Foraging Habitat of Sepia officinalis at STARESO Research Station
advertisement

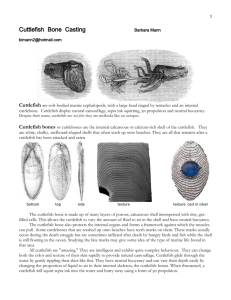
Foraging Habitat of Sepia officinalis at STARESO Research Station in Calvi, Corsica, France. Colin Gaylord and Tyler Hubbell ABSTRACT The goal of our observational field study was to discover if the cryptic species Sepia officinalis displays a habitat association while foraging at night. We predicted that S. officinalis would show a strong preference for cobble and boulders covered in turfy algae in the coves in and around STARESO research station. To test our hypothesis, we compared the available habitat to where we located Sepia officinalis during night sampling. Our study found that Sepia does show an association to at least the turfy algae. This association is potentially due to Sepia officinalis’s mimetic abilities that may enhance its foraging abilities and reduce the risk of predation. INTRODUCTION It is widely recognized that cephalopods represent the pinnacle of invertebrate evolution. Animals in this class have adapted incredibly complex sensory systems and have been shown to possess intelligence unrivaled among other invertebrates. A centralized nervous system, including a relatively large, lobed brain found in numerous cephalopods, allow for fined tuned interpretation and rapid response in regards to certain environmental conditions (Boycott, 1961). This advanced centralized nervous system provides the basis for several physiological characteristics that drastically increase the likelihood of survival for many cephalopods. One of the most intriguing abilities in the cephalopod class if found in Sepia officinalis, the common cuttlefish. Cuttlefish are referred to by many as having the most highly adapted form of camouflage in the animal kingdom. The skin of an adult specimen contains on the order of 2 million pigmented chromatophore organs that are innervated to the brain (Hanlon & Messenger, 1988). Dilation and contraction of the yellow, orange and brown chromatophore organs allow for a widespread and nearly instantaneous change in body color and patterning (Hanlon & Messenger, 1988; Hanlon & Messenger, 1996). Along with chromatophore organs, Sepia possess’ reflecting iridiphores and light scattering leucophores, which collectively allow for superior crypsis and signaling abilities (Mäthger et. al, 2008). The cryptic abilities of Sepia officinalis are well studied. It is accepted that, though colorblind, cuttlefish visually cue in on their surroundings, particularly edges, contrast, brightness, size etc., and use this information to adopt appropriate color and textural patterns (Hanlon & Messenger, 1988, 1996; Mathger, et. al, 2006). It is particularly interesting that even though cuttlefish do not perceive color, they effectively display coloration that allows them to be cryptic with their surroundings. This is directly attributable to the fact that the color variation between chromatophores, (orange, yellow and dark brown) closely match that of the substrate (Mäthger et. al, 2008). Previous papers have shown Sepia utilize three categories of camouflage patterns used in varying substrates: uniform, mottle and disruptive. (Hanlon and Messenger, 1988; Hanlon 2007). Substrate spatial scale is an important factor in informing which pattern a cuttlefish will adopt. Camouflage is the primary defense mechanism of numerous animals throughout the animal kingdom. The complexity of the crypsis displayed by cuttlefish suggests the immense selection pressure placed upon the organism by visual predators (Endler, 1986). Not only do they alter their coloration, they often alter morphological characteristics to more closely resemble their surroundings. These abilities offer two fold benefits for their survival and proliferation. First off, they drastically decrease the likelihood of predation, as predators simply will not see them. Secondly increased crypsis will make them more efficient predators, expending less energy to catch prey as they can more effectively hunt and ambush when they remain inconspicuous. With this in mind, it is evident that cuttlefish crypsis and visual acuity of their predators have coevolved over time. As cuttlefish are so finely attuned to their environment, it is reasonable to wonder if they show specific habitat associations to certain substrates or primary placeholders. The ornate displays of cuttlefish crypsis may allow them to be more or less vulnerable in differing habitat, as in theory they can blend into some backgrounds better than others. We attempt to show that cuttlefish do show habitat association in their natural environment and offer explanations about why this might occur. METHODS General Approach The goal of our study was to see whether Sepia officinalis associate to specific habitat during foraging activities. In order to test our hypothesis, we conducted an observational field study of Sepia officinalis. We characterized the habitat of the surroundings of the research station, as well as where we found Sepia in the field. Species Description Sepia officinalis, otherwise known as the common cuttlefish, is found in the waters of the Mediterranean to the eastern Atlantic from southern Norway to the northwest coast of Africa as well as Madeira and the Canary islands (Khromov et al. 1998). They are a mobile species with greatest abundance found at depths of less than 100m. This species, like many other cephalopods, is primarily nocturnal but has been observed during the day with some frequency. The common cuttlefish is usually found on or close to the bottom and its morphology suggests adaptation to a complex environment. It has been shown that cuttlefish visually cue on their surroundings in order to change patterning to become more cryptic (J.J. Allen et al. 2010). Predation pressures are suggested to have played a large role in their adaptation of textural, chromatic and postural components of defensive strategies (Hanlon and Messenger 1996). Upon observation, it becomes apparent that cuttlefish have the ability to rapidly change their color and patterning using chromatophores. This is important to quickly and adaptively respond to different habitats. Site Description We conducted our field study at STARESO Research Station Calvi, Corsica, France (+42.580246, +8.72411). The marine environment around the station primarily consists of Posidonia oceanica meadows and granitic bedrock reefs that extend off the headlands. Surveys were performed to the North and South of the field station as well as in the harbor of the station in order to characterize the largest amount of habitat available for foraging. The surveys we performed all took place October 9-30, 2012. Our habitat characterization transects were run during the day, while our opportunistic sampling of Sepia officinalis was conducted primarily at night. Study Design (Specific Hypotheses) Sepia officinalis associates to the shallow rocky subtidal environment while hunting during the night. Specifically S. officinalis shows a preference to cobble and boulder covered in turfy algal species. In order to show this we surveyed at night, which is Sepia officinalis’s primary feeding, using a similar assessment of the habitat as used in our UPC sampling but with S. officinalis individuals as our point contacts. Data Collection Surveys were conducted with another group to characterize available habitat in and around STARESO harbor. Permanent transect lines were laid out in the harbor, in the cove immediately south of the harbor and just to the north of the harbor. Non-fixed transects were conducted perpendicularly off of these main transect line at intervals of 5 meters starting at the surface or as close to the surface as possible. Meter tape was run out off the permanent transect line and depth was recorded at every meter. At every other meter, uniform point contact was conducted with a 9 point quadrat to classify the substrate composition as well as primary placeholders. These non fixed transect lines extended either to the surface or to a length of 15 meters when it would be infeasible to sample to the surface. Using this method, a profile of depth, substrate and primary placeholders was established. Substrate was classified into distinct groups ranging from sand, cobble, boulder, bedrock, concrete and jacks. As for primary placeholders a large list of distinct categories of algae, sponges, encrusting species and Posidonia was established. We decided to classify algae into 2 distinct groups, Turf and Bushy, based on sizes less than 3 centimeters and greater than 3 centimeters respectively. Opportunistic sampling was conducted on SCUBA primarily at night to assess Sepia officinalis’ habitat association. These surveys consisted of meandering dives over various substrate types from shallow subtidal depths to depths of 12 meters. When individuals were found, they were sized, depth was recorded and a 9 point UPC quadrat was employed to sample substrate and primary placeholder that the cuttlefish was found over. Using this information, a clearer idea of available and utilized habitat will emerge for the population of cuttlefish in the vicinity of STARESO. For the statistical analysis of our data, the help of Pete Raimondi and his knowledge of statistics was enlisted. With his help, our data was put into a PERMANOVA table, which compared the average abundances of the available habitat characteristics and the average abundance of sampled point of Sepia. This allowed the data points of S. officinalis to be compared to the available habitat characteristics across the 3 regions we sampled. Our dataset was separated into two tables: one being for substrate and the other for the primary placeholders. RESULTS General Results After analyzing our data, the results supported half of our initial hypothesis of the habitat association of Sepia officinalis. Our prediction had two distinct features: one being that S. officinalis associated with cobble and boulder substrates and that they also associated with the Turfy algae types. Primary Placeholder Results The most conclusive evidence for the habitat associations of Sepia officinalis comes from our data on the primary placeholders of our study site. As we predicted in our hypothesis, Sepia show a disproportionate tendency to be found on the Turfy algae type (Figure 1). We can say this with a high degree of certainty based of the p-value of 0.001, which is far below our critical p-value of 0.05 (Figure 2). 7 Average Abundances 6 5 4 Series1 3 Series2 2 1 0 T B POS PD Primary Placeholders EL Figure 1: Bar graph comparing the average abundances of the primary placeholders in the available habitat (Red, Series 2) versus the average abundances of the primary placeholders that Sepia officinalis (Blue, Series 1) was found on. Sepia was found disproportionately more frequently on Turfy algae types. Average abundances are out of 9 total in reference to the 9 point UPC quadrats that were used in sampling. Primary placeholder PERMANOVA table of results Source TR Res Total Species T B df SS MS 1 244 245 21646 6.66E+05 6.88E+05 Group Cuttlefish Av.Abund 6.36 1.43 Group Available Av.Abund 2.58 3.77 21646 2730.2 Av.Diss 28.34 21.94 Pseudo-F Unique perms P(perm) 7.9283 Diss/SD 0.001 999 39.99 30.96 Cum.% 39.99 70.95 Contrib% 1.51 1.15 POS PD EL 0 0.86 0.14 1.3 0.17 0.51 7.24 5.51 3.3 0.42 0.4 0.53 10.21 7.77 4.66 81.16 88.93 93.59 Figure 2: Table showing the average abundances of the available habitat and average abundances of primary placeholders that Sepia officinalis was found on. Percent of contributions to the average abundances are also shown, along with the p-value of this PERMANOVA table. Substrate Results Our results for the substrate habitat association of Sepia officinalis proved to be inconclusive. The results showed that there were no strong associations to any substrate type (Figure 2). Also, with a p-value of 0.055, which is above our critical p-value of 0.05, there is not a significant amount of certainty in these particular results (Figure 4). Thus, our hypothesis for the substrate association of Sepia is unsupported by our data. 4.5 Average Abundances 4 3.5 3 2.5 2 Series1 1.5 Series2 1 0.5 0 BO BD CO SA Substrate Figure 3: Bar graphs showing the average abundances of the available substrate types in the study site (Red, Series 2) versus the average abundances of substrate types that Sepia officinalis was found on (Blue, Series 1). Average abundances are out of 9 total in reference to the 9 point UPC quadrats that were used in sampling. Substrate PERMANOVA table of results Source df TR 1 Res 511 Total 512 Groups Cuttlefish & Available Average dissimilarity = 74.63 Group Cuttlefish Species Av.Abund BO 3.21 BD 3 CO 2.14 SS MS 7426.9 1.66E+06 1.66E+06 Group Available Av.Abund 3.96 0.71 1.38 7426.9 3241.1 Av.Diss 24.09 18.73 15.33 Pseudo-F 2.2914 P(perm) 0.055 Diss/SD Contrib% 32.28 25.1 20.54 1.14 0.76 0.89 Unique perms 996 Cum.% 32.28 57.38 77.92 SA 0.64 1.66 11.67 0.56 15.63 93.55 Figure 4: Table showing the average abundances of the available habitat and average abundances of substrates that Sepia officinalis was found on. Percent of contributions to the average abundances are also shown, along with the p-value of this PERMANOVA table. DISCUSSION The substrate portion of our results proved to be the least significant and did not support our specific hypothesis for Sepia. There are a couple potential reasons as to why we did not get the results we were predicting. The first issue comes from our sample size and the comparison of the habitat data to the data on Sepia. Essentially we were trying to compare around 900 habitat quadrats to 14 quadrats for Sepia officinalis. Ideally we would have had more samples for Sepia; however, attempting to study a less common cryptic species yielded a much smaller dataset than we were expecting. The next issue may have come from how we decided to sample with the quadrat. We were taking 9 points of data for substrate for each individual Sepia, this may have led to an over representation of the surrounding habitat as opposed to what the cuttlefish was actually located on. As we predicted, Sepia officinalis showed an association to the Turfy algae species of the primary placeholders we surveyed. This result makes sense for reasons that we mentioned in the introduction. Sepia’s cryptic abilities have distinct characteristics and limitations. Their ability to mimic texture in the three dimensional habitat is limited by the extent their musculature can shift. Thus it makes sense that Sepia officinalis associate to the Turfy algae, which it can mimic best based on, “the animal’s texture perception” (Kelman et al. 2007). Our study showed that Sepia officinalis does associate to a particular habitat while foraging at night. Potential reasoning behind this association may have to do with the ability of Sepia to mimic its environment in color and, to a degree, in its form. As we mentioned earlier, there are a number of benefits to remaining cryptic while foraging. First of all, it could allow Sepia to be a more effective ambush predator and expend less energy while hunting. Second, having exceptional camouflage, “thwarts a wide array of visual predators” (Mäthger et al 2008). Many studies have been performed to study the disruptive body patterns and matching of natural substrates (Kelman et al 2007, Mäthger et al 2008). These studies have shown that Sepia do in fact have the extraordinary ability to mimic their environment, and with our study we aimed to answer how they use this trait in their habitat. By showing that Sepia officinalis do prefer a certain aspect of their environment while foraging, the door is left open to a whole host of other questions regarding Sepia’s interaction with its environment. Future studies could attempt to follow up on our question by analyzing Sepia officinalis’s preferred environment for potential prey items or hiding locations. Studying a predatory species such as Sepia officinalis is very important because so little is truly known about their role in the ecosystem. It is unknown the level of impact these predators have on their environment, but the first step to understanding this question is to find our where they happen to be found. LITERATURE CITED Allen, J.J., et al. 2010. Cuttlefish dynamic camouflage: responses to substrate choice and integration of multiple visual cues. Proceedings of the Royal Society of London B. 277. 1031-1039. Boycott, B., 1961. The Functional Organization of the Brain of the Cuttlefish Sepia officinalis. Proceedings of the Royal Society of London. Series B, Biological Sciences, Vol 153, No. 953: 503-534 Endler, J.A. 1986. Natural Selection in the Wild. Monographs in Population Biology 21. Princeton University Press. 336pp. Hanlon, R.T. ,2007. Cephalopod dynamic camouflage. Current Biology 17(11): R400R404 Hanlon, R.T. , J.B. Messenger. 1988. Adaptive Coloration in Young Cuttlefish (Sepia officinalis L.): The Morphology and Development of Body Patterns and Their Relation to Behavior. Philosophical Transactions of the Royal Society of London. Series B, Biological Sciences, Vol. 320, No. 1200: 437-487 Hanlon, R.T., J.B. Messenger. 1996. Cephalopod behavior. Cambridge University Press Kelman, E.J., A.J. Shohet, D. Osorio, R.J. Baddley. 2007. Perception of visual texture and the expression of disruptive camouflage by the cuttlefish, Sepia officinalis. Proceedings of the Royal Society B. 274. 1369-1375 . Khromov, D.N., Lu, C.C., Guerra, A., Dong, Z. & Boletzky, S.V., 1998. A synopsis of Sepiidae outside Australian waters (Cephalopoda: Sepioidea). In Systematics and biogeography of cephalopod, vol. I (ed. N.A. Voss et al.). pp.77^57. Smithsonian Contributions to Zoology, no. 586. Mäthger et al., 2006. Color blindness and contrast perception in cuttlefish (Sepia officinalis) determined by visual sensorimotor assay. Vision Research, 46(11): 1746-1753 Mäthger et al., 2008. Color matching on natural substrates in cuttlefish, Sepia officinalis. Journal of Comparative Physiology, 194: 577-585 APPENDIX It should be noted that our project did not turn out exactly how we planned. Initially we intended to have both laboratory and field portions of our study. With numerous studies showing that substrate composition affects cuttlefish patterning and behavior, we wanted to examine if this patterning is context specific. We planned on examining cuttlefish behavior and patterning over varying substrates and how it changed in response to varying stimuli (i.e. predators, prey, other cuttlefish). We planned on taking video of our trials and analyzing the chromatophoric pattern shifts over different parts of the body and quantifying them by scoring the type of display seen. Several cuttlefish were captured over the course of our trip and housed individually in a roughly 35cm^3 clear aquarium. Cuttlefish were fed triple fin and blennies to establish a prey preference to use in the study. Because feeding the cuttlefish every trial seemed infeasible for our design, we put the triple fin in a 5cm long clear glass cylinder and attached fishing wire to drop it into the tank to attain replicates. Because the cuttlefish were very responsive to triple fin outside the jar, quickly capturing and consuming the prey, we figured they would do the same at least a few times with the contained prey item. However, we never witnessed one response from the test subjects even though dozens of trials were run. We ran into similar problems testing response to predators. This design consisted of placing a 35cm^3 tank adjacent to the cuttlefish tank and adding an octopus much larger than the cuttlefish. Again, no response was measured. A blind was added to try and attain replicates in this test. Sadly, we ran into problems trying to keep our specimens alive over the duration of our stay. Francis, the first cuttlefish we caught lived in the tank for nearly a week before tragically passing away. However, in this unfortunate event we were able to capture a definitive cuttlefish stress response of two black dots on the dorsal posterior of the mantle. We thought this display was attributable to adding a larger than prey size fish to the tank, however it probably was also effected by the vulnerable, weak state of Francis. During the course of our project we learned quite a bit, primarily that cuttlefish are hard to maintain in a lab setting. They are very sensitive creatures that need a lot of space. They are also have a voracious appetite and can become cannibalistic if left in close quarters with other cuttlefish without enough food. In the field they are easiest seen and captured at night over turfy algae. In the laboratory setting, more, larger tanks, closed off from the stimulus of in the room would have aided in our ability to successfully maintain the specimens. We could have benefited from more time to figure out how to work with the animals. All in all, we were lucky we had a field portion of our project to fall back on. A contingency plan is a good idea when working with a less common, unpredictable cryptic species. Acknowledgements Thank you Pete, Giacomo, Kristen, Jimmy, and Gary. This class was an amazing, eye opening learning experience. Thank you for sharing it with us.