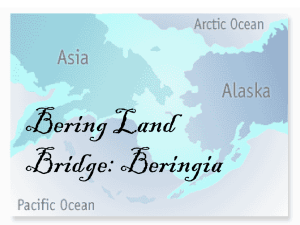
complete - North Pacific Research Board
advertisement

1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18 19 20 21 22 NORTH PACIFIC RESEARCH BOARD FINAL REPORT Biophysical Moorings NPRB Project B52 Final Report Phyllis J. Stabeno1, Jeffrey M. Napp2, Terry Whitledge3 1 NOAA Research, PMEL, 7600 Sand Point Way NE, Seattle, WA, 98115, 206-526-6453, 23 Phyllis.stabeno@noaa.gov 24 2 25 3 26 27 28 29 30 31 NOAA Fisheries, AFSC, 7600 Sand Point Way NE, Seattle, WA, 98115 UAF/SFOS, Institute of Marine Science, PO Box 757220 October 2013 i 32 Abstract 33 This project resulted in collection of three years of nearly continuous data at four mooring sites, 34 and hydrographic and plankton sampling around the sites. Major findings from these data 35 include: 36 37 38 Identified warm and cold years in southeastern Bering Sea - warm (cold) years have little (extensive) ice in March – April. Identified transition from strong interannual variability in warm/cold years before 2000 39 and multi-year variability (stanzas) after 2000 - stanzas of warm years resulted in 40 decreased concentrations of large zooplankton -populations rebounded during cold 41 period. 42 Identified sharp division between northern and southern shelves at ~60 °N. 43 During cold years, the monthly-mean currents over the southeastern shelf are largely 44 45 westward, while in warm years the flow is northward during winter. 46 47 Flow on southern middle shelf is complex and not completely dominated by winds winter bottom currents at M2 are complex, summer bottom currents at M4 are southward. 48 Temperature dominates stratification in the south, and temperature and salinity are equally important in the north. 49 The strength of stratification is not related to warm or cold years. 50 Approximately 1 m of ice melts over the southern (northern) shelf during ice advance 51 52 (retreat). If ice is present after mid-March, spring phytoplankton bloom is related to ice retreat; if 53 ice is absent after mid-March, a spring bloom occurs in May-June, a fall bloom often 54 occurs in late September. 55 The extensive mooring data provided information for model validation. 56 57 Key words: Bering Sea, shelf domains, currents, fluorescence, zooplankton, nutrients, 58 stratification, warm-cold years, north-south structure, sea ice 59 60 Citation: Stabeno, P. J., N. B. Kachel, C. W. Mordy, J. M. Napp, and T. E. Whitledge. 2013. 61 Four Biophysical Mooring sites on the eastern Bering Sea shelf. NPRB Project B52 Final Report, 62 204 pages. 63 64 ii 65 Table of Contents 66 Abstract ii Keywords ii Citation ii Table of Contents iii Study Chronology 1 Introduction 3 Objectives 4 Manuscripts 12 Published Manuscripts 13 Chapter 1 14 Chapter 2 15 Chapter 3 16 Chapter 4 18 Chapter 5 19 Chapter 6 20 Chapter 7 21 Chapter 8 22 Chapter 9 23 Chapter 10 25 Chapter 11 26 Submitted Manuscripts Chapter 12 27 28 Introduction 29 Data and Methods 31 Results 33 Conclusions and Discussion 43 Literature Cited 49 Chapter 13 52 Introduction 53 Methods 56 Results 64 Discussion 73 iii Conclusions 80 Literature Cited 82 Chapter 14 98 Introduction 99 Data and Methods 101 Results 106 Discussion 110 Conclusions 118 Literature Cited 119 Chapter 15 142 Introduction 143 Methods 146 Results and Discussion 150 Conclusions 164 Literature Cited 167 Papers in Preparation 179 Paper 1 179 Paper 2 182 Conclusions 186 Management or Policy Implications 188 List of Publications 189 In Preparation 189 Submitted/in press 189 Published 190 Outreach 193 Websites 193 Presentations in schools 193 Conference Presentations 194 Acknowledgements 199 67 68 69 70 iv 71 72 List of Tables Table 1. Table 2. Table 3. Table 4. Table 5. Table 6. 73 74 75 Study Chronology Design of Moorings Deployments at M2 Deployments at M4 Deployments at M5 Deployments at M8 1 5 6 7 8 9 List of Figures Figure 1. Location of the four biophysical moorings at the 70 m isobath. Figure 2. The annual Bering Sea Report Card from the Ecosystem Considerations Chapter, including 10 key indicators of the status of the Bering Sea. Figure 3. Depth averaged temperature measured at M2 during the last 18 years and the temperature anomaly, with the annual cycle (1995-2009) removed. Figure 4. Eastern Bering Sea and M2 study site where the acoustic instruments collected zooplankton volume backscatter data. Figure 5. ADCP-derived zooplankton volume backscatter. Top – 2007, Middle – 2009, Bottom – 2010. Figure 6. Mean currents during summer (May – September) derived from satellitetracked drifters drogued at 45 m. There are at least 10 independent estimates of velocity in each grid cell. Figure 7. Bottom currents at M2 divided into octents determined by wind directions (e.g. for 0 – 45 °T the vector is the average velocity when winds are toward that direction). Note currents are not 45 – 90 ° to the right of the winds (lower left). Number of points in each bin shown in lower right. Figure 8. Bottom currents at M5 divided into octents determined by wind directions (e.g. for 0 – 45 °T the vector is the average velocity when winds are toward that direction). Note currents are not 45 – 90 ° to the right of the winds (middle panel). Number of points in each bin shown in bottom. Figure 9. Bottom currents at M4 divided into octents determined by wind directions (e.g. for 0 – 45 °T the vector is the average velocity when winds are toward that direction). Note currents are not 45 – 90 ° to the right of the winds (middle panel). Number of points in each bin shown in bottom Figure 10. Schematic of the pathway for management action for walleye pollock 3 11 11 180 181 183 183 184 185 188 76 77 v 78 Study Chronology 79 This report documents the activities, analytical methods, results and findings of Project B52 of the 80 Bering Sea Integrated Ecosystem Research Program. The project started in 2008 and ended in April 81 2013. Relationships identified in this analysis were used to inform or parameterize FEAST (B70, 82 Aydin et al.). In addition, there were strong collaborations with several NSF funded components of 83 the Bering Sea Project (NSF Project 1107250, Synthesis, Mordy et al.; NSF Project 84 0732640 Hydrographic Structure and Nutrients over the Eastern Bering Sea Shelf During 85 Summer, Mordy et al.; NSF Project 0732430 Impacts of Sea-ice on Hydrographic Structure and 86 Nutrients over the Eastern Bering Sea Shelf, Mordy et al.; and NSF project 0732534, Downscaling 87 global climate projections, Bond et al.). There were no no-cost extensions to the program. Progress 88 reports were submitted biannually and can be found on the North Pacific Research Board website. 89 90 91 Table 1. Study Chronology 2008-2010 Tasks, Assignments, Timeline What Who/Ship Prep equipment, design and Stabeno, Napp, build moorings, develop Whitledge, Mordy cruise plans. (SNWM) Recover/deploy moorings NOAA ship Miller M2 and M4 (ice permitting) Freeman SNWM Process and disseminate SNWM data Data Delivery to data manager Send plankton samples to Napp Poland to be counted Deploy moorings at M5 and USCG Healy M8 SNM Recover/Deploy moorings Miller Freeman M2, M4, M5 and M8 and/or Melville SNWM Process and disseminate data Data Delivery to data manager Sent plankton samples to Poland to be counted Prepared equipment and built moorings Failed to deploy moorings at M2 SNWM Napp Oscar Dyson, Stabeno Dates 2008 Other information May 1 – May 10, 2008 Conducted CTD/bongos along southern hydro. line May 15-Nov 15 Nov. 2008 phys/chem. Sep 2009 zoop Jun 1, 2008 – Jun 1, 2009 July, 2008 Sep, 2008 Oct – Dec, 2008 March 2009 phys/chem Jan 2010 zoop Oct 1, 2008 – Oct 1, 2009 Jan – Feb. 2009 March 2009 The actual time for processing varies. CTD/bongos along the 5 primary hydro. lines (70m isobath, & 4 cross shelf transects) The actual time for processing varies. Failed because of ice and weather. 1 Submitted data to data manager Recover/deploy moorings M2 and M4 Processed and disseminated data Sent plankton samples to Poland to be counted Deploy moorings at M5 and M8 Prepared equipment and built moorings Recover/Deploy moorings M2, M4, M5 and M8 Processed and disseminated data Sent plankton samples to Poland Recover/deploy moorings M2 (over half the mooring was missing) Deployed moorings at M5, did not recover moorings at site. Oscar Dyson SNWM Apr 24 – May 7, 2009 SNWM May 15 – Sept 30, 2009 July 30, 2009 Napp Conducted some hydrographic stations Actual time for processing varies Jun/Jul 2009 R/V Knorr SW Stabeno Jun - Aug. 2009 Napp Sep 23 – Oct 11, 2009 Oct 2009– Apr, 2010 Nov 2009 Oscar Dyson SNWM Apr 23 – May3 , 2010 SNWM June 2010 Processed and disseminated data and data delivered to data manager Conducted hydro and zooplankton survey SNWM Apr – Sep 2010 Wecoma SNWM Aug 23 – Sep 10, 2010 Recover/Deploy moorings M2, M4, M5 and M8 Freeman SNWM Sep 23 – Oct 3, 2010 Processed and disseminated data and data delivered to data manager Sent plankton samples to Poland Synthesis of data and ideas, complete manuscripts 6 manuscripts were submitted and accepted for SNWM Oct 2010 – Jul 2011 Napp November 2010 SNWM 2010 - 2012 SNWM Oct – Dec 2011 Miller Freeman SNWM SNWM CTD/bongos along the 5 primary hydro lines Actual time for processing varies Due to extensive ice M4, M5 and M8 could not be reached. M4 could not be located and M5 was not recovered because of limited space on the boat. Conducted last sampling of 70-m isobath and 3 cross lines M4 could not be found. It was likely dragged by ice or a fishing vessel. Will continue searching for it on future cruises. Actual time for processing varies This is an ongoing task. 2 92 93 publication in Special issue of DSR II 6 manuscripts published 2 papers published in 2nd special issue of DSR II 8 manuscripts submitted for 3rd special issue >3 manuscripts being prepared for submission to 4th special issue of DSR II July 2012 In the out years, the prepping, mooring design and construction, and cruise plan development 94 occur during the 3 months before the beginning of the cruise. Similarly the data processing, 95 qcing etc. take approximately 3 months, with the exception of the counting of zooplankton which 96 can take as long as 1 year. Note, the delivery date of data is contingent upon the successful 97 recovery of the moorings. 98 99 Introduction 100 Project B52 is a continuation of a long-term partnership 101 between NOAA and NPRB. List of other NPRB grants 102 which supported the moorings are 203, 315, 410, 517, 103 602, 701, 1202. Moorings have been maintained on 104 the southeastern Bering Sea shelf at four sites: M2 105 (56.9°N, 164.1°W) since 1995, M4 (57.9°N, 168.9°W) 106 since 1996, M5 (59.9°N, 171.7°W) and M8 (62.2°N 107 174.7°W) since 2004 (Fig. 1). This project, together 108 with research by the NOAA program North Pacific 109 Climate 110 (NPCREP), continued these measurements through 111 2013. The moorings, together with observations along 112 114 the 70-m isobath, are core to the long-term observations Fig. 1. Location of the 4 biophysical moorings and the 70-m on the Bering Sea shelf. B52 continued the time series of isobath. temperature, salinity, fluorescence, currents, zooplankton 115 abundance (TAPS-8 at M2), nitrate, and oxygen (at M2) at four mooring sites on the Bering Sea 116 shelf. To avoid damage by ice, most of these moorings were subsurface. During the late spring and 117 summer a surface mooring was deployed at M2, which measured meteorological variables (air 118 temperature, humidity, barometric pressure, wind velocity and solar radiation – PAR). 113 Research and Ecosystem Productivity 3 119 This project supports all five BSIERP hypotheses, by providing critical data to identify long-term 120 physical, chemical, and biological changes in the ecosystem: 121 1. Physical forcing affects food availability: Climate-induced changes in physical forcing 122 will modify the availability and partitioning of food for all trophic levels through bottom- 123 up processes; 124 2. Ocean conditions structure trophic relationships: Climate and ocean conditions 125 influencing water temperature, circulation patterns, and domain boundaries impact fish 126 reproduction, survival and distribution, the intensity of predator-prey relationships, and 127 the location of zoogeographic provinces through bottom-up processes; 128 3. Ecosystem controls are dynamic: Later spring phytoplankton blooms resulting from 129 early ice retreat will increase zooplankton production, thereby leading to increased 130 abundances of piscivorous fish (walleye pollock, Pacific cod, and arrowtooth flounder) 131 and a community controlled by top-down processes with several trophic consequences; 132 4. Location matters: Climate and ocean conditions influencing circulation patterns and 133 domain boundaries will affect the distribution, frequency, and persistence of fronts and 134 other prey-concentrating features and, thus, the foraging success of marine birds and 135 mammals largely through bottom-up processes; 136 5. Commercial and subsistence fisheries reflect climate: Climate-ocean conditions will 137 change and, thus, affect the abundance and distribution of commercial and subsistence 138 fisheries. 139 140 Objectives 141 1) Provide near continuous, water column temperature, salinity, fluorescence, nitrate, and 142 currents at the four primary mooring sites, plus zooplankton and oxygen at selected sites. 143 Moorings were deployed at all four sites at least once a year. Below are the mooring designs 144 (Table 2) and the list of moorings deployed for each site (Tables 3 – 6), with the date of 145 deployment and recovery, the number of instruments on the moorings and any comments. 146 Hydrography and net tows were done at the mooring site and at the four corners of the 20 km x 147 20 km box around each mooring site, whenever possible. In spring, with extensive ice some of 148 these stations were skipped because of ice cover. 149 150 4 151 Table 2. The design of the moorings. The extensive sea ice precluded the recovery /deployment 152 of M5 and M8 in May of 2009. To resolve this, a simple mooring with temperature at 15, 10 and 153 5 m was deployed in July at M8 to obtain measurements in the upper column. A summer 154 mooring was also deployed at M5 in July when possible. See Tables 3-6 for details of moorings. 155 Sensors M4 M5 Summer & Winter 15, 20, 25, 30, 35, 40, 45, 50, 55, 60, 65, 69 M8 Summer & Winter 20, 25, 30, 35, 40, 45, 50, 55, 60, 65, 69 m Summer Winter Summer Winter Temperature 1, 5, 8, 10, 15, 20, 25, 30, 35, 40, 45, 50, 55, 60, 65, 69 5, 11, 15, 20, 25, 30, 35, 40, 45, 50, 55, 60, 65, 69 11 15 20, 25, 30, 35, 40, 45, 50, 55, 60, 65, 69 Salinity 11, 35, 60 11, 35, 60 11, 35, 60 15, 35, 60 20, 35, 60 Nitrate Fluorescence 12, 59 11, 22, 44 11, 15, 20, 25, 30, 35, 40, 45, 50, 55, 60, 65, 69 11, 35, 60 12, 59 11, 22, 44 11 11 15 20 CDOM Zooplankton biovolume,size composition Currents ADCP Meteorological variables 156 157 M2 Bottom depth 11 m 35 35 every 4 m every 4m every 4m every 4 m every 4 m every 4 m 73 70 70 71 70 surface 73 158 5 159 Table 3. Moorings were deployed twice a year at M2. The first set of moorings in 2008, replaced 160 moorings, which had been deployed the previous September. Moorings indicated by “bsm” were 161 all surface moorings. Those indicated by “bst” contained the TAPS-8 and selection of other 162 instruments. Selected data on bst and bsm were reported back in real time and put on the web. 163 Those indicated by “bsp” are bottom-mounted, upward looking ADCP moorings. Moorings 164 indicated by just “bs” were overwintering moorings with top instrument at 11 m. Mooring Deployed Recovered M2 Number of Comments Instr. 08bsm2a 5/6/2008 9/28/2008 26 07bs2c deployed on Sept. 2007 and recovered May 7, 2008 with 15 instruments 08bsp2a 5/6/2008 Not recovered 1 07bsp2c deployed on Sept. 2007 and recovered May 7, 2008 with 15 instruments 08bs2c 9/29/2008 4/30/2009 15 08bsp2c 9/29/2008 4/30/2009 11 09bsm2a 4/30/2009 9/25/2009 22 09bst2a 4/30/2009 9/24/2009 7 09bsp2a 4/30/2009 9/25/02009 1 09bs2c 9/26/2009 4/30/2010 4 09bsp2b 9/26/2009 4/27/2010 1 10bsm2a 5/12/2010 10/4/2010 21 10bst2a 4/29/2010 10/3/2010 4 10bsp2a 4/30/2010 10/4/2010 1 10bs2c 10/5/2010 5/19/2011 18 10bsp2b 10/5/2010 5/19/2011 1 The top of this mooring was trawled and we only recovered the bottom 4 instruments. Extensive ice required the 2 week delay of deployment of surface mooring 165 166 6 167 Table 4. Moorings were generally deployed twice a year at M4. The first set of moorings in 168 2008, replaced moorings ,which had been deployed the previous September. Moorings indicated 169 by “bsm” were all surface moorings. Those indicated by “bsp” are upward looking ADCP 170 moorings. Moorings indicated by just “bs” were overwintering moorings with top instrument at 171 11 m. Mooring 09bsp4b was not recovered and all instruments and data were lost. 172 Mooring Deployed Recovered M4 Number of Comments Instr. 5/9/2008 5/2/2009 14 07bs4b deployed on Sept. 2007 and recovered May 7, 2008 with 15 instruments 5/9/2008 9/26/2008 1 08bsp4a 08bs4b 07bsp4b deployed on Sept. 2007 and recovered May 7, 2008 with 1 instruments 9/26/2008 5/2/2009 13 08bsp4b 9/26/2008 5/2/2009 1 09bsm4a 5/2/2009 8/18/2009 19 09bsp4a 5/2/2009 9/27/2009 2 09bs4b 9/27/2009 Not recovered 21 09bsp4b 9/2872009 9/24/2010 1 08bs4a This surface mooring was dragged by ice and had to be recovered early. 10bs4a, Due to very extensive and persistent ice, these moorings were not deployed. The 10bsp4a previous fall deployments were expected to supply necessary data. 10bs4b 9/24/2010 5/20/2011 17 10bsp4b 9/24/2010 5/20/2011 2 173 174 7 175 Table 5. Moorings were generally deployed once a year at M5. The first set of moorings in 2008 176 replaced moorings, which had been deployed the previous September. Those indicated by “bsp” 177 are upward looking ADCP moorings. Moorings indicated by just “bs” were overwintering 178 moorings with top instrument at 15 m. 179 Mooring M5 Deployed Recovered 9/25/2008 5/29/2009 Number of Instr. 12 9/25/2008 5/29/2009 1 08bsp5b 09bs5a 7/1/2009 9/29/2009 15 09bsp5a 09bs5b 09bsp5b 10bs5a 10bsp5a 10bs5b 10bsp5b 5/29/2009 9/29/2009 9/29/2009 7/4/2010 7/4/2010 9/27/2010 9/27/2010 9/29/2009 9/26/2010 9/26/2010 9/26/2010 9/26/2010 5/20/2011 5/20/2011 1 13 1 18 1 15 1 08bs5b 180 181 Comments 07bs5b was deployed in Sept. 2007 and recover on 9/25/08 with 11 instruments 07bsp5b was deployed in Sept. 2007 and recover on 9/25/08 with 1 instrument Weather and time constraints prevented the deployment of full water column mooring in May. Ice prevented recovery in May. Ice prevented deployment in May 8 182 Table 6. Moorings were generally deployed once a year at M8, with a second mooring with 183 thermistors in the upper 20 m deployed in July. The first set of moorings in 2008 replaced 184 moorings, which had been deployed the previous September. Those indicated by “bsp” are 185 upward looking ADCP moorings. Moorings indicated by “bsv” only contained temperature 186 measurements in the upper 20 m. Moorings indicated by just “bs” were overwintering moorings 187 with top instrument at 20 m. 188 Mooring M8 Deployed Recovered 7/26/2008 8/31/2008 Number of Instr. 2 8/31/2008 10/1/2009 11 8/31/2008 10/1/2009 1 7/7/2009 10/1/2009 10/1/2009 9/29/2010 9/29/2010 10/1/2009 9/29/2010 9/29/2010 8/16/2011 8/16/2011 11 2 13 1 08bsv8a 08bs8a 189 08bsp8a 09bsv8a 09bs8a 09bsp8a 10bs8a 10bsp8a Comments This simple mooring was deployed to obtain near-surface temperatures. 07bs8a was deployed in Sept. 2007 and recover on 8/31/08 with 11 instruments 07bsp8a was deployed in Sept. 2007 and recover on 8/31/08 with 1 instruments 190 191 2) Provide derived products: mixed layer depth, timing of the spring phytoplankton bloom and 192 other blooms, heat content, mixed layer depth, and advection at the four sites. 193 Products from the moorings include mixed layer depth, heat content at M2 and M4, temperature, 194 position of the transition between southern pelagic-dominated shelf and northern benthic- 195 dominated shelf, advection, tidal energy, nitrate concentration, the timing of the spring 196 phytoplankton bloom, estimates of zooplankton from TAPS-8 and backscatter from the ADCPs. In 197 addition, a number of annual cycles were derived using the mooring data, including: total heat 198 content (depth averaged temperature) at M2 and M4, vertical temperature structure at M2, bottom 199 and near-surface temperature at each mooring site, mean currents (near surface and bottom) at 200 each site. A number of derived indices of ice cover were develop to better understand the 201 response of the water column and lower trophic level to sea ice. 202 9 203 3) Use mooring and shipboard measurements as indicators of ecosystem status and to apply the 204 observations to metrics/indices developed during Southeast Bering Sea Carrying Capacity 205 (SEBSCC). Possible indices include wind mixing, ocean stratification, mixing events (strength, 206 date, number of), mixed layer temperature, photosynthetically active radiation, sea ice 207 characteristics, extent of the cold pool, timing of the spring phytoplankton bloom, zooplankton 208 biomass and estimated larval advection. 209 EcoFOCI contributes to the Ecosystem Status and Management Indicators section of the 210 Ecosystem Considerations Chapter. Our mooring and shipboard measurements contribute to the 211 following indicators: 212 213 214 215 216 217 1. North Pacific Climate Overview, Bering Sea Regional Highlights contributed by N. Bond. 2. Eastern Bering Sea Climate – FOCI contributed by J. Overland, P. Stabeno, C. Ladd, S. Salo, M. Wang and N. Bond. 3. Bering Sea Zooplankton, contributed by J. Napp, P. Ressler, P. Stabeno, and A. Yamaguchi 218 219 220 EcoFOCI also contributes indicators to the annual Eastern Bering Sea Report Card (Fig. 2). 221 averaged temperature at M2 and the derived anomaly (e.g. Fig. 3) are used to define warm and 222 cold years for the eastern Bering Sea shelf. The chlorophyll-a fluorescence time series at all the 223 moorings provides an index of the timing of the spring and fall phytoplankton blooms, which can 224 be related to both ice-retreat and storminess. The vertical temperature profiles at the moorings 225 provide indicators of mixed layer depth. Sudden increases in mixed-layer depth coupled with 226 winds and fluorescence time series provides an index of how many events during the summer 227 were strong enough to mix nutrients into the euphotic zone. 228 Mooring M8 is located in Region 1 of the Distributed Biological Observatory (DBO). Data from 229 this site provide a range of statistics to characterize that region and how it varies from year to 230 year. Information includes: minimum and maximum bottom temperature, timing of that site 231 becoming well mixed, strength of vertical density gradient, and direction of currents. Long-term mooring data can be used to create a number of indices. For instance the depth 232 10 Fig. 2. The annual Bering Sea Report Card from the Ecosystem Considerations Chapter, including 10 key indicators of the status of the Bering Sea. Ocean temperature is not one of the items because of its high correlations with sea ice extent in the spring. The copepod index comes partially from data collected as part of this program. 233 234 235 236 237 238 239 240 241 242 4) 243 Use 244 data Fig. 3. Top panel: Depth averaged temperature measured at M2 during the last 18 years. Bottom Panel: The temperature anomaly, with the annual cycle (1995 – 2009) removed. 245 from the moorings and shipboard observations to validate and tune the physical and nutrient- 11 246 phytoplankton-zooplankton (NPZ) models that are being developed. 247 The data from the biophysical mooring data were frequently used to calibrate biophysical 248 models of the Bering Sea. These included side-by-side model/data comparisons of the vertical 249 profiles of currents, temperature, salinity, and chlorophyll-a at multiple locations, from hourly 250 to interdecadal time scales. The data were especially useful in demonstrating that initial 251 parameter choices for the model produced unrealistically warm conditions in the Bering Sea. A 252 time series of surface shortwave radiation at M2 demonstrated that shortwave forcing was in 253 fact *not* the source of that bias. Subsequent adjustments to other factors – albedo and bulk 254 flux parameters in particular - yielded a much better fit to the extensive mooring dataset. The 255 data were especially powerful in providing multivariate time series at many locations. These 256 allow the examination of the covariance structure among different locations, and among 257 different variables, which are compared with their model equivalent. Note that this is a far 258 more rigorous method of model-data comparison than is possible with any single time series. 259 260 Manuscripts 261 There are ~30 peer reviewed publications (published, submitted and in preparation) as results of 262 this grant. All published manuscripts are listed with the link to the appropriate web pages from 263 which they can be down loaded and the abstract. The complete texts of manuscripts that are 264 submitted, but not published are presented in Chapters 12 –15. Finally, two manuscripts that are 265 presently in preparation for the fourth special issue are presented with detail currently available. 266 267 12 268 269 270 271 272 273 274 275 276 277 278 279 280 281 282 283 Published Manuscripts 284 (alphabetized by first author) 285 13 286 Chapter 1 287 288 Danielson, S., E. Curchitser, K. Hedstrom, T. Weingartner, and P. Stabeno (2011): On ocean and 289 sea ice modes of variability in the Bering Sea. J. Geophys. Res., 116(C12), C12034, doi: 290 10.1029/2011JC007389. 291 292 http://onlinelibrary.wiley.com/doi/10.1029/2011JC007389/full 293 294 ABSTRACT: Results from a 35 year hindcast of northeast Pacific Ocean conditions are 295 confronted with observational data collected over the Bering Sea shelf within the integration time 296 period. Rotary power spectra of the hindcast currents near NOAA mooring site M2 site fall within 297 the 95% confidence bounds for the observational spectra, except for a high bias in the counter- 298 clockwise rotating component at 10 m depth in the high frequencies (periods <24 h). The model 299 exhibits the most skill in reproducing anomalies of the integrated annual sea ice concentration and 300 monthly subsurface (60 m depth) temperature fields, accounting for 85% and 50% of their 301 observed variability. Analysis of the integrated ice concentration time series reveals evolution in 302 the mean duration of ice-free waters (40 year trend of +6.8 days/decade) and changes in this 303 parameter's variance with time. Correlation and empirical orthogonal function (EOF) analyses 304 reveal the primary temporal-spatial patterns of variability in the temperature and salinity fields 305 over the Bering Sea and northern Gulf of Alaska for near-surface (0–20 m) and subsurface (40– 306 100 m) depth layers. Correlation analysis between the EOF principal components and various 307 climate index and observed time series shows that the Pacific Decadal Oscillation, the North 308 Pacific Gyre Oscillation, and the Bering Sea annually integrated ice area anomalies are important 309 indices of thermohaline variability; the spatial structures of these modes give insight to their 310 potential impacts upon the ecosystem. We identify a number of ecologically and economically 311 important species whose temporal variability is significantly correlated with the identified spatial 312 patterns. 313 314 14 315 Chapter 2 316 317 Hermann, A.J., G.A. Gibson, N.A. Bond, E.N. Curchitser, K. Hedstrom, W. Cheng, M. Wang, 318 P.J. Stabeno, L. Eisner, and K.D. Cieciel (2013): A multivariate analysis of observed and 319 modeled biophysical variability on the Bering Sea shelf: Multidecadal hindcasts (1970-2009) and 320 forecasts (2010-2040). Deep-Sea Res. II, doi: 10.1016/j.dsr2.2013.04.007. 321 322 http://www.sciencedirect.com/science/article/pii/S096706451300146X 323 324 ABSTRACT: Coupled physical/biological models can be used to downscale global climate 325 change to the ecology of subarctic regions, and to explore the bottom-up and top-down effects of 326 that change on the spatial structure of subarctic ecosystems—for example, the relative dominance 327 of large vs. small zooplankton in relation to ice cover. Here we utilize a multivariate statistical 328 approach to extract the emergent properties of a coupled physical/biological hindcast of the 329 Bering Sea for years 1970–2009, which includes multiple episodes of warming and cooling (e.g. 330 the recent cooling of 2005–2009), and a multidecadal regional forecast of the coupled models, 331 driven by an IPCC global model forecast of 2010–2040. Specifically, we employ multivariate 332 empirical orthogonal function (EOF) analysis to derive the spatial covariance among physical and 333 biological timeseries from our simulations. These are compared with EOFs derived from spatially 334 gridded measurements of the region, collected during multiyear field programs. The model 335 replicates observed relationships among temperature and salinity, as well as the observed inverse 336 correlation between temperature and large crustacean zooplankton on the southeastern Bering Sea 337 shelf. Predicted future warming of the shelf is accompanied by a northward shift in both pelagic 338 and benthic biomass. 339 340 15 341 Chapter 3 342 343 Hunt, Jr., G.L., B.M. Allen, R.P. Angliss, T. Baker, N. Bond, G. Buck, G.V. Byrd, K.O. Coyle, 344 A. Devol, D.M. Eggers, L. Eisner, R.A. Feely, S. Fitzgerald, L.W. Fritz, E.V. Gritsay, C. Ladd, 345 W. Lewis, J. Mathis, C.W. Mordy, F. Mueter, J. Napp, E. Sherr, D. Shull, P. Stabeno, M.A. 346 Stepanenko, S. Strom, and T.E. Whitledge (2010): Status and trends of the Bering Sea region, 347 2003-2008. In Marine Ecosystems of the North Pacific Ocean, 2003-2008, S. M. McKinnell and 348 M. J. Dagg (eds.), PICES Special Publication 4, 196-267. 349 350 http://www.pices.int/publications/special_publications/NPESR/2010/NPESR_2010.aspx 351 352 353 HIGHLIGHTS: o The Bering Sea experienced four years with low sea ice cover and extraordinarily warm 354 summers (2002-2005), followed by four years with some of the heaviest sea ice cover 355 since the early 1970s and cold summers (2006-2009). During the warm period, integrated 356 water column temperatures were elevated, bottom temperatures were higher, and the cold 357 pool over the southeastern shelf was small and not as cold as in the cold period. During the 358 cold period, integrated water column temperatures were anomalously low, bottom 359 temperatures were below the long term mean, and the cold pool consisted of cold, -1.7 .C 360 water that extended across most of the Middle Shelf Domain with cool waters extending to 361 Bristol Bay and the Alaska Peninsula. 362 o 363 364 Water column stratification varied spatially, and was sometimes stronger in the warm years and sometimes stronger in the cold years, depending upon location. Net primary production and surface chlorophyll a were positively affected by temperature. 365 The size distribution of crustacean zooplankton became smaller in years of warmer 366 temperature. 367 In the warm years of 2002-2005, small neritic species of crustacean zooplankton thrived 368 whereas the medium-large copepod, Calanus marshallae, and the shelf euphausiid, 369 Thysanoessa raschii, were scarce. In the cold years of 1999 and 2006- 2008, both C. 370 marshallae and T. raschii were abundant. 371 Catches of both eastern and western Bering Sea groundfish stocks declined in recent years. 372 In the eastern Bering Sea and Aleutian Islands, Pacific cod, yellowfin sole, flathead sole 373 and Greenland turbot, and particularly walleye pollock, have shown declines while Pacific 374 ocean perch, northern rockfish, rock sole, Alaska plaice, and especially arrowtooth 16 375 flounder (whose biomass has quadrupled since the late 1970s) have increasing biomass 376 trends. 377 Walleye pollock recruitment may be adversely affected by unusually warm conditions; 378 strong year-classes failed to emerge in the warm years of 2002-2005 despite a declining 379 biomass of pollock. Possibly, there were strong year-classes in the cold years of 2006 and 380 2008. In the cold years of 2006-2008, age-0 pollock were in good condition and energy- 381 rich, though not as abundant as they were in the warmer years, when they were less 382 energy-rich. 383 Age-0 pollock consumed mostly small crustacean zooplankton and smaller pollock in the 384 warmer years, and neither C. marshallae nor T. raschii were important dietary 385 components. In the cold years, their diets were dominated by C. marshallae and 386 euphausiids, and there was less cannibalism. 387 Comparison of fisheries statistics between the eastern and western Bering Sea showed 388 that, overall, the aggregated catches of all major fisheries tended to be positive, suggesting 389 that the two sides of the Bering Sea were responding similarly to shared climate forcing. 390 391 392 Both size-at-age and weight-at-length of groundfish tended to be above the long term mean in the warmer years, and below the mean in the cold years. Catches of crabs in the eastern and western Bering Sea declined after the early 1990s 393 (eastern Bering Sea) or the late 1990s (western Bering Sea), but catches began to increase 394 on both sides of the Bering Sea after 2004. The biomass of both red king crab and snow 395 crab have increased in the eastern Bering Sea, although snow crab catches remain low due 396 to conservative catch limits. 397 398 399 Catches of Pacific herring on both sides of the Bering Sea have been stable since 2001, whereas the Togiak stock in the eastern Bering Sea has declined. Catches of salmon in both the eastern and western Bering Sea were above average and 400 dominated by pink and chum salmon in the western Bering Sea and sockeye salmon in the 401 eastern Bering Sea. In both the eastern and western Bering Sea, the catches of chinook 402 salmon are in decline when compared to catches from the mid-1960s to the early 1990s. 403 404 17 405 406 407 Chapter 4 408 Sea Res. II, 65–70, doi: 10.1016/j.dsr2.2012.02.009, 72–83. Ladd, C., and P.J. Stabeno (2012): Stratification on the Eastern Bering Sea Shelf revisited. Deep- 409 410 http://www.sciencedirect.com/science/article/pii/S0967064512000136 411 412 ABSTRACT: The timing and magnitude of stratification can have profound influences on the 413 marine ecosystem. On the Eastern Bering Sea shelf, in the absence of strong wind mixing, 414 stratification can be initiated by the melting of seasonal sea ice or by springtime warming of the 415 surface. Temperature and salinity both influence the stratification of the Eastern Bering Sea shelf 416 with their relative importance varying spatially and temporally. In the northern middle shelf 417 domain (north of ∼60°N), salinity stratification is often as important as temperature stratification. 418 On the southern middle shelf, while the influence of temperature on stratification dominates 419 during summer, the influence of salinity stratification plays a role in the interannual variability. 420 Mooring 2 (M2; 56.9°N, 164.1°W) has been deployed at ∼70 m depth in the southern middle 421 shelf domain since 1995. Data from this mooring show that stratification typically begins to set 422 up in May and to break down in September/October, but these dates can vary by >30 d. While no 423 trend is found in the timing of the spring setup, the fall stratification breakdown exhibited a trend 424 toward later breakdown (∼2 d later per year from 1996 to 2009). Results suggest that it may be 425 difficult to forecast stratification on the Eastern Bering Sea shelf from climate models as simple 426 indices of wind mixing or heat fluxes are not correlated with stratification. Contrary to intuition, 427 the strength of summer stratification is not correlated with depth averaged temperature. Warm 428 years such as 2000 and 2001 can have low stratification and cold years such as 2007 can have 429 very high stratification. This decoupling of stratification and temperature has implications for 430 forecasting the ecosystem in the face of climate change, as we cannot assume that projections of a 431 warmer climate simply imply higher stratification in the future. 432 433 18 434 Chapter 5 435 436 Miksis-Olds, J.L., P.J. Stabeno, J.M. Napp, A.I. Pinchuk, J.A. Nystuen, J.D. Warren, and S.L. 437 Denes (2013): Ecosystem response to a temporary sea ice retreat in the Bering Sea: Winter 2009. 438 Prog. Oceanogr., 111, doi: 10.1016/j.pocean.2012.10.010, 38–51. 439 440 http://www.sciencedirect.com/science/article/pii/S0079661112001590 441 442 ABSTRACT: Adding acoustic systems onto ocean moorings and observatories provides 443 additional data to more fully document ecosystem responses to environmental perturbations. A 444 passive acoustic recorder and three-frequency echosounder system were integrated into a 445 biophysical mooring on the central eastern Bering Sea continental shelf. An unexpected, 446 transient, mid-winter retreat of the seasonal sea ice was observed over the mooring for a 2-week 447 period in March 2009. Interpretation of the passive acoustic data provided information about sea 448 ice conditions and included the detection and identification of vocalizing marine mammals, while 449 the acoustic backscatter provided information on relative zooplankton and fish abundance before, 450 during, and after the retreat. Hydrographic data confirmed the acoustic signal was associated with 451 changing surface ice conditions, and the combined information from the biophysical mooring 452 sensors revealed changes in winter trophic level dynamics during the retreat, which would have 453 otherwise been undetected by traditional ship-based observations. Changes in the acoustic 454 environment, zooplankton dynamics, and acoustic detection of marine mammals were observed 455 amidst a physically stable and uniform water column with no indication of a phytoplankton 456 bloom. These data demonstrate the value of acoustic technologies to monitor changing 457 ecosystems dynamics in remote and hazardous locations. 458 459 19 460 Chapter 6 461 462 Mordy, C.W., E.D. Cokelet, C. Ladd, F.A. Menzia, P. Proctor, P.J. Stabeno, and E. Wisegarver 463 (2012): Net community production on the middle shelf of the Eastern Bering Sea. Deep-Sea Res. 464 II, 65–70, doi: 10.1016/j.dsr2.2012.02.012, 110–125. 465 466 http://www.sciencedirect.com/science/article/pii/S0967064512000264 467 468 ABSTRACT: To estimate temporal changes of nutrients and calculate the seasonal net 469 community production (NCP) on the eastern shelf of the Bering Sea, hydrographic sampling 470 along the 70-m isobath of the middle shelf was conducted in spring (2007–2009), summer (2008– 471 2009), and fall (2007). These were cold years, with sea ice covering much of the eastern Bering 472 Sea in April. Each spring, there was a region with relatively low nitrate in the middle portion 473 (59°–60°N) of the transect prior to the spring phytoplankton bloom. This water appeared to have 474 originated in the coastal domain and was advected offshore into the middle domain. Seasonal 475 NCP (mean±standard deviation) in this region was low (26±12 g C m−2), and may be indicative of 476 a portion of the middle shelf ecosystem that is chronically short of fixed carbon in spring. In 477 2007, the post-bloom cruise occurred during the fall transition when deep mixing, 478 remineralization, and denitrification/anammox compromised seasonal estimates of NCP. In other 479 years (2008–2009), the post-bloom cruise occurred in summer. On those cruises, the euphotic 480 zone, elevated chlorophyll fluorescence, and oxygen supersaturation were occasionally deeper 481 than the pycnocline, and there was a seasonal loss of nitrate and phosphate in the bottom layer. In 482 2008, preferential uptake of ammonium may have sustained sub-surface production in the north. 483 Therefore, seasonal estimates of NCP were not only evaluated in the upper mixed layer, but 484 throughout the water column. During summer, denitrification/anammox in bottom waters did not 485 appear to compromise seasonal estimates of NCP. Seasonal NCP averaged for 2008 and 2009 486 was slightly but significantly higher (p<0.0041) in the south (47±9 g C m−2, n=80) than in the 487 north (41±16 g C m−2, n=78). In the south, interannual variability of seasonal NCP was related to 488 the wind mixing in spring rather than the presence or absence of ice. 489 490 20 491 Chapter 7 492 493 Nystuen, J.A., S.E. Moore, and P.J. Stabeno (2010): A sound budget for the southeastern Bering 494 Sea: Measuring wind, rainfall, shipping, and other sources of underwater sound. J. Acoust. Soc. 495 Am., 128(1), doi: 10.1121/1.3436547, 58−65. 496 497 http://scitation.aip.org/content/asa/journal/jasa/128/1/10.1121/1.3436547 498 499 ABSTRACT: Ambient sound in the ocean contains quantifiable information about the marine 500 environment. A passive aquatic listener (PAL) was deployed at a long-term mooring site in the 501 southeastern Bering Sea from 27 April through 28 September 2004. This was a chain mooring 502 with lots of clanking. However, the sampling strategy of the PAL filtered through this noise and 503 allowed the background sound field to be quantified for natural signals. Distinctive signals 504 include the sound from wind, drizzle and rain. These sources dominate the sound budget and their 505 intensity can be used to quantify wind speed and rainfall rate. The wind speed measurement has 506 an accuracy of ±0.4 m s−1 when compared to a buoy-mounted anemometer. The rainfall rate 507 measurement is consistent with a land-based measurement in the Aleutian chain at Cold Bay, AK 508 (170 km south of the mooring location). Other identifiable sounds include ships and short 509 transient tones. The PAL was designed to reject transients in the range important for 510 quantification of wind speed and rainfall, but serendipitously recorded peaks in the sound 511 spectrum between 200 Hz and 3 kHz. Some of these tones are consistent with whale calls, but 512 most are apparently associated with mooring self-noise. 513 514 21 515 Chapter 8 516 517 Sigler, M.F., H.R. Harvey, C.J. Ashjian, M.W. Lomas, J.M. Napp, P.J. Stabeno, and T.I. Van Pelt 518 (2010): How does climate change affect the Bering Sea ecosystem? Eos Trans. AGU, 91(48), doi: 519 10.1029/2010EO480001, 457–458. 520 521 http://onlinelibrary.wiley.com/doi/10.1029/2010EO480001/abstract 522 ABSTRACT: The Bering Sea is one of the most productive marine ecosystems in the world, 523 sustaining nearly half of U.S. annual commercial fish catches and providing food and cultural 524 value to thousands of coastal and island residents. Fish and crab are abundant in the Bering Sea; 525 whales, seals, and seabirds migrate there every year. In winter, the topography, latitude, 526 atmosphere, and ocean circulation combine to produce a sea ice advance in the Bering Sea 527 unmatched elsewhere in the Northern Hemisphere, and in spring the retreating ice; longer 528 daylight hours; and nutrient-rich, deep-ocean waters forced up onto the broad continental shelf 529 result in intense marine productivity (Figure 1). This seasonal ice cover is a major driver of 530 Bering Sea ecology, making this ecosystem particularly sensitive to changes in climate. Predicted 531 changes in ice cover in the coming decades have intensified concern about the future of this 532 economically and culturally important region. In response, the North Pacific Research Board 533 (NPRB) and the U.S. National Science Foundation (NSF) entered into a partnership in 2007 to 534 support the Bering Sea Project, a comprehensive $52 million investigation to understand how 535 climate change is affecting the Bering Sea ecosystem, ranging from lower trophic levels (e.g., 536 plankton) to fish, seabirds, marine mammals, and, ultimately, humans. The project integrates two 537 research programs, the NSF Bering Ecosystem Study (BEST) and the NPRB Bering Sea 538 Integrated Ecosystem Research Program (BSIERP), with substantial in-kind contributions from 539 the U.S. National Oceanic and Atmospheric Administration (NOAA) and the U.S. Fish and 540 Wildlife Service. 541 542 22 543 544 545 Chapter 9 546 and M.F. Sigler (2012): A comparison of the physics of the northern and southern shelves of the 547 eastern Bering Sea and some implications for the ecosystem. Deep-Sea Res. II, 65–70, doi: 548 10.1016/j.dsr2.2012.02.019, 14–30. Stabeno, P.J., E. Farley, N. Kachel, S. Moore, C. Mordy, J.M. Napp, J.E. Overland, A.I. Pinchuk, 549 550 http://www.sciencedirect.com/science/article/pii/S0967064512000331 551 552 ABSTRACT: Sufficient oceanographic measurements have been made in recent years to 553 describe the latitudinal variation in the physics of the eastern Bering Sea shelf and the potential 554 impact of climate change on the species assemblages in the two ecosystems (north and south). 555 Many of the predicted ecosystem changes will result from alterations in the timing and extent of 556 sea ice. It is predicted that the sea ice in the northern Bering Sea will be less common in May, but 557 will continue to be extensive through April. In contrast, the southern shelf will have, on average, 558 much less sea ice than currently observed, but with large interannual and multiyear variability 559 until at least 2050. Thus, even under current climate warming scenarios, bottom temperatures on 560 the northern shelf will remain cold. Based on biophysical measurements, the southern and 561 northern ecosystems were divided by a North–South Transition at ∼60°N. The northern middle 562 shelf was characterized by a freshwater lens at the surface, cold bottom temperatures, and a 563 thicker pycnocline than found on the southern shelf. Subsurface phytoplankton blooms were 564 common. In contrast, the southern shelf stratification was largely determined by temperature 565 alone; the pycnocline was thin (often<3 m) and subsurface blooms were uncommon. Biological 566 responses to climate warming could include greater north–south differences in zooplankton 567 community structure, the transport of large Outer Shelf Domain crustacean zooplankton to the 568 middle shelf, and the disappearance of two principal prey taxa (Calanus spp. and Thysanoessa 569 spp.) of planktivorous fish, seabirds and whales. The response of commercially and ecologically 570 important fish species is predicted to vary. Some species of fish (e.g., juvenile sockeye salmon, 571 Oncorhynchus nerka) may expand their summer range into the northern Bering Sea; some (e.g., 572 pink salmon, O. gorbuscha) may increase in abundance while still other species (e.g., walleye 573 pollock and arrowtooth flounder; Theragra chalcogramma and Atheresthes stomias, respectively) 574 are unlikely to become common in the north. The projected warming of the southern shelf will 575 limit the distribution of arctic species (e.g., snow crab, Chionoecetes opilio) to the northern shelf 576 and will likely permit expansion of subarctic species into the southern Bering Sea. The 23 577 distribution and abundance of baleen whales will respond to shifts in prey availability; for 578 instance, if prey are advected northward from the southeastern Bering Sea, an extension of range 579 and an increase in seasonally migratory baleen whale numbers is anticipated. Thus, alteration of 580 this ecosystem in response to climate change is expected to result in something other than a 581 simple northward shift in the distribution of all species. 582 583 24 584 Chapter 10 585 586 Stabeno, P.J., N.B. Kachel, S.E. Moore, J.M. Napp, M. Sigler, A. Yamaguchi, and A.N. Zerbini 587 (2012): Comparison of warm and cold years on the southeastern Bering Sea shelf and some 588 implications for the ecosystem. Deep-Sea Res. II, 65–70, doi: 10.1016/j.dsr2.2012.02.020, 31–45. 589 590 http://www.sciencedirect.com/science/article/pii/S0967064512000343 591 592 ABSTRACT: The southeastern, middle shelf of the Bering Sea has exhibited extreme variability 593 in sea ice extent, temperature, and the distribution and abundance of species at multiple trophic 594 levels over the past four decades. From 1972–2000, there was high interannual variability of areal 595 extent of sea ice during spring (March–April). In 2000, this shifted to a 5-year (2001–2005) 596 period of low ice extent during spring, which transitioned to a 4-year (2007–2010) period of 597 extensive sea ice. High (low) areal extent of sea ice in spring was associated with cold (warm) 598 water column temperatures for the following 6–7 months. The ocean currents also differed 599 between warm and cold years. During cold years, the monthly-mean currents over the shelf were 600 largely westward, while in warm years the direction of currents was more variable, with 601 northward flow during December–February and relatively weak flow during the remainder of the 602 year. The types and abundance of zooplankton differed sharply between warm and cold years. 603 This was especially true during the prolonged warm period (2001–2005) and cold period (2007– 604 2010), and was less evident during the years of high interannual variability. During the warm 605 period, there was a lack of large copepods and euphausiids over the shelf; however, their 606 populations rebounded during cold period. Small crustacean zooplankton taxa did not appear to 607 vary between and warm and cold years. For both walleye pollock and Pacific cod, year-class 608 strength (recruitment) was low during the prolonged warm period, but improved during the 609 following cold period. Year-class strength did not appear to vary as a function of warm and cold 610 years during the period of high year-to-year variability. Also, recruitment of arrowtooth flounder 611 (a predator of pollock and cod) did not appear influenced by the warm or cold years. Finally, the 612 distribution and relative abundance of fin whales appeared to differ in warm and cold years, with 613 fewer whales on the southeastern, middle shelf during warm years. 614 615 25 616 Chapter 11 617 Stafford, K.M., S.E. Moore, P.J. Stabeno, D.V. Holliday, J.M. Napp, and D.K. Mellinger (2010): 618 Biophysical ocean observation in the southeastern Bering Sea. Geophys. Res. Lett., 37, L02606, 619 doi: 10.1029/2009GL040724. 620 621 http://onlinelibrary.wiley.com/doi/10.1029/2009GL040724/full 622 623 ABSTRACT: Integrated ocean observation, from physical and atmospheric forcing mechanisms 624 to the distribution and abundance of top-level predators, is critical to the investigation of marine 625 ecosystems and the impact of climate change on them. We integrated data from a biophysical 626 mooring in the southeast Bering Sea to create a one-year snapshot of ocean dynamics in this 627 remote large marine ecosystem. Distinct patterns in production (chlorophyll), zooplankton 628 biovolume (copepods and euphausiids) and the occurrence of zooplankton predators (fin and 629 right whales) were defined and related to discrete features in the annual physical cycle. Peaks in 630 prey and predator cycles were linked to spikes in fluorescence that occurred at the onset of water 631 column stratification in late spring 2006 and the appearance of sea ice in late winter 2007. These 632 data illustrate the capability and potential of integrated ocean observing systems (IOOS) to 633 describe seasonal variability and linkages in a remote marine ecosystem. 634 635 636 637 26 638 639 640 641 642 643 644 645 646 647 648 649 650 651 652 653 654 Submitted manuscripts 27 655 Chapter 12 656 657 Cheng, W., E. Curchitser, C. Ladd, and P.J. Stabeno (2013): Ice–ocean interactions in the eastern 658 Bering Sea: NCAR CESM simulations and comparison with observations. Deep-Sea Res. II. 659 660 Abstract 661 We examine ice-ocean interactions in the Eastern Bering Sea (EBS) using output from the 662 National Center for Atmospheric Research-Community Earth System Model and satellite 663 observations, and draw comparisons with published results based on ocean in situ data. The 664 model simulation generally captures the observed seasonal advance and retreat of sea ice in the 665 EBS and spatial patterns. In the northern EBS, however, modeled sea ice annual maximum occurs 666 in April instead in March, as observed by satellite observations. Sea ice also retreats slower in 667 spring in the simulations than observed. Simulations indicate that early winter changes in ice 668 coverage in the north are due to a combination of freezing and advection processes, while in the 669 south changes are primarily due to advection from the north and melting. Via this seasonal transit, 670 sea ice cools the entire EBS middle shelf relatively uniformly, but its influence on ocean salinity 671 has greater spatial variability, as suggested by both observations and simulations. On inter-annual 672 time scales, high ice cover in the EBS leads to cold anomalies in the bottom water, especially on 673 the middle shelf, with the strongest anomalies in the southern domain. The corresponding salinity 674 anomalies are positive, with the strongest signal in the northern near-shore locations. The 675 associated current anomalies at 50m are generally southward on the shelf, and northwestward in 676 the slope region. Comparing years 1961-2005 versus years 2005-2050, the Probability 677 Distribution Function of ice time series on the EBS has shifted northward by ~ 2° latitude. The 678 implications of these results on marine ecosystem responses are discussed. 679 680 Keywords: Bering Sea, sea ice, numerical model, ocean temperature, salinity 681 28 682 1. Introduction 683 The Bering Sea is a North Pacific 684 subarctic ocean bounded to the north by 685 Bering Strait, which connects the Bering Sea 686 with the Arctic Ocean, and to the south by the 687 Aleutian Island chain (Fig. 1). The Eastern 688 Bering Sea (EBS) shelf is wide (approximately 689 500 km) and flat (with depth less than 180m). 690 As early as November, sea ice begins forming 691 in the northern EBS, and under the prevailing 692 wind conditions, is transported southward 700 area. The black dots mark the ocean moorings in the 693 – 1000 km, often covering much of the EBS shelf. Maximum ice extent typically occurs in 694 March, but it can occur as early as February or as late as April (Stabeno et al., 2012a). During 695 melting season, sea ice retreats and by June the Bering Sea is usually ice free. Sea ice extent is 696 highly variable on interannual scales, but thus far, shows no significant trends since the beginning 697 of the satellite record in 1979 (Brown and Arrigo, 2012). Figure 2. Geographical information of the study Eastern Bering Sea middle shelf. 698 The seasonal changes of sea ice profoundly influence the physical oceanography of the 699 EBS and its ecosystem. During extensive ice years, with sea ice persisting on the southern shelf 700 after mid March, the water column over the southern shelf (south of 60° N) tends to be colder and 701 fresher (Stabeno et al., 2010, 2012b; Ladd and Stabeno, 2012; Sullivan et al., this issue). The 702 timing of ice retreat also influences the timing of the spring phytoplankton bloom (Sigler et al., 703 this issue), zooplankton species composition, and fish recruitment. For instance, a period of 704 limited ice over the southern shelf (2001 – 2005) was characterized by fewer large crustacean 705 zooplankton the following summer (Stabeno et al., 2012a). As a result the young of the year 706 walleye pollock (Theragra chalcogramma) had less lipids and failed to survive the following 29 707 winter (Heintz, personal communication) resulting in very low recruitment (Stabeno et al., 708 2012b). 709 The EBS shelf is divided into three domains (Coachman, 1986). The inner or coastal 710 domain (water depth < 50 m) is characterized as well mixed or weakly stratified during the 711 summer. The middle domain (50 – 100 m) has well defined two layers during the summer with a 712 wind mixed surface layer and tidally mixed bottom layer. The outer domain (100 – 180 m) is 713 more oceanic, with a well-mixed surface layer and bottom layer separated by an intermediate 714 layer. On the middle domain, the influence of the sea ice from the winter and spring often persists 715 through the following summer. As a part of a biophysical observing network on the EBS shelf, 716 four moorings on the middle domain (M2, M4, M5 and M8) (Fig. 1) provide long time series (9 – 717 18 years) of temperature and salinity data that have been published (e.g. Stabeno et al., 2010, 718 2012a,b; Sigler et al., this issue; Sullivan et al., this issue). 719 The purpose of this study is to examine the impacts of seasonal sea ice on the EBS shelf 720 water properties, using output from a global climate model and compare it with in situ ocean 721 (primarily data from moorings) and satellite observations. We will examine sea ice variability in 722 the EBS on seasonal and inter-annual time scales. Pan-Arctic sea ice is projected to decline in the 723 future under Greenhouse Gases (GHGs) forcing but there are large uncertainties (Wang and 724 Overland, 2009). For instance since 2007, winter/spring sea ice extent has been well above 725 average in the EBS even though the summer minimum ice extent in the Arctic has reached record 726 minimums (Stroeve et al. 2007; Stabeno et al., 2012 a,b; Brown and Arrigo 2012). 727 The paper is organized as the following. In section 2 we describe sea ice satellite 728 observational data and the numerical model output used in this study. In section 3 we present the 729 main results, including comparing model simulations with satellite and in situ observations. In 730 section 4 we draw conclusions and discuss the implications of the results on the EBS marine 731 ecosystem. 732 30 733 2. Data and methods 734 2.1. Satellite and sea ice data 735 Ice concentration images are derived using data from the Bootstrap algorithm files of 736 daily ice concentration we download from the National Snow and Ice Data Center (NSIDC). The 737 files use data from the Scanning Multichannel Microwave Radiometer (SMMR) aboard the 738 Nimbus-7 satellite or the Special Sensor Microwave/Imager (SSM/I) aboard Defense 739 Meteorological Satellite Program (DMSP) satellites, depending on the year. We used data from 740 years 1987 to 2005. 741 To examine how the ice cover varies along the EBS middle shelf, a 100 km by 100 km 742 box was defined centered on each of four biophysical moorings (M2, M4, M5, and M8) that have 743 been maintained by NOAA since 1990s. Percent of ice cover from satellite was averaged over 744 these 100 km by 100 km regions and monthly climatologies were calculated. 745 746 2.2. Numerical model output 747 The global climate model we used is the NCAR-CESM (National Center for Atmospheric 748 Research-Community Earth System Model). The NCAR-CESM (and its predecessors) is a global 749 coupled ocean-sea ice-atmosphere-land model (Gent et al. 2012) participating in the IPCC 750 (Intergovernmental Panel on Climate Change) assessment reports. Specific formulations of the 751 sea ice model used in CESM are summarized in Holland et al. (2012). CCSM4 (the version of 752 CESM used in up-to-date publications) simulated Arctic ice extent seasonal cycle in the 20 th 753 century follows observations closely (Jahn et al., 2012). Arctic sea ice simulated by CCSM4 in 754 the 21st century has been investigated in a number of studies (e.g., Vavrus et al., 2012). 755 The particular configuration of the CESM used in this study has 0.9° latitude by 1.25° 756 longitude resolution for the atmosphere and land components, and nominal 1° horizontal 757 resolution for the ocean and sea-ice components. The external climate forcing factors (including 31 758 natural and anthropogenic components) vary with time. Because it is a coupled model, the 759 feedbacks between different components of the climate system, such as the ocean and sea ice, are 760 preserved. Monthly averaged ocean and sea ice states from the CESM “historical” (from 1850 to 761 2005) and “RCP” (Representative Concentration Pathways) (year 2005 onward) simulations were 762 downloaded from NCAR data mass storage. The “historical” simulations of the CESM were 763 initialized from a spun-up state of the climate system under pre-industrial GHGs forcings. The 764 “historical” simulations cover years 1850 to 2005 (we only used output from the late 20 th 765 century), while its “RCP” simulations were initialized from the climate state of the model at the 766 end of the “historical” simulations (Gent et al. 2012), and RCP simulations cover years 2006- 767 2100 and later. We use results from the RCP6.0 forcing (radiative forcing stabilizes at 6.0 W/m2 768 by year 2100), a medium emission scenario. 769 To study the inter-annual variability of sea ice and its impact on the EBS shelf, we extract 770 Empirical Orthogonal Functions (EOFs) of physical variables from multiple years in March. 771 Mean states of the shelf water property and circulation averaged over “high” and “low” ice years 772 are computed and compared against each other. The “high” and “low” ice years are defined based 773 on March ice concentration at M5: if a year’s value is above +1STD of the multi-year time series, 774 that year is a “high” ice year; if it is below -1STD, it’s “low” ice year. 775 To facilitate the comparison with observations, we carried out an additional CESM 776 RCP6.0 simulation with daily sea ice and ocean output, from December 1, 2016 to June 30, 2017. 777 This time period is used as a “representative” year with substantial ice cover in the EBS (shown 778 later). The comparison between this model output and observations are process oriented. Due to 779 the internal variability of the coupled system, simulated climate state at any given year is not 780 guaranteed to “match” with observations for that year. 781 When applicable, the model simulations are compared with existing figures in Sullivan et 782 al. (this issue) which summarizes the ocean in situ observations at the four moorings (M2, M4, 783 M5 and M8) (Fig. 1). 32 784 785 3. Results 786 3.1. Sea ice climatology 787 Monthly mean climatology 788 (defined over years 1987-2005) of sea ice 789 concentration was computed from the 790 CESM 791 compared with the satellite data (Fig. 2). 792 Satellite data show a gradual increase in 793 ice cover from December through March 794 (growth in December is small), with the 1987-2005) of sea ice concentration (percentage) at the “historical” simulation and Figure 3. Monthly mean climatology (averaged over years mooring sites from CESM simulation (red) and SSM/I data (green). Names of the moorings are marked on each panel. Vertical bars show one standard deviation. 795 percent 796 maximum in March on the middle shelf 797 regardless of latitude. After March, ice cover decreases rapidly. At the southern two sites, ice 798 cover is reduced to zero by May. Farther north, an additional month is required for complete loss 799 of sea ice (June at M5 and M8). The magnitude of the standard deviations at each location 800 suggests that ice cover is more variable at M4 than at the other locations examined. of ice cover reaching its 801 While the model simulations show same general patterns as the observations, there are 802 marked differences in timing and magnitude. The CESM captures the March maximum at the 803 southern two sites (Fig. 2, M2 and M4); at the northern sites (M5 and M8), CESM reaches annual 804 maximum ice cover in April, a month later than observed. The model simulation underestimates 805 ice cover at all four locations leading up to the March maximum and overestimates ice cover after 806 March. Complete loss of ice occurs one month later than observed in CESM simulations, except 807 at the southernmost site. The magnitude of the variability is well reproduced by CESM except at 808 the M4 location where the model significantly underestimates the interannual variability. 33 809 The modeled spatial pattern of ice 810 concentration in March, the month of maximum 811 annual ice extent, is similar to observations 812 (Fig. 3). The biggest discrepancy is that the 813 simulated ice concentration gradient is weaker 814 than observations, especially in the western part 815 of the domain. The model results show 816 significant ice cover over the deep basin in the 817 northern ice is 818 constrained to the shelf in observations. In 819 addition, in the southeast corner of the EBS 820 shelf, the influence of warm inflow from the 821 Pacific is observed to result in less ice close to 822 the Alaskan Peninsula (Fig. 3 upper panel). Figure 4. Multi-year (1987-2005) mean March sea ice 823 Due to the low spatial resolution, the CESM 824 model cannot be expected to reproduce such regional features. Bering Sea, while sea concentration (percentage) from SSM/I data (upper panel) and CESM simulation (lower panel). 825 826 3.2. Ice formation: dynamic versus thermodynamic processes 827 The mechanisms that result in changes in the areal extent of ice can be divided into two 828 broad categories: dynamic processes and thermodynamic processes. Dynamic processes include 829 advection and mechanical ridging/rafting, while thermodynamic processes are ice melting and 830 freezing. Not surprisingly, ice cover changes seasonally in different locations because of different 831 processes. 34 832 At all four mooring 833 locations, dynamical processes (for 834 simplicity, 835 “advection”) contribute to positive 836 ice growth in January (Fig. 4). At 837 the southern two moorings (M2 and 838 M5), 839 concentration 840 advection of ice into the region 841 offset by local melting (Fig. 4b, c). Figure 5. Time integrals of ice area tendency we will call instantaneous is a it ice result of t A t dt,T 0 Dec1 T0 842 The 843 advection and the local melt terms instantaneous ice area A (black) at the moorings (names of them are 844 are weaker at M5 than at M2. 845 Further north at M8, ice growth is dominated by advection into the region, with a small 846 contribution from local formation (Fig. 4a). Thus, ice melt contributes to water column properties 847 throughout the winter over a majority of the shelf. Ice formation is only important in the 848 northernmost region from January through April. After April, local melting contributes to the 849 decline in ice cover even in the north. magnitudes of both the (here A is the ice area and t is time) due to dynamics (green) and thermodynamics (red), and their comparisons with the daily marked on the panels). Ice area is defined as percentage coverage. they are similar to results at Results at M4 are not shown because M2. 850 Spatially, near Bering Strait and along the inner shelf, ice formation (Fig. 5a) is largely 851 offset by advection out of the region (Fig. 5b). This balance is reversed in the southern and 852 offshore locations, where advection into the region is offset by local ice melt. The pattern thus 853 emerged is that, sea ice is thermodynamically formed in the northern domain and along the coast, 854 and then transported southward and offshore, and subsequently melted in the warmer ambient 855 environment. This is the well-known “freshwater conveyor” effect of seasonal sea ice on the 856 Bering Sea shelf (Pease 1980; Sullivan et al, this issue). During this seasonal transit, the largest 857 ice-ocean exchanges in heat and freshwater occur at the ice edge (Figs. 5 c, d). 35 858 The above ice growth and 859 retreat processes have strong interannual 860 variabilities, which are influenced by 861 wind 862 empirical orthogonal functions (EOFs) 863 of March ice advective growth (ice area 864 tendency 865 formation/melt (ice area tendency due to 866 thermodynamics), and wind stress are 867 extracted 868 another (Fig. 6). The dominant mode of 869 March 870 variability (accounting for over 60% of 871 the variability) is an oscillation between 872 northeasterly and southwesterly winds 873 (Fig. 6c,d). The principal component 874 (PC) of this wind stress mode (black line 875 on Fig. 6d) is highly correlated with PCs 876 of 877 thermodynamics and dynamics (Fig. 6d, 878 red and green lines): the correlation to dynamics (a) and thermodynamics (b), and spatial pattern of 879 coefficients between the three PCs on denote the 50m, 100m, and 200m ocean topography. Contours of forcing. due To to illustrate dynamics), this, ice Figure 6. Colors show the spatial patterns of ice area tendency ice and wind area compared stress with one (percent/day) due to thermodynamics (a) and dynamics (b), and inter-annual tendencies due ice to ocean heat (c, in W/m2) and freshwater (d, in cm/day) fluxes. White areas are ice-free. Positive signs in c) and d) mean into the ocean. Contours in c) and d) mark the 0.8, 0.6, 0.4, and 0.2 ice area (in fraction). All results are averaged over February of the model year shown in Figure 4. Circles mark the mooring locations. to Figure 6. Spatial patterns of EOF1 of the ice area tendency due EOF1 of the surface wind stress (c). Thin black lines on a) and b) a) is redrawn in c). d) Corresponding principal components (PCs) of the spatial patterns shown in a)–c). CESM March data is used for this analysis. Percentage of variance explained by EOF1 is marked on a)–c). 880 Fig. 6d range from 0.80 to 0.97. When 881 the winds are from the northeast, more 882 sea ice is advected into the middle shelf (Fig. 6a), where it melts (Fig. 6b). At the same time, in 883 the northern and coastal regions, freezing (Fig. 6b) and ice removal by advection (Fig. 6a) are 36 884 enhanced under northeasterly winds (Fig. 6c). This strong coupling between ice advection and 885 melting is a characteristic of the EBS. 886 Next we examine relationships 887 between seasonal ice growth/decline 888 (via 889 formation/melting) and ice-ocean heat 890 and freshwater exchange (Fig. 7). 891 Advection of ice into the region begins 892 in December in the north (M8) and 893 January farther south (M2 and M5) and Figure 7a. Modeled daily ice area tendency due to 894 cooling of the ocean occurs soon after. 895 At the northern sites (M5 and M8), there 896 is a period in which ice area tendency and ice-ocean fluxes are relatively constant between 897 January, the beginning of the seasonal ice increase, and March or April, the beginning of seasonal 898 decrease (Fig. 7). Advection induced ice area tendency (red lines in Fig. 7) is generally smaller at 899 the northern mooring M8 than at moorings farther south (M5 and M2). advection and local thermodynamics (red) and dynamics (green) and their comparisons with the simultaneous ice to ocean heat fluxes (black). Positive (negative) heat flux means into (out of) the ocean. Panels a)–c) correspond to M8, M5, and M2 respectively. M4 results are similar to M2 and not shown. 900 In terms of ice-ocean heat exchange, at all sites and throughout the season, the ocean 901 loses heat. At M2 and M5, during the ice growth season (January or February), the heat loss 902 appears correlated with ice area tendency but lags the latter by a few days (Fig. 7a). The lag-0 903 correlation coefficients between black and red (green) lines on Fig. 7 are 0.97 (-0.52) and 0.74 (- 904 0.54) for M2 and M5, respectively, which are statistically significant with the 95% confidence 905 level. Heat loss in these locations is controlled by both latent and sensible heat exchange. During 906 ice formation, latent heat would result in a positive heat flux to the ocean. At M8, freezing (Fig. 907 7a; positive red line) is accompanied by negative heat flux to the ocean (Fig. 7a; black line), 908 implying that sensible heat flux dominates latent heat flux. This is consistent with model output 909 (Fig. 9) showing that the ocean temperature is above the freezing point in January. Because ice 37 910 bottom temperature is at the freezing point, there is a sensible heat exchange at the ice-ocean 911 interface. 912 The relationship between ice 913 tendency and ice-ocean freshwater 914 exchange (Fig. 7b) is straightforward. 915 At all locations, the water column 916 freshens (becomes saltier) whenever ice 917 melts (forms). Thus, the ocean gains 918 freshwater in the south throughout the 919 season while, in the north, the ocean Figure 7b. Same as Fig. 7a except the black lines are ice to oean 920 gains salt during ice formation (mainly of) the ocean. 921 in January and February) and freshwater during ice melting (in April and May). Ice advective 922 growth (green lines in Fig. 7) plays an implicit role in ice-ocean freshwater exchange through its 923 control on ice thermodynamic growth (Fig. 5). freshwater fluxes (cm/day), positive (negative) means into (out 924 925 3.3. 926 temperature and salinity 927 Influences on The vertical ocean structure and 928 temporal 929 temperature and salinity are related to 930 ice cover. At M5 and M8, the water 931 column in CESM is vertically well 932 mixed 933 temperature 934 beginning of the winter (December) as 935 a result of wind mixing (Fig. 8). The weak stratification at M8 (and to a lesser extent at M5) is variability or weakly and of ocean stratified salinity at in the Figure 8. Colors show modeled ocean temperature (left) and sality (right) as a function of depth (0-100m) and time at the moorings. The thick black lines on each panel show ice area (percentage) at that mooring. M4 results are not shown because they are similar to M2 results. 38 936 maintained through spring under significant ice cover, and at M8, there is a hint of brine rejection 937 into the bottom layer when ice is formed. This is not dissimilar to observations at the 938 moorings. The observed water column at M8 tends to be well mixed under the ice, with a winter 939 season increase in salinity of ~1 psu, which is a result of brine rejection or on-shelf advection 940 during the cold winter months (e.g., Fig. 12 in Sullivan et al., this issue). This observed increased 941 in salinity contrasts with a small (~0.2 psu) depth-averaged decrease in salinity in the model. 942 At M2, however, the differences between the modeled and observed water column 943 structures are greater. In December, the modeled water column is stratified near the bottom, albeit 944 weakly, especially in salinity (Fig. 8, bottom panels). In observations, during years with little or 945 no ice over the mooring, the water column is well mixed usually by November (Table 2 and Fig. 946 6 in Sullivan et al., this issue). During years in which ice is extensive over the southern domain, 947 the ice often arrives when the water column is well mixed, but above freezing. The ice cools and 948 freshens the surface waters, but the near bottom waters remain warmer and saltier for several 949 weeks before the water column becomes 950 well mixed. 951 cooling and freshening at M2 after ice 952 arrival, similar to observations, but 953 stratification persists in the model. In 954 CESM the ocean stratification begins to 955 increase during the melting season at all 956 mooring locations (Fig. 8). Stratification 957 measured at the moorings usually begins 958 after ice concentrations have decreased ocean temperature (red, units: °C) and salinity (green, units: 959 CESM also simulates Figure 9. Modeled daily ice area (black,, units: percentage) and (Sullivan et al., this issue). psu) at 10m depth at moorings M2, M5 and M8. M4 result is similar to M2 result and therefore not shown. 960 The near-surface ocean gradually cools starting from fall (not shown), but the strongest 961 cooling is concurrent with largest sea ice increase at all moorings (Fig. 9, red lines). This result is 39 962 consistent with observations (Sullivan et al., this issue, their Figs. 7-12). In the model, as in 963 observations, cooling at M8 and M5 continues even under significant ice coverage (Fig. 9ab, red 964 line, late January), until the temperature reaches the freezing point and remains constant 965 thereafter. The quasi-steady thermal ocean lasted a couple months at M8, but only briefly at 966 M5. In observations, warming generally begins in March, although if ice retreats, the water 967 column can warm through advection especially at the southern moorings M2 and M4 (Stabeno et 968 al., 2010, 2012a; Sullivan et al., this issue). The model shows a similar pattern, in that after the 969 sea ice diminishes (which occurs sooner in the lower latitudes), the near-surface ocean warms 970 rapidly (Fig. 9). 971 The modeled seasonal evolution of near-surface salinity is more variable and differs from 972 north to south (Fig. 9, green lines). Starting in December, modeled near-surface salinity at the two 973 northern mooring sites generally decreases. At M8, the linear trend continues throughout the 974 season, although it is also punctuated by episodes of salinity increase. At M5, salinity decreases 975 from December to early February along with ice increase, it then exhibits a period of weak 976 changes from late February to March, followed by a gradual increase. At M2, large synoptic scale 977 variability is observed in the near-surface salinity. In contrast, at each of the mooring sites, 978 observed salinity increases from December to March because of brine rejection and/or on-shelf 979 advection during the cold winter months, after which ice begins to melt, thus freshening the water 980 column (Sullivan et al., this issue). 981 982 983 984 3.4. Contrast in ocean properties between “high” and “low” ice years Change in bottom temperature, salinity, and density in high ice years relative to low ice years indicates cooling anomalies over the entire bottom layer, with the strongest 40 985 anomaly occurring in the southern 986 domain (Fig. 10a); concurrently, a 987 large part of the inner and middle 988 shelf domain has positive salinity 989 anomalies (Fig. 10b). While the 990 gradient 991 anomalies is the strongest in the 992 north-south direction, the salinity 993 anomaly has the strongest gradient 994 in the onshore-offshore direction. As Figure 10. Shading indicates anomalies in March bottom 995 expected, in this subarctic ocean, the 996 bottom density anomaly (Fig. 10d) is 997 dominantly controlled by salinity. 998 The in the temperature temperature (a), salinity (b), and density (d) averaged over high ice years minus that over low ice years. Thin black lines in a) and b) mark the 50m, 100m, and 200m isobath. Vectors in d) show the ocean ciruculation anomalies at 50m depth, dark blue ones are the results on the shelf (depth< 200m), while the light blue ones are results in the slope region (200m<depth<2000m). March bottom density (shade) and 50m currents (vectors) averaged over high ice years are shown in c) as a reference. CESM output from years 1961-2005 is used in this analysis. corresponding upper 999 ocean circulation is anomalously southward on the shelf except in the Gulf of Anadyr (Fig. 10d, 1000 vectors). The anomalous circulation is close to the shelf-break between 59°N and 60°N with a 1001 slightly offshore direction, but is located further in-shore in the southern domain (south of 59°N), 1002 and shows a strong on-shore movement toward the southeast corner of the EBS when it reaches 1003 the Aleutian Island chain. However, the circulation anomalies next to the shelf-break in the slope 1004 region (200m < depth < 2000m) are opposite to circulation anomalies on the shelf, and move 1005 1006 northwestward. 41 1007 3.5. 1008 We compare March ice cover at the mooring sites between two periods: years 1961-2005 vs. 1009 years 2006-2050, since March is near 1010 the annual maximum ice extent (Fig. 1011 2). During the earlier period, ice 1012 cover at M2 and M4 is usually low (< 1013 50%) while ice cover at M8 is usually 1014 high (>50%) (Fig. 11, left panels). Ice 1015 cover at M5 fluctuates between low 1016 and high conditions on interannual 1017 time scales (Fig. 11, left panels). Ice Figure 11. Time series of March sea ice area (percentage) at the 1018 cover at all locations in the later 1019 mooring locations during years 1961-2005 (left panels) and years 2006-2050 (right panels) from CESM. Year 2005 is the end of “historical” period, while year 2006 is the beginning of “RCP” period. period (Fig. 11, right panels) is 1020 smaller relative to the earlier period. 1021 M2 and M4 are mostly ice-free in 1022 the later period, only occasionally 1023 having significant ice cover. In the 1024 earlier 1025 distribution functions (PDFs) of ice 1026 cover at M2 and M4 show increased 1027 occurrence of low ice cover events 1028 while the PDF at M8 has more Figure 12. Probability distribution functions (PDFs) of March ice 1029 occurrence of high ice cover events 1030 (Fig. 12, left panels); PDF at M5 is approximately flat among all categories (Fig. 12, left panel, 1031 M5). In the later period, PDF at M5 has more occurrence of low ice cover events while PDF at Decadal changes: “historical” versus “RCP” simulations period, the probability area (percentage) at the mooring locations. Left panels are results from period 1 (years 1961-2005), right panels are from period 2 (years 2006-2050). 42 1032 M8 is more or less flat among all categories (Fig. 12, right panels). In this sense, PDF at M8 in 1033 the later period is similar to PDF at M5 in the earlier period. 1034 1035 4. Conclusions and Discussion 1036 Seasonal cycle and spatial pattern of the annual maximum ice cover in the EBS modeled 1037 by CESM are reasonable compared to satellite observations (Figs. 2, 3). However, biggest 1038 discrepancies include delayed onset of the annual maxima (by a month) and slower summer 1039 melting in the northern Bering Sea in the simulation (Fig. 2). 1040 One possible cause for the slower summer melt could be that the simulation maintains 1041 weak stratification of the water column under the ice (Fig. 8), while observations show that the 1042 water column is well mixed throughout the winter. The EBS shelf loses heat and gains freshwater 1043 during the melting season. If the ocean layer receiving these fluxes is too shallow (due to 1044 stratification), this cooling and freshening effect on the surface layer would be too strong. A 1045 cooled ocean slows down further melting, causing a negative feedback on ice melting. An 1046 anomalously fresh surface layer further increases water column stability. In addition, an initial 1047 delay in ice retreat allows ice to remain into the period when spring winds are weakening, further 1048 delaying ice break-up and melt. These feedbacks may contribute to the slower melting in the 1049 model than in observations. 1050 Modeled seasonal ice advance in the EBS is consistent with the “freshwater conveyor” 1051 conceptual model suggested by observations (Pease 1980; Sullivan et al., this issue). Sea ice is 1052 both advected in and formed locally in the northern domain. In the southern domain, sea ice is 1053 advected from the north and melts locally. The regional differences in the partition between 1054 advection and formation/melt influence water column properties, particularly affecting local 1055 salinity budgets. The boundary between the northern domain where formation dominates and the 1056 southern domain where advection and melt dominate changes seasonally (Fig. 5) and 43 1057 interannually (Fig. 6). On interannual time scales, the division is strongly influenced by surface 1058 wind. 1059 At all mooring sites and throughout the season, modeled heat flux at the ice-ocean 1060 interface is from the ocean to the ice (Fig. 7a) while freshwater flux can be either into the ocean 1061 (during melting) or out of the ocean (brine rejection during freezing). The salt/freshwater flux in 1062 nature is influenced by island polynyas in the Bering Sea. St. Lawrence Island, to the north of 1063 M8, and to a lesser extent St. Matthews Island to the north of M5, is associated with important 1064 polynyas. CESM does not resolve either polynya. Thus, the model will underestimate ice 1065 formation due to the polynyas and the associated salinity flux into the ocean at these locations. 1066 Despite these limitations, in CESM, higher spring ice concentration in the northern EBS 1067 leads to positive salinity anomalies (relative to years with less ice concentration) on a large part of 1068 the bottom water (Fig. 10b), and the anomaly is the strongest in the northern domain near-shore 1069 locations. Weak negative salinity anomalies are seen in the southern end of the middle- and outer- 1070 shelf (Fig. 10b). Correspondingly, bottom water on the entire shelf has cold anomalies in high ice 1071 years relative to low ice years (Fig. 10a), and the anomalies are the strongest in the southern 1072 domain. Both temperature and salinity anomalies persists from spring into summer (not shown). 1073 According to the model, circulation on the middle shelf is anomalously southward during 1074 high ice years; however, next to the shelf-break in the slope region, circulation is anomalously 1075 northwestward during high ice years, opposite to situation on the shelf (Fig. 10 d). Broadly 1076 speaking, the southward circulation anomalies on the shelf are consistent with the southward 1077 wind anomalies during high ice years (Fig. 6). Such wind anomalies are also expected to cause 1078 the Anadyr current to flow stronger in the on-shelf direction toward Bering Strait (Danielson et al. 1079 2012). The reason for the northwestward anomalies in the slope region is unclear. The strength of 1080 the Bering Slope Current is correlated with the multivariate ENSO index and North Pacific Index 1081 on interannual time scales (Ladd, this issue), it is possible that these climate indices have 1082 manifestations in the EBS surface wind and sea ice variability. 44 1083 Over a 45-year time span (between period 1: 1961-2005 and period 2: 2006-2050), the 1084 EBS ice concentration PDF have shifted northward by 2 degrees. PDF of Ice concentration at M8 1085 in the later period is more or less flat, similar to PDF of ice concentration at M5 in the earlier 1086 period, while PDF at M5 in the later period becomes similar to PDF at M2 and M4 in the earlier 1087 period. 1088 Sea ice has profound effect on the EBS primary production (Hunt et al., 2002; Stabeno et 1089 al. 2010; Brown and Arrigo, 2012; Sigler et al., this issue), zooplankton structure (Hunt et al. 1090 2002; Coyle et al., 2011; Eisner et al., this issue), and upper trophic level species (e.g., Wyllie- 1091 Echeverria and Wooster, 1998). Timing of ice retreat determines spring bloom. Over the southern 1092 shelf, if ice is present after mid-March, an early ice-associated bloom occurs; otherwise the spring 1093 bloom usually occurs in May (Stabeno et al. 2001; Hunt et al. 2002). In this regard, CESM 1094 captures well the March maximum ice cover in the southern domain in a climatological mean 1095 sense. 1096 Lower water temperatures associated with more sea ice reduce energy required for 1097 zooplankton overwinter survival, resulting in increased spawning biomass and production (Sigler 1098 et al., this issue). In addition, the distribution of low bottom temperatures associated with ice 1099 distribution (the cold pool) determine available habitat for some fish species (Hollowed et al. 1100 2012). Conversely, warmer temperatures increase metabolic requirements, leading to food 1101 limitation. This could result in reduced net primary production (Lomas et al., 2012), leading to 1102 poor overwinter survival of zooplankton (Coyle et al., 2011). 1103 In addition to influences on temperature and salinity, ice melt results in input of ice algae 1104 to the water column, and ice algae provides an early sources of food for zooplankton growth and 1105 spawning (Sigler et al., this issue). Thus, the “freshwater conveyor” associated with seasonal sea 1106 ice in the EBS can also be thought of as conveyor belt for ice alge, and possibly zooplankton. The 1107 balances between ice advection and ice formation/melt are important to the ecosystem, and 1108 understanding what processes controls the seasonal to interannual variability in these balances 45 1109 will have important implications. Our results suggest surface winds in the EBS should be closely 1110 monitored because of its strong influences on ice advection and local formation. 1111 Global climate models are more often used to study global and basin scale processes, 1112 their applications on regional scales are relatively rare. Understanding how climate models work 1113 on all scales, however, is critically important to our ability to predict future changes in the 1114 physical environment and marine ecosystems, the latter tend to operate on regional scales. Our 1115 next step is to investigate the Bering Sea ice-ocean system in a coupled biophysical ocean-ice 1116 circulation model. This class of models can afford higher spatial resolutions, although they 1117 misrepresent atmosphere-ocean or atmosphere-sea ice interactions. Combining knowledge gained 1118 from different models and observations should ultimately improve our understanding of the 1119 system. 1120 1121 Acknowledgements 1122 We thank W. Floering, C. Dewitt and S. McKeever for maintaining, deploying and recovering the 1123 instruments on the mooring, and D. Kachel, and P. Proctor for data processing. The research was 1124 generously supported by grants from NPRB (grant numbers), and the NPRB-sponsored BSIERP 1125 and NOAA’s North Pacific Climate Regimes and Ecosystem Productivity (NPCREP) programs. 1126 The CESM project is supported by the National Science Foundation and the Office of Science 1127 (BER) of the U.S. Department of Energy. Computing resources were provided by the Climate 1128 Simulation Laboratory at NCAR's Computational and Information Systems Laboratory (CISL), 1129 sponsored by the National Science Foundation and other agencies. This research was enabled by 1130 CISL compute and storage resources. Bluefire, a 4,064-processor IBM Power6 resource with a 1131 peak of 77 TeraFLOPS provided more than 7.5 million computing hours, the GLADE high-speed 1132 disk resources provided 0.4 PetaBytes of dedicated disk and CISL's 12-PB HPSS archive 1133 provided over 1 PetaByte of storage in support of this research project. This is contribution 1134 EcoFOCI-0806 to the Ecosystems & Fisheries Oceanography Coordinated Investigations; 2093 to 46 1135 Joint Institute for the Study of Atmosphere and Ocean, University of Washington, and 3995 to 1136 NOAA/Pacific Marine Environmental Laboratory. This is the BEST_BSIERP Bering Sea Project 1137 publication number XX. 1138 1139 Figure Captions 1140 Figure 1. Geographical information of the study area. The black dots mark the ocean moorings in 1141 the Eastern Bering Sea middle shelf. 1142 Figure 2. Monthly mean climatology (averaged over years 1987-2005) of sea ice concentration 1143 (percentage) at the mooring sites from CESM simulation (red) and SSM/I data (green). Names of 1144 the moorings are marked on each panel. Vertical bars show one standard deviation. 1145 Figure 3. Multi-year (1987-2005) mean March sea ice concentration (percentage) from SSM/I 1146 data (upper panel) and CESM simulation (lower panel). 1147 Figure 4. Time integrals of ice area tendency t A t dt,T 0 Dec1 (here A is the ice area and t is T0 1148 time) due to dynamics (green) and thermodynamics (red), and their comparisons with the daily 1149 instantaneous ice area A (black) at the moorings (names of them are marked on the panels). Ice 1150 area is defined as percentage coverage. Results at M4 are not shown because they are similar to 1151 results at M2. 1152 Figure 5. Colors show the spatial patterns of ice area tendency (percent/day) due to 1153 thermodynamics (a) and dynamics (b), and ice to ocean heat (c, in W/m2) and freshwater (d, in 1154 cm/day) fluxes. White areas are ice-free. Positive signs in c) and d) mean into the ocean. 1155 Contours in c) and d) mark the 0.8, 0.6, 0.4, and 0.2 ice area (in fraction). All results are averaged 1156 over February of the model year shown in Figure 4. Circles mark the mooring locations. 1157 Figure 6. Spatial patterns of EOF1 of the ice area tendency due to dynamics (a) and 1158 thermodynamics (b), and spatial pattern of EOF1 of the surface wind stress (c). Thin black lines 1159 on a) and b) denote the 50m, 100m, and 200m ocean topography. Contours of a) is redrawn in c). 47 1160 d) Corresponding principal components (PCs) of the spatial patterns shown in a)–c). CESM 1161 March data is used for this analysis. Percentage of variance explained by EOF1 is marked on a)– 1162 c). 1163 Figure 7a. Modeled daily ice area tendency due to thermodynamics (red) and dynamics (green) 1164 and their comparisons with the simultaneous ice to ocean heat fluxes (black). Positive (negative) 1165 heat flux means into (out of) the ocean. Panels a)–c) correspond to M8, M5, and M2 respectively. 1166 M4 results are similar to M2 and not shown. 1167 Figure 7b. Same as Fig. 7a except the black lines are ice to oean freshwater fluxes (cm/day), 1168 positive (negative) means into (out of) the ocean. 1169 Figure 8. Colors show modeled ocean temperature (left) and sality (right) as a function of depth 1170 (0-100m) and time at the moorings. The thick black lines on each panel show ice area 1171 (percentage) at that mooring. M4 results are not shown because they are similar to M2 results. 1172 Figure 9. Modeled daily ice area (black,, units: percentage) and ocean temperature (red, units: °C) 1173 and salinity (green, units: psu) at 10m depth at moorings M2, M5 and M8. M4 result is similar to 1174 M2 result and therefore not shown. 1175 Figure 10. Shading indicates anomalies in March bottom temperature (a), salinity (b), and 1176 density (d) averaged over high ice years minus that over low ice years. Thin black lines in a) and 1177 b) mark the 50m, 100m, and 200m isobath. Vectors in d) show the ocean ciruculation anomalies 1178 at 50m depth, dark blue ones are the results on the shelf (depth< 200m), while the light blue ones 1179 are results in the slope region (200m<depth<2000m). March bottom density (shade) and 50m 1180 currents (vectors) averaged over high ice years are shown in c) as a reference. CESM output from 1181 years 1961-2005 is used in this analysis. 1182 Figure 11. Time series of March sea ice area (percentage) at the mooring locations during years 1183 1961-2005 (left panels) and years 2006-2050 (right panels) from CESM. Year 2005 is the end of 1184 “historical” period, while year 2006 is the beginning of “RCP” period. 48 1185 Figure 12. Probability distribution functions (PDFs) of March ice area (percentage) at the 1186 mooring locations. Left panels are results from period 1 (years 1961-2005), right panels are from 1187 period 2 (years 2006-2050). 1188 1189 References 1190 Brown, Z.W., Arrigo, K.R., 2012. Contrasting trends in sea ice and primary production in the 1191 Bering Sea and Arctic Ocean. ICES J. Mar. Sci. 69, 1180-1193, 10.1093/icesjms/fss113. 1192 Coachman, L.K., 1986. Circulation, water masses and fluxes on the southeastern Bering Sea 1193 shelf. Cont. Shelf Res. 5(1–2), 23–108. 1194 Coyle, K.O., Eisner, L.B., Mueter, F.J., Pinchuk, A.I., Janout, M.A., Cieciel, K.D., Farley, E.V., 1195 Andrews, A.G., 2011. Climate change in the southeastern Bering Sea: impacts on pollock 1196 stocks and implications for the oscillating control hypothesis. Fish. Oceanogr. 20, 139-156, 1197 10.1111/j.1365-2419.2011.00574.x. 1198 Danielson, S., Hedstrom, K., Aagaard, K., Weingartner, T., Curchitser, E., 2012. Wind-induced 1199 reorganization 1200 http://dx.doi.org/10.1029/2012GL051231. of the Bering Shelf circulation. Geophys. Res. Lett. 39(8), 1201 Eisner, L.B., Napp, J.M., Mier, K.L., Pinchuk, A.I., Andrews, A.G., submitted. Climate-mediated 1202 changes in zooplankton community structure for the eastern Bering Sea. Deep-Sea Res. II. 1203 Gent, P. R., G. Danabasoglu, L. J. Donner, M. M. Holland, E. C. Hunke, S. R. Jayne, D. M. 1204 Lawrence, R. B. Neale, P. J. Rasch, M. Vertenstein, P. H. Worley, Z. -L. Yang, and M. 1205 Zhang, 2011: The Community Climate System Model version 4. J. Climate, 24, 4973-4991, 1206 doi: 10.1175/2011JCLI4083.1. 1207 Holland, M. M., D. A. Bailey, B. P. Briegleb, B. Light, and E. Hunke, 2012: Improved Sea Ice 1208 Shortwave Radiation Physics in CCSM4: The Impact of Melt Ponds and Aerosols on Arctic 1209 Sea Ice. J. Climate, 25, 1413-1430, doi: 10.1175/JCLI-D-11-00078.1. 49 1210 Hollowed, A.B., Barbeaux, S.J., Cokelet, E.D., Farley, E., Kotwicki, S., Ressler, P.H., Spital, C., 1211 Wilson, C.D., 2012. Effects of climate variations on pelagic ocean habitats and their role in 1212 structuring forage fish distributions in the Bering Sea. Deep-Sea Res. II 65–70, 230-250, 1213 http://dx.doi.org/10.1016/j.dsr2.2012.02.008. 1214 Hunt, G.L., Jr., Stabeno, P., Walters, G., Sinclair, E., Brodeur, R.D., Napp, J.M., Bond, N.A., 1215 2002. Climate change and control of the southeastern Bering Sea pelagic ecosystem. Deep- 1216 Sea Res. II 49, 5821-5853. 1217 Jahn, A., D.A. Bailey, C.M. Bitz, M.M. Holland, E.C. Hunke, J. E. Kay, W. H. Lipscomb, J.A. 1218 Maslanik, D. Pollak, K. Sterling, J. Strœve, 2012: Late 20th Century Simulation of Arctic Sea 1219 Ice and Ocean Properties in the CCSM4. J. Climate, 25, 1431-1452, doi: 10.1175/JCLI-D-11- 1220 00201.1. 1221 Vavrus, S. J., M. M. Holland, A. Jahn, D. A. Bailey, and B. A. Blazey, 2012: 21st-Century 1222 Arctic Climate Change in CCSM4. J. Climate, 25, 2696-2710, doi: 10.1175/JCLI-D-11- 1223 00220.1. 1224 1225 Ladd, C.A., Stabeno, P.J., 2012. Stratification on the Eastern Bering Sea shelf revisited. Deep-Sea Res. II 65–70, 72–83. 1226 Lomas, M.W., Moran, S.B., Casey, J.R., Bell, D.W., Tiahlo, M., Whitefield, J., Kelly, R.P., 1227 Mathis, J.T., Cokelet, E.D., 2012. Spatial and seasonal variability of primary production on 1228 the 1229 http://dx.doi.org/10.1016/j.dsr2.2012.02.010. Eastern Bering Sea shelf. Deep-Sea Res. II 65–70, 126-140, 1230 Pease, C.H., 1980. Eastern Bering Sea ice processes. Mon. Weather Rev. 108(12), 2015–2023. 1231 Sigler, M.F., Stabeno, P.J., Eisner, L.B., Napp, J.M., Mueter, F.J., this issue. Spring and fall 1232 phytoplankton blooms in a productive subarctic ecosystem, the eastern Bering Sea, during 1233 1955-2011. 50 1234 Stabeno, P.J., Bond, N.A., Kachel, N.B., Salo, S.A., Schumacher, J.D., 2001. On the temporal 1235 variability of the physical environment over the south-eastern Bering Sea. Fish. Oceanogr. 1236 10, 81-98. 1237 Stabeno, P.J., Napp, J., Mordy, C., Whitledge, T., 2010. Factors influencing physical structure 1238 and lower trophic levels of the eastern Bering Sea shelf in 2005: Sea ice, tides and winds. 1239 Prog. Oceanogr. 85(3–4), 180–196. 1240 Stabeno, P.J., Farley Jr., E.V., Kachel, N.B., Moore, S., Mordy, C.W., Napp, J.M., Overland, J.E., 1241 Pinchuk, A.I., Sigler, M.F., 2012a. A comparison of the physics of the northern and southern 1242 shelves of the eastern Bering Sea and some implications for the ecosystem, Deep-Sea Res. II 1243 65–70, 14–30. 1244 Stabeno, P.J., Kachel, N.B., Moore, S.E., Napp, J.M., Sigler, M., Yamaguchi, A., Zerbini, A.N., 1245 2012b. Comparison of warm and cold years on the southeastern Bering Sea shelf and some 1246 implications for the ecosystem, Deep-Sea Res. II 65–70, 31–45. 1247 1248 1249 1250 1251 1252 1253 1254 Stroeve, J., M. M. Holland, W. Meier, T. Scambos, and M. Serreze, 2007: Arctic sea ice decline: Faster than forecast. Geophys. Res. Lett., 34, L09501, doi:10.1029/2007GL029703. Magaret Sullivan, M., C. Mordy, S. Salo, and P. Stabeno, this issue: The influence of sea ice on seasonal water column changes on the eastern Bering Sea shelf. Wang, M., and J. E. Overland, 2009: A sea ice free summer Arctic within 30 years? Geophys. Res. Lett., 36, L07502, doi:10.1029/2009GL037820. Wyllie-Echeverria, T., Wooster, W.S., 1998. Year-to-year variations in Bering Sea ice cover and some consequences for fish distributions. Fish. Oceanogr. 7, 159-170. 1255 1256 51 1257 Chapter 13 1258 1259 Eisner, L. B., J. M. Napp, K. L. Mier, A. I. Pinchuk, A. G. Andrews III. (2013). Climate- 1260 mediated changes in zooplankton community structure for the eastern Bering Sea , Deep-Sea Res. 1261 II. 1262 1263 Climate-mediated changes in zooplankton community structure for the eastern Bering Sea 1264 1265 Lisa B. Eisnera*, Jeffrey M. Nappa, Kathryn L. Miera, Alexei I. Pinchukb, Alexander G. Andrews 1266 IIIc. 1267 1268 a 1269 98115, Lisa.Eisner@NOAA.gov, 001-206-526-4060; Jeff.Napp@NOAA.gov; 1270 Kathy.Mier@NOAA.gov 1271 b 1272 Loop Road, Juneau, Alaska 99801, aipinchuk@alaska.edu 1273 c 1274 Alaska 99801, Alex.Andrews@NOAA.gov 1275 1276 1277 1278 *Corresponding Author Address 1279 Zooplankton are critical to energy transfer between higher and lower trophic levels in the eastern 1280 Bering Sea ecosystem. Previous studies from the southeastern Bering Sea shelf documented 1281 substantial differences in zooplankton taxa in the Middle and Inner Shelf Domains between warm 1282 and cold years. Our investigation expands this analysis into the northern Bering Sea and the south 1283 Outer Domain, looking at zooplankton community structure during a period of climate-mediated, 1284 large-scale change. Elevated air temperatures in the early 2000s resulted in regional warming and NOAA-Fisheries, Alaska Fisheries Science Center, 7600 Sand Point Way NE, Seattle, WA University of Alaska Fairbanks, School of Fisheries and Ocean Sciences, 17101 Point Lena NOAA-Fisheries, Alaska Fisheries Science Center, 17109 Point Lena Loop Road, Juneau, Abstract: 52 1285 low sea ice extent in the southern shelf whereas the late 2000s were characterized by cold 1286 winters, extensive spring sea ice, and a well- developed pool of cold water over the entire Middle 1287 Domain. The abundance of large zooplankton taxa such as Calanus spp. (C. marshallae and C. 1288 glacialis), and Parasagitta elegans, increased from warm to cold periods, while the abundance of 1289 gelatinous zooplankton (Cnidaria) and small taxa decreased. Biomass followed the same trends as 1290 abundance, except that the biomass of small taxa in the southeastern Bering Sea remained 1291 constant due to changes in abundance of small copepod taxa (increases in Acartia spp. and 1292 Pseudocalanus spp. and decreases in Oithona spp.). Statistically significant changes in 1293 zooplankton community structure and individual species were greatest in the Middle Domain, but 1294 were evident in all shelf domains, and in both the northern and southern portions of the eastern 1295 shelf. Changes in community structure did not occur abruptly during the transition from warm to 1296 cold, but seemed to begin gradually and build as the influence of the sea ice and cold water 1297 temperatures persisted. The change occurred one year earlier in the northern than the southern 1298 Middle Shelf. These and previous observations demonstrate that lower trophic levels within the 1299 eastern Bering Sea respond to climate-mediated changes on a variety of time scales, including 1300 those shorter than the commonly accepted quasi-decadal time periods. This lack of resilience or 1301 inertia at the lowest trophic levels affects production at higher trophic levels and must be 1302 considered in management strategy evaluations of living marine resources. 1303 1304 Keywords: Zooplankton, community structure, climate change, eastern Bering Sea 1305 1306 1.0 Introduction: 1307 In recent years, climate change in the western Arctic has led to rapid changes in the eastern 1308 Bering Sea shelf resulting in variations in seasonal sea ice coverage and water column 1309 temperatures, and these variations have affected the entire ecosystem (e.g. Napp and Hunt, 2001; 1310 Stabeno et al., 2012b). Zooplankters are essential prey for many fish, seabirds, and marine 53 1311 mammals, therefore there is considerable interest in changes in zooplankton abundance and how 1312 these changes may propagate through the food web and impact higher trophic levels (Coyle et al., 1313 2011; Hunt et al., 2011). Many planktivorous fishes, seabirds, and marine mammals inhabit both 1314 the northeastern and southeastern Bering Sea during summer (e.g. Piatt and Springer, 2003; 1315 Friday et al., 2012; Hollowed et al., 2012; Stevenson and Lauth, 2012). Historical research 1316 documented variations in single species abundance and cross shelf patterns of zooplankton in the 1317 southeastern Bering Sea (Cooney and Coyle, 1982; Coyle and Pinchuk, 2002; Napp et al., 2002; 1318 Stabeno et al., 2010; Coyle et al., 2011; Stabeno et al., 2012a & b). However, there are few 1319 modern descriptions of the broad-scale distribution of eastern Bering Sea zooplankton, 1320 particularly for the northern Bering Sea (Motoda and Minoda, 1974; Coyle et al., 1996 and 1321 references therein). To better understand how climate variability and change are affecting the 1322 recruitment and broad-scale distributions of planktivores, we need a more thorough understanding 1323 of how climate and other forcing affects the production and distribution of zooplankton across the 1324 entire region. 1325 1326 The eastern Bering Sea is a large marine ecosystem characterized by a broad continental shelf > 1327 500 km wide and > 1000 km long oriented in a north westerly direction from the Alaskan 1328 Peninsula in the south, to below St. Lawrence Island, approximately 63° latitude, in the north. 1329 Near the northern boundary, the system shifts from a pelagic to benthic dominated system (e.g. 1330 Grebmeier et al., 2006; Stevenson and Lauth, 2012; Sigler et al., this issue). The shelf can be 1331 divided into regions or domains, each with their characteristic hydrography, circulation, and 1332 fauna (Iverson et al., 1979; Cooney and Coyle, 1982; Coachman, 1986; Kachel et al., 2002). 1333 Cross shelf domains are most distinct during summer and fall in the southeastern Bering Sea 1334 where there are 3 domains approximated by water depth: Outer (100 – 180 m), Middle (50 – 100 1335 m) and Inner (< 50 m). The Inner Domain is well mixed, the Middle Domain is stratified into 2 1336 layers, and the Outer Domain is stratified, but with 3 layers. In addition to cross shelf variations, 54 1337 there are latitudinal variations in wind fields, water column properties, and most importantly, sea 1338 ice coverage (Stabeno et al., 2010, 2012a). The northeastern Bering Sea is ice covered every year; 1339 whereas coverage in the south historically varies by more than 100 km (Stabeno et al., 2012a). 1340 The cold pool (a layer of < 2 °C water that resides on the shelf bottom), is formed as a result of 1341 winter cooling and mixing that most often precedes sea ice formation), and typically persists 1342 through the following summer. Its extent and magnitude varies with climatic conditions affecting 1343 the distribution of many upper trophic level organisms in the region (e.g. Wyllie-Echeverria, 1344 1998; Hollowed et al., 2012, Kotwicki and Lauth, 2013). 1345 1346 The goal of this manuscript is to describe the broad-scale spatial variations in large and small 1347 zooplankton community composition in the north and southeastern Bering Sea during warm and 1348 cold climate states. Additionally, we address several important questions regarding zooplankton 1349 ecology affected by changing climate conditions (see Table 1). The data for these analyses come 1350 from broad-scale surveys of zooplankton conducted by the Bering-Aleutian Salmon International 1351 Survey (BASIS) program at NOAA, and provide the opportunity to describe the spatial cross- 1352 shelf and latitudinal distribution of zooplankton for the entire eastern Bering Sea shelf. These 1353 surveys began in 1999 before the Bering Sea Project (Bering Sea Ecosystem Study [BEST] and 1354 Bering Sea Integrated Ecosystem Research Program [BSIERP]) was launched in 2007 (Wiese et 1355 al., 2012) and have continued into the present after the field components of BEST and BSIERP 1356 ended. During the 2000s the region experienced different multi-year climate regimes (Stabeno et 1357 al., 2012b) above average sea water temperatures and very low sea ice coverage (2000 – 2005), a 1358 single year of average sea water temperatures and sea ice extent (2006), and cold years with 1359 extensive sea ice (2007 – 2009). Thus, in contrast to most regional scientific programs in the last 1360 50 years whose timing or duration allowed limited sampling in a single ecosystem state, our 1361 dataset allows us to evaluate changes in the eastern Bering Sea shelf pelagic ecosystem during 1362 multiple thermal phases. 55 1363 2.0 Methods: 1364 2.1 Survey station locations and 1365 oceanographic sample collection 1366 From mid August to early October, 1367 2003 – 2009, samples were collected in 1368 the eastern Bering Sea, at stations 1369 located from 54.5 – 63.0 N and 159.0 1370 – 174.0 W, and spaced approximately 1371 60 km apart (Cieciel et al., 2009; 1372 Farley and Moss, 2009; Fig 1). Stations 1373 were divided into shelf regions for 1374 analyses using the Bering Sea 1375 Integrated Ecosystem Program (BSIERP) designations based on oceanographic/hydrographic, 1376 fisheries and ecosystem characterizations (Ortiz et al., 2012). We further grouped these regions 1377 into five major domains using approximately 60°N to split the shelf into south and north (Stabeno 1378 et al. 2010) and approximating cross-shelf domains defined by Coachman (1986) yielding: S 1379 Inner (< 50 m bathymetry, BSIERP regions 2 and 7), S Middle (50 – 100 m, regions 3 and 6), S 1380 Outer (100 – 200 m, region 4), N Inner (<~40 m, region11), and N Middle (~40 – 100 m, regions 1381 9 and 10). Figure 7. Bering Sea study area showing stations used for analyses in each oceanographic shelf domain for: A) large zooplankton and B) small zooplankton. Black lines indicate spatial “blocks” which are thiessen polygons around points. Blocks containing colored points (reduced data set) were visited at least two years during warm (2003 – 2005) and two years during cold (2006 – 2009) regimes; colors signify domain. 1382 1383 Vertical profiles of temperature, salinity, and chlorophyll a (Chla) fluorescence were collected at 1384 each station with a Sea-Bird 1Model 25 or Model 9plus CTD. A rosette sampler was used to 1385 obtain discrete water samples for surface and bottom nutrients, total Chla (Whatman GF/F) and 1386 large size-fractionated Chla (> 10µm, Millipore Isopore polycarbonate membrane filters); all 1 Use of trade names does not signify an endorsement by the U.S. National Oceanic and Atmospheric Administration. 56 1387 samples were stored frozen (-80 C) for 6 months maximum and analyzed using standard 1388 procedures at shore-based facilities (Parsons et al., 1984; Gordon et al., 1993). Vertical profiles of 1389 Chla concentrations were derived from regressions between in vivo fluorescence and discrete 1390 Chla samples, mean r2 = 0.65). Water column stability (energy required to mix the water column 1391 to 70 m, J m-3) was calculated at each station from CTD temperature and salinity data (Simpson et 1392 al., 1977). Ice coverage (number of days of ice at each station during the prior winter) and timing 1393 of retreat (the last day of ice in spring) for a 60 km2 square box centered on each station was 1394 obtained from the Advanced Microwave Scanning Radiometer (AMSR) on MODIS Aqua from 1395 the National Snow and Ice Data Center (NSIDC) (S. Salo, NOAA PMEL, personal comm.). We 1396 assumed no ice when concentrations dropped below 15%. Wind mixing data (wind velocity3, u*3) 1397 for August were obtained from National Centers for Environmental Prediction (NCEP) reanalysis 1398 data set (N. Bond, NOAA PMEL, personal comm.). Wind mixing data were averaged over a 2° 1399 latitude by 5° longitude box centered on a NOAA PMEL mooring at site M4 (57. 9°N, 168. 9°W; 1400 Stabeno et al., 2010). We assumed M4 winds approximated wind fields over our entire survey 1401 area, since winds at all four 70 m moorings in the northeast and southeast Bering Sea (M2, M4, 1402 M5 and M8) have shown strong coherence during recent years (Stabeno et al., 2010). Winter 1403 wind fields for the prior period (October to April) based on shelf-wide model results were used to 1404 estimate potential onshore and offshore flow, assuming that southeasterly winds promote stronger 1405 onshore flows than northwesterly winds (Danielson et al., 2012). 1406 1407 2.2 Zooplankton sample collection and lab analysis 1408 Zooplankton samples were collected and analyzed using methods described in Coyle et al. 1409 (2011). Briefly, small zooplankton assemblages were sampled at every other station with a 0.1 m2 1410 Juday net with 168 µm mesh (Volkov, 1984; Volkov et al., 2007). The net was towed vertically 1411 from within 5 – 10 m of the bottom (or maximum depth of 200 m) to the surface at about 1 m s-1. 1412 Juday net samples were sieved into size fractions prior to counting on board ship (Volkov et al. 57 1413 2007); however, data from all size fractions combined were used to quantify small zooplankton 1414 taxa. Large zooplankton assemblages were collected at every station with a 60 cm MARMAP- 1415 style bongo frame with a 505 µm mesh net. Oblique tows were conducted from the surface to 1416 within 5–10 m of the bottom, and volume filtered was measured with calibrated General Oceanics 1417 flowmeters. All samples were preserved in 5% formalin: buffered with seawater. Zooplankton 1418 collections from 2003 – 2004 were sorted at the Polish Plankton Sorting and Identification Center 1419 (Szczecin, Poland); collections from 2005 – 2009 were processed at the University of Alaska 1420 (Coyle et al., 2008). The lowest taxonomic level of sorting varied between labs, so we used the 1421 lowest taxonomic stage available across all years. Zooplankton tows were collected primarily 1422 during the daytime. Many euphausiids on the Bering Sea shelf stay within 1 – 2 m of the bottom 1423 during the day (Coyle and Pinchuk, 2002) and those in the water column are difficult to 1424 quantitatively capture with nets (e.g. Sameoto et al., 1993; Wiebe et al., 2004); therefore 1425 euphausiids were not included in analyses. It is also possible that under representation of other 1426 diel migrants (e.g. Metridia spp.) occurred. The small volume filtered by Juday and bongo nets 1427 compared to larger nets (e.g. Multinet or MOCNESS) could have resulted in under sampling of 1428 relatively large and rare taxa. Accordingly, very large medusae such as Chrysaora melanaster 1429 were excluded since they were not quantitatively sampled by our bongo nets. Unidentified 1430 copepod nauplii were also excluded, although they were fairly numerous in the Juday net samples 1431 (~ 11% of total abundance). A total of 424 Juday and 717 bongo net samples were collected for 1432 all years combined. 1433 1434 2.3 Statistical analyses 1435 Separate analyses were conducted for large (bongo samples) and small (Juday samples) 1436 zooplankton taxa. Zooplankton data were 4th root transformed (unless otherwise indicated), to 1437 down weight the contribution of dominant taxa to similarity/dissimilarity patterns (Clarke and 1438 Warwick, 2001). Initial analyses were conducted using both abundance (no. m-3) and integrated 58 1439 abundance (no. m-2) for comparisons among regions with large changes in bathymetry (Questions 1440 1 and 2, Table 1). We found no substantial differences in the outcome when using number 1441 normalized by volume or area; therefore we chose to present abundances normalized by volume 1442 filtered (no. m-3) to allow for comparisons with most prior studies on the eastern Bering Sea shelf 1443 which reported their abundance data normalized by volume filtered. Zooplankton biomass wet 1444 weights (g m-3, Coyle et al., 2011) were used in Question 2 analyses. The warm regime was 1445 designated as 2003 – 2005 and the cold regime as 2006 – 2009 based on late summer /fall mean 1446 water column temperature anomalies for 2003 – 2009, estimated from CTD data collected during 1447 our surveys (cf. Stabeno et al., 2012b, which categorizes 2006 as a year of average temperatures 1448 and sea ice extent). For all analyses, excluding those for Question 1, it was necessary to account 1449 for uneven sampling across years. Therefore, a reduced data set was used that included only 1450 station locations within equally spaced geographically partitioned regions (Blocks) that were 1451 sampled a minimum of two warm and two cold years (Fig. 1). Blocks were created in 1452 ArcMap/ArcInfo, 10.1 (ESRI, 2012) using thiessen polygons, polygons constructed around 1453 stations by calculating the midline between each pair of adjacent stations (Thiessen and Alter, 1454 1911; Gold, 1991). Blocks were approximately 60 x 60 km for the large taxa (equivalent to the 1455 spacing of stations) and approximately 80 x 80 km for the small (equivalent to the diagonal 1456 spacing of every other station). The small taxa required larger blocks due to sparser sampling. 1457 Separate analysis were conducted for each of the five major domains (S Middle, N Middle, S 1458 Inner, N Inner, S Outer) for Questions 3and 4 analyses (Table 1). 1459 1460 Statistical analyses included cluster analysis, similarity percentage contribution of individual taxa 1461 (SIMPER), and permutational, multivariate analysis of variance (PERMANOVA) in the software 1462 package, PRIMER-E,version 6.1.15 with PERMANOVA+ version 1.0.5 (Clarke and Warwick, 1463 2001; Clarke and Gorley, 2006; Anderson et al., 2008). All procedures used Bray-Curtis 1464 dissimilarities, a statistic commonly used to quantify the dissimilarity in composition between 59 1465 samples (Bray and Curtis 1957). In our study, these procedures were used to evaluate variations 1466 in community composition (the relative abundances of zooplankton taxa). SIMPER calculates the 1467 average contribution of each taxon to the overall Bray-Curtis dissimilarity in community 1468 composition. PERMANOVA allows the use of distance measures, in our case, Bray-Curtis 1469 dissimilarity in community composition, to test for the significance of specific effects, similar to a 1470 multivariate ANOVA. The advantage of using a permutation test is that it does not require the 1471 data to follow a particular distribution and is therefore more robust than parametric statistics. This 1472 procedure also allows for the partitioning of the multivariate variation in abundance according to 1473 nested sampling designs and 2-way designs with interactions, as well as the addition of 1474 covariates, such as environmental variables (Anderson et al. 2008). Statistical procedures used for 1475 each of our research questions are detailed below. 1476 1477 Q1) The taxonomic composition and distribution of zooplankton were evaluated qualitatively 1478 using abundance data from all stations and years. For grouping stations, we applied a hierarchical 1479 cluster analysis using group average linkage of Bray-Curtis dissimilarities among stations. We 1480 then used SIMPER analysis to determine the average percent contribution of each taxa in each 1481 station cluster. For grouping taxa, we applied a separate cluster analysis of Bray-Curtis 1482 dissimilarities using mean abundance (standardized untransformed data) of each taxon across 1483 station clusters. Standardization rather than transformation was necessary for grouping species 1484 since we were more interested in relative similarity/dissimilarity patterns rather than down 1485 weighting abundant species, which was necessary when grouping stations (Clarke and Warwick, 1486 2001).Clusters were identified by drawing a line across branches on each dendrogram (one for 1487 station and one for taxa) determined by at least 50% similarity, length of branches (indicating 1488 stability of groups), a permutation procedure that objectively tests for grouping a priori 1489 (SIMPROF, Clarke and Warwick, 2001; Clarke and Gorley, 2006), and subjective biological 60 1490 interpretation of clusters. For the station cluster analysis, we designated station clusters as outliers 1491 if they contained less than 1% of the sampled stations. 1492 1493 Q2) The total abundance and total biomass of all large and all small zooplankton in the north and 1494 south eastern Bering Sea for each year were estimated by summing the abundance or biomass 1495 over all taxa within each block and dividing by number of stations. The average abundance or 1496 biomass was computed over all blocks within each region, i.e. the south (< 60 °N, Inner, Middle 1497 and Outer Domains) and north (60 – 63 °N, Inner and Middle Domains). Then, we estimated the 1498 mean and standard error (SE) of the 4th root transformed data, then back-transformed the data to 1499 estimate geometric means and SE of untransformed total abundance and biomass. 1500 1501 Q3a) To test for significant differences in zooplankton community composition between Regimes 1502 (warm vs. cold), a PERMANOVA was applied to all domains combined and then to each domain 1503 separately. In addition to the Regime effect, a geographic blocking factor (Block) was included as 1504 a random effect to account for the uneven sampling across years (defined above). Year was also 1505 included in the model as a random nested effect within Regime. Interactions among these effects 1506 were also included. A permutational dispersion (PERMDISP) analysis was applied to test for any 1507 significant dispersion effects between Regimes (Anderson et al. 2008). This is analogous to a 1508 univariate Levene test for homogeneity of variances in ANOVA and tests for differences in 1509 distances between the yearly centroids (center of points in multivariate space). 1510 1511 Q3b) A 2-way (Regime and Block) SIMPER analysis was run to determine which zooplankton 1512 taxa contributed most to the differences between regimes for domains where significant 1513 differences were found between Regimes. 1514 61 1515 Q3c) A PERMANOVA was applied to each domain to determine if zooplankton communities 1516 were significantly different for one or more years. In this case, Year was considered a fixed 1517 effect, and Block was considered a random effect accounting for spatial variability and the 1518 uneven sampling across years. To determine which consecutive years had significantly different 1519 community composition, pairwise multiple comparison tests were applied to consecutive years 1520 only. No Bonferroni correction was applied here as PERMANOVA provides exact tests, and a 1521 Bonferroni correction would be overly conservative in this case (Anderson et al., 2008). 1522 1523 Q3d) The mean untransformed zooplankton abundances for the most common taxa (ones that 1524 jointly contributed to ≥ 97% of the total untransformed abundance for all years combined) were 1525 calculated for the north and southeastern Bering Sea to determine which taxa contributed to the 1526 interannual variations in community structure and total abundance. 1527 1528 Q4) A set of local environmental variables measured concurrently at the same stations as 1529 zooplankton abundance and regional variables measured yearly over the eastern Bering Sea were 1530 used to evaluate the conditions most often associated with variations in community composition 1531 (dependent variable), for warm vs. cold Regimes within each shelf domain. Variables were 1532 removed if they were correlated with other variables (R > 0.8) or were determined to be of lesser 1533 biological importance. The variables of interest were included as covariates (independent 1534 variables) in PERMANOVA tests for domains where significant differences were found between 1535 warm and cold years (based on results of Question 3a). The local covariates included temperature, 1536 Chla (a measure of phytoplankton biomass), nitrate, and ammonium measured at the surface (5 1537 m) or above the mixed layer depth; stability over the top 70 m; integrated Chla and mean large 1538 size-fractionated Chla/total Chla over the top 50 m; day of ice retreat in spring prior to our late 1539 summer/ fall sampling; and temperature, salinity, and ammonium measured at the bottom or 1540 below the mixed layer depth. The regional covariate tested was the yearly winter (October-April) 62 1541 wind direction (% of time winds were from the SE and NW; Danielson et al., 2012). We also 1542 tested latitude and longitude to see if there were significant interannual geographic gradients 1543 within oceanographic domains. 1544 1545 We first removed the within year 1546 spatial variability by including a -------------------------Middle Shelf ------------------------------------Inner Shelf-----------------Outer Shelf --- Large Zooplankton Clusters of Stations Eukrohnia hamata Calanus pacificus Neocalanus spp. Eucalanus bungii Metridia pacifica Ctenophora Clione limacina Caridea Epilabidocera amphitrites Anomura Mysida Limacina helicina Brachyura Hyperiidae Gammaridae Cumacea Cnidaria Parasagitta elegans Calanus spp. Appendicularia predominant regime L1 L2 L3 L4 L5 L6 L7 L8 W W/C W C W/C C W/C C blocking factor (described 1548 previously). Then, each covariate 1549 was sequentially added using 1550 Type I sum of squares which 1551 tests for the significance of the 1552 addition of that variable with all 1553 prior covariates accounted for. 1554 The best fit model was chosen 1555 by including the most significant 1556 covariates in a particular order 1557 that gave the lowest mean square 1558 error. The Regime effect and 1559 interactions were included as the 1560 last covariates in the model. Whenever a Regime effect (added last) became insignificant (P > 1561 0.05) compared to the Regime effect before adding environmental covariates (P < 0.05), we 1562 concluded that the environmental covariates were explaining at least some of the variability in 1563 community composition between warm and cold years. To examine yearly changes in 1564 temperature and sea ice variations we plotted surface and bottom temperature, day of sea ice 1565 retreat and number of days of ice coverage the prior winter, averaged over stations in the N and S 1566 Middle Domains. Large Zooplankton Clusters of Taxa 1547 Abundance (no. m¯³)0.25 =0 0.1 - 0.5 0.5 - 1.0 1.0 - 2.0 2.0 - 3.0 >3 Figure 8. Heat map of large zooplankton showing clusters of stations (x axis) by taxa (y axis) using all stations sampled. Shade of each cell indicates the 4th root transformed abundance of each taxon. Approximate domain designations are shown above clusters. Designations at bottom indicate climate regime (W = warm; C = cold; W/C = both) when each cluster was commonly observed. Horizontal lines separate clusters of species (indicating taxa which tended to co-occur over all years and domains combined). 63 1567 1568 3.0 Results: 1569 3.1 Spatial and temporal patterns of large and small zooplankton (Q1). 1570 The clusters of stations (station clusters) show which stations had similar taxonomic 1571 compositions over the eastern Bering Sea shelf for all years combined. For large 1572 1573 1574 1575 Figure 3. Spatial variation of large zooplankton clusters for each year using 4th root transformed abundance data. Warm years highlighted with red and cold with blue. Refer to Fig. 2 to see taxa abundances for each cluster. Intended for color reproduction on the Web and in print 64 1576 1577 zooplankton, the multivariate analyses yielded eight station clusters (Fig. 2, 3). The taxa that 1578 made up approximately 80% of the total abundance are listed from high to low for the clusters 1579 described below. One pattern that stands out is the replacement of Middle Domain clusters during 1580 the transition from warm to cold years (Fig. 3). Cluster L3 characterized by Cnidaria (gelatinous 1581 zooplankton dominated by Aglantha digitale in our study), Parasagitta elegans (Chaetognatha) 1582 and Appendicularia (Larvacea) extended over a broad area in the warm years, particularly 2003 – 1583 2004, and was replaced in cold years by Cluster L4 characterized by Calanus spp.2 (Copepoda), 1584 Cnidaria and P. elegans. In addition, in warm years, there was a cluster confined primarily to the 1585 Inner Domain, Cluster L1, characterized by P. elegans, Cnidaria, Epilabidocera amphitrites 1586 (Copepoda) and Caridea (shrimp), but in cold years this group was observed less often. 1587 Latitudinal variations in the timing of changes were also seen, with increased coverage for 1588 Cluster L4 in the northeastern Bering Sea in 2004 – 2005, and expansion southward and inshore 1589 into the Inner Domain in the following cold years. In contrast, in the Outer Domain, Cluster L8 1590 characterized by Limacina helicina (Pteropoda), Calanus spp., Eucalanus bungii (Copepoda), and 1591 Metridia pacifica (Copepoda), was observed during all years. 1592 1593 For small zooplankton, the analyses yielded nine station clusters (Fig. 4 and 5). Similar to large 1594 taxa, in the Middle Domain there was one cluster with broad spatial coverage in warm years, 1595 Cluster S4, characterized by Oithona spp.(Copepoda), Pseudocalanus spp.(Copepoda), 1596 Echinodermata larvae, Polychaeta (annelid worms), molluscan Bivalvia larvae and Cirripedia 1597 (barnacle larvae), which was replaced in cold years by Cluster S7, characterized by 1598 2 Calanus spp. referred to in this work is most likely a mixture of C. marshallae and C. glacialis based on recent genetic analyses by Campbell et al., personal comm. and J. Nelson (2009). 65 Pseudocalanus spp., Oithona spp., 1600 Acartia spp. (Copepoda) and 1601 Polychaeta (Fig. 5), with fewer taxa 1602 making up the bulk of abundance in 1603 cold years. In the Inner Domain, a 1604 cluster with broad coverage in the 1605 southeastern Bering Sea, Cluster 1606 S1, characterized by Oithona spp., ----------- Middle Shelf ---------------------------------- Inner Shelf -------------------- Outer Shelf Small Zooplankton Clusters of Taxa 1599 Metridia spp. copepodites Globigerina sp. Tortanus discaudatus Eurytemora herdmani Podon sp. Evadne sp. Harpacticoida Cirripedia Echinodermata Microcalanus pygmaeus Centropages abdominalis Bivalvia Pseudocalanus spp. Acartia spp. Oithona spp. Polychaeta Fritillaria sp. Scolecithricella sp. predominant regime S1 Small Zooplankton Clusters of Stations S2 S3 S4 S5 S6 S7 S8 W W C W W C C S9 W/C W/C Abundance (no. m¯³)0.25 =0 0-1 1-2 2-4 4-6 >6 1607 Pseudocalanus spp., larval 1608 Bivalvia, Centropages abdominalis 1609 (Copepoda), Polychaeta, Acartia 1610 spp., Echinodermata larvae and Podon sp. (Cladocera) was partially replaced in the coldest years, 1611 2008 – 2009, by Cluster S6, characterized by Pseudocalanus spp. and Acartia spp. and C. 1612 abdominalis and by Cluster S7. Again, similar to the large taxa, in the Outer Domain there was 1613 one cluster common during all years, Cluster S9, characterized by Oithona spp., Pseudocalanus 1614 spp., Metridia spp. copepodites, Acartia spp., Microcalanus spp. (Copepoda), and Bivalvia 1615 larvae. Small taxa clusters did not show large north – south variations. Figure 4. Heat map of small zooplankton clusters of stations (x axis) by taxa (y axis). Refer to Fig. 2 for details. 1616 66 1617 Clusters of co-occurring taxa (taxa clusters) 1618 show how taxa grouped together over all 1619 years and spatial areas combined (Fig. 2, 4, 1620 y-axis). Large zooplankton taxa that tended 1621 to co-occur included Neocalanus spp., E. 1622 bungii, M. pacifica; these are oceanic 1623 copepods resident in the Outer Domain or 1624 brought in from beyond the shelf break 1625 (Gibson et al. 2013). Other taxa that co- 1626 occurred were P. elegans, Calanus spp., and 1627 Appendicularia; all are common taxa with 1628 relatively high abundances in the Middle 1629 Domain (Coyle et al. 2008, 2011). Small taxa 1630 that tended to co-occur included Evadne sp. 1631 (Cladocera), Podon sp., Eurytemora 1632 herdmani (Copepoda), and Tortanus 1633 discaudatus (Copepoda), taxa typically 1634 neritic and common nearshore (Gieskes 1971, Johnson 1934, Kos 1977); and Oithona spp., 1635 Acartia spp., Pseudocalanus spp., Polychaeta, Bivalvia larvae, and C. abdominalis, taxa that 1636 often reside in upper layers of the water column (Marlowe and Miller, 1975). Figure 5. Spatial variation of small zooplankton clusters by year using 4th root transformed abundance data. Warm years highlighted with red and cold with blue. Refer to Fig. 4 to see taxa abundances for each cluster. Intended for color reproduction on the Web and in print. 1637 1638 3.2 Total abundance and total biomass of large and small zooplankton during warm and cold 1639 regimes (Q2). 1640 Trends in total abundance and biomass varied for large and small zooplankton and between the 1641 north and south eastern Bering Sea (Fig. 6). In the south, large zooplankton increased in 1642 abundance starting in 2007, peaked in 2008 and declined in 2009; in contrast, small zooplankton 67 1643 steadily decreased in abundance 1644 from 2003 – 2009 (Fig. 6A). In the 1645 north, total abundances of large 1646 zooplankton increased between 2004 1647 and 2005 and decreased between 1648 2007 and 2009; small zooplankton 1649 abundances did not show clear 1650 trends with time, and were only 1651 noticeably higher in 2004 than in 1652 other years (Fig. 6B). The overall 1653 magnitude of large zooplankton abundances was similar in the north and south, however, for 1654 small zooplankton, abundances were lower in the north compared to the south. Trends in biomass 1655 mirrored those in abundance except for small taxa in the south where despite the observed 1656 declines in abundance from 2003 to 2009, the biomass remained constant (Fig. 6C, D). It is also 1657 worth noting that in spite of high abundance of small taxa during the warm years, their biomass 1658 was similar to that of large taxa during the warm period in the south and during all years in the 1659 north (Fig. 6). Figure 6. Small and large zooplankton total mean abundance (no. m-3) in the eastern Bering Sea in the A) south (< ~60 N) and B) north (~60-63 N), and total mean biomass (g m-3) in the C) south and D) north. Geometric means and standard errors are shown. 1660 1661 3.3 Statistical differences among assemblages, shelf domains, and years (Q3) 1662 3.3.1 Zooplankton abundances between regimes by domain (Q3a) 1663 We next evaluated if the observed differences in zooplankton assemblages between regimes were 1664 statistically significant for the five major shelf domains. For all domains combined, 1665 PERMANOVAs showed significant interactions between Regime and Domain (P = 0.012, P = 1666 0.001, for large and small taxa, respectively), indicating that community structure varied by 1667 Regime depending on Domain. Therefore, each domain was evaluated separately for all further 1668 analyses. There were significant differences in the community structure between warm and cold 68 1669 regimes for all domains except the N Inner for large taxa and N Inner and S Outer for small taxa, 1670 after accounting for significant spatial block effects (Table 2). The strongest differences were 1671 observed in the S Middle and S Inner Domains. 1672 1673 3.3.2 Zooplankton taxa differences between warm and cold regimes (Q3b) 1674 Large zooplankton taxa that substantially decreased in abundance (> 2-fold) during the transition 1675 from warm to cold regimes include Cnidaria, Gammaridae amphipods, Mysida, the neritic 1676 copepod Epilabidocera amphitrites, and the temperate copepod Calanus pacificus (Table 3). 1677 Taxa that substantially increased in abundance from warm and cold regimes include Anomuran 1678 and Brachyuran crab larvae, Calanus spp., Parasagitta elegans, the oceanic copepods 1679 Neocalanus spp. Eucalanus bungii, Hyperiidea amphipods and Cumacea. Taxa that showed both 1680 increases and decreases depending on domain include Appendicularia, Caridea decapods and 1681 Limacina helicina. Calanus spp. were the top contributor to changes in community composition 1682 between regimes (13% – 23% of total dissimilarity), particularly in the Middle Domain (both 1683 north and south). High contributions to dissimilarity (> 10%) were also seen for Caridea decapods 1684 in the S Inner, P. elegans and Appendicularia in the S Middle; Limacina helicina in the S Outer, 1685 and Appendicularia and Cnidaria in the N Middle Domains. 1686 1687 Most small zooplankton taxa decreased in abundance in the S Inner, S Middle and N Middle 1688 Domain from warm to cold years (Table 3). The only small taxa showing increases in cold years 1689 were Acartia spp. in all three domains, Metridia spp. copepodites and Pseudocalanus spp. in the 1690 S Middle Domain, and Fritillaria spp. (Appendicularia) in the N Middle Domain. Oithona spp. 1691 and larval bivalves had high contributions to dissimilarity in community composition between 1692 regimes for all three domains, with Acartia spp., Polychaeta, and Echinodermata larvae also 1693 important, particularly in the Middle Domain. 1694 69 1695 3.3.3 Zooplankton abundances between consecutive years (Q3c and d) 1696 Next, we evaluated which consecutive years showed the greatest changes in community 1697 composition in each domain to determine if the differences between regimes occurred during our 1698 designated break point between warm to cold regimes (i.e. between 2005 and 2006) or if the 1699 changes in community composition 1700 took place over a period of years. 1701 We also evaluated which taxa 1702 showed the largest interannual 1703 variations in the south and north 1704 eastern Bering Sea. 1705 1706 For large and small zooplankton 1707 communities, most regions with the 1708 exception of the S Outer showed 1709 significant differences (based on 1710 pairwise multiple comparisons) 1711 between pairs of consecutive years 1712 (Table 4) indicating that interannual 1713 changes in community composition 1714 were observed over our entire study 1715 period, not just for the years when the temperature regime changed most rapidly from warm to 1716 cold (eg. 2005-2007 in south). For large zooplankton, the greatest interannual variations, in the 1717 south were seen by Calanus spp., which increased in cold years starting in 2007 (Fig. 7); while in 1718 the north, increases were due to Cnidaria in 2005 and 2007, and Calanus spp. starting in 2006. 1719 The small taxa that decreased in abundance from warm to cold years in the south were Oithona 1720 spp. and Bivalvia larvae, while in the north, Oithona spp. decreased only in 2005, 2007 and 2009 Figure 7. Untransformed mean zooplankton abundances in the eastern Bering Sea for large taxa in the A) south (< ~60 south and D) north. Intended for color reproduction on the Web and in print. 70 1721 (Fig. 7). Small zooplankton composition in the north was most different in 2009, with higher 1722 levels of polychaetes and Acartia 1723 spp.; no data exist for 2008 to 1724 determine if this change was 1725 gradual. Overall, L. helicina, M. 1726 pacifica, Pseudocalanus spp., and 1727 Bivalvia larvae appear to be more 1728 abundant in the south, and Cnidaria 1729 and Polychaeta more abundant in 1730 the north. Some of these north – 1731 south variations can be explained by 1732 the lack of sampling in the Outer 1733 Domain in the northeastern Bering 1734 Sea. 1735 1736 3.4 Environmental factors 1737 contributing to observed differences 1738 in zooplankton community 1739 composition (Q4) 1740 The interannual variations in 1741 zooplankton abundance and 1742 community composition appear to relate to changes in temperature and sea ice retreat in the 1743 eastern Bering Sea. Temperature above and below the mixed layer depth in the N and S Middle 1744 Domains showed a declining trend from 2003 – 2009, concurrent with increases in the date of ice 1745 retreat and duration of ice coverage (Fig. 8). The greatest difference in timing of ice retreat and 1746 ice coverage was found in the S Middle Domain, with increases seen from 2005 – 2008, which Figure 8. A) Mean temperature above (surface) and below (deep) the mixed layer depth during our surveys, B) mean day of year for sea ice retreat during the prior spring, and C) mean number of days of sea ice coverage during the prior winter, in the south (S) and north (N) Middle Domains. Standard errors are shown. 71 1747 overlaps the increase in Calanus spp. (Fig. 7, 8). Decreases in surface and bottom temperatures 1748 coincided with decreases in small taxa such as Oithona spp. and increases in Pseudocalanus spp. 1749 and Acartia spp. in the S Inner and S Middle Domains. Temperatures and small taxa abundance 1750 were lower in the north than the south (Fig. 7, 8). For large zooplankton taxa, models with 1751 environmental co-variates were constructed for the S Inner, S Middle, S Outer and N Middle 1752 Domains (where significant Regime effect was found). Environmental covariates were able to 1753 account for at least some of the variability in zooplankton community composition between 1754 regimes for all but the S Middle Domain based on P values for Regime with and without 1755 environmental covariates (Table 5 vs. 2). Physical and geographic covariates explained the 1756 greatest amount of variability in large zooplankton community composition (Table 5). 1757 Temperature below the mixed layer depth was important in the N and S Middle Domains 1758 although temperature above the mixed layer depth, salinity below the mixed layer depth, ice 1759 retreat timing, latitude and longitude were also highly significant (P < 0.01), depending on 1760 domain (Table 5). For small taxa, environmental covariates explained some of the variability in 1761 community composition between regimes in the S Inner, S Middle and N Middle Domains (Table 1762 2 vs. 5). Similar to large taxa, physical covariates were highly significant, with temperature above 1763 and below the mixed layer depth important for the S Inner and S Middle Domains as well as 1764 stability for the S Middle Domain (Table 5). Overall, temperature explained the greatest amount 1765 of temporal variability in community structure, and was significant in models for all domains 1766 except the S Outer for large and the N Middle for small zooplankton. Most of the biologically 1767 related covariates, nutrients (ammonium and nitrate) at surface and bottom, surface Chla, 1768 percentage of large phytoplankton, and August wind mixing, were not significant. 1769 1770 1771 1772 72 1773 4.0 Discussion: 1774 4. 1 Overview 1775 This study provides a rare look into how stanzas of warm and cold years affect the zooplankton 1776 community structure in both the north and south eastern Bering Sea and the environmental 1777 variables associated with these changes. Our analyses demonstrate the considerable cross-shelf 1778 variability in zooplankton communities as seen in prior studies (e.g. Cooney and Coyle, 1982), 1779 but also shows variability between assemblages in the north and south both in timing of changes 1780 in abundance of key taxa and total abundance and biomass. Water temperature showed the 1781 strongest relationship with large and small zooplankton community structure. We hypothesize 1782 that large taxa abundance has a direct or indirect relationship to bottom temperature (depending 1783 on taxa) and a direct relationship to the winter/spring southerly extent of sea ice; while small taxa 1784 abundance has a direct relationship to surface and bottom temperature. Ice algae and sea-ice 1785 related phytoplankton blooms in cold years may provide an early food source for large 1786 zooplankton on the southern shelf (Siger et al. this issue). In contrast, lower temperatures may 1787 reduce temperature-dependent growth (e.g. Hirst and Lampitt 1998), negatively impacting 1788 reproduction rates of small zooplankton with fewer cohorts produced during the growing season. 1789 However, temperature may also affect zooplankton survival, so multiple generations do not 1790 guarantee higher abundances. The observed changes in community composition were not abrupt, 1791 but showed gradual changes over a period of years, concordant with changes in sea ice and water 1792 temperature. Species-specific responses in cold years include both increases in omnivorous 1793 Calanus spp., Neocalanus spp., and in predators such as P. elegans, and decreases (with large 1794 interannual variability) in predatory Cnidaria (A.digitale). 1795 1796 4.2 Temporal and spatial patterns in community structure between regimes 1797 Assemblages often were clustered together based on domain with separate groups of oceanic (e.g. 1798 Neocalanus spp., Metridia pacifica, Eucalanus bungii) and neritic taxa (Podon sp., Evadne sp., 73 1799 Polychaeta, Bivalvia larvae). These differences are attributed to, and maintained by, the presence 1800 of hydrographic fronts that separate the domains during summer and the consistency of 1801 hydrographic and biological processes that are inherent to the different domains (Iverson et al., 1802 1979). The extension of a Middle Domain cluster into the Inner Domain in cold years for large 1803 zooplankton , suggests that the Inner Front may not be as strong (or is more leaky) in cold years, 1804 possibly resulting in increased transport across the Inner Shelf. Alternatively, the Inner Front may 1805 be established later in the season in cold than in warm years allowing more time for Middle 1806 Domain taxa to become established in the areas that eventually become the Inner Domain. 1807 Previous studies suggest the Inner Front was established later in the season during years with low 1808 sea surface temperatures (SSTs) (1998 and 1999), due to storminess and late ice retreat, than 1809 during a year with high SSTs (1997) (Kachel et al., 2002). Additionally, winter wind direction 1810 was a significant co-variate for large zooplankton in the S Inner Domain, suggesting the 1811 differences in wind fields may be related to onshore transport or timing of set up for the Inner 1812 Front (Kachel, et al. 2002, Danielson et al., 2012). Inshore – offshore changes in centers of 1813 distribution within domains are also suggested by the importance of latitude or longitude in our 1814 best-fit models for small taxa within the N and S Middle Domains and large taxa within the N 1815 Middle and S Outer Domains. 1816 1817 The recent regime shifts influenced large and small zooplankton community structure over most 1818 shelf domains, but variations were greatest in the S Middle Domain, where the largest physical 1819 changes (e.g. sea ice retreat timing, temperature) were observed (Stabeno et al., 2012a). The high 1820 significance of surface temperature (and higher abundances) for small taxa in the south but not in 1821 the north may be because in the north, the Middle Domain summer surface temperature oscillated 1822 around 8 ºC, whereas in the south, it declined from almost 12 ºC in 2004 to below 8 ºC in 2009. 1823 The surface layer may be where small taxa predominate, as found in the Gulf of Alaska (Marlowe 1824 and Miller, 1975). 74 1825 1826 North – south variations in Middle Domain zooplankton assemblages were also lower in cold 1827 years compared to a single warm year (2005) (Stabeno et al., 2012a). We found a similar result 1828 for 2005, but the other warm years (2003, 2004) showed much lower latitudinal variations in 1829 zooplankton assemblages. These differences may relate to interannual variability in the N-S 1830 strength of the temperature gradients. After the recent period of extended warming, even the north 1831 received heat so that in 2003 and 2004 the surface temperature gradients between the north and 1832 south were diminished. However, the north in 2005 was colder than in 2003 – 2004, while the 1833 south in 2005 was still warm (Danielson et al., 2011). 1834 1835 1836 4.3 Interannual variations in total abundance and biomass 1837 Total abundance and total biomass of zooplankton also varied by N – S region and regime. In the 1838 south, total abundances of large taxa were highest in cold years, and similarly total biomass for 1839 large zooplankton taxa increased 5 – 10 fold between 2005 – 2007 and 2008 – 2009. This is 1840 relevant to the feeding of small age-0 fishes, such as Walleye Pollock (Gadus chalcogramma, 1841 herein referred to as pollock). The improvement in age-0 pollock condition in cold years as 1842 evidenced by increased total energy content (kJ gm-1 wet weight) is attributed to the higher 1843 availability of large, lipid bearing zooplankton prey such as Calanus spp. and Thysanoessa spp. 1844 (Euphausiacea) in cold years (Coyle et al. 2011, Hunt et al., 2011; Heintz et al., 2013). Calanus 1845 spp., and Neocalanus spp. are also important prey for other forage fish such as juvenile Pacific 1846 salmon (Oncorhynchus spp.), Pacific Herring (Clupea pallasi) and Capelin (Mallotus villosus), 1847 planktivorous sea birds and endangered baleen whales such as the North Pacific Right Whale 1848 (Eubalaena japonica) (Russell et al., 1999; Springer et al., 2008; Sheffield-Guy et al., 2009; 1849 Coyle et al., 2011; Wade et al. 2011; Heintz et al., 2013). Therefore decreases in large lipid-rich 1850 crustacean zooplankton in warm years may be detrimental to a variety of predators, not just 75 1851 pollock. In contrast to large taxa, biomass for small zooplankton in the south eastern Bering Sea 1852 was not different between regimes, even though abundances were lower in cold than warm years. 1853 This indicates that reductions in abundances of very small taxa such as Oithona spp. were 1854 compensated for by increases in abundances of larger taxa such as Pseudocalanus spp. and 1855 Acartia spp. Future work is needed to determine its impact on trophic transfer and production at 1856 higher trophic levels. 1857 1858 The north eastern Bering Sea did not follow the same pattern as the south. In the north, the 1859 abundance of large taxa increased 4-fold between 2004 and 2005, and in 2007 reached the same 1860 levels as observed in the south a year later (2008). Increases in the north were due to both 1861 Cnidaria and Calanus spp., while in the south they were due primarily to Calanus spp. Unlike in 1862 the south, total biomass for large zooplankton remained fairly constant in the north, even though 1863 taxonomic composition changed. The abundance and biomass of small taxa in the north declined 1864 from 2004 to 2005, coincident with a drop in water temperature, and then remained at that level 1865 for the remainder of our time series. 1866 1867 Total zooplankton biomass (sum of the small and large taxa) was highest in the coldest years 1868 (2008 – 2009) in the south due to increases in large zooplankton biomass, but remained constant 1869 across all years in the north. Large and small taxa contributed equally to total biomass except in 1870 the south in 2008-2009, where the contribution to total biomass was 2 – 4 times higher for large 1871 compared to small taxa. This result for the southeastern Bering Sea was previously shown by 1872 Coyle et al. (2011), however biomass levels for the north and their relation to the south in warm 1873 and cold years is new. This suggests that as the climate warms, the total biomass available for 1874 higher trophic levels will decrease in the south, but may not change in the northeastern Bering 1875 Sea. 1876 76 1877 4.4 Proposed mechanisms driving variations in abundances of key taxa 1878 Finally, to synthesize the information on changes in abundances and the environmental factors 1879 responsible for these changes between regimes and north – south regions, we describe potential 1880 mechanisms for some of the key herbivorous and predatory zooplankton taxa: Calanus spp., 1881 Neocalanus spp., P. elegans and A. digitale (Cnidaria). 1882 1883 Calanus spp. had the greatest impact on large zooplankton community structure between warm 1884 and cold regimes. Increases in Calanus spp. in cold years have been documented for the 1885 southeastern Bering Sea (Hunt et al., 2008; Coyle et al. 2011; Hunt et al. 2011, Stabeno et al., 1886 2008, 2010, 2012b), but data were not available for the north. One proposed mechanism driving 1887 increases in cold years in the south is that when ice algae are available there is an early source of 1888 food for Calanus spp. reproduction (Hunt et al., 2002; Baier and Napp, 2003, Sigler et al., this 1889 issue). An early season ice-associated phytoplankton bloom can also provide a source of energy 1890 for metamorphosis and growth (Baier and Napp, 2003; Sigler et al., this issue). Lower winter 1891 water temperatures will also lower metabolic rates which allow Calanus spp. to reduce the energy 1892 required for survival. Therefore, a series of cold years may facilitate increases in spawning 1893 biomass and reproductive output resulting in a stronger year class (Sigler et al., this issue). A 1894 corollary of this is that warmer summer temperatures lead to increased food limitation due to 1895 increased metabolic requirements and reductions in net primary production (Lomas et al., 2012), 1896 which result in reductions in zooplankton lipid (energy) stores, poor overwinter survival, and 1897 fewer copepodites produced the following spring (Coyle et al., 2011). 1898 1899 In the southeastern Bering Sea, late summer abundance of Calanus spp. increased in 2007, the 1900 first year the majority of the S Middle Domain was ice covered, so the availability of an early 1901 spring algal food source, may have been important in this region. In the north, increases in ice 1902 coverage and retreat were seen from 2003 – 2006, but had a smaller range of variability than seen 77 1903 in the south. We do not yet understand what processes promoted increases in Calanus spp. in the 1904 northeastern Bering Sea starting in 2006; because this region was at least partially ice covered 1905 until mid-April in all years and presumably ice algae were available all years. Finally, because the 1906 Calanus spp. we sampled were a combination of Calanus marshallae and Calanus glacialis, 1907 changes in advection in the north and south of these different taxa may also be at play, and these 1908 two taxa may respond to changes in climate and temperature in different ways. C. marshallae was 1909 previously thought to be dominant in the southeastern Bering Sea, while C. glacialis dominated 1910 further northward in the Bering, Chukchi and Beaufort Seas (Frost, 1974). The highest 1911 abundances of Calanus spp. were observed in 2007 in the north and in 2008 in the south, a year 1912 after Calanus spp. began to increase in each region. This suggests that sequential years of cold 1913 conditions are advantageous to build large biomass of the species. Decreases in Calanus spp. in 1914 2009 in the current study could be due to increases in predation from Themisto libellula, an arctic 1915 hyperid amphipod (Pinchuk et al., 2013). 1916 1917 Parasagitta elegans also increased in cold years with the largest abundances observed in 2007. P. 1918 elegans is a voracious predator of the early developmental stages of large and small copepods in 1919 the eastern Bering Sea (Baier and Terazaki, 2005), so this increase may be a response to increases 1920 in prey availability. On the south eastern shelf for 1995 – 1999, the most frequently consumed 1921 prey of P. elegans was Calanus nauplii in cold icy years when Calanus spp. were in higher 1922 abundance, with Pseudocalanus spp. predominantly consumed in relatively warmer years (Baier 1923 and Terazaki, 2005). 1924 1925 Neocalanus spp. overwinter off the shelf at depths of 200-2000 m (e.g. Miller et al., 1984). The 1926 newly hatched nauplii and copepodites ascend to the upper layer in late winter to early spring, 1927 where they complete their development by early summer (Vidal and Smith, 1986), and start to 1928 descend to deeper layers. Recent modeling suggests that wind is a primary factor controlling 78 1929 inter-annual variability in on-shelf transport of Neocalanus spp. (Gibson et al, 2013). 1930 Southeasterly winds, which prevailed during warm years, enhanced the on-shelf transport of 1931 oceanic zooplankton over the southern shelf, while northwesterly winds, more common in cold 1932 years, reduced onshore transport. This suggests that Neocalanus spp. abundance would be lower 1933 during cold years since it would be more difficult for this taxa to move onto the southern shelf in 1934 spring (Danielson et al 2012, Gibson et al. 2013). However, Neocalanus spp. abundance 1935 increased in cold years over the S Middle Domain in our data set and there was no significant 1936 difference in its abundance between warm and cold years over the Outer shelf. Thus, the 1937 explanation for variations in Neocalanus spp. abundance over the shelf is likely more complicated 1938 with multiple factors important (e.g. transport, food availability, predation). 1939 1940 Aglantha digitale, a small (10-40 mm long) hydromedusan jellyfish, was the most numerous 1941 Cnidarian collected in our bongo nets from 2005-2009, making up 93% (ranged 87 – 97% by 1942 year) of the total Cnidarian abundance. A digitale is an ambush predator capable of retaining 1943 small, motile prey (Costello and Colin, 2002). This taxon has been shown to ingest adults and 1944 larvae of small copepods (Oithona spp., Acartia spp., Pseudocalanus spp., Temora longicornis), 1945 Evadne sp., Oikopleura spp., zooplankton eggs, veligers, tintinnids and dinoflagellates (Matsakis 1946 and Conover, 1991; Pages et al., 1996; Costello and Colin, 2002) with an estimated maximum 1947 prey size of 1.5 mm (Prudkovsky, 2006). Matsakis and Conover (1991) found that medusan 1948 carbon weight and total zooplankton carbon biomass varied reciprocally in studies in Bedford 1949 Basin, Nova Scotia. For the current study, in the northeastern Bering Sea with the exception of 1950 2009, small copepod abundances tended to be higher when Cnidaria (A. digitale primarily) 1951 abundances were lower and vice versa (Fig. 7). Thus, predation by A. digitale on small copepods 1952 may be one factor to help explain these interannual changes in abundances. Higher numbers of 1953 Cnidarians in the northeastern Bering Sea may also partially account for lower numbers of small 1954 taxa in the north compared to the south. Average abundances of A. digitale and total copepods, 79 1955 respectively, were 42.5 m-3 and 3800 m-3 in the north and 4.3 m-3 and 6600 m-3 in the south, for 1956 2005 – 2009 for the Inner and Middle Domains combined. If A. digitale consumes an average of 1957 13.9 copepod prey per day (based on feeding experiments at 4°C , Matsakis and Conover, 1991), 1958 then for the north and south respectively, they can consume 15% and 1% of the total copepod 1959 standing stock per day. In other studies, gelatinous zooplankton often consume < 10% of prey 1960 resources, but can consume over 50% during episodic events (Matsakis and Conover, 1991 and 1961 references there in; Pages et al., 1996). We also found that A. digitale abundances were 63% and 1962 72% higher in a warm year (2005) than in cold years (2006 – 2009) for the north and southeastern 1963 Bering Sea, respectively. 1964 1965 5.0 Conclusions 1966 Changes in eastern Bering Sea zooplankton community composition in response to climate 1967 change will vary by oceanographic domain and by latitude, with larger effects predicted for the 1968 south compared to the northeastern Bering Sea. This impact will be stronger on large, than on 1969 small zooplankton community structure. Reductions in sea ice coverage and changes in ice retreat 1970 timing and subsequent decreases in food resources in the spring, increases in temperature and 1971 basal metabolic requirements, and potential changes in advection may be detrimental for large 1972 crustacean taxa such as Calanus spp. and Neocalanus spp. The negative impact of warming on 1973 large, lipid-rich zooplankton taxa is expected to propagate through the food web affecting 1974 commercial fisheries, seabirds, and protected species. In the eastern Bering Sea, statistical 1975 relationships between water temperature and recruitment of pollock have been used in 1976 combination with accepted climate change scenarios to project a future decrease in one of the 1977 world’s largest fisheries (Ianelli et al., 2011 and Mueter et al., 2011). Our work helps to elucidate 1978 the mechanisms behind that statistical relationship. 1979 80 1980 Alterations in the composition of small zooplankton assemblages from climate change may also 1981 contribute to anticipated effects at higher trophic levels. In the eastern Bering Sea, Oithona spp., 1982 dominant during the warm regime, decreased in abundance in the cold regime, while 1983 Pseudocalanus and Acartia spp. increased. This species replacement had no net effect on the total 1984 biomass, but we speculate that it did affect trophic pathways, as Pseudocalanus and Acartia are 1985 considered more favorable as food for other planktivores (e.g. Oithona spp. nauplii are under- 1986 represented in larval pollock stomachs). Accordingly under future warming scenarios, global food 1987 webs, which include humans, (sensu Rice et al., 2011) may be impacted to a higher degree in 1988 regions that become ice-free compared to those which retain seasonal ice coverage. 1989 1990 Acknowledgements: 1991 We thank the captain and crews of the NOAA ship Oscar Dyson, and charter vessels, Sea Storm, 1992 NW Explorer, and Epic Explorer, for their years of hard work and diligence on our surveys. We 1993 are grateful for the assistance in field sampling, data processing and analysis from NOAA 1994 scientific staff and volunteers. In particular we’d like to thank K. Cieciel, J. Gann and F. Van 1995 Tulder for sampling, processing and database maintenance of oceanographic data, M. Auburn- 1996 Cook, M. Courtney, A. Feldman, E. Farley, J. Murphy and B. Wing for the many years of survey 1997 participation and assistance with zooplankton collection and processing. We are deeply 1998 appreciative of the TINRO scientists, in particular N. Kusnetsova and A.Volkov, who conducted 1999 the onboard taxonomic analysis of Juday samples. We thank K. Coyle and his lab for assistance 2000 and guidance regarding zooplankton sampling and analysis, as well as the Polish Plankton Sorting 2001 and Identification Center for their contributions to the early part of this time series. K. Coyle also 2002 provided insightful comments on an earlier version of this manuscript. We greatly appreciate the 2003 helpful comments from the three anonymous reviewers and the journal editor, C. Ashjion. 2004 Funding was provided by the North Pacific Research Board, Bering Sea Fisherman’s Association, 2005 Arctic-Yukon-Kuskokwim-Sustainable-Salmon-Initiative, and NOAA National Marine Fisheries 81 2006 Service including the Fisheries and the Environment (FATE) and the North Pacific Climate 2007 Regimes and Ecosystem Productivity (NPCREP) programs. This is contribution EcoFOCI-0807 2008 to NOAA’s Ecosystems Fisheries Oceanography Coordinated Investigations, contribution ##### 2009 from BEST/BSIERP. 2010 References: 2011 Anderson, M.J., Gorley, R.N., Clarke, K.R., 2008. PERMANOVA+ for PRIMER: guide to 2012 software and statistical methods. PRIMER-E, Plymouth. 2013 2014 2015 Baier, C.T. Napp, J.M., 2003. Climate-induced variability in Calanus marshallae populations. J. Plankton Res. 25, 771-782. 2016 2017 Baier, C. T.,Terazaki, M., 2005. Interannual variability in a predator-prey interaction: climate, 2018 chaetognaths and copepods in the southeastern Bering Sea. J. Plankton Res. 27(11), 2019 1113-1125. 2020 2021 2022 Bray J.R., Curtis J.T., 1957. An ordination of the upland forest communities of Southern Wisconsin. Ecological Monographies 27:325-349 2023 2024 Cieciel, K., Farley, E.V. Jr, Eisner, L.B., 2009. Jellyfish and juvenile salmon associations with 2025 oceanographic characteristics during warm and cool years in the eastern Bering Sea. 2026 North Pacific Anadromous Fish Commission Bulletin 5, 209–224. 2027 2028 Clarke, K.R., Gorley, R.N., 2006. PRIMER v6: user manual/tutorial. PRIMER-E, Plymouth. 2029 2030 2031 Clarke, K.R., Warwick, R.M., 2001. Change in marine communities: an approach to statistical analysis and interpretation, 2nd edn. PRIMER-E, Plymouth. 2032 2033 2034 Coachman, L.K., 1986. Circulation, water masses and fluxes on the southeastern Bering Sea shelf. Cont. Shelf Res. 5, 23–108. 2035 2036 2037 Cooney, R.T., Coyle, K.O., 1982. Trophic implications of cross-shelf copepod distributions in the southeastern Bering Sea. Mar. Biol. 70(2), 187-196. 82 2038 2039 2040 Costello, J.H., Colin, S.P., 2002. Prey resource use by coexistent hydromedusae from Friday Harbor, Washington. Limnol. Oceanogr. 47(4), 934-942. 2041 2042 Coyle, K.O., Chavtur, V.G., Pinchuk, A.I., 1996. Zooplankton of the Bering Sea: A review of 2043 Russian-language literature, pp. 97-133. In: Mathisen, O. and Coyle, K.O. (Eds.), 2044 Ecology of the Bering Sea: A review of Russian Literature, Alaska Sea Grant Program 2045 Report No. 96-01, University of Alaska, Fairbanks. 2046 2047 Coyle, K.O., Eisner, L.B., Mueter, F.J., Pinchuk, A.I., Janout, M.A., Cieciel, K.D., Farley, E.V., 2048 Andrews, A.G., 2011. Climate change in the southeastern Bering Sea: impacts on pollock 2049 stocks and implications for the oscillating control hypothesis. Fish. Oceanogr. 20, 139– 2050 156. 2051 2052 Coyle, K.O., Pinchuk, A.I., 2002. Climate-related differences in zooplankton density and growth 2053 on the inner shelf of the southeastern Bering Sea. Prog. Oceanogr. 55, 177–194. 2054 2055 Coyle, K.O., Pinchuk, A.I., Eisner, L.B., Napp, J.M., 2008. Zooplankton species composition, 2056 abundance and biomass on the eastern Bering Sea shelf during summer: the potential role 2057 of water column stability and nutrients in structuring the zooplankton community. Deep 2058 Sea Res. II 55, 1775–1791. 2059 2060 Danielson, S., Eisner, L., Weingartner, T. and Aagaard, K., 2011. Thermal and haline variability 2061 over the central Bering Sea shelf: Seasonal and interannual perspectives. Cont. Shelf Res. 2062 31, 539-554. 2063 2064 Danielson, S., Hedstrom, K., Aagaard, K., Weingartner, T., Curchitser, E., 2012. Wind-induced 2065 reorganization of the Bering shelf circulation. Geophys. Res. Lett. 39, L08601, 2066 doi:10.1029/2012GL051231 2067 2068 2069 ESRI, 2012. ArcGIS Desktop: Release 10.1. Redlands, CA: Environmental Systems Research Institute 2070 83 2071 Farley, E.V. ,Moss, J.H., 2009. Growth rate potential of juvenile chum salmon on the eastern 2072 Bering Sea shelf: an assessment of salmon carrying capacity. North Pacific Anadromous 2073 Fish Commission Bulletin 5, 265–277. 2074 2075 Friday, N.A., Waite, J.M., Moore, S.E., Zerbini, A.N., 2012. Cetacean distribution and abundance 2076 in relation to oceanographic domains on the eastern Bering Sea shelf: 1999–2004. Deep 2077 Sea Res. II 65–70, 260–272. 2078 2079 2080 Frost, B.W., 1974. Calanus marshallae, a new species of Calanoid copepod closely allied to the sibling species C. finmarchicus and C. glacialis. Marine Biology 26, 77-99. 2081 2082 Gibson, G. A., Coyle, K.O., Hedstrom, K, Curchitser, E. N. 2013. A modeling study to explore 2083 on-shelf transport of oceanic zooplankton in the Eastern Bering Sea. Journal of Marine 2084 Systems. 121, 47-64 2085 2086 2087 Gieskes, W.W.C., 1971. Ecology of the cladocera of the North Atlantic and the North Sea. Netherlands Journal of Sea Research. 5(3), 342-376. 2088 2089 2090 Gold, C.M., 1991. Problems with handling spatial data--the Voronoi approach. CISM Journal 45, 65-80 2091 2092 Gordon, L.I., Jennings, J.C. Jr., Ross, A.A., Krest, J.M., 1993. A suggested protocol for 2093 continuous flow automated analysis of seawater nutrients (phosphate, nitrate, nitrite and 2094 silicic acid) in the WOCE Hydrographic Program and the Joint Global Ocean Fluxes 2095 Study. WOCE Hydrographic Program Office, Methods Manual WHPO 91-1, Oregon 2096 State University, Corvallis. 2097 2098 Grebmeier, J.M., Overland, J.E., Moore, S.E., Farley, E.V., Carmack, E.C., Cooper, L.W., Frey, 2099 K.E., Helle, J.H., McLaughlin, F.A., McNutt, S.L., 2006. A major ecosystem shift in the 2100 northern Bering Sea. 311(5766), 1461-4. 2101 2102 Heintz, R.A., Siddon, E.C., Farley, E.V. Jr., Napp, J.M., 2013. Climate-related changes in the 2103 nutritional condition of young-of-the-year walleye pollock (Theragra chalcogramma) 2104 from the eastern Bering Sea. Deep Sea Res. II BEST-BSIERP Special Issue 2. 84 2105 2106 2107 Hirst, A.G., Lampitt, R.S., 1998. Towards a global model of in situ weight specific growth in marine planktonic copepods. Mar. Biol. 132, 247-257. 2108 2109 Hollowed, A.B., Barbeaux, S.J., Cokelet, E.D., Farley, E., Kotwicki, S., Ressler, P.H., Spital, C., 2110 Wilson, C.D., 2012. Effects of climate variations on pelagic ocean habitats and their role 2111 in structuring forage fish distributions in the Bering Sea. Deep Sea Res. II, 65-70, 230- 2112 250. 2113 2114 Hunt, G.L. Jr., Coyle, K.O., Eisner, L.B., Farley, E.V., Heintz, R., Mueter, F., Napp, J.M., 2115 Overland, J.E., Ressler,P.H., Salo, S., Stabeno, P.J., 2011. Climate impacts on eastern 2116 Bering Sea food webs: A synthesis of new data and an assessment of the Oscillating 2117 Control Hypothesis. ICES J. Mar. Sci. 68, 1230-1243. 2118 2119 Hunt, G.L., Jr., Stabeno, P.J., Walters, G., Sinclair, E., Brodeur, R.D., Napp, J.M., Bond, N.A., 2120 2002. Climate change and control of the southeastern Bering Sea pelagic ecosystem. 2121 Deep Sea Res. II 49, 5821-5853. 2122 2123 Hunt, G.L., Stabeno, P.J., Strom, S., and Napp, J.M., 2008. Patterns of spatial and temporal 2124 variation in the marine ecosystem of the southeastern Bering Sea, with special reference 2125 to the Pribilof Domain. Deep Sea Res. II, 55, 1919–1944. 2126 2127 Ianelli, J.N, Hollowed, A.B., Haynie A.C., Mueter, F.J., Bond, N.A., 2011. Evaluating 2128 management strategies for eastern Bering Sea walleye pollock (Theragra chalcogramma) 2129 in a changing environment. ICES J. Mar. Sci. 68(6), 1297-1304. 2130 2131 Iverson, R.L., Coachman, L.K., Cooney, R.T., English, T.S., Goering, J.J., Hunt, Jr., G.L., 2132 McCauley, M.C., McRoy, C.P., Reeburg, W.S., Whitledge, T.E., 1979. Ecological 2133 significance of fronts in the southeastern Bering Sea, pp. 437-466. In: Ecological 2134 Processes in Coastal and Marine Systems, R.J. Livingston (Ed.), Plenum Press. 2135 2136 2137 Johnson, M., 1934. The life history of the copepod Tortanus discaudatus (Thomspon and Scott). The Biological Bulletin, Woods Hole 67, 182-200. 2138 85 2139 Kachel, N.B., Hunt Jr., G.L., Salo, S.A., Schumacher, J.D., Stabeno, P.J., Whitledge, T.E., 2002. 2140 Characteristics and variability of the inner front of the southeastern Bering Sea. Deep Sea 2141 Res. II 49 (26), 5889–5909. 2142 2143 Kos, M.S., 1977. Species of the genus Eurytemora (Copepoda, Calanoida) in the northern part of 2144 The Pacific Ocean: systematics, distribution, variability. Explorations of the Fauna of the 2145 Seas 20, 20-53. 2146 2147 Kotwicki, S. Lauth,R.R., 2013. Detecting temporal trends and environmentally-driven changes in 2148 the spatial distribution of groundfishes and crabs on the eastern Bering Sea shelf. Deep 2149 Sea Res. II. 2150 2151 Lomas, M.W., Moran, S.B., Casey, J.R., Bell, D.W., Tiahlo, M., Whitefield, J., Kelly, R.P., 2152 Mathis, J.T., Cokelet, E.D., 2012. Spatial and seasonal variability of primary production 2153 on the Eastern Bering Sea shelf. Deep Sea Res II 65-70, 126-140. 2154 2155 2156 Marlowe, C.J., Miller, C.B., 1975. Patterns of vertical distribution and migration of zooplankton at ocean Station “P”. Limnol. Oceanogr. 20(5), 824-844. 2157 2158 Matsakis, S., Conover, R. J., 1991. Abundance and feeding of medusae and their potential impact 2159 as predators on other zooplankton in Bedford Basin (Nova Scotia, Canada) during spring. 2160 Can. J. Fish. Aquat. Sci. 48(8), 1419-1430. 2161 2162 Miller, C. B., Frost, B. W., Batchelder, H. P., Clemons, M. J., Conway, R. E., 1984. Life histories 2163 of large, grazing copepods in a subarctic ocean gyre: Neocalanus plumchrus, Neocalanus 2164 cristatus and Eucalanus bungii in the northeast Pacific. Prog. Oceanogr. 13, 201-243. 2165 2166 Motoda, S., Minoda, T., 1974. Plankton of the Bering Sea, pp. 207-241. In: D.W. Hood and E.J. 2167 Kelley, (Eds), Oceanography of the Bering Sea with Emphasis on Renewable Resources. 2168 University of Alaska Press. 2169 2170 Mueter, F. J., Bond, N.A., Ianelli, J.N., Hollowed, A.B., 2011. Expected declines in recruitment 2171 of walleye pollock (Theragra chalcogramma) in the eastern Bering Sea under future 2172 climate change. ICES J. Mar. Sci. 68(6), 1284-1296. 86 2173 2174 Napp, J.M., Baier, C.T., Brodeur, R.D., Coyle, K.O., Shiga, N., Mier, K., 2002. Interannual and 2175 decadal variability in zooplankton communities of the south- east Bering Sea shelf. Deep 2176 Sea Res. II 49, 5991–6008. 2177 2178 2179 Napp, J.M., Hunt Jr., G.L., 2001. Anomalous conditions in the southeastern Bering Sea, 1997: linkages among climate, weather, ocean, and biology. Fish. Oceanogr. 10, 61–68. 2180 2181 Ortiz, I,. Weise, F., Greig, A., 2012. Marine Regions Boundary Data for the Bering Sea Shelf and 2182 Slope. UCAR/NCAR - Earth Observing Laboratory/Computing, Data, and Software 2183 Facility. Dataset. doi:10.5065/D6DF6P6C 2184 2185 Pages, F., Gonzalez, H.E., Gonzalez, S.R., 1996. Diet of the gelatinous zooplankton in 2186 Hardangerfjord (Norway) and potential predatory impact by Aglantha digitale 2187 (trachymedusae). Mar. Ecol. Prog. Ser. 139(1-3), 69-77. 2188 2189 2190 Parsons, T.R., Maita, Y., Lalli, C.M., 1984. A Manual of Chemical and Biological Methods for Seawater Analysis. Pergamon Press, Oxford, England, 173 pp. 2191 2192 2193 Piatt, J.F., Springer, A.M., 2003. Advection, pelagic food webs and the biogeography of seabirds in Beringia. Mar. Ornithol. 31, 141–154. 2194 2195 Pinchuk, A.I., Coyle, K.O., Farley, E.V., Renner, H.M., 2013. Emergence of the Arctic Themisto 2196 libellula (Amphipoda: Hyperiidae) on the southeastern Bering Sea shelf as a result of the 2197 recent cooling and their potential impact on pelagic food web. ICES J. Mar. Sci. 2198 DOI###### 2199 2200 2201 Prudkovsky, A.A., 2006. Food selectivity in Aglantha digitale (Hydrozoa, Trachymedusae) in the White Sea. Zoologicheskii Zhurnal 85(11), 1294-1302. 2202 2203 Rice, J.C., Garcia, S.M., 2011. Fisheries, food security, climate change, and biodiversity: 2204 characteristics of the sector and perspectives on emerging issues. ICES J. Mar. Sci. 68, 2205 1343-1353. 2206 87 2207 Russell, R.W., Harrison, N.M., Hunt, G. J., 1999. Foraging at a front: Hydrography, zooplankton, 2208 and avian planktivory in the northern Bering Sea. Mar. Ecol. Prog. Ser.182, 77-93. 2209 2210 Sameoto, D., Cochrane, N. and Herman, A., 1993. Convergence of acoustic, optical, and net- 2211 catch estimates of euphausiid abundance: Use of artificial light to reduce net avoidance. 2212 Can. J. Fish. Aquat. Sci. 50, 334-346. 2213 2214 Sheffield-Guy, L., Roby, D.D., Gall, A.E., Irons, D. B., Rose, I. C., 2009. The influence of diet 2215 and ocean conditions on productivity of auklets on St Lawrence Island, Alaska. Mar. 2216 Ornithol. 37(3), 227-236. 2217 2218 Sigler, M.F. Stabeno, P., In review, this issue. Spring and fall phytoplankton blooms in a 2219 productive subarctic ecosystem, the eastern Bering Sea, during 1995-2011. Deep-Sea 2220 Res. II 2221 2222 2223 Simpson, J.H., Hughes, D.G., Morris, N.C.G., 1977. The relation of seasonal stratification to tidal mixing on the continental shelf. Deep Sea Res. 24(Suppl.), 327–340. 2224 2225 2226 Springer, A.M., Byrd, G.V., Iverson, S.J., 2008. Hot oceanography: Planktivorous seabirds reveal ecosystem responses to warming of the Bering Sea. Mar. Ecol. Prog. Ser. 352, 289-297. 2227 2228 Stabeno, P.J., Farley, E.V., Kachel, N., Moore, S., Mordy, C., Napp, J.M., Overland, J.E., 2229 Pinchuk, A.I., Sigler, M., 2012a. A comparison of the physics, of the northern and 2230 southern shelves of the eastern Bering Sea and some implications to the ecosystem. Deep 2231 Sea Res. II 65-70:14-30. 2232 2233 Stabeno, P.J., Kachel, N.B., Moore, S.E., Napp, J.M., Sigler, M., Yamaguchi, A., Zerbini, A.N., 2234 2012b. Comparison of warm and cold years on the southeastern Bering Sea shelf and 2235 some implications for the ecosystem. Deep Sea Res. II 65-70:31-45. 2236 2237 Stabeno, P. J., Kachel, N., Mordy, C., Righi, D., Dalo, S., 2008. An examination of the physical 2238 variability around the Pribilof Islands in 2004. Deep Sea Res. II 55(16-17), 1701-1716. 2239 88 2240 Stabeno, P.J., Napp, J., Mordy, C., Whitledge, T., 2010. Factors influencing physical structure 2241 and lower trophic levels of the eastern Bering Sea shelf in 2005: sea ice, tides and winds. 2242 Prog. Oceanogr. 85 (3-4), 180–196. 2243 2244 Stevenson, D.E., Lauth,R.R., 2012. Latitudinal trends and temporal shifts in the catch 2245 composition of bottom trawls conducted on the eastern Bering Sea shelf. Deep Sea Res. 2246 II, 65-70, 251-259. 2247 2248 2249 Thiessen, A. J., Alter, J.C., 1911. Precipitation Averages for Large Areas. Monthly Weather Rev. 39, 1082-84. 2250 2251 Vidal, J., Smith, S.L., 1986. Biomass, growth, and development of populations of herbivorous 2252 zooplankton in the southeastern Bering Sea during spring. Deep Sea Res. 33, 523-556. 2253 2254 2255 Volkov, A.F., 1984. Recommendations for Expedited Zooplankton Sample Processing at Sea. Vladivostok: TINRO, 31 p (in Russian). 2256 2257 Volkov, A.F., Efimkin, A.Y. Kuznetseva, N.A., 2007. Characteristics of the plankton community 2258 of the Bering Sea and several regions of the North Pacific in 2002 – 2006. Izvestiya 2259 TINRO 151, 338–364 (in Russian). 2260 2261 Wade, P. R., De Robertis, A., Hough, K.,. Booth, R , Kennedy, A., LeDuc, R., Munger, L., Napp, 2262 J., Shelden, K.E.W., Rankin, S., Vasquez, O., Wilson., C., 2011. Rare detections of North 2263 Pacific right whales in the Gulf of Alaska, with observations of their potential prey. 2264 Endang. Species Res. 13:99-109. 2265 2266 Wiebe, P.H., Ashjian, C.J., Gallager, S.M., Davis, C.S., Lawson, G.L., and Copley, N.J., 2004. 2267 Using a high-powered strobe light to increase the catch of Antarctic krill. Mar. Biol. 144, 2268 493-502. 2269 2270 2271 Wiese, F.K., Wiseman, W.J., van Pelt, T.I., 2012. Bering Sea linkages. Deep Sea Res. II 65-70, 25. 2272 89 2273 2274 Wyllie-Echevarria, T., Wooster, W.S., 1998. Year-to-year variations in Bering Sea ice cover and some consequences for fish distributions. Fish. Oceanogr. 7, 159–170. 2275 90 Table 1. Questions addressed in manuscript. Q1 What are the spatial and temporal patterns of large and small zooplankton taxa in the eastern Bering Sea for 2003–2009? Q2 How do the total abundances and biomasses of large and small zooplankton vary between warm and cold years for the southeastern and northeastern Bering Sea? Q3a Are the community compositions of large and small taxa different between warm and cold regimes within each domain? b When these communities differed between warm and cold regimes which taxa contributed most to the differences? c Did the observed differences in zooplankton community composition between regimes occur exactly during the break between low and high ice years? d Which taxa showed the largest variations among years? Q4 For each domain, which environmental factors explained the greatest amount of variability in zooplankton community composition overall and between warm and cold regimes? 91 Table 2. PERMANOVA results (P-values from permutation test) for differences in community structure of large and small zooplankton taxa (abundance data) in the warm (2003 – 2005) versus cold (2006 – 2009) years by domain (Fig. 1). Other effects in model are not shown (Block, Year (Regime), and interactions). *Significant at P < 0.05. Domain S Inner S Middle S Outer N Inner N Middle Large P(perm) 0.002* 0.001* 0.008* 0.152 0.024* Small P(perm) 0.005* 0.001* 0.731 0.438 0.029* 92 93 Table 4. PERMANOVA results (P-values) showing differences in community composition for large and small zooplankton taxa (abundance data) by domain. P-values for at least one difference among years (All years) are followed by pairwise multiple comparisons for consecutive years. * P < 0.05. Large zooplankton: S Inner S Middle S Outer N Inner N Middle All years < 0.001* < 0.001* 0.015* < 0.001* < 0.001* 2003 2004 0.005* 0.007* 0.382 0.290 0.157 2004 2005 < 0.001* < 0.001* 0.084 0.001* 0.014* 2005 2006 < 0.001* < 0.001* 0.364 0.026* 0.007* 2006 2007 0.001* < 0.001* 0.764 0.023* 0.002* 2007 2008 0.108 < 0.001* 2008 2009 0.043* zooplankton: S Inner S Middle S Outer N Inner N Middle All years < 0.001* < 0.001* 0.011* < 0.001* < 0.001* 0.043* Small 2003 2004 0.003* 0.296 0.743 0.028* 0.154 2004 2005 0.001* 0.045* 0.044* 0.026* 0.003* 2005 2006 0.160 0.031* 0.149 0.715 0.020* 2006 2007 0.438 0.084 0.243 0.734 0.133 2007 2008 0.038* 0.320 0.770 2008 2009 0.087 0.057 0.204 94 Table 5. Results of best-fit PERMANOVA models using Type I (sequential) sum of squares showing P-values of environmental covariates for large and small zooplankton community composition. Order of inclusion in the model is shown in parentheses. An insignificant (P > 0.05) Regime (factor) indicates covariates explained some of the variability between regimes. *Significant at P < 0.05. Large zooplankton Variable longitude latitude Integrated Chla Winter wind direction S below MLD T below MLD T above MLD Ice retreat timing Regime (factor) Small Zooplankton Variable longitude latitude T below MLD T above MLD Ice retreat timing stability Regime (factor) S Inner 0.012 (1)* 0.014 (2)* S Middle - S Outer 0.006 (1)* - N Middle 0.006 (1)* - 0.030 (3)* 0.079 0.001 (1)* 0.001 (2)* 0.001 (3)* 0.001* 0.006 (2)* 0.383 0.001 (2)* 0.082 (3)* 0.335 S Inner 0.001 (1)* 0.001 (2)* 0.393 S Middle 0.025 (4)* 0.003 (3)* 0.001 (2)* 0.001 (1)* 0.061 N Middle 0.020 (1)* 0.042 (2)* 0.202 (3) 0.252 95 2276 Figure Captions: 2277 Figure 1. Bering Sea study area showing stations used for analyses in each oceanographic shelf 2278 domain for: A) large zooplankton and B) small zooplankton. Black lines indicate spatial “blocks” 2279 which are thiessen polygons around points. Blocks containing colored points (reduced data set) 2280 were visited at least two years during warm (2003 – 2005) and two years during cold (2006 – 2281 2009) regimes; colors signify domain. 2282 2283 Figure 2. Heat map of large zooplankton showing clusters of stations (x axis) by taxa (y axis) 2284 using all stations sampled. Shade of each cell indicates the 4th root transformed abundance of each 2285 taxon. Approximate domain designations are shown above clusters. Designations at bottom 2286 indicate climate regime (W = warm; C = cold; W/C = both) when each cluster was commonly 2287 observed. Horizontal lines separate clusters of species (indicating taxa which tended to co-occur 2288 over all years and domains combined). 2289 2290 Figure 3. Spatial variation of large zooplankton clusters for each year using 4th root transformed 2291 abundance data. Warm years highlighted with red and cold with blue. Refer to Fig. 2 to see taxa 2292 abundances for each cluster. Intended for color reproduction on the Web and in print 2293 2294 Figure 4. Heat map of small zooplankton clusters of stations (x axis) by taxa (y axis). Refer to 2295 Fig. 2 for details. 2296 2297 Figure 5. Spatial variation of small zooplankton clusters by year using 4th root transformed 2298 abundance data. Warm years highlighted with red and cold with blue. Refer to Fig. 4 to see taxa 2299 abundances for each cluster. Intended for color reproduction on the Web and in print. 2300 96 2301 Figure 6. Small and large zooplankton total mean abundance (no. m-3) in the eastern Bering Sea 2302 in the A) south (< ~60N) and B) north (~60-63 N), and total mean biomass (g m-3) in the C) 2303 south and D) north. Geometric means and standard errors are shown. 2304 2305 Figure 7. Untransformed mean zooplankton abundances in the eastern Bering Sea for large taxa 2306 in the A) south (< ~60 N) and B) north (~60-63 N) and for small taxa in the C) south and D) 2307 north. Intended for color reproduction on the Web and in print. 2308 2309 Figure 8. A) Mean temperature above (surface) and below (deep) the mixed layer depth during 2310 our surveys, B) mean day of year for sea ice retreat during the prior spring, and C) mean number 2311 of days of sea ice coverage during the prior winter, in the south (S) and north (N) Middle 2312 Domains. Standard errors are shown. 97 2313 Chapter 14 2314 2315 Sigler, M.F., P.J. Stabeno, L.B. Eisner, and J.M. Napp (2012): Spring and fall phytoplankton 2316 blooms in a productive subarctic ecosystem, the eastern Bering Sea, during 1995-2011. Deep-Sea 2317 Res. II. 2318 2319 Spring and fall phytoplankton blooms in a productive subarctic ecosystem, the eastern Bering 2320 Sea, during 1995-2011 2321 Michael F. Sigler1, Phyllis J. Stabeno2, Lisa B. Eisner1, Jeffrey M. Napp3, Franz J. Mueter4 2322 2323 1 2324 Atmospheric Administration, 17109 Pt. Lena Loop Rd., Juneau, AK 99801 USA 2325 (907) 789-6037; mike.sigler@noaa.gov and (907) 789-6602; lisa.eisner@noaa.gov Alaska Fisheries Science Center, National Marine Fisheries Service, National Oceanic and 2326 2327 2 2328 and Atmospheric Administration, 7600 Sand Point Way NE, Seattle, WA 98115-0070 USA 2329 (206) 526-6453; phyllis.stabeno@noaa.gov Pacific Marine Environmental Laboratory, Oceans and Atmospheric Research, National Oceanic 2330 2331 3 2332 Atmospheric Administration, 7600 Sand Point Way NE, Seattle, WA 98115-0070 USA 2333 Phone: (206) 526-4148; jeff.napp@noaa.gov Alaska Fisheries Science Center, National Marine Fisheries Service, National Oceanic and 2334 2335 3 2336 Rd., Juneau, AK 99801 USA 2337 Phone: (907) 790-5448; fmueter@alaska.edu University of Alaska Fairbanks, Juneau Center for Fisheries and Oceans, 17309 Pt. Lena Loop 2338 2339 2340 Abstract The timing and magnitude of phytoplankton blooms in subarctic ecosystems often 2341 strongly influence the amount of energy that is transferred through each trophic pathway. In the 2342 eastern Bering Sea, spring bloom timing has been linked to ice retreat timing and production of 2343 zooplankton and fish. A large part of the eastern Bering Sea shelf (~500 km wide) is ice-covered 2344 during winter and spring. Four oceanographic moorings have been deployed along the 70-m 2345 depth contour of the eastern Bering Sea shelf with the southern location occupied annually since 2346 1995, the two northern locations since 2004 and the remaining location since 2001. Chlorophyll 98 2347 a fluorescence data from the four moorings provide 37 realizations of a spring bloom and 33 2348 realizations of a fall bloom. We found that in the eastern Bering Sea: if ice was present after 2349 mid-March, spring bloom timing was related to ice retreat timing (p < 0.001, df = 1,24); if ice 2350 was absent or retreats before mid-March, a spring bloom usually occurred in May or early June 2351 (average day 148, SE = 3.5, n = 11). A fall bloom also commonly occurred, usually in late 2352 September (average day 274, SE = 4.2, n = 33), and its timing was not significantly related to 2353 the timing of storms (p = 0.88, df = 1,27) or fall water column overturn (p = 0.49, df = 1,27). 2354 The magnitudes of the spring and fall blooms were correlated (p = 0.011, df = 28). The interval 2355 between the spring and fall blooms varied between four to six months depending on year and 2356 location. We present a hypothesis to explain how the large crustacean zooplankton taxa Calanus 2357 spp. likely responds to variation in the interval between blooms (spring to fall and fall to spring). 2358 2359 1. Introduction 2360 The timing and magnitude of phytoplankton blooms in subarctic ecosystems often 2361 strongly influence the amount of energy transferred through each trophic pathway. In the 2362 southeastern Bering Sea, spring bloom timing has been linked to production of large crustacean 2363 zooplankton and walleye pollock (Theragra chalcogramma) (Hunt et al., 2002, 2011; Coyle et 2364 al., 2011); if ice is present after mid-March, an early ice-associated bloom occurs there; otherwise 2365 a spring bloom usually occurs in May (Hunt et al., 2002; Stabeno et al., 2001). Although spring 2366 bloom timing is well-characterized in the southeastern part of the shelf (Brown and Arrigo, 2011, 2367 2013; Hunt et al., 2002, 2011; Rho and Whitledge 2007; Stabeno et al., 2001), less is known 2368 about the spring bloom elsewhere in the eastern Bering Sea (Brown and Arrigo 2013), as well as 2369 the characteristics of the fall bloom (Rho and Whitledge 2007). 2370 The eastern Bering Sea is dominated by a broad continental shelf (~500 km wide), a large 2371 part of which is ice-covered during winter, with the maximum extent varying >100 km among 2372 years. In ice-covered areas, the seasonal cycle of primary production begins with ice algae 2373 (primarily large diatoms), which begin to grow in the spring when light level becomes adequate. 2374 Ice algae are adapted to lower light levels than pelagic phytoplankton (Kirst and Wiencke, 1995) 2375 and grow within the ice and at the ice-water interface depending on the amount of overlying snow 2376 cover. Ice algae begin to grow in mid-February in the Bering Sea (R. Gradinger, University of 2377 Alaska, Fairbanks, pers. comm.) and may provide an early concentrated food source (chlorophyll 2378 a maximum ~300 µg l-1; Mock and Gradinger, 2000; Niemi et al., 2011) for zooplankton (e.g. 2379 O’Brien, 1987; Runge and Ingram, 1991). Sea ice algae that are not grazed can seed the spring 2380 phytoplankton bloom or may aggregate and sink out of the upper water column (Tremblay et al., 99 2381 1989, Riebesell et al., 1991; Haecky et al., 1998; Fortier et al., 2002) with seeding more likely if 2382 the release from ice is not followed by a strong mixing event that breaks down stratification (Jin 2383 et al., 2007). Sedimentation of ice algae occurs during and immediately after the bloom starts 2384 over the Bering Sea middle shelf (R. Gradinger, University of Alaska, Fairbanks, pers. comm.). 2385 Ice algae increases within the ice typically are terminated during ice melt in late spring or early 2386 summer. 2387 Phytoplankton in the Bering Sea begin to bloom in the spring once the water becomes 2388 stratified and the day length becomes long enough (sensu Sverdrup 1953). Prior to this, 2389 phytoplankton are considered to be light-limited, but have adequate nutrients due to the advection 2390 of nutrient rich slope water onto the shelf during the previous winter, which is mixed throughout 2391 the water column (Niebauer et al., 1995); nutrient recycling on the shelf also is important 2392 (Granger et al., 2011). The phytoplankton spring bloom typically ends when the surface nutrient 2393 supply is exhausted and phytoplankton growth becomes nutrient limited (typically below 1 µM 2394 nitrate). Grazing pressure from mesozooplankton and microzooplankton also increases as the 2395 spring progresses, which lowers phytoplankton standing stocks. 2396 In the summer, phytoplankton concentration in the surface mixed layer is typically low 2397 due to nutrient limitation and continued grazing pressure. Episodic wind events can break down 2398 stratification and mix nutrients and viable phytoplankton cells to the surface during this period 2399 (Sambrotto 1986; Stabeno et al., 2010). During fall, increased storminess and overall cooling of 2400 the water column reduces stratification and deepens the mixed layer so that nutrients are mixed to 2401 the surface to fuel fall phytoplankton blooms. The fall bloom ends when phytoplankton become 2402 light limited, due to decreased day length and deepening of the mixed layer, and when 2403 zooplankton grazing reduces standing stocks. Pelagic-benthic coupling (Grebmeier et al., 2006) 2404 and luxury consumption by diatoms of nutrients near the sediment may also be important (Droop 2405 1973), since the Bering Sea shelf is a shallow shelf system with 1% light levels located 10-20 m 2406 below the pycnocline (Mordy et al., 2012). Phytoplankton present in the water column during late 2407 fall, when ice begins to form, can be incorporated into the ice with large diatom cells 2408 preferentially selected (Gradinger and Ikavalko1998, Niemi et al., 2011). Ice algal cells may grow 2409 slowly during winter when light levels are very low (Melnikov 1998) and then begin to grow 2410 faster when light increases in spring. 2411 In this paper we focus on the middle domain of the eastern Bering Sea shelf where four 2412 oceanographic moorings have been located. The measurements on the moorings include 2413 temperature and chlorophyll a fluorescence. In summer, the middle domain is strongly stratified 2414 into two layers, with a wind-mixed upper layer and tidally-mixed lower layer. The middle domain 100 2415 typically extends from the 50 m isobath to the 100 m isobath, and is bounded by oceanic fronts or 2416 transition zones (Iverson et al., 1979). In winter, the middle domain is usually well mixed and 2417 cold, with a large part (>50%) ice-covered. These four oceanographic moorings provide the 2418 longest, daily record of in situ oceanographic measurements in the eastern Bering Sea. This paper 2419 is the first examination of the chlorophyll a fluorescence data, excepting previous analyses of the 2420 spring bloom at the southernmost mooring (Hunt et al., 2002, 2011; Stabeno et al., 2001). In this 2421 paper our objectives are to: characterize spring and fall blooms over the eastern Bering Sea 2422 middle shelf; relate their timing and strength to physical characteristics including spring ice 2423 retreat and fall overturn; and discuss some implications of these results for one of the large 2424 crustacean zooplankton taxa characteristic of that domain (Calanus spp.). 2425 2426 2. Data and methods 2427 2.1. Moorings 2428 Four oceanographic moorings have been 2429 deployed along the 70-m depth contour of the eastern 2430 Bering Sea shelf with two southern locations sampled 2431 almost continually since 1995 (M2) and 1999 (M4), and 2432 two northern locations since 2004 (M8) and 2005 (M5) 2433 (ure 1). Prior to 2005, moorings were recovered and 2434 redeployed twice a year, once in the spring (April/May) 2435 and again in the late summer or early fall (September / 2436 October). Since 2006, there has been extensive ice on 2437 the northern shelf in spring and M8 (and sometimes M5) 2438 has only been recovered and redeployed once a year in 2439 August or September. Data collected by instruments on 2440 the moorings included temperature (miniature temperature recorders, SeaBird3 SBE-37 and SBE- 2441 39) and chlorophyll a fluorescence (WET Labs DLSB ECO Fluorometer). A transition to 2442 fluorometer sensors with wipers that sharply reduced fouling occurred during 2001-2004. Data 2443 were collected at least hourly and instruments were calibrated according to manufacturer’s 2444 specification prior to deployment. During autumn, winter, and early spring, the shallowest 2445 instrument was at 11 m at M2 and M4, at 15 m at M5 and at 20 m at M8. During late spring to 2446 early autumn (the ice-free period), the mooring at M2 included a surface toroid measuring 3 Figure 9. Study area and mooring locations. Use of trade names does not constitute an endorsement by NOAA 101 2447 temperature at a depth of 1 m, and the upper instrument at M4 was at 11 m, as was the upper 2448 instrument at M5 and M8 if a summer mooring was deployed. For consistency, our analyses 2449 focus on data recorded at 11 m (or the shallowest instrument at M5 and M8 during autumn, 2450 winter and early spring). This standardization is reasonable because for parallel data sets collected 2451 at both 1 and 11 m at M2 and M4, the data recorded at 11 m captured 96% of variability recorded 2452 at 1 m (Stabeno et al., 2007). At these mooring locations, the water column is well-mixed during 2453 winter and develops a 15-35 m wind-mixed layer once stratification develops during spring 2454 (Stabeno et al., 2007). Farther north, at M8 and possibly M5, the spring bloom can occupy a 2455 narrower depth range and slowly sink, so that identified bloom strengths and timings are 2456 approximate. 2457 Additional data were collected when the moorings were deployed and recovered and 2458 were used for quality control of the chlorophyll a fluorescence data collected by mooring 2459 instruments. The additional data included chlorophyll a fluorescence measurements made with a 2460 Seabird SBE 911 plus system with chlorophyll a fluorescence sensors (WET Labs WETStar). 2461 Data were recorded during the downcast, with a descent rate of usually 15 m min-1 to a depth of 2462 35 m, and 30 m min-1 below that. The additional data also included water samples for extracted 2463 chlorophyll a collected during CTD casts. The water samples were filtered through glass fiber 2464 filters (nominal pore size 0.7 µm), then frozen at -80 °C until analysis. Frozen chlorophyll a 2465 samples were analyzed with a calibrated benchtop Turner TD-700 fluorometer using standard 2466 acidification methods (Parsons et al., 1984). 2467 Conversion of in vivo chlorophyll a fluorescence (volts) to chlorophyll a concentration 2468 (µg l-1) was performed for both the moored and CTD chlorophyll a fluorescence sensors using 2469 relationships provided by the manufacturer for each instrument during annual service. We did not 2470 attempt to create a relation between mooring chlorophyll a fluorescence and extracted chlorophyll 2471 a from water samples due to the limited number of water samples at deployment and recovery of 2472 the moorings and the difficulty of matching the timing and location of the water samples and 2473 mooring measurements. The chlorophyll a estimates based on the fluorescence sensors were 2474 compared to the chlorophyll a water samples collected during CTD casts for quality control of the 2475 mooring-based measurements; when high (>40 µg l-1) values of chlorophyll a occurred or isolated 2476 spikes occurred, these measurements were excluded from the analysis. In addition, the mooring– 2477 based measurements also were reviewed for measurement drift due to fouling and when drift 2478 occurred, these measurements were excluded from the analysis. Typically drift did not occur 2479 during winter and was more common during summer, which was mostly eliminated for 2480 instruments with wipers. 102 2481 2482 2.2. Sea Ice 2483 Two sources of sea-ice data were used. The first source was the National Ice Center 2484 (NIC), with data available from 1972 to 2005; the second source was the Advanced Microwave 2485 Scanning Radiometer EOS (AMSR), with data available from 2002 to 2012 (Spreen et al., 2008). 2486 These two data sets provide data over the entire period (1972–2012) for which high-quality data 2487 on sea-ice extent and areal concentration are available. We downloaded data available from the 2488 NIC website (http://www.natice.noaa.gov) for our study period (1995-2005) which were 2489 interpolated to a 0.25 degree grid. NIC data are derived from a variety of sources including the 2490 Advanced Very High Resolution Radiometer (AVHRR) aboard the Polar Orbiting Environmental 2491 Satellites (POES). AMSR data consist of daily ice concentration at 12.5 km resolution, which are 2492 available from the National Snow and Ice Data Center (NSIDC; http://nsidc.org). To examine 2493 how the ice cover varies along the 70-m isobath, a 100 km by 100 km box was defined around 2494 each of our biophysical moorings (M2, M4, M5, and M8) maintained by NOAA. AMSR and NIC 2495 data overlap during the four-year period 2002-2005, during which time they have very similar 2496 values (M2, Stabeno et al., 2012b). To span the period 1972-2012, we used both NIC and AMSR 2497 data, using the average value in the overlap years to derive the annual cycle of percent ice cover 2498 for each mooring location. 2499 2500 2.3. Wind 2501 Winds were estimated using daily data from the National Centers for Environmental 2502 Prediction (NCEP)/National Center for Atmospheric Research (NCAR) Reanalysis (Kalnay et al., 2503 1996). We follow the procedure used in Bond and Adams (2002) to specify particular elements of 2504 the atmospheric forcing on a daily basis for selected periods and interpolated wind velocity to the 2505 locations of four moorings (Fig. 1). The daily winds from the reanalysis are reliable in this region 2506 based on a comparison to independent buoy measurements from 1995 to 2000 (Ladd and Bond, 2507 2002). 2508 2509 2510 2.4. Data analysis Ice cover for each mooring and year was examined to determine if ice was present at any 2511 time that winter or spring and if present, when the ice retreated for the last time. Ice retreat was 2512 considered to have occurred when ice cover fell below 15% for the last time during that spring. 2513 The temperature records for each mooring and year were examined to determine: 1) when the 2514 ocean began warming following ice retreat; and 2) when fall overturn occurred. When ice was 103 2515 present the temperature was approximately -1.7 °C. Warming was considered to have started 2516 when the near surface temperature rose above -1 °C for the last time that spring. Fall overturn 2517 was considered to have occurred once temperature (at 11 m) fell 2 °C below the summer 2518 maximum. 2519 We also considered basing timing 2520 of the fall overturn on timing of mixed 2521 layer depth deepening. Unfortunately, the 2522 mooring data at M5 and M8 were 2523 inadequate to do this. We tested whether 2524 timing of mixed layer depth deepening and 2525 fall temperature drop were related, which 2526 would support using fall temperature drop 2527 as a measure of stratification breakdown 2528 and the resultant nutrient replenishment 2529 that could support a fall bloom. We found 2530 that they are related (Fig. 2) (Pearson’s 2531 product-moment correlation = 0.61, df = 20, p = 0.002). The relationship was stronger for M4 2532 (Pearson’s product-moment correlation = 0.95, df = 6, p < 0.001) than M2 (Pearson’s product- 2533 moment correlation = 0.50, df = 12, p = 0.07), but the latter relationship was strongly influenced 2534 by one data point. In 1997, the mixed layer depth deepened about 40 days after the temperature 2535 dropped, which is later than usual and may have occurred because winds were weak that late 2536 summer and early fall. If this data point is excluded, the relationship for M2 is much better 2537 (Pearson’s product-moment correlation = 0.76, df = 11, p = 0.003). Figure 10. The relationship of mixed layer depth deepening (day) and temperature drop (day) for moorings M2 and M4. The mooring data at M5 and M8 were inadequate to compute mixed layer deepening. 2538 The average wind speed records by calendar day and location were examined to 2539 determine when the first stormy period occurred after day 210 (ca. July 27). The wind data were 2540 divided into 10-day periods; if daily average wind was greater than 11 m s-1 for half or more of 2541 the ten-day period, then this period was defined as stormy (Sullivan et al., this issue). 2542 Fluorescence for each mooring and year was examined to determine the time and 2543 magnitude of the maximum value in spring and fall. These records were plotted and the times and 2544 magnitudes of the spring and fall blooms were assigned (Appendix Figure 1). Each year, the 2545 spring bloom was assigned to the maximum fluorescence value before day 180 (ca. June 27) and 2546 the fall bloom was assigned to the maximum fluorescence value after day 210 (ca. July 27). In 2547 some years the fluorescence record was discontinuous and the maximum value could not be 2548 determined (e.g., spring bloom at M2 in 1996. The values are tabled in Appendix Table 1. 104 2549 2550 2. 5. Comparison to shelf-wide patterns 2551 for fluorescence 2552 Satellite ocean color data were 2553 examined to understand the spatial 2554 pattern of chlorophyll a over the middle 2555 shelf. In particular, we wanted to find 2556 out whether it was reasonable to assume 2557 that the mooring fluorescence data were 2558 representative of fluorescence values 2559 over a much larger area. To do so, we 2560 retrieved eight-day composite Level-3 2561 SeaWiFS and MODIS-Aqua 2562 chlorophyll a data at 9-km spatial 2563 resolution using the Giovanni online data system, developed and maintained by the NASA 2564 Goddard Earth Sciences Data and Information Services Center (Acker and Leptoukh 2007). Data 2565 for a rectangular region (54 – 66°N and 157 – 180°W) that encompasses the eastern Bering Sea 2566 shelf were extracted from the global coverage 2567 datasets (units: mg m-3). Open ocean values 2568 (offshore of the 500 m isobaths) and nearshore 2569 values (inshore of 10 m isobaths) were excluded. 2570 We compared eight full years of overlap between 2571 the two data sets (2003-2010) based on correlations 2572 and absolute differences (bias) of the 2573 logarithmically transformed data on a pixel-by- 2574 pixel basis. Estimates from the two satellites were 2575 strongly correlated (r = 0.65) and bias was found to 2576 be negligible (median bias < 0.01, 88% of 1.4 2577 million paired observations had absolute bias < 2578 0.05). The SeaWiFS data from 1998-2002 were 2579 combined with the MODIS-Aqua data from 2003- 2580 2011 to produce the longest possible continuous Figure 11. A boxplot of ice retreat (day) by mooring. The box extends from the first quartile to the third quartile, the heavy line dividing the box is the median and the whiskers are the smallest and largest values. The number of years with ice present were 12 (M2), 15 (M4), 17 (M5) and 17 (M8). Figure 4. Scatterplots of maximum spring bloom strength (ug Chl l-1), spring bloom timing (day), minimum winter-spring temperature (day), day temperature rose above -1 ºC and day ice cover fell below 15%. If ice was absent that year, then the ice retreat day is zero. 105 2581 time series of remotely-sensed chlorophyll a data. Finally, we correlated eight-day composites of 2582 the fluorescence data with 8-day remotely-sensed chlorophyll a data for all non-missing pairs on 2583 a pixel-by-pixel basis to produce correlation maps for each of the four moorings. 2584 2585 2.6. Zooplankton Metabolism 2586 We examined some implications of these results for a large crustacean zooplankton taxa 2587 characteristic of that domain (Calanus spp.).The effect of temperature on the respiration of late 2588 stage Calanus spp. was approximated from literature values. We used a Q10 of 2.77 and nitrogen- 2589 specific respiration of Calanus finmarchicus (130 µmol O2 gN-1 h-1; Saumweber and Durbin, 2590 2006), average bottom temperatures in cold and warm years (-1.8 and 2°C, respectively; Stabeno 2591 et al., 2012a), wet weight measurements on preserved Calanus spp. C5 obtained from the eastern 2592 Bering Sea during August and September (1.82 x 10-3 g), and literature values for the relationship 2593 of dry to wet weight (15%) and nitrogen to dry weight (8%) for copepods to estimate respiration 2594 rates. 2595 2596 3. Results 2597 3.1. Spring 2598 Ice was present for about two 2599 out of three years at M2 (12 of 17), 2600 nearly all years at M4 (15 of 17), and all 2601 years at M5 and M8 (17 of 17). When 2602 ice was present, retreat was significantly 2603 (ANOVA, p < 0.001, df = 3, 57) earlier 2604 in the south (mean M2: Day 88; M4: 99; 2605 M5: 130; M8: 138) (Fig. 3). In some 2606 years when ice was present, retreat 2607 occurred before mid-March (M2 during 2608 1998, 2000, 2002, 2006 and M4 during 2609 2000 and 2002) (Appendix Table 1). Ice 2610 was absent at M2 during 1996, 2001 and 2611 2003-2005 and at M4 during 2001 and 2612 2005. Ice presence reduced ocean 2613 temperature below -1ºC in all cases, but 2614 one (ice was present at M2 in 2002, but for only 11 days, and minimum temperature was -0.1°C). Figure 5. Scatterplots of spring bloom maximum (day) versus ice retreat (day) by mooring. If ice was absent that year, then the ice retreat date is zero. The diagonal dashed line is the 1:1 line to compare timings of spring bloom and ice retreat. The vertical dashed line is March 15. 106 2615 The timing of warming (near- 2616 surface temperature > -1°C) was 2617 strongly correlated with the time of ice 2618 retreat (Pearson’s r = 0.96, df = 40, p < 2619 0.001). The interval between ice retreat 2620 and warming was longer in the north 2621 (M2 warming averaged Day 100; M4: 2622 110; M5: 153; M8: 181, corresponding 2623 to intervals for M2 of 12 days; M4: 11; 2624 M5: 23; M8: 43). 2625 The spring bloom usually Figure 6. Scatterplot of the observed (number) and fitted (line, based on simple linear regression, y-intercept = 53, slope = 0.66, df = 1, 24, p < 0.001) values of spring bloom maximum (day) versus ice retreat (day) for all moorings when ice was present after March 15. The number indicates mooring number. 2626 reached only one peak (34 of 37 cases) 2627 (Appendix Figure 1), so that the 2628 maximum value and when the 2629 maximum value occurred are reasonable measures of the strength and timing of the spring bloom. 2630 Double rather than single spring peaks occurred at M2 in 2003 and 2011 and at M4 in 1997. 2631 During years when ice was absent (or was present, but retreated before mid-March), the 2632 spring bloom maximum occurred in late May to early June (average day 148, SE = 3.5, n = 11) 2633 (Fig. 4, panel row 2, column 5). This pattern occurred for M2 and M4 but not M5 or M8 where 2634 ice always was present after mid-March 2635 (Fig. 5). There was no statistically 2636 significant difference in timing of the spring 2637 bloom maximum in years when ice was 2638 absent (average day 141) and when ice was 2639 present, but retreated before mid-March 2640 (average day 153) (two-way t-test, df = 9, p 2641 = 0.10). During years when ice was absent 2642 or retreated before mid-March, spring bloom 2643 maximum averaged 19 µg Chl l-1 (log- 2644 transformed, SE = 1.2, n = 11) (Fig. 4, panel 2645 row 1, column 5). There was no statistically 2646 significant difference in spring bloom 2647 maximum in years when ice was absent (average 25 ug Chl l-1) and when ice retreated before 2648 mid-March (average 15 ug Chl l-1) (log-transformed, two-way t-test, df = 9, p = 0.14). Figure 7. A boxplot of fall overturn (day) by mooring. The box extends from the first quartile to the third quartile, the heavy line dividing the box is the median and the whiskers are the smallest and largest values. The numbers of years when fall overturn could be determined were 15 (M2), 10 (M4), 16 (M5) and 7 (M8). 107 2649 In contrast to the late May to 2650 early June timing of the spring bloom 2651 in years when ice was absent or 2652 retreated early, if ice was present after 2653 mid-March (day 75), an ice-associated 2654 bloom occurred between early April 2655 and mid-June and the bloom timing 2656 was related to ice retreat timing (Fig. 4, 2657 panel row 2, column 5), regardless of 2658 mooring (Fig. 5). Later blooms 2659 occurred when ice retreat was later 2660 (linear regression, y-intercept = 53, 2661 slope = 0.66, df = 1, 24, p < 0.001) 2662 (Fig. 6). This relationship implies that 2663 bloom day is 119 when ice retreat 2664 occurs on day 100, 152 when ice 2665 retreat occurs on day 150; and 172 when ice retreat occurs on day 180. There was no statistically 2666 significant difference in spring bloom strength whether ice was absent or present, but retreated 2667 before mid-March (average 19 µg Chl l-1) or ice was present after mid-March (average 17 µg Chl 2668 l-1) (log-transformed, two-way t-test, df = 35, p = 0.70); the overall average spring bloom strength 2669 was 17 µg Chl l-1 (log-transformed, SE = 1.2, n = 37). Figure 8. Scatterplots of maximum fall bloom strength (ug Chl l-1), fall bloom timing (day), maximum summer-fall temperature (ºC), fall overturn (day) and first storm (day). 2670 2671 3.2. Fall 2672 Fall overturn timing was similar among moorings on average (M2 mean was Day 259; 2673 M4: 261; M5: 259; M8: 268 (ANOVA, df = 3, 34, p = 0.49)), but was more variable at M2 and 2674 M4 (Fig. 7). Sometimes an early major storm prompted early fall overturn; on five occasions 2675 when overturn was earlier than usual (approximately day 240 versus a range of 250-280), the day 2676 of the first major storm was earlier than usual (about day 240 versus a range of 240-320) (Figure 2677 8, panel row 4, column 5). 108 2678 The fall bloom usually reached 2679 only one peak (30 of 33 cases) (Appendix 2680 Figure 1), so that the maximum value and 2681 when the maximum value occurred are 2682 reasonable measures of the strength and 2683 timing of the fall bloom. Double rather 2684 than single fall blooms occurred at M2 2685 once (1998) and M4 twice (2004, 2007). 2686 In six cases (1996 M4, 2002 M2, 2005 2687 M4, 2007 M2, 2009 M5, 2010 M8), only 2688 the descending limb of the fall 2689 fluorescence data is available due to 2690 instrument failure, hence the fall bloom 2691 peak is uncertain and no values for 2692 magnitude or timing were assigned. 2693 There was no significant Figure 9. Scatterplots of fall bloom maximum (day) versus fall overturn (day) by mooring. The diagonal dashed line is the 1:1 line to compare timings of fall bloom maximum and fall overturn. The horizontal dashed line is August 15. 2694 relationship between fall overturn timing and fall bloom timing (linear regression, df = 1, 27, p = 2695 0.88) (Fig. 8, panel row 2, column 4) or fall bloom strength (linear regression, log-transform, df = 2696 1, 27, p = 0.49) (Fig. 8, panel row 1, column 4). 2697 Fall bloom timing was similar among moorings (M2 mean: Day 276; M4: 277; M5: 258; 2698 M8: 281) (ANOVA, p = 0.32, df = 3, 29) (Fig. 9). Fall bloom strength also was similar among 2699 moorings (ANOVA, log-transform, p = 0.93, df = 3, 30) (Fig. 9). On average, the fall bloom 2700 occurred on day 274 (late September) (SE = 4.2, n = 33) with strength 8 µg Chl l-1 (SE = 1.2, n = 2701 34). 2702 The strengths of the spring and fall blooms were correlated (Pearson’s r = 0.46, df = 28, p 2703 = 0.011, log-transformed values) (Fig. 10). The interval of time between the spring and fall 2704 blooms ranged from about 4-6 months, with longer intervals occurring for earlier spring blooms 2705 (linear regression, intercept = 294, slope = -1.16, df = 1, 27, p < 0.001) (Fig. 11). 2706 2707 3.3. Comparison to shelf-wide patterns 109 2708 Eight-day composites of chlorophyll a values at both M2 and M4 were moderately to 2709 strongly correlated with remotely-sensed chlorophyll a values over broad regions of the eastern 2710 shelf (Appendix Figure 2), including the middle shelf in the southeast and most of the northern 2711 Bering Sea except Norton Sound. In contrast, 2712 chlorophyll a values at M5 were positively 2713 correlated with remotely-sensed values over 2714 much of the shelf north of about 60°N, but 2715 negatively correlated with remotely-sensed 2716 values to the south of M5. Chlorophyll a 2717 values at M8 were positively correlated with 2718 remotely-sensed values over the northwestern 2719 part of the shelf and much of the outer shelf 2720 and negatively correlated with remotely- 2721 sensed values on much of the inner shelf. 2722 These negative correlations are likely due to 2723 differences in bloom timing. Figure 10. Scatterplot of maximum spring bloom strength (ug Chl l-1) and maximum fall bloom strength. The number indicates mooring number. 2724 2725 4. Discussion 2726 4.1. Spring 2727 We found that in the eastern Bering 2728 Sea: if ice was present after mid-March, 2729 spring bloom timing was related to ice 2730 retreat timing; if ice was absent or retreats 2731 before mid-March, a spring bloom usually 2732 occurred in May or early June. Our findings 2733 generally match Brown and Arrigo (2013) 2734 based on their satellite-based observations 2735 (see exception below), as well as those of 2736 Hunt et al. (2002, 2011) and Stabeno et al. 2737 (2001). In general, ice-associated 2738 phytoplankton blooms are observed near the 2739 retreating ice edge on shipboard surveys and in ocean color data, due to melting ice increasing the 2740 stability of the water column (Alexander and Niebauer, 1981; Niebauer et al., 1995; Hunt et al., 2741 2010; Brown et al., 2011). While the spring bloom usually moves northward as the eastern Bering Figure 11. Scatterplot of observed (number) and fitted (line, based on linear regression, intercept = 294, slope = -1.16, df = 1, 27, p < 0.001) values for all moorings. The y=axis is the interval between the spring and fall bloom maximum (days). The x-axis is the spring bloom maximum (day). The number indicates mooring number. 110 2742 Sea becomes ice free, sometimes ice melts in the north before disappearing farther south and a 2743 spring bloom occurs in the northern ice-free area before it occurs in the south if light level is 2744 sufficient. For example in 2010, the northern Bering Sea melted earlier than in the south leaving 2745 an area of persistent ice between M2 and M4. Percent ice cover also influences whether or not 2746 ice-associated blooms occur by modulating both light and stratification (Sullivan et al., this 2747 issue). 2748 The relationship between the timing of the spring bloom and ice retreat when ice retreat 2749 occurred after mid-March was statistically significant for the eastern Bering Sea (pooled data 2750 from the north and south; Figure 6), which contradicts Brown and Arrigo (2013), who concluded 2751 that ice retreat timing was a good predictor of spring bloom timing for only the northern Bering 2752 Sea. The different conclusions for the southeastern Bering Sea likely resulted from how the two 2753 data sets were analyzed. Brown and Arrigo (2013) classified their data into four regions 2754 corresponding to M2, M4, M5 and M8 (i.e., similar geographic grouping to our approach). In one 2755 analysis, Brown and Arrigo (2013) regressed spring bloom timing versus ice retreat timing and 2756 found significant relationships for the northern, but not southeastern Bering Sea. In a second 2757 analysis, they classified the southeastern Bering Sea data based on whether or not ice retreat 2758 occurred before (early) or after (late) the mean ice retreat date for the two regions (M2 and M4) 2759 and whether the bloom occurred within (ice-edge bloom) or after (open-water bloom). Open- 2760 water blooms were defined as those occurring 20 days or more after ice retreat. They found no 2761 significant difference in mean timing between open-water blooms of early retreat (or ice free) 2762 years and ice-edge blooms of late retreat years. 2763 Like their first analysis, we regressed spring bloom timing versus ice retreat timing, but 2764 instead classified the data into two groups: ice absent or ice retreat before March 15; and ice 2765 retreat after March 15 following Hunt et al., (2002, 2011). Mid-March was chosen as the time 2766 when sufficient light becomes available for a phytoplankton bloom in the southeastern Bering Sea 2767 (Hunt et al. 2002). We pooled all mooring data for years when ice was present after mid-March 2768 (Figure 6) to test whether ice retreat and the spring bloom move northward as fson progresses. 2769 We suspect that if the Brown and Arrigo (2013) data set were similarly analyzed (regress bloom 2770 timing versus ice retreat timing when ice retreat occurred after mid-March), our results would 2771 match and both data sets would support the conclusion that the spring bloom generally moves 2772 northward as the eastern Bering Sea becomes ice free, and that the bloom occurs later when the 2773 ice retreats later. 2774 2775 The present work corrects and clarifies the relationship of spring bloom timing and ice retreat timing reported by Hunt et al. (2002) for the southeastern Bering Sea. Their conclusion of 111 2776 late ice retreats with early ice-related blooms oversimplifies the data; while supported by early 2777 ice-related blooms for 1995, 1997 and 1999, later ice-related blooms occurred for 1975 and 1976 2778 (Hunt et al., 2002, their Figure 7). There is some evidence that there were early blooms under the 2779 ice in these two years, but the data are not very reliable. With more years of data, Hunt et al. 2780 (2011) observed that because ice can remain in the southeastern Bering Sea until May or rarely 2781 June, the calendar date of an ice-associated bloom could be later than that of an open-water 2782 bloom (i.e., if the ice retreat is late, the spring bloom also is late). 2783 In addition to ice retreat timing, wind patterns can affect spring bloom timing. When ice 2784 was absent or retreated before mid-March, spring bloom timing averaged day 148 (May 28) and 2785 varied from day 132 (May 12) to 166 (June 15). Short-lived surface pulses of chlorophyll a can 2786 occur with early ice retreat during a temporary relaxation of winds without significant 2787 stratification (Brown and Arrigo, 2013; Stabeno et al., 2010) and precede the larger primary 2788 bloom in May or early June. When ice retreat occurs after mid-March in the southeastern Bering 2789 Sea, a very early bloom can occur when strong winds advect ice which melts in place during a 2790 temporary relaxation of winds (Brown and Arrigo, 2013; Stockwell et al., 2001). This scenario 2791 matched the two earliest spring blooms we observed in the southeastern Bering Sea (1997 at M2; 2792 2003 at M4, Appendix Table 1). An open-water bloom subsequent to the ice-related spring bloom 2793 also can occur after a strong mixing event. This occurred in 1997, with a strong ice associated 2794 bloom in April and then a very strong storm in late May that resulted in an open water bloom. 2795 We summarize these findings on spring bloom timing and ice retreat timing (ours; Brown 2796 and Arrigo 2013; Hunt et al., 2002, 2011) as follows. If ice retreats after mid-March, an ice- 2797 associated bloom occurs; this pattern applies throughout the eastern Bering Sea. If the ice retreats 2798 before mid-March, there is no ice-associated bloom because sunlight is insufficient to initiate an 2799 ice-associated bloom. To date, early ice retreat occurs only in the southeastern Bering Sea and not 2800 in the northern Bering Sea; this pattern of persistent ice in the northern Bering Sea is expected to 2801 continue into the foreseeable future (Stabeno et al., 2012a). An open-water bloom occurs if ice 2802 retreats before March 15 or ice is absent; this pattern applies only in the southeastern Bering Sea. 2803 The strengths of spring blooms were similar for years when ice retreated after mid- 2804 March, and for years when ice retreated before mid-March or was absent. We used the maximum 2805 chlorophyll a value to measure spring bloom strength; its timing usually coincides with that of ice 2806 retreat, indicating that these values represent both ice algae and phytoplankton. We anticipated 2807 spring blooms for late retreat years would be weaker due to consumption of nutrients by ice algae 2808 prior to the spring bloom maximum; ice algae growth and the spring phytoplankton bloom 2809 together can create a prolonged period of production or two spring blooms (Niebauer et al., 1995, 112 2810 Stabeno et al., 2001). We found some evidence that ice algae production reduces the subsequent 2811 value of maximum chlorophyll a. The spring bloom maximum averaged 25 (µg chlorophyll a l-l) 2812 when ice was absent, 15 when ice retreated before mid-March and 17 when ice retreated after 2813 mid-March. Although not significant, these differences imply that phytoplankton bloom strength 2814 is diminished if preceded by ice cover and ice algal growth. In contrast to our findings, satellite- 2815 based estimates of open-water blooms of early retreat years were >70% more productive than the 2816 ice-edge blooms of late retreat years, but under-ice production was not quantified (Brown and 2817 Arrigo, 2013). An analysis of measurements of nutrient draw-down would help to resolve this 2818 question. 2819 2820 2821 4.2. Fall A fall bloom commonly occurred in both the northern and southeastern Bering Sea, on 2822 average in late September. Winds at M2, M4, M5, and M8 are significantly correlated (Stabeno et 2823 al., 2010), which will tend to synchronize fall blooms, as was observed. However bloom timing 2824 was not significantly related to either storm or fall overturn timing. A timing effect for the fall 2825 bloom may be difficult to detect because the month-long interval when blooms usually occurred 2826 is short and because the timing is affected by several interacting factors including wind strength, 2827 stratification, fall cooling and light level. For example, fall overturn requires strong winds, but 2828 once cooling begins in late September and early October, less wind energy is necessary to 2829 overturn the ocean. A fall bloom also depends on sufficient light, introduction of nutrients from 2830 below the pycnocline and short periods of stabilization that allow phytoplankton to remain in the 2831 sunlit waters and grow. 2832 The timing of the fall overturns at M2 and M4 were much more variable than at M5 and 2833 M8 which may be due to north-south differences in stratification. In the south, stratification is 2834 almost completely controlled by temperature, while in the north it is equally controlled by 2835 temperature and salinity so variations in temperature have more effect on stratification in the 2836 south than in the north. This contrast implies that the southeastern Bering Sea is more sensitive to 2837 the timing of fall cooling than the northern Bering Sea. 2838 Our mooring data and previous middle-shelf observations (Rho and Whitledge, 2007; 2839 Brown et al., 2011) indicated that the strengths of fall blooms were weaker than spring blooms on 2840 average. It may be that nutrients are limiting and storm mixing can’t provide sufficient 2841 replenishment of nutrients for a strong fall bloom or the bloom is limited by grazing. For instance, 2842 if the lower layer has 20 μM l-1 of nitrate and the surface mixed layer is 20 m deep and depleted 2843 of nitrate, then deepening the mixed layer by 5 m will result in only 4 μM l-1 of nitrate, compared 113 2844 to 18 μM l-1 available for consumption in the top 20 m of water column in spring. In addition, 2845 variations in chlorophyll a reflect the result of multiple processes including phytoplankton 2846 production, grazing, sinking, and advection. Chlorophyll a will not increase until cell growth 2847 exceeds losses by grazing and other factors. While grazing impact has been measured for spring 2848 (Cooney and Coyle 1982; Sherr et al., 2013), grazing impact has not been measured for fall, so 2849 comparing grazing pressure on spring and fall blooms is not possible. Finally, phytoplankton 2850 physiological status will impact the bloom intensity, with healthier cells growing faster, so 2851 phytoplankton populations in spring may grow faster than those in fall. 2852 Broad-scale sampling of the eastern Bering Sea shelf found that late summer and early 2853 fall chlorophyll a values were higher during warm years compared to cold years (Eisner et al., 2854 2012). Ocean color data for 1998-2007 in the Bering Sea also show that annual mean chlorophyll 2855 a and total primary production were higher in warm years than cold years (Brown et al., 2011, 2856 Mueter et al., 2009). We used the warm-average-cold year classification of Stabeno et al. (2012) 2857 to classify our fall bloom data. Likewise we found that fall bloom strength was stronger in warm 2858 (average = 12.3 µg Chl l-1, log-transform) versus cold (6.3) and average (8.6) years, but the 2859 differences were not statistically significant (ANOVA, log-transform, df = 2, 31, p = 0.32). 2860 2861 4.3 Spring/fall comparisons 2862 Spring and fall bloom strengths were related, implying that a common factor influences 2863 spring and fall primary production (e.g., overwinter replenishment of nutrients). In addition, this 2864 relationship likely amplifies secondary production during good years (both spring and fall blooms 2865 tend to be strong) and vice versa, bad years (both spring and fall blooms tend to be weak). An 2866 analysis of nutrient information, comparing spring-fall differences by year, would help us to 2867 understand the mechanism for this relationship, but has not yet been published. 2868 The fall bloom occurred in late September on average, and the timing was less variable 2869 than for the spring bloom (varies over ~60 day compared to ~120 day period) regardless of 2870 location, so the spring-fall interval largely depends on spring bloom timing. In the northern 2871 Bering Sea, where ice is present every year and usually retreats in late May (Fig. 5), the interval 2872 typically lasted four months (Fig. 11). The interval also typically lasted four months in the 2873 southeastern Bering Sea in years when ice was absent or retreated before March 15. In contrast, 2874 the interval lasted up to six months in the southeastern Bering Sea in years when ice was present 2875 after March 15, but retreated soon thereafter. 2876 2877 4.4. The representativeness of the fluorescence data 114 2878 Distinct spring and fall blooms occurred at these moorings located in the middle domain 2879 with the spring bloom stronger than the fall bloom. Likewise, a bimodal spring-fall pattern was 2880 found for net primary production for the middle domain with spring stronger than fall for both 2881 satellite-based (Brown et al., 2011) and temporally and spatially scattered in situ measurements 2882 (Rho and Whitledge 2007). During the summer interval between the spring and fall blooms, we 2883 found that chlorophyll a fluorescence was low in the middle domain, similar to Goes et al. (this 2884 issue). We also found that the mooring observations of chlorophyll a fluorescence were 2885 consistently correlated with satellite observations of chlorophyll a fluorescence in the middle 2886 domain at the same approximate latitude (Appendix Figure 2). This pattern implies that the 2887 mooring information represents conditions of the middle domain over a range of ca. 150-200 km. 2888 The bimodal pattern was replicated for the inner domain except that spring and fall bloom 2889 strengths appear similar for satellite-based measurements (Brown et al., 2011). In contrast, the 2890 fall bloom was less distinct for the outer domain (Rho and Whitledge 2007; Brown et al., 2012). 2891 These patterns imply that bloom patterns differ somewhat among the inner, middle and outer 2892 domains, and that the mooring information do not well represent the oceanographic conditions of 2893 the inner (influence of the coastal current, river runoff, and tidal and wind mixing ) or outer 2894 domains (influence of slope, episodic on-shelf flow and upwelling). The broad-scale data 2895 collected during the late summer BASIS sampling also indicates that chlorophyll a concentrations 2896 and phytoplankton particle size (large >= 10 um and small < 10 um) varied among shelf domains 2897 (Eisner et al., 2012). The relatively high concentrations found in the outer domain were primarily 2898 comprised of small taxa (< 25% large); chlorophyll a concentrations on average were lower in the 2899 middle domain during these surveys, but when increases in chlorophyll a were observed (usually 2900 near M4), there was a higher proportion of large taxa (40-70% large). Besides phytoplankton, 2901 mixotrophic ciliates (a taxonomic group of microzooplankton that also photosynthesize) also can 2902 contribute to total chlorophyll a measurements in summer (Stoecker et al., this issue). 2903 2904 4.5 Biological Implications 2905 4.5.1 Lower trophic levels 2906 Spring bloom timing predictably varied between early April and mid-June depending on 2907 ice presence/absence and ice retreat timing. In contrast, we found no statistically significant 2908 difference in spring bloom maximum among the independent factors examined including ice 2909 presence/absence and ice retreat timing. Thus phytoplankton biomass is the same, but the 2910 secondary community (zooplankton species) that benefits will depend on how close timing of 2911 their spring energy-intensive needs such as reproduction and awakening from winter diapause 115 2912 matches the timing of the spring bloom. Species that require an early pulse of energy will benefit 2913 from years when ice is present, but retreats in late March (conditions that tend to result in early 2914 April phytoplankton bloom). In contrast, species with a phenology timed for a late energy pulse 2915 will benefit from years with no ice (conditions that tend to result in a late May to early June 2916 bloom). 2917 These observations also imply that climate, through its connection to the production, 2918 transport, and dissipation of sea ice has the potential to affect the success of zooplankton 2919 populations and the strength of coupling between primary production and higher trophic levels. 2920 For example, the large crustacean zooplankton taxa Calanus spp. may benefit in years when ice is 2921 present after March 15, but retreats relatively early. Baier and Napp (2003) observed that spring 2922 Calanus concentrations in the southeastern Bering Sea were higher in cold years than in warm 2923 years, and that these individuals likely metamorphosed from naupliar to copepodite stages during 2924 the early spring bloom. They hypothesized that metamorphosis was a recruitment bottleneck and 2925 that an early spring bloom benefitted copepodite recruitment. The spring bloom occurs during 2926 April in years when ice is present, but retreats relatively early, which may promote strong 2927 recruitment of copepodites. In addition, colder winters with ice present likely reduces metabolism 2928 and lipid utilization by Calanus spp. and thus may promote winter survival (Coyle et al., 2011). 2929 Recruitment of Calanus spp. on the eastern Bering Sea shelf is complicated, involving 2930 many different processes. Life history strategies of large crustacean zooplankton (LCZ) in the 2931 eastern Bering Sea may be able to take advantage of the oscillations between warm and cold 2932 periods, regardless of whether or not there are stanzas of multiple years of the same conditions 2933 (Overland et al., 2012). The timing of reproductive events for the Calanus congener Calanus 2934 glacialis varies between different physical and biological environments of the Arctic (Daase et al. 2935 2013). The following scenarios were developed as a working hypothesis to explain some of the 2936 present observations and to serve as a guide for future research. In these scenarios, we simplified 2937 Calanus life history into four major steps: spawning, metamorphosis, accumulation of depot 2938 lipids and overwintering. These scenarios make the following assumptions: 1) the timing of 2939 spawning by Calanus spp. is protracted (from February to May) (Baier and Napp 2003 and 2940 references therein) and the longer that conditions are suitable, the more total eggs each individual 2941 will produce (E. Durbin, U. Rhode Island, pers. comm.); 2) early-spawned individuals enter 2942 diapause before late-spawned individuals; and 3) overwinter respiration is a function of 2943 temperature (Saumweber and Durbin 2006). The working hypothesis does not address the role of 2944 advection of individuals into or out of the area. 116 2945 Case A. Cold/Ice-Replete Years 2946 with Early Ice Retreat (Fig. 12). We 2947 hypothesize that access to under ice 2948 algae (beginning as early as mid 2949 February) increases egg production rates 2950 of early spawning Calanus (Runge and 2951 Ingram, 1991). Metamorphosis to the 2952 copepodite stage benefits from an early 2953 ice retreat and spring bloom production 2954 (Baier and Napp, 2003). Summer 2955 accumulation of depot lipids is 2956 dependent on summer primary and 2957 microzooplankton production 2958 (Sambrotto et al., 1986; Sherr et al., 2959 2013; Stoecker et al., this volume). 2960 Overwintering for individuals spawned 2961 in late winter begins relatively early and 2962 these individuals may not be able to take 2963 advantage of the fall bloom. The ability 2964 of individuals (C5 copepodites) to 2965 survive the following winter benefits 2966 from low respiration rates (RC5 copepodite = 5.66 µm 02 d-1 at -1.8 °C (see methods)) resulting from 2967 cold bottom temperatures. Reproductive output during the following year(s) is maximized when 2968 there are subsequent ice-replete years and under ice algae. 2969 Figure 12. Timing of Calanus reproduction relative to the presence of ice and ice algae, and the spring and fall blooms. The Calanus life history is simplified and the timings are approximate. Black is the period of ice cover; gray is the 50th percentiles for the timing of ice retreat. White rectangles are the 50th percentile dates for bloom maximum in the spring and fall. Ice algae blooms during ice cover (solid gray line) and ice retreat (dashed gray line) and begin as early as mid-February (R. Gradinger, personal comm.). A – a cold icy year, with ice retreat shortly after March 15; B – a cold icy year with ice retreat after April 14; C - no ice after March 15. Case B. Cold/Ice-Replete Years with Late Ice Retreat. In this scenario, both early and late 2970 spawning by Calanus spp. females benefits, as reproduction timing coincides with either the ice 2971 algae or the spring bloom. The nauplii and copepodites from late spawning would likely develop 2972 post-spring bloom (i.e., metamorphosis is not coincident with the spring bloom), and would 2973 depend upon summer primary and microzooplankton production to grow and accumulate depot 2974 lipids. The onset of overwintering occurs latest for the late-spawned progeny which may take 2975 advantage of the fall bloom to accumulate additional lipids before entering diapause. Ability to 2976 survive the overwintering period benefits from the cold bottom temperatures present in late fall 2977 through early spring. 117 2978 Case C. Warm/Ice-Deplete Years. Egg production by early spawning of Calanus is 2979 reduced by the absence or short duration of sea ice and under ice algae. The timing of the spring 2980 bloom is late; it benefits late spawning of Calanus spp. females. The late spring bloom may 2981 coincide with metamorphosis of the early-spawned nauplii to copepodites, but is mismatched 2982 with the metamorphosis from the late spawners. Growth and development are dependent upon 2983 summer primary and microzooplankton production. Diapause that is delayed until after the fall 2984 bloom benefits late spawners. Warm bottom temperatures result in higher respiration rates -- 47% 2985 higher daily respiration than during cold years (RC5 copepodite = 8.34 µm 02 d-1 at 2.0 °C). The impact 2986 on lipid reserves is exacerbated if the period without food (beginning of diapause to the following 2987 spring bloom) is longer in warm years relative to cold. Thus we hypothesize that survival of 2988 Calanus spp. over a warm winter is low relative to a cold winter. Predation losses would also 2989 likely be higher in warm winters at a given predator abundance due to higher metabolism and 2990 food requirements of fishes. Absence of sea ice for a second year results in low reproductive 2991 output by the early spawners, and a decrease in the population. 2992 2993 2994 4.4.2. Connections to Higher Trophic Levels The six to eight month variation in the fall to spring bloom interval also may contribute to 2995 the peak of the dome-shaped relationship between juvenile pollock production and sea surface 2996 temperature observed by Mueter et al. (2006, 2011) and Coyle et al. (2011). Pollock recruitment 2997 was generally high when ice retreated early and adult biomass was low (Mueter et al., 2006); we 2998 found that a six-month fall to spring bloom interval occurs in years with an early ice retreat. The 2999 likely mechanism is the relationship between early ice retreat, delayed spring bloom, and the 3000 production of large crustacean zooplankton as pollock recruitment was generally high when large 3001 crustacean zooplankton were abundant (Hunt et al., 2011). The mechanism linking secondary 3002 production and pollock recruitment has been attributed to enhanced energy content and 3003 overwinter survival of age-0 pollock during cold years when the lipid-rich, large crustacean 3004 zooplankton biomass is highest (Heintz et al., accepted). 3005 3006 3007 5. Conclusions ● In the eastern Bering Sea: if ice is present after mid-March, spring bloom timing is 3008 related to ice retreat timing; if ice is absent or retreats before mid-March, a spring bloom 3009 usually occurs in May or early June. 3010 3011 ● Spring and fall bloom strengths are related, implying that a common factor influences spring and fall primary production. 118 3012 ● We hypothesize that large crustacean zooplankton such as Calanus spp. benefit from 3013 cold, icy winters in the southeastern Bering Sea because ice algae or ice associated 3014 phytoplankton blooms provide an early spring food source and respiration rates are lower 3015 during cold winters. 3016 3017 Acknowledgements 3018 We thank D. Kachel for data analysis, S. Salo for providing the ice data and W. Floering and C. 3019 Dewitt for deploying and recovering the moorings. We thank the officers and crews of the NOAA 3020 ships Miller Freeman and Oscar Dyson for their invaluable assistance in deploying and 3021 recovering the moorings. We thank Zach Brown, Rolf Gradinger and one anonymous reviewers 3022 whose comments improved our manuscript. The research was generously supported by the North 3023 Pacific Research Board and NOAA’s North Pacific Climate Regimes and Ecosystem Productivity 3024 (NPCREP) and Fisheries Oceanography Coordinated Investigations programs. This is BEST- 3025 BSIERP publication no. XXX and NPRB publication no. YYY. This is contribution FOCI-B*** 3026 to the Ecosystems & Fisheries Oceanography Coordinated Investigations; and 3953 to Pacific 3027 Marine Environmental Laboratory. The findings and conclusions in this paper are those of the 3028 authors and do not necessarily represent the views of NOAA’s National Marine Fisheries Service 3029 or Oceans and Atmospheric Research. 3030 3031 References 3032 Acker, J.G., Leptoukh, G. 2007. Online analysis enhances use of NASA Earth science data. Eos, 3033 Transactions of the American Geophysical Union 88(2): 14. 3034 3035 Alexander V., Niebauer, H.J.1981. Oceanography of the eastern Bering Sea ice edge zone in 3036 spring. Limnology & Oceanography. 26: 1111-1125. 3037 3038 Baier, C.T., Napp, J.M. 2003. Climate-induced variability in Calanus marshallae populations. J. 3039 Plankton Res. 25: 771–782. 3040 3041 Baier, C.T., Terazaki, M. 2005. Interannual variability in a predator-prey interaction: climate, 3042 chaetognaths and copepods in the southeastern Bering Sea. J. Plankton Res., 27: 1113–1125. 3043 3044 Bond, N.A., Adams, J.M. 2002. Atmospheric forcing of the southeast Bering Sea Shelf during 3045 1995–99 in the context of a 40-year historical record. Deep-Sea Research II 49: 5869–5887. 119 3046 3047 Brown, Z.W., Arrigo, K.R. 2013. Sea ice impacts on spring bloom dynamics and net primary 3048 production in the eastern Bering Sea. J. Geophys. Res. 118, 1-20, doi: 10.1029/2012JC008034. 3049 3050 Brown, Z.W., van Dijken, G.L., Arrigo, K.R. 2011. A reassessment of primary production and 3051 environmental change in the Bering Sea. J. Geophys. Res. 116, C08014, doi: 3052 10.1029/2010JC006766. 3053 3054 Cooney, R.T., Coyle, K. O. 1982. Trophic implications of cross-shelf copepod distributions in the 3055 southeastern Bering Sea. Mar. Biol. 70: 187-196. 3056 3057 Coyle, K.O., Eisner, L.B., Mueter, F.J., Pinchuk, A.I., Janout, M.A., Cieciel, K.D., Farley, E.V., 3058 Andrews, A. G. 2011. Climate change in the southeastern Bering Sea: impacts on pollock stocks 3059 and implications for the Oscillating Control Hypothesis. Fish. Oceanogr. 20: 139–156. 3060 3061 Daase, M., Falk-Petersen, S., Varpe, Ø., Darnis, G., Søreide, J.E., Wold, A., Leu, E., Berge, J., 3062 Philippe, B., Fortier, L. 2013. Timing of reproductive events in the marine copepod Calanus 3063 glacialis: a pan-Arctic perspective. Can. J. Fish. Aquat. Sci. 70: 1-14. 3064 3065 Droop, M. R. 1973. Some thoughts on nutrient limitation in algae. J. Phycol. 9(3): 264-272. 3066 3067 Eisner, L.B., Cieciel, K., Gann, J. 2012. Phytoplankton biomass and size structure during late 3068 summer to early fall in the eastern Bering Sea. In Zador, S.[Ed.], Ecosystem Considerations 2012, 3069 Stock Assessment and Fishery Evaluation Report, North Pacific Fisheries Management Council, 3070 605 W 4th Ave, Suite 306, Anchorage, AK. 3071 http://www.afsc.noaa.gov/REFM/docs/2012/ecosystem.pdf 3072 3073 Fortier, M., Fortier, L., Michel, C., and Legendre, L. 2002. Climatic and biological forcing of the 3074 vertical flux of biogenic particles under seasonal Arctic sea ice. Mar. Ecol. Prog. Ser. 225:1-16. 3075 3076 Goes, J.I., Gomes, H.R., Haugen, E., McKee, K., D’Sa, E., Chekalyuk, A.M., Stoecker, D., 3077 Stabeno, P., Saitoh, S., Sambrotto, R. This issue. Fluorescence, pigment and microscopic 3078 characterization of Bering Sea phytoplankton community structure and photosynthetic 3079 competency in the presence of a cold pool during summer. Deep-Sea Res. II. 120 3080 3081 Gradinger, R., Ikavalko, J. 1998. Organism incorporation into newly forming Arctic sea ice in the 3082 Greenland Sea. J. Plankton Res. 20: 871-886. 3083 3084 Granger, J., Prokopenko, M.G., Sigman, D.M., Mordy, C.W., Morse, Z.M., Morales, L.V., 3085 Sambrotto, R.N., Plessen, B. 2011. Coupled nitrification-denitrification in sediment of the eastern 3086 Bering Sea shelf leads to 15N enrichment of fixed N in shelf waters. J. Geophys. Res.: Oceans 3087 (1978–2012) 116(C11) doi: 10.1029/2010JC006751 3088 3089 Grebmeier, J.M., Cooper, L.W., Feder, H.M., & Sirenko, B.I. 2006. Ecosystem dynamics of the 3090 Pacific-influenced northern Bering and Chukchi Seas in the Amerasian Arctic. Prog. 3091 Oceanogr. 71: 331-361. 3092 3093 Haecky, P., Jonsson, S., Andersson, A. 1998. Influence of sea ice on the composition of the 3094 spring phytoplankton bloom in the northern Baltic Sea. Polar Biol. 20: 1-8. 3095 3096 Heintz, R.A., Siddon, E.C., Farley, E.V., Napp, J.M. Accepted. Climate related changes in the 3097 nutritional condition of young-of-the-year pollock (Theragra chalcogramma) from the eastern 3098 Bering Sea. Deep-Sea Res. II 3099 3100 Hunt, G.L, Allen, B.M., Angliss, R.P., Baker, T., Bond,N., Buck, G. and others. 2010. Status and 3101 trends of the Bering Sea region, 2003-2008, pp. 196-267 In Mckinnell, S.M. and Dagg M.J. 3102 [Eds.], Marine Ecosystems of the North Pacific Ocean, 2003-2008. PICES Special Publication 4, 3103 393 p. 3104 3105 Hunt, G.L., Coyle, K.O., Eisner, L.B., Farley, E.V., Heintz, R.A., Mueter, F.J., Napp, J.M., 3106 Overland, J.E., Ressler, P.H., Salo, S., Stabeno, P.J. 2011. Climate impacts on eastern Bering Sea 3107 foodwebs: a synthesis of new data and an assessment of the Oscillating Control Hypothesis. ICES 3108 J. Mar. Sci. 68: 1230–1243. 3109 3110 Hunt, G.L., Stabeno, P.J., Walters, G., Sinclair, E., Brodeur, R.D., Napp, J.M., Bond, N.A. 2002. 3111 Climate change and control of the southeastern Bering Sea pelagic ecosystem. Deep-Sea Res. II. 3112 49: 5821–5853. 3113 121 3114 Iverson, R.L., Coachman, L.K., Cooney, R.T., English, T.S., Goering, J.J., Hunt, Jr., G.L., 3115 McCauley, M.C., McRoy, C.P., Reeburg, W.S., Whitledge, T.E. 1979. Ecological significance of 3116 fronts in the southeastern Bering Sea, pp. 437-466. In: Ecological Processes in Coastal and 3117 Marine Systems, R.J. Livingston (Ed.), Plenum Press. 3118 3119 Jin, M., Deal, C., Wang, J., Alexander, V., Gradinger, R., Saitoh, S., Iida, T., Wan, Z., Stabeno, 3120 P. 2007. Ice-associated phytoplankton blooms in the southeastern Bering Sea, Geophys. Res. 3121 Lett., 34, L06612, doi:10.1029/2006GL028849. 3122 3123 Kalnay, E., Kanamitsu, M., Kistler, R., Collins, W., Deaven, D., Gandin, L., Iredell, M., Saha, S., 3124 White, G., Woollen, J., Zhu, Y., Kirst G. and Wienchke C. 1995. Ecophysiology of polar algae. J. 3125 Phycol. 31: 181-199. 3126 3127 Kirst, G.O., Wiencke, C.1995. Ecophysiology of polar algae. Journal of Phycology, 31: 181–199. 3128 doi: 10.1111/j.0022-3646.1995.00181.x 3129 3130 Ladd, C., Bond, N.A., 2002. Evaluation of the NCEP-NCAR Reanalysis in the northeast Pacific 3131 and Bering Sea. J. Geophys. Res. 107, 3158. doi:10.1029/2001JC001157. 3132 3133 Melnikov, I.A. 1998. Winter production of sea ice algae in the western Weddell Sea. J. Mar. Sys. 3134 17: 195-205. 3135 3136 Mock T., and Gradinger R., 2000. Changes in photosynthetic carbon allocation in algal 3137 assemblages of Arctic sea ice with decreasing nutrient concentrations and irradiance. Mar. Ecol. 3138 Prog. Ser. 202: 1-11. 3139 3140 Mueter, F.J., Bond, N.A., Ianelli, J.N., Hollowed, A.B. 2011. Expected declines in recruitment of 3141 walleye pollock (Theragra chalcogramma) in the eastern Bering Sea under future climate change. 3142 ICES J. Mar. Sci. 68: 1284-1296. 3143 3144 Mueter, F.J., Broms, C., Drinkwater, K.F., Friedland, K.D., Hare, J.A., Hunt, G.L., Melle, W., 3145 Taylor, M. 2009. Ecosystem responses to recent oceanographic variability in high-latitude 3146 Northern Hemisphere ecosystems. Prog. Oceanography 81: 93-110. 3147 122 3148 Mueter, F.J., Ladd, C., Palmer, M.C., Norcross, B.L. 2006. Bottom-up and top-down controls of 3149 walleye pollock (Theragra chalcogramma) on the Eastern Bering Sea shelf. Prog. Oceanography 3150 68: 152-183. 3151 3152 Niebauer, H.J., Alexander, V., Henrichs, S.M. 1995. A time-series study of the spring bloom at 3153 the Bering Sea ice edge I. Physical processes, chlorophyll and nutrient chemistry. Cont. Shelf 3154 Res. 15: 1859-1877. 3155 3156 Niemi, A., Michel, C., Hille, K., Poulin, M. 2011 Protist assemblages in winter sea ice: setting the 3157 stage for the spring ice algal bloom. Polar Biol. 34: 1803-1817. 3158 3159 O'Brien, D.P. 1987. Direct observations of the behavior of Euphausia superba and Euphausia 3160 crystallorophias (Crustacea: Euphausiacea) under pack ice during the Antarctic spring of 1985. J. 3161 Crustacean Biol. 7: 437-448. 3162 3163 Overland, J.E., Wang, M., Wood, K.R., Percival, D.B., Bond, N.A. 2012. Recent Bering Sea 3164 warm and cold events in a 95-year context. Deep-Sea Res. II 65-70: 6-13. 3165 3166 Parsons, T.R., Maita, Y., Lalli, C.M. 1984. A manual of biological and chemical methods for 3167 seawater analysis. Pergamon Press, Oxford, 173pp. 3168 3169 Rho, T., Whitledge, T.E. 2007. Characteristics of seasonal and spatial variations of primary 3170 production over the southeastern Bering Sea shelf. Continental Shelf Research 27: 2556-2569. 3171 3172 Riebesell U., Schloss I., Smetacek V. 1991. Aggregation of algae form melting ice: implications 3173 for seeding and sedimentation. Polar Biol. 11: 239-248. 3174 3175 Runge, J.A., Ingram, R.G. 1991. Under-ice feeding and diel migration by the planktonic 3176 copepods Calanus glacialis and Pseudocalanus minutus in relation to the ice algal production 3177 cycle in southeastern Hudson Bay, Canada. Mar. Biol. 108: 217-225 3178 3179 Sambrotto R.N., H.J. Niebauer, J.J. Goering, and R.L. Iverson, 1986. Relationships among 3180 vertical mixing nitrate uptake and phytoplankton growth during the spring bloom in the southeast 3181 Bering Sea middle shelf. Cont. Shelf Res. 5: 161–198. 123 3182 3183 Saumweber, W.J., Durbin, E.G., 2006. Estimating potential diapause duration in Calanus 3184 finmarchicus. Deep-Sea Res. II 53: 2597-2617. 3185 3186 Sherr, E.B., Sherr, B.F., Ross, C. 2013. Microzooplankton grazing impact in the Bering Sea 3187 during spring sea ice conditions. Deep-Sea Res. II http://dx.doi.org/10.1016/j.dsr2.2013.03.019 3188 3189 Spreen, G., Kaleschke, L., Heygster, G. 2008. Sea ice remote sensing using AMSR-E 89 GHz 3190 channels. J. Geophys. Res. 113(C2), C02. 3191 3192 Stabeno, P.J., Bond, N.A., Kachel, N.B., Salo, S.A., 2007. On the recent warming of the 3193 Southeastern Bering Sea Shelf. Deep-Sea Res. II 54: 2599–2618. 3194 3195 Stabeno, P.J., Bond, N.A., Kachel, N.B., Salo, S.A., Schumacher, J. D. 2001. On the temporal 3196 variability of the physical environment over the southeastern Bering Sea. Fish. Oceanogr. 10: 81– 3197 98. 3198 3199 Stabeno, P.J., Farley, E., Kachel, N.B., Moore, S.E., Mordy, C.W., Napp, J.M., Overland, J.E., 3200 Pinchuk, A.I., Sigler, M.F. 2012a. A comparison of the physics of the northern and southern 3201 shelves of the eastern Bering Sea and some implications to the ecosystem. Deep-Sea Res. II. 3202 doi:10.1016/j.dsr2.2012.02.019 3203 3204 Stabeno, P.J., Kachel, N.B., Moore, S.E., Napp, J.M., Sigler, M.F., Yamaguchi, A., Zerbini, A.N. 3205 2012b. Comparison of warm and cold years on the southeastern Bering Sea shelf and some 3206 implications to the ecosystem. Deep-Sea Res. II. doi:10.1016/j.dsr2.2012.02.020 3207 3208 Stabeno, P.J., Napp, J., Mordy, C., Whitledge, T., 2010. Factors influencing physical structure 3209 and lower trophic levels of the eastern Bering Sea shelf in 2005: Sea ice, tides and winds. Prog. 3210 Oceanogr. 85(3–4), 180–196. 3211 3212 Stoecker, D.K., Weigel, A.C., Stockwell, D.A., Lomas, M.W. This issue. Microzooplankton: 3213 Abundance, biomass and contribution to chlorophyll in the eastern Bering Sea in summer. Deep- 3214 Sea Res. II. 3215 124 3216 Sullivan, M.E., Mordy, C.W., Salo, S.A., Stabeno, P.J. This issue. The influence of sea ice on 3217 seasonal water column changes on the southeastern Bering Sea Shelf. Deep-Sea Res. II. 3218 3219 Sverdrup, H. U. 1953. On conditions for the vernal blooming of phytoplankton. J. Conseil perm. 3220 int. Explor. Mer. 18: 287-295. 3221 3222 Tremblay, C., Runge, J., Legendre, L. 1989. Grazing and sedimentation of ice algae during and 3223 immediately after a bloom at the ice-water interface. Mar. Ecol. Prog. Ser. 56: 291-300. 3224 3225 Figure Caption 3226 3227 Figure 1. Study area and mooring locations. 3228 3229 Figure 2. The relationship of mixed layer depth deepening (day) and temperature drop (day) for 3230 moorings M2 and M4. The mooring data at M5 and M8 were inadequate to compute mixed layer 3231 deepening. 3232 3233 Figure 3. A boxplot of ice retreat (day) by mooring. The box extends from the first quartile to the 3234 third quartile, the heavy line dividing the box is the median and the whiskers are the smallest and 3235 largest values. The number of years with ice present were 12 (M2), 15 (M4), 17 (M5) and 17 3236 (M8). 3237 3238 Figure 4. Scatterplots of maximum spring bloom strength (ug Chl l-1), spring bloom timing (day), 3239 minimum winter-spring temperature (day), day temperature rose above -1 ºC and day ice cover 3240 fell below 15%. If ice was absent that year, then the ice retreat day is zero. 3241 3242 Figure 5. Scatterplots of spring bloom maximum (day) versus ice retreat (day) by mooring. If ice 3243 was absent that year, then the ice retreat date is zero. The diagonal dashed line is the 1:1 line to 3244 compare timings of spring bloom and ice retreat. The vertical dashed line is March 15. 3245 3246 Figure 6. Scatterplot of the observed (number) and fitted (line, based on simple linear regression, y- 3247 intercept = 53, slope = 0.66, df = 1, 24, p < 0.001) values of spring bloom maximum (day) versus ice 3248 retreat (day) for all moorings when ice was present after March 15. The number indicates mooring 3249 number. 125 3250 3251 Figure 7. A boxplot of fall overturn (day) by mooring. The box extends from the first quartile to 3252 the third quartile, the heavy line dividing the box is the median and the whiskers are the smallest 3253 and largest values. The numbers of years when fall overturn could be determined were 15 (M2), 3254 10 (M4), 16 (M5) and 7 (M8). 3255 3256 Figure 8. Scatterplots of maximum fall bloom strength (ug Chl l-1), fall bloom timing (day), 3257 maximum summer-fall temperature (ºC), fall overturn (day) and first storm (day). 3258 3259 Figure 9. Scatterplots of fall bloom maximum (day) versus fall overturn (day) by mooring. The 3260 diagonal dashed line is the 1:1 line to compare timings of fall bloom maximum and fall overturn. 3261 The horizontal dashed line is August 15. 3262 3263 Figure 10. Scatterplot of maximum spring bloom strength (ug Chl l-1) and maximum fall bloom 3264 strength. The number indicates mooring number. 3265 3266 Figure 11. Scatterplot of observed (number) and fitted (line, based on linear regression, intercept 3267 = 294, slope = -1.16, df = 1, 27, p < 0.001) values for all moorings. The y=axis is the interval 3268 between the spring and fall bloom maximum (days). The x-axis is the spring bloom maximum 3269 (day). The number indicates mooring number. 3270 3271 Figure 12. Timing of Calanus reproduction relative to the presence of ice and ice algae, and the 3272 spring and fall blooms. The Calanus life history is simplified and the timings are approximate. 3273 Black is the period of ice cover; gray is the 50th percentiles for the timing of ice retreat. White 3274 rectangles are the 50th percentile dates for bloom maximum in the spring and fall. Ice algae 3275 blooms during ice cover (solid gray line) and ice retreat (dashed gray line) and begin as early as 3276 mid-February (R. Gradinger, personal comm.). A – a cold icy year, with ice retreat shortly after 3277 March 15; B – a cold icy year with ice retreat after April 14; C - no ice after March 15. 126 3278 Appendix Table 1. Annual values compiled for this manuscript; methods for compiling these 3279 values are described in the methods section. In all cases “Day” refers to Julian day. Day ice 3280 retreat: the day that percent ice cover fell below 15% (a zero value indicates that ice cover never 3281 fell below 15%); Day temp >-1 ºC: the day that ocean temperature rose above -1 ºC (a zero value 3282 indicates that ocean temperature never fell below -1 ºC); Min WS temp: Minimum temperature 3283 during winter and spring; Day max spring bloom: Day that the chlorophyll a (µg l-1) values 3284 reached their maximum (NA indicates missing data); Max spring bloom: Maximum chlorophyll a 3285 value during winter and spring; Max fall temp: Maximum temperature during summer and fall; 3286 Day first storm: first day a storm occurred during summer and fall as defined in the methods; Day 3287 overturn: first day that temperature drop indicated overturn was underway; Max SF temp: 3288 Maximum temperature during summer and fall; Day max fall bloom: Day that the chlorophyll a 3289 values reached their maximum; Max fall bloom: Maximum chlorophyll a value during summer 3290 and fall. 127 Moor Year M2 1995 M2 1996 M2 1997 M2 1998 M2 1999 M2 2000 M2 2001 M2 2002 M2 2003 M2 2004 M2 2005 M2 2006 M2 2007 M2 2008 M2 2009 M2 2010 M2 2011 M4 1995 M4 1996 M4 1997 M4 1998 M4 1999 M4 2000 M4 2001 M4 2002 M4 2003 M4 2004 M4 2005 Day ice retreat 104 0 99 60 129 35 0 53 0 0 0 36 98 110 119 103 114 112 44 100 72 128 40 0 60 87 101 0 Day temp >-1C 110 0 92 56 134 58 0 0 0 0 0 36 111 128 125 127 120 NA NA 120 NA NA NA 0 65 87 100 0 Min WS temp -1.60 -0.90 -1.56 -1.68 -1.73 -1.20 1.67 -0.14 1.63 1.31 2.12 -1.61 -1.67 -1.69 -1.73 -1.38 -1.38 NA NA -1.69 NA NA NA 0.86 -1.68 -1.24 -1.28 1.34 Day max Max spring spring bloom bloom NA NA NA NA 106 39.6 146 24.5 113 94.8 132 4.2 144 26.7 153 13.1 138 22.4 143 23.8 132 30.8 165 28.1 115 9.7 122 20.0 155 9.0 126 16.1 158 18.5 NA NA NA NA 127 27.2 NA NA 124 6.0 166 12.5 149 21.9 155 22.0 94 76.2 144 17.6 NA NA Max fall temp 10.5 9.6 13.6 11.8 9.7 10.3 11.6 12.5 12.1 13.6 13.0 10.4 12.2 9.8 10.1 8.4 10.4 NA 7.6 NA NA 8.6 10.3 10.3 11.4 11.2 11.7 12.0 Day first Day storm overturn 295 NA 245 242 245 277 235 228 295 263 285 240 295 246 265 247 295 262 275 265 255 235 285 262 255 268 325 284 315 NA 265 251 265 263 325 NA 245 NA 245 NA 235 NA 215 259 245 242 305 267 295 268 275 274 285 266 255 239 Max SF temp 239 207 220 217 195 195 229 242 251 224 220 235 246 254 237 213 231 NA 266 NA NA 200 232 232 231 258 241 218 Day max Max fall fall bloom bloom NA NA 306 6.9 293 16.6 275 9.1 238 22.6 248 7.6 NA NA NA NA 294 59.3 271 19.8 261 15.5 268 4.6 NA NA 300 3.2 NA NA 301 4.6 261 6.3 NA NA NA NA NA NA NA NA 249 8.4 263 5.4 272 14.4 288 59.6 274 9.7 286 8.3 NA NA 128 M4 M4 M4 M4 M4 M4 2006 2007 2008 2009 2010 2011 135 100 135 117 138 121 120 123 127 125 NA 124 -1.70 -1.70 -1.74 -1.03 NA -1.62 134 104 129 145 NA 129 32.1 10.6 52.3 2.9 NA 2.8 9.9 10.8 9.6 9.7 8.1 NA 255 255 285 315 265 265 258 254 279 223 NA NA 242 223 236 230 268 NA 270 298 282 NA 291 NA 24.6 9.1 5.4 0.1 8.9 NA 129 Moor Year M5 1995 M5 1996 M5 1997 M5 1998 M5 1999 M5 2000 M5 2001 M5 2002 M5 2003 M5 2004 M5 2005 M5 2006 M5 2007 M5 2008 M5 2009 M5 2010 M5 2011 M8 1995 M8 1996 M8 1997 M8 1998 M8 1999 M8 2000 M8 2001 M8 2002 M8 2003 M8 2004 M8 2005 M8 2006 Day ice retreat 132 120 129 124 156 127 93 109 102 108 128 153 151 166 137 136 136 127 131 138 142 158 134 141 136 117 128 122 144 Day temp >-1C NA NA NA NA NA NA NA NA NA NA 130 177 166 158 160 139 141 NA NA NA NA NA NA NA NA NA NA 131 176 Min WS temp NA NA NA NA NA NA NA NA NA NA -1.43 -1.74 -1.74 -1.75 -1.74 -1.71 -1.71 NA NA NA NA NA NA NA NA NA NA -1.76 -1.79 Day max Max spring spring bloom bloom NA NA NA NA NA NA NA NA NA NA NA NA NA NA NA NA NA NA NA NA NA NA 162 6.0 141 14.5 156 35.3 NA NA 146 6.3 144 2.4 NA NA NA NA NA NA NA NA NA NA NA NA NA NA NA NA NA NA NA NA 133 17.0 144 10.3 Max fall temp NA NA NA NA NA NA NA NA NA NA 11.3 9.3 11.0 9.3 9.4 9.5 7.7 NA NA NA NA NA NA NA NA NA 11.2 10.0 9.1 Day first Day storm overturn 255 NA 245 NA 265 NA 315 NA 295 NA 315 NA 295 NA 285 NA 275 NA 285 NA 255 NA 255 NA 265 NA 325 NA 335 NA 265 NA 215 NA 255 NA 245 NA 265 NA 295 NA 265 NA 285 NA 275 NA 295 NA 285 NA 285 NA 315 NA 335 NA Max SF temp NA NA NA NA NA NA NA NA NA NA 217 242 223 247 232 223 215 NA NA NA NA NA NA NA NA NA 225 239 241 Day max Max fall fall bloom bloom NA NA NA NA NA NA NA NA NA NA NA NA NA NA NA NA NA NA NA NA 281 6.4 298 10.7 279 14.5 246 14.8 NA NA 218 10.0 223 4.1 NA NA NA NA NA NA NA NA NA NA NA NA NA NA NA NA NA NA 290 2.8 259 4.8 315 10.3 130 M8 M8 M8 M8 M8 2007 2008 2009 2010 2011 144 152 155 135 141 186 187 185 185 217 -1.76 -1.80 -1.78 -1.77 -1.76 161 160 166 NA NA 35.4 49.6 49.3 NA NA 9.9 8.6 7.9 9.7 1.8 295 275 265 305 265 NA NA NA NA NA 260 257 249 254 NA 244 298 NA NA NA 15.3 5.7 NA NA NA 131 3282 3283 Appendix Figure 1. Plots of chlorophyll a fluorescence by mooring, year and Julian day. The x- 3284 axis is Julian day and the y-axis is chlorophyll a (µg l-1). Each plot is labeled with mooring and 3285 year. The open symbols indicate the assigned spring (circle) and fall (triangle) blooms. 3286 132 3287 133 3288 134 3289 135 3290 136 3291 137 3292 138 3293 139 3294 3295 3296 140 3297 Appendix Figure 2. Maps of Pearson's product moment correlations between 8-day averages of 3298 log-transformed fluorescence at four mooring sites and 8-day averages of log-transformed 3299 chlorophyll a as estimated from satellite data (SeaWiFS 1998-2002, Modis Aqua 2003-2011). 3300 Black squares denote reference mooring locations for each map. Only correlations individually 3301 significant at approximately 95% are shown. 3302 3303 141 3304 Chapter 15 3305 3306 Sullivan, M., C. Mordy, S. Salo, and P.J. Stabeno (2013): The influence of sea ice on seasonal 3307 water column changes on the eastern Bering Sea shelf. Deep-Sea Res. II. 3308 Sea ice and water column structure on the eastern Bering Sea shelf Margaret E. Sullivanab*, Nancy B. Kachel ab, Calvin W. Mordyab. Sigrid A. Salob, and Phyllis J. Stabenob a Joint Institute for the Study of the Atmosphere and Ocean, University of Washington, 3737 Brooklyn Ave NE, Box 355672, Seattle, WA 98105-5672 USA b Pacific Marine Environmental Laboratory, NOAA, 7600 Sand Point Way NE, Seattle, WA 98115 USA *Corresponding Author 7600 Sand Point Way NE, Seattle, WA 98115 USA msulliva@u.washington.edu, Phone: (206) 526-6185, Fax: (206) 526-6485 Revised: September 5, 2013 For submission to Deep-Sea Research Part II: Topical Studies in Oceanography 3309 142 3310 Abstract 3311 Seasonal sea ice is a defining characteristic of the eastern Bering Sea shelf, and plays a critical 3312 role in determining the vertical structure of temperature and salinity over this shelf. This paper 3313 examines ice movement relative to local winds, and the impact of the arrival and retreat of ice on 3314 the water column at four mooring sites on the middle shelf. Ice forms primarily in coastal regions 3315 and is advected over the southern and outer shelves. Ice drift was ~2% of local wind speed and 3316 oriented 44 ° to the right of the winds (R-squared = 0.25). Measurements from 30 ice cores 3317 collected in 2006 -2009 gave an average salinity of ~6, and an average nitrate concentration of ~1 3318 µM. Time series data collected at the biophysical moorings in the Bering Sea (1995 – 2012) were 3319 used to explore the evolution of stratification under the ice. At mooring M8 in the north, the 3320 water column mixed and cooled prior to the arrival of ice, hence, little melt occurred when the ice 3321 arrived. At the other three moorings, the ocean temperature was 2 – 4 °C above freezing, resulting 3322 in extensive ice melt. The melting ice freshened and cooled the upper part of the water 3323 column. The water column remained stratified for 10 to 25 days. Mixing was a slower process 3324 under the ice than in open water. An estimated 1.3 m of ice melted with the first arrival of ice at 3325 three southern moorings and the latent heat of cooling of this ice accounted for approximately 3326 half the observed cooling. During ice retreat there appeared to be little ice melt around the 3327 southern two moorings, but an estimated 0.8 m at M5 and M8. The extent of ice melt sets up the 3328 water column for the following summer. 3329 3330 Keywords: Bering Sea, sea ice, ocean temperature, salinity, currents, stratification, ice melt 3331 3332 1. Introduction 3333 Situated between the North Pacific and the Arctic Ocean, the eastern Bering Sea is a 3334 marginal sea ice zone. During summer it is ice free, with ice beginning to form along the coast in 3335 the northern Bering Sea as early as November (Pease, 1980). During subsequent months, the sea 3336 ice is advected southward with maximum ice extent typically occurring in March or April 3337 (Stabeno et al., 2012b). In extensive ice years, sea ice can advance more than 1000 km, which, 3338 before the extensive melting now occurring in the Arctic Ocean, was the largest ice advance in 3339 any arctic or subarctic region (Niebauer, 1998). Sea ice typically begins retreat in late winter or 3340 early spring, and by late June the Bering Sea is ice free. 3341 Pease (1980) describes the expansion of ice into the southern Bering Sea as a conveyor belt 3342 in which ice forms in polynyas in the northern Bering Sea and is advected southward by winds. 3343 Polynyas are most common off southern coastlines due to prevailing northerly winds in winter, 143 3344 but they also can form when winds are southerly off northern and western coastlines (Niebauer 3345 and Schell, 1993, Niebauer et al., 1999). The St. Lawrence polynya occurs along the southern 3346 coast of St. Lawrence Island, and is a dominant ice-producing feature (Schumacher et al., 1983; 3347 Stringer and Groves, 1991; Danielson et al., 2006; Drucker et al., 2003), with ice being advected 3348 mostly south and southwestward (McNutt, 1981). The leading edge of the ice melts as it 3349 encounters warmer ocean temperatures in the south or along the shelf break (McNutt, 1981; 3350 Zhang et al., 2010). 3351 The eastern Bering Sea continental shelf is broad (>500 km), less than 180 meters deep, 3352 and extends more than 1000 km south from Bering Strait to the Aleutian Islands. The shelf is 3353 divided into inner (water depth < 50 m), middle (~50-100 m), and outer (~100-180 m) domains 3354 (Coachman, 1986), which are separated by transition zones or fronts (Schumacher, et al., 1979; 3355 Stabeno, et al., 2001; Kachel, et al., 2002). Ice typically appears first in the coastal domain, and 3356 latest in the outer domain. On average, sea ice covers the middle shelf north of 58 °N for five 3357 months each year, with ice persisting, on average, for less than one month on the southern shelf 3358 (south of 57 °N) (Stabeno et al., 2012a). 3359 These north-south differences in timing of ice retreat and advance, together with a north 3360 (weaker) – south (stronger) gradient in tidal energy, result in distinct differences in the water 3361 column structure on the northern and southern shelves during the summer (Stabeno et al., 2012a). 3362 The northern shelf with its more extensive and persistent ice cover remains cold until solar input 3363 warms the ocean, initiates ice melt and sets up summer stratification. This results in a warmer, 3364 fresher surface layer overlaying a colder, saltier bottom layer separated by a ~10 m interface. The 3365 cold bottom layer (< 2 °C) over the middle shelf is known as the cold pool, and remains largely 3366 well mixed due to the tides. Ice retreat in the south typically occurs in March or April with 3367 southerly winds, when winds are usually still strong enough to mix the water column (Stabeno et 3368 al, 2010). The water column becomes thermally stratified when winds weaken and solar 3369 insolation increases (Stabeno et al, 2007). In years with extensive ice in March or April, the 3370 bottom temperatures in ice-covered areas in the south are < 2 °C, and part of the cold pool 3371 (Wyllie-Echeverria, et al., 1998; Stabeno et al., 2012b). During years when the sea ice does not 3372 extend onto the southern shelf, the bottom temperatures remain above 2 °C, and the cold pool 3373 exists only over the northern shelf (Stabeno et al., 2012a,b). 3374 The presence or absence of sea ice impacts spatial and temporal variability of biological 3375 production across the shelf and slope (Stabeno et al., 2012a,b; Sigler, et al., this volume). 3376 Constituents of sea ice, including iron and phytoplankton, are advected southward, and influence 3377 the amount and fate of spring production (Aguilar-Islas et al., 2008; Schandelmeier and 144 3378 Alexander, 1981). In addition, sea ice impacts the timing of the spring phytoplankton bloom, 3379 with a bloom occurring under the ice if ice is present after mid-March, and an open-water bloom 3380 occurring in May or June if no ice is present after mid-March (Brown and Arrigo, 2013; Stabeno 3381 et al., 2012a; Sigler et al., this volume). Vertical stratification of the water column is important to 3382 3383 3384 3385 3386 3387 Figure 12. a) A MODIS True-Color image is combined with MODIS sea surface temperature in the eastern Bering Sea for March 11 - 13, 2007. The large, red letters indicate the following: A polynyas; B heavy ice; and C cloud cover. b) A shelf map showing distribution of ice sampling sites per year (2006-2009), and the bathymetry. The four mooring locations (M2, M4, M5, M8) and geographic names are indicated in both panels. 3388 Support under-ice phytoplankton blooms (Alexander and Niebauer, 1981; Mundy et al., 2009). 3389 The southern extent of the cold pool and the timing of spring primary production influence the 3390 food chain and higher trophic level predator-prey relationships for the following 6 - 8 months 3391 (Sigler et al., this volume), and over longer time scales (Mueter and Litzow, 2008). 3392 The purpose of this paper is to examine the impact of sea ice on the vertical structure of the 3393 water column. Measurements from satellites, ice cores and four long-term biophysical moorings 3394 distributed along the 70-m isobath from 56.9 °N to 62 °N were used to determine the rate and 3395 direction of ice movement, the arrival of ice at moorings, and the extent of ice melt and resulting 3396 stratification at each of the mooring sites (Fig. 1). First, the rate and direction of ice movement in 145 3397 the region were calculated using satellite images. Next, the characteristics of 30 ice cores 3398 collected from 2006 – 2009 are presented. Using time series of temperature, salinity and currents 3399 from the biophysical moorings (M2, M4, M5 and M8; Fig. 1), the impact of sea ice on the water 3400 column structure is examined. Using selected time series from the ~42 yearly records, the 3401 thickness of the ice melted at the mooring sites is estimated. 3402 3403 2. Methods 3404 2.1. Satellite Data 3405 Satellite measurements of ocean color and sea surface temperature (SST) were obtained 3406 using MODIS (Moderate Resolution Imaging Spectroradiometer) data files from the ocean color 3407 website at NASA (http://oceancolor.gsfc.nasa.gov, assessed 2012). SeaDAS, a suite of programs 3408 created and maintained by NASA, was used to map true-color and SST images. Individual floes 3409 were identified in a series of true-color images and tracked from one image to another, until they 3410 were no longer recognizable (Holt, et al, 1992, Leberl, et al, 1983). Ocean color and SST 3411 observations cannot be made through clouds, severely limiting the number of useful MODIS 3412 images from the Bering Sea. In addition, increased cloud cover is associated with southerly 3413 winds, so the set of floes that were tracked were biased against times when southerly winds were 3414 prevalent. This limited the analysis to years with a sufficient number of clear days to follow a floe 3415 over a period of time, and to larger floes in regions where horizontal shear was weak enough not 3416 to deform the floes. 3417 For 2002–2011, ice concentration data from the AMSR_E (Advanced Microwave Scanning 3418 Radiometer-Earth Observing System) sensor aboard the MODIS Aqua satellite were downloaded 3419 from the National Snow and Ice Data Center website (http://nsidc.org/data/amsre/, assessed 2012; 3420 Cavalieri et al., 2003). Ice concentrations from the NASA Special Sensor Microwave Imager 3421 (http://nsidc.org/data/docs/daac/ssmi_instrument.gd.html; Maslanik and Stroeve, 1999) were used 3422 for fall 1994–2002 to extend the time span prior to the start date of AMSR-E data. Daily-mean ice 3423 concentration values were used in this paper. The data were used to calculate mean daily ice 3424 concentrations within boxes centered on the four moorings (M2, M4, M5, and M8 in Fig. 1). The 3425 grid size of the ice data is approximately 12.5 km × 12.5 km. Following Stabeno et al. (2012a) we 3426 used 100 km × 100 km boxes. 3427 3428 2.2 Wind stress and winds 146 3429 National Center for Environmental Prediction (NCEP) – Department of Energy Reanalysis 3430 2 uses a state-of-the-art analysis/forecast system to perform data assimilation with data ranging 3431 from January 1979 to August 2012 (http://www.cpc.ncep.noaa.gov/products/wesley/reanalysis2/; 3432 Kanamitsu et al., 2002), and is an update to the NCEP/NCAR (National Center for Atmospheric 3433 Research) reanalysis. Six-hourly wind and wind-stress data were extracted from the NCEP 3434 Reanalysis 2 at the grid points nearest to the four mooring locations. NCEP Reanalysis 2 data 3435 were obtained from the website maintained by NOAA Earth System Research Laboratory, 3436 Physical Sciences Division in Boulder, Colorado, USA (http://www.esrl.noaa.gov/psd/). NCEP 3437 winds are well correlated with the observed winds in the Bering Sea (Ladd and Bond, 2002). 3438 QuikSCAT wind data were downloaded from the Jet Propulsion Laboratory, Physical 3439 Oceanography Distributed Active Archive Center (http://podaac.jpl.nasa.gov). Scatterometer 3440 winds are derived from ripple patterns on the surface of the ocean, and are therefore limited to 3441 ice-free areas. Although the instrument sees through clouds, measurements are affected by heavy 3442 rain. The records identify pixels where rain was suspected, and these records were excluded from 3443 our analysis. Grid spacing for QuikSCAT winds is 12.5 km (0.25 ° longitude), but the data were 3444 binned into 2° bins for regional plots. 3445 3446 3447 2.3. Ice core measurements During four springs (2006-2009), a total of 30 ice cores were collected by NOAA scientists 3448 from floes distributed over the eastern Bering Sea shelf (Fig. 1b). In 2006, three floes were 3449 sampled on the R/V Thomas G. Thompson (Cruise TN193). The Thompson had limited 3450 maneuverability in the ice, so sampling was restricted to floes at the ice edge. Cruises in 2007- 3451 2009 were conducted on the icebreaker, U.S. Coast Guard Cutter Healy as part of the Bering Sea 3452 Project (http://bsierp.nprb.org/). The Healy is capable of breaking 1.4 meters of ice at 3 knots 3453 continuously, and 2.4 meters by backing and ramming. This permitted sampling deep into the 3454 pack-ice in water depths between 30 and 100 meters. 3455 Floes were chosen for accessibility and for ease of data collection. Sampling was biased 3456 against ice that was too thin to walk on or highly rafted, and to times when air temperature was 3457 above -15 °C. All samples were taken in spring when both melting and refreezing were prevalent. 3458 In 2006, the three ice stations were located along the ice edge in the outer domain, while the 3459 seven stations in 2007, the 11 in 2008, and the nine in 2009 were mostly located deeper within the 3460 ice field in the middle domain north of 58 °N (Fig. 1b; Table 1). 3461 3462 At most sites, air temperature, freeboard, ice thickness, and snow cover were recorded; coordinates were recorded at the beginning and end of each site visit using a GPS unit. All cores 147 3463 were obtained using a Kovacs4 Mark-II, 9-cm diameter, 1.15-meter-long ice corer driven by an 3464 attachable gas engine. Cores were positioned on a black plastic surface, then measured and 3465 photographed. Top and bottom conditions and banding were noted. Small holes were drilled and 3466 temperatures were recorded every 10 cm along the core using either an Oakton Acorn or an 3467 Omega thermometer. All cores were sliced into 10-cm sections and double-bagged for later 3468 analysis. On the ship, sections from each core were thawed in the dark. Volume of the melt water 3469 was measured and then sub-sampled to measure salinity, and nutrient concentrations. Salinities 3470 were measured at sea using a salinometer referenced against IAPSO standard water. Ice density 3471 was determined from the dimensions of each ice core, and the volume and density of the ice melt. 3472 Samples for nutrient analysis were filtered using syringe with 0.45 μm cellulose acetate 3473 membranes, and collected in 30 ml acid washed, high-density polyethelene bottles after being 3474 rinsed three times with sample water. Samples were analyzed shipboard within 1 - 12 hrs of 3475 collection. Nitrate and nitrite concentrations were determined using a combination of analytical 3476 components from Alpkem, Perstorp and Technicon. WOCE-JGOFS standardization and analysis 3477 procedures specified by Gordon et al. (1994) were followed, including reagent preparation, 3478 calibration of labware, preparation of primary and secondary standards, and corrections for blanks 3479 and refractive index. 3480 For determination of chlorophyll and phaeopigment content a second core was taken at the 3481 sites, and sections of ice were melted in ~1.5 times the ice volume in filtered seawater to prevent 3482 cell lysis. Sub-samples (~50 ml) were filtered though Whatman GF/C filters, and the filters were 3483 frozen at -80 °C, remaining frozen until analysis several months later. Pigments were extracted in 3484 cold 90% acetone in the dark for 24 hrs, and determined using a calibrated fluorometer according 3485 to Strickland and Parsons (1968). 3486 3487 2.4. Moorings 3488 Four biophysical mooring sites (M2, M4, M5, and M8) are the cardinal locations of the 3489 Bering Sea biophysical observing network (Fig. 1). When possible, the moorings are recovered 3490 and redeployed twice a year, once in the spring (April/May) and again in the late summer or early 3491 fall (September/October). During years with extensive ice, M8 and sometimes M5 were only 3492 recovered/deployed in September. Moorings at the M2 site have been maintained almost 3493 continually since 1995, while moorings have been deployed at M4 since 1996 (continuously since 3494 2000), at M5 since 2005 and at M8 since 2004. At the writing of this manuscript, these moorings 3495 provided 42 fall-through-spring sets of time series describing water column conditions. 4 Use of trade names does not constitute an endorsement by NOAA 148 3496 Data collected by instruments on the moorings included temperature (miniature 3497 temperature recorders, SeaBird SBE-37 and SBE-39) and salinity (SBE-37). Currents were 3498 measured using an upward-looking, bottom-mounted, 300 or 600 kHz Teledyne RD Instruments 3499 acoustic Doppler current profiler (ADCP) deployed within 500 m of the main mooring. All 3500 instruments were calibrated prior to deployment, and the data were processed according to 3501 manufacturers’ specifications. The main mooring “line” at each site was chain and the 3502 instruments were mounted in-line in metal frames. Such construction is necessary to reduce the 3503 chances of losing the mooring due to sea ice and heavy fishing pressure in the region. 3504 Sampling intervals varied among the different instruments and ranged from 10 to 60 3505 minutes. Small data gaps (< 1 day) were filled using 3506 linear interpolation or spectral methods. Unless 3507 otherwise noted, data presented here have been filtered 3508 using a 35-hour, cosine squared, tapered Lanczos filter to 3509 remove semidiurnal and diurnal tidal signals, and re- 3510 sampled to 6-hour intervals. 3511 The top instrument at each mooring was at a depth 3512 between 1 and 20 m. When the mixed layer was deeper 3513 than the depth of the top instrument, the data from the 3514 top instrument could be reliably extrapolated to the 3515 surface (Stabeno et al., 2007). However, when the top 3516 instrument was below ~20 m and the ice had retreated 3517 from the mooring in the spring, the mixed layer was 3518 usually shallower than the top instrument and there was 3519 no reliable way of estimating localized near-surface 3520 temperatures. 3521 3522 2.5. Calculation of volume of ice melt 3523 Estimates of magnitude of ice melt can be made 3524 at the mooring sites from the change in salinity in the 3525 water column using the following equation: 3526 hw Si + (H-hw) Sw = H Sf 3527 or (1) Figure 13. Change in ice concentration (color filled) associated with 2-day mean QuikSCAT winds (vectors) are shown for three time periods: a) February 21 - 23, 2003; b) March 2 3, 2003; and c) March 12 - 13, 2003. 149 3528 hw = H (Sw - Sf) / (Sw – Si) (2) 3529 where hw is the thickness of water resulting from ice melt, Si the salinity of the ice, H the depth of 3530 the upper mixed layer (which may be the whole water column), Sw the initial salinity of the water 3531 column and Sf the final salinity of the water column. This is an approximation, since an 3532 introduced piece of ice does not melt completely, but rather the ice is advected over the water, 3533 with only a portion of it melting at that location. When the water reaches the freezing point, there 3534 is no further net melting of the ice. 3535 The thickness of the ice melted, hi, was calculated using hi = hw 3536 i w i w is the is the density of ice. Using the value of hw calculated from salinity, the 3537 change in temperature of the water column per unit area due to latent heat can be calculated using 3538 the following equation 3539 3540 i hi (3) 3541 where L is the latent heat of fusion of sea ice and T is 3542 temperature. 3543 3544 3. Results and Discussion 3545 3.1. Satellite floe-tracking and winds 3546 Ice in the Bering Sea can be broadly 3547 characterized using true-color satellite imagery from 3548 which polynyas (dark shadows leeward of the 3549 coastline), thin ice (gray, often seaward of the open 3550 water of the polynya), and thicker ice (white) can be 3551 identified (Fig. 1a). Ice conditions, including the 3552 location of polynyas and the direction and extent of 3553 ice advance, can change rapidly and are largely forced 3554 by winds (Fig. 2). For example, in late February 2003, 3555 northeasterly winds near Siberia and easterly winds 3556 off the Alaskan coast reduced the ice concentration 3557 near Siberia, St. Lawrence Island, and the Alaskan 3558 coast (Fig. 2a). Nine days later, southerly winds over 3559 the eastern shelf resulted in ice edge retreat in the east Figure 14. Ice floe trajectories tracked using MODIS True Color images are shown for two years: a) February 19 – April 15, 2003; and b) March 1 – April 20, 2007. The colors indicate the dates of observations. See color bar at right of plots for dates. Mooring locations are indicated. 150 3560 and compression of the ice along those coastlines (Fig. 2b). In mid-March, northerly winds 3561 opened polynyas in the north, advanced the ice edge, and increased the ice concentration at the 3562 ice edge (Fig. 2c). 3563 Floe trajectories obtained from true-color satellite images are shown for 2003 and 2007 3564 (Fig. 3), a warm and a cold year, respectively (Stabeno et al., 2012b). These trajectories are 3565 primarily from advancing ice. While many floes could only be tracked for a few days, several 3566 were tracked for 1–2 weeks while they moved southward more than 100 km. The maps from both 3567 years show a corridor in which ice floes moved from the shelf north and east of St. Lawrence 3568 Island through Shpanberg Strait, with more westward ice motion in 2003 than 2007 as floes 3569 moved beyond Shpanberg Strait. These observations are similar to the conveyor belt corridor 3570 observations in Pease (1980). With southerly winds, ice produced from a polynya north of 3571 Nunivak Island could also contribute to the conveyor belt corridor (Figs. 2 and 3a). South of the 3572 Chukotka Peninsula, floes moved eastward with prevailing currents (Danielson, 2000), and those 3573 nearest the coast continued north through Anadyr Strait. A few floes originating southwest of St. 3574 Lawrence were tracked to M8 (Fig. 3). 3575 These floe trajectories were used to infer the origin of ice in each of the four mooring areas 3576 (Figs. 1 and 3). The origin of the ice at M8 was a combination of ice from the conveyor belt and 3577 from the polynya south of St. Lawrence Island (McNutt, 1981; Schumacher et al., 1983), and 3578 occasionally Anadyr Strait (Schumacher, et al., 1983). M4 and M5 locations were directly 3579 influenced by ice from the conveyor belt corridor (Muench and Ahlnas,1976; Pease, 1980). The 3580 area around M2 was outside the dominant influence of the conveyor belt corridor, and subject to 3581 ice formation from polynyas south of Nunivak Island and along the northern Bristol Bay coast 3582 during northerly or northeasterly 3583 winds. 3584 Tracked floes, which had 3585 typical speeds of 0.1 to 0.3 m s-1, 3586 were influenced by both winds and 3587 currents. These findings are 3588 comparable to earlier observations 3589 (Shapiro and Burns, 1975; Muench 3590 and Ahlnas, 1976; McNutt, 1981; 3591 Weeks and Weller, 1984), as well as 3592 results from a recent ice model 3593 (Zhang et al, 2010), but slower than Figure 15. Wind vector plots, from 6-hourly NCEP-2 Reanalysis data centered at mooring site M4, for the cold seasons of a) 2005 – 2006 and b) 2006 – 2007. Northward is upward. Ice extent in the 100-km x 100-km around M4 is included as a blue line. 151 3594 the 0.5 m s-1 average reported by Pease (1980) and the 0.45 m s-1 average reported by Macklin et 3595 al. (1984). Ice drift was compared to NCEP2 winds interpolated in space and time, and found to 3596 be ~2% of local wind speed (n = 440), and oriented 44 ° to the right of the wind direction with an 3597 R-squared value of 0.25, which is higher than the findings of Macklin et al. (1984). 3598 While wind largely controls the movement of sea ice (Figs. 2, 3), the timing of storms is 3599 also important. The influence of wind events on ice formation varied dependent upon the season. 3600 Northerly winds during active freeze-up (January into March) created more ice than similar winds 3601 would have generated during warming (mid April to May). Nonetheless, northerly winds later in 3602 the season could prolong ice cover by continuing to transport ice southward. For example, in the 3603 winter of 2005-2006, northerly winds in December and January (Fig. 4a) pushed the ice to its 3604 most southern extent in late January. This was in spite of the fact that water column was ~2.8 °C. 3605 The next episode of sustained northerly winds did not occur until mid-April (initial water 3606 temperature of 0.5 °C), and although the ice advanced at this time, the ice edge did not regain its 3607 earlier position, because the increase solar insolation. At the beginning of April the water column 3608 warmed from -1 °C to >0 °C. In contrast, during the winter of 2006 – 2007, there were several 3609 storms in December and January (Fig. 4b), but the longest period of northerly winds was during 3610 March and April, and sea ice reached its maximum extent in late March (Rodionov et al., 2007). 3611 The water column cooled in early March (from 1.1 C to -1.7 °C) and did not begin warming until 3612 the ice began retreating. 3613 3614 3.2. Sea-ice cores 3615 Thirty ice stations were 3616 sampled on the eastern Bering Sea 3617 shelf in April and early May on four 3618 cruises between 2006 and 2009 (Fig. 3619 1b). Ice thickness ranged from 37 to 3620 127 cm with a single outlier of >329 3621 cm of rafted ice at an inner domain 3622 station (Table 1). Mean ice thickness 3623 was calculated excluding this outlier. 3624 Surface water from nearby 3625 hydrographic casts had mean salinity of 31.9, mean water temperature of -1.7 °C, and mean Figure 16. a) Vertical profiles of temperature, salinity and nitrate from an ice core collected at Ice Station 2 in 2008 (Table 1). b) Ice core photograph from same station. c) Selected vertical profiles of salinity from 2009 ice cores. 3626 alinities in the ice were low (mean ~6, Table 1), reflecting brine rejection 3627 during ice formation and brine drainage. There was considerable variability in the shapes of the 152 3628 salinity profiles (Fig. 5c). Some salinity profiles showed the well-defined C-shape (e.g., green 3629 line, Fig. 5c) of marginal ice zones and young ice (Malmgren, 1927, Eicken, 2003). Another 3630 common pattern was a double C-shape (e.g., red line, Fig. 5c), often a result of rafting. Mean ice 3631 density was 0.89 ± 0.03 x 103 kg m-3, which is in agreement with the Table 1 summary in Timco 3632 and Frederking (1996). Nutrient concentrations were low in the ice cores (mean nitrate ~1 µM), 3633 suggesting that ice was not a conveyor of macro nutrients in the Bering Sea. Melting sea ice, 3634 however, does provide a significant contribution of iron to the water column (Aguilar-Islas et al., 3635 2008). 3636 Horizontal banding was evident in the ice cores and often, dark coloration occurred at the 3637 ice-water interface (Fig. 5). Some banding was correlated with elevated chlorophyll-a and 3638 phaeopigments, while in other cores the banding appeared to be sediment. Highest chlorophyll 3639 concentrations were most commonly found in the bottom section of each ice core. For example, 3640 in 2009, bottom chlorophyll was as high as 209 µg l-1, with an average of ~80 µg l-1 which was 3641 approximately 20 times greater than the average concentration (4 µg l-1) found in the rest of the 3642 core slices. Mean snow depth on floes was variable, but generally less than 10 cm (Table 1). 3643 3644 3645 3.3. Influence of sea ice on the water column Typically, the depth-averaged water column temperature reaches a maximum in September 3646 (Stabeno et al., 2012a, b), when the mixed layer begins to deepen. In examining the 18 years of 3647 data at M2, the depth-averaged temperature is almost as warm in August as in September, and 3648 usually the water column has not mixed. Temperature data from August are used to represent 3649 maximum summer near-surface temperature. During August, the mixed layer is usually 20-30 m 3650 deep. The highest mean August near-surface temperature was 12.5 °C (M2, 2002, a warm year) 3651 and the lowest monthly mean bottom temperature was -1.7 °C (M8, 2009, a cold year) (Table 2). 3652 The differences between monthly mean surface and bottom temperatures in August range from 3653 5.2 to 10.8 °C. 3654 3655 153 3656 3657 3658 3659 3660 3661 3662 3663 3664 3665 3666 3667 3668 3669 3670 3671 3672 3673 3674 3675 3676 3677 Figure 6. Time series from M2 showing a) wind stress, b) temperature at 12 depths distributed through the water column, and c) salinity at four depths. The breaks in the time series (b and c) occurred when the winter mooring was replace by the summer mooring. The gray shading indicates times when the water column was well mixed. There was no ice cover at M2 during this period. 3678 3679 3680 154 3681 The time at which the water 3682 column became well mixed varied 3683 from north to south (Table 2), with M8 3684 remaining stratified the longest of the 3685 four mooring areas. At M8 during 3686 summer, both temperature and salinity 3687 contribute equally to stratification, 3688 while at the southern moorings 3689 stratification is controlled by 3690 temperature. Since the vertical 3691 differences in temperature are similar 3692 north-to-south, the stratification is 3693 almost twice as strong at M8 than at 3694 the other mooring sites (Stabeno et al., 3695 2010, 2012a; Ladd and Stabeno, 2012), 3696 thus delaying mixing. 3697 While there is considerable Figure 6. Time series from M2 showing a) wind stress, b) temperature at 12 depths distributed through the water column, and c) salinity at four depths. The breaks in the time series (b and c) occurred when the winter mooring was replace by the summer mooring. The gray shading indicates times when the water column was well mixed. There was no ice cover at M2 during this period. 3698 information on the timing and 3699 characteristics of ice retreat and melting (Alexander and Niebauer, 1981; Hunt and Stabeno, 3700 2002; Stabeno et al., 1998; Stabeno et al., 2012b), less is known about the short-term response of 3701 the water column to melting sea ice. Instruments at each of the four biophysical moorings in the 3702 Bering Sea have measured temperature, salinity and currents in the water column as ice was 3703 advected over the moorings. In total, there were 42 sets of data showing the water column 3704 response to the ice. Several patterns were evident in the time series at the various moorings. To 3705 explore these patterns, seven examples were selected (two from M2, three from M4, one each 3706 from M5 and M8). 3707 3708 3709 3.3.1. No ice (Example: M2, autumn 2004 – spring 2005) From 1995 to the present, there was little (areal coverage of < 20% concentration) or no ice 3710 present for five years of observations at M2 and two years at M4. Data from these seven 3711 examples show how the water column mixes, cools and becomes more saline without the 3712 influence of ice. In October 2004, the water column at M2 was cooling and mixing (Fig. 6). 155 3713 From a mean August surface temperature of 12.5 °C and mean bottom temperature of 3.2 °C (not 3714 shown), the water column became well mixed at 5.7 °C at the end of October. The mean wind 3715 stress during the fall of 2004 (mean 0.23 N m-2) was not significantly stronger than the long-term 3716 (1995 – 2012) mean (0.20 N m-2). The water column remained well-mixed through the winter 3717 until April 8, with the bottom temperature reaching a minimum of ~2.0 °C in mid April. 3718 Minimum water-column temperature in 2004 was anomalously high, and was surpassed by only 3719 two other warm-years: January, 2001 (3.0 °C) and January, 2003 (3.4 °C). All three instances are 3720 from an unusually warm period, 2001 to 2005 (Stabeno et al., 2012b). Mean minimum winter 3721 temperature from 1995 to 2011 at M2 was -0.6 °C, while the mean for the nine years excluding 3722 this warm window (2001 – 2005) was -1.7 °C (Table 2). Salinity increased from ~31.7 in early 3723 October to ~32.3 in mid February most likely from advection, of more saline water from the outer 3724 shelf and slope (Sullivan et al., 2008). 3725 3726 3727 3.3.2. A single ice event (Example: M2, fall 2007 – spring 2008) During the winter and spring 2007 3728 ice was present near M2 for 3729 approximately one month (Fig. 7). 3730 During this period, winds were moderate, 3731 with isolated storms interspersed with 3732 periods of weak winds. The water 3733 column became well mixed (defined as 3734 when the vertical temperature difference 3735 was < 0.04 °C, and the salinity difference 3736 was < 0.02) at 3.6 °C in mid November, 3737 2006 (not shown). Ice arrived in the 3738 vicinity of the mooring on March 2, and 3739 four days later, as the ice coverage 3740 reached ~20%, the water column began 3741 to stratify with colder, fresher water 3742 overlying warmer, saltier water. The top 3743 39 m of the water column cooled to the 3744 freezing point (-1.7 °C) by mid March 3745 and ice melt freshened the surface salinity by ~0.5. The water column remained stratified for ~20 3746 days. There were four moderate storms during the ice period (Fig. 7, top, 1 – 4). The first storm Figure 7. Time series from M2, February 27 – April 17, 2007, showing a) wind stress, b) temperature at 12 depths distributed through the water column and the percent areal ice cover in the 100-km x 100-km box around M2, and c) salinity at three depths. The gray shading indicates times when the water column was well mixed. 156 3747 (March 6-9) coincided with the arrival of the ice; the second (March 18-22) occurred during a 3748 period of >90% ice cover; and the third (March 25 – 27) occurred during a period of decreasing 3749 ice. The second storm coincided with a slight mixing at 39 – 44 m and the third event resulted in 3750 the total mixing of the water column. The primary cause of the greater mixing associated with 3751 the third storm, which was only slightly stronger than the second, was likely reduced ice cover. 3752 Less areal ice cover permits the more rapid movement of ice floes and also greater direct energy 3753 transfer between the atmosphere and the water. 3754 The duration and ice concentration at M2 in 2007 (Fig. 7) typified ice coverage in the 3755 south. Ice cover at M2 is more variable, has lower areal concentration and persists for shorter 3756 periods of time than at the other three mooring areas (Stabeno et al., 2012a). M2 is outside the 3757 direct path of the ice conveyor belt corridor and it is likely that ice near M2 arrived from Nunivak 3758 Island and the coastal polynya to the east. Over 17 year period (late winter 1995 to spring, 2011), 3759 single winter ice events occurred 6 times at M2 (lasting 20 - 83 days) and 7 times at M4 (lasting 3760 11 - 121 days). 3761 3762 3763 3.3.3. Multiple ice events with incomplete mixing (Example: M4, fall 2010 – spring 2011) During some years, ice can be intermittent, with ice advancing and retreating several 3764 times during the cold season (December – May). The winter and spring of 2011 was such a 3765 season at M4. The water column became well mixed at ~1.7 °C in late November 2010 (not 3766 shown) and was still well mixed when northerly winds forced the ice over the mooring on 3767 January 20 (Fig. 8). Strong winds kept the water column well mixed as the ice cooled and 3768 freshened the water. The ice persisted for approximately one week and then retreated with 3769 southeasterly winds. Following this initial ice advance and retreat, the winds remained variable, 3770 forcing four major ice events over the next three months. As expected, periods of northerly or 3771 northeasterly winds resulted in ice advancing, while periods of southerly winds resulted in ice 3772 retreat. Maximum areal ice concentration remained below 70%, except for the final ice event. 3773 While variable, salinity (especially at 31 m) generally increased from January into late 3774 March in spite of the melting ice evidenced by the cooling of the surface waters. Large 3775 freshening events (> 0.3) of the complete water column occurred (e.g., early March, early April), 3776 but freshening of just the surface waters (from ice melt) occurred more often. The greatest 3777 decrease in salinity (0.6) occurred in late April when ice concentrations approached 100%. The 3778 157 3779 Figure 8. Time series from M4, January 19 – May 1, 2011, showing a) 6-hourly wind stress, b) 12-hourly wind velocity, c) temperature at 16 depths distributed through the water column and the percent areal ice cover in the 100-km x 100-km box around M4, and d) salinity at three depths. The gray shading indicates times when the water column was well mixed. Asterisks (*) indicate periods of increased temperature and salinity, likely a result of advection. Two storms are indicated in each panel by a red bar. 158 3780 3781 temperature time series showed recurring, albeit weak, stratification (< 1.5 °C difference between 3782 top and bottom), interspersed with well-mixed periods (Fig. 8, gray shading). The cooling and 3783 freshening of the upper 20 - 27 m of water with the advent of ice was similar to the event 3784 described at M2 in 2007 (Fig. 7). The 3785 water column began mixing after each 3786 ice retreat and was usually well mixed 3787 before the next arrival of ice, only to 3788 become stratified again. The two wind 3789 events (A and B in Fig. 8 a) were similar 3790 in magnitude. During the first storm the 3791 water column was uniformly stratified 3792 (Fig. 8c; ~1 °C in temperature, 0.3 in 3793 salinity). The winds during the first 3794 storm mixed the water column, but not 3795 completely. During the second storm, 3796 the water column mixed completely. 3797 Wind mixing became most effective in 3798 both cases when ice concentration fell 3799 below 30%. 3800 The bottom temperatures often 3801 warmed after mixing episodes (Fig. 8b), 3802 an indication of advection. Similarly the 3803 0.7 increase in salinity from mid January 3804 through late March was also indicative of 3805 advection. Mean winter currents at M4 3806 are southward (Stabeno et al., 2010). 3807 Currents, which flow north along the 3808 100-m isobath, can circle around St. Paul Island (Sullivan et al., 2008) and advect warmer, more 3809 saline water onto the middle-shelf near M4. 3810 Figure 9. Time series from M4, February 2 – April 24, 2008, showing a) wind stress, b) temperature at 12 depths distributed through the water column and the percent areal ice cover in the 100-km x 100-km box around M4, and c) salinity at three depths. Low-pass filtered currents are shown at d) 11 m and e) 55 m. The gray shading indicates times when the water column was well mixed. The red bar at the bottom of each panel represents the time period shown in Fig. 10. Multiple instances of ice advance and retreat (>2 events per year) were not uncommon at 3811 the southern moorings: 4 of 17 years at M2; 1 of 17 at M4; and 3 of 17 at M5. The example 3812 discussed here (Fig. 8) had the most ice events. This example, with a persistent series of 159 3813 advances and retreats, serves as a good example of both wind and advection influencing ice melt 3814 and winter stratification events. 3815 3816 3.3.4. A cold year with persistent ice (Example: M4, fall 2007 – spring 2008) 3817 During 2007-2008 (both cold years, Stabeno et al., 2012b), the water column became well 3818 mixed on November 15 at 2.8 °C and cooled to -0.4 °C by January 13 (not shown). Ice reached 3819 the area in early February coincident with strong winds (Fig. 9). The water column was well 3820 mixed for 3 - 4 days in early February, and then became stratified as winds weakened and ice 3821 concentration increased to > 20%. The ice-covered water column became well mixed again in 3822 late February and remained well mixed or weakly stratified until late March. Once again, there 3823 are strong indications that advection played a role in stratifying the water column. In both early 3824 February and late March there was a marked increase in temperature and salinity below 40 m, 3825 while the temperature and salinity in the upper water column remained relatively constant. Low- 3826 pass filtered, near-bottom currents during the February event were > 15 cm s-1 from the south. 3827 During the March event, currents were variable and relatively weak (< 10 cm s-1) (Fig. 9 d, e). 3828 While the daily mean currents were relatively weak in mid-March (Fig. 9), the tidal 3829 currents were significantly stronger, 3830 regularly exceeding 40 cm s-1 (Fig. 10). 3831 The first and second increase in bottom 3832 temperature and salinity occurred when 3833 the currents began to turn 3834 eastward. Once the boundary (front) 3835 between the warmer, more saline bottom 3836 water and the cooler, fresher bottom 3837 water passed the mooring, the 3838 relationship between the events and the 3839 direction of the current became complex. 3840 Bottom frontal structures are often not 3841 straight lines, but rather are crooked, 3842 sometimes following the bottom 3843 bathymetry. The low-pass filtered data 3844 show a relatively steady increase in 3845 stratification in late March (Fig. 9b, c), 3846 but the unfiltered data show that the Figure 10. Hourly time series at M4 of a) temperature at 12 depths, b) hourly salinity at three depths, and hourly current velocity at c) 11 m and d) 55 m. The 2008 time period shown is the same as that indicated by the red bars in Fig. 9. The blue shading indicates periods when the bottom currents have a northward component. Northward is upward. 160 3847 increase in stratification occurred as a series of events related to the tidal currents. That the 3848 surface temperatures remain near freezing is not surprising since there was ~100% ice cover in 3849 the 100-km x 100-km box around M4 keeping the upper water column cold and fresh. 3850 3851 3852 3.3.5. Two ice events, one stratified and one well mixed (Example: M5, 2005 – 2006) During the cold season of 2005 – 2006 extensive ice arrived at M5 in December and finally 3853 departed in early June. Four periods of strong ice retreat occurred over this time (Fig. 11). The 3854 water column became fully mixed in early November at ~3.1 °C (not shown), and remained so 3855 until mid December when ice cover reached ~30%. Two distinct periods of > 90% ice 3856 concentration occurred. During the first ice event, late December through late February, the 3857 winds were relatively weak for the first 6 weeks and strengthened for the last 2 weeks. In 3858 contrast, during the second ice event, the winds were stronger during the first 5 weeks and weaker 3859 during the last 2 – 3 weeks. Ice, combined with low wind stress during the first ice event, cooled 3860 and freshened only the upper water column (upper 3861 24 – 42 m) without greatly impacting the deeper 3862 water. The sharp increase in bottom salinity in early 3863 February, and the accompanying increase in 3864 temperature was likely due to advection. Areal ice 3865 concentration fell to <30% in late February, and 3866 under the influence of relatively strong winds, the 3867 water mixed. The storms on February 12 with 3868 >50% ice cover had little impact mixing the water 3869 column, while a week later after the ice began 3870 retreating, winds of similar magnitude readily 3871 mixed the water column. A second large influx of 3872 ice occurred in mid-March. In sharp contrast to the 3873 first event, the water column remained largely well 3874 mixed through two months of 60-90% ice 3875 concentration. During this time the temperature (~ - 3876 1.8 °C) and the salinity (~ 31.8) remained relatively 3877 constant. That the water column temperature was at 3878 the freezing point with constant salinity supports the conclusion that there was little net ice melt 3879 during the second ice event. Figure 11. Time series from M5, December 14, 2005 – May 31, 2006, showing a) wind stress, b) temperature at 11 depths distributed through the water column and the percent areal ice cover in the 100-km x 100-km box around M5, and c) salinity at three depths. The gray shading indicates times when the water column was well mixed. 161 3880 M5, similar to M4, is solidly within the ice conveyor belt (Fig. 1). Ice cover was often 80- 3881 100%, either steadily retaining a high concentration of ice all season, or partially retreating for a 3882 periods of days to weeks and returning to a high concentration. Areal ice concentration data at 3883 M4 show five years of the single ice event, and 10 years of multiple ice events for the period fall 3884 1994 – spring 2011. As such, the cold season of 2005 – 2006 was fairly typical at M5. 3885 3886 3887 3888 3.3.6. Months of ~100% ice cover (Example: M8, 2008 – 2009) M8 is the farthest north of the four moorings (Fig. 1), and representative of the northern 3889 shelf (Stabeno et al., 2012a). The water column at M8 from 2005-2011 mixed notably later (early 3890 December to late January) and at colder temperatures (average -1.0 °C) than at M2, M4, or M5 3891 (Table 2). The average minimum winter temperature over those years was -1.8 °C, which was 3892 colder than at the southern moorings as a result of higher salinity. In 2008 the water column 3893 became well mixed at -0.1 °C in early December (Fig. 12). Sea ice arrived in late December and 3894 stratified the water column for a few days until the water column cooled to < -1.7 °C. The high 3895 areal ice cover (>90%) and a well-mixed or weakly stratified water column persisted for ~165 3896 days. Surface water began to warm 3897 and stratify ahead of May ice melt, 3898 and ice concentration was < 20% by 3899 the end of May, 2009. 3900 The patterns of strong pre- 3901 ice cooling, minimal ice melt, 3902 minimal decrease in water 3903 temperature, and areal ice 3904 concentrations >80% for more than 4 3905 months were typical at M8. The 3906 duration of ice cover (Fig. 12) was 3907 typical for the period (1995 – 2011) 3908 for the northern shelf. Over the 17- 3909 year period, ice was present an 3910 average of 127 days, with the longest 3911 single, near-100% ice concentration 3912 event lasting 189 days in 2005 – 3913 2006. Twelve years of the 17 years Figure 12. Time series from M8, November 15, 2008 – June 30, 2009, showing a) wind stress, b) temperature at 11 depths distributed through the water column and the percent areal ice cover in the 100-km x 100-km box around M8, and c) salinity at three depths. The gray shading indicates times when the water column was well mixed. 162 3914 examined had only one ice event (80 – 100% coverage) during the cold season, and the remaining 3915 five had two. 3916 3917 3918 3.4 Estimating the magnitude of ice melt and its direct influence on temperature The thickness of the total ice melt can be estimated using the time series from the moorings 3919 and Equations (2) and (3). The largest problem with selecting data for this calculation was 3920 advection. Care was made in selecting examples where freshening was well defined and 3921 advection did not play an important role in modifying the water column. Of the 42 sets of winter 3922 and spring data (1995 – 2012), 15 sets were suitable for this calculation (Table 3). For each of 3923 these data sets the water column was well mixed at the time of the ice’s arrival; the water column 3924 (or at least the upper layer) cooled to the freezing point within a month of ice arrival; and there 3925 was measurable cooling and freshening of the water column. Advection did not appear to 3926 influence the system in these selected instances. 3927 Among the 14 annual examples (the two sets from M2, 2009 were combined together to 3928 provide total cooling for that year) most were from the southern moorings with only three from 3929 M5 and none from M8. The lack of examples from M8 was primarily because the water column 3930 was already at or near the freezing point when ice first appeared at that mooring site and any 3931 stratification resulting from the arrival of ice was minimal and short lived. 3932 From each of the examples, the average initial salinity (Sw) and initial ocean temperature (Tw) 3933 were determined (Table 3). After the arrival of ice, the water column often became stratified and 3934 then re-mixed to a depth of H. Usually it mixed to the bottom, but in certain instances only the 3935 surface was mixed before the retreat of ice. The change in salinity and temperature of this mixed 3936 layer was calculated. Using Equation (2) and an ice salinity of 6 (Table 1), hw was calculated. 3937 Using the mean density (0.89 x103 kg m-3) from Table 1 the thickness (hi) of ice melted was 3938 estimated for each data set, giving a mean thickness of 1.3 m. At -1.7 °C and a salinity of 6, the 3939 latent heat of fusion of sea ice (L) is 65 cal g-1 (Weeks and Weller, 1984). Using Eq. 3, the 3940 L) was calculated for each example 3941 (Table 3). The latent heat of fusion accounted for an estimated 26-100 % of total observed 3942 cooling. On average almost half of the cooling of the water column was due solely to ice melt; the 3943 remainder was from air-sea interaction either directly or through the ice. 3944 A similar estimate can be made when ice melted in place in the spring (Table 4). At the 3945 southern moorings most ice retreated through advection. At the northern two moorings, however, 3946 often there was a sharp decrease in salinity associated with loss of sea ice. This decrease in 3947 salinity resulted in a fresh surface layer with a mean depth of 28 m (Table 4). This fresh water 163 3948 layer persists at M8 throughout the summer season (Stabeno et al., 2012a). The estimated amount 3949 of ice melted ranged from 0.31 m to 1.51 m, with an average of 0.84 m. The balance between 3950 atmospheric warming and cooling due to ice melt (latent heat) resulted in the upper-layer 3951 temperature remaining cold. 3952 In the winter, the arrival of ice freshened and cooled the southern shelf, while in spring the 3953 retreat of ice freshened the northern shelf. These estimates of ice melt are probably 3954 underestimates, because ice melt resulting from the advection of warmer water was not included 3955 and numerous minimal changes in temperature/salinity were also neglected. The estimation that 3956 >1.3 m of ice melted over the middle domain of the southern shelf could not be used to calculate 3957 the total ice melt over the southern shelf. Once the water column reached the freezing point, ice 3958 continued to be transported southward without modifying the water column and melted at the 3959 leading edge often over the slope. 3960 3961 3962 4. Summary and Conclusions While variability in the winds resulted in sudden changes in the distribution and areal 3963 concentration of the ice over the cold season, the winds were primarily from the north resulting in 3964 the creation of polynyas along the leeward coasts and a south- or southwestward transport of sea 3965 ice over the Bering Sea shelf. The wind-driven conveyor belt of Pease (1980) originated north- 3966 northeast of St. Lawrence Island fanning south and westward over the shelf. This main corridor 3967 provided ice primarily to the areas around moorings M4 and M5. The St. Lawrence polynya and 3968 the region around Anadyr contributed ice to the area around M8. Finally, the Alaskan coastal 3969 polynyas in Bristol Bay and south of Nunivak Island were the primary suppliers to sea ice in the 3970 vicinity of M2. 3971 The moorings extend along the 70-m isobath of the middle shelf, with M8 on the northern 3972 shelf, M5 in the transition zone between north and south, and M2 and M4 on the southern shelf. 3973 Thus, examining data from these moorings gives insight on how the middle shelf responded 3974 spatially and temporally to ice melt. Over the southern part of the middle shelf (south of ~ 60 3975 °N; Stabeno et al., 2012a), the primary ice melt occurred when the ice was advected southward. 3976 Approximately 1.3 m of ice melted, freshening the water column and reducing the concentration 3977 of nutrients. Over the northern part of the middle shelf, as represented by M8, there was little ice 3978 melt with the first arrival of ice. Most of the melt at this site occurred when ice retreated, and ice 3979 melt during retreat left behind a fresh surface layer, which persisted at M8 through the summer. 3980 This increased the strength of stratification and delayed the fall mixing of the water column on 164 3981 the northern shelf by ~1.3 months relative to M2 (Table 2). The transition zone at M5 had 3982 substantial ice melt during both ice advance and retreat (a total ~2.1 m; Tables 3 and 4). 3983 Typically when ice first arrived, the water column (or the surface mixed layer) cooled 3984 and freshened, and remained well mixed. Once the areal ice cover exceeded 20 – 25 %, the 3985 melting ice stratified the water column, with a fresher, colder layer overlaying warmer, more 3986 saline water. This surface layer was usually > 20 m deep. The vertical stratification usually was 3987 t = ~ 0.18 (using average temperature and salinity values from Table 3 of -1.7 3988 °C and 31.7, respectively, for surface layer, and 1.3 °C and 32.2 for bottom layer), but persisted 3989 for 10 - 25 days. Storms quickly re-mixed the water column if ice retreated (e.g. Fig. 8); storms 3990 had less influence under extensive ice, but the water column would eventually become well 3991 mixed (e.g. Figs. 7, 9 and 10). Advection plays a role in the stratification of the water column, 3992 particularly in the vicinity of the ice edge front. 3993 Without sea ice, the water column at M2 and M4 usually cools by ~4 °C to ~2 °C, while 3994 in years with extensive ice the water column cools on average more than 6 °C to -1.7 °C (Stabeno 3995 et al., 2012b). Multiple consecutive cold (warm) years modify this, making the depth-averaged 3996 summer temperatures colder (warmer). Average cooling in a series of cold years was ~5.5 °C, 3997 while average cooling in a series of warm years was ~4.5 °C (from Fig. 4A in Stabeno et al., 3998 2012b). For comparison, more cooling (~ 7.5 °C) occurred between warm and cold years. The 3999 greater cooling during cold years rather than warm years comes from two sources. First, the air 4000 temperature during years with extensive ice is colder than years with little or no ice; thus, cooling 4001 due to sensible heat flux is greater. The second and probably more important source of cooling is 4002 the latent heat flux due to ice melt. The cooling from melting 1.3 m of ice (probably an 4003 underestimate) accounts for approximately half the observed cooling of the water column. In 4004 sharp contrast, on the northern shelf near M8, the water column cooled to ~ -1 °C before ice 4005 arrived (Table 2). Ice arrival cooled water temperatures to ~ -1.8 °C with little melt because water 4006 temperatures were already low. 4007 This discussion of sea-ice melt applies to the middle shelf domain, and not to the coastal 4008 or outer shelf domains. In the coastal domain, ice is formed and shore fast ice and rafting are 4009 common. The water column is typically well mixed by tides which are stronger on the 4010 southeastern shelf than in the north. In the outer shelf domain advection dominates, and intrusions 4011 of warmer, more saline slope water are common (e.g. Stabeno and van Meurs, 1999; Sullivan et 4012 al., 2008; Coachman, 1986). The water column is deeper (> 100 m) and not well mixed even in 4013 the winter (Stabeno et al., 1998). A few hypotheses, however, can be made about how the coastal 4014 and outer domains may respond to ice melt. Probably little ice is melted in the coastal domain 165 4015 until ice retreat at the end of the cold season. The coastal domain, with its weak alongshore flow, 4016 retains the signature of lower salinity through the summer as a result of ice melt (Danielson et al., 4017 2012; Kachel et al., 2002). With the breakdown of the Inner Front in late summer, this freshwater 4018 signature spreads out over the shelf (Ladd and Stabeno, 2012). The signature of ice melt on the 4019 outer domain is ephemeral; it does not persist through the summer because of the persistent 4020 alongshore currents, and onshore intrusions of more saline, nutrient rich water (Stabeno et al., 4021 2010). 4022 It is becoming evident that the presence or absence of sea ice in spring is the single most 4023 important component determining the physical and biological structure of the eastern Bering Sea 4024 shelf ecosystem, not only in the winter and spring when it is present, but also during the warm 4025 season (Stabeno et al., 2010, 2012a, b; Hunt et al., 2002, 2008, 2010). The advance of sea ice 4026 early in the season transports fresh water southward, which then melts at the ice edge reducing 4027 the salinity of the middle shelf by ~0.4 (Table 3). In contrast late in the cold season, the retreat of 4028 ice transports fresh water northward, where it melts providing approximately half of the summer 4029 stratification on the northern shelf (Stabeno et al., 2012a; Ladd and Stabeno, 2012). Sea ice had 4030 low concentrations of nitrate and, thus, could not contribute to the water column as the ice 4031 melted. There are significant amounts of iron, however, in the sea ice (Aguilar-Islas, 2008), 4032 which contributes to the production along the slope. Significant concentrations of chlorophyll-a 4033 were found in bottom sections of the ice cores. This ice algae, located at the ice-seawater 4034 interface, may provide an early concentrated food source (Mock and Gradinger, 2000; Niemi et 4035 al., 2011) for zooplankton (e.g. O’Brien 1987, Runge and Ingram 1991). While chlorophyll-a is 4036 transported by the ice toward the ice edge, it is not known if the ice also transports large amounts 4037 of zooplankton which feed on the ice algae. The absence of ice over the southern shelf during 4038 consecutive warm years (2001-2005) likely contributed, directly or indirectly, to a drop in 4039 populations of large crustacean zooplankton (e.g. Calanus marshallae, Thysanoessa raschii) and 4040 a resultant reduction in the pollock fishery (Hunt et al., 2008, 2010; Coyle et al., 2008, Stabeno et 4041 al., 2012b). 4042 4043 Acknowledgements 4044 We thank the officers and crews of the NOAA ships Miller Freeman and Oscar Dyson, U. S. 4045 Coast Guard Cutter Healy, and R/V Thomas G. Thompson for the invaluable assistance in making 4046 these oceanographic measurements. Special thanks go to the personnel who collected the ice 4047 cores including E. Cokelet, D. Kachel, P. Proctor, J. Cross, and members of the National Marine 4048 Mammal Lab during the 2006 ice cruise lead by M. Cameron and J. Bengston. We thank W. 166 4049 Floering, C. Dewitt and S. McKeever for maintaining, deploying and recovering the instruments 4050 on the mooring, D. Kachel and P. Proctor for processing the mooring and ship board 4051 measurements, P. Proctor for nutrient analysis on some samples, and A. Hermann for comparison 4052 of ice drift data to interpolated NCEP winds. Graphics were provided by K. Birchfield. This 4053 research was funded by grants from The North Pacific Research Board (grant B52) and the 4054 National Science Foundation (grants 0732430, 1107250, and 0813985), and support from 4055 NOAA’s North Pacific Climate Regimes and Ecosystem Productivity (NPCREP) program. This 4056 publication was partially funded by the University of Washington Joint Institute for the Study of 4057 the Atmosphere and Ocean (JISAO) under NOAA Cooperative Agreements NA17RJ1232 and 4058 NA10OAR4320148, and is contribution EcoFOCI-0802 to NOAA’s Ecosystems and Fisheries- 4059 Oceanography Coordinated Investigations, contribution 2157 to JISAO, contribution 3975 to 4060 NOAA’s Pacific Marine Environmental Laboratory, and BEST-BSIERP publication number XX. 4061 The findings and conclusions in this paper are those of the authors and do not necessarily 4062 represent the views of NOAA’s Oceans and Atmospheric Research. 4063 4064 References 4065 Aguilar-Islas, A.M., Rember, R.D, Mordy, C.W., Wu, J., 2008. Sea ice-derived dissolved iron 4066 and its potential influence on the spring algal bloom in the Bering Sea. Geophys. Res. Lett. 4067 35, http://dx.doi.org/10.1029/2008GL035736. 4068 4069 4070 Alexander, V., Niebauer, H.J., 1981. Oceanography of the eastern Bering Sea ice-edge zone in spring, Limnol. Oceanogr. 26 (6), 1111–1125. Brown, Z. W., Arrigo, K. R., 2013, Sea ice impacts on spring bloom dynamics and net primary 4071 production in the Eastern Bering Sea, J. Geophys. Res. J. Oceans, 118, 43–62, 4072 doi:10.1029/2012JC008034. 4073 Cavalieri, D.J., Markus, T., Comiso, J.C., 2003. AMSR-E/Aqua Daily L3 12.5 km Brightness 4074 Temperature, Sea Ice Concentration, & Snow Depth Polar Grids. Version 2. 2002-2011. 4075 Boulder, Colorado USA: NASA DAAC at the National Snow and Ice Data Center, 4076 http://nsidc.org/data/amsre/. 4077 4078 Coachman, L.K., 1986. Circulation, water masses and fluxes on the southeastern Bering Sea shelf. Cont. Shelf Res. 5 (1–2), 23–108. 4079 Coyle, K. O., Pinchuk, A. I., Eisner, L. B., Napp, J. M., 2008. Zooplankton species composition, 4080 abundance and biomass on the eastern Bering Sea shelf during summer: the potential role of 4081 water column stability and nutrients in structuring the zooplankton community. Deep Sea 4082 Research II, 55: 1755–1791. 167 4083 Danielson, S., Aagaard, K., Weingartner, T., Martin, S., Winsor, P., Gawarkiewicz, G., 4084 Quadfasel, D., 2006. The St. Lawrence polynya and the Bering shelf circulation: New 4085 observations that test the models, J. Geophys. Res., J111, C09023, 4086 doi:10.1029/2005JC003268. 4087 Danielson, S., Hedstrom, K., Aagaard, K., Weingartner, T., Curchitser, E., 2012. Wind-induced 4088 reorganization of the Bering Shelf circulation. Geophys. Res. Lett. 39 (8), 4089 http://dx.doi.org/10.1029/2012GL051231. 4090 Drucker, R., Martin, S., Moritz, R., 2003. Observations of ice thickness and frazil ice in the St. 4091 Lawrence Island polynya from satellite imagery, upward looking sonar, and 4092 salinity/temperature moorings, J. Geophys. Res., 108(C5), 3149, doi:10.1029/2001JC001213. 4093 Eicken, H., 2003. From the microscopic to the macroscopic to the regional scale: Growth, 4094 microstructure and properties of sea ice, in: Thomas, D.N., Dieckmann, G.S. (Eds.), Sea 4095 Ice—An Introduction to its Physics, Biology, Chemistry and Geology. Blackwells Scientific 4096 Ltd., London, pp. 22–81. 4097 Gordon, L.I., Jennings Jr, J.C., Ross, A.A., Krest, J.M., 1994. A suggested protocol for 4098 continuous flow automated analysis of seawater nutrients (phosphate, nitrate, nitrite and 4099 silicic acid) in the WOCE Hydrographic Program and the Joint Global Ocean Fluxes Study. 4100 WHP Operations and Methods. WOCE Hydrographic Program Office, Methods Manual 91- 4101 1. November. 4102 Holt, B., Rothrock, D.A., Kwok, R., 1992. Determination of sea ice motion from satellite images, 4103 in: Carsey, F.D. (Ed.), Microwave Remote Sensing of Sea Ice, Geophys. Monogr. Ser., Vol. 4104 68, AGU, Washington, D.C., doi:10.1029/GM068p0343, pp. 343–354. 4105 Hunt, Jr., G.L., B.M. Allen, R.P. Angliss, T. Baker, N. Bond, G. Buck, G.V. Byrd, K.O. Coyle, 4106 A. Devol, D.M. Eggers, L. Eisner, R.A. Feely, S. Fitzgerald, L.W. Fritz, E.V. Gritsay, C. 4107 Ladd, W. Lewis, J. Mathis, C.W. Mordy, F. Mueter, J. Napp, E. Sherr, D. Shull, P. Stabeno, 4108 M.A. Stepanenko, S. Strom, T.E. Whitledge (2010): Status and trends of the Bering Sea 4109 region, 2003-2008. In Marine Ecosystems of the North Pacific Ocean, 2003-2008, S. M. 4110 McKinnell and M. J. Dagg (eds.), PICES Special Publication 4, 196-267. 4111 4112 4113 Hunt Jr., G.L., Stabeno, P.J., 2002. Climate change and the control of energy flow in the southeastern Bering Sea. Prog. Oceanogr. 55 (1–2), 5–22. Kachel, N.B., Hunt Jr., G.L., Salo, S.A., Schumacher, J.D., Stabeno, P.J., Whitledge, T.E., 2002. 4114 Characteristics and variability of the inner front of the southeastern Bering Sea. Deep-Sea 4115 Res. II 49 (26), 5889–5909. 168 4116 Kanamitsu, M., Ebisuzaki, W., Woollen, J., Yang, S.-K., Hnilo, J.J., Fiorino, M., Potter, G.L., 4117 2002. NCEP-DOE AMIP-II Reanalysis (R-2). Bull. Amer. Meteor. Soc. 83, 1631–1643. 4118 Ladd, C., Bond, N.A., 2002. Evaluation of the NCEP/NCAR reanalysis in the NE Pacific and the 4119 4120 4121 4122 Bering Sea. J. Geophys. Res., 107 (C10), 3158, doi: 10.1029/2001JC001157. Ladd, C.A., Stabeno, P.J., 2012. Stratification on the Eastern Bering Sea shelf revisited. Deep-Sea Res. II 65–70, 72–83. Leberl, F., Raggam, J., Elachi, C., Campbell, W.J., 1983. Sea ice motion measurements from 4123 SEASAT SAR images, J. Geophys. Res. 88 (C3), 1915–1928, 4124 doi:10.1029/JC088iC03p01915. 4125 Leu, E., Søreide, J. E., Hessen, D. O., Falk-Petersen, S., Berge, J., 2011, Consequences of 4126 changing sea-ice cover for primary and secondary producers in the European Arctic shelf 4127 seas: timing, quantity, and quality. Prog. Oceanogr. 90(1), 18-32. 4128 4129 4130 4131 Macklin, S.A., Pease, C.H., Reynolds, R.M., 1984. Bering Air-Sea-Ice Study (BASICS), February and March 1981. NOAA Tech Memo ERL PMEL-52, 104 pp. Malmgren, F., 1927. On the Properties of Sea Ice, Norwegian North Polar Expedition with the “Maud” 1918–1925: Scientific Results, Vol. 1, No. 5, 67 pp. 4132 Maslanik, J., Stroeve, J., 1999. Near-Real-Time DMSP SSM/I-SSMIS Daily Polar Gridded Sea 4133 Ice Concentrations. 1994-2002. (Updated daily.) Boulder, Colorado USA: NASA DAAC at 4134 the National Snow and Ice Data Center, 4135 http://nsidc.org/data/docs/daac/ssmi_instrument.gd.html. 4136 McNutt, S.L., 1981. Remote sensing analysis of ice growth and distribution in the Eastern Bering 4137 Sea. Chapter 10 in Eastern Bering Sea Shelf: Oceanography and Resources, D.W. Hood and 4138 J.A. Calder (eds.), Vol. 1, USDOC/NOAA/OMPA, 141–165. 4139 Michel, C., Legendre, L., Ingram, R. G., Gosselin, M., Levasseur, M., 1996. Carbon budget of 4140 sea-ice algae in spring: evidence of a significant transfer to zooplankton grazers. J. Geophys. 4141 Res. 101(C8), 18345-18 4142 Mock T., Gradinger R., 2000. Changes in photosynthetic carbon allocation in algal assemblages 4143 of Arctic sea ice with decreasing nutrient concentrations and irradiance. Mar. Ecol. Prog. Ser. 4144 202: 1-11. 4145 4146 4147 4148 Muench, R.D., Ahlnas, K., 1976. Ice movement and distribution in the Bering Sea from March to June 1974. J. Geophys. Res. 81, 4469–4476. Muench, R.D., Schumacher, J.D., Salo, S. A., 1988: Winter currents and hydrographic conditions on the northern central Bering Sea shelf. J. Geophys. Res., 93 (C1), 516-526. 169 4149 4150 4151 Mueter, F.J., Litzow, M.A., 2008. Sea ice retreat alters the biogeography of the Bering Sea continental shelf. Ecol. Appl. 18, 309–320, doi:10.1890/07-0564.1. Mundy, C.J., et al., 2009. Contribution of under-ice primary production to an ice-edge upwelling 4152 phytoplankton bloom in the Canadian Beaufort Sea. Geophys. Res. Lett. 36, L17601, 4153 doi:10.1029/2009GL038837. 4154 Niebauer, H.J., 1998. Variability in Bering Sea ice cover as affected by a regime shift in the 4155 North Pacific in the period 1947–1996. J. Geophys. Res. 103(C12), 27,717–27,737. 4156 Niebauer, H.J., Bond, N.A., Yakunin, L.P., Plotnikov, V.V., 1999. An update on the climatology 4157 and sea ice of the Bering Sea, in: Loughlin, T.R., Ohtani, K. (Eds.), The Bering Sea: A 4158 Summary of Physical, Chemical and Biological Characteristics and a Synopsis of Research. 4159 North Pacific Marine Science Organization, PICES, Alaska Sea Grant, pp. 29–60. 4160 Niebauer, H.J., Schell, D.M., 1993. Physical environment of the Bering Sea population, in: Burns, 4161 J.J., Montague, J.J., Cowles, C.J. (Eds.), The Bowhead Whale. The Society for Marine 4162 Mammalogy Special Pub. No. 2, Seattle, Washington, pp. 23–43. 4163 4164 Niemi, A., Michel, C., Hille, K., Poulin, M. 2011 Protist assemblages in winter sea ice: setting the stage for the spring ice algal bloom. Polar Biol. 34: 1803-1817. 4165 O'Brien, D.P. 1987. Direct observations of the behavior of Euphausia superba and Euphausia 4166 crystallorophias (Crustacea: Euphausiacea) under pack ice during the Antarctic spring of 4167 1985. J. Crustacean Biol. 7: 437-448. 4168 Pease, C.H., 1980. Eastern Bering Sea ice processes. Mon. Weather Rev. 108 (12), 2015–2023. 4169 Rodionov, S.N., Bond, N.A., Overland, J.E., 2007. The Aleutian Low, storm tracks, and winter 4170 4171 climate variability in the Bering Sea. Deep-Sea Res. II 54, 2560–2577. Runge, J.A., Ingram, R.G. 1991. Under-ice feeding and diel migration by the planktonic 4172 copepods Calanus glacialis and Pseudocalanus minutus in relation to the ice algal production 4173 cycle in southeastern Hudson Bay, Canada. Mar. Biol. 108: 217-225 4174 Schandelmeier, L., Alexander, V., 1981. An analysis of the influence of ice on spring 4175 phytoplankton population structure in the southeast Bering Sea. Limnol. Oceanogr. 26 (5), 4176 935–943. 4177 Schumacher, J.D., Aagaard, K., Pease, C. H., Tripp, R.B., 1983. Effects of a shelf polynya on 4178 flow and water properties in the Northern Bering Sea. J. Geophys. Res., 88, 2723-2732; 4179 Schumacher, J.D., Kinder, T.H., Pashinski, D.J., Charnell, R.L., 1979. A structural front over the 4180 4181 4182 continental shelf of the eastern Bering Sea. J. Phys. Oceanogr. 9 (1), 79–87. Shapiro, L.H., Burns, J.J., 1975. Satellite observations of sea ice movement in the Bering Strait region, in: Weller, G., Bowling, S.A. (Eds.), Climate of the Arctic. Twenty-fourth Alaska 170 4183 Science Conference, Fairbanks, Alaska, August 15 to 17, 1973. Geophysical Institute, 4184 University of Alaska, Fairbanks, pp. 379–386. 4185 Sigler, M.F., Stabeno, P.J., Eisner, L.B., Napp, J.M., Mueter, F.J., this issue. Spring and fall 4186 phytoplankton blooms in a productive subarctic ecosystem, the eastern Bering Sea, during 4187 1955–2011. 4188 4189 Stabeno, P.J., Bond, N.A., Kachel, N.B., Salo, S.A., 2007. On the recent warming of the southeastern Bering Sea shelf. Deep-Sea Res. II 54, 2599–2618. 4190 Stabeno, P.J., Bond, N.A., Kachel, N.B., Salo, S.A., Schumacher, J.D., 2001. On the temporal 4191 variability of the physical environment over the south-eastern Bering Sea. Fish. Oceanogr. 4192 10(1), 81–98. 4193 Stabeno, P.J., Farley Jr., E.V., Kachel, N.B., Moore, S., Mordy, C.W., Napp, J.M., Overland, J.E., 4194 Pinchuk, A.I., Sigler, M.F., 2012a. A comparison of the physics of the northern and southern 4195 shelves of the eastern Bering Sea and some implications for the ecosystem, Deep-Sea Res. II 4196 65–70, 14–30. 4197 Stabeno, P.J., Kachel, N., Mordy, C., Righi, D., Salo, S., 2008. An examination of the physical 4198 variability around the Pribilof Islands in 2004. Deep-Sea Res. II 55 (16–17), doi: 4199 10.1016/j.dsr2.2008.03.006, 1701–1716. 4200 Stabeno, P.J., Kachel, N.B., Moore, S.E., Napp, J.M., Sigler, M., Yamaguchi, A., Zerbini, A.N., 4201 2012b. Comparison of warm and cold years on the southeastern Bering Sea shelf and some 4202 implications for the ecosystem, Deep-Sea Res. II 65–70, 31–45. 4203 Stabeno, P.J., Napp, J., Mordy, C., Whitledge, T., 2010. Factors influencing physical structure 4204 and lower trophic levels of the eastern Bering Sea shelf in 2005: Sea ice, tides and winds. 4205 Prog. Oceanogr. 85 (3–4), 180–196. 4206 Stabeno, P.J., Schumacher, J.D., Davis, R.F., Napp, J.M., 1998. Under-ice observations of water 4207 column temperature, salinity and spring phytoplankton dynamics: Eastern Bering Sea shelf. J. 4208 Mar. Res. 56 (1), 239–255. 4209 4210 4211 4212 4213 4214 Stabeno, P.J., van Meurs, P., 1999. Evidence of episodic on-shelf flow in the southeastern Bering Sea. J. Geophys. Res., 104 (C12), doi: 10.1029/1999JC900242, 29,715–29,720. Strickland, J.D.H., Parsons, T.R., 1968. A Practical Handbook of Seawater Analysis, 1st Ed. Bull. 167, Fish. Res. Bd Can., Ottawa, 293 pp. Stringer, W. J., Groves, J. E., 1991. Location and areal extent of polynyas in the Bering and Chukchi Seas, Arctic, 44, 164-171. 171 4215 Sullivan, M.E., Kachel, N.B., Mordy, C.W., Stabeno, P.J., 2008. The Pribilof Islands: 4216 Temperature, salinity and nitrate during summer 2004. Deep-Sea Res. II 55 (16–17), 4217 http://dx.doi.org/10.1016/j.dsr2.2008.03.004, 1729–1737. 4218 4219 Timco, G.W., Frederking, R.M.W., 1996. A review of sea ice density. Cold Regions Science and Technology 24 (1), 1–6. 4220 Weeks, W.F., Weller, G., 1984. Offshore oil in the Alaskan Arctic. Science 225, 371–378. 4221 Wyllie-Echeverria, T., Wooster, W.S., 1998. Year to year variations in Bering Sea ice cover and 4222 4223 4224 some consequences for fish distributions. Fisheries Oceanography 7, 159–170. Zhang, J., Woodgate, R., Moritz, R., 2010. Sea ice response to atmospheric and oceanic forcing in the Bering Sea. J. Phys. Oceanogr. 40 (8), 1729–1747. 4225 172 4226 4227 Tables 4228 4229 Table 1. Ice stations, 2006-2009, with average data from the core. April 19, 2007 Ice Core 2 ice 4230 thickness value was excluded from the mean thickness. 4231 Date Ice Snow station depth (cm) Ice Air Mean thickness temp. salinity (cm) (°C) Mean Mean Mean temp. ice nitrate (°C) density (µM) (103 kg m-3) 29-Apr-06 01 (D3) 2 50 -5.5 4.86 -1.72 0.89 0.4 30-Apr-06 02 (D4) 4 37 -5.9 7.23 -2.48 0.85 0.5 3-May-06 03 (E3) 27 100 -2.6 6.00 -1.58 0.97 2.2 17-Apr-07 1 1 51 -9.8 5.83 -3.75 0.91 0.3 19-Apr-07 2 -- 4.05 -4.14 0.89 1.1 21-Apr-07 3 2 30 -1.0 -- -2.00 -- 0.2 24-Apr-07 4 3 73 -1.2 5.79 -1.46 0.91 1.2 30-Apr-07 5 2 64 -- 4.74 -1.39 0.89 0.4 6-May-07 6 2 62 -1.0 5.26 -1.43 0.90 0.1 7-May-07 7 0 69 2.6 4.00 -0.10 0.82 3.1 3-Apr-08 1 10 44 0.9 6.57 -2.23 0.93 1.2 4-Apr-08 2 43 92 -10.0 5.64 -3.44 0.89 1.1 6-Apr-08 4 8 55 -5.3 5.77 -2.56 0.89 1.1 8-Apr-08 5 8 41 -13.8 5.86 -3.20 0.92 3.5 8-Apr-08 5a 6 40 -- 5.44 -3.25 0.91 1.5 12-Apr-08 6 4 76 -8.0 7.24 -2.86 0.83 1.0 15-Apr-08 8 17 85 -- -- -2.73 -- 0.3 16-Apr-08 9 11 59 -1.1 -- -2.38 -- 0.8 28-Apr-08 10 5 41 -2.5 6.34 -2.10 0.88 0.2 29-Apr-08 11 7 69 -4.0 6.04 -2.19 0.85 0.7 -- >329 173 30-Apr-08 12 5 71 -3.0 5.29 -2.13 0.88 2.1 5-Apr-09 1 4 73 1.5 5.70 -2.97 0.92 1.5 7-Apr-09 2 17 87 -0.2 5.05 -3.09 0.89 0.3 14-Apr-09 3 5 93 -10.6 6.39 -5.46 0.91 0.5 16-Apr-09 4 9 43 -4.2 7.21 -4.35 0.92 1.2 18-Apr-09 5 28 87 -5.3 4.59 -1.60 0.92 0.7 20-Apr-09 6 30 86 -3.6 5.63 -2.56 0.88 0.4 1-May-09 8 16 64 -- -- -1.76 -- 1.4 2-May-09 9 4 42 -0.2 4.70 -0.60 0.84 0.3 3-May-09 10 21 127 -0.6 5.04 -1.78 0.86 0.3 MEAN 10 66 -3.8 5.62 -2.44 0.89 0.99 Std. 10 23 4.1 0.88 1.11 0.03 0.83 Deviation 4232 4233 4234 4235 4236 174 4237 Table 2. Bering Sea shelf moorings: statistics and information. The location of the moorings and 4238 the years during which they were deployed are given on the left. Mixing information includes the 4239 range of time, and average date, at which the water column became well mixed with a 4240 temperature difference of < 0.02 °C. The years during which areal sea ice concentration was < 4241 20% for the entire cold season were 1996, 2001, 2003 – 2005 at M2, and 2001 and 2005 at M4. 4242 Mooring sites M5 and M8 always had ice present during the cold season. 4243 Mooring M2 56.9 °N, 164.1 °W 1995 – 2011 M4 57.9 °N, 168.9°W 1996, 19992011 M5 59.9 °N, 171.7 °W 2005 – 2011 M8 62.2 °N, 174.7 °W 2005 – 2011 Mean August Temp 2005 – 2011 (all years) Fall Water Column Mixing Minimum date and temperature Temp. 2005 – 2011 (all Date Range Temperature years) (mean) 2005 – 2011 (°C) (all years) (°C) Surface (°C) Bottom (°C) 10.1 (10.4) 1.4 (2.0) 10/21 – 12/4 (Nov. 9) 3.6 (4.1) -1.7 (-0.6) 9.4 (9.7) 1.2 (2.2) 10/18 – 11/24 (Nov. 1) 3.4 (4.4) -1.7 (-1.1) 8.9 -0.8 10/16 – 12/21 (Nov. 20) 1.7 -1.7 5.0 -1.5 12/4 – 1/25 (Dec. 19) -1.0 -1.8 4244 4245 4246 4247 4248 4249 4250 175 4251 4252 Table 3. Observations and calculations of the impact of melting ice on the water column. Tw (Sw) 4253 is the depth- 4254 the average change in temperature (salinity) in the part of the water column that was modified by 4255 melting ice. H is depth of the water column cooled (usually the entire water column), h i is the 4256 thickness of ice melted as calculated from changes in salinity, and hw is the equivalent depth of 4257 L is the change in temperature due to latent heat of fusion and the final column is 4258 the fraction of the observed cooling that is caused by the latent heat of fusion. Note that 2009 at 4259 M2 had two distinct melting events (a and b) which were combined together (2009t). 4260 Mooring Tw Sw H hw hi – year (°C) (°C) (m) (m) (m) (°C) M2 – 1998 2.0 3.4 31.95 0.45 25 0.43 0.51 1.17 0.34 2000 1.1 2.9 31.94 0.55 70 1.48 1.75 1.42 0.49 2006 2.6 4.3 32.12 0.71 27 0.73 0.86 1.82 0.43 2007 1.2 2.8 32.30 0.50 70 1.33 1.57 1.27 0.45 0.35 70 0.95 1.11 0.91 0.29 3.1 2009t L L a 2.2 2.0 32.00 0.20 70 0.54 0.63 0.52 0.26 b -0.6 1.1 31.92 0.15 70 0.41 0.48 0.39 0.35 2012 0.8 2.5 31.94 0.32 70 0.86 1.02 0.83 0.33 M4 – 2004 1.2 2.2 32.24 0.34 70 0.91 1.07 0.87 0.40 2006 2.5 3.8 32.38 0.65 70 1.72 2.03 1.65 0.43 2007 0.7 2.4 32.18 0.46 70 1.23 1.45 1.18 0.49 2009 1.6 3.5 32.11 0.45 70 1.21 1.42 1.15 0.33 2012 0.2 1.9 31.96 0.34 70 0.92 1.08 0.88 0.46 0.9 2.6 32.20 0.92 32 1.12 1.32 2.35 0.90 2007 2.0 3.6 32.20 0.50 70 1.34 1.57 1.28 0.36 2010 -0.1 1.8 31.55 0.70 70 1.92 2.25 1.83 1.00 1.15 1.36 1.33 0.48 M5 – 2006 Mean 2.9 0.52 4261 4262 4263 176 4264 Table 4. Statistics for the amount of ice that melts at moorings sites when the ice retreats. 4265 Column headings are the same as in Table 3. 4266 4267 Mooring - year H Sw hw hi M5 - 2007 0.7 25 31.5 0.69 0.81 2008 0.7 25 31.4 0.69 0.81 2010 1.3 25 31.3 1.28 1.51 2011 0.3 30 31.4 0.35 0.42 2012 0.9 25 31.4 0.89 1.04 M8 - 2006 0.2 35 32.4 0.27 0.31 2008 0.7 32 32.8 0.84 0.98 Mean 0.7 28 31.7 0.71 0.84 4268 4269 4270 4271 4272 177 4273 4274 4275 4276 4277 4278 4279 4280 4281 4282 4283 4284 4285 4286 4287 4288 4289 Papers in preparation 178 4290 4291 4292 Chapter 16 Acoustically-Determined Patterns of Summer Zooplankton Biomass and Vertical Migration 4293 Behavior from the Bering Sea Middle Shelf during a Cold Regime 4294 4295 4296 Jeffrey M. Nappa, Adam H. Spearb, Phyllis .J. Stabenoc 4297 Benjamin Blossc and David Strauszc 4298 4299 4300 a 4301 7600 Sand Point Way NE, Seattle, WA 98115, USA; Jeff.Napp@NOAA.gov; Tel:+1-206-526- 4302 4148; Fax: +1-206-526-6723 Corresponding author: NOAA-Fisheries, Alaska Fisheries Science Center, RACE Division, 4303 4304 b 4305 Seattle, WA 98115, USA; Adam.Spear@NOAA.gov NOAA-Fisheries, Alaska Fisheries Science Center, RACE Division, 7600 Sand Point Way NE, 4306 4307 c 4308 WA 98115, USA; Phyllis.Stabeno@NOAA.gov 4309 4310 4311 4312 Abstract 4313 variations in climate, and influence recruitment success of planktivorous fishes (e.g walleye 4314 pollock, Gadus chalcogramma). The broad continental shelf in this region is remote and not 4315 easily sampled by ships, particularly during cold winters and springs when sea ice covers much of 4316 the region. Two active acoustic sensors were moored over the eastern Bering Sea shelf during the 4317 summers of 2007, 2009, and 2010 and were used to gain insight into the temporal variability of 4318 summer zooplankton on this highly productive marine ecosystem. The first sensor, a 300 kHz 4319 Acoustic Doppler Current Profiler (ADCP) was intended for current measurements, sampled the 4320 entire water column, but was not calibrated for acoustic detection of small scatterers. The second 4321 sensor, a Tracor Acoustic Profiling System (TAPS) was calibrated, contained 8 unique 4322 frequencies between 100 and 3000 kHZ, but only sampled a very small volume. Volume 4323 backscatter data from the two samplers each provided indices of zooplankton biomass and 4324 distribution through different processing techniques with differing sets of assumptions. The 4325 temporal patterns of scattering by small particles varied both seasonally and among years, even NOAA-Research, Pacific Marine Environmental Laboratory, 7600 Sand Point Way NE, Seattle, Zooplankton biomass and distribution in the eastern Bering Sea are strongly connected to 179 4326 though all three years had similar environmental forcing. The backscatter volume from the ADCP 4327 and the estimated zooplankton biovolume from the TAPS were correlated. Acoustically-derived 4328 estimates of large copepod biovolume was highest in 2009 and 2010 and lowest in 2007 in 4329 agreement with other data. The diel signal dominated temporal variability and we were unable to 4330 detect additional modes of high variability at frequencies below 0.5 day-1. The TAPS-8 estimated 4331 sizes of the organisms responsible for diel vertical migration roughly matched our expectation of 4332 the identity of those organisms from nets and the literature. Length of time migrators spent in the 4333 surface waters was inversely related to the number of daylight hours and increased vulnerability 4334 to predation by age-0 fishes. This latter observation, while not new, provides an advantage to 4335 planktivores like pollock which must accumulate sufficient depot lipids by the end of summer to 4336 survive their first winter. 4337 4338 Key Words: Bering Sea, zooplankton, bioacoustics, diel vertical migration, walleye pollock, 4339 climate regimes 4340 4341 4342 4343 4344 4345 Figure 4. Eastern Bering Sea and M2 study site where the acoustic instruments collected 4346 zooplankton volume backscatter data. 4347 4348 180 4349 4350 4351 Figure 5. ADCP-derived zooplankton volume backscatter. Top – 2007, Middle – 2009, Bottom 4352 – 2010. 4353 4354 4355 181 4356 4357 4358 Chapter 17 Patterns of flow on the eastern Bering Sea shelf: 20 years of observations 4359 4360 P. J. Stabenoa and N. B. Kachelb 4361 4362 a 4363 Point Way NE, Seattle, WA 98115, USA; Phyllis.Stabeno@NOAA.gov Corresponding author: cNOAA-Research, Pacific Marine Environmental Laboratory, 7600 Sand 4364 4365 b 4366 WA 98115, USA JISAO/UW and Pacific Marine Environmental Laboratory, 7600 Sand Point Way NE, Seattle, 4367 4368 Abstract 4369 Currents are examined on the eastern Bering Sea shelf using data from moorings, 4370 satellite-tracked drifters and hydrographic surveys. The major source of water on the 4371 Bering Sea shelf is Unimak Pass and the canyons that intersect the shelf break. Absolute 4372 geostrophic transport through Unimak Pass varies from an average of 0.25 x 106 m3 s-1 4373 in the warm months to 0.43 x 106 m3 s-1 in the cold months. Flow along the 50-m isobath 4374 is weak, with a transport of < 0.1 x 106 m3 s-1 in both summer and fall. The transport 4375 along the 100-m isobath is more than twice that along 50-m isobath: 0.2 x 106 m3 s-1 in 4376 the summer at the Pribilof Islands; and a northward geostrophic transport (referenced to 4377 the bottom) of 0.31 x 106 m3 s-1 during spring and summer at 60°N. Northward transport 4378 along the 100-m and 50-m isobaths accounts for approximately half of the northward 4379 flow through Bering Strait. The transit time north from Unimak Pass to Bering Strait is 4380 greater than 13 months. Thus, the source of most of the heat transported into the Arctic 4381 through Bering Strait is a result of air-sea interactions in the Bering Sea. Examination of 4382 the currents and water properties on the southern shelf indicates that ~50% of the shelf 4383 water is exchanged with slope in October – January each year, replenishing >50% of the 4384 nutrients consumed the previous growing season. 4385 4386 Key Words: Bering Sea, currents, middle shelf, climate regimes 4387 182 Fig. 6. Mean currents during summer (May – September) derived from satellite-tracked drifters drogued at 45 m. There are at least 10 independent estimates of velocity in each grid cell. 4388 4389 4390 Fig. 7. Bottom currents at M2 divided into octents determined by wind directions (e.g. for 0 – 45 °T the vector is the average velocity when winds are toward that direction). Note currents are not 45 – 90 ° to the right of the winds (lower left). Number of points in each bin shown in lower right. 4391 183 4392 4393 4394 Fig. 8. Bottom currents at M5 divided into octents determined by wind directions (e.g. for 0 – 45 4395 °T the vector is the average velocity when winds are toward that direction). Note currents are not 4396 45 – 90 ° to the right of the winds (middle panel). Number of points in each bin shown in 4397 bottom. 4398 184 4399 4400 4401 4402 4403 Figure 9. Relationship between winds and currents at M8. In the top panels, vectors 4404 indicate the mean direction of the flow when the winds are toward the octant indicated 4405 (e.g. 270-315). The bottom panels show current direction as a function of wind direction 4406 (black line) and the number of points in each direction octant (shaded). The period 4407 examined is winter when there is no ice for a) near-surface (~12-17 m), and b) the 4408 difference between near-surface and near-bottom flow. 185 4409 4410 Conclusions Major results from moorings, accompanying CTD and the analyses that resulted from these 4411 findings. 4412 1. Almost 18 years of data from M2 were used to identify warm and cold years over the 4413 southeastern Bering Sea. There is close relationship between ice extent in March and 4414 April and depth averaged temperature at M2, which permitted the extension of the 4415 definition of warm and cold back to 1972. 4416 2. Prior to 2000, the amount of ice in March and April was highly variable, but beginning 4417 in spring 2000, the SE Bering Sea enter 5.5 year window of low ice extent and warm 4418 conditions. This was followed by 8+ year period (2007 – present) more extensive ice in 4419 March and April and very cold conditions on the southern shelf. 4420 3. The types and abundance of zooplankton differed sharply between warm and cold years. 4421 This was especially true during the prolonged warm period (2001–2005) and cold period 4422 (2007–2010), and was less evident during the years of high interannual variability. 4423 During the warm period, there was a lack of large copepods and euphausiids over the 4424 shelf, however, their populations rebounded during cold period. Small crustacean 4425 zooplankton taxa did not appear to vary between and warm and cold years. 4426 4. Contrary to historical views, there is a sharp division between the northern and southern 4427 Bering Sea shelves which occurs at ~60 °N. These differences include tidal strength, 4428 sharpness of the interface, cause of vertical stratification, presence of subsurface 4429 chlorphyll maxima, and influence of ice melt. These are discussed below. 4430 5. The ocean currents differed between warm and cold years. During cold years, the 4431 monthly-mean currents over the southeastern shelf were largely westward, while in 4432 warm years the direction of currents was more variable, with northward flow during 4433 December–February and relatively weak flow during the remainder of the year. 4434 6. Unlike the more coastal domain on the northern shelf where winds dominate the flow, 4435 the flow on the southern middle shelf is more complex. During the winter, the bottom 4436 currents at M2 are not always couple to the wind,. This could have important 4437 implication for transport of plankton, such as snow crabs larvae. 4438 7. Temperature and salinity both influence the stratification of the Eastern Bering Sea 4439 shelf: temperature dominates stratification on the southern shelf, and temperature and 4440 salinity are equally important on the northern shelf. The strength of stratification does 4441 not vary as a function of warm or cold years. 4442 8. Approximately, 1 m of ice melts over the southern shelf during the ice advance, 186 4443 modifying the salinity. A similar amount melts over the northern shelf with ice retreat. 4444 Ice cover inhibits the vertical mixing of the water column. Approximately 50% of the 4445 cooling of the southern shelf is caused by ice melt, with the remainder a result of air-sea 4446 interaction. 4447 9. Chlorophyll a fluorescence data from the four biophysical moorings provided 37 4448 realizations of a spring bloom and 33 realizations of a fall bloom. Basic findings: If 4449 ice was present after mid-March, spring bloom timing was related to ice retreat 4450 timing; if ice was absent or retreated before mid-March, a spring bloom usually 4451 occurred in May or early June. A fall bloom also commonly occurred, usually in late 4452 September, and its timing was not significantly related to the timing of storms or fall 4453 water column overturn. The magnitudes of the spring and fall blooms were correlated. 4454 10. The breadth of the mooring instrumentation from thermistors, conductivity cells, 4455 current meters and ADCPs, nitrate sensors, fluorometers, TAPS-8 to measure 4456 zooplankton biovolume, and acoustic systems to listen for marine mammals. Passive 4457 acoustic data provided information about sea ice conditions and detected and 4458 identified of vocalizing marine mammals; the acoustic backscatter provided 4459 information on relative zooplankton and fish abundance before, during, and after the 4460 retreat; and the hydrographic data confirmed the acoustic signal was associated with 4461 changing surface ice conditions. The combined information from the biophysical 4462 mooring sensors revealed changes in winter trophic level dynamics during the ice 4463 retreat, which would have otherwise been undetected by traditional ship-based 4464 observations. 4465 11. The nitrate sensors and nutrient measurements around the moorings provided 4466 information of the seasonal draw down of nutrients associated with primary production, 4467 and winter replenishment. Nutrients are typically replenished on the shelf from 4468 November – January. Summer storms result in mixing of nutrients into the photic zone 4469 and a phytoplankton bloom 5 – 7 days later. 4470 12. Subsurface blooms in the region around M8 are common during the summer. 4471 13. The biophysical mooring data were frequently used to calibrate biophysical models of 4472 the Bering Sea. These included side-by-side model/data comparisons of the vertical 4473 profiles of currents, temperature, salinity, and chlorophyll-a at multiple locations, from 4474 hourly to interdecadal time scales. 4475 4476 BSIERP and Bering Sea Project Connections 187 4477 We found this to be a highly successful program. Collaborations among groups 4478 was extensive and the sharing of data and resources strongly to contributed to the 4479 success of this program. By sharing ship time each research group gained a first- 4480 hand understanding of the goals, needs and problems of other groups. The PI 4481 meetings were comprehensive and productive. The results of these collaborations 4482 are seen in the papers published and the fact that individuals from variety of groups 4483 co-author manuscripts. The success in the program is evident in the series of 4484 special issues that have come out of it. 4485 Ship time is always a problem in high latitudes. Participation in this program 4486 provided us the opportunity to collect data during the winter/spring when the 4487 Bering Sea is covered in ice. In turn, data and samples were collected on our late 4488 summer cruises, which extended the sampling period into late summer. This 4489 benefited the other scientists of this program. 4490 EcoFOCI had extensive physical, chemical and biological data (moorings and 4491 shipboard measurements) collected during the warm period. These data provided 4492 the ability to make critical comparisons between warm period (2001 – 2005) and 4493 the cold period sampled by Bering Sea Project. In addition, the long time series at 4494 the moorings (M2, M4, M5 and M8) provided context for many of the other 4495 measurements made as part of the Bering Sea Project. 4496 This is the best large integrated program that we have participated in. 4497 4498 188 4499 Management or policy implications 4500 Our long-term biophysical mooring array provides critical and highly cited measurements of the 4501 physical drivers of the southeastern Bering Sea ecosystem. These measurements are used by the 4502 North Pacific Fishery Management Council (Fig. 10) to inform stock assessments and guide total 4503 allowable catch for some of the most economically important fisheries in the U.S. For example, 4504 during a recent warm phase in the Bering Sea (2000 – 2005) pollock recruitment was extremely 4505 poor and assessments of pollock stocks and environmental conditions warned of possible 4506 continued declines. 4507 ecosystem considerations advice by changing existing management practices – reducing pollock 4508 harvest levels. This occurred during a period when the pollock population was facing a variety of 4509 environmental stressors, and potentially averted a collapse of the fish stock and the fisheries, 4510 communities and economies that depend on it. Resource managers responded to the combined stock assessment and 4511 4512 4513 4514 4515 Figure 10. Schematic of the pathway for management action for walleye pollock. 4516 4517 189 4518 4519 4520 4521 Publications 4522 Jeffrey M. Napp, J. M., A. H. Spear, P. .J. Stabeno, B. Bloss and D. Strausz, Acoustically- In Preparation 4523 Determined Patterns of Summer Zooplankton Biomass and Vertical Migration Behavior 4524 from the Bering Sea Middle Shelf during a Cold Regime, For submission to the fourth 4525 special issue. 4526 4527 4528 Stabeno, P. J. and N. B. Kachel, Patterns of flow on the eastern Bering Sea shelf: 20 years of data. For submission to the fourth special issue. 4529 4530 Submitted/in press 4531 4532 Cheng, W., E. Curchitser, C. Ladd, and P.J. Stabeno (2013): Ice–ocean interactions in the eastern 4533 Bering Sea: NCAR CESM simulations and comparison with observations. Deep-Sea Res. II. 4534 4535 Cross, J.N., J.T. Mathis, M.W. Lomas, S.B. Moran, J. Grebmeier, D. Shull, C.W. Mordy, P.J. 4536 Stabeno, R. Sambrotto, R. Gradinger, L. Juranek, M. Prokopenko, M.S. Baumann, and M. 4537 Ostendorf (2013): Integrated assessment of the carbon budget in the Southeastern Bering 4538 Sea. Deep-Sea Res. II. 4539 4540 Goes, J.I., H. Rosario Gomes, E. D'Sa, D. Stoecker, C.W. Mordy, R. Sambrotto, and P.J. Stabeno 4541 (2013): Distribution of phytoplankton communities in the Bering Sea during the summer of 4542 2008 that followed a cold spring. Deep-Sea Res. II. 4543 4544 Napp, J.M., R.D. Brodeur, D. Demer, R. Hewitt, P.J. Stabeno, and G.L. Hunt, Jr. (2008): 4545 Acoustic determination of nekton and zooplankton distributions at tidally generated shelf 4546 fronts in the eastern Bering Sea. Mar. Ecol. Prog. Ser. 4547 4548 Prokopenko, M.G., J. Granger, M. Long, C. Mordy, E. Barkan, N. Cassar, E. Cokelet, P. DiFiore, 4549 C. Ladd, R. Sambrotto, and B. Moran (2013): Mixed layer depth controls the instantaneous 4550 Net to Gross Production ratios and potential export efficiency in spring blooms on the 4551 eastern Bering Sea shelf. Deep-Sea Res. II. 4552 190 4553 Sigler, M.F., P.J. Stabeno, L.B. Eisner, and J.M. Napp (2012): Spring and fall phytoplankton 4554 blooms in a productive subarctic ecosystem, the eastern Bering Sea, during 1995-2011. 4555 Deep-Sea Res. II. 4556 4557 Stauffer, B., J.I. Goes, H. Rosario Gomes, K. McGee, and P.J. Stabeno (2013): Interannual 4558 variability of spring-time phytoplankton community composition across the continental shelf 4559 of the southeastern Bering Sea. Deep-Sea Res. II. 4560 4561 4562 Sullivan, M., C. Mordy, S. Salo, and P.J. Stabeno (2013): The influence of sea ice on seasonal water column changes on the eastern Bering Sea shelf. Deep-Sea Res. II. 4563 4564 Published 4565 4566 Cooper, L.W., M. Sexson, J.M. Grebmeier, R. Gradinger, C.W. Mordy, and J.R. Lovvorn (2013): 4567 Linkages between sea-ice coverage, pelagic-benthic coupling, and the distribution of 4568 spectacled eiders: Observations in March 2008, 2009 and 2010, Northern Bering Sea. Deep- 4569 Sea Res. II, doi: 10.1016/j.dsr2.2013.03.009. 4570 4571 Coyle, K.O., A. I. Pinchuk, L. B. Eisner, J. M. Napp (2008). Zooplankton species composition, 4572 abundance and biomass on the eastern Bering Sea shelf during summer: The potential role of 4573 water-column stability and nutrients in structuring the zooplankton community. Deep-Sea 4574 Res. II. doi:10.1016/j.dsr2.2008.04.029. 4575 4576 Danielson, S., E. Curchitser, K. Hedstrom, T. Weingartner, and P. Stabeno (2011): On ocean and 4577 sea ice modes of variability in the Bering Sea. J. Geophys. Res., 116(C12), C12034, doi: 4578 10.1029/2011JC007389. 4579 4580 Granger, J, M.G. Prokopenko, C.W. Mordy, and D.M. Sigman (2013): The proportion of 4581 remineralized nitrate on the ice-covered eastern Bering Sea shelf evidenced from the oxygen 4582 isotope ratio of nitrate. Global Biogeochem. Cycles, doi: 10.1002/gbc.20075. 4583 4584 Hermann, A.J., G.A. Gibson, N.A. Bond, E.N. Curchitser, K. Hedstrom, W. Cheng, M. Wang, 4585 P.J. Stabeno, L. Eisner, and K.D. Cieciel (2013): A multivariate analysis of observed and 191 4586 modeled biophysical variability on the Bering Sea shelf: Multidecadal hindcasts (1970- 4587 2009) and forecasts (2010-2040). Deep-Sea Res. II, doi: 10.1016/j.dsr2.2013.04.007. 4588 4589 Hunt, Jr., G.L., B.M. Allen, R.P. Angliss, T. Baker, N. Bond, G. Buck, G.V. Byrd, K.O. Coyle, 4590 A. Devol, D.M. Eggers, L. Eisner, R.A. Feely, S. Fitzgerald, L.W. Fritz, E.V. Gritsay, C. 4591 Ladd, W. Lewis, J. Mathis, C.W. Mordy, F. Mueter, J. Napp, E. Sherr, D. Shull, P. Stabeno, 4592 M.A. Stepanenko, S. Strom, and T.E. Whitledge (2010): Status and trends of the Bering Sea 4593 region, 2003-2008. In Marine Ecosystems of the North Pacific Ocean, 2003-2008, S. M. 4594 McKinnell and M. J. Dagg (eds.), PICES Special Publication 4, 196-267. 4595 4596 Hunt, Jr., G. L., K.O. Coyle, L. Eisner, E.V. Farley, R. Heintz, F. Mueter, J.M. Napp, J. E. 4597 Overland, P.H. Ressler, S. Salo, and P. J. Stabeno (2011): Climate impacts on eastern Bering 4598 Sea foodwebs: A synthesis of new data and an assessment of the Oscillating Control 4599 Hypothesis. ICES J. Mar. Sci., 68(6), doi: 10.1093/icesjms/fsr036, 1230–1243. 4600 4601 Hunt, Jr., G.L., P.J. Stabeno, S. Strom, and J.M. Napp (2008): Patterns of spatial and temporal 4602 variation in the marine ecosystem of the southeastern Bering Sea, with special reference to 4603 the Pribilof Domain. Deep-Sea Res. II, 55(16–17), doi: 10.1016/j.dsr2.2008.04.032, 1919– 4604 1944. 4605 4606 4607 Ladd, C., and P.J. Stabeno (2009): Freshwater transport from the Pacific to the Bering Sea through Amukta Pass. Geophys. Res. Lett., 36, L14608, doi: 10.1029/2009GL039095. 4608 4609 4610 Ladd, C., and P.J. Stabeno (2012): Stratification on the Eastern Bering Sea Shelf revisited. DeepSea Res. II, 65–70, doi: 10.1016/j.dsr2.2012.02.009, 72–83. 4611 4612 4613 Ladd, C., P.J. Stabeno, and J.E. O'Hern (2012): Observations of a Pribilof eddy. Deep-Sea Res. I, 66, doi: 10.1016/j.dsr.2012.04.003, 67–76. 4614 4615 Mathis, J.T., J.N. Cross, N.R. Bates, S.B. Moran, M.W. Lomas, C.W. Mordy, and P.J. Stabeno 4616 (2010): Seasonal distribution of dissolved inorganic carbon and net community production 4617 on the Bering Sea shelf. Biogeosciences, 7, doi: 10.5194/bg-7-1769-2010, 1769–1787. 4618 192 4619 Mathis, J.T., J.N. Cross, N. Monacci, R.A. Feely, and P.J. Stabeno (2013): Evidence of prolonged 4620 aragonite undersaturations in the bottom waters of the southern Bering Sea shelf from 4621 autonomous sensors. Deep-Sea Res. II, doi: 10.1016/j.dsr2.2013.07.019. 4622 4623 Miksis-Olds, J.L., P.J. Stabeno, J.M. Napp, A.I. Pinchuk, J.A. Nystuen, J.D. Warren, and S.L. 4624 Denes (2013): Ecosystem response to a temporary sea ice retreat in the Bering Sea: Winter 4625 2009. Prog. Oceanogr., 111, doi: 10.1016/j.pocean.2012.10.010, 38–51. 4626 4627 Mordy, C.W., E.D. Cokelet, C. Ladd, F.A. Menzia, P. Proctor, P.J. Stabeno, and E. Wisegarver 4628 (2012): Net community production on the middle shelf of the Eastern Bering Sea. Deep-Sea 4629 Res. II, 65–70, doi: 10.1016/j.dsr2.2012.02.012, 110–125. 4630 4631 Mordy, C.W., L. Eisner, P. Proctor, P.J. Stabeno, A. Devol, D.H. Shull, J.M. Napp, and T. 4632 Whitledge (2010): Temporary uncoupling of the marine nitrogen cycle: Accumulation of 4633 nitrite on the Bering Sea Shelf. Mar. Chem., 121, doi: 10.1016/j.marchem.2010.04.004, 4634 157–166. 4635 4636 Nystuen, J.A., S.E. Moore, and P.J. Stabeno (2010): A sound budget for the southeastern Bering 4637 Sea: Measuring wind, rainfall, shipping, and other sources of underwater sound. J. Acoust. 4638 Soc. Am., 128(1), doi: 10.1121/1.3436547, 58−65. 4639 4640 Sigler, M.F., H.R. Harvey, C.J. Ashjian, M.W. Lomas, J.M. Napp, P.J. Stabeno, and T.I. Van Pelt 4641 (2010): How does climate change affect the Bering Sea ecosystem? Eos Trans. AGU, 4642 91(48), doi: 10.1029/2010EO480001, 457–458. 4643 4644 Stabeno, P.J., E. Farley, N. Kachel, S. Moore, C. Mordy, J.M. Napp, J.E. Overland, A.I. Pinchuk, 4645 and M.F. Sigler (2012): A comparison of the physics of the northern and southern shelves of 4646 the eastern Bering Sea and some implications for the ecosystem. Deep-Sea Res. II, 65–70, 4647 doi: 10.1016/j.dsr2.2012.02.019, 14–30. 4648 4649 Stabeno, P.J., N.B. Kachel, S.E. Moore, J.M. Napp, M. Sigler, A. Yamaguchi, and A.N. Zerbini 4650 (2012): Comparison of warm and cold years on the southeastern Bering Sea shelf and some 4651 implications for the ecosystem. Deep-Sea Res. II, 65–70, doi: 10.1016/j.dsr2.2012.02.020, 4652 31–45. 193 4653 4654 Stabeno, P.J., J. Napp, C. Mordy, and T. Whitledge (2010): Factors influencing physical structure 4655 and lower trophic levels of the eastern Bering Sea shelf in 2005: Sea ice, tides and winds. 4656 Prog. Oceanogr., 85(3–4), doi: 10.1016/j.pocean.2010.02.010, 180–196. 4657 4658 Stafford, K.M., S.E. Moore, P.J. Stabeno, D.V. Holliday, J.M. Napp, and D.K. Mellinger (2010): 4659 Biophysical ocean observation in the southeastern Bering Sea. Geophys. Res. Lett., 37, 4660 L02606, doi: 10.1029/2009GL040724. 4661 4662 4663 Outreach 4664 Web-page 4665 EcoFOCI maintains a suite of NPRB related web sites that can found at 4666 http://www.pmel.noaa.gov/foci/. 4667 4668 Schools, mentoring, radio and press 4669 Carol Ladd made a presentation to four 5thgrade classrooms at Sunset Elementary, Issaquah 4670 4671 4672 4673 School District, about the Bering Sea Ice Expedition 2008. May 22, 2008. Ned Cokelet contributed to a public presentation after the spring BEST cruise in May 2009 and discussed with the public goals and findings of the research. Members of PMEL/Eco-FOCI collaborated with Aleut Community of St. Paul Island in 4674 maintaining their temperature and salinity measurements in their harbor and on St. George 4675 Island from 2009-2012. 4676 4677 4678 4679 4680 4681 4682 4683 4684 4685 Nancy and Dave Kachel gave Science Day Presentations about BEST/BSIERP at Jane Adams K8 School, Seattle, WA. December 8, 2010. Nancy and Dave Kachel gave Science Day Presentations about BEST/BSIERP at Jane Adams K8 School, Seattle, November 30, 2011. Lisa Guy, Calvin Mordy and Scott McKeever. Satellite-Tracked Drifters, Polar Science Weekend at the Pacific Science Center, March 1-2, 2012. Seattle, WA. Scott McKeever. Volunteered as a judge at the Orca Bowl, School of Oceanography, University of Washington, March, 2012. Nancy and Dave Kachel participated as Science Fair Judges at Jane Adams K-8 School, Seattle, WA. April 2012. 194 4686 4687 4688 Margaret Sullivan, Speed Networking, Young Leadership Program/NOAA Science Camp, Seattle, WA. July 2012. Guy, Lisa. Press Release titled "New Bering Sea research reveals how changing ecosystem 4689 impacts America's most valuable fisheries" Link to article: 4690 http://www.noaanews.noaa.gov/stories2012/20120530_beringsea.html 4691 4692 Guy,Lisa N.Kachel and Scott McKeever provided outreach to Sea Grant Fellow Kristen Jabanoski via in-person discussion, slide presentation and a tour on 29 January 2013. 4693 Nancy and Dave Kachel gave Science Day Presentations about BEST/BSIERP at Jane Adams K- 4694 8 School, Seattle, Kachel, N. Science Day Presentations at Jane Adams K-8 School, Seattle, 4695 WA. Feb 6, 2013. 4696 Margaret Sullivan provided EcoFOCI animations to Danielle Dickson (NPRB) for a presentation. 4697 The animation shows Bering Sea satellite ice (Modis true-color) with ice-floe tracking (dots) 4698 for 2003 (warm year, less ice) and 2007 (cold year, more ice, 20 Feb 2013. 4699 4700 4701 4702 4703 4704 4705 Scott McKeever. Volunteered as a judge at the Orca Bowl, School of Oceanography, University of Washington, February 23, 2013. Nancy and Dave Kachel participated as Science Fair Judges at Jane Adams K-8 School, Seattle, WA. April 2013. Nancy Kachel and Jeff Napp made presentations and Scott McKeever gave a facilities tour for undergraduate Seattle University Marine Science class on 22 May 2013. Carol Ladd, is a mentor for the MPOWIR program, providing mentoring to physical 4706 oceanographers from late graduate school through their early careers. The overall goal of 4707 MPOWIR is to make mentoring opportunities for junior physical oceanographers universally 4708 available and of higher quality by expanding the reach beyond individual home 4709 institutions. The aim is to reduce the barriers to career development for all junior scientists 4710 in the field, with a particular focus on improving the retention of junior women. 4711 Carol Ladd, Hosted a high school senior (Zach Hall) to discuss a career in science (6/28/2012). 4712 Jeff Napp and Phyllis Stabeno both served on the Science Advisory Board. 4713 4714 Conference Presentations 4715 Stabeno, P., J. Napp, C. Mordy and T. Whitledge, Southeast Bering Sea shelf: a contrast between 4716 a warm year and a cool year. Alaska Marine Science Symposium, Anchorage Alaska. 4717 January 2008. 4718 4719 Ladd, C. North Pacific meets the Bering Sea: transport through Amukta Pass. Alaska Marine Science Symposium, Anchorage Alaska. January 2008. 195 4720 4721 Stabeno, P. and J. Napp. EcoFOCI: The Bering Sea ecosystem. PMEL Site Review. Seattle, Washington. August 2008. 4722 Stabeno, P.J., J.M.Napp, T.E.Whitledge, and C. Mordy. The influence of seasonal sea ice on the 4723 eastern Bering Sea shelf ecosystem. AGU / ASLO Ocean Sciences Meeting, Orlando, 4724 Florida, February 2008. 4725 Napp, J. and P. Stabeno. Bottom-up control of eastern Bering Sea ecosystem: implications for 4726 integrated ecosystem assessment during a period of climate change. Library Lunch Seminar. 4727 Washington DC. 4728 4729 4730 4731 4732 Napp, J. The Bering Sea: Current Status and Recent Events. PICES Press, 16 (1), January 2008. p. 31- 33. Napp, J. The Bering Sea: Current Status and Recent Events. PICES Press, 16 (2), July 2008. p. 30-31. Stabeno, P. J., N. Kachel, C. Mordy, D. Moore and J. Napp. Bering Sea Observing Network: 4733 Physics to Whales. Poster presented at OceanObs09, Venice, Italy, September 2009. 4734 Buckley, T.W., TenBrink, T., Napp, J. and Aydin, K. “Comparing zooplankton consumed by 4735 walleye pollock and caught by nets in the eastern Bering Sea.” AGU/ASLO Ocean Sciences 4736 Meeting, Portland, OR, February 2010. 4737 Cross, J. N.; J. T. Mathis; N. R. Bates; B. Moran; M. W. Lomas; P. J. Stabeno Seasonal 4738 Distribution and Controls on Dissolved Inorganic Carbon and Net Community Production 4739 on the Eastern Bering Sea Shelf. 2010 Ocean Science Meeting Portland OR 4740 Hermann, A. J.; K. Aydin; N. A. Bond; W. Cheng; E. N. Curchitser; G. A. Gibson; K. Hedstrom; 4741 I. Ortiz; M. Wang; P. J. Stabeno The Bering Sea Integrated Ecosystem Research Program: 4742 downscaling climate change to a subarctic region with coupled biophysical models. 2010 4743 Ocean Science Meeting Portland OR 4744 4745 Kachel, N. B.; C. W. Mordy; P. J. Stabeno Comparison of four decades of hydrographic structure over the SE Bering Sea Shelf. 2010 Ocean Science Meeting Portland OR 4746 Mordy, C. W.; C. Ladd; M. W. Lomas; R. Sambrotto; P. J. Stabeno. The Annual Nutrient Cycle 4747 on the Bering Sea Shelf: Seasonal Drawdown vs. Winter Replenishment. 2010 Ocean 4748 Science Meeting, Portland OR 4749 Napp, J.M., Eisner, L.B., Farley, E.W., Stabeno, P.J., Hunt, Jr., G.L. Recent changes in 4750 zooplankton abundance and biomass in the eastern Bering Sea, Alaska Marine Science 4751 Symposium, Anchorage, AK, January 2010. 4752 4753 Napp, J.M., Sigler, M.F., and Stabeno, P.J. Understanding ecosystem processes in the Bering Sea. NOAA Headquarters Lunch Seminar, Silver Spring, MD, March 2010. 196 4754 4755 Salo, S. A and P. J. Stabeno, Spatial Scales of Correlation on the Bering Sea Middle Shelf, 2010 Ocean Science Meeting Portland OR 4756 Stabeno, P. J.; N. B. Kachel; C. Mordy; J. M. Napp; M. C. Spillane, The north - south structure of 4757 eastern Bering Sea shelf ? Implications for a changing ecosystem. 2010 Ocean Science 4758 Meeting Portland OR 4759 Stabeno, P.J. N. A. Bond, C. M. Mordy, and J. M. Napp. Comparison between a cold year (2009) 4760 and a warm year (2005) and implications for the ecosystem on response to changes in 4761 climate, Alaska Marine Science Symposium, Cook Hotel, Anchorage, Alaska. 4762 4763 4764 K. Stafford; S. E. Moore; P. J. Stabeno; V. Holliday; J. M. Napp. Biophysical Ocean Observation in the Southeastern Bering Sea. 2010 Ocean Science Meeting Portland OR Sullivan, M. E.; C. W. Mordy; P. J. Stabeno; N. B. Kachel; D. G. Kachel; S. A. Salo A Brief 4765 History of an Ecosystem Spring Setup as Seen in Annual Ice Cover. 2010 Ocean Science 4766 Meeting Portland OR. 4767 Hunt, G.L. Jr., Eisner, L.B., Farley, E.W., Moss, J.H., Napp, J.M., and Stabeno, P.J. “The 4768 oscillating control hypothesis: Reassessment in view of new information from the eastern 4769 Bering Sea.” PICES/ICES Climate Change Effects on Fish and Fisheries, Sendai Japan, 4770 April, 2010. 4771 Napp, J.M., Eisner, L.B., Farley, E.W., Stabeno, P.J., Hunt, Jr., G.L. “Recent changes in 4772 zooplankton abundance and biomass in the eastern Bering Sea.” Alaska Marine Science 4773 Symposium, Anchorage, AK, January 2010. 4774 4775 4776 Napp, J.M., Sigler, M., and Stabeno, P.J. ”Understanding ecosystem processes in the Bering Sea.” NOAA Headquarters, Silver Spring, MD, March 2010. Napp, J.M., Hunt, Jr., G.L., Eisner, L.B., Farley, E.W., Stabeno, P.J., Andrews, A., and 4777 Yamaguchi, A. The response of eastern Bering Sea zooplankton communities to climate 4778 fluctuations: Community structure, biodiversity, and energy flow to higher trophic levels. 4779 PICES/ICES Climate Change Effects on Fish and Fisheries, Sendai Japan, April, 2010. 4780 Stabeno, P.J., Bond, N.A., Napp, J.M., and Mordy, C.W. “Comparison between a cold year 4781 (2009) and a warm year (2005) and implications for the ecosystem on response to changes in 4782 climate. Alaska Marine Science Symposium, Anchorage, AK, January 2010. 4783 Bond, N.A. “Climate and modeling the Bering Sea Ecosystem”. Bering Sea Ecosystem 4784 4785 4786 4787 Professional Development Workshop, Anchorage, Alaska. 11-15 October 2010. Bond, N.A., Stabeno, P.J., Hermann, A.J., and M. Wang. “What controls the extent of Bering Sea Ice in spring?” PICES Annual Meeting, Portland, Oregon. 25-29 October 2010. Hermann, A.J., Aydin, K., Bond, N.A., Cheng, W., Curchitser, E.N., Gibson, G.A., Hedstrom, K., 197 4788 Ortiz, I., Wang, M., and Stabeno, P.J. “Modes of biophysical variability on the Bering Sea 4789 shelf”. PICES Annual Meeting, Portland, Oregon. 25-29 October 2010. 4790 Hinckley, S., Burgos, J., Szuwalski, C., Parada, C., Orensanz, J.M., Punt, A., Armstrong, D., 4791 Ernst, B., Hermann, A., Megrey, B., and J. Napp. Can we use a coupled biophysical IBM 4792 model experiment to predict recruitment of snow crab in the Bering Sea – The Gate 4793 Hypothesis. Alaska Marine Science Symposium, Anchorage, AK, January 2011. 4794 Hunt, G.L. Jr., Coyle, K.O., Eisner, L., Farley, E.V., Heintz, R., Mueter, F., Napp, J.M., 4795 Overland, J.E., Ressler, P.H., Salo, S., and P.J. Stabeno. “Climate impacts on eastern Bering 4796 Sea food webs: A synthesis of new data and an assessment of the Oscillating Control 4797 Hypothesis. PICES Annual Meeting, Portland, Oregon. 25-29 October 2010. 4798 4799 4800 4801 4802 Ladd, C., Stabeno, P., and O’Hern, J. “The Pribilof eddy in the eastern Bering Sea”. PICES Annual Meeting, Portland, Oregon. 25-29 October 2010. Ladd, C., Stabeno, P.J., and J. O’Hern. “The Pribilof eddy in the eastern Bering Sea”. Alaska Marine Science Symposium, Anchorage, AK, January 2011. Mordy, C., Ladd, C., Menzia, F.A., Nabor, D., Proctor, P., Stabeno, P., Whitledge, T., and E. 4803 Wisegarver. Temporal and spatial variability of primary production on the eastern Bering 4804 Sea Shelf. Alaska Marine Science Symposium, Anchorage, AK, January 2011. 4805 Napp, J.M., Ashjian, C., Harvey, R., Lomas, M., Sigler, M., and Stabeno, P.J. ”Understanding 4806 ecosystem processes in the Bering Sea.” PICES Annual Meeting, Portland, Oregon. 25-29 4807 October 2010. 4808 Stabeno, P.J., Bond, N.A., and J.M. Napp. “Eastern Bering Sea shelf: Comparison between a cold 4809 period (2007-2010) and a warm period (2001-2005). PICES Annual Meeting, Portland, 4810 Oregon. 25-29 October 2010. 4811 4812 4813 Stabeno, P.J. and J.M. Napp. A continuation of cold conditions in the Bering Sea: 2010. Alaska Marine Science Symposium, Anchorage, AK, January 2011. Wang, M., Overland, J.E., and P. Stabeno. Examples of using global climate models for Bering 4814 Sea marine ecosystem projection. Alaska Marine Science Symposium, Anchorage, AK, 4815 January 2011. 4816 Napp, J.M., Eisner, L.B., Farley, E.V., Mier, K.L., Pinchuk, A.I., and Stabeno, P.J. ”North-south 4817 variation in eastern Bering Sea shelf spring and late summer zooplankton assemblage.” 4818 ESSAS Annual Science Meeting, Seattle, WA, May 2011 4819 4820 4821 Napp, J.M., Stabeno, P.J., and Sigler, M.F. “From climate to whales: Components of an Integrated Ecosystem Assessment.” NOAA Headquarters, Silver Spring, MD, June 2011 Stabeno, P.J., Napp, J.M., and Sigler, M.F. “North Pacific Climate Regimes and Ecosystem 198 4822 Productivity (NPCREP): Ecosystem-based management in the Bering Sea.” Adaptations 4823 2011 Workshop, Invited Speakers, Washington, D.C., June 2011. 4824 4825 4826 4827 Napp, J.M., and the BEST/BSIERP PIs. “Integrated ecosystem research in the Bering Sea.” AFS Annual Meeting, Seattle, WA, September 2011. Stabeno, P., J. Napp, and N. Bond. The Status Of The Bering Sea In 2011: Cool. Alaska Marine Science Symposium. January 16 – 19, Anchorage, AK. 4828 Hermann, A., Gibson, Bond, Cuchitser, Hedstrom, Chen, Want, Stabeno, Eisner, Janout, Aydin, 4829 A multivariate analysis of observed and modeled biophysical variability on the Bering Sea 4830 shelf: multidecadal hindcasts (1969-2009) and forecasts (2010-2040), Alaska Marine 4831 Science Symposium. January 16 – 19, Anchorage, AK. 4832 P. Stabeno and P. Ressler. Advection. Bering Sea Synthesis. February 6 -9, 2012, Bermuda. 4833 Sullivan, P., S. Salo, P. Stabeno, C. Mordy. Spring Ice and Salt Flux in the Eastern Bering Sea 4834 Marginal Ice Zone. 2012 Ocean Sciences Meeting, February, 2012, Salt Lake City, UT. 4835 P. Stabeno. Physics, Bering Sea PI Meeting. March 28 -30, 2012, Anchorage, AK. 4836 Stabeno, Danielson. Physics of the Bering Sea. Bering Sea Project PI meeting. Anchorage, AK. 4837 March, 2012. Oral Presentation 4838 Stabeno. EcoFOCI. Presentation to head of OAR. Seattle, WA, April 2012. Oral Presentation 4839 Stabeno and Napp. 4840 4841 4842 4843 The Bering Sea. EcoFOCI Retreat. Seattle WA, June 2012. Oral Presentation Stabeno and Overland. The New Normal in the U. S. Arctic. NOAA Senior Research Council. Seattle, WA, July, 2012. Oral Presentation Stabeno, Farley, Kachel, Moore, Mordy, Napp, Overland, Pinchuk and Sigler. Climate-mediated 4844 processes on the northern and southern shelves of the eastern Bering Sea and some 4845 implications for the ecosystem. Yeosu, Korea, May 2012. Oral Presentation 4846 4847 Reports 4848 Napp, Jeffrey. The Bering Sea: Current Status and Recent Events. PICES Press Volume 20 4849 Number 2. July 2012. 4850 4851 Napp, Jeffrey. The Bering Sea: Current Status and Recent Events. PICES Press Volume 20 4852 Number 1. January 2012. 4853 4854 Napp, Jeffrey. The Bering Sea: Current Status and Recent Events. PICES Press Volume 19 4855 Number 2. July 2011. 199 4856 4857 Napp, Jeffrey. The Bering Sea: Current Status and Recent Events. PICES Press Volume 18 4858 Number 2. July 2010. 4859 4860 Napp, Jeffrey. The Bering Sea: Current Status and Recent Events. PICES Press Volume 18 4861 Number 1. January 2010. 4862 4863 Napp, Jeffrey. Current Status of the Bering Sea Ecosystem. PICES Press Volume 17 Number 2. 4864 July 2009. 4865 4866 Napp, Jeffrey. The Bering Sea: Current Status and Recent Events. PICES Press Volume 17 4867 Number 1. January 2009. 4868 4869 Napp, Jeffrey. The Bering Sea: Current Status and Recent Events. PICES Press Volume 16 4870 Number 2. July 2008. 4871 4872 Napp, Jeffrey. The Bering Sea: Current Status and Recent Events. PICES Press Volume 16 4873 Number 1. January 2008 4874 4875 Acknowledgements 4876 Special thanks goes to the Program Manager for the Bering Sea Project, Thomas Van Pelt, for his 4877 hard work, insight and support to this program. We thank all the scientists from EcoFOCI at both 4878 Alaska Fisheries Science Center and the Pacific Marine Environmental Laboratory for their hard 4879 work at sea, in processing data and in assisting in analysis. We thank the officers and crews of the 4880 NOAA ships Miller Freeman and Oscar Dyson, R/V Thomas G. Thompson, R/V Knorr and 4881 USCG Healy for invaluable assistance in deploying and recovery of the moorings, and collecting 4882 oceanographic measurements. Special thanks goes to NOAA’s Alaska Fisheries Science Center’s 4883 Resource Assessment and Conservation Engineering Division Groundfish and Shellfish 4884 Assessment Program who shared shiptime during particularly icy years to deploy moorings at M5 4885 and M8 on several occasions. NOAA’s North Pacific Climate Regimes and Ecosystem 4886 Productivity and Fisheries Oceanography Coordinated Investigations provided support including 4887 equipment, salaries and shiptime that helps to maintain these long-term moorings. 4888 4889 200