and expiration. The aggregate of cell bodies in the ventral region of the medulla is the ventral respiratory group (VRG). The VRG consists of expiratory and inspiratory neurons. This region is inactive during normal, quiet breathing. (Recall that expiration at this time is a passive process.) However, the VRG is active when the demands for ventilation are increased, such as during exercise. Under these conditions, action potentials in the expiratory neurons cause forced, or active, expiration. These neurons descend to the spinal cord where they stimulate the neurons that supply the expiratory muscles, including those that innervate the muscles of the abdominal wall and the internal intercostal muscles. Contractions of these muscles cause a more rapid and more forceful expiration. Inspiratory neurons of the VRG augment inspiratory activity. These neurons descend to the spinal cord where they stimulate neurons that supply the accessory muscles of inspiration including those that innervate the scalenus and sternocleidomastoid muscles. Contractions of these muscles cause a more forceful inspiration. In summary, the regulation of ventilation by the medullary respiratory center determines the: • Interval between the successive groups of action potentials of the inspiratory neurons, which determines the rate or frequency of breathing (as the interval shortens, the breathing rate increases) • Frequency of action potential generation and duration of this electrical activity to the motor neurons, and therefore the muscles of inspiration and expiration, which determines the depth of breathing, or the tidal volume (as the frequency and duration of stimulation increase, the tidal volume increases) The medullary respiratory center receives excitatory and inhibitory inputs from many areas of the brain and peripheral nervous system, including: • Lung receptors • Proprioceptors • Pain receptors • Limbic system • Chemoreceptors
 0
0
advertisement
Related documents









Download
advertisement
Add this document to collection(s)
You can add this document to your study collection(s)
Sign in Available only to authorized usersAdd this document to saved
You can add this document to your saved list
Sign in Available only to authorized users