Photosynthesis notes
advertisement

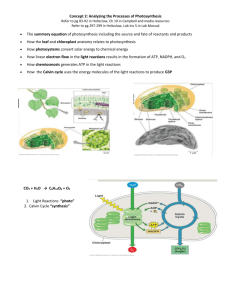
Photosynthesis Autotrophs Plants and some other types of organisms that contain chlorophyll are able to use light energy from the sun to produce food. Autotrophs include organisms that make their own food Autotrophs can use the sun’s energy directly Heterotrophs Heterotrophs are organisms that can NOT make their own food Heterotrophs can NOT directly use the sun’s energy Energy Energy Takes Many Forms such as light, heat, electrical, chemical, mechanical Energy can be changed from one form to another Energy can be stored in chemical bonds & then released later ATP – Cellular Energy Adenosine Triphosphate Contains two, high-energy phosphate bonds Also contains the nitrogen base adenine & a ribose sugar ADP o Adenosine Diphosphate o ATP releases energy, a free phosphate, & ADP when cells take energy from ATP Sugar in ADP & ATP Called ribose Pentose sugar Also found on RNA Importance of ATP Principal Compound Used To Store Energy In Living Organisms Releasing Energy From ATP ATP is constantly being used and remade by cells ATP provides all of the energy for cell activities The high energy phosphate bonds can be BROKEN to release energy The process of releasing ATP’s energy & reforming the molecule is called phosphorylation Releasing Energy From ATP Adding A Phosphate Group To ADP stores Energy in ATP Removing A Phosphate Group From ATP Releases Energy & forms ADP Cells Using Biochemical Energy Cells Use ATP For: Active transport Movement Photosynthesis Protein Synthesis Cellular respiration All other cellular reactions More on ATP Cells Have Enough ATP To Last For A Few Seconds ATP must constantly be made ATP Transfers Energy Very Well ATP Is NOT Good At Energy Storage Glucose Glucose is a monosaccharide C6H12O6 One Molecule of glucose Stores 90 Times More Chemical Energy Than One Molecule of ATP Photosynthesis Involves the Use Of light Energy to convert Water (H20) and Carbon Dioxide (CO2) into Oxygen (O2) and High Energy Carbohydrates (sugars, e.g. Glucose) & Starches Investigating Photosynthesis o Many Scientists Have Contributed To Understanding Photosynthesis o Early Research Focused On The Overall Process o Later Researchers Investigated The Detailed Chemical Pathways Pigments In addition to water, carbon dioxide, and light energy, photosynthesis requires Pigments Chlorophyll is the primary light-absorbing pigment in autotrophs Chlorophyll is found inside chloroplasts Light and Pigments Energy From The Sun Enters Earth’s Biosphere As Photons Photon = Light Energy Unit Light Contains A Mixture Of Wavelengths Different Wavelengths Have Different Colors Different pigments absorb different wavelengths of light Photons of light “excite” electrons in the plant’s pigments Excited electrons carry the absorbed energy Excited electrons move to HIGHER energy levels Chlorophyll There are 2 main types of chlorophyll molecules: Chlorophyll a Chlorophyll b A third type, chlorophyll c, is found in dinoflagellates Chlorophyll a Found in all plants, algae, & cyanobacteria Makes photosynthesis possible Participates directly in the Light Reactions Can accept energy from chlorophyll b Chlorophyll b Chlorophyll b is an accessory pigment Chlorophyll b acts indirectly in photosynthesis by transferring the light it absorbs to chlorophyll a Like chlorophyll a, it absorbs red & blue light and REFLECTS GREEN Inside A Chloroplast Structure of the Chloroplast Double membrane organelle Outer membrane smooth Inner membrane forms stacks of connected sacs called thylakoids Thylakoid stack is called the granun (grana-plural) Gel-like material around grana called stroma Function of the Stroma Light Independent reactions occur here ATP used to make carbohydrates like glucose Location of the Calvin Cycle Thylakoid membranes Light Dependent reactions occur here Photosystems are made up of clusters of chlorophyll molecules Photosystems are embedded in the thylakoid membranes The two photosystems are: o Photosytem I o Photosystem II Energy Carriers Nicotinamide Adenine Dinucleotide Phosphate (NADP+) NADP+ = Reduced Form Picks Up 2 high-energy electrons and H+ from the Light Reaction to form NADPH NADPH carries energy to be passed on to another molecule Light Dependent Reactions Occurs across the thylakoid membranes Uses light energy Produce Oxygen from water Convert ADP to ATP Also convert NADP+ into the energy carrier NADPH Photosystem I o Discovered First o Active in the final stage of the Light Dependent Reaction o Made of 300 molecules of Chlorophyll o Almost completely chlorophyll a Photosystem II o Discovered Second o Active in the beginning stage Of the Light Dependent Reaction o Contains about equal amounts of chlorophyll a and chlorophyll b Photosynthesis Begins Photosystem II absorbs light energy Electrons are energized and passed to the Electron Transport Chain Lost electrons are replaced from the splitting of water into 2H+, free electrons, and Oxygen 2H+ pumped across thylakoid membrane Photosystem I High-energy electrons are moved to Photosystem I through the Electron Transport Chain Energy is used to transport H+ from stroma to inner thylakoid membrane NADP+ converted to NADPH when it picks up 2 electrons & H+ Phosphorylation Enzyme in thylakoid membrane called ATP Synthase As H+ ions passed through thylakoid membrane, enzyme binds them to ADP Forms ATP for cellular Light Reaction Summary Reactants: o H2O o Light Energy Energy Products: o ATP o NADPH Light Independent Reaction ATP & NADPH from light reactions used as energy Atmospheric C02 is used to make sugars like glucose and fructose Six-carbon Sugars made during the Calvin Cycle Occurs in the stroma The Calvin Cycle Two turns of the Calvin Cycle are required to make one molecule of glucose 3-CO2 molecules enter the cycle to form several intermediate compounds (PGA) A 3-carbon molecule called Ribulose Biphosphate (RuBP) is used to regenerate the Calvin cycle Factors Affecting the Rate of Photosynthesis Amount of available water Temperature Amount of available light energy Alternative mechanisms of carbon fixation have evolved in hot, arid climates One of the major problems facing terrestrial plants is dehydration. At times, solutions to this problem conflict with other metabolic processes, especially photosynthesis. The stomata are not only the major route for gas exchange (CO2 in and O2 out), but also for the evaporative loss of water. On hot, dry days plants close the stomata to conserve water, but this causes problems for photosynthesis. In most plants (C3 plants) initial fixation of CO2 occurs via rubisco and results in a three-carbon compound, 3-phosphoglycerate. These plants include rice, wheat, and soybeans. When their stomata are closed on a hot, dry day, CO2 levels drop as CO2 is consumed in the Calvin cycle. At the same time, O2 levels rise as the light reaction converts light to chemical energy. While rubisco normally accepts CO2, when the O2/CO2 ratio increases (on a hot, dry day with closed stomata), rubisco can add O2 to RuBP. When rubisco adds O2 to RuBP, RuBP splits into a three-carbon piece and a two-carbon piece in a process called photorespiration. The two-carbon fragment is exported from the chloroplast and degraded to CO2 by mitochondria and peroxisomes. Unlike normal respiration, this process produces no ATP, nor additional organic molecules. Photorespiration decreases photosynthetic output by siphoning organic material from the Calvin cycle. A hypothesis for the existence of photorespiraton (a inexact requirement for CO2 versus O2 by rubisco) is that it is evolutionary baggage. When rubisco first evolved, the atmosphere had far less O2 and more CO2 than it does today. The inability of the active site of rubisco to exclude O2 would have made little difference. Today it does make a difference. Photorespiration can drain away as much as 50% of the carbon fixed by the Calvin cycle on a hot, dry day. Certain plant species have evolved alternate modes of carbon fixation to minimize photorespiration. The C4 plants fix CO2 first in a four-carbon compound. Several thousand plants, including sugercane and corn, use this pathway. In C4 plants, mesophyll cells incorporate CO2 into organic molecules. The key enzyme, phosphoenolpyruvate carboxylase, adds CO2 to phosphoenolpyruvate (PEP) to form oxaloacetetate. PEP carboxylase has a very high affinity for CO2 and can fix CO2 efficiently when rubisco cannot, i.e. on hot, dry days when the stomata are closed. The mesophyll cells pump these four-carbon compounds into bundle-sheath cells. The bundle-sheath cells strip a carbon, as CO2, from the four-carbon compound and return the three-carbon remainder to the mesophyll cells. The bundle-sheath cells then use rubisco to start the Calvin cycle with an abundant supply of CO 2. In effect, the mesophyll cells pump CO2 into the bundle sheath cells, keeping CO2 levels high enough for rubisco to accept CO2 and not O2. C4 photosynthesis minimizes photorespiration and enhances sugar production. C4 plants thrive in hot regions with intense sunlight. A second strategy to minimize photorespiration is found in succulent plants, cacti, pineapples, and several other plant families. These plants, known as CAM plants for crassulacean acid metabolism (CAM), open stomata during the night and close them during the day. Temperatures are typically lower at night and humidity is higher. During the night, these plants fix CO2 into a variety of organic acids in mesophyll cells. During the day, the light reactions supply ATP and NADPH to the Calvin cycle and CO2 is released from the organic acids. Both C4 and CAM plants add CO2 into organic intermediates before it enters the Calvin cycle. In C4 plants, carbon fixation and the Calvin cycle are spatially separated. In CAM plants, carbon fixation and the Calvin cycle are temporally separated. Both eventually use the Calvin cycle to incorporate light energy into the production of sugar.