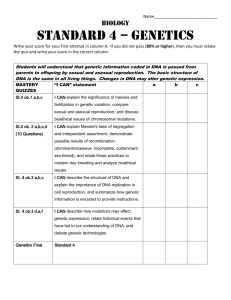
2. History of the genetics and biotechnology
advertisement

Applied Genetics and Biotechnology Pepó, Pál Created by XMLmind XSL-FO Converter. Applied Genetics and Biotechnology Pepó, Pál TÁMOP-4.1.2.A/1-11/1-2011-0009 University of Debrecen, Service Sciences Methodology Centre Debrecen, 2013. Created by XMLmind XSL-FO Converter. Tartalom Tárgymutató ....................................................................................................................................... 1 Terminology (important terms) .......................................................................................................... ii 1. 1. Basis and history ......................................................................................................................... 4 1. Fundamentals and the history of genetics ............................................................................. 4 2. History of the genetics and biotechnology ............................................................................ 8 3. Fundamentals of biotechnology .......................................................................................... 10 4. Timeline of plant biotechnology ......................................................................................... 11 2. 2. Biotechnology of asexual reproduction .................................................................................... 12 1. Tissue culture ...................................................................................................................... 12 2. Micropropagation ................................................................................................................ 14 3. Meristerm cultrure ............................................................................................................... 15 4. Shoot culture and adventitious shoot culture ...................................................................... 16 3. 3. Biotechnology of sexual reproduction ...................................................................................... 17 1. Embryo culture .................................................................................................................... 17 2. Microspore culture .............................................................................................................. 18 4. 4. Manipulating of DNA ............................................................................................................... 20 1. Molecular cloning ............................................................................................................... 20 2. Genetic transformation ........................................................................................................ 21 3. Agrobacterium mediated gene transformation .................................................................... 22 4. Direct gene transformation .................................................................................................. 25 5. Chemical methods ............................................................................................................... 25 6. Physical gene transfer methods ........................................................................................... 26 5. 5. Plant regeneration/molecular selection ..................................................................................... 28 1. In vitro plant regeneration ................................................................................................... 28 2. Influence of carbon sources on plant regeneration in wheat ............................................... 28 3. Molecular marker ................................................................................................................ 31 4. Cryopreservation ................................................................................................................. 34 6. 6. Genetically modified plants (GM) ............................................................................................ 38 1. Types of genetically modified plants .................................................................................. 39 2. GMOs in the U.S. ................................................................................................................ 45 3. GMOs in Europe ................................................................................................................. 46 4. GMOs in Hungary ............................................................................................................... 49 7. 7. Basis of plant breeding ............................................................................................................ 50 1. Plant genomics .................................................................................................................... 50 2. Fundamentals of plant breeding .......................................................................................... 52 8. 8. Methods of plant breeding ........................................................................................................ 55 1. Selective breeding ............................................................................................................... 55 2. Polyploidy ........................................................................................................................... 59 3. Hybridization/heterosis ....................................................................................................... 60 4. Mutation breeding ............................................................................................................... 63 9. 9. New alternatives in plant breeding ........................................................................................... 67 1. The genotypes, fertilization and irrigation from environmental point of view ................... 67 2. Regeneration of maize using tissue culture and inhibition of pollen grain germination ..... 69 3. Haploid production in wheat ............................................................................................... 70 10. 10. Resistance breeding .............................................................................................................. 73 1. Stress resistance .................................................................................................................. 73 1.1. Indirect breeding for stress environments ............................................................... 73 1.2. Direct breeding for stress environments ................................................................. 73 1.3. Breeding under precise, laboratory-controlled conditions ...................................... 74 1.4. Breeding for fundamental causes of stress resistance ............................................ 74 2. Genetic engineering for stress resistance ............................................................................ 74 3. Resistance to abiotic stress ................................................................................................. 75 3.1. Salt tolerance of potato ........................................................................................... 75 3.2. Adaptation of maize lines and hybrids to abiotic/biotic stresses ............................ 78 11. 11. Resistance to biotic stress .................................................................................................... 79 1. Resistance to aphids in maize (Zea mays L.) ..................................................................... 79 iii Created by XMLmind XSL-FO Converter. Applied Genetics and Biotechnology 2. Investigation of obligate parasites (Striga Spp.) in vitro cirumtances ................................. 80 12. 12. Environmentally-sound plant breeding ................................................................................. 81 1. Genetic manipulation in agricultural environment .............................................................. 81 2. Wheat and maize improvement for sustainable agriculture ................................................ 82 13. 13. Wheat and sorghum breeding ............................................................................................... 83 1. Evaluation of prevailing tendencies wheat quality .............................................................. 83 2. Amino acid compositions in wheat species with different genomes .................................. 87 3. Effects of amino acid fertilizers on yield and quality of winter wheat ............................... 87 4. Improvement of Sorghum breeding .................................................................................... 88 14. 14. MAIZE BREEDING ............................................................................................................ 90 1. Improve of maize quality (chemical component) ................................................................ 90 2. Morphology of tassel components and their relationship in maize ..................................... 92 3. Chemical contents and thousand kernel weight in maize breeding ..................................... 92 4. Evaluation of blue and red corn landraces under Hungarian conditions ............................. 95 5. Importance of blue/red kernel corns .................................................................................... 96 6. Antioxidant activities in blue, purple and red kernel corns ............................................... 100 15. 15. SELECTION OF HIGH ENERGY FIBER PLANTS ........................................................ 102 16. 16. PLANT IDENTIFICATION FOR SELECTION ............................................................... 107 1. Genetic diversity ............................................................................................................... 107 2. Genetic similarity and hybrid performance in irradiated maize ........................................ 109 iv Created by XMLmind XSL-FO Converter. Az ábrák listája 1.1. Figure 1.: Centromeric position and arm length .......................................................................... 5 1.2. Figure 2.: Gregor Johan Mendel (1822-1884) ............................................................................. 5 1.3. Figure 3.: Mendel analysed seven characters of garden pea ........................................................ 6 1.4. Figure 4.: Mendel crossed wrinkled yellow peas with round green peas .................................... 7 1.5. Figure 5.: F2 generation all possible combinations of traits, in a characteristic 9:3:3:1 phenotypic ratio ............................................................................................................................................................. 7 1.6. Figure 6.: The ‘Golden rice’ ...................................................................................................... 11 2.1. Figure 7.: Overview of plant tissue culture process ................................................................... 12 2.2. Figure 8.: Technology of plant tissue culture ............................................................................ 14 4.1. Figure 9.: Restriction endonucleases cut DNA .......................................................................... 20 4.2. Figure 10.: Agrobacterium-mediated transformation ................................................................ 22 4.3. Figure 11.: Genes in the T-DNA ............................................................................................... 23 4.4. Figure 12.: Plants that have undergone transformation with Agrobacterium ............................ 24 4.5. Figure 13.: The scheme of the GENEBOOSTER (Gene Gun) and the principles of genetic bombardment .................................................................................................................................... 27 5.1. Figure 14.: Alternative ways of plant regenaration .................................................................... 28 5.2. Table 4.: Induction and regeneration in vitro with different carbon sources in media. ............. 29 5.3. Figure 15.: Induction frequency (%) in parallel with sugar concentrations ............................... 30 5.4. Figure 16.: Proportions of plant regeneration in maltose ........................................................... 30 5.5. Figure 17.: Proportions of plant regeneration in sucrose ........................................................... 31 5.6. Figure 18. Figure 19. ......................................................................... 34 5.7. Figure 20. Figure 21. ........................................................................ 35 5.8. Figure 22. Figure 23. ......................................................................... 36 5.9. Figure 24. Figure 25. ......................................................................... 36 5.10. Figure 26. Figure 27. ....................................................................... 36 5.11. Figure 28. Figure 29. ....................................................................... 37 6.1. Table 6.: Generations and possible uses of GM plants .............................................................. 39 6.2. Table 7.: Possible advantages of GM production for the preservation of the environment ....... 41 6.3. Figure 30.: Phytoremediation strategies .................................................................................... 41 6.4. Table 8.: Potential risks of GMO production ............................................................................. 42 6.5. Figure 31.: Authorisation process of GMO’s ............................................................................ 44 6.6. Figure 32.: Gene technological marking .................................................................................... 45 6.7. Figure 33.: Approaches of the public to fields of biotechnology in Europe and the U.S. 1. remedies, 2. genetic testing, 3. crops, 4. food, 5. organ transplant ........................................................................ 48 7.1. Figure 34.: Arabidopsis thaliana ................................................................................................ 50 7.2. Figure 35.: Genome size, chromosome and genes number of Arabidopsis thaliana ................. 51 7.3. Table 13.: Gene complement of Arabidopsis ............................................................................ 51 7.4. Figure 36.: Oryza sativa ............................................................................................................. 51 8.1. Figure 37.: Mass selection ......................................................................................................... 55 8.2. Figure 38.: Pedigree method ...................................................................................................... 56 8.3. Table 15.: Full diallel cross method ........................................................................................... 58 8.4. Figure 39.: Ear crops of lines of ‘A’ complete diallel system ................................................... 58 8.5. Figure 40.: Parental lines of ‘A’ complete diallel system at the time of flowering ................... 58 8.6. Table 17.: Changes of heterosis values in our maize hybrids (Debrecen) ................................. 61 8.7. Figure 41.: Scheme of mutation breeding .................................................................................. 63 8.8. Figure 42.: The cyclotron (Institute for Nuclear Research, Debrecen) ...................................... 63 8.9. Figure 43.: The effect of the mutation in Sorghum panicle ....................................................... 65 8.10. Figure 44.: Double panicle in the case of ‘12 Gy Zádor’ ......................................................... 66 9.1. Figure 45.: Types of winter wheats according to their fertilizer responses ............................... 67 9.2. Figure 46. : Complex maize breeding program in Debrecen ..................................................... 71 10.1. Figure 47.: Somaclones propagated at different NaCl concentrations ..................................... 75 10.2. Figure 48.: Length changes of in vitro cultivated plants by the effect of NaCl ....................... 76 11.1. Table 19.: Pathological data of middle winter wheat varieties (infected surface%) (National Institute for Agricultural Quality Control) (2005-2010) ................................................................................. 79 13.1. Table 22.: Flour and dough properties of ‘HP Pusztaszél’ cultivar (Triticum aestivum L.). Central Agricultural Office, 2001-2003) ....................................................................................................... 86 v Created by XMLmind XSL-FO Converter. Applied Genetics and Biotechnology 13.2. Figure 49.: Male sterile female and restorer male lines ........................................................... 88 13.3. Figure 50.: Process of the bioethanol production (experimental) ............................................ 89 14.1. Figure 51.: Protein, starch and oil content data of the maize genotypes (Debrecen) ............... 90 14.2. Figure 52.: Correlation between starch and protein content in maize hybrids (Debrecen) ...... 90 14.3. Figure 53.: Trends in the protein content of the investigated maize (Debrecen) ..................... 94 14.4. Figure 54.: Ash contents of the investigated maize (Debrecen) .............................................. 94 14.5. Table 28.: Macroelement contents (mg kg-1) of investigated corn varieties (mean±SD; dry weight basis) ................................................................................................................................................. 97 14.6. Table 29.: Some microelements (mg kg-1) of investigated corn varieties I. (mean±SD, dry weight basis) ................................................................................................................................................. 97 14.7. Table 30.: Some microelements (mg kg –1) of investigated corn varieties II. (mean±SD, dry weight basis) ................................................................................................................................................. 98 15.1. Figure 55.: Miscanthus .......................................................................................................... 102 15.2. Figure 56 .: Shoot weight development (g) during 40 days of Miscanthus x giganteus (M118) 103 15.3. Figure57.: Shoot weight development (g) during 40 days of Miscanthus sinensis (Goliath) 103 15.4. Figure 58.: Relative increase in shoot weight (M118/Goliath) .............................................. 104 15.5. Figure 59.: Change of nitrate content in the nutrient solution in comparison with growth of shoot of two Miscanthus genotypes (G1, G3) during the 60 days culturing. ................................................ 105 15.6. Figure 60.: Change of phosphate content in the nutrient solution in comparison with growth of shoot of two Miscanthus genotypes (G1, G3) during the 60 days culturing. ........................................... 105 16.1. Figure 61.: Low plant height, increased sprouting activity and double panicle in the case of ‘12 Gy Zádor’ ............................................................................................................................................. 107 16.2. Figure 62.: Low plant height and panicle sterility in ‘12 Gy Rib .......................................... 108 16.3. Figure 63.: Shorghum albino specimens with extreme chlorophyll defects - Irradiated plants ‘10 Gy Zádor’ ............................................................................................................................................. 108 16.4. Figure 64.: Dendogram obtained from cluster analysis based on the morphological descriptions (DUS) of four maize inbred lines ................................................................................................... 110 16.5. Figure 65.: Dendogram obtained from cluster analysis based on the zein patterns of four maize inbred lines. .................................................................................................................................... 110 16.6. Figure 66. Dendogram obtained from cluster analysis based on the AFLP amplification patterns of four maize inbred lines. .................................................................................................................. 110 vi Created by XMLmind XSL-FO Converter. A táblázatok listája 1.1. Table 1.: Diploid chromosome numbers in plant species ........................................................... 4 1.2. Table 2.: Timetable of Genetics and Biotechnology History ...................................................... 9 2.1. Table 3.: Advantages and limitations of different methods of micropropagation ...................... 14 6.1. Table 5.: Strategies for developing herbicide resistance ............................................................ 38 6.2. Table 9.: Potential risks of GMO ............................................................................................... 44 6.3. Table 10.: Average pesticide pounds applied per acre planted to conventional, herbicide-tolerant (HT) and Bt transgenic varieties ................................................................................................................ 46 6.4. Table 11.: Cultivation areas (hectares) of Bt corn in European countries ................................. 47 7.1. Table 12.: Nuclear genome size of plants .................................................................................. 50 7.2. Table 14.: Genome size, chromosome and genes number of Oryza sativa ................................ 52 8.1. Table 16.: Polyploid plants ........................................................................................................ 59 8.2. Table 18.: Some characterization of irradiated Sorghum genotypes ......................................... 65 13.1. Table 20.: General characteristics of cereal breeding trends for a continuous of organic faming systems (Verhoog et al. (2003) and Lammerts van Bueren and Struik (2004)) ................................ 84 13.2. Table 21.: Traits for winter wheat that is relevant for the prevailing tendencies (Lammerts van Bueren et al., 2002). .......................................................................................................................... 85 13.3. Table 23.: Dough quality of winter wheat cultivars ................................................................. 86 14.1. Table 24.: Correlation matrix .................................................................................................. 91 14.2. Table 25.: Characteristics of investigated maize ..................................................................... 93 14.3. Table 26.: Trends in thousand kernel weight in the investigated maize .................................. 93 14.4. Table 27.: Trends in starch contents (%) of the investigated maize (Debrecen) ...................... 94 14.5. Table 31.: Pearson’s correlation coefficients between investigated elements (without Li) ..... 99 15.1. Table 32.: Shoot induction of Miscanthus x giganteus (M118) in different culture media ... 104 15.2. Table 33 .: Shoot induction of Miscanthus sinensis (Goliath) in different culture media ...... 104 16.1. Table 34.: Origin and identification of the investigated lines ................................................ 109 16.2. Table 35.: A genetic background of the full diallel system ................................................... 109 16.3. Table 36.: Effect of general (GCA) and specific (SCA) combining ability to yield (kg/plot) in full diallel system .................................................................................................................................. 111 vii Created by XMLmind XSL-FO Converter. Tárgymutató 1 Created by XMLmind XSL-FO Converter. Terminology (important terms) Allel: An allele is an alternative form of a gene that is located at a specific position in a chromosome. Chromatide: One of the members of the chromosome called chromatid. Chromosome: Chromosome is an organized structure of DNA and protein in cells. Diploid: Term used when a cell contains both sets of homologous chromosomes. DNA (deoxyribonucleic acid): A nucleic acid that carries the genetic information in the cell and is capable of self-replication and synthesis of RNA. DNA consists of two long chains of nucleotides twisted into a double helix and joined by hydrogen bonds between the complementary bases adenine and thymine or cytosine and guanine. The sequence of nucleotides determines individual hereditary characteristics. Dominant allel: It is an allele that can take over a recessive allele, so if you have a dominant allele and a recessive allele, then the offspring will most likely have a dominant allele over a recessive allele. The dominant allele is expressed over the recessive allele. Gamete: A reproductive cell or sex cell that contains the haploid set of chromosomes, e.g. sperm cell (male reproductive cell) and egg cell or ovum (female reproductive cell). Gene: The functional and structural molecular unit of the DNA (deoxyribonucleic acid) is the gene. The gene that particulate hereditary determiner located is chromosome and responsible for traits and carries genetic information from one generation to the other. It is the basic unit of heredity. Gene mapping: Determination of the positions of genes on a DNA and of the distance, in ulinkage units or physical units, between them. General Combining Ability (GCA): The average performance of a parent in a series of crosses. Genome: a kind of an ‘instruction manual’ inherited from the parents of the plant as a combination of their genomes All the genetic material in a cell. Genomics: It is the mapping and sequencing of all the genetic material in the DNA of a particular organism. Genomics are used of information derived from genome sequence data to further elucidate what genes do, how they are controlled, and how they work together. Genotype: It is the set of genes which expresses the phenotypic character. GMO (Genetically Modified Organisms): Organisms, genetically engineered by inserting a gene from another organism, are called genetically modified organisms. In vitro: In an artificial environment outside the living organism. In vivo: In the living organism. An experiment that is done in vivo is done in the body of a living organism. F1: The first generation of hybrid between two parents is called first filial generation, abbreviated F 1. The next and subsequent generations are referred to as F2, F3, F4, etc. Haploid: Term used when a cell contains only a single set of chromosomes and therefore only a single set of genes. Heterozygote: An organism having two different alleles of a particular gene. Homozygote: An organism having two identical alleles of a particular gene. Hybrid: The progeny of cross-breeding two pure-breeding lines. Locus: The location of a gene on a chromosome called locus. ii Created by XMLmind XSL-FO Converter. Terminology (important terms) Molecular marker: Molecular marker is a gene or DNA sequence with a known location on a chromosome that can be used to identify individuals or species. Mutation: Any inheritable change in DNA sequence. Plasmid: Plasmids are double-stranded DNA (dsDNA) molecules which are separate from a cell’s chromosomal DNA (extrachromosomal DNA). Plasmids usually are circular. Some plasmids can integrate into the host genome, can be artificially constructed in the laboratory, and serve as cloning vectors. Phenotype: Phenotypes result from the expression of an organism's genes as the influence of environmental factors and the interactions between the two. Recessive allel: A recessive allele is one in which the effect is not tangible, or is masked by the effects of the dominant gene. The recessive trait may be expressed when the recessive genes are in homozygous condition or when the dominant gene is not present. That happens when an organism inherits a pair of recessive genes from its parents. Restriction enzymes: Enzymes (proteins) that recognize specific, short nucleotide sequences in DNA and cut at those sites. Specific combining ability (SCA): Deviation in the performance of a cross from the performance predicted on the basis of general combining ability of its parents. Tissue culture: A process of growing a plant in the laboratory from cells. This technique is used in traditional plant breeding as well as when using techniques of plant biotechnology. Transformation: The genetic transformation is a process by which the genetic material carried by an individual cell is altered by the incorporation of foreign (exogenous) DNA into its genome. Vector: Vector is a vehicle for transferring genetic material into a cell. iii Created by XMLmind XSL-FO Converter. 1. fejezet - 1. Basis and history 1. Fundamentals and the history of genetics Genetics is the branch of biology that studies heredity and variation in organisms. Genetics is about genes and chromosomes, their structure and function, their behaviour and mis behaviour, their evolution, and methods of studying them. Main task of genetics is to clarify the really strange fact why life appears in so many different forms and to analyse the causes of similarities between parents and offspring. DNA is the basic hereditary material located in the chromosomes. Most DNA is located in the cell nucleus (it is called nuclear DNA), but a small amount of DNA can also be found in the mitochondria (it is called mitochondrial DNA or mtDNA). Mostly in plant cells and other eukaryotic organisms that perform photosynthesis, chloroplast also contains circular double stranded DNA molecules (it is called chloroplast DNA or cpDNA). In 1953 the British biochemist Francis Crick and his American colleague James Watson discovered the molecular structure of the deoxyribonucleic acid (DNA), later in 1962 they got the Nobel Prize in Medicine and Physiology. DNA is a polymer which is known as a "polynucleotide." The monomer units of DNA are nucleotides. Each nucleotide consists of a 5-carbon sugar (deoxyribose), a nitrogen containing base attached to the sugar, and a phosphate group. There are four different types of nucleotides found in DNA, differing only in the nitrogenous base. The sugar-phosphate backbone is on the outside of the helix, and the nitrogenous bases are on the inside. Each base pair is formed from two complementary nucleotides (purine with pyrimidine). The base pairs in DNA are adenine with thymine and cytosine with guanine bound together by weak hydrogen bonds. The two strands of the double helix are anti-parallel, which means that they run in opposite directions. The functional and structural molecular unit of the DNA (deoxyribonucleic acid) is the gene. The gene is located in chromosome and responsible for traits and carries genetic information from one generation to the other. Danish scientist Johansen used first the term “gene”. The location of a gene on a chromosome called locus. An allele is an alternative form of a gene that is located at a specific position in a chromosome. The process by which alleles are transmitted was discovered by Gregor Mendel and formulated in what is known as Mendel's law of segregation. These DNA codings determine distinct traits that can be passed on from parents to offspring. Chromosome is an organized structure of DNA and protein in cells. Eukaryotic cells contain large linear chromosomes in nuclei. Prokaryotic cells (cells without defined nuclei) have smaller circular chromosomes in cytoplasm. Plants, animals and humans have their chromosomes in pairs, one member of each from the mother and one from the father (Table 1.). This chromosome pairs are separated at the reduction division of meiosis. One of the members of the chromosome called chromatid. 1.1. táblázat - Table 1.: Diploid chromosome numbers in plant species Plant species Diploid or zygotic number (2n) Durum wheat (Triticum durum) 28 Wheat (Triticum aestivum) 42 Rye (Secale cereale) 14 Barley (Hordeum vulgare) 14 Oat (Avena sativa) 42 4 Created by XMLmind XSL-FO Converter. 1. Basis and history Rice (Orysa sativa) 24 Maize (Zea mays) 20 Sunflower (Helianthus annuus) 34 Squash (Cucurbita pepo) 40 Line (Linum usitatissiumum) 30, 32 Potato (Solanum tuberosum) 48 Tomato (Lycopersicum esculentum) 24 Sugar-beet (Beta vulgaris) 18 Onion (Allium cepa) 16 Bean (Phaseolus vulgaris) 22 Apple (Malus silvestris) 34, 51 Pear (Pyrus communis) 34, 51, 68 Cherry (Prunus cerasus) 32 Apricot (Prunus armeniace) 16 Under a microscope, chromosomes are visible in the cell’s nucleus when the cell is dividing. Each chromosome is made up of DNA tightly coiled many times around proteins called histones that support its structure. Centromere ulinks sister chromatids and divides the chromosome into two arms. The short arm is labelled the “p” arm, and the long arm of the chromosome is labelled the “q” arm. The position of the centromere gives the chromosome its characteristic shape. Chromosomes vary by size of “arm” and centromere location (Figure 1.). 1.1. ábra - Figure 1.: Centromeric position and arm length Gregor Johan Mendel (Figure 2.) an Augustinian monk, who lived in Brünn, Austria (now Brno, Czech Republic), he worked on heredity and became the father of the modern theory of genetics. 1.2. ábra - Figure 2.: Gregor Johan Mendel (1822-1884) 5 Created by XMLmind XSL-FO Converter. 1. Basis and history Mendel's research was with common garden pea (Pisum sativum); the basic underlying principles of heredity that he discovered also apply to animals and people because the mechanisms of heredity are essentially the same for all complex life forms. It is necessary to introduce some technical terms. The parents of any cross are designated by the capital letter P. The first generation of hybrid between two parents is called first filial generation, abbreviated F 1. A mating of two F1 hybrids, or self-fertilization of the F1 produces the F2 generation. The dominant alleles being indicated by capital letters, the recessive alleles by small ones. Garden pea plants were great object of the heredity because they can be grown easily in large numbers and their reproduction can be manipulated, they can either self-pollinate themselves or cross-pollinate with another plants. Pea plants have both male and female reproductive organs. Mendel carefully analysed seven characters of garden pea for comparison, such as (Figure 3.): 1. Seed colour: yellow or green 2. Seed shape: round or wrinkled 3. Flower colour: purple or white 4. Flower position: axial or terminal 5. Pod colour: yellow or green 6. Pod shape: inflated or constricted 7. Stem length: tall or short 1.3. ábra - Figure 3.: Mendel analysed seven characters of garden pea From crosses, he found that the hybrids were not all intermediate between the parents, but in most cases corresponded to one or the other of them. When he crossed a yellow pea seed by a green pea seed variety, the hybrid was yellow pea seed. The “yellow” parent seemed to “dominate” the appearance of the hybrid, Mendel 6 Created by XMLmind XSL-FO Converter. 1. Basis and history called it dominant. The opposite of this, the green characteristic he called the recessive. Mendel experienced that the first offspring generation (F1) always has yellow seeds. Furthermore, the following generation (F 2) consistently has a 3:1 ratio of yellow to green. 1.4. ábra - Figure 4.: Mendel crossed wrinkled yellow peas with round green peas To determine the behavior of the two genes simultaneously (a dihybrid cross), Mendel crossed wrinkled yellow peas with round green peas. The F1 generation all showed the dominant traits for both characters, round and yellow (Figure 4.). When he crossed two F1 hybrids, he obtained in the F2 generation all possible combinations of traits, in a characteristic 9:3:3:1 phenotypic ratio (Figure 5.). 1.5. ábra - Figure 5.: F2 generation all possible combinations of traits, in a characteristic 9:3:3:1 phenotypic ratio 7 Created by XMLmind XSL-FO Converter. 1. Basis and history Mendel's contribution was unique because he was able to demonstrate that traits were passed from each parent to their offspring through the inheritance of genes. MENDEL's first law, it is also called the principle of uniformity. This statement says that if two heterozygote plants that differ from each other in just one characteristic are crossed, then the resulting F1 hybrids are uniform in the phenotype and genotype as well. Example: when we cross a yellow pea seed by a green pea seed variety, the F1 hybrid is always yellow and heterozygote. Depending on the traits is the uniform features either one of the parents' traits (a dominant-recessive pair of characteristics - like pea colour) or it is intermediate – like incomplete dominance in flowers of Mirabilis jalapa. MENDEL's second law it is also called the principle of segregation. The individuals of the F2 generation are not uniform, instead different types are visible. The characteristics of the parental generation do always occur at a certain ratio. Depending on a dominant-recessive or an intermediate crossing, they segregate in the ratio 3:1 or 1:2:1. MENDEL's third law, it is also called the principle of „pure” gametes. The gametes are pure means that they always carry only one allele of the gene. 2. History of the genetics and biotechnology 8 Created by XMLmind XSL-FO Converter. 1. Basis and history 1.2. táblázat - Table 2.: Timetable of Genetics and Biotechnology History 1665 Robert Hooke Discovered cells under a microscope. 1838 Matthias Schleiden Stated all plants are made of cells. 1839 Theodor Schwann Stated all animals are made of cells. 1859 Charles Darwin Published his book “Origin of Species,” which explained units of heredity and variations in species. 1866 Gregor Mendel Published the results of inheritance of "factors" in garden pea. Later he would become known as the father of genetics. 1880 Walther Fleming Discovered mitosis. 1890 Heinrich Waldeyer 1900 Carl Correns von Discovered chromosomes. They independently rediscovered Mendel's principles and verified, marking the beginning of modern genetics. Hugo de Vries Erich von Tschermak 1902 Walter Sutton Pointed out the interrelationships between cytology and Mendelism, closing the gap between cell morphology and heredity. 1904 Gregory Bateson He was the first person to use the term genetics and discovered ulinkage between genes. 1905 Nettie Stevens They independently discovered X & Y sex chromosomes. Edmund Wilson 1951 Rosalind Franklin 1910 Thomas Morgan Made x-ray photographs of DNA. Hunt Proved that genes are located on the chromosomes (using Drosophila). 1927- Hermann J. Muller 28 Showed that X-rays induce mutations in a dose-dependent fashion in Drosophila. 1931 Barbara McClintock Demonstrated the crossing-over in maize. 1953 Francis Crick and Solved the three-dimensional structure of the DNA molecule. James Watson 1958 Arthur Kornberg Discovered the DNA polymerase I. 1966 Marshall Nirenberg Solved the genetic code, showed that 3 DNA bases code for one amino acid 1970 Hamilton Smith Isolated the first restriction enzyme, which cut the DNA molecules within specific recognition sites. Kent Wilcox 9 Created by XMLmind XSL-FO Converter. 1. Basis and history 1972 1972 The DNA composition of chimpanzees and gorillas is discovered to be 99% similar to that of humans. Paul Berg They made the first recombinant DNA molecules. Combined DNA from two different species in vitro. Herb Boyer 1977 Fred Sanger Developed the DNA sequencing technology. 1977 The Genentech the first genetic engineering company was founded made medically important drugs using recombinant DNA methods. 1978 The first human hormone was produced using recombinant DNA technology. 1981 Human cancer genes were discovered. 1982 Humulin, Genentech's human insulin drug produced by genetically engineered bacteria for the treatment of diabetes. 1985 Kary B. Mullis Discovered the polymerase chain reaction (PCR). 1988 The Human Genome Project began. 1993 FlavrSavr tomatoes, genetically modified plants engineered for longer shelf life, were marketed in USA. 1995 Haemophilus influenzae is the first bacterial genome sequence, which was completely determined. 1996 2000 Ian Wilmut Cloned the first mammal sheep Dolly. Pigs are the next animal cloned to help produce organs for human transplant. Golden Rice, modified to make vitamin A, promises to help third-world countries alleviate blindness. 2001 The sequence of the human genome is published in Science and Nature. 3. Fundamentals of biotechnology Biotechnology is scientific manipulation of living organisms like microorganisms, such as bacteria or yeasts, or biological substances, such as enzymes, to produce specific industrial or manufacturing processes. Biotechnology is “the manipulation of living organisms to produce goods and services”. Traditional biotechnologies include domesticating plants and animals and then selectively breeding them for specific characteristics. Traditional biotechnologies mean bake bread, brew alcoholic beverages. But recent developments in molecular biology have given biotechnology new meaning, new prominence, and new potential. Modern biotechnology is referring to biotechnological techniques for the manipulation of genetic material. The most obvious example is genetic engineering to create genetically modified/engineered organisms (GMOs/GEOs) through transgenic technology involving the insertion or deletion of genes. The major techniques of biotechnology are gene splicing and use of recombinant DNA. Genetic engineering, recombinant DNA technology, genetic modification/ manipulation (GM), gene splicing are terms that apply to the direct manipulation of an organism's genes. The term "genetic engineering" was coined in Jack Williamson's science fiction novel Dragon's Island, published in 1951. Two years before James Watson and Francis Crick showed that DNA could be the medium of transmission of genetic information. Genetic engineering uses the techniques of molecular cloning and transformation. 10 Created by XMLmind XSL-FO Converter. 1. Basis and history Biotechnology has applications in four major industrial areas, including health care (medical), crop production and agriculture, non food (industrial) uses of crops and other products (e.g. biodegradable plastics, vegetable oil, biofuels), and environmental uses. Applications include the production of certain drugs, synthetic hormones, and foodstuffs as well as the bioconversion of organic waste and the use of genetically altered bacteria. The goal of plant breeding is to combine desirable traits from different varieties of plants to produce plants of superior quality. This approach to improving crop production has been very successful over the years. Advances in scientific discovery and laboratory techniques during the last half of the 20th century led to the ability to manipulate the deoxyribonucleic acid (DNA) of organisms, which accelerated the process of plant improvement through the use of biotechnology. The ability to move genes into plants from other organisms, thereby producing new proteins in the plant, has resulted in significant achievements in plant biotechnology, that were not possible using traditional breeding practices. 4. Timeline of plant biotechnology 1700s: Naturalists identify hybrid plants. 1860s: Gregor Mendel discovers the rules of inheritance. 1900: European botanists begin to improve productivity of plants using theories based on Mendel’s work. 1922: Farmers purchase hybrid seed corn created by cross-breeding with two corn inbred lines. 1953: Structure of DNA is discovered. 1973: Genetic engineering, manipulation of bacterial DNA. 1983: The creation of antibiotic resistant GM tobacco. 1990: First successful field trial of GM cotton (herbicide resistant). 1994: FlavrSavr Tomato becomes the first GM food to be approved for sale. 1995: Herbicide resistant canola, corn. 2000: Herbicide resistant cotton, soybeans, sugar beet as well as insect or virus resistant corn, cotton, papaya, potato, squash, tomato approved in the U.S. 2001: ‘Golden rice’ which may help preventing millions of cases of blindness and death caused by Vitamin A and iron deficiencies undergoes continued testing (Figure 6.). 1.6. ábra - Figure 6.: The ‘Golden rice’ 11 Created by XMLmind XSL-FO Converter. 2. fejezet - 2. Biotechnology of asexual reproduction 1. Tissue culture Tissue culture is a biological technique in which fragments of plant or animal tissue or cells are transferred to an artificial environment, free of other organisms, in which they continue to survive and function for reproduction, chemical production and medical research. Plant tissue culture now has direct commercial applications as well as value in basic research into cell biology, genetics and biochemistry. Plant tissue culture techniques used to maintain or grow plant cells, tissues under sterile conditions on a nutrient culture medium. Explant is the part of an organism used in „in vitro" culture. Plant tissue cultures can be initiated from almost any part of a plant (Figure 7.). The parent plant must be healthy and free from diseases. Younger tissue contains a higher proportion of actively dividing cells and is more responsive to a callus initiation programme. Callus is an undifferentiated tissue that develops on the cutted plant surface or in tissue culture. Explants from several parts of large intact plants can be used to form callus. Callus formation is investigated and applied tissue culture procedures. Callus can be multiplied and later used to clone numerous whole plants. Nowadays callus is used to genetic engineering protocols after DNA has been inserted into cells, transgenic plants are then regenerated from transformed callus. 2.1. ábra - Figure 7.: Overview of plant tissue culture process The plant tissue culture required a very specialized laboratory and a sterile cabinet (laminar flow box). Physical factors, such as temperature, pH, light (quality and duration), and osmotic pressure are very important in tissue culture. The most commonly used culture media are based on the formulations of Murashige and Skoog medium (MS) which was invented by plant scientists Toshio Murashige and Folke K. Skoog in 1962. Two types of medium are used in tissue culture: the callus growth medium and the plant regeneration medium. The growth medium 12 Created by XMLmind XSL-FO Converter. 2. Biotechnology of asexual reproduction contains all the essential mineral ions required for growth and development of the callus. The plant tissue cultures media are generally contain macronutrients and micronutrients, organic supplements such as vitamins and amino acids (or other nitrogen supplements). Many plant cell cultures, as they are not photosynthetic, also require the addition of a fixed carbon source in the form of sugars (most often sucrose). One other vital component that must also be supplied is water, the principal biological solvent. As other undefined organic supplements, solidifying agents and hormones are in culture medium. The addition of suitable hormones can then induce root growth, and the plants can then be placed in soil and grown in the normal circumstances. Plant hormones are relatively small molecules that are at low tissue concentrations. The most commonly used plant hormones are cytokinins and auxins. Cytokinins stimulate the DNA synthesis, increase cell division and produce a delay response in undifferentiated tissue which is the formation of shoot. Auxins promote and regulate the growth and development of plants. Auxins are produced in the meristem of shoot tips. Auxins cause the cells below the shoot apex to expand or elongate, and this is what causes the plant to increase in height. The most common used auxins are 2,4-dichlorophenoxy-acetic acid (2,4-D), naphthaleneaacetic acid (NAA) and indoleacetic acid (IAA). Plant growth, development and organogenesis can be controlled with the manipulation of the levels and ratios of auxin and cytokinin. Plant tissue culture techniques include culture of cells, anthers, ovules and embryos, and protoplast isolation and fusion, cell selection and meristem and bud culture. Plant tissue culture applications include: • micropropagation using meristem and shoot culture to produce large numbers of identical individuals, • screening programmes of plant cells, • crossing distantly related species by protoplast fusion and regeneration of the novel hybrid, • production of dihaploid plants from haploid cultures to achieve homozygous lines more rapidly in breeding programmes, • as a tissue for transformation, followed by either short-term testing of genetic constructs or regeneration of transgenic plants, • removal of viruses by propagation from meristematic tissues. The exact conditions require initiating and sustaining plant cells in culture, or to regenerate intact plants from cultured cells, are different for each plant species. Each variety of a species will often have a particular set of cultural requirements. Despite all the knowledge that has been obtained about plant tissue culture during the twentieth century, these conditions have to be identified for each variety through experimentation. Technology of plant tissue culture (Figure 8.): 1. Selection of mother plant: healthy, identical. 2. Testing the mother plant: has to be pathogen-free. 3. Sterile starting culture: sterile isolates, inoculation, elimination of endogenous pathogens, DNA pattern for molecular identification. 4. Maintenance of starting culture: minimal conditions, genetic stability, genetic identity, pathogen-free. 5. Sterile micropropagation: inoculation and propagation cycles until the suitable amount of seed material, pathogen-free conditions. 6. Preparations for plantation: modification of physical parameters, helping of shoot elongation, in vitro rooting. 7. Plantation and in vivoculturing: acclimatization in glass houses, less light, higher temperature and humidity, in vivo propagation after the development of photo-synthesis, roots. 13 Created by XMLmind XSL-FO Converter. 2. Biotechnology of asexual reproduction 2.2. ábra - Figure 8.: Technology of plant tissue culture Critical points: • pathogen-free conditions • human error: changes, bad labelling or inoculation • failure of rooting • low surviving rate after plantation 2. Micropropagation Micropropagation is a special type of asexual propagation where a very small piece of tissue (shoot apex, leaf, or even an individual cell) is excised (cut-out) and placed in sterile (aseptic) culture in a petri dish or test tube, or tissue culture container which are containing a special culture medium. The major techniques of the micropropagation are meristem and shoot culture, axillary shoot production and somatic embryogenesis (Table 3.). In meristem culture the explant, up to 1 mm in length, may consist of the shoot apical meristem. An advantage of utilizing a small explant is the potential for eliminating pathogenic organisms that may be present in the donor plant. 2.1. táblázat - Table 3.: Advantages and limitations of different methods of micropropagation Plant regeneration via Advantages Limitations 14 Created by XMLmind XSL-FO Converter. 2. Biotechnology of asexual reproduction I. Axillary shoot production a) Shoot-tips b) Axillary and terminal buds Wide species applicability Unsuitable for virus elimination Phenotypically homogeneous Initial low multiplication rate Genetically stable Low survival rate High survival and growth rates Genetically stable c) Meristem-tips Phenotypically homogeneous Useful for pathogen elimination II. Adventitious shoot production Organogenesis Positive somaclonal variation Staged process involving callus-, meristerm induction, and organ differentiation Initial high multiplication rate III Somatic embryogenesis Developmentally zygotic embryos analogous to High cost of production Synthetic seed. Continuous production Circumvent long breeding cycles Circumvent self-incompatibility Decrease hybrid seed cost Be used conservation for germplasm 3. Meristerm cultrure Meristem isolation In case of the meristem isolation the size of the apex of meristem is critical. The smallest possible apex is used which is suitable for developing a plant. We generally use Murashige-Skoog (MS) medium. The growth regulators are auxins, cytokinins, e.g. naphtyl acetate, indolyl acetate, indolyl butirate, gibberellic acid, benzyl adenine, zeatine, kinetine. In this method there are some important conditions such as: temperature, time, light intensity, and wavelength. Meristerm cultrure’s possible applications are: • micropropagation • elimination of viruses • maintenance of plants • cryopreservation • experimental procedures Meristem isolation 15 Created by XMLmind XSL-FO Converter. 2. Biotechnology of asexual reproduction 1. The size of the apex of meristem is critical. The smallest possible apex is used which is suitable for developing a plant. 2. Medium generally used is Murashige-Skoog (MS). 3. The growth regulators are auxins, cytokinins, e.g. naphtyl acetate, indolyl acetate, indolyl butirate, gibberellic acid, benzyl adenine, zeatine, kinetine. 4. Conditions like temperature, time, light intensity, wavelength are important. 4. Shoot culture and adventitious shoot culture Shoot culture 1. The development of shoots without roots on medium under sterile conditions. 2. During propagation normal development is maintained: no dedifferentiation, no redifferentiation. 3. It is widespread method for in vitro gene banks. 4. It is genetically stable cloning technique. Adventitious shoot culture 1. Inoculates: parts containing meristem, parenchima, cambium, e.g. segments of leaves, stalks, roots, flower parts, embryos, cotyledon, epicotyl, hypocotyl, etc. 2. The younger parts are used, the easier the induction of organogenesis. 3. Sterility of medium is crucial. 4. Commonly used for plant regeneration. 16 Created by XMLmind XSL-FO Converter. 3. fejezet - 3. Biotechnology of sexual reproduction • Embryo culture • Microspore culture • Anther culture (androgenesis in vitro) • Culture of sexual organs • Culture of sexual tissues • Culture of sexual cells 1. Embryo culture Embryos can be used as explants to generate callus cultures or somatic embryos. Both immature and mature embryos can be used as explants. Immature, embryo-derived embryogenic callus is the most popular method of monocot plant regeneration. Embryo culture is an in vitro technique that has been used to save the hybrid products of fertilization when they might otherwise degenerate. In 1904 Hannig was the first who obtained viable plants from mature embryos of two crucifers that were isolated aseptically and grown on a mineral salt medium supplemented with sugar. In 1924, Dietrich cultured mature and immature embryos of various plant species. He reported that the mature embryos grew immediately, circumventing dormancy. The immature embryos germinated precociously without further embryo development. Zygotic embryo culture for interspecific hybridization was described by Laibach in 1925. He observed that seeds from interspecific crosses between Linum perenne L. x Linum austriacum L. were nonviable; however, if embryos were excised early during seed development and cultured in vitro, then embryo abortion was overcome. Datura hybrid embryos could be grown in culture on media containing coconut milk. This discovery ultimately led to understanding the importance of reduced N in the form of amino acids for embryo culture. Since the early 1940s, embryo culture has been used increasingly to understand the physical and nutritional requirements for embryonic development, bypass seed dormancy, shorten the breeding cycle, test seed viability, provide material for micropropagation, and rescue immature hybrid embryos from incompatible crosses. Medium composition of embryo culture: carbohydrates - sucrose nitrogen - in the form of NH4NO3 and KNO3 plant extracts - e. g. coconut milk, tomato juice growth regulators - abscisic acid, auxins, cytokinins agar - for solidification of the medium Aims and results of developed embryo culture • shortening of the ripening time of seeds, that results in fastest generation change • elimination of germ silence • elimination of sterile seed development • in all cases, when normal germination has difficulties in situ Culturing method 17 Created by XMLmind XSL-FO Converter. 3. Biotechnology of sexual reproduction The general method of embryo culture follows the following steps: • Healthy and mature donor plant is needed. • Surface sterilize with 0.01% Tween-20 for 15 min, rinse seeds several times with distilled water. • Finally rinse it with sterile distilled water. • Break seeds aseptically and isolate the embryo. • Culture embryo on callus proliferation medium. Murashige and Skoog (1962) the basal medium supplemened with different combinations and concentrations of vitamin, sugar, hormones and other growth adjuvants for callus proliferation and shoot regeneration. • Incubate the cultures at 20-25°C under a 16 h photoperiod of 2000 lux luminous intensity. • After two weeks of inoculation the embryo begins to swell on callus proliferation medium. Distinct callus growth is observed after 4 weeks. • After 8 weeks of inoculation transfer the callus on shoot regeneration medium. • Within 4 weeks of transfer into second medium the callus turns green and produces soft spongy tissue. Some of these tissues are differentiated into embryoids. • The embryoids produce cluster of budlets when subcultured onto shoot regeneration medium. The budlets grow into shoots and produce 2-3 leaf appendages within 12 weeks. • Thereafter, they are separated into individual shoots and then subcultured into a fresh medium of the same composition until shoots develop. 2. Microspore culture Microspores are immature pollen cells. Buds are surface sterilized and homogenized to release the microspores. In many microspore culture procedures, a heat treatment causes the microspores to undergo equal mitosis and essentially behave like egg cells and develop into an embryo. The microspore culture technique is so efficient that thousands of plants can be produced. Haploid tissue can be cultured in vitro by using pollen or anthers as an explant. Pollen contains the male gametophyte, which is termed the ‘microspore’. Both callus and embryos can be produced from pollen. Immature pollen can also be extracted from developing anthers and cultured directly, although this is a very time-consuming process. In microspore culture, the condition of the donor plant is of critical importance, as is the timing of isolation. Pretreatments, such as a cold treatment, are often found to increase the efficiency. Plant species can be divided into two groups, depending on whether they require the addition of plant growth regulators to the medium for pollen/anther culture; those that do also often require organic supplements, e.g. amino acids. Many of the cereals (rice, wheat, barley and maize) require medium supplemented with plant growth regulators for pollen/anther culture. Regeneration from microspore explants can be obtained by direct embryogenesis, or via a callus stage and subsequent embryogenesis. Anther culture (androgenesis in vitro) • isolation of pollen • induction of androgenesis in the microspore • maintenance on sterile medium • production of pollen haploids in vitro Androgenous microspores Capable of androgenesis (S or P pollens) - genetically suitable for developing an embryo then a whole plant. 18 Created by XMLmind XSL-FO Converter. 3. Biotechnology of sexual reproduction Induction of morphogenesis The totipotent cell need stress for the activation of the ontogenetic program: 2,4-D, cold or heat, osmotic stress. Anther culture: the technique • reduction of genotypic determination • environmental conditions • cold pre-treatment of donor flowers and inflorescence • stress treatment of anthers, microspores • composition of medium • incubation time and temperature • production of doubled haploids Application in genetics and plant breeding • opportunity for the reduction of ploidity: gametoclonal variability - production of plant without fertilisation • in haploids and doubled haploids recessive features can be manifested and selected phenotypically • inbreeding can reduce the production time • addition and substitution hybrids (e. g. Triticale) • fusion of monohaploids • production of homozygous transgenes • production of „super males” (e. g. asparagus) • genetically pure lines for marker selection 19 Created by XMLmind XSL-FO Converter. 4. fejezet - 4. Manipulating of DNA 1. Molecular cloning Gene cloning is the isolation and amplification of an individual gene sequence by insertion of that sequence into a bacterium where replicated. Molecular cloning refers to the procedure of isolating a defined DNA sequence and obtaining multiple copies of it in vivo. Cloning is frequently employed to amplify DNA fragments containing genes, but it can be used to amplify any DNA sequence such as promoters, non-coding sequences, chemically synthesised oligonucleotides and randomly fragmented DNA. Cloning is used in a wide array of biological experiments and technological applications such as large scale protein production. The process of gene cloning: • Isolation of the gene of interest. • Incorporation of the gene into a vector (small replicating DNA molecule, usually circular). • Transformation of this construct into the host cell. • Selection for the cells that have acquired the recombinant DNA molecule. • Multiplication of the recombinant vector within the host cell to produce a number of identical copies of the cloned gene. The key figure of gene cloning is the vector. Vector is a vehicle for transferring genetic material into a cell. There are four major types of vectors: • plasmids • viral vectors • cosmids • artificial chromosomes Plasmids are double-stranded DNA (dsDNA) molecules which are separate from a cell’s chromosomal DNA (extrachromosomal DNA). Plasmids usually are circular. Plasmids occur naturally in bacteria, yeast, and some higher eukaryotic cells. Size of the plasmids varied between a few thousand base pairs to more than 100 kilobases (kb). Plasmid DNA is duplicated before every cell division. Plasmids as transcription vectors, incubating bacteria with plasmids generates hundreds or thousands of copies of the vector within the bacteria in hours, and the vectors can be extracted from the bacteria, and the multiple cloning site can be cut by restriction enzymes to excise the hundredfold or thousandfold amplified insert. The promoter is the part of a transgene, determines where and when the transgene will function. The gene can be Tissue Specific or Constitutive. Restriction endonucleases recognize specific DNA sequences (4, 6 or more base-pairs) and cut DNA (Figure 9.). 4.1. ábra - Figure 9.: Restriction endonucleases cut DNA 20 Created by XMLmind XSL-FO Converter. 4. Manipulating of DNA A viral vector is a virus that has been modified to transduct specific genetic material into a cell, e.g., for gene therapy. A plasmid vector is made by splicing a DNA construct into a plasmid. Various techniques are used to transfect the cell. • adenovirus Adenoviral DNA does not integrate into the genome. It is not replicated during cell division. This limits their use in basic research, although adenoviral vectors are occasionally used in in vitro experiments. Their primary applications are in gene therapy and vaccination. • adeno-associated virus Adeno-associated virus (AAV) is nonpathogenic, nonenveloped, single-stranded DNA viruses in development as gene therapy vectors. AAV is a small virus that infects humans and some other primate species and can infect both dividing and non-dividing cells. It may incorporate its genome into that of the host cell, these features make AAV a very attractive candidate for creating viral vectors for gene therapy. • Tobacco mosaic virus (TMV) TMV is an RNA virus that infects plants, especially tobacco and other members of the family Solanaceae. The infection causes characteristic patterns (mottling and discoloration) on the leaves (thence the name). TMV was the first virus to be discovered. • cytomegalovirus • bacteriophage Bacteriophage is any one of a number of viruses that infect bacteria, by injecting genetic material, which they carry enclosed in an outer protein capsid. The genetic material can be RNA or DNA (single-strand or doublestrand) with either circular or linear arrangement. Transient and stable transformation In transient transformation the transfected DNA is not integrated into host chromosome. DNA is transferred into a recipient cell in order to obtain a temporary but high level of expression of the target gene. Stable transformation is also called permanent transformation. By the stable transformation the transferred DNA is integrated (inserted) into chromosomal DNA and the genetics of recipient cells is permanently changed. 2. Genetic transformation The genetic transformation is a process by which the genetic material carried by an individual cell is altered by the incorporation of foreign (exogenous) DNA into its genome. The genetic transformation is the genetic 21 Created by XMLmind XSL-FO Converter. 4. Manipulating of DNA alteration of a cell resulting from the uptake, genomic incorporation, and expression of foreign genetic material (DNA). Transduction - is the process by which DNA is transferred from one bacterium to another by a virus. It also refers to the process whereby foreign DNA is introduced into another cell via a viral vector. Transduction does not require cell-to-cell contact, which occurs in conjugation. Transfection is a transformation of eukaryotic cells in tissue culture. RNA also be transferred into cells using similar methods, but this does not normally produce heritable change, so is not true transformation. Organisms, genetically engineered by inserting a gene from another organism, are called transgenic organisms. Transformation is usually more difficult with multicellular organisms, such as plants. Transformation of higher organisms is to transform an individual plant cell and then regenerate it into a whole organism. The whole plant can grow from a single cell (totipotent nature of plant cells). Genetic transformation occurs naturally. Bacteria and viruses carry DNA (or RNA) into an organism and cause profound changes. Agrobacterium tumefaciens and Agrobacterium rhizogenes (soil bacteria) examples of natural transformation systems. The two types of genetic transformation methods are in plants: 1. vector-mediated or indirect gene transformation 2. direct gene transformation Various vectors are used in plant transformation. 1. Agrobacterium mediated 2. Virus mediated 3. Agrobacterium mediated gene transformation Agrobacterium tumefaciens is a Gram negative, soil bacterium, which is a widespread naturally occurring soil bacterium that causes crown gall, and has the ability to introduce new genetic material into the plant cell. Fridiano Cavara identified bacilloid bacterium, as a casual agent of crown gall of grape in 1897. This organism is Agrobacterium vitis, causing the growth of neoplastic tumors on the stem and crown of grapevines and inducing necrotic lesions on grape roots. The genetic material that is introduced is called T DNA (transferred DNA) which is located on a Ti plasmid (tumour-inducing plasmid). A Ti plasmid is a circular piece of DNA found in almost all bacteria. The natural ability of Agrobacterium tumefaciens to transfer parts of its genetic material to plant cells that has thus been used as a tool for genetically engineering plants. The bacterium is used as a means of transporting foreign genes into plants (vector). To do this, the bacterial T-DNA is cut out of the bacterial Ti-plasmid (tumor indicated) and replaced with the desired foreign gene (Figure 10.). Any desired genes, such as insecticidal toxin genes, virus or herbicide-resistance genes, can be engineered into the bacterial DNA and thereby inserted into the plant genome. 4.2. ábra - Figure 10.: Agrobacterium-mediated transformation 22 Created by XMLmind XSL-FO Converter. 4. Manipulating of DNA Several components of the Ti plasmid enable effective transfer of the genes of interest into the plant cells: • T-DNA border sequences, which demarcate the DNA segment (T-DNA) to be transferred into the plant genome. • Vir genes (virulence genes), which are required for transferring the T-DNA region to the plant but are not themselves transferred. • Modified T-DNA region where the genes that cause crown gall formation are removed and replaced with the genes of interest. Genes in the T-DNA (Figure 11.) 4.3. ábra - Figure 11.: Genes in the T-DNA Hormones 23 Created by XMLmind XSL-FO Converter. 4. Manipulating of DNA In order to cause gall formation, the T-DNA encodes genes for the production of auxin or indole-3-acetic acid via the IAM pathway. This biosynthetic pathway is not used in many plants for the production of auxin, so it means the plant has no molecular means of regulating it and auxin will be produced constitutively. Genes for the production of cytokinins are also expressed. This stimulates cell proliferation and gall formation. Opines The T-DNA contains genes for encoding enzymes that cause the plant to create specialized amino acids which the bacteria can metabolize called opines. Opines are a class of chemicals that serve as a source of nitrogen for A. tumefaciens, but not for most other organisms. The specific type of opine produced by A. tumefaciens C58 infected plants is nopaline. Two nopaline type Ti plasmids, pTi-SAKURA and pTiC58, were fully sequenced. A. tumefaciens C58, the first fully sequenced pathovar was first isolated from a cherry tree crown gall. Then the genome was simultaneously sequenced. The genome of A. tumefaciens C58 consists of a circular chromosome, two plasmids, and a linear chromosome. The presence of a covalently bonded circular chromosome is common to Bacteria, with few exceptions. However, the presence of both a single circular chromosome and single linear chromosome is unique to a group in this genus. The two plasmids are pTiC58, responsible for the processes involved in virulence, and pAtC58, coined the “cryptic” plasmid. The pAtC58 plasmid has been shown to be involved in the metabolism of opines and to conjugate with other bacteria in the absence of the pTiC58 plasmid. If the pTi plasmid is removed, the tumor growth that is the means of classifying this species of bacteria does not occur. The Agrobacterium-mediated transformation process involves a number of steps: 1. Isolation of the genes of interest from the source organism. 2. Development of a functional transgenic construct including: • the gene of interest • promoters to drive expression • codon modification, if needed to increase successful protein production • marker genes to facilitate tracking of the introduced genes in the host plant 3. Insertion of the transgene into the Ti-plasmid. 4. Introduction of the T-DNA-containing-plasmid into Agrobacterium. 5. Mixture of the transformed Agrobacterium with plant cells to allow transfer of T-DNA into plant chromosome (Figure 12 .). 6. Regeneration of the transformed cells into genetically modified (GM) plants. 7. Testing for trait performance or transgene expression at lab, greenhouse and field level. 4.4. ábra - Figure 12.: Plants that have undergone transformation with Agrobacterium 24 Created by XMLmind XSL-FO Converter. 4. Manipulating of DNA Transferring genes with agrobacteria is a commonly used and reliable method. It works especially well for dicotelydenous plants like potato, tomato, and tobacco. Agrobacteria are less suitable for introducing foreign genes to crops like wheat and maize. Virus mediated Viruses are highly adapted to the process of gene transfer. Viral vectors have the ability to transfer DNA to a high fraction of cells. 4. Direct gene transformation Methods of gene transfer A variety of direct gene transfer strategies have been developed during the last decade, which can be grouped into the two major categories: the chemical and physical gene transfer methods. 5. Chemical methods PEG, Polyvinil alcohol, DEAE-Dextran, Calcium-phosphate mediated PEG-mediated gene transformation The PEG (polyethylene glycol)-mediated transformation is simple and efficient. It was the first methods to report the successful integration of foreign genes into plant cells. Polyethylene glycol can be used to induce DNA uptake into plant protoplasts. One advantages of PEG-mediated DNA uptake is that it does not seem to be species-specific, as is Agrobacterium mediated transformation. PEG is also useful tool for the functional analysis of genes via transient expression. The availability of efficient and reproducible protoplast culture methods limits PEG mediated transformation in Gramineae. 25 Created by XMLmind XSL-FO Converter. 4. Manipulating of DNA DEAE-Dextran transformation One of the oldest chemical techniques is the DEAE-Dextran transfection, which was developed to transfer DNA or RNA to cultured cells. This reagent is generally used for transient expression studies, because long-term stable transfections are less successful. In the standard protocol, positively charged DEAE-dextran polymer binds with the negatively charged nucleic acids. The complex adsorbs to negatively charged cellular membranes and is taken into the cell, presumably by endocytosis. The addition of chloroquine may improve transfection in cells by reducing degradation of the DNA in the lysosome. The concentration and the exposure time of the chloroquine are very important, because the chloroquine is cytotoxic. Improvement of transfection can be increased by adding glycerol or dimethyl sulfoxide (DMSO) to the cells following exposure to DEAE-Dextran and DNA. It is more reproducible than calcium phosphate transfection but only works in a select few cell lines. Calcium-phosphate mediated transformation The Calcium-phosphate mediated transfection technique relies on precipitates of plasmid DNA formed by its interaction with calcium ions. It is a very inexpensive and simple technique to perform. Plasmid DNA is mixed in a solution of calcium chloride, and then is added to a phosphate- buffered solution. After 20 minutes, the solution added directly to the cells in culture. Transfection efficiencies can be improved in some cell lines by 'shocking' the cells with DMSO or glycerol. Cells can be either transiently or stably transfected using this technique. Levels of stable transfectants can be improved using bis-hydroxyethylaminoethansulfonate (BES). The DNA precipitates enter the cell by endocytosis. Although this technique has minimal cellular toxicity, and is both simple and inexpensive, the low level of transgene expression prompted development of other techniques. 6. Physical gene transfer methods Electroporation Electroporation is a simple, rapid and efficient technique for introducing DNA, RNA, proteins into cells and tissues. Electroporation involves the creation of transient, electrically induced membrane pores through which molecules pass into cytoplasmic and nuclear compartments. Because electroporation involves physical process acting on cell membranes, it is effective with a wide range of cell types including bacterial, fungal, plant and animal cells. Electroporation is used to transiently or stably increase gene expression. Low voltage long pulses method causes transient transformation. High voltage short pulses method causes stable transformation. These techniques are increasingly used in clinical applications including gene therapy, cancer drug delivery and vaccination. Microinjection Microinjection is a simple mechanical process. Microinjection delivers plasmid DNA directly into the cell's nucleus. Using the light microscope, a glass pipette is guided into the nucleus and a small amount of DNA or RNA injected. Microinjection is normally performed under a specialized optical microscope setup called a micromanipulator. The process is frequently used as a vector in genetic engineering and transgenics to insert genetic material into a single cell. Microinjection can also be used in the cloning of organisms, and in the study of cell biology and viruses. Microcapillary and microscopic devices are used to deliver DNA into a protoplast. Both cytoplasmic and lysosomal degradation of the injected material is avoided and efficient gene expression can be expected from the surviving cells. Unfortunately, this technique is extremely labor intensive and requires well isolated cells. Particle bombardment (Gene gun) method The gene gun is part of the gene transfer method called the biolistic (also known as biobalistic or particle bombardment) method and it was initially designed to transform the plants; however, many other types of organisms are successfully transformed. In this method, DNA or RNA adhere to biological inert particles (such as gold or tungsten). By this method, DNA-particle complex is put on the top location of target tissue in a vacuum condition and accelerated by powerful shot to the tissue, then DNA will be effectively introduce into the target cells. The efficiency of the gene gun transfer could be depended on the following factors: cell type, cell growth condition, culture medium, gene gun ammunition type, gene gun settings and the experimental experiences, etc (Figure 13.). 26 Created by XMLmind XSL-FO Converter. 4. Manipulating of DNA 4.5. ábra - Figure 13.: The scheme of the GENEBOOSTER (Gene Gun) and the principles of genetic bombardment Some cells, tissues and intracellular organelles are impermeable to foreign DNA, especially plant cells. biolistics, including particle bombardment, is a commonly used method for genetic transformation of plants and other organisms. Each of these methods has merits and limitations and is used in specific situations. At this time there is no single technique that is suitable for all species. However, developments of plant molecular biology and genomics now give us access to knowledge and understanding of plant genomes and the possibility of modifying them. There are two most powerful technologies for transferring gene into plants: Agrobacterium-mediated transformation and biolistics. As plants have cell wall, the biolistics is very useful in the plant gene transfer. 27 Created by XMLmind XSL-FO Converter. 5. fejezet - 5. Plant regeneration/molecular selection 1. In vitro plant regeneration In vitro plant regenaration accomplished via organogenesis or somatic embryogenesis. The regeneration of plants from tissue cultures was first reported in maize (Zea mays L.) by Green and Philips in 1975, who utilised immature embryos as the tissue source. Using the same tissue system, Springer et al. in 1979 demonstrated that plant regeneration took place by means of organogenesis. Rice et al. in 1978 found that plant regeneration could also occur by somatic embryogenesis. Both types of regeneration arise from hard, white or yellow callus, which can be clearly distinguished from the granular, greyish-yellow, translucent callus that is incapable of plant regeneration. The regeneration from cell and tissue cultures has been limited to a few specific genotype and medium combinations. Medium improvements boosted the average regeneration, but genotypic differences remained. In the following figure we summerized the alternative ways of the plant regeneration (Figure 14.). 5.1. ábra - Figure 14.: Alternative ways of plant regenaration 2. Influence of carbon sources on plant regeneration in wheat The haploid cultures provide a rapid route to complete homozigosity and to expression of recessive genes. In wheat, the low level of callus induction from microspores and subsequent plant regeneration, and high percentage of albino plants in vitro limited the application of haploids in plant breeding and genetic research for cereal crops. Most success in haploid production through anther culture has been obtained by manipulating the growth conditions of anther culture donor plants, the culture medium, and culture conditions. 28 Created by XMLmind XSL-FO Converter. 5. Plant regeneration/molecular selection The other major limitation to wider practical application of anther culture is that green plants occur too infrequently. Researchers have therefore shifted their efforts to investigating the influence of medium components on the proportion of green plants produced. A spring wheat cultivar, 'Pavon 76' as donor material were grown in a controlled growth chamber equipped with mixed fluorescent and incandescent lights which provided 350 to 400 μmol m-2 s-1 at the plot level in a 16-h photoperiod. The day/night temperatures were 20/15 °C. The first two to three spikes from each plant were collected when the owns were just visible from the flag leaf shealth. The tillers were sampled when the microspores from the anthers in the central portion of the spikes were at the mid to late uninucleate stages. The sampled spikes were placed in flasks containing tap water and stored in a dark refrigerator at - 4 °C for 48 h of cold pretreatment. Anthers with microspores at the uninucleate stage were excised with fine forceps, after the spikes were surface sterilized with 800 ml L-1 ethanol, and plated aseptically on the liquid induction medium. Liquid potato 4 (P4) medium can be used as a standard induction medium. Spikes were selected at random from a bulk from which 100 anthers were excised for each petri dish, with four replicates per treatment. After anther plating, the dishes were sealed with parafilm and placed in a darken incubator at 26 to 28 °C for ~ 40 d. The callus induction frequency was the number of calli obtained from 100 anthers plated 40 d following induction. Embryoids induced from the anthers were transferrred to a '190-2' medium for plant regeneration when they reached 1 mm in diameter. The standard regeneration medium contained 30 gL-1 sucrose and was solified using 6 gL-1 agar. No plant growth regulator (PGR) was included in the regeneration medium. The time of transfer was based on the size of the calli that developed between 30 to 40 d after anther plating. Calli were cultured at room temperature with 150 to 180 µmol m-2 s-1 illumination provided by fluorescent lights in a 16-h photoperiod. After 30 d of regeneration culture the number of calli that developed into albino or green plants was recorded. Plant regeneration frequency was the number of calli producing plantlets per 100 calli transferred. The green plant regeneration frequency was number of calli producing green plantlets divided by the number of total calli producing either green or albino plants. Study was conducted to determine the influence of several medium components on the percentage of green plants from wheat anther culture. The study was designed to examine the effects of four sucrose concentrations (45, 60, 75 and 90 gL -1) and four maltose concentrations (65, 100, 135, 170 gL-1) on callus induction, plant regeneration and green plant proportions. Anthers of Pavon 76 were cultured on these induction media and embryoids induced were transferred to the standard regeneration medium to compare for differences in green plant percentage and the influence of carbon sources on it. Because sucrose is rapidly degraded into fructose and glucose in P4 induction medium, remarkably increasing medium osmolality, higher concentrations of sucrose than 90 gL-1 were not included in this experiment. The callus induction response to sucrose for v. 'Pavon 76' was higher at all sucrose levels than that of maltose (Table 4.). 5.2. ábra - Table 4.: Induction and regeneration in vitro with different carbon sources in media. 29 Created by XMLmind XSL-FO Converter. 5. Plant regeneration/molecular selection Pavon 76 showed basically linear responses to sucrose and maltose concentrations. Some trends emerged from the response for callus induction across increasing sucrose and maltose concentrations: (i) the callus induction increased dramatically for both induction media when either sucrose or maltose were increased from 45 to 90 gL-1 and from 65 to 170 gL-1, respectively, (ii) callus induction from Pavon 76 can be increased at sucrose levels > 90 gL-1 and at maltose levels > 135 gL-1 significantly in the induction medium (Figure 15.). 5.3. ábra - Figure 15.: Induction frequency (%) in parallel with sugar concentrations 66,0 % embryoids from the 170 gL-1 and 64,5 % embryoids from the 135 gL-1 maltose medium produced green plants (Figure 16.), whereas only 42,9 % embryoids from the 90 gL -1 sucrose medium produced green plants (Figure 17.). 5.4. ábra - Figure 16.: Proportions of plant regeneration in maltose 30 Created by XMLmind XSL-FO Converter. 5. Plant regeneration/molecular selection 5.5. ábra - Figure 17.: Proportions of plant regeneration in sucrose Sugar has two functions in culture media; as a carbon source and as an osmotic regulator. Since the sugar content of media was found to change little during the culture period, the major function of sugar may be as regulator of medium osmolality. Thus, the difference in green plant percentages between maltose concentrations demonstrates the effect of medium osmolality on albinism. Sucrose in induction media is rapidly hydrolyzed to fructose and glucose, increasing the medium osmolality, whereas no detectable osmotic change occurs in maltose containing medium. A distinct response pattern types were found for plant regeneration across increasing concentration in the induction medium; the plant regeneration for 100 calli transferred was 50,4-51,4 % in medium containing various amount of sucrose and 40.9-48.0 % for that with maltose. However, the green plant proportion showed a linear response with concentration and was the highest one for sucrose (22,3 green plant number/100 calli transferred) at 90 gL-1, and that for maltose (31,7 %) at 170 gL-1, which were not significantly different from 75 gL-1 sucrose and 135 gL-1 maltose concentration, respectively. Results from this study have profound implications on the choice of carbon sources and concentrations. Many of the reported effects of medium modifications and pretreatments may be related to osmotic potential. In addition, if the hypothesis about the importance of medium osmotic potential is correct, more attention also should be paid to establishing the optimal osmotic potential for regeneration media. Currently, most researchers use 90 gL 1 sucrose in incubation media, but only 30 gL-1 sucrose in regeneration media. Because of the hydrolysis of sucrose in induction media, difference in osmotic potentials between the induction and regeneration media during the transition phase of culture is much greater than expected. This difference also may have a significant impact on green plant percentage. 3. Molecular marker Molecular marker is a gene or DNA sequence with a known location on a chromosome that can be used to identify individuals or species. It can be described as a variation (which may arise due to mutation or alteration in the genomic loci) that can be observed. 31 Created by XMLmind XSL-FO Converter. 5. Plant regeneration/molecular selection The use of molecular markers to enhance plant breeding efforts is being widely studied. Even though marker – assisted selection now plays a prominent role in the field of plant breeding, examples of successful, practical outcomes are rare. It is clear that DNA markers hold great promise, but realizing that promise remains elusive. Despite innovations like better marker systems and improved genetic mapping strategies, most marker associations are not sufficiently robust for successful marker-assisted selection. Molecular breeders must reassess their research programs so that DNA marker work leads to useful selection tools and valuable germplasm. As molecular breeders adapt more rigorous experimental guidelines and ambitious goals, they also need to integrate the growing body of knowledge from genomics and bioinformatics. A genetic marker may be a short DNA sequence, such as a sequence surrounding a single base-pair change (single nucleotide polymorphism, SNP), or a long one, like minisatellites. Three common technologies used as molecular markers: • restriction fragment length polymorphisms • simple sequence repeats • single nucleotide polymorphisms Types of genetic markers: • RFLP (or Restriction Fragment Length Polymorphism) • SSLP (or Simple Sequence Length Polymorphism) • AFLP (or Amplified Fragment Length Polymorphism) • RAPD (or Random Amplification of Polymorphic DNA) • VNTR (or Variable Number Tandem Repeat) • Microsatellite polymorphism, SSR (or Simple Sequence Repeat) • SNP (or Single Nucleotide Polymorphism) • STR (or Short Tandem Repeat) • SFP (or Single Feature Polymorphism) • DArT (or Diversity Arrays Technology) • RAD markers (or Restriction Site Associated DNA markers) RFLP (or Restriction fragment length polymorphism) A site in a genome, where the distance between two restriction sites varies among different individuals. These sites are identified by restriction enzyme digests of chromosomal DNA, and the use of Southern blotting to identify the specific fragments. It requires a radioactive probe. AFLP markers provide a larger number of polymorphic bands. It is based GS, which are not sensitive to ecological factors and can be easily detected. It is an efficient technique for comparing genetic similarities or differences among different genotypes. RAPD (or Random Amplification of Polymorphic DNA) This method based on PCR reactions and has several advantages in contrast with other mapping methods. RAPD analysis quick to perform, the DNA does not need to be as pure as for other mapping techniques, there is no need for species specific genomic or cDNA derived probes, and radioactive materials are not involved. From a technical point of view, this is the simplest PCR method using arbitrary primers. In our experiment, we extracted DNA samples from the leaves of maize plants being in 3-4 leaves stadium. Extraction was carried out by the Potassium Acetate Procedure. This strategy of plant DNA isolation was introduced by Dellaporta et al. (1983). Its key step relies on the simultaneous precipitation of proteins and polysaccharides by high concentration of potassium acetate in the presence of SDS. 32 Created by XMLmind XSL-FO Converter. 5. Plant regeneration/molecular selection Polymerase chain reaction (PCR) Polymerase chain reaction (PCR) enables researchers to produce millions of copies of a specific DNA sequence in approximately two hours. This automated process bypasses the need to use bacteria for amplifying DNA developed by Kary Mullis in the 1980s. PCR is based on using the ability of DNA polymerase to synthesize new strand of DNA complementary to the offered template strand. Because DNA polymerase can add a nucleotide only onto a preexisting 3'-OH group, it needs a primer to which it can add the first nucleotide. This requirement makes it possible to delineate a specific region of template sequence that the researcher wants to amplify. At the end of the PCR reaction, the specific sequence will be accumulated in billions of copies (amplicons). This method is based on the in vitro amplification of DNA with the help of specific or arbitrary and Taq polymerase. Starting from a very low amount of template DNA (in the nanogram range), millions of copies of one or more particular target DNA fragments are produced, which can be electrophoresed and visualized by staining or autoradiography. PCR is characterized by its high speed, selectivity and sensitivity. The selectivity of the reaction is determined by the choice of the primers. Primers are single-stranded pieces of DNA with sequence complementary to template sequences flanking the targeted region. For exponential amplification primers must anneal in opposite directions, so that their 3’ ends face the target. Amplification is most efficient when the two promoter binding sites are not further apart than 4 kb. The PCR requires the following components: DNA template - the sample DNA that contains the target sequence. At the beginning of the reaction, high temperature is applied to the original double-stranded DNA molecule to separate the strands from each other. DNA polymerase - a type of enzyme that synthesizes new strands of DNA complementary to the target sequence. The first and most commonly used of these enzymes is Taq DNA polymerase (from Thermis aquaticus), whereas Pfu DNA polymerase (from Pyrococcus furiosus) is used widely because of its higher fidelity when copying DNA. Primers - short pieces of single-stranded DNA that are complementary to the target sequence. The polymerase begins synthesizing new DNA from the end of the primer. Nucleotides (dNTPs or deoxynucleotide triphosphates) - single units of the bases A, T, G, and C, which are essentially "building blocks" for new DNA strands. RT-PCR (Reverse Transcription PCR) is PCR preceded with conversion of sample RNA into cDNA with enzyme reverse transcriptase. Applications of PCR: cloning, genetic engineering, sequencing PCR can be performed in a volume of 20μl containing 50 mM MgCl2, 10 mM dNTP mix, 15 μM primer, Taq polymerase (5 unit/μl), 20ng /ml DNA. Amplification may be performed in Programmable Thermal Cycler. The thermal cycle are: 94°C (2min) for initial strand separation, then 40 cycles at 37°C (15 sec) for annealing, 72°C (30 sec) for extension. Amplification products can be analyzed by electrophoresis in 1,5% agarose (Reanal) gels and can be detected by staining with ethidium bromide. Marker Assisted Selection/ Marker Aided Selection (MAS) The use of molecular markers to enhance plant breeding efforts is being widely studied. Even though marker – assisted selection now plays a prominent role in the field of plant breeding, examples of successful, practical outcomes are rare. It is clear that DNA markers hold great promise, but realizing that promise remains elusive. Despite innovations like better marker systems and improved genetic mapping strategies, most marker associations are not sufficiently robust for successful marker-assisted selection. Molecular breeders must reassess their research programs so that DNA marker work leads to useful selection tools and valuable gerplasm. As molecular breeders adapt more rigorous experimental guidelines and ambitious goals, they also need to integrate the growing body of knowledge from genomics and bioinformatics. • Marker is used for indirect selection of a genetic determinant of a trait or interest (i.e. productivity, disease resistance, abiotic stress tolerance, and/or quality) • Many different genes can influence a desirable trait in plant breeding. Molecular markers or DNA fingerprinting can map thousand of genes. 33 Created by XMLmind XSL-FO Converter. 5. Plant regeneration/molecular selection • Screening large populations of plants (based presence/absence of a certain gene → no need for visual identification of the expressed trait) 4. Cryopreservation Cryopreservation – storage in liquid nitrogen – is an ideal solution for long term potato maintenance. This method seems especially suitable for preservation of rarely used clones. According to researches, cryopreservation of shoot tips proved to be useful in case of long term storage of vegetatively multi-plicapable embryonic plant plasms. For the preservation of clones isolated shoot tips have an advantage, since they can be easily regenerated into plants identical with the initial prototype material. We can find a few references in literature to the freezing of certain Solanum tuberosum genotypes and of a few Solanum species and also concerning the improvement of the method adapted to varieties Freezing at a degree of -196°C was applied for more than 40 species and their different tissues and survival was observed. In most cases cryoprotectant additives were used in the course of the freezing procedure, most often as single components. Vitrification is an alternative method used for cryopreservation and can be applied with success for cells, protoplasts and shooting tips of different plant species. In case of meristems the aim of preserving the whole macroscopic size structure is to achieve re-growth without adventitive organogenesis. In case of potato and date-palm meristems the survival of meristems increased when prior to their treatment they were placed into standard medium for 1-3 days. Pre-growth in the presence of cryoprotectants is quite often needed. There is no general standard for freezing procedures; both ultra-rapid freezing and controlled, slow pace freezing can be used depending on species. Artichoke (Solanum tuberosum L.) and potato meristems survived being directly dipped into liquid nitrogen as well. We can observe new growth even following slow cooling (0,2-0,3°C min-1-35°C) of meristems originating from in vitro potato plantlets. The type of development following defrosting depends on the method of freezing. Potato meristems show callus formation following ultra-rapid freezing. On the contrary they show direct re-growth following slow freezing. The vitrifification method was developed not too long ago for cell, suspension and protoplast cultures of different plant varieties, for somatic embryos and meristems. In most cases the treatment of meristems with vitrification solution happens in two phases: first the meristems are placed in cryoprotective solutions of low concentration then the concentration is increased until reaching the desired degree of cell shrinkage. High concentration of PVS 2 reduces the survival of potato meristems similar to the vitrification of sweet potato. To achieve an optimal vitrification precisely timed pretreatments in the presence of cryoprotectants of different concentrations and of different chemical structures are needed. Cryopreservation seems to be as an applicable solution for maintenance of potato germplasms. The protocol described above is suitable for vitrification and preserve meristems. With the histo-cytological study, we can observe the modifications at the cellular level and understand the adaptive mechanism to low temperatures. In the control potato meristem tissue we found a number of meristematic cells with a gradient of differentiation. After freezing there were a lot of vacuolated cells and some of them showed broken cell wall and plasmolysis. We observed thickening of cell wall with sinuous aspect in the frozen-thawed meristem with ruptured cuticle and epidermal layer. When freezing meristem, the objective is to preserve the whole structure and to obtain its re-growth without adventitious organogenesis. Surviving cells were mainly located at the superficial layers of apical dome and central region of foliar primordia, mitosis and cell proliferation could be detectable. In Hungary there is a tradition of in vitro storage of potato germplasmm and pollen preservation of different cereal species. 5.6. ábra - Figure 18. Figure 19. 34 Created by XMLmind XSL-FO Converter. 5. Plant regeneration/molecular selection Figure 18.: Structure of the control potato (Adagin) sample (longitudinal section of the apex). Magnification: 250x Figure 19.: Histological structure of the apex after freezing. Magnification: 250x Figure 18 shows the structure of a control sample on a longitudinal section of the apex, indicating the apical dome with two differently aged foliar primordia. The section shows the characteristics of meristematic cells (high nucleoplasmic ratio, dense cytoplasm with small vacuoles). There is a gradient of differentiation from the superficial layers to the central region, with a lower nucleoplasmic ratio and more accentuated vacuolisation. No polysaccharide reserves can be observed. Figures 19 (apex) and 20 (meristematic cells) demonstrate the state of the structures after freezing. Cellular heterogeneity is a conspicuous phenomenon. Some cells conserved their meristematic characters (in the cellular layers corresponding to the meristem itself). In the underlying zone the cells are more vacuolated and some are damaged, exhibiting broken cell walls and cells where the cytoplasrn has contracted away from the cell wall. Figure 21 shows apices during recovery (4-6 days after thawing). The thickening of the cell wall can be observed in some cells in various areas, and from the 6th day onwards both the meristematic and the non-meristematic cells had a sinuous appearance. 5.7. ábra - Figure 20. Figure 21. Figure 20.: Histological structure of meristematic cells after freezing. Magnification: 400x Figure 21.: Histological evaluation of apecis during recovery (4-6 days after thawing). Magnification: 400x On the phase-contrast micrograph massive damage to a frozen and thawed meristem can be observed (Figure 22.). A number of cells are ruptured with the total loss of the protoplast; the cuticle and epidermal layer are also ruptured. Damaged cells do not retain the toluidine blue stain. Figure 23 shows various types of damage in a section through portions of several damaged cells. The plasmalemmas have pulled away from the cell wall and there is evidence of the leakage of cytoplasmic materials through it. In many cases the protoplasts have completely broken down and are separated from the cell wall in places randomly scattered throughout the thawed explant. During recovery many changes can be observed in the apices. Meristematic cells are located around a central zone of damaged cells (cellular zonation). 35 Created by XMLmind XSL-FO Converter. 5. Plant regeneration/molecular selection 5.8. ábra - Figure 22. Figure 23. Figure 22.: Phase-contrast micrograph of a frozen and thawed meristem. Magnification: 400x Figure 23.: Section through portions of damaged cells. Magnification: 400x In the leaf primordia surviving and dividing cells are mainly located in the centre. Figure 24 shows outer cells without a cytoplasm, while the interior cells divide fastest. In the apical dome the first mitosis occurs in the superficial layer. Images of apices after thawing and culturing in MSTo medium can be seen in Figure 25 which illustrates the structure of the apical dome, demonstrating mitosis and cell proliferation. 5.9. ábra - Figure 24. Figure 25. Figure 24.: Histological evaluation of apices during recovery. Magnification: 250x Figure 25.: Apices after thawing and culturing in MSTo medium. Magnification: 250x Figure 26 shows that although the outer cells are damaged, the layer under them is developing rapidly, and a number of cells contain starch grains. The synthesis of starch increased in intensity mostly in cells in the underlying zone, and despite the damage the cells retained a very meristematic appearance. A section through the leaf primordium and dome of an explant that survived freezing and thawing revealed a number of large 'vesicles' in the damaged cells, together with darkly-staining nuclei and cytoplasm. These 'vesicles' are presumably related to ice-crystal formation, which obscures most of the protoplast detail in Figure 27. 5.10. ábra - Figure 26. Figure 27. 36 Created by XMLmind XSL-FO Converter. 5. Plant regeneration/molecular selection Figure 26.: Richness in starch content in the underlying zone of the meristem. Magnification: 400x Figure 27.: Section through a leaf primordium and dome an explant that survived freezing and thawing. Magnification: 250x Somatic embryogenesis (secondary cell division) can be observed in Figure 28 which shows apices (meristems) with apical domes and leaf primordia. Figure 29 shows the histological structure of a non-recovered apex, in which most cells are highly vacuolated, the majority of the cells have condensed a poorly stained cytoplasm and the nuclei are only just visible. 5.11. ábra - Figure 28. Figure 29. Figure 28.: Histological evaluation after thawing. Magnification: 250x Figure 29.: Histological structure of a noon-recovered apex. Magnification: 400x When freezing meristems, the objective is to preserve the whole structure and to induce re-growth without adventitious organogenesis. Surviving cells mainly located in the superficial layers of the apical dome and in the central region of foliar primordia, where mitosis and cell proliferation could be detected. At present cryopreservation, which basically means keeping the explants in liquid nitrogen, is a possibitity to guarantee the safe long-term preservation of plant germplasm. In Hungary there is a tradition of in vitro storage of potato germplasm and pollen preservation for various cereal species. 37 Created by XMLmind XSL-FO Converter. 6. fejezet - 6. Genetically modified plants (GM) Currently molecular engineering exceeds biotechnological laboratories’ frames, because it is pursued in large scale industries. In the commerce there are living organisms especially plants which are created by transformation. They can adapt to unfavorable climate conditions perfectly, and resistant to the pest/diseases and/or produce more and better quality of foods. Especially successful is therapeutic applications of gene technology. Besides advantages, we may not forget that transgenic plants are going out to the environment, it can cause genetic-ecological risk which has never been seen. We usually speak about damage of biodiversity. We do not know the effect of new or modified proteins which would be appeared in our nutriment. Appearance and speading out of genetically modified organisms set off discussion in sciences and in public opinion. Using gene technology methods in plant selection and animal breeding have plenty of advantages although at the same time their irresponsible application could have danger for living organisims. Because danger or safe of genetic transformation have not been proved, it is required to control by quantitative analysis the foodstuff and animal feed originated from GM organisims. In spite of speading of GMO in laboratories they can not been able to prove the effects of GMOs’. That is why both scientists and consumers think that GMO can be dangerous for heath and environmental point of view. Genetically modified or transgenic plants are plants which DNA is modified using genetic engineering techniques. In most cases the aim is to introduce a new trait to the plant which does not occur naturally in this species. DNA integrates into the cell’s genome and begins to function. Crop plants created for human or animal consumption by molecular biological techniques are referred as GMOs (genetically-modified organisms). Genetic engineering can create plants in the laboratory with desired trait(s), e.g. increased resistance to herbicides (Table 5.), weather extremes (droughts, cold) or enhanced nutritional value (protein/sugar content, etc.). This process is much faster than conventional breeding methods. During genetic modification, the intended gene is built-in the genome of the plant with the help of a vector that contains other genes, e.g. viral promoters, transcription terminator elements, genes of antibiotic resistance and reporter genes. 6.1. táblázat - Table 5.: Strategies for developing herbicide resistance Mechanism Effect transgene Building in the mutant gene Building in a detoxifying Gene expression (base pair change) gene of Changes in the amino acid sequence of the damaged protein, it inhibits the engagement of the herbicide’s active material Chemical modification of the Overexpression herbicide’s active material, damaged protein. which inhibits the engagement with the proper protein. The enzyme deactivates the active material of the herbicide. of Changes in mutant EPSP protein /enol- acetylase, nitrilase enzymes amplification the piruvyl-shikimate-3-phosphate/ deactivate the herbicide’s metabolism agent mutant ALS protein /acetolactate synthetase/ mutant AHAS protein /hydroxy acetate synthetase/ Result of glyphosate resistance glufosinate resistance transgenesis, the herbicide e.g.: Round-Up Ready systems e. g. Liberty ulink resistance 38 Created by XMLmind XSL-FO Converter. phosphinotricine resistance e. g.: Finale resistant wheat the 6. Genetically modified plants (GM) sulphonylurea resistance GM rape e. g.: SUMO maize hybrids bromoxinyl resistance imidazolinol resistance e. g.: cotton containing bxn gene e. g.: IMI maize hybrids 1. Types of genetically modified plants Biotic stress resistant plants • insect resistant • fungal infection resistant • bacterial infection resistant • viral infection resistant Abiotic stress resistant plants • herbicide resistant • heat stress resistant Plants with modified metabolism • amino acid and protein metabolism • carbohydrate metabolism • fatty acid metabolism The three generations of genetically-modified plants (also called ‘transgenic plants’) with their possible advantages are shown in Table 6 . 6.1. ábra - Table 6.: Generations and possible uses of GM plants 39 Created by XMLmind XSL-FO Converter. 6. Genetically modified plants (GM) Application of transgenic plants for environment protecting purposes: The transgenic plants can be used for promotion of phosphorus mobilisation. In plants phosphorus is found in the form of phytic acid (inositol hexaphosphate), which compound can not be digested by animals. With the use of manure phosphate content in the soil increases, which results in the eutrophization of natural aquatic environments. With the help of transgenic plants expressing higher phytase activity, phosphorus consumption in plant can be increased which is accompanied by the increased uptake of iron and zinc. The production of cadmium and copper tolerant plants • The binding and vacuolar transport of Cd and Cu generating toxic oxigen radicals are done by phytokelatins in the cells. • The tolerance can be enhanced by the amplification or the overexpression of the phytokelatin synthetase gene. • Heavy metals can be eliminated from the soil but they remain in the plant. • Phytokelatin synthetase synthesises peptide with a mechanism independent of the ribosome. In the peptide bond the γ-carboxyl group of Glu and the α-amino group of Cys take part. Mercury tolerant and mercury detoxifying transgenic plants • In the soil mercury is methylated by micro-organisms. • CH3-Hg+ is highly toxic and accumulates in the nutrition chain • Bacteria are capable of converting organic mercury into a less toxic ionic form, ionic mercury into the least toxic and volatile form by enzymatic reactions. • MerB (organic mercury liase): bacterial enzyme that brakes the C-Hg bond and therefore ionic Hg(II) is formed • MerA (ionic mercury reductase): bacetrial enzyme that converts mercury ions into volatile Hg • These built into Arabidopsis, canola, tobacco, poplar, etc. 40 Created by XMLmind XSL-FO Converter. 6. Genetically modified plants (GM) • With the simultaneous input of MerA and MerB organic Hg compounds can be disclosed from the nutrition chain. In 2020 an estimated 8 billion people will live on Earth, which will mean massive changes in the production, distribution and stability of food products. With no doubt, new methods in production are needed to feed the increased population of the planet. GM crops could significantly improve crop yields because more food can be grown on less land area. An environmental fact is that after the first some years of using herbicide tolerant GM soybean, seed rape, cotton, corn varieties and insect protected GM cotton, a dramatic reduction of pesticide use was observed (in 2000 total global reduction in pesticide use was 22.3 million kg of formulated product). Reduced use of pesticides can significantly decrease their effects on water quality through run-off and leaching of residues into surface and groundwater. Deployment of insect resistant Bacillus thuringiensis (Bt) varieties was estimated to have reduced the total world use of insecticides by 14%. Plants could be modified directly to be used for environment preservation purposes, such as (Table 7.): 6.2. ábra - Table 7.: Possible advantages of GM production for the preservation of the environment • Phytoremediation: removals of pollution from the environment with the help of plants, e.g. poplar trees have been genetically engineered to clean up heavy metal pollution from contaminated soil (Figure 30.). • Phytoextraction (or phytoaccumulation) uses plants to remove contaminants (heavy metals) from soils, sediments or water into harvestable plant biomass. • Phytostabilization focuses on long-term stabilization and containment of the pollutant. Mainly focuses on sequestering pollutants in soil near the roots but not in plant tissues. Pollutants become less bioavailable and livestock, wildlife, and human exposure is reduced. An example application of this sort is using a vegetative cap to stabilize and contain mine tailings. 6.3. ábra - Figure 30.: Phytoremediation strategies 41 Created by XMLmind XSL-FO Converter. 6. Genetically modified plants (GM) In contrast with the possible advantages of GM plants there are a lot of potential risks; among these we summarize some environmental/ecological problems (Table 8.): 6.4. ábra - Table 8.: Potential risks of GMO production 42 Created by XMLmind XSL-FO Converter. 6. Genetically modified plants (GM) • Escape of transgene: • during the harvest, seeds can be scattered, mixed with seeds of non-GM plants • in the case of vegetatively propagated plants, vegetative plant parts (tubers) can remain in the soil and new plants can develop from them • Canola in Europe can cross with weeds and make fertile hybrids • Development of new viruses: in the cells of GM plants viral RNA synthesized from the virus genes can recombine with the infecting viral RNAs • Development of weeds that can be killed: enhanced effect of them • Effect on non-target organisms: e.g. pollens of Bt species with high endotoxin content can escape into the environment, killing the useful insects Besides the problems mentioned above, risks on human health have to be considered, such as (Table 9.): • digestibility problems, • development of allergic reactions, • toxicity. 43 Created by XMLmind XSL-FO Converter. 6. Genetically modified plants (GM) 6.2. táblázat - Table 9.: Potential risks of GMO HUMAN/HEALTH PROBLEMS Toxicity, protein allergy JUDICIARY PROBLEMS Monopolistic (patents) POLITICAL PROBLEMS Genetic colonialism Plants with modified metabolism overexpress proteins, enzymes express and status Global patents Defenceless areas Plant with modified metabolism express/overexpress proteins, enzymes and their effects can not be predicted. Authorisation of the GMO’s Food and feed produced from genetically modified organisms can be allowed on the market if they have received authorisation. The authorisation process is carried out by the Europian Union, and the resulting decision applies to all EU Member States. The process for authorising a GMO is based on the EU regulation on genetically modified food and feed (1829/2003). The authorisation process of GMO’s is summerized in the Figure 31. 6.5. ábra - Figure 31.: Authorisation process of GMO’s ‘Vienna Declaration for a GMO-free Europe’ „ Transgenic agriculture will have an unacceptable impact on the survival of conventional and organic agriculture in Europe. Without clear prohibition of genetic pollution, the quality of our agriculture can not be guaranteed. We now face the challenge of protecting our natural and agricultural biodiversity.” „ Coexistence must not mean contamination. The objective of any legislation on coexistence must be to ensure guaranteed GMO-free agriculture and food production. This means: Who applies genetically modified organisms must strive for zero-contamination. Coexistence measures have to be tailored in such a way, that contaminations remain the absolute exception.” Labelling of genetically modified foods 44 Created by XMLmind XSL-FO Converter. 6. Genetically modified plants (GM) In the Europian Union, if a food consists or contains of genetically modified organisms or contains ingredients produced from GMOs, it has to be indicated on the label. The GM Food and Feed Regulation (EC) No. 1829/2003 lays down rules to cover all GM food and animal feed, regardless of the presence of any GM material in the final product. Products have to be labelled as GM if they are from a GM source. Products are produced with GM technology (for example: cheese produced with GM enzymes) or food (for example: milk, eggs) is from animals fed on GM animal feed do not have to be labelled. The EU regulations lays down that if the ingredient contains, consists of or is produced from GMO products and contains the GMO more than 0.9 % must be labelled (Figure 32.). 6.6. ábra - Figure 32.: Gene technological marking „Contains component produced with gene technology" „Produced with gene technology, but does not contain component produced with gene technology” Regulation of GMO’s European Community Directive 90/220/EEC (1990), amended for several times (2001/18/EC) • deliberate releases of GMO’s • environmental evaluation and step-by step approval for dissemination of GMO’s • case-by case assesment of the risks • framework for Member State to accomplish Regulation (EC) No. 1829/2003 and (EC) No. 1830/2003 of the European Parlament and of the Council: „For food or feed products -including those intended directly for processing- traces of GMO’s will continue to be exempt from the labelling obligation if they do not exceed the threshold of 0.9% and if they present is adventitious and technically unavoidable.” • consumer’s safety • compulsory labeling • transmition of information 2. GMOs in the U.S. Genetically-modified foods are prevalent on the U.S. markets; we speak about not whole vegetables or fruits but processed ones like vegetable oils or breakfast cereals that may contain genetically-modified ingredients in a very small portion. Soybean derivatives also can be present in foods. From 1996 to 2006 (the first ten years of commercially available genetically-modified plants), herbicide tolerance has consistently been the dominant trait followed by insect resistance. In 2006, GM plants in the U.S. were cultivated on 54.6 million hectares. The major biotech crops were soybean, maize (these two plants are the most widely grown ones, not only in the U.S. but all over the world), cotton, canola, squash, papaya, alfalfa. Plant varieties meeting the requirements of commercialization determined by the Food and Drug Administration 45 Created by XMLmind XSL-FO Converter. 6. Genetically modified plants (GM) (FDA) and the United States Department of Agriculture (USDA) can be found on ‘The List of Completed Consultations on Bioengineered Foods’. Genetically-modified varieties have been widely adopted by farmers in the U.S. They expect higher yields and lower pesticide costs. A number of surveys were conducted in the U.S. during recent years on public acceptance of foods containing GE (genetically-engineered) ingredients. In 2003-04, almost half (47%) of the population asked opposed the introduction of GE foods, while only 27% favored. 47% approved or leaned toward approval of the use of GE to make plant-based foods, 41% disapproved or leaned toward disapproval, and 12% were unsure. In 2005, 50% said likely to buy and 45% said not likely to buy GE products modified to taste better or fresher; 64% said likely to buy and 32% said not likely to buy GE produce modified to require fewer pesticide applications. Despite the expectancies, pesticide use has not decreased since the introduction of GMOs in the first eight years in the U.S. (Table 10.). 6.3. táblázat - Table 10.: Average pesticide pounds applied per acre planted to conventional, herbicide-tolerant (HT) and Bt transgenic varieties 1996 2000 2003 Herbicides 2.67 2.13 1.99 Insecticides 0.18 0.18 0.06 Herbicide-tolerant 1.87 1.83 2.32 Bt transgenic 0.16 0.05 0.02 1.20 0.99 0.87 0.84 1.10 1.34 Herbicides 1.93 1.86 1.42 Insecticides 0.56 0.41 0.35 Herbicide-tolerant 1.58 2.09 2.43 Bt transgenic 0.10 0.10 0.10 Conventional corn GE corn Conventional soybean Herbicides GE soybean Herbicide-tolerant Conventional cotton GE cotton Across the three crops, HT varieties increased herbicide use, the two Bt transgenic crops reduced insecticide use and all GE crops planted since 1996 have increased corn, soybean, and cotton pesticide use. 3. GMOs in Europe 46 Created by XMLmind XSL-FO Converter. 6. Genetically modified plants (GM) To date, the only type of GMO grown in the EU is Bt corn. Bt corn contains a gene from a bacterium (Bacillus thuringiensis) that produces a toxin (Bt-toxin) to defend it from the European corn borer (Ostrinia nubilalis). The insect pest is present primarily in southern and middle Europe, and is slowly making its way north. Genetically-modified crops are grown in six countries of the European Union and their cultivation areas are increasing. Data on the last two years are shown in Table 11. 6.4. táblázat - Table 11.: Cultivation areas (hectares) of Bt corn in European countries Country 2006 2007 Spain 53,700 75,150 France 5,000 21,200 Czech Republic 1,290 5,000 Portugal 1,250 4,199 Germany 950 2,685 Slovakia 30 900 In Spain, a significant amount of the corn production is genetically modified - an estimated 25% of the current production. Bt corn was first grown in 1998. In France, biotech corn has gained strong support among the farmers, who stand to gain more from the crop than any other EU country. Bt corn was first grown in the Czech Republic in 2005. Portugal also began producing Bt corn in 2005. Since the 2006 growing season, Bt corn cultivars have full approval in Germany and are now ready for commercial cultivation. All areas must be declared in a site register. Slovakia became the newest country in 2006 to plant biotech crops. In 2006, nearly 80% of Romania’s soybean production consisted of herbicide-tolerant varieties. Romanian Government decided to discontinue cultivation of GM soybean upon joining the European Union in January 2007. Most of the European countries are against GMO. On the 5th of April 2006, the ‘Vienna Declaration for a GMO-free Europe’ was pronounced by the platform organizing the ‘March for a GMO-free Europe’ - prepared by the many NGO’s (non-governmental organizations) like Global 2000, Greenpeace, and other environmental organizations -, saying: “Transgenic agriculture will have an unacceptable impact on the survival of conventional and organic agriculture in Europe. Without clear prohibition of genetic pollution, the quality of our agriculture cannot be guaranteed. We now face the challenge of protecting our natural and agricultural biodiversity.” (…) “Coexistence must not mean contamination. The objective of any legislation on coexistence must be to ensure guaranteed GMO-free agriculture and food production. This means: Who applies genetically modified organisms must strive for zero-contamination. Coexistence measures have to be tailored in such a way that contaminations remain the absolute exception.” Regulations are strict for the authorization of GMOs in Europe. The main instrument is European Community Directive 90/220/EEC - prepared in April 1990, amended for several times (e.g. 2001/18/EC). It regulates the deliberate releases of GMOs for research and development and the placing on the market of geneticallymodified products. It provides for an environmental evaluation and a step-by-step approval for the dissemination of GMOs. A case-by-case assessment of the risks to human health, animal health and the environment is carried out prior to a release and the placing of a GMO on the market. It also provides a framework that Member States 47 Created by XMLmind XSL-FO Converter. 6. Genetically modified plants (GM) must accomplish. A national authority in every Member State has to regulate the release of GMOs. Decisionmakers have to find balance between interests and opinions of consumers, NGOs, producers, retailers and farmers who influence the authorization process. The consumer group has the most influence on the authorization procedure. Labeling and traceability is among the very strict regulations in Europe. Regulations (EC) No. 1829/2003 and (EC) No. 1830/2003 of the European Parliament and of the Council concerning the traceability and labeling of genetically modified organisms and the traceability of food and feed products produced from genetically modified organisms require the following: • Consumers’ safety has to be guaranteed as a result of the traceability of products consisting of or containing GMOs. • All products are subject to compulsory labeling. • Operators should transmit the following information in writing: 1. an indication that the products consist of or contain GMOs 2. the unique alphanumerical identifiers assigned to the GMOs contained in the products • Operators who place on the market a pre-packaged product consisting of or containing GMOs must, at all stages of the production and distribution chain, ensure that the words “This product contains genetically modified organisms” or “Product produced from GM (name of organism)” appear on a label affixed to or transmitted with the product. • When placing a product on the market, the industrial operator must transmit the following information in writing to the operator receiving the product: 1. an indication of each food ingredient produced from GMOs, 2. an indication of each raw material or additive for feeding stuffs produced from GMOs, 3. if there is no list of ingredients, the product must nevertheless bear an indication that it is produced from GMOs. • For food or feed products, including those intended directly for processing, traces of GMOs will continue to be exempt from the labeling obligation if they do not exceed the threshold of 0.9% and if their presence is adventitious and technically unavoidable (European Union website). European consumer opinion is expressed and enhanced through NGO’s activities. It is dominated by a negative attitude towards GM products. According to the results of several surveys the issue of genetic engineering ranks high up in the list of potential risks caused by food. Considering the above mentioned, the future of this field in Europe cannot be predicted. The resistance is very strong, so it seems that GM food will not spread the markets in the next years. Figure 33 shows a comparison of people’s opinion about different applications of biotechnology in Europe and the USA. 6.7. ábra - Figure 33.: Approaches of the public to fields of biotechnology in Europe and the U.S. 1. remedies, 2. genetic testing, 3. crops, 4. food, 5. organ transplant 48 Created by XMLmind XSL-FO Converter. 6. Genetically modified plants (GM) 4. GMOs in Hungary Hungary joined the European Union on 1st May 2004. The country has moratorium in place for the commercial cultivation of GM crops that can be applied with reference to new scientific evidence for security risks. The Commission’s motion to lift the Hungarian moratorium was outvoted by more than half of the Member States in September 2006. It is not easy to see the clear picture. On one side, there are opinions that most of Hungarian consumers (7080%) do not want GMOs. These figures come from green movements and NGO’s. Multinational biotechnological companies in the country state the opposite that almost three-fourth of farmers want to grow GMOs. As it is seen, two sides are present opposite each other about the question. The ‘pros’ - academics interested in conducting research for multinationals and the big farmers’ group both claim the right to choose. This kind of development in biotechnological research could place Hungarian agriculture on the top rank of the region in view of the application of the new technology. Other arguments are that energetic sector could profit from the new technology, it is environmentally friendly, high added value and many jobs could be created in regions where unemployment is a real serious problem. Their communication is a bit one-sided and neglects the resistance of consumers saying that consumers have no business defining what the product comes from. The ‘contras’ are mostly politicians - there is a 5-party consensus in the Parliament -, consumers, environmental organizations, organic farmers, leading scientists from the fields of nutritional and ecological sciences. Farmers say that GMO-free status of the country would mean better prices on markets, while coexistence would result in the loss of markets; and since Hungary is an agricultural country, these losses would adversely affect its whole economy. Scientists emphasize the unnecessary and unpredictable risks of the technology. The most debates were on the buffer zones - the separation of conventional and biotech crops. Finally, the size became 400 meters. This kind of strict restriction is exceptional in Europe. Not only crops but infrastructure for the two technologies would be needed to avoid contamination from treatments after harvests and the economical sources for this can be questionable. The opinion of consumers and professionals about gene technology is mostly negative and figures are a bit different but similar. Professionals: 37.61% ‘rather negative than positive’, 17.95% ‘predominantly negative’. Consumers: 35.04% ‘rather negative than positive’, 13.25% ‘predominantly negative’. More than half (51%) of the consumers would refuse GM food even if it was cheaper, more tasty, had better appearance and longer shelf-life than the traditional one. Labeling also seems to be very important for Hungarian consumers as mainly three-fourth of them (73%) thinks it compulsory to indicate GM content on the packages of foodstuffs. Even if Hungarian consumers predominantly refuse GM products, this proportion is still much smaller than in Western Europe. The approach to genetic modification in plant breeding and the coexistence of traditional and biotech crops is not uniform all over the world. While in the U.S. the ratio of the GM-production reached 30-40%, from which it made no longer sense to differentiate between GM and conventional, in Europe there is great resistance to the new technology. Standpoints are also diverse about the environmental advantages, mainly knowing the facts that e.g. the tendency of herbicide/insecticide consumption in some places was the opposite than expected (increased) or the potential health risks possibly caused by GM foodstuffs. In Hungary the possible future application of GM plants has more disadvantages just right now instead of providing benefits to farmers from environmental, ecological and economical point of view. It seems that consensus is still far away, the debates will go on; the counterparts will abide by their opinions for a long time. 49 Created by XMLmind XSL-FO Converter. 7. fejezet - 7. Basis of plant breeding 1. Plant genomics Plant genomics allows scientists to understand the genetic architecture of plant genomes, to analyze thousand of genes in parallel, and also to isolate the genes responsible for mutations. The whole plant genomes can now be sequenced. The study of the way genetic information and genes are organized within the genome, the methods of analyzing and collecting this information and how this organization determines their biological functionality is referred to as genomics. Plant genomes are described in terms of gene content, genome size, extent of repetitive sequences and polyploidy/duplication events. Although plants also have mitochondrial and chloroplast genomes, their nuclear genome is the largest and most complex. There is variation in nuclear genome size of plants (Table 12.). 7.1. táblázat - Table 12.: Nuclear genome size of plants Common name Scientific name Nuclear genom size (in Megabases) Wheat Triticum aestivum 15,966 Corn Zea mays 2,292 Garden pea Pisum sativum 3,947 Tomato Lycopersicum esculentum 907 Sugar beet Beta vulgaris 758 Plant genomes contain various repetitive sequences. Corn genome contains over 70% repetitive sequences and 5% protein encoding regions. Many economically important plant species, such as corn, wheat, potato, and oat are either ancient or more recent polyploids, comprising more than one, and in cases such as wheat, three different homologous genomes within a single species. Bread wheat is a hexaploid, with three complete genomes termed A, B and D in the nucleus of each cell. Each of these genomes is almost twice of the human genome and consists of around 5,500 million letters. The genetic code of wheat contains estimated 80,000 genes. The wheat genome is five times larger than the human genome. Duplicated segments also account for a significant fraction of the rice genome. About 60% of the Arabidopsis genome is present in 24 duplicated segments, each more than 100 kilobases (kb) in size. The species used as model organisms for mono- and dicotyledonous plants are rice (Oryza sativa) and Arabidopsis (Arabidopsis thaliana) respectively. Arabidopsis thaliana as a model plant Arabidopsis is a member of the mustard (Brassicaceae) family, which includes cultivated species such as cabbage and radish (Figure 34.). 7.1. ábra - Figure 34.: Arabidopsis thaliana 50 Created by XMLmind XSL-FO Converter. 7. Basis of plant breeding Arabidopsis is not of major agronomic significance. It has small genome (114.5 Mb/125 Mb total) which has been sequenced in the year 2000 (SequenceViewer, AGI). Arabidopsis thaliana has only five chromosomes and it has a rapid life cycle (about 6 weeks from germination to mature seed) (Figure 35. and Table 13.). Arabidopsis can be genetically transformed on a large scale with Agrobacterium tumefaciens and biolistics. 7.2. ábra - Figure 35.: Genome size, chromosome and genes number of Arabidopsis thaliana 7.3. ábra - Table 13.: Gene complement of Arabidopsis Oryza sativa as a model plant 7.4. ábra - Figure 36.: Oryza sativa 51 Created by XMLmind XSL-FO Converter. 7. Basis of plant breeding Oryza sativa (rice) is the staple food for 2.5 billion people, and after maize, the yield with the second highest worldwide production. It has 12 chromosomes, a genome size of 420 Mb (Table 14.). It can be transformed through biolistics method and Agrobacterium tumefaciens. 7.2. táblázat - Table 14.: Genome size, chromosome and genes number of Oryza sativa Organism: Oryza sativa Genome size: 420 Mb Number of chromosomes: 12 Number of genes predicted: 32-50,000 Organization initiative: Beijing Genomics Institute, Zhejiang University and the Chinese Academy of Sciences Year of completion: 2002 2. Fundamentals of plant breeding Plant breeding is the art and science of improving the genetic pattern of plants, and changing the genetics of plants in order to produce desired characteristics. The goal of plant breeding is to combine desirable traits from different varieties of plants to produce plants of superior quality. This approach to improving crop production has been very successful over the years. At the beginning of plant breeding farmers started to select plants for specific features such as larger seeds, faster growth, or sweeter fruits. Classical plant breeding dates from Gregor Mendel’s discoveries. 52 Created by XMLmind XSL-FO Converter. 7. Basis of plant breeding Advances in scientific discovery and laboratory techniques during the last half of the 20th century led to the ability to manipulate the deoxyribonucleic acid (DNA) of organisms, which accelerated the process of plant improvement through the use of biotechnology. The ability to move genes into plants from other organisms, thereby producing new proteins in the plant, has resulted in significant achievements in plant biotechnology, that were not possible using traditional breeding practices. Basis material and sources of plant breeding: • Wild species and varieties • Natural population, native species • Artificial population • Breeding species • Mutant lines • Polyploidy • Transgenic hybrids • Doublehaploid (DH) lines Traits that breeders have tried to incorporate into crop plants in the last 100 years include: • increased quality and yield of the crop, • increased tolerance of environmental pressures (salinity, extreme temperature, drought), • resistance to viruses, fungi and bacteria, • increased tolerance to insect pests, • increased tolerance of herbicides. Major activities of plant breeding are the following: • creation variation • selection • evaluation • release • multiplication • distribution of the new variety Application in plant breeding • opportunity for the reduction of ploidity: gametoclonal variability - production of plant without fertilisation • in haploids and doubled haploids recessive features can be manifested and selected phenotypically • inbreeding can reduce the production time • addition and substitution hybrids (e. g. Triticale) • fusion of monohaploids 53 Created by XMLmind XSL-FO Converter. 7. Basis of plant breeding • production of homozygous transgenes • production of „super males” (e. g. asparagus) • genetically pure lines for marker selection 54 Created by XMLmind XSL-FO Converter. 8. fejezet - 8. Methods of plant breeding 1. Selective breeding It is the most ancient and basic procedure in plant breeding. The breeding of selected plants and animals to produce offspring with desired traits. The offspring with the desired traits are then used as breeding stock for the next generation and so on, until offspring that express the desired traits are obtained. Mass selection It is the oldest, simplest and easiest method which is based on the phenotype selection. Plants are selected according to their phenotype and performance from a mixed population. The seed from selected plants are bulked and used to grow the next generation (Figure 37.). 8.1. ábra - Figure 37.: Mass selection Mass selection method is effective for characters with high heritability. It can be used in self pollinated and cross pollinated plants. In case of self pollinated plants, mass selection varieties are a mixture of several pure lines. It means homozygous and heterogeneous population. In case of cross pollinated plants, mass selection 55 Created by XMLmind XSL-FO Converter. 8. Methods of plant breeding varieties are mixture of several homozygotes and heterozygous. It means heterozygous and heterogeneous population. There are two types of this selection, negative and positive mass selection. Positive mass selection means when we select the desirable crops from a mixed population, then we mix their seeds together to raise next generation. In this method the base material can be land races or old varieties. Negative mass selection means when we remove undesirable crops from a mixed population and the rest are allowed to grow further. In case of self pollinated varieties the negative mass selection method helps to maintain the genetic purity. The mass selection can be applied to improve local varieties and purify the existing pureline varieties. Advantages of mass selection • simple, rapid and cheep procedure • plants procedured by mass selection have wide adaptability, wide genetic base • use for the improvement of land races Pedigree selection in plant breeding The pedigree method was developed by Vilmorin (French plant breeder) in the second part of last century. In this manner selected plant seeds grown in their own row the next generation (ear to row, plant to row). The first step in pedigree method is hybridization when we cross the selected parent plants. F1 seeds are harvested in bulk and forwarded to raise F2 generation. In F2 generation the main process is the selection. The selected plants are harvested separately. Desirable plants are selected in F4 generation. Individual plant progenies planted in multi row (3 or more) plots in F5 and F6 generations. Superior plants are selected by comparison in F5 generation and visual evaluation selections are in F6 generation, in this way we can eliminate progenies showing segregation. Quality test and preliminary yield trials with minimum 3 replications are in F7 generation. Multi-location yield trials with replications, quality and disease resistance test are in F8 to F12 generations. Seed increase for distribution are in F9 to F10 generations (Figure 38.). 8.2. ábra - Figure 38.: Pedigree method 56 Created by XMLmind XSL-FO Converter. 8. Methods of plant breeding Cross breeding In the beginning plant breeder cross (interbreed) the closely or distantly related individuals to produce new plant lines or varieties with desirable characteristics. Plants also are crossed with themselves to produce inbred varieties. Test cross a cross between an organism whose genotype for a certain trait is unknown and an organism that is homozygous recessive for that trait so the unknown genotype can be determined from that of the offspring Single cross is when a cultivar or a line is crossed with another line or cultivar (AxB). In case of every cross we try to get lines which are better than the existing cultivars. Double cross is the mating of two F1 generations. Top cross is the cross of a single hybrid (AxB) to a variety or inbred line. Backcross breeding Backcross breeding is a crossing of a hybrid with one of its parents or an individual genetically similar to its parent, to introduce a desirable gene. This method is modified the existing varieties and it is not created new varieties. The recurrent parent is an elite line that can be made better by crossing with the donor parent. The donor parent contains one particularly valued gene e.g. for disease resistance. The progenies backcrossed to the recurrent parent, and this process is repeated until about the seventh or eighth backcross generation. Backcrossing is more efficient in case of the self-pollinating crops but it can be used the cross pollinated crops as well. Diallel cross breeding All parents are crossed to make hybrids in all possible combinations in the full diallel cross method (Table 15.). It allows for testing for maternal and paternal effects. It can be used to estimate general or specific combining 57 Created by XMLmind XSL-FO Converter. 8. Methods of plant breeding ability, interactions with testing environments and years, or estimates of additive, dominant, and epistatic genetic effects and genetic correlations (Figure 39., 40.). 8.3. ábra - Table 15.: Full diallel cross method 8.4. ábra - Figure 39.: Ear crops of lines of ‘A’ complete diallel system 8.5. ábra - Figure 40.: Parental lines of ‘A’ complete diallel system at the time of flowering 58 Created by XMLmind XSL-FO Converter. 8. Methods of plant breeding 2. Polyploidy The base number of nuclear chromosomes is called the haploid number (n). Haploid plants have only one set of chromosomes.The most species are diploid, having double the haploid number of chromosomes (2n) in each somatic cell. Aneuploid and polyploid organisms have abnormal numbers of whole chromosomes. In aneuploidy, one or more whole chromosomes lost or gained from the diploid state. Trisomy (2n+1) and monosomy (2n-1) are two forms of aneuploidy. Polyploid plants contain more than two haploid (n) sets of chromosomes, that is, their chromosome number is some multiple of n greater than the 2n content of diploid cells (Table 16.). Triploid (3n), tetraploid (4n), hexaploid (6n) oktoploid (8n) and etc. cells are polyploid. 8.1. táblázat - Table 16.: Polyploid plants Plants Ploidy level Chromosome number Haploid number (n) wheat 6n 42 7 59 Created by XMLmind XSL-FO Converter. 8. Methods of plant breeding oat 6n 42 7 potato 4n 48 12 tobacco 4n 48 12 sugar cane 8n 80 10 Polyploids are very common in plants. Polyploid plants often have larger flowers or fruit and more productive than diploid plants, and well-adapted to their environments, furthermore plant breeders often produce such plants by crossing species. Polyploidy can be induced in the plant breeding by treating dividing cells with colchicine. Colchicine is an alkaloid derivative from the autumn crocus (Crocus veneris). The polyploid species are of two types: • Autopolyploid species which are contain more than two copies of a single genome. • Allopolyploid species which are contain two or more genomes and each genome is ordinarily present in two copies. Allopolyploidy is more common in nature than autopolyploidy, about 80% of plants are allopolyploids. The common bread wheat (Triticum aestivum) is an allohexaploid (AABBDD). Triticum aestivum contains three distinct sets of chromosomes derived from three different diploid species. Durum wheat (Triticum turgidum durum) (AABB) is used to make pasta, is tetraploid (it has four sets of chromosomes). 3. Hybridization/heterosis It is a very important method in plant breeding. The production of offspring, known as hybrids, from genetically dissimilar parents. The object of hybridization is to combine desirable genes found in two or more different varieties to produce pure-breeding offspring superior in many respects to the parental types. In generally, the production of hybrid is known as hybridization. Types of hybrid • single cross SC (AxB) F1 • three way cross TC (AxB) F1 x C • double cross DC (AxB) F1 x (CxD) F1 • sister line cross SLC = A1xA2 • modified single cross MSC (A1xA2) x B • reciprocal single cross SC(R) BxA Heterosis is the superiority of an F1 hybrid over its parents. The high crop productivity that results from heterosis has been exploited through the development of hybrid varieties in many crop species, most notably maize. Heterosis as it applies to crop breeding was first recognized by Shull. The genetic basis of heterosis, however, has not been completely explained. The two main hypotheses that have been proposed as the genetic basis of heterosis are the dominance hypothesis and the overdominance hypothesis. Heterosis has been exploited extensively in crop production and has been a powerful force in the evolution of plants. Heterosis is the difference between the hybrid performance and the mean value of the inbred parents, or alternatively, the value of the best parent, for a trait of interest normally expressed as a percentage. The heterosis breeding in maize caused gene erosion by using uniform inbred lines. In order to avoid it can be estabilished a maize in vivo gene bank, this contains inbred lines with graeter genetic potential, better resistance and more ecological adaption. 60 Created by XMLmind XSL-FO Converter. 8. Methods of plant breeding A few studies have documented the presence of heterosis for the physical, compositional, and wet-milling characteristics of inbred lines and their hybrids in maize. Zehr studied 15 adapted inbred lines and 20 related hybrids and reported significant divergence of hybrids from mid-parent values for wet-milling fraction values. The expression of heterosis by crossing 10 exotic populations with the public inbred lines B73 and Mo17 and reported higher levels of protein and reduced starch contents of the hybrids relative to the inbred lines. Fifteen phenotypically diverse white maize inbred lines were crossed and grain yield recorded heterosis up to 78.83% (mean value of 41%). The mean heterosis was 21% for TKW. Harvest index and 100-seed weight had the highest correlation with grain yield in terms of heterotic expression 0.518 and 0.495, respectively. Positive mid-parent and high-parent heterosis for starch content and negative values for protein content were expressed by the grain yield obtained via self-pollination from specific hybrids between lines with partial exotic background and commercial inbred lines. There was a positive correlation between starch content and starch yield and starch recovery and a strong negative correlation between protein content and starch yield and starch recovery. This indicated that genotypes with high starch and low protein contents produce hybrids with better milling ability. Component analysis was used to quantify the effect of heterosis for kernel traits. The average heterosis is the most important heterosis component in the studied material. And the average heterosis is depending on dominant genes and gene frequencies of the population. The average heterosis had a significant effect on yield in dent-type maize. Reasearhers experienced in a 7x7 diallel analysis that the heterosis of starch content was the highest. The mid-parent heterosis of protein content was significant, negatively correlated with that of starch content. The heterosis of all characters had a wide range of variability among crosses. The correlation between parents and F1 for protein and oil content was significant or highly significant. In the experiments with 28 hybrids from 8 inbred lines, 4 hybrids showed significant positive heterobeltiosis and heterosis for grain yield per plant and other yield component traits. The aim of our breeding program was to determinate the heterosis and heterobeltiosis in the case of our maize lines and hybrids (Table 17.). 8.6. ábra - Table 17.: Changes of heterosis values in our maize hybrids (Debrecen) 61 Created by XMLmind XSL-FO Converter. 8. Methods of plant breeding Maize genotypes with increased starch content could serve as breeding materials of high promise to meet the requirements of bioethanol production and as the raw material supply of food and feed. The heterosis effect study The levels of heterosis can be determinated. There are two heterosis types: average heterosis and heterobeltiosis with their absolute and percentage values. When heterosis is estimated from the average of two parents it is referred as average or relative heterosis. When heterosis is estimated over better parent it is called as heterobeltiosis. Heterosis and heterobeltiosis can be calculated with the following two formulas. Heterosis (average %) = [F1-(P1+P2)/2]*100 / ((P1+P2)/2) F1=average of the studied trait of the hybrid P1 = average of the studied trait of one of the parents P2 = average of the studied trait of the other parent Heterobeltiosis (average %) = [F1-HP]*100 / HP 62 Created by XMLmind XSL-FO Converter. 8. Methods of plant breeding HP = average of the highest parental performance 4. Mutation breeding Mutation breeding is a great help for widen the genotypes suitable in plant selection. Application of induced mutants increases the gene reserves which have a great importance because of the reduction in variation of populations. With the help of mutation we can increase the variability and select lines favourable for plant production. We can produce new hybrid combinations to satisfy the production requirements. Figure 41 shows the scheme of the plant mutation breeding. 8.7. ábra - Figure 41.: Scheme of mutation breeding We can complete diversification of the basic breeding material with neutron radiation. Use of neutron sources in maize breeding programs in a larger scale is well based because of its great genetic affinity. After the seed treatment with fast neutron source we can self-pollinate the segregated population, create genetically homozygote lines and select the stable inbred lines with favourable agronomic characters. In order to decrease the genetic vulnerability, it is very important to widen the genetic base for maize breeding by the application of various methods. After more than twenty-five years of research, it has been proved that such lines can be produced by mutation. In Debrecen, mutation breeding was initiated in 1958 by Károly Pásztor. Within the programme started in 1979–80, F1 maize hybrid seeds were treated by radiation of Co60 isotope. Later, in 1985 and in 1991, the trial was expanded and the seeds were treated by radiation of fast neutron at the Atomic Research Institute of the Hungarian Academy of Sciences in Debrecen. The cyclotron can be successfully applied in widening the genetic variability (Figure 42.) 8.8. ábra - Figure 42.: The cyclotron (Institute for Nuclear Research, Debrecen) 63 Created by XMLmind XSL-FO Converter. 8. Methods of plant breeding The selected mutant lines – developed in this way – have been self – pollinated for several years. In 1995, we established a maize gene bank with 1,500 lines, which are registered by the IBPGR. We can produce a number of inbred lines with wide genetic variability using mutation breeding: with application of diallele analysis, the lines can be selected with the best general and specific combining ability. Using these lines we can create F1 hybrids which have the most valuable economic features. In order to complete these tasks we have to examine diallele experiments with many replications in a number of vegetation periods and investigate many factors. After summarising results we can get conclusions to further selections. Only those lines and their hybrids can form the base of further breeding programs which give prominent results and fulfill the requirements. It is an important task to increase the productivity and improve the economical characters of the crops. It is important to take into account not only the yield and its elements, but their physiological processes. Heterosis breeding in maize has caused gene erosion by using uniform inbred lines. In order to avoid the genetic diversity in vivo maize gene bank has been established, which contains inbred lines with greater genetic potential, better resistance and better ecological adaptation. After more than twenty years of research it has been proved that such lines can be produced not only by polyploidization, or though intergeneric and interspecific crosses, but also by mutation. There is large genetic variation in the germplasm utilised, the exploitation of which is only possible using suitable methods of selection and evaluation. Radiation generated conspicuous changes in the plant characters described by UPOV in comparison with the basic material. The most pronounced aberrations were observed for the expression of anthocyanin coloration in various plant organs. Genetic improvements have been estimated to account for about 50 % of the total realised gains in yield per unit area of major crops over the past fifty years. Rates of gain attributable to genetic improvement have generally been linear and show no sign of slackening. The role of breeding methodology to improve cereals for grain yield, disease resistance and other agronomic traits is significant. Conventional breeding methods however are laborious and time and space consuming. To expand the scope of our mutation breeding at the Department of Genetics and Breeding the grain sorghum hybrid Zádor and the restorer B line Rib (Karcag Research Institute) were subjected to fast neutron treatment at the Institute of Nuclear Research of the Hungarian Academy of Sciences (Debrecen). The seeds of the hybrid base material and their lines were irradiated in the cyclotron of the Institute of Nuclear Research with doses of 5, 7.5, 10.0 and 12.5 Gy (Gray). The biological effect of the radiation is characterized with the absorbed dose marked with the symbol D. Its unit is the Gray (symbol: Gy), defined as the absorption of 1 joule of radiation by 1 kilogram of matter. 1 Gy = 1 J per kg. The changes in the M0, M1 and M2 generations were monitored biweekly and the following traits were studied: shooting data, plant height (cm), productive tillering (pieces), truss length (cm), collaterals of truss (pieces) and thousand kernel weight (Table 18.). The obtained data were used for the calculation of the following values: leaf area index (LAI, m2m-2) was determined with Montgomery’s 64 Created by XMLmind XSL-FO Converter. 8. Methods of plant breeding formula: Σ leaf length (cm) multiplied with leaf width (cm) and with 0.75. The truss density was determined with the formula of trashing percentage. Calli were induced from the leaf basis of the albino plantlets on media with the concentrations of 4.5, 12.5 and 22.5 micromol, 2.4 D and the changes were compared with those of the controls. 8.2. táblázat - Table 18.: Some characterization of irradiated Sorghum genotypes Trait Plant height Harvest(cm) index (average) Panicle: length Panicle: 1000-kernel of branches number of weight (gr) (cm) branches (db) Grain (kg/m-2) Rib (control) 105-117 0,4886 17-22 28-37 32,33 0,408 Rib 5 Gy 99-129 0,4620 15-33 13-32 27,65 0,422 Rib 7,5 Gy 70-124 0,6106 14-26 9-27 20,91 0,644 Rib 1n0 Gy 88-125 0,5264 12-25 14-32 30,21 0,4648 Rib 12,5 Gy 65-105 0,2338 14-23 14-36 25,64 0,183 Zádor (control) 100-120 0,4532 21-27 23-35 32,82 0,606 Zádor 5 Gy 80-120 0,4625 13-26 13-32 27,97 0,407 Zádor 7,5 Gy 90-118 0,5807 19-27 12-26 31,48 0,521 Zádor 10 Gy 68-106 0,4234 12-25 11-33 31,99 0,471 Zádor 12,5 Gy 48-119 0,2265 16-30 17-32 32,98 0,225 We experienced significant changes in the plants of Sorghum bicolor L. (Figure 43.). The ratio of germinated planlets reduced to 50 % in the 12 Gy treatment of Zador variety in comparison to the control. Double panicles developed in these plants (Figure 44.). 8.9. ábra - Figure 43.: The effect of the mutation in Sorghum panicle 65 Created by XMLmind XSL-FO Converter. 8. Methods of plant breeding 8.10. ábra - Figure 44.: Double panicle in the case of ‘12 Gy Zádor’ The shortest plant height was 48 cm. In the Rib population irradiated with 12,5 Gy the Harvest-index varied from 0,2265 to 0,2368 because of sterility. The period from planting date to panicle appearance was 7-8 days longer than the control. The highest Harvest-index was measured in the Rib and Zador populations both treated with 10 Gy fast neutron. It varied from 0,4234 to 0,5807. The leaf area index (LAI) was 3,87 m2/m2 due to the intensive productive tillering. In the case of Zador and Rib originated from populations irradiated with 7,5 Gy fast neutron th thousand kernel mass (TKM) varied from 22 g to 38 g. The kernel yield measured started at 0,5204 kg/m 2 and ended at 0,6443 kg/m2. There were only light changes in the 5 Gy treatments compard to the control. 66 Created by XMLmind XSL-FO Converter. 9. fejezet - 9. New alternatives in plant breeding 1. The genotypes, fertilization and irrigation from environmental point of view Among the agrotechnical elements of cereals fertilization and irrigation are the most important and efficient means of increasing the yields. The influence of these elements does not separately effect on yield but there are several interactions among them and other ecological, agrotechnical and biological factors. Strong correlation exists between the yields and the quantity of fertilizers used. In spite of the varying environmental conditions the specific nutrient demand of winter wheat is not too variable. In the references connected to fertilization a wide range of data can be found regarding the specific nutrient demand of winter wheat. In most of the cases the optimal species-specific dose of fertilizers varies, within the range of 150-300 kg/ha of NPK active substance. In studies concerning the nutrient-supply of winter wheat, the problems of variety-specific fertilization have come into the focus of interest and have published experimental result in that field. In addition to fertilization, irrigation is another important factor increasing the yields under the ecological conditions in Hungary. According to experimental results the extra crops gained by irrigation were strongly related to the weather of the actual year, to the applied water-norm, to the timing of irrigation and to the variety used. Regarding the special interaction of fertilization and irrigation in winter wheat some authors found the demand of fertilizers to increase while others found it to decrease as a result of irrigation. In recent years the variety assortment of winter wheat has enlarged in Hungary which caused a big pressure of practical-farming on the scientific research to work out the variety-specific technology of winter wheat. The most important elements of variety-specific technology of winter wheat are fertilization and irrigation. The demand of N-fertilizer of winter wheat varieties varies 60-180 kg/ha. Three different N-requesting groups of winter wheat varieties (intensive, semi-intensive and extensive variety groups) are specified (Figure 45.). 9.1. ábra - Figure 45.: Types of winter wheats according to their fertilizer responses 67 Created by XMLmind XSL-FO Converter. 9. New alternatives in plant breeding Big differences can be found in water demand and irrigation response of plants. Winter wheat varieties with excellent irrigation response have to be chosen into irrigation farming. The internationally outstanding increase of yields in the Hungarian plant growing until the middle 1980s was based on the improvement of the biological (genetic) properties. Of the agrotechnological factors, though other elements were also of importance, the decisive role could be assigned to the nutrient supply; an especially strict, consequent connection could be established between the mostly extensive, recently also intensive development of fertilization and the yields of corns. Due to the intensive fertilization involving high doses the degree of phosphorus and potassium supply in our soils has increased markedly, depending on the type of soil, fertilizer doses, growing and ecological conditions. The quantitative determination of these elements can be considered from the point of view of fertilization extension. A good supply of phosphorus and potassium in most of our soils gives us favourable conditions for the determination of these elements and for rational farming. For most field plants (especially for winter wheat) the most reactive element that determines the yield is nitrogen, depending on condition of harmonic nutrient supply. However, the determination of dose, prediction of transformation during the vegetation period, elucidation of the degree of uptake are the problems most seriously occurring in the case of nitrogen of all elements regularly replaced. Especially it applies to winter 68 Created by XMLmind XSL-FO Converter. 9. New alternatives in plant breeding wheat where the N-optimum range is relatively narrow. Nitrogen fertilizer in quantities less that the optimum results in a decrease of yield, while N-doses higher than the potentially usable quantity leads to a significant decrease in the economic effectiveness of fertilization and primarily in soils of favourable N-supply to the increase of the damage in the environment. Loss of N-fertilizer in soil varies between 30-70% depending on rainfall and irrigation physical-chemical characteristics of soil quantity and timing of N-fertilizer. There are differences of nitrogen demand of winter wheat varieties manifested in unfavourable ecological and agrotechnical conditions. The environmental aspects of N fertilization are getting more and more important. Unfortunately the nitrogen doses over the optimum point, bad application can cause serious environmental pollution in the soil and ground water. It means we have to focus our attention on nitrogen fertilizer doses and application. 2. Regeneration of maize using tissue culture and inhibition of pollen grain germination Hungarian maize production utilizes many foreign hybrids, which are derived from only a few inbred lines. Because of this genetic vulnerability production can be in a serious danger and quick gene erosion is a possibility. During the past ten years maize production level has remained unchanged and the opportunity of further development would be to develop new inbreds with desirable agronomic attributes. The regeneration of plants from tissue cultures of corn (Zea mays L.) was first reported by utilizing immature embryos as the tissue source. Regeneration of monocotyledonous plants (e.g. maize) via somatic embryogenesis from primary callus is poorly understood and progress in this area has been very slow. Both types of regeneration arise from hard, white or yellow callus that appears distinctly different from the granular, gray-yellow and translucent callus incapable of plant regeneration. Consequently, visual selection of plant regeneration-competent calli has become the method of choice for detecting and maintaining plant regeneration capability in Zea mays L. tissue cultures. Plant regeneration from maize tissue culture has mainly been obtained from callus derived from immature embryo. Successful plant regeneration has also been reported from calli initiated from anthers, glumes, immature tassels, leaf bases, mesocotyls, seedling segments and mature embryos. Immature embryo-derived callus is more embryogenic and efficient for plant regeneration than calli from other explant tissues. All the plant tissues used to date must be obtained from living plants which require time and space to grow. In maize the ability of leaf cell to express morphogenic capability decreases as the leaf matures. A method for the induction of embryogenic and organogenic calli from maize mature embryos could be satisfy the time and space requirements. The possible genetic mechanisms for controlling plant regeneration were studied for corn. It was concluded that regeneration from immature embryo-derived calli was controlled by a few nuclear genes or a few gene clusters. A large proportion of genotypic variation for formation of type 1 and type 2 callus in maize was due to additive gene effects while heterosis positively increased culture response. A significant maternal effect was also noted for B 73, but not for Lancester type lines. Callus growth from mature embryos on a modified LS medium was also controlled mainly by additive gene effects (general combining ability). The additive gene effects to be more important than the dominant gene effects for the percent of somatic embryogenesis and percent of number of plants regenerated per embryo when generation means were analyzed. Cytoplasmic, maternal and/or paternal effects were significant for frequency of somatic embryos as well as frequency or number of plants regenerated per embryo. Analysis of genetic variances suggested that at least one gene or block of genes controlled the expression of the frequency of somatic embryogenesis. Regeneration of maize from cell and tissue cultures has been limited to few specific genotypes and media combinations. Media improvements boost the general mean of the entries tested but genotypic differences remained. Studies on the genotype component can be used to predict probabilities for a desired response level and to describe in more detail the nature of the tissue culture response. 69 Created by XMLmind XSL-FO Converter. 9. New alternatives in plant breeding Mycotoxins, in general, reportedly contaminate about one-quarter of the world's yearly food and feed crops. Effects of mycotoxin consumption on animal health range from decreased growth rates and reproductive efficiency to mortality. In addition, there is an increasing concern about the effects of aflatoxin on human health, as aflatoxins have been ulinked to liver cancer. The occurrence of aflatoxin in food is viewed as a potential threat to the food supply. It has been deemed necessary to develop efficient methods that will prevent aflatoxin contamination in crops. Host-plant resistance is the most logical and useful method of control. Since the early 1970s, corn breeders and pathologists have made a concentrated effort to eliminate or reduce aflatoxin contamination in corn. To develop proper breeding strategies for incorporating resistance to a disease into germplasm, a breeder needs to (i) identify and/or develop sources of resistance, and (ii) determine the genetic nature of resistance. Identification of aflatoxin-resistant genotypes has been difficult due to the erratic nature of infection by Aspergillus. Natural infection has not been reliable enough to screen for resistance. Attempts have been made to identify effective artificial inoculation techniques. An efficient inoculation technique is required for improving populations for resistance Aspergillus group fungi or aflatoxin via recurrent selection procedures. Success from recurrent selection for improving resistance to the fungus or aflatoxin has not been reported. One of the reasons might be the nonavailability of a suitable inoculation technique for that purpose. In addition, thus far, screening has only been conducted at the sporophytic (plant or seed) level. So far, only a few studies on inheritance of resistance have been conducted. Progress in elucidating genetic mechanisms and identifying sources of resistance to aflatoxin has been slow primarily because genotype evaluation in the field (sporophytic selection) is laborious, expensive, and time consuming. Selection at the pollen level (male gametophytic selection) for screening for resistance to aflatoxin did not receive much attention until recently. The gametophytic generation has appropriately been called 'the forgotten generation'. A total of 60-70 % of structural genes controlling traits of the sporophytic generation (plant) are expressed in the gametophytic generation (pollen). This genetic overlap between the sporophytic and gametophytic generations offers a tremendous potential for modifying the sporophyte by applying selection pressure on the gametophyte. A corn plant produces 2 to 5 million pollen grain that can be subjected to selection. Selection pressure applied to pollen produced by a genetically homogeneous, heterozygous plant is expected to produce genetic changes in the sporophytic population. If a significant part of the physiological processes controlling pollen development, pollen germination, and pollen tube elongation were under the control of the haploid gametophytic genome, it would be elongation expected that the genetic variability expressed in the pollen population of a single heterozygous plant would lead to important selection effects due to pollen competition. This expectation was based on the assumption that not only the genetic variability expressed by the male gametophytic population is of haploid origin, but also that several (60-70 % of the total) genes expressed in the gametophytic phase are equally expressed in the sporophyte ("haplo-diploid" gene expression or gametophytic-sporophytic genetic overlap). 3. Haploid production in wheat The anther/microspore culture technique has opened a new dimension for plant breeding and genetic applications. The haploid cultures provide a rapid route to complete homozygosity and to expression recessive genes. However, cereals were known as recalcitrant species for anther culture response for many years. In wheat, the low level of callus induction from microspores and subsequent plant regeneration, and high percentages of albino plants in vitro limited the application of haploids in plant breeding and genetic research for cereal crops. Various efforts have been made to improve the effectiveness of haploid production. Liquid culture of microspores is one method that has been exploited recently for gramineae and dycotyledones. Liquid media improved callus induction from cultured anthers in wheat. The regeneration ability of calli and the ratio of green to albino haploids in liquid culture, however, were usually lower than for agar culture. The increased callus induction also was not consistently attained by other researches. Moreover, the methods recommended in previous reports involved complicated procedures, such as microspore preculture or subculture, serial transfer of anthers or pollen isolation, centrifugation, and resuspension. Most success in haploid production through anther culture has been obtained by manipulating the growth conditions of anther culture donor plants, the culture medium, and culture conditions. However, major antherculture response factors are still strongly under genotypic control, and some genotypes are only weekly responsive to current techniques. 70 Created by XMLmind XSL-FO Converter. 9. New alternatives in plant breeding The yield of green haploid plants in anther culture depends on three independent components: embryoid production from cultured anthers, plant regeneration from the embryoids and the percentage of green plants. These components are heritable characteristics of donor materials, but are affected by environmental condition. The production of haploid plants from wheat anther culture is a practice included in many plant breeding programs because of recent progress in anther culture methodology. The yield of anther culture production is under strong genetic control, affected also by environmental factors. Because of the strong genetic effects on culturability, only a few wheat genotypes have been shown to be efficient for application of anther culture techniques. The other major limitation to wider practical application of anther culture is that green plants occur, too infrequently. Researchers have therefore shifted their efforts to investigating the influence of medium components on the proportion of green plants produced. Adjustments of environmental factors to increase green plant frequency include (i) increasing the sucrose concentration of the induction media, (ii) increasing the temperature during the callus induction phase, (iii) use of alternative carbon sources and concentrations, and (iv) improving the growth conditions of anther donor plants. Many of the reported effects of medium modifications and pretreatments may be related to osmotic potential. In addition, if the hypothesis about the importance of medium osmotic potential is correct, more attention also should be paid to establishing the optimal osmotic potential for regeneration media. Currently, most researchers use 90 gL-1 sucrose in incubation media, but only 30 gL-1 sucrose in regeneration media. Because of the hydrolysis of sucrose in induction media, difference in osmotic potentials between the induction and regeneration media during the transition phase of culture is much greater than expected. This difference also may have a significant impact on green plant percentage. The broad variety of diploid and double haploid lines produced in vitro are increasingly used in practical breeding programs (Figure 46.). We have employed anther culture and tissue culture techniques to develop lines with resistance to aflatoxins and herbicides, and other desirable agronomic traits such as fast drydown rate, better stalk and seed quality, and weevil resistance. As figure shows we used a complex maize breeding program that was developed to obtain desirable agronomic attributes and speed up the breeding process and enhance the genes responsible for them. Depending on the nature of the source material, e.g., synthetics, Fl, open-pollinated varieties, as well as the breeding aims, one or more haploid (pollen) or diploid (tissue) steps have been made and the Fl hybrids or selfed progenies in later generations have served as source material for haploidization or tissue cultures. One haploid step followed by selection in the greenhouse/field or diploid step after selection on a cell/plant level during the first androgenetic (Al) or diploid progenies (Dl) and two subsequent selfed generations (A2, A3, D2, D3) proved to be the most efficient procedure, if characters from related varieties were to be combined. To make this genetic manipulation system more efficient, we have combined it with several backcrosses. 9.2. ábra - Figure 46. : Complex maize breeding program in Debrecen 71 Created by XMLmind XSL-FO Converter. 9. New alternatives in plant breeding D1: first diploid generation S1, S2, S3: self pollinated progenies D2, D3: two subsequent generation s: selection in greenhouse/field A1: first androgenetic generation A2, A3: two subsequent generation Combination of conventional and new genetic recombination methods (in vivo and in vitro genetic manipulation) may result in such cereal varieties and hybrids as could meet production demands better. The efficiency of anther and tissue cultures in most cereals such as maize and wheat have reached the stage where it can be used in breeding programs to some extent and many new cultivars produced by this system have now reached the market. 72 Created by XMLmind XSL-FO Converter. 10. fejezet - 10. Resistance breeding 1. Stress resistance Plant growth and development is a result of the interplay between the genetically governed potential of the plant and the plant environment in which it grows. This may be symbolized by the equation: P= G x E, where P = the phenotype (what actually develops) G = the genotype (the genetic potential) E = the environment (soil, water, air, light, and other nonliving factors influencing plant growth and development) Fitting crops to environments is as old as plant culture in agriculture. Plants are grown where they are grown for good reasons. Each species has its limits of adaptability, and if environmental stress becomes severe enough, it is better to turn to species more suited to the environment than to attempt to grow unadapted ones. Agricultural research in Hungary and other countries in Europe focus on improving the stress resistance of crops and increasing the competitiveness of production. In this respect, it is important to rationalize the conventional technological elements, reduce the application of pesticides and fertilizers and produce new varieties suitable for environmentally-sound production. For that reson we examined quality and resistance in our winter wheat breeding programme, emphasizing those progenies which originated from wheat varieties which can give high yields without any application of fertilizers. We selected these descendants from F2 generation. The strict selection used was built on the ear-to-row procedure and the crossed generations were tested in microreplications. We have taken off the intermediers which have favourable characteristics for organic agriculture in a recombinant form. 1.1. Indirect breeding for stress environments The first major category of breeding for stress environments is the indirect method, in which the material, although not being tested directly for environmental stresses, is exposed to such stresses in regular field performance trials. If stress environments are a problem within the territory a cultivar is supposed to serve, the testing program should screen for those stresses; therefore, strains with the ability to perform well will excel under such conditions. In selecting for high yield and quality, the breeder automatically will choose those strains with stress-resistant characteristics. As such, breeders generally do not make any measurement or select directly for stress tolerance. Cultivars bred without direct selection pressure for stress resistance nevertheless have considerable variability in resistance to stress. This is probably due to the indirect selection pressure imposed by performance trials conducted in the area of growth where the stress conditions exist. If breeders place test locations on soil that is atypical of the region in which the crop is normally grown, they may release cultivars that are susceptible to some stress in the environment. 1.2. Direct breeding for stress environments The second category of breeding for stress environments involves deliberately choosing testing sites that represent stress conditions reliably and uniformly. Temperature and moisture are notorious for their unpredictable variability from location to location and year to year. Soil problems generally are not variable from year to year but may vary greatly from location to location. This makes it difficult for the breeder to get an adequate sample of environments within the scope of his testing program. Therefore, breeders select fields that have a stress factor at a level that will discriminate between resistant and susceptible genotypes. There is no value to a test if it is located in an environment that is stress-free or if the stress is so severe that nothing survives. Pathologists and nematologists use this technique by screening plants in fields known to be infested with pathogens and nematodes, respectively. 73 Created by XMLmind XSL-FO Converter. 10. Resistance breeding In short, better knowledge about the mechanism for resistance and genetics of the response should enhance breeding progress for stress resistance. 1.3. Breeding under precise, laboratory-controlled conditions The third category of breeding for stress environments involves providing precisely controlled test conditions. Field environments, even those specifically chosen for the purpose, are highly variable. To bring more precision into the selection process, particularly in the early screening and selection phases, solution culture tanks or potting mixtures have been developed with a range in salt, pH, or mineral content levels. Controlled environment chambers may be employed to provide closely regulated temperature, moisture, and light regimes. 1.4. Breeding for fundamental causes of stress resistance The fourth category in breeding for stress environments involves breeding for the fundamental causes of stress resistance rather than measuring the phenotypes of plants. Generally, breeders have knowledge about the chain of biochemical and physiological events between genes and phenotype, so some key step in the chain is known to be highly correlated with stress resistance, the breeder can select directly for it rather than for the final phenotype. 2. Genetic engineering for stress resistance The final category in breeding for stress environments involves genetic engineering. That is, genetic engineering is genetic manipulation (by-passing the sexual cycle) by which an individual having a new combination of inherited properties is established. Genetic engineering presently follows two major approaches: (1) The cellular approach involving the in vitro culturing of (haploid) cells and the hybridization of somatic cells; (2) The molecular approach involves the direct manipulation of DNA. Direct manipulation involves recombinant DNA molecules (a) molecules which are constructed outside living cells by joining natural or synthetic DNA segments to DNA molecules that can replicate in a living cell, or (b) DNA molecules that result from the replication of those described in (a) above. These genetic engineering techniques possibly would be useful for combining germplasms across species and genera which are presently insurmountable by sexual means. However, the technique most likely to be useful in breeding plants for stress environment would be cell culture. For certain stresses, such as salinity, it would seem possible to subject millions of cells in a flask to an elevated level of saline solution with the result that only resistant or tolerant cells would survive. The surviving cells would be treated to regenerate plants that were saline tolerant. Selection for stress resistance The world demand for food and fiber means that crops must be grown on marginal land that is likely to have stress problems. Land now in cultivation experiences stresses from widely fluctuating temperature and moisture conditions, but marginal lands present chronic problems with salinity, acidity, alkalinity, air pollution, and mineral deficiency and toxicity. Breeding for stress resistance and good agronomic procedures are two approaches to increase agricultural production and stabilize productivity. Germplasm offers useful heritable variation for breeding for stress resistance, and this variability has only begun to be exploited. Considerable success has already been achieved in breeding for stress environments, largely by the indirect method. Different alternatives of breeding approaches ranging from indirect selection through field performance testing can be applied. Lot of progress in each category suggest that much more can be accomplished if breeders and agronomists cooperate to codify stress environments and breed plants more tolerant of stress conditions. It seems clear that scientists are becoming more deliberate in breeding for stress resistance; they are looking for ways to measure resistance and seeking the underlying mechanisms for resistance. True, plants perform better in optimum than in suboptimum environments, but plant genotypes can be developed that perform better in stress conditions than cultivars now available; and cultivars can be developed that make agricultural production practical on marginal lands. 74 Created by XMLmind XSL-FO Converter. 10. Resistance breeding Because the first defense of plants against stress is genetic resistance, the work in stress physiology has a primary goal of providing breeders with quick, efficient selection tools to identify resistant germplasms. The terminology used by physiologists may differ slightly from that of plant breeders, but our common goals are to improve the efficiency and productivity of plants under extreme environments. Scientists who work on research in environmental stress seek to define what types of environmental regimes cause stress, the nature of plant injury from stress, and how the stress factors can be ameliorated. And, the use of genetics is the first approach for amelioration. Cultural techniques, chemical therapy, and the like, are other approaches. And, the ultimate goal is to grown productive crop where no crop grows today. To date, plant breeders have done a remarkable job in breeding crops for stress environments, but given better screening tools, the rate of progress will be accelerated markedly. If pests, that is, diseases and insects, as factors that limit crop production are assigned to pathologists and entomologists, the remaining major environmental factors that limit crop productivity and delimit cropping areas are temperature, water, and soil. These factors determine where crops are grown and how much they produce. With the great changes in the world energy situation, however, ideas and viewpoints may need to be altered. Researchers currently are discussing fertilizer efficiency and genotype tolerance to extremes in soil chemical factors. 3. Resistance to abiotic stress 3.1. Salt tolerance of potato Potato production plays an important role in Hungary and the other countries of Europe. Consumption of potato products has increased to a large extent during the past several years. We can satisfy market demands with high quality and virus-free varieties. Results of potato production depend on tolerance/resistance to abiotic stresses. In many cases, increased concentration of NaCl causes yield loss. Selection of salt tolerant varieties proved to be a difficult problem. Nowadays, the salt tolerance of potato varieties can be determined by cell/tissue/ protoplast techniques. Somaclonal variation provides a great potential for selection of lines resistant to salt stress (Figure 47.). In vitro shoots and callus, derived plantlets selected for salt tolerance/resistance provide material for micropropagation. 10.1. ábra - Figure 47.: Somaclones propagated at different NaCl concentrations 75 Created by XMLmind XSL-FO Converter. 10. Resistance breeding In vitro shoot development of potato (Solanum tuberosum L. cv. Kuroda) was investigated under salt stress (40 mM, 80 mM, 120 mM NaCl) conditions. Shoot heights of plantlets cultured under salt conditions were lower than the control through the investigation (Figure 48.). However, the shoot development of plantlets originated from in vitro meristems was almost at the same level as the control under 40 mM NaCl concentration. 10.2. ábra - Figure 48.: Length changes of in vitro cultivated plants by the effect of NaCl 76 Created by XMLmind XSL-FO Converter. 10. Resistance breeding There was no significant difference in the in vitro biomass production between control and treatment with 40 mM NaCl concentration. The dry matter mass decreased upto 120 mM NaCl concentration. There is a need for more investigation of different genotypes and for a conclusion as to whether in vitro tolerance could occur under in vivo circumstances in plants originated from somaclones as well. 77 Created by XMLmind XSL-FO Converter. 10. Resistance breeding Under in vitro conditions, we investigated shoot and leaf callus initiation using different culture media with different 2,4-D concentrations. Under dark conditions, callus induction of shoot/leaf decreased as the 2,4-D concentrations increased. In light conditions, there was a little callus induction, while callus initiation from the shoot from 5 μM to 12 μM 2,4-D concentration showed a significant increase. 3.2. Adaptation of maize lines and hybrids to abiotic/biotic stresses Plant resistance is a promising control method for the most damaging insect pest of maize, such as Diabrotica virgifera virgifera LeConte. We used maize lines derived from a backcross breeding program to transfer resistance into inbred lines. Maize hybrids may inherently differ in their ability to tolerate rootworm injury and partition biomass in response to injury and other stresses. Transient reductions in photosynthetic rate occurred in rootworm infested maize, which after a lag period, led to significant reductions in plant height. High variability among European maize varieties was occurred. Fusarium species reduce yield and seed quality of maize in Hungary and worldwide by causing stalk, root and ear rot. Damage from Fusarium sp. depends on principally factors of cultivation technique/weather (e.g. insects control, hail etc.) and the genotype of maize. Recent herbicides with a great effectiveness, particular spectrum and quick degradability affect adversely plant in many cases. Solution of safe herbicide application is to select mutant lines/hybrids. 78 Created by XMLmind XSL-FO Converter. 11. fejezet - 11. Resistance to biotic stress Winter wheat is the most important cereal in Hungary. The average yield of this crop is very high and was among the world 's highest yields. In spite of the high production potential of the Hungarian soils and wheat crops, there are a number of problems. Wheat production is limited by soil moisture conserved during the summerfallow year. And several diseases can limit the quantity and quality of wheat crop. Among these diseases, the soil-borne diseases are particularly important and the different varieties do not have resistance to the cereal pathogens involved. All the research in Hungary and other countries in Europe focusing improving the quality of crops and increasing the competitiveness of production. In this respect we have to rationalize the conventional technological elements, reduce the application of pesticides and fertilizers and produce new resistant varieties suitable for environmentally sound production. During the past decades because of lack of financial resources, the level of Hungarian wheat production has decreased and the amount of outputs was much lower from year to year. The serious problem was the small quantity of applied fertilizers and the other externals, which focused our research to develop new wheat varieties for sustainable development. We have a large number of winter wheat varieties grown in Hungary (approximately 100). They are very different not only in their genetical background but their reaction to the environment and agrotechnology. That is the reason we have to investigate the behavior to the ecological conditions. We are convinced that the future in cereal breeding is the selection of adaptable varieties which are produced under unfavourable circumstances. In the breeding programs, physiological parameters of the wheat varieties are investigated, such as nitrate reductase activity. There are large differences among the resistance of different genotypes. Some new techniques are used in order to breed varieties with better characters. Our challange in the sustainable winter wheat production is to decrease the susseptibility of varieties to diseases and increase their resistances. In case of winter wheat, our disease problem is dryland foot rot (Fusarium culmorum), root rot (Gibberella zea), Cercosporella foot rot which also causes a disease called scab or head blight. We have leaf and stem rust (stripe rust), snow mold (Fusarium and Typhula) and different bunts (Tilletia, ssp). In the following Table 19. shows some pathological data of middle winter wheat varieties in our experiments. 11.1. ábra - Table 19.: Pathological data of middle winter wheat varieties (infected surface%) (National Institute for Agricultural Quality Control) (2005-2010) 1. Resistance to aphids in maize (Zea mays L.) There are serious yield loss caused by aphids in forage and seed maize production in certain years with a high precipitation and humidity. As a subsequence of aphid epidemy, physiological processes of plants are slowing and particular phenophases are late with 4-7 days. The male/female flowering will start later and last longer. The final solution would be to select tolerant/resistant maize inbred lines and their F 1 hybrids for conventional/bio 79 Created by XMLmind XSL-FO Converter. 11. Resistance to biotic stress protection without application of any insecticides. In case of larger fall amount and relative humidity aphid injury can occur in maize in view of feed or seed production. Tassel and pistil flowering happens later and the process is longer also. Grain development and ripening happen also later. Without prevention, one can count with significant yield and quality loss. In addition to aphids, Fusarium spp. and Ustilago maydis can cause secondary injuries. An alternative, environmental friendly solution of the very effective commercially available insecticides is to produce aphid resistant lines and hybrids (mainly organic production). Aphids of maize are the following: Aphis fabae Scopoli, Rhopalosiphum maidis Fitch, Schizapis graminum Rondani, Rophalosiphum padi Linnaeus, Anoecia corni F.) and Tetraneura ulmi Linnaeus. Most of the aphid species has only one host (monoecious), they live one or some species of a genus. About 10% of aphids are heteroecius: these species live on primary, xyloid host plants during fall, winter and spring. During summer, they injure secondary, taxonomically different hosts. Heteroecious aphids are considered ad polyphags, despite they live on one host. Aphis fabae Scopoli is a heteroecious species for which plant abundant in water give favourable conditions in the second half of the summer, just like maize and sunflower. Aphids and other parasites cause significant yield losses all over the world. It is important to study genetic resources of maize in view of complex inheritance of tolerance/resistance. Also very important is to choose resistant sources correctly. Modern molecular techniques can help to transfer resistance genes into crops. Aphids spread on more and more plants and cause economic damages. In the maize zone of the USA, more attention is paid on them. A lot of publications dealt with the biology, cytogenetics, molecular genetics of aphids and the symbiotic genome. So far, only a few results were published about breeding against aphids despite their enhanced population causes yield loss and the spread of viral diseases. Some authors consider the inheritance against aphids complex, some consider it simple. In inheritance multiplex genes play roles, significantly influenced by environmental conditions. Development of resistance lines with breeding are also considered as important. 2. Investigation of obligate parasites (Striga Spp.) in vitro cirumtances Obligate parasites, among them the Striga species (witchweed) that belong to the Scrophulariaceae family cause serious damages to the majority of our most important crops (maize, sorghum, panicum, rice, sugar cane). Parasite weeds belonging to the Striga and Orobanche species influence the lives of over 400 million people living in Africa, the Near-East and India by reducing the crop of species belonging to the Gramineae family and the Legumineae order. Parasite seeds being able to be viable in soil for as long as 20 years have dormancy and germinate in case the stimulating effect of chemicals exuded from the root of host plant prevails. In case of the Striga, the seeds that started germinating attach themselves to the roots of the host plant with haustoriums, drawing the majority of its nutriments away from the host plant therefore reducing crop yield. In the course of investigation several groups of stimulants were isolated: sorgoalectone from sorghum; strigo from cotton, maize and panicum; alectrol from cowpea (Vigna unguiculata L.). Quite a few synthetic strigol analogues posses various structures (GR5, GR7, GR24) and regulating activity. In vitro techniques offer new possibilities for the analysis of host-parasite interrelationship. Most Striga species have a complex life cycle that can be divided into the following separable phases: dispersion, after-ripening, conditioning, induction of haustorium, attachment, penetration, development of seedling, emergence, and flowering. Our researches on the discovery of the system of host-parasite interrelationship included sprouting Sorghum (bicolor) seeds using normal and modified CC fostering soil (CCDM, CCE) , concerned the quantity of the exudate originating from seeds and artificially fostered roots, their repeated usage. We conditioned different solutions (distilled water, GR 24 and S Bicolor exudate) and different Striga hermonthica branches. In the system of Sorghum-Striga interactions in vitro circumstances we examined the question if parasite seeds germinate in different fostering soils. We made experiments with Fumonisin in the phases of conditioning and early development in order to examine the inhibitor effect. Plant tissue cultures have more advantages over the intact plant systems when host-parasite interactions are investigated in vitro. The identification of resistance genes against obligate parasites and building them into cultivated plants is the only possible solution for developing countries lacking resources. The development of plant regeneration systems makes the application of stable gene transfer possible. 80 Created by XMLmind XSL-FO Converter. 12. fejezet - 12. Environmentallysound plant breeding 1. Genetic manipulation in agricultural environment Genetic manipulation may not replace any conventional method in crop breeding programs, it can be an important adjunct to them. Plant regeneration via tissue culture is becoming increasingly more common in monocats such as corn (Zea mays L.). In vitro culturability and regeneration ability of corn decreased as homozygosity increased, which suggested that these two attributes were controlled primarily by dominant gene action. Pollen (gametophytic) selection for resistance to aflatoxin in com can greatly facilitate recurrent selection and screening of germplasm for resistance at a much less cost and shorter time than field testing. Integration of in vivo and in vitro techniques in maize breeding program is necessary to obtain desirable agronomic attributes, speed up the breeding process and enhance the genes responsible for them. The anther/micropore culture technique has opened a new dimension for plant breeding and genetic applications. However in wheat, the low level of callus induction from microspores and subsequent plant regeneration and high percentages of albino plants in vitro limited the application of haploids in wheat breeding and genetic research. Components of culture media influence the physical environments related to anther culture ability, as well as the nutrient balance and availability. Components in the culture media have a significant influence on the percentage of green plants from wheat anther culture. A higher concentration of maltose produced significantly higher percentage of green plants than the medium containing sucrose. Culture medium effects on albinism indicating that the percentage of green plants in wheat anther culture can be increased by optimizing medium osmotic potential. The production of haploid plants from wheat anther culture has become a practice included in many plant breeding programs because of progress in anther culture methodology. The efficiency of anther and tissue cultures in most cereals such as maize and wheat have reached the stage where it can be used in breeding programs to some extent and many new cultivars produced by genetic manipulation have now reached the market. In the past, conventional breeding methods have been quite successful in improving crop yields throughout the world. Genetic improvements have been estimated to account for about 50 % of the total realized gains in yield per unit area of major crops over the past fifty years. Rates of gain attributable to genetic improvement have generally been linear and show no sign of slackening. The role of breeding methodology to improve cereals for grain yield, disease resistance and other agronomic traits is significant. Conventional breeding methods, however are laborious and time and space consuming. Successful employment of anther or pollen culture could accomplish development of plants with 100 % homozygozity in much shorter time, thus making the breeding process more efficient in use of critical resources. Whereas, in vitro methods, such as anther or tissue culture, may not replace a conventional breeding program, they can be an important adjunct to it. The discovery of viable haploid plants and doubling of their chromosomes mean that homozygous plants could be obtained in one generation. Nevertheless, haploidization techniques applied to many species appeared only during the midsixties with the in vitro culture of pollen grains inside the anthers. The double haploids may allow: 1. Production of strictly homozygous lines with a progeny as homogenous as possible. 2. Considerable saving in time for homozygous line development. 3. Direct choice of individuals for genetic characters. The other possible option for increasing variability is to establish regenerable cell cultures and to improve the economically important traits of cereals. Although all plant cells are derived from fertilized egg cell and contain identical information, callus derived from somatic cells varies in its competence to express totipotency. Cell and tissue culture per se generates genetic variation detected in the progeny of regenerated plants (somaclonal variation). Success in the selection of mutants resistant to pathotoxins, aminoacids and their analogs, and herbicides/insecticides demonstrates the usefulness of tissue culture selection of biochemical mutants for crop improvement. 81 Created by XMLmind XSL-FO Converter. 12. Environmentally-sound plant breeding 2. Wheat and maize improvement for sustainable agriculture Sustainable agriculture has emerged as a key issue in agricultural development, because of widespread of growing concern about the seriousness of degradation of the environment and ever-increasing pressures on agricultural production from continuing rapid population growth. Lately, there has been emphasis for development of maize and wheat genotypes for sustainable production both in developed and developing countries. In the former case, identification of maize and wheat genotypes, which can sustain high productivity at a reduced rate of nitrogen application, has been made because of environmental consideration; while in the latter case socio-economic factors compel application of a low dose of nutrients. The growing concern about the negative impact of present-day agricultural practices on the environment is a driving force behind the development of varieties, which are friendly to the environment and provide many of the answers to the problems of today and tomorrow. Agricultural research in Hungary and other countries in Europe focus on improving the stress resistance of crops and increasing the competitiveness of production. In this respect, we have to rationalize the conventional technological elements, reduce the application of pesticides and fertilizers and produce new varieties suitable for environmentally-sound production. In modern maize production, we have enhanced productivity primarily by changing the cropping environment to fit the hybrids' demands. This strategy is becoming increasingly expensive, both in the cost of inputs and in environmental impact of these products, particularly when inappropriately applied to the production field. In sustainable corn production, hybrids are needed that are productive under a range of environmental conditions. We have used a cyclotron, providing fast neutron irradiation and different biotechnological methods to induce parent lines with sustainable features. We have found several suitable traits of interest in high yielding, welladapted inbred lines that have superior combining ability, as compared to bringing those characters from nonadapted introductions. In our opinion to develop maize hybrids for sustainable production is the identification of genetic combinations that not only tolerate stresses, but also respond to favourable growing conditions. We use integrated methods of in vivo and in vitro breeding in order to increase the efficiency of selection for sustainable agriculture. In our winter wheat improvement program, we have made crosses of clustered varieties with different genetic background. Plants of each of the crosses were grown individually in the second generation and selected as a single plant followed by ear rows. They were selected rigorously by the criteria of sustainable production and efficiency parameters. In profitable sustainable wheat production, input use efficiency and tolerance to minor agrotechnical deficiencies are important, since much of the world's wheat is grown under low input conditions. Though semi-dwarf wheat are relatively more efficient, than older varieties in using nitrogen at all level of nitrogen availability, they too show considerable variability with regard to nitrogen use efficiency. Genetic variation of our winter wheat varieties has been attributed to two multifactorial components: several genotypes have differed in the efficiency with which the nutrients in the plant are utilized to produce a yield (utilization efficiency) and/or they varied in their effectiveness in absorbing nutrients from the soil (uptake efficiency). Different conventional and biotechnological methods have been used for creating combinations, applicable to sustainable production. The world wide demands for sustainability in general are related to the sustainable development of rural areas in particular. Our maize and winter wheat breeding programs compare basic requirements of sustainable development with some features of recent strategies, such as integrated agriculture (integrated pest management and integrated plant nutrition systems) and low input sustainable agriculture (LISA). Special attention is given to the principles underlying autonomous ecosystem management (AEM) as applied in organic types of agriculture (OA). 82 Created by XMLmind XSL-FO Converter. 13. fejezet - 13. Wheat and sorghum breeding 1. Evaluation of prevailing tendencies wheat quality Currently, the Hungarian wheat industry is operating in a buyers’ market place and is facing strong competitive pressures. These pressures are not only due to an oversupply wheat situation and changes in political marketing strategies in some of the major wheat exporting countries, but also because of changes in the economic and agricultural policies of many former large wheat importing countries. Coupled with these changes there have been significant technological developments throughout the world in the flour milling and flour processing industries which are aimed to producing a better quality product more economically. These advances have exerted additional pressure on the seller, not only in terms of having to supply a more uniform product but also in being required to provide more precise specification in contracts. The international wheat market is becoming increasingly more demanding in terms of quality, and it is taking a keener interest in specifications being guaranteed for wheat purchases. Breeding for baking quality in wheat is determined largely by the common negative correlation between yield and grain protein. Over recent decades, wheat breeding for conventional agriculture has concentrated on yields, so that newer varieties generally have higher yields and lower grain protein. Next to yield the most important basic breeding aim for wheat is quality for milling and baking. The precise needs vary, however, depending on the market use. For the global model, supermarkets usually depends on industrialized milling and baking, using cereals with a constant and high protein content, with relatively hard gluten. Cereals for regional and local markets are often produced for artisanal milling and baking, in which there is more flexibility, for example to adjust the baking process to the quantity and quality of the proteins, or to mixtures of different types of flour. However, for whole meal bread products, the process can be complicated, because the high fiber content itself can modify the behavior of the gluten. Breeding for baking quality in wheat is determined largely by the common negative correlation between yield and grain protein. Over recent decades, wheat breeding for conventional agriculture has concentrated on yield, so that newer varieties, generally, have higher yields and lower grain protein. To compensate for this, there has been selection for higher gluten quality, together with improved fertilizer distribution over the season. As regard the quality, it means that in the spreading sustainable agriculture – with limited opportunities for improved fertilizer distribution – the same modern varieties have lower yields together with levels of protein that often do not fulfill the requirements of the baking industry. The effect of commonly applied, reduced N input may vary however with the climatic conditions: in continental/mediterranean climates, where drought occurs often during grain filling, protein contents and consequently baking quality, may be considerably higher than in temperate climates. More recently the main breeding aim for wheat is to dissociate yield from grain protein, so that, even at relatively low yield levels, the grain produced can have acceptable baking quality. Since there is little genotype x environment interaction (GEI) for grain protein content and gluten quality, specific selection for the latter, may prove difficult. However the lack of GEI also means that selection for quality traits can be indirect for example, under conventional agriculture conditions, including also the use of HMWGS (High Molecular Weight Glutenin Subunits) markers. A problem might be that most varieties with high protein content often have softer gluten, which reduces baking quality. A future challenge in breeding quality production will be, therefore, to develop good lines combining high protein content with high gluten quality. Nutritional quality is one of the critical questions in marketing any wheat products. Although it is hard to find definitive data in the literature, the available information suggests that newly released varieties contain significantly more antioxidants (especially fat soluble antioxidants). Indeed, in organic produce and more healthy products, increased antioxidants and bioactive compounds important in plant defense systems seem to be a general feature making them an excellent source of functional and dietary food. Concerning mineral nutrients, breeders should be able to increase mineral concentration in modern cultivars without negatively affecting yield. 83 Created by XMLmind XSL-FO Converter. 13. Wheat and sorghum breeding The major long term marketing problems confronting the wheat industry are, firstly, the reduction of protein levels. There is also increasing evidence that due to the higher yield of newly released wheat varieties and declining soil fertility, there is the potential that lower tonnages of excellent wheat grades will be segregated in the future. The second problem is the reduction in dough strength in some of the premium wheat grades. It is difficult to quantify the environmental effect on protein content and dough strength compared to the varietal effect. However, it is a fact that the majority of recent varietal releases have exhibited inherent protein levels of 0.1 to 1% below the protein content of the wheat variety to be replaced, and many of the varieties released for the premium grades have dough strengths below the required minimum. The need for increased sustainability of performance in cereal varieties, particularly in organic agriculture (OA) is limited by the lack of varieties adapted to organic conditions. In the Table 20., the kinds of breeding strategy and their application to the three generalized forms of OA relate to the three approaches to naturalness. A large number of cereal varieties are available across Europe from conventional breeding programmes, some of which will fulfill partly the requirements of OA. However better varieties are needed to optimize OA systems and to comply with the required product quality under low input, OA conditions, particularly for regional and local marketing. 13.1. táblázat - Table 20.: General characteristics of cereal breeding trends for a continuous of organic faming systems (Verhoog et al. (2003) and Lammerts van Bueren and Struik (2004)) Classification of Orientation organic farming breeding system/product Marker: commodity of Genetic resources global BFCA* Priority of traits Naturalness component in focus Advanced breeding Industrial quality Non-chemical lines and varieties traits, high grain approach yield, targeted agronomic traits BFOA Driver: economics OPB Product: uniform Marker: regional BFOA Advanced breeding Artisan and regional Agroecology lines and former quality traits, approach regional varieties agronomics robustness Driver: agroecology OPB Product: variable more Market: local Driver: social Product: crop. livestock, farm, landscape, society as a whole OPB including evolutionarybreeding, maintenance of genetic diversity and cultural heritage Locally genotypes; adapted Multifunctional crop/ Integrity interspecific crop approach populations conservation integrated with local varieties and environmental needs, landraces environmental flexibility and robustness of life The basic aim of wheat breeding programmes is the improvement of yield and the incorporation of resistance. This has led to a philosophy in the release of wheat that it is sufficient to maintain “quality” and that small decrease in “quality” (for one or more parameters) are acceptable. This philosophy from the marketing viewpoint is not acceptable as most wheat flour products have a minimum protein level in the flour below which it is impossible to guarantee the production of a satisfactory product. Assuming a satisfactory balance of dough properties, this minimum figure for flour protein (except for biscuit flour) is approximately 9.0 – 9.5% (14% moist). At normal flour extraction rates this equates to a wheat protein in the range of 10.0 to 11.0% (11% moist). 84 Created by XMLmind XSL-FO Converter. 13. Wheat and sorghum breeding The traits (Table 21.) required reflect the fundamental differences between OA and CA in the management of soil fertility, weeds, diseases and pests, together with the different demands on product quality and yield stability under organic conditions. 13.2. táblázat - Table 21.: Traits for winter wheat that is relevant for the prevailing tendencies (Lammerts van Bueren et al., 2002). Trait Organi Remarks c Tillering capacity +++ Better differentiation found in ORG by the authors Regeneration ability after +++ harrowing In low input systems harrowing is ralely practised Crop ground cover* +++ Important for weed suppression ability, see below Rapid nutrient uptake +++ Periods of drought and nutrient deficiency can be overcome Weed suppression ability* +++ Plant height genotypes* – taller +++ Tallness can enhance stability in yield and quality in both systems Good shading ability* *** Advantageous also for soil water conservation Competition ability* +++ In high input systems sometimes disadvantageous for yield level Nutrient efficiency* +++ Approximated by measurement of protein yield per unit area Good rooting ++ Disease resistance* +++ Wide adaptation ecological +++ More and other diseases are important in ORG Could enhance stability in more diverse ORG environments Adaptation to local climate ++ Is measured through yield level and yield stability in relation to adapted standard varieties Specific adaptation example: special quality traits in (local) specialities +++ Adaptation to variability +++ of influencing factors A variety for ORG must not have any major weekness Grain yield* ++ Yield has to be seen in relation to quality; protein yield is usually influenced to a higher degree by grain yield than by protein content Yield stability +++ Eventually less cultivation measures exist for control in ORG High protein content* +++ Minimum requirements are common in ORG marketing Grain quality* quality/baking +++ Mostly higher, but sometimes different kind of demand for quality in ORG 85 Created by XMLmind XSL-FO Converter. 13. Wheat and sorghum breeding *Traits that are scored/measured in organic Value for Cultivation and Use (VCU) test +, ++ and +++ are given for increasing importance In our selection programme of the flour and dough properties of the ‘HP Pusztaszél’ cultivar (Triticum aestivum L.) was measured (Table 22.). 13.1. ábra - Table 22.: Flour and dough properties of ‘HP Pusztaszél’ cultivar (Triticum aestivum L.). Central Agricultural Office, 2001-2003) We investigated the dough quality with farinograph and valorigraph in our registered and patented varieties (Table 23.). Out of the valorigraph results the FQN was of high importance which was the highest one in case of ‘HP Pusztaszél’ (110). It reflects to the dough resistance and the capability to apply this cultivar in intensive baking industry. In terms of gluten evaluated in Glutomatic system the Gluten Index (GI) proved to be the highest one in ‘HP Pusztaszél’ (72.85) which indicated excellent hard gluten. Looking at the alveograph parameters ‘HP Pusztaszél’ had a high W value (210.3·10 -4 J) coupled with 1.43 P/L. This is strong evidence that this cultivar’s dough is outstanding for baking industry to produce high quality bread. In cultivar ‘HP Pusztaszél’ the P value - which stands for maximum pressure that was withstood - was 89.5, while L value (stands for the height of the bubble) was 63.5. Additional advantage is that ‘HP Pusztaszél’ cultivar has about the same yieldibility as the standards. 13.3. táblázat - Table 23.: Dough quality of winter wheat cultivars Cultivar GI FQN HP Árkus 60.25 80 HP Mező 56.90 HP Rárós P(mm) L(mm) P/L W(10-4J) 119.0 52.5 2.27 242.2 75 45.1 102.0 0.44 133.1 56.30 70 43.1 113.5 0.38 143.0 HP Pusztaszél 72.85 110 89.6 63.5 1.43 210.3 86 Created by XMLmind XSL-FO Converter. 13. Wheat and sorghum breeding 2. Amino acid compositions in wheat species with different genomes The issue of analysing diploid, tetraploid and hexaploid wheats is topical since ancient varieties can play significant roles in contemporary agriculture. Some can be gene sources and valuable crops. The domestication began over ten thousand years ago with "einkom" (Triticum monococcum L.) while the modem hexaploid hard wheat for bread making emerged eight thousand years ago. In addition to wheat varieties of the aestivum type, durum wheat (T. durum Desf.) (contains genomes A and B, 2n=28) is produced on the remaining approximately 5% of the wheat production area. Recently, primarily in the hilly regions of Europe the production of the spelta (T. spelta L.) wheat variety, which contains hexaploid genomes (A, B and D genomes, 2n=42) has come to play a major role and is used in organic farming due to its smaller nitrogen requirement. Einkom (genome A and 2n=14) is only grown occasionally for animal nutrition. The einkom or diploid wheat is the earliest grown bread variety in production and one, which is supposed to have been the donor for genome "A" of the hexaploid (Triticum aestivum L.) and tetraploid (Triticum durum L.) varieties. Today the ancient varieties are only grown in marginal areas of former Yugoslavia, Turkey and Italy. Protein contents of the hexaploid and durum wheat varieties are high in Glu but low in Lys and Thr, which are especially important for human diets. Reaserchers compared 15 T. monococcum strains, two modern hexaploid and two durum wheat varieties as regards their protein contents and amino acid compositions. In the case of the einkom strains there was positive correlation found between Glu and Pro values and protein contents while the same relations exhibited negative correlations as regards Thr, Ile, Leu, Asp, Ser, Gly and Ala. The protein contents of grains ranged between 1425% when analysing T. monococcum strains but no types high in Lys concentrations could be selected during tests. When analysing the amino acid compositions, in the varieties (44 spring wheat varieties) analysed, there were found considerable variability in Lys, Leu, Ile, Thr, Val, Met, His, Arg, Ala, Asp, Glu, Gly, Pro, Ser and Tyr contents. When analysing the protein and amino acid contents of wheat varieties, we found significant differences in protein concentrations, which findings were similar to those in certain amino acids. Amino acid content of wheat is also influenced by several other factors: amount of nitrogen, damage by pests. When analysing grain crops, the highest protein contents (18.2-20.9%) can be found in varieties T. monococcum, T. spelta and an old Hungarian bread-wheat variety Bánkúti 1201. Similarly, the highest total amino acid contents were found in these varieties. When analysing the total protein amount it was found that in the so-called ancient varieties that the amount of total protein per total milled volume was varied in the case of einkom varieties and the highest values (17.7%) were provided by the hard-grained spelta variety. When analysing biochemical parameters of stored proteins it was similar amino acid compositions in the wheat varieties and strains of the aestivum type and the essential amino acid level also showed a negative correlation with the total protein amount. The quality of plant and thus that of wheat is shaped by the genotype, the environmental conditions and their interactions. The contemporary rich Hungarian choice of varieties offered gives producers an opportunity to use an appropriate genetic base best suited the location of production and the type of management chosen. 3. Effects of amino acid fertilizers on yield and quality of winter wheat Different amino acids influence different vital processes of plants, such as protein synthesis, stress resistance, photosynthesis, effects on stomata. Amino acids are precursors and activators of different plant hormones and growth factors. L-methionine and ethylene are precursors of growth processes, L-tryptophan is of auxin synthesis, while L-arginine induces syntheses of hormones related to flowering and crop development. Regular high scale fertilization raised up the question of trace element retrieval. Primary solution for this can be foliar fertilization since nutrient supply of a plant is better through leaves than roots, according to the literature. Nitrogen mainly has significant effects on yield quality during stalk development and flowering. It was observed that the effect of foliar fertilization can lead to reliable extra yield (increase of thousand kernel weight and seed number of ears) during droughty years. 87 Created by XMLmind XSL-FO Converter. 13. Wheat and sorghum breeding Late carbamide foliar fertilization caused decline in yield. Pharinographic values were endangered mainly by inappropriate fertilization. In contrast, Finney et al. (1957) found that applying carbamide foliar fertilizer after ear development caused yield decrease only altogether with severe absence of N. It was applied spring and late N fertilization in case of winter wheat and concluded that protein contents increased, depending significantly on N amount and time point of fertilization. The protein content was higher in case of application of carbamide during flowering; in contrast, rapid decline in protein amount happened when foliar fertilizer was applied in a different period. There was a clear positive effect on the protein content of the gluten, but there was no advantage of the highest dose of fertilizer compared to the low one. Highest increase in sulphur contents were observed after treatment with 2.5% and 5% DL-methionine - above 2000 mg/kg in both cases while the control’s was 1516 mg/kg. Highest protein contents were the results of 2.5 and 5% ammonium nitrate treatments, above 18%. L-arginine HCl and L-tryptophan (both concentrations) treatments resulted in 17% protein contents. In cases of gluten contents and W values, the tendencies were similar, ammonium nitrate treatments made the best effects - 2.5%: 41.5% gluten content, 5%: 42.3%, while control: 36.9%. 5% L-arginine HCl and 5% L-tryptophan increased gluten content to 41%. W (energy needed for the deformation of dough) was above 400 (*10 -4 J) after treatments with ammonium nitrate and 5% L-tryptophan. Falling numbers exceeded 300 as results of L-tryptophan, ammonium nitrate and L-arginine HCl, so flour can be classified as excellent. After other treatments, falling numbers stayed below 300, so flour can be marked as average. It can be concluded that treatments were effective in cases of protein and gluten contents of the grain. Differences were significant between treatments, except protein contents. Significant differences can be observed in case of W values compared to controls, so treatments were also effective. Except L-lysine HCl treatments, sulphur contents were significantly different compared to controls. Summarizing the observations, it can be concluded that amino acid foliar fertilization treatments effected yield amount and some quality parameters to different extents. Lowest effects were experienced on yield averages (lowest significant differences compared to controls). In addition, traditional ammonium nitrate increased the quality of yield (especially gluten and protein contents) more effectively than any other amino acid foliar fertilizer. 4. Improvement of Sorghum breeding As a result of the declining tendency in the quantity of livestock, the crop structure changed while the milk and meat production intensified in the countries of the European Union. Bioenergy production may open up new perspectives in the future as fossil energy sources decrease, while industrialized countries face growing energy demand. Mutation breeding and heterosis breeding are appropriate methods for the quality improvement of the genetic material base. Silage maize breeding programs aim at the decreasing of lignin content. The dry-matter of the silage made of Brown Midrib maize hybrids is better in digestibility. The optimal dry matter content at harvest (30-40%), the high dry matter yield, the high protein and energy contents and the low fibre content are of great importance in terms of silage quality. The digestibility of silage hybrids is influenced by the selection of the hybrid, the breeding and the time of harvest. The digestibility of the whole silage is determined by the neutral detergent fibre (NDF), the acid detergent fibre (ADF) and the acid detergent lignin (ADL) contents of plant cells. Sorghum is one of the top five imported cereals worldwide. The genetic variability of the sorghum types decrease with the increase of distance from the gene centre, therefore the hybrids produced in Hungary have narrow genetic base. Sorghum hybrids are produced by the application of cytoplasmic male sterility. Single cross hybrids with high carbohydrate content were produced by the Comstock-Robinson crossing model. For the production of the hybrids, male sterile female (A) and restorer male lines were crossed (Figure 49.). The male sterile female lines were re-produced by restorer male lines (B). 13.2. ábra - Figure 49.: Male sterile female and restorer male lines 88 Created by XMLmind XSL-FO Converter. 13. Wheat and sorghum breeding The sorghum hybrids were applicable to the production of good quality silage and bioethanol too (Figure 50.). 13.3. ábra - Figure 50.: Process of the bioethanol production (experimental) Bioenergies (among them e.g. bioalcohol) can be a solutions for replacement of fossil fuels. For its production, plants with high sugar or starch content can be used. Juice pressed from the stalk of sugar sorghum has high sugar content (14-17%) that makes it suitable for bioethanol production. In the case of male lines the following traits were studied: juice content of stalk, dry content, sugar-, reducing, and non-reducing sugar content of stalk juice. We examined the correlation of the above mentioned parameters. The stalk medullas were wet, and the sugar content was between 8,27-18,10%. The highest sugar content was in the case of 4. and 5. nodus at the restorer male lines. The highest sugar content was in the case of RL12. We found strong positive correlation between the dry content and total sugar content (r=0,898**), and between dry content and non-reducing sugar content (r=0,889**), and between the total sugar content and non-reducing sugar content (r=0,953**). Sugar accounts for 82-87% of the refractometrically measured dry matter in the sorghum stalk juice. During drought water soluble sugars have important role in maintaining the osmotic concentration of the cells. The juice pressed form stalks were fermented into ethyl alcohol with the yeast Saccharomyces cerevisiae. Bioethanol may partially substitute the fuel of engines. By the co-cultivation of modern silage maize and sorghum genotypes in 2+2 rows, higher dry matter yield per unity area can be harvested than by the alternative utilization of grain maize as silage. 89 Created by XMLmind XSL-FO Converter. 14. fejezet - 14. MAIZE BREEDING 1. Improve of maize quality (chemical component) The importance of maize production in Hungary is indisputable. The major chemical component of the maize kernel is starch, which usually provides up to 72 to 73 percent of the kernel weight in the commercial hybrids. After starch, the next largest chemical component of the kernel is protein. Protein content varies in common varieties from about 8 to 11 percent of the kernel weight (Figure 51.). Most of it is found in the endosperm. The oil content of the maize kernel comes mainly from the embryo. It was reported that the reached grain oil content in the Illinois High Oil population was 22%, but the upper plateau from selection had not been achieved yet. 14.1. ábra - Figure 51.: Protein, starch and oil content data of the maize genotypes (Debrecen) Among the chemical components decisive for quality, the protein, oil and starch contents can be influenced to a substantial extent by plant breeding, and one of the main aims of maize breeding is to produce hybrids with high starch content. If plant breeders intend to exploit existing genotypes for various end-use applications, a rapid and serial method of determining kernel constituents will be necessary. Traditional wet chemistry tests for protein, oil, and starch are too laborious and expensive for the large numbers of samples generated in a typical plant breeding program. Near-infrared reflectance spectroscopy (NIRS) has been used extensively to measure major constituents in maize and other crops. Significant negative correlations between protein and starch concentrations in maize grain (Figure 52.) have generally been observed in previous investigations. 14.2. ábra - Figure 52.: Correlation between starch and protein content in maize hybrids (Debrecen) 90 Created by XMLmind XSL-FO Converter. 14. MAIZE BREEDING Highly significant negative correlations were observed between protein concentration and starch concentration; protein concentration and grain weight per plant in two populations, while highly significant positive but weak correlation was observed between starch concentration and grain weight per plant. The correlation can be calculated between pairs of traits including starch, protein and TKW in maize lines and hybrids (Table 24.). 14.1. táblázat - Table 24.: Correlation matrix Correlation Coefficient Interpretation 0.00 - 0.25 No correlation 0.26 – 0.50 Low, quite small correlation 0.51 – 0.75 Moderate correlation 0.75 – 1.00 High correlation The protein content may differ in maize hybrids cultivated in our country e.g. 9.7% and 14.3% of PR37D25 hybrid and Furio hybrid, respectively. The hybrid itself had a significant (p<0.001) effect on the starch and crude protein content of maize grain. The crude protein content decreased with increasing starch content. Generally, protein concentration is negatively and positively correlated with the other two grain components, starch and oil concentration, respectively. It was concluded that higher average yield increased the accumulation of starch in the kernel but protein and oil content were decreased. It was found a positive, moderately strong correlation (0.68) between the yield and the starch content, a negative, moderate strong correlation (−0.52) between the yield and the protein content, and a loose negative correlation (−0.19) between the yield and the oil content. Yield increase of maize could be promoted by increasing either kernel weight or kernel number per ear, while that of spring maize mainly depends on increasing kernel number per ear relatively. Spring maize had 19.925.4% more kernel number per ear, and 9.7-16.5% higher TKW compared to summer maize. TKW depended primarily on the genotype and secondly on the weather conditions and fractionation did not have a significant effect on the TKW. In S-2 families (produced from a Pioneer hybrid) the ear length, the grain number per row and the kernel length were in positive, significant correlation with the grain yield. From the same yield 91 Created by XMLmind XSL-FO Converter. 14. MAIZE BREEDING components of S-2 families and of their hybrids, the correlation only between the ear length and thousandkernel-weight was positive and reliable. The strongest correlation between grain yield and TKW (S(1) progenies r(g) = 0.684**; HS progenies r(g) = 0.633**). Desirable, high significant influence on grain yield, in path coefficient analysis, was found for TKW. In case of tropical maize genotypes positive correlations was not found between grain yield and TKW (r = 0.10). Maize genotypes with increased starch content could serve as breeding materials of high promise to meet the requirements of bioethanol production and as the raw material supply of food and feed. 2. Morphology of tassel components and their relationship in maize Discovering the relationships among plant morphological and quantitative features is very important in maize breeding and production, particularly if they can be altered by selection or agro-technical methods. In maize breeding, increased attention is being paid to the selection of features that can help to reach maximum yield with regulation of energy conversion. Such phenomena are plant height, ear height, leaf number, leaf area. In addition, tassel characteristics can influence plant performance and productivity significantly. The male inflorescence of maize is a tassel, components of which are frequently studied. Maize is monoecious, so its reproductive success largely depends on the development of male flower. Morphology of tassel components influencing primarily pollen amount can be significant factors of successful seed production and selection. Several researchers studied relations between pollen and tassel components. A number of authors examined the inheritance of tassel characteristics. It was found 86.1, 45.8 and 28.8% hereditability for tassel weight, tassel branch number and tassel length, respectively. Inheritance of tassel characteristics are not fully clarified. Tassel branch number is related to grain yield and there is negative correlation between them. Tassel weight also is negatively correlated with grain yield; consequently it is positively correlated with tassel branch number. These results imply that selection targeted on the decrease of tassel branch number and tassel size may have indirectly influenced the increase in yield. Selection on smaller tassels decreases the energy of plant consumed for tassel and the shading of flag and upper leaves. Smaller tassel size in the case of male parental lines, however, can cause problems in Fl seed production and the maintenance of male line due to the unsatisfactory pollen production and shed. Tassel branch number is a determinant of pollen amount. Greater tassel branch number is dominant over lesser branch number. It was found negative correlation between individual productions of lines and tassel branch numbers. Tassel components and some quantitative features were compared with Pearson's correlation coefficient determining method. Strength of relations between traits and directions of interactions were determined. With the knowledge of correlations between characteristics one can predict change of features related to each other. Correlations between measured traits may be useful as a means to simplify selection, if the correlation is consistent across genotypes and environments. We find correlation between the following traits. Tassel weight was positively correlated with total number of branches (0.59**), number of primary branches (0.53**) and tassel area index (0.63**), and negatively correlated with plant height (-0.45**) and yield (-0.39**). Tassel area index was negatively correlated with plant height (-0.63**) and yield (-0.55**), as were number of branches and Leaf Area Index (0.39**). Number of primary branches showed medium correlation with plant height (-0.43**). Test weight correlated negatively with plant height, ear height, yield, leaf area index (LAI) and 1000-kernel weight. Because a number of these traits are highly heritable, these correlations could be used as a basis for indirect selection of special features in maize breeding programs. 3. Chemical contents and thousand kernel weight in maize breeding The cultivation of speciality maize has an important role in the word agriculture. The food processors demand maize with increased yields and quality to improve product attributes. Some unique types such as blue corn are becoming increasingly popular in speciality food stores. The analysis of maize grain yields is extremely important for both animal feeding and human nutrition, with the letter coming more and more to the foreground. Some authors studied chemical composition in speciality colour maize. The most important characteristics of the yield components are the number of ears per plant and the thousand-kernel weight. We determined some 92 Created by XMLmind XSL-FO Converter. 14. MAIZE BREEDING chemical parameters of various – special colour and conventional (yellow) – maize trends in the thousand kernel weight of their maize in our breeding program. Protein, ash, starch contents of seed samples were analysed in two parental inbred (UDL1 and UDL 6) lines and their two F1 hybrids (UDH3 and UDH 10), one control (DK 471) and three special colour maize landraces (Hopi Blue, Blaumais, Blau Babyfinger) Table 1. The colour of investigated maize kernels is also presented in the Table 25. Moisture, protein, ash, and starch content of kernel samples were determined using an NIR analyser. Nearinfrared reflectance (NIR) method is commonly used to measure moisture, protein, ash, and starch content in maize. All measurements were reported at 14% moisture adjustment. 14.2. táblázat - Table 25.: Characteristics of investigated maize Lines, hybrids and landraces Kernel colour UDL1 Yellow UDL6 Yellow UDH 3 Yellow UDH 10 Yellow DK 471 (control) Yellow Blaumais1 Blue Hopi blue1 Blue Blau Babyfinger1 Blue 1 Oridin: Austrian Agency for Health and Food Safety (AGES) Genbank, Linz Thousand kernel weights are presented in Table 26. Hybrid UDH 10 had the highest kernel weight (435.6 g) while landrace Blau Babyfinger had the lowest kernek weight with 87.9 g, but landrace Blau Babyfinger had the relative high individual ear/plant (3-4 ears per plant) production. 14.3. táblázat - Table 26.: Trends in thousand kernel weight in the investigated maize Lines, hybrids and landraces Mean (g) UDL1 205.1 UDL6 318.7 UDH 3 413.6 UDH 10 435.6 DK 471 (control) 412.7 Blaumais 373.5 Hopi blue 378.4 93 Created by XMLmind XSL-FO Converter. 14. MAIZE BREEDING Blau Babyfinger 87.9 Differences in protein, starch and ash content among the investigated maize are presented in Table 3., Figure 53. and Figure 54. Landrace Blau Babyfinger had higher protein (13.32%) (Figure 1.). Protein content range from 6.5 to 13.32. Nowadays maize – hybrid – (under Hungarian conditions) kernel contains between 8-11%. Landraces had higher protein content from 10.75 to 13.32. 14.3. ábra - Figure 53.: Trends in the protein content of the investigated maize (Debrecen) No significant differences were detected among investigated maize for ash content determined by NIR (Figure 54.). Landraces Blau Babyfinger had the highest ash content (1.63%), while hybrid UDH 10 had the lowest. Our researches show a range of 1.2-1.63% for ash content. 14.4. ábra - Figure 54.: Ash contents of the investigated maize (Debrecen) Significant differences were observed in the starch content (Table 27.) Hybrids UDH 3 and UDH 10 had the highest starch content, while Blaumais (with 44,98%) had the lowest. 14.4. táblázat - Table 27.: Trends in starch contents (%) of the investigated maize (Debrecen) Starch 44.98*** 59.4*** - 61.22*** 59.24*** 69.11*** 66.39+ 65.88 ***= Significant at 0.001 probability levels for comparison with control (DK 471), += non significant Starch content of Blau Babyfinger proved to be unlikely high with the applied method. 94 Created by XMLmind XSL-FO Converter. 14. MAIZE BREEDING Knowledge of quality characteristics among available genetic materials is also fundamental for the successful maize breeding. For the maize breeder the variability of protein content of investigated landraces is valuable and provides a good genetic material for effective selection. We can use this information to select quality maize hybrid, which can be integrated into breeding programs. 4. Evaluation of blue and red corn landraces under Hungarian conditions A significant part in the food industry of the future will be the production of the so called functional foods which are of natural origin and not only nutritive but enhance the immune system and suitable for health preservation in many other ways. Nowadays there is intensive research worldwide on the production and testing of these types of foods. Blue, purple and red kernel corns could match these criteria and make the basis of functional, health preserving foods with their good nutritional values (protein, antioxidants, micro-elements) and exotic taste. Maize is a very diverse plant with hundreds of races. Nearly all kinds of colours can be experienced among the varieties. These days mainly the yellow and white kernel corns are cultivated, for forage or industry and for food, respectively. The only perspective is the hybrid breeding, utilization of the heterosis effect that reformed maize breeding and cultivation during the last hundred years. With the appearance of hybrid maize varieties productivity and agronomical features increased but nutritional values decreased. Fortunately, old landraces with diverse genetic origin remained on many locations. In Peru and Mexico foods made of corn are still the most important nutrients. Different blue corns are the most widely cultivated ones in these areas. In the last decade scientific interest increased about these plants, a number of studies were made on the investigation of their colourings, on the health protecting effects of their antioxidants. There are only a few papers published about their cultivation or breeding perspectives. Origins, utilizations and characteristics of corns Corn was in connection with many great cultures in the New World such as the Inca, Maya and Aztec civilisations. Nowadays several North American Indian tribes - Zuni, Hopi, Navajo - prefer blue corn. The Southwestern American Indian tribes used blue corn as basic material for foods. The New Mexican cuisine is significantly based on these ancient demands so the cultivation and products of blue corn are from these areas. The nutritional values of the blue corn exceed those of the traditional, widely cultivated yellow ones. Most of the commercial hybrid dent corns are homogenous and can produce a yield of 8-13 tons/ha. The blue corns can give a significantly higher crop so their cost is much larger. Blue corn varieties are very diverse in the features of the plant and the grains. There are ones with shorter and longer flowering times and their ripening times are also can be different. Because of the weakness of the stalks blue corns frequently lean before harvest. Stormy winds can break the stalks causing notable damages. This leads to the decrease of crop and grain quality. Harvest with machines is heavy due to the leaning of the stalks. The different varieties are very susceptible to pests when they are cultivated far from their original locations. In general, the cultivation of the blue is similar to that of the dent corn but in the lack of adaptation it does not react to the intensive technologies. It is favourable to cultivate it under organic conditions, with the application of less fertilizer or other technological elements. Nowadays principally varieties are cultivated but hybrid breeding is also going on; there are some hybrids on the market as well. Blue corn cannot tolerate the excessive fertilization easily. Generally the density of the population is 30-45.000 plants/ha and the application of excessive fertilizer doses lead to problems of stalk strength and quality. During cultivation it is very important to keep the minimum isolation distance (at least 230 m) from other maize types. The number of blue corn races is still unknown. The most frequently cultivated population or landraces are Hopi blue, Navajo and Black-Aztec. Hybrid breeding is going on and hybrids exist. There are problems with the availability of breeding and cultivation basic materials; only some organically cultivated landraces can be purchased from North American companies; the prices of seeds are high because of this. Gene banks can be alternatives. It is practical to get the genetically most diverse materials from the more sources in favour of the most successful breeding avoid drift. It was found significant differences in the ex situ and in situ maintenance of two Hopi corn varieties. We studied 11 blue, purple and red exotic maize varieties on two locations in Hungary to collect the more information about their cultivation and breeding. There is little knowledge about their cultivation in Europe. 95 Created by XMLmind XSL-FO Converter. 14. MAIZE BREEDING Cultivation under organic conditions can support latter stages of breeding (line and hybrid production) and the authorization of the organic seeds. As their antioxidant contents are high, they could be used as functional nutrients, supplements or basic materials for the food industry. With their large height and abundant pollen amounts special blue varieties can play as markers in the studies of GMO (genetically modified organism) versus conventional cultivation, co-cultivation, since the blue colour as a dominant feature make the detection easy. The selection of varieties meeting the requirements of organic cultivation has already begun which will satisfy the increasing needs of bio-nutrient production. Nowadays there is an ever increasing demand on foods of natural origin and a growing ignorance of synthetic additives and colouring substances. The spread of functional nutrients is up-and-coming since health preserving materials can make their effects in natural ways. Antioxidant and antocyanin researches are getting more and more important in the case of vegetables, fruits and cereals. There are intense studies on the colouring materials of the blue corn with special respect to their role in food industry and health care. 5. Importance of blue/red kernel corns Consumption of whole grain cereals can be connected to the decreased risk of some chronic diseases. Several studies dealt with the positive effects of wheat and rye on health. Corn and corn products have little literature despite the fact that corn is consumed in the USA, Central and South America to a great extent. Production of nutrients of natural origin will play a significant role in the food industry of the future. Besides their nutritional values, these foods are good for health preservation and the immune system. They are called functional foods; red and blue kernel corns are among them. It is topical to study mineral contents of special corn types. More than 2 billion people - especially women, newborns and children suffer from diseases caused by improper mineral intake. Mainly lacks of iron, zinc, selenium, calcium and magnesium in diets impose risks for health all over the world. Concentration and distribution of elements in plant organs change according to environmental factors and genotype. B, Co, Cu, Mn, Mo, Se, V and Zn as essential elements for plants, while Ag, As, Ba, Be, Bi, Cd, Cr, F, Hg, Ni, Pb, Sb, Sn and Ti as toxic ones or essentials only for animals. In the case of animals, about 40 elements take part in mineral metabolism. Among them Ca, P, Mg, K, Na, S and CI are macroelements, I, Fe, Cu, Zn, Co, Se, Mo, Ni, and Mn are microelements, while ultra-microelements are F, Cr and Li. Although microelements are vital for biological processes in very little amounts, absence of them in food can cause deficiency diseases. It was compared mineral content of 8 New Mexican blue kernel corn varieties to those of 1 yellow and 1 white dent commercial hybrids. Zinc contents were found to be higher in an of the blue varieties, 7 out of 8 blue varieties had higher iron levels, while boron and aluminium were commonly higher in the 2 dent types. The mineral content of blue corn was more favourable than those of yellow and white hybrids - latter is cultivated for human consumption. In the last decade at CIMMYT, preliminary studies have begun on iron and zinc concentrations of kernels. In Zimbabwe, 1045 breeding lines were evaluated in 1995/96. In the case of iron, values varied between 17.7 and 61.8 μg g-1 while in the case of zinc, the range was 12.9-28.5 μg g-l. It was examined microelement contents of corn samples from different states of USA and found the following concentrations: Zn: 19.0-27.0 ppm, Mn: 4.5-7.7 ppm, Cu: 1.75-2.0 ppm, Cr: 0.02-0.15 ppm. Mineral contents of yellow hybrids cultivated in monoculture and in crop rotations were investigated using various fertilizer doses. Zinc contents of specimens grown in crop rotation were 10.0-19.0 ppm, in mono culture 4.0-7.0 ppm lower, while Mu contents 3.0-5.0 ppm and 1.0-2.0 ppm higher, respectively. During breeding of field plants, little attention is paid on the enhancement of mineral content favourable for human diet. There are many limited data on iron and zinc contents of tropical corn. Based on our results, we can state that genotypes play an important role in mineral content of maize kernels. The mineral content may be improved in maize seeds with application of these genotypes in conventional breeding. In view of macroelement contents, we observed significant differences between red and blue varieties (Table 28.). 96 Created by XMLmind XSL-FO Converter. 14. MAIZE BREEDING 14.5. ábra - Table 28.: Macroelement contents (mg kg-1) of investigated corn varieties (mean±SD; dry weight basis) Average phosphorus value of blue corns was 3527.7±280.0 mg kg-1. Black Mexican performed a surpassing 3859.5±562.1 mg kg-1, as well as in case of potassium content (4325.0±469.5 mg kg-1). Except Ca, blue corns showed higher macroelement contents than red ones. Santo Domingo Blue and Black Mexican had the two highest S contents (1555.0±128.6 mg kg-1 and 1453.0±164.0 mg kg-1, respectively). Mean of calcium content (126.9±27.1 mg kg-1) was higher in red cultivars. Two genotypes, Rotmais (147.5±27.5 mg kg-1) and Black Mexican (143.0±45.2 mg kg-1) exceeded this rate. We experienced this kind of positive difference in case of Black Mexican’s magnesium content (1450.0±104.6 mg kg-1). Ca and Mg values were above those found in literature. In the corn literature, the most results can be found on iron, zinc and selenium in addition to toxic heavy metals, e.g. cadmium, lead and mercury, while there is less information on detailed microelement contents of red and blue varieties. In our study, iron played the leading role (Table 29. and 30.). 14.6. ábra - Table 29.: Some microelements (mg kg-1) of investigated corn varieties I. (mean±SD, dry weight basis) 97 Created by XMLmind XSL-FO Converter. 14. MAIZE BREEDING 14.7. ábra - Table 30.: Some microelements (mg kg –1) of investigated corn varieties II. (mean±SD, dry weight basis) 98 Created by XMLmind XSL-FO Converter. 14. MAIZE BREEDING Except one case, iron contents were above 30 mg kg-1, in Blaumais, Hopi Turquoise and Hopi Blue above 40 mg kg-1 (41-46.3 mg kg-1), which exceeds value in the literature with the average of 10 mg kg -1. Copper contents varied between 2.8 and 3.7 mg kg-1. Zinc values were also ordinary (15.2-31.5 mg kg-1). Manganese contents (6.2-9.3 mg kg-1) were near the averages published (5-8 mg kg-1), while boron contents (<0.1-1.5 mg kg-1) were significantly below the previously experienced (7-26 mg kg-1). Aluminium contents in red varieties (9.4-17.2 mg kg-1) turned out to be higher than in blue ones (mean: 7.4 mg kg-1). Averages of chromium contents were the same in both types (0.4 mg kg -1), Only Santo Pueblo Black, Hopi Turquoise and Black Mexican performed higher values (0.7 mg kg-1). Corn is the most important selenium source for animals and as an antioxidant for humans as well, but in a lot of cases it has to be supplemented in forage since several areas of the world (e.g. Corn Belt (USA) or Hungary) are poor in this element. Genotypes showed broad genetic variability in view of the amount of accumulated selenium too. Each variety performed 0.1 mg kg-1 for lithium content, while in case of sodium values varied between 117 and 181.5 mg kg1, only Hopi Blue had higher sodium content (181.5±75.6 mg kg -1). The strongest positive and significant associations among minerals were between Mg-P, Mg-K, Mg-S, Zn-K, Zn-Mg, Mn-Fe, Se-B, Se-Al concentration seems to be highest positively associated with other mineral concentrations (Table 31.). 14.5. táblázat - Table 31.: Pearson’s correlation coefficients between investigated elements (without Li) P K S Ca Mg Fe Cu Zn Mn 99 Created by XMLmind XSL-FO Converter. B Al Cr Se Na 14. MAIZE BREEDING P 1.000 K 0.675* 1.000 S 0.431 0.620* 1.000 Ca 0.208 0.235 0.202 1.000 Mg 0.726* 0.777* 0.681 0.273 1.000 * * Fe 0.285 0.151 0.270 0.171 0.355 1.000 Cu 0.404 0.133 0.447 -0.393 0.396 0.280 1.000 Zn 0.370 0.728* 0.549 0.275 0.813* 0.052 -0.068 1.000 * * Mn 0.541 0.369 0.351 0.489 0.508 0.707 -0.033 0.373 1.000 * B -0.351 -0.358 -0.470 -0.175 -0.018 0.034 1.000 0.403 0.200 0.407 Al .0.186 -0.293 0.356 -0.222 0.083 0.179 0.402 1.000 0.393 0.163 0.663* Cr 0.120 0.328 0.141 0.605* 0.492 0.057 -0.209 0.511 0.222 -0.240 0.188 1.000 Se -0.358 -0.396 -0.279 0.364 0.666 -0.446 0.034 0.840* 0.706* 1.000 0.581 0.150 * 0.121 * Na 0.601* 0.153 0.097 0.141 0.278 0.081 0.134 -0.030 0.521 -0.115 -0.195 -0.229 1.000 0.227 In the last decades, interest has increased in potential health preserving effects of whole grain cereals; however, corn and corn products were not really among them. A possible explanation for this may be that corn plays a smaller role in human nutrition than wheat or rice. Little is known about the nutritional values of corns although many popular foods are made of them (e.g. tortilla), especially their element compositions. In this study, data on macro- and microelement contents of red and blue kernel varieties were shown. Presences of iron and selenium are notable in the investigated cultivars. Variability of element contents among genotypes sheds light on increasing these features during plant breeding. 6. Antioxidant activities in blue, purple and red kernel corns Nowadays an increased attention is paid on plants that have favourable effects on health due to their special material contents. Food industry has been changed radically during the last fifty years. Foods contain more and more artificial additives and synthetic components. As there is even more light shed on food safety and healthy diets, a lot of consumers try to desist from these products. To find alternative solutions is a great challenge for food industry. As many times in different fields of life, the key is in nature where the possible sources could be found for the substitution of synthetic materials. Natural pigment materials could be the alternatives for artificial colourings. Plants are rich in antocyanins and flavonoids. These materials play role not only in the colouration 100 Created by XMLmind XSL-FO Converter. 14. MAIZE BREEDING but have antioxidant activities; in addition they are neither toxic nor mutagenic so their consumption is risk-free. Blue and red kernel corns are also among these plants. American Indians have been utilized the pigments of the plant for textile staining for centuries. The number of races of blue corn is still unknown and it does not differ from the other varieties botanically. The Peruvian variety Morado has the deepest purple kernel colouration and this tone cannot be found in any other plants. Purple corn contains cell protecting antioxidants which materials proved to be effective against carcinogenic tumours in rats. A lot of plants have this kind of traits but only a small part of them takes anti-inflammatory effects. The nutritional values of blue corn exceed such phenomenon of the generally produced yellow corn in many ways. Its nutritional values colour and taste can contribute to the formation of a healthy diet. Foods made of blue corn are tastier than those made of white. The nutritional value of blue corn is also higher than that of yellow or white because the ratio of the germ is higher than the ratio of endosperm. The high flavonoid content of the blue corn has antioxidant activity in the foods made of it. People living in the Peruvian Andes have been using purple corns (maiz morado) for a long time as food and drink, while in the developed countries its popularity has just begun. The alcoholic drink made of the kernels is called chicha morada. Antocyanins are well-known alternatives of synthetic colouring materials. There are a lot of patents for the processing and usage of these colouring materials of corn. From the last few years more attention are paid on the studies of antioxidant contents and activities of pepper, onion, sour cherry, beetroot and cereals. Antioxidants can react with free radicals produced in the organisms and can decrease their adverse effects. Due to different enzymatic reactions and deleterious effects of the outer environment free radicals are produced in the human body; these compounds can enhance ageing and can cause diseases of the immune and vascular systems, cancer, etc. It was found the highest total activity in corn (181.42±0.83 µmol vitamin C equivalent/g), followed by wheat (76.70±1.38 µmol vitamin C equivalent/g), oat and rice (55.77±1.62 µmol vitamin C equivalent/g). There is still little knowledge on the antioxidant activities of different corn genotypes. 101 Created by XMLmind XSL-FO Converter. 15. fejezet - 15. SELECTION OF HIGH ENERGY FIBER PLANTS Miscanthus which is a temperate perennial cross-pollinating grass has a great potential for increasing the biomass production and using as a novel energy crop (Figure 55.). 15.1. ábra - Figure 55.: Miscanthus There have been great efforts for establishment and improving in vitro propagation system at Miscanthus. Different carbon sources in the media play an important role on the induction, maintenance and plant regeneration of different Miscanthus genotypes, derived from various explant types. For instance concluded that on the medium containing mannitol or sorbitol, only a small amount of callus were induced and no embryogenic callus was formed. Furthermore callus induction and embryogenic callus formation on shoot apices and immature inflorescenses did not differ significantly between media containing saccharose, glucose, fructose, maltose or a mixture of glucose and fructose. However, callus induction and embryogenic callus formation from leaf explants were best on glucose. A higher percentage of leaf explants formed callus on autoclaved saccharose, as opposed to the other carbon sources where filter-sterilization in general resulted in a higher callus percentage. The growth rate of embryogenic callus was influenced both by carbon source and sterilization method when less than 1g of callus was inoculated. None of the tested carbon sources could considerably improve plant regeneration from Miscanthus giganteus callus, but a higher number of plants tended to be regenerated per callus piece from filter-sterilized carbon sources. It was found stimulating affect of different proline concentrations on callus initiation, embryogenic callus formation, growth and regeneration in callus and suspension cultures of Miscanthus x ogiformis Honda Giganteus. In a further experiment investigating the growth characteristics and nutrient depletion of the same genotype mentioned above in a suspension culture- that saccharose was hydrolysed into its monosaccharide components in the culture medium. Glucose was depleted farter than fructose indicating a preference for glucose as a carbohydrate source of the M. x ogiformis cultures. Plant materials of two different Miscanthus x giganteus origin were obtained from field plants for in vitro shoot induction. During the whole culture duration (10th-60th days) the shoot growth of two genotypes was determined. The results showed in vitro shoot induction and development was dependent on genotypes. There has been a lot of work to study the effects of different cytokinins, i. e. benzyladenine, kinetin, isopentenyladenine or thidiazuron in Miscanthus. More research has to be done on the effects of different carbon sources in vitro Miscanthus cultures. Plant material of Miscanthus x giganteus and Miscanthus sinensis was obtained from field plantations. The plant material was surface sterilized with 80% alcohol. Axillary buds were placed on a shoot inducing nutrient solution. These medium was a modified MS basal medium (Murashige-Skoog, 1962) supplemented with 20gl-1 102 Created by XMLmind XSL-FO Converter. 15. SELECTION OF HIGH ENERGY FIBER PLANTS saccharose, glucose, fructose respectively and 0.3 mgl-1 6-Benzylaminopurin for the shoot induction. In the nutrient solution the pH was adjusted to 5.7 prior to autoclaving. Explants were incubated with a day length of 16h at 21 Co in glass culture dishes. During a particular period of time we measured the weight of shoots in 5., 10., 20., 40. days of Miscanthus x giganteus and Miscanthus sinensis originated from their axillary buds. By this method we wanted to know how the carbon sources effect the shoot formation of Miscanthus and the differences between the hybrid and genotypes of this species. In a modified MS basal medium supplemented with the same amount (20gl-1) of different carbon sources (saccharose, glucose, fructose) and the equal amount of BAP (0,3 mgl -1) we induced shoot formation from axillary buds. In a particular time (in the 5., 10., 20., 40. days after plating) we measured the shoot weight and developed further the induced shoots (Figure 56. and 57.). 15.2. ábra - Figure 56 .: Shoot weight development (g) during 40 days of Miscanthus x giganteus (M118) 15.3. ábra - Figure57.: Shoot weight development (g) during 40 days of Miscanthus sinensis (Goliath) Figure 56. and 57. show that in the Miscanthus x giganteus (M118) and Miscanthus sinensis "Goliath" the best carbon source proved to be the D+ saccharose. In our experiments we found that the disaccharide gave better results in different Miscanthus genotypes and can use more afficiently for optimising in vitro propagation system than the monosaccharides. In the Table 32. the increase in shoot weight of Miscanthus x giganteus (M118) in a certain period of times ( 5, 10, 20 days interval). Not respect to the carbon source of MS medium, the shoot development is very rapid. However the increase of shoot was the highest in medium containing D+ saccharose at the beginning of shoot induction. The shoot increase was accelerated the time being and very high in the case of fructose. However neither the shoot weight values of glucose medium nor the ones of fructose medium were higher than those of saccharose medium. 103 Created by XMLmind XSL-FO Converter. 15. SELECTION OF HIGH ENERGY FIBER PLANTS 15.1. táblázat - Table 32.: Shoot induction of Miscanthus x giganteus (M118) in different culture media Carbon source Difference in shoot weight, g 10-5 20-10 40-20 Days D+saccharose 0,385 1,700 2,618 glucose 0,388 0,878 2,395 fructose 0,410 0,600 3,320 We found the same trend in Miscanthus sinensis "Goliath". The only difference that the difference in shoot weight was the higher in the explants plated on medium containing D+ saccharose (Table 33.). In those experiments, which in the medium glucose and fructose were the carbon sources, not only the developments of in vitro plants were lower but the depletion of nutrients as well than in the case of saccharose. 15.2. táblázat - Table 33 .: Shoot induction of Miscanthus sinensis (Goliath) in different culture media Carbon source Difference in shoot weight, g 10-5 20-10 40-20 Days D+ saccharose 0,418 0,805 3,163 glucose 0,375 0,328 3,165 fructose 0,380 0,558 2,873 In Figure 58. we can see the relative increase in shoot weight in comparison of M 118 to Goliath. 15.4. ábra - Figure 58.: Relative increase in shoot weight (M118/Goliath) The development of plant in vitro culture conditions shoot showed a very strong dependence to genotypes. In our investigated genotypes the development of M 118 hybrid was better than in the case of M. sinensis 104 Created by XMLmind XSL-FO Converter. 15. SELECTION OF HIGH ENERGY FIBER PLANTS "Goliath". The growth of M 118 Hybrid was higher in the same in vitro conditions and its development was more efficient then that of variety "Goliath". Explants were incubated with a day length of 16 h at 21 °C in glass culture dishes. During the whole culture duration (10th.-60th day, every 10th days) the nutrient content of the culture medium (N, P, K, Ca, Mg) of 2 genotypes were determined by Kjeldahl-method, AAS and photometer. After 40 days of culturing the axillary buds three times more shoots could be harvested. The results showed that after 35 days nitrogen and phosphate in the medium was nearly completely taken up. That means from that time on no increase of shoot growth will happen. The nitrate content in the nutrient solution decreased steadily up to the 40th day but stagnated thereafter. This was the same for both genotypes (Figure 59.). 15.5. ábra - Figure 59.: Change of nitrate content in the nutrient solution in comparison with growth of shoot of two Miscanthus genotypes (G1, G3) during the 60 days culturing. The phosphate content in the medium of both genotypes has already decreased from the beginning of the culture (Figure 60.). 15.6. ábra - Figure 60.: Change of phosphate content in the nutrient solution in comparison with growth of shoot of two Miscanthus genotypes (G1, G3) during the 60 days culturing. Therefore, phosphate is obviously the limited element of the in-vitro propagation. During the first 20 days potassium, calcium and magnesium was hardly taken up by the plants. However after that time a slow decrease 105 Created by XMLmind XSL-FO Converter. 15. SELECTION OF HIGH ENERGY FIBER PLANTS of these nutrients could be observed. Up to the 60th day the plants had enough K, Ca and Mg for their development. After the shoot propagation they were transferred into a nutrient solution for root formation. After about 14 days the rooted shoots could be potted in soil. 106 Created by XMLmind XSL-FO Converter. 16. fejezet - 16. PLANT IDENTIFICATION FOR SELECTION 1. Genetic diversity Genetic diversity refers to the total number of genetic characteristics in the genetic makeup of a species. It is distinguished from genetic variability, which describes the tendency of genetic characteristics to vary. Genetic diversity serves as a way for populations to adapt to changing environments. With more variation, it is more likely that some individuals in a population will possess variations of alleles that are suited for the environment. Knowledge of genetic diversity among available parental lines is fundamental for successful hybrid maize breeding. Since the 1920s, the hybrid maize production in the world has been based on the development and crossing of inbred lines. Currently in maize, many new hybrids have been developed from crosses of a limited number of parent lines. This represents a great risk for the loss of genetic diversity in elite germplasm. For these reasons and others, maize breeders have a keen interest in the characterization of genetic diversity among parental lines. The genetic diversity among breeding materials could help to prevent the great risk of increasing uniformity in the germplasm and could also ensure long-term selection gains. Due to specific breeding aims and gene erosion, the genetic basis of maize breeding has decreased significantly in Hungary in recent decades, with well-known unfavourable effects. The determination of the most similar variety of groups or similar varieties can be carried out using variety descriptions obtained as the result of DUS testing. The success of plant breeding depends mainly on the genetic diversity of the basic material. Crossing and mutation are different methods which are applied to generate genetic diversity. In the interests of greater yield stability, the aim is for each maturity group in each growing area to be represented by hybrids of different genetic origins. When creating hybrids, lines with diverse genetic origin should be used in order to achieve a greater heterosis effect. Much research has been done concerning the amount of variation for AFLP markers in maize inbred lines, examined the usefulness of AFLP markers for assigning inbred lines to heterotic groups, compared the genetic similarity (GS) based on AFLP markers. AFLP molecular marker is efficient in assigning maize lines to heterotic groups and that AFLP-based genetic distance (GD) is suitable for predicting the maize single cross performance for intra population crosses of broad-based populations. AFLP markers were applied in maize to compare genetic distance and hybrid performance. For instance due to climatic barriers, the genetic variability of Sorghum varieties decreases away from the gene centre, therefore hybrids cultivated in Hungary are restricted to a relatively limited genetic basis. This limited genetic basis results in a lower rate of genetic improvement, so the genetic basis of cytoplasm factors are to be extended. Genetic diversity can be enhanced by fast neutron treatment. As a result of this treatment it was detected sterile, deformed specimens with double panicles in the stock treated with a dose of 12.5 Gy in the M1 generation (Figure 61., 62.) 16.1. ábra - Figure 61.: Low plant height, increased sprouting activity and double panicle in the case of ‘12 Gy Zádor’ 107 Created by XMLmind XSL-FO Converter. 16. PLANT IDENTIFICATION FOR SELECTION 16.2. ábra - Figure 62.: Low plant height and panicle sterility in ‘12 Gy Rib Agronomically useful specimens were found in the stock treated with 10 Gy and 7.5 Gy doses, where the leaf area index (LAI) and Harvest- index gave higher values than those of the control group. In the stock treated with 5 Gy dose, variations were less significant. In the reseeded M2 generation we detected albino specimens with extreme chlorophyll defects in the 10 Gy Zádor stock (Figure 63.). 16.3. ábra - Figure 63.: Shorghum albino specimens with extreme chlorophyll defects Irradiated plants ‘10 Gy Zádor’ 108 Created by XMLmind XSL-FO Converter. 16. PLANT IDENTIFICATION FOR SELECTION 2. Genetic similarity and hybrid performance in irradiated maize The aims of our study were to estimate genetic similarity (GS) and genetic distance (GD) in four maize inbreed lines; to classify the lines according to their GD and GS in 4x4 full diallel system. We used morphological description and AFLP and zein (IEF) analysis for estimation genetic polymorphism in four maize inbred lines. The dendogram based on GS and GD and morphological description separated four inbred lines into welldefined groups. Morphological description just with AFLP and zein analysis showed reliable results. We can conclude that the AFLP markers provide a large number of polymorphic bands. AFLP and zein analysis seem to be most efficient technique for comparing genetic similarities/differences among different genotypes. Maize inbred lines applied were originated from earlier mutation breeding program. Identification and the origin of these genotypes are given in Table 34. 16.1. táblázat - Table 34.: Origin and identification of the investigated lines Lines Origin Type of irradiation Dose [Gy] UDL1 F1*(NK-PX14)M2** fast neutron 15 UDL4 F1*(Pi3978SC)M3** fast neutron 5 UDL5 F1*(Pi3764MTC)M3** fast neutron 5 UDL6 F1*(Pi3478)M2** fast neutron 5 Genomic DNA of the leaves, and was isolated by QIAGEN Dneasy Plant Mini Kit.. AFLP analysis profiles were then performed. For AFLP analysis we applied 3 primer combinations which were the following: M48E32*; M49-E32*; M50-E32* (* by KeygeneR nomenclatura). Estimates of genetic similarity/distance among all genotypes were calculated according to Jaccard’s similarity coefficient. Cluster analysis based on the similarity matrix was carried out using SPSS program. 16.2. táblázat - Table 35.: A genetic background of the full diallel system 109 Created by XMLmind XSL-FO Converter. 16. PLANT IDENTIFICATION FOR SELECTION Parental lines UDL 1 UDL 4 UDL 5 UDL 6 UDL 1 UDL 1 UDH 1 UDH 2 UDH 3 UDL 4 UDH 4 UDL 4 UDH 5 UDH 6 UDL 5 UDH 7 UDH 8 UDL 5 UDH 9 UDL 6 UDH 10 UDH 11 UDH 12 UDL 6 The examination of the full diallel system (Table 35.) was carried out by the application of the further developed DIALLEL Analysis and Simulation programme according to Griffing 1 method. Morphological observations are containing during the whole vegetation period and ecological conditions effect these characters are to a large extent. Because of it found phenological characters not suitable to estimate genetic distance among inbred lines. The genetic similarity can be established with morphological parameters together with isoenzyme patterns. We got a dendrogram (Figure 64.) based on 28 morphological characters of CPVO-TP/2/2. Our result indicated that there are two clusters: UDL5/UDL6 and UDL1/UDL4 based on their genetic distance. 16.4. ábra - Figure 64.: Dendogram obtained from cluster analysis based on the morphological descriptions (DUS) of four maize inbred lines As a result of the zein analysis (Figure 65.), we found two groups according to genetic similarity of investigated lines. 16.5. ábra - Figure 65.: Dendogram obtained from cluster analysis based on the zein patterns of four maize inbred lines. We use cluster analysis and the dendrogram - obtained from it - based on the AFLP amplification patterns of four maize inbred lines which is shown in Figure 66. 16.6. ábra - Figure 66. Dendogram obtained from cluster analysis based on the AFLP amplification patterns of four maize inbred lines. 110 Created by XMLmind XSL-FO Converter. 16. PLANT IDENTIFICATION FOR SELECTION Based on dendrogram we can cluster two groups: I: UDL1 and II.: UDL4, UDL5, UDL6 which are different from cluster based only in morphological characters. These features are modified by ecological condition but the AFLP based genetic similarity (GS) are not sensitive to ecological factors and can be easily reproduced. The genetic distance (GD) among the four inbred lines based on Jaccard index was the following: UDL4UDL1:0,75; UDL6-UDL1:0,78; UDL5-UDL1:0,74; UDL6-UDL4:0,59; UDL5-UDL4:0,57; UDL5UDL6:0,51). 16.3. táblázat - Table 36.: Effect of general (GCA) and specific (SCA) combining ability to yield (kg/plot) in full diallel system Parental lines Specific combining ability (SCA) UDL 1 UDL 4 UDL 5 UDL 6 2004 2005 2004 2005 2004 2005 2004 2005 UDL 1 -5,4431 -13,368 2,3750 0,8050 -0,2550 -0,8975 0,8400 19,492 UDL 4 -0,8500 -1,5387 -1,8718 -0,9693 -1,7450 -0,2800 -0,7125 -0,1025 UDL 5 1,9681 -0,3593 0,2812 3,9575 -3,2456 -1,2906 0,5750 0,1325 UDL 6 2,6250 15,266 0,7406 -1,4493 0,9962 -2,3075 -4,3618 -11,509 GCA* (3) 0,9956 3,4431 0,8975 -3,0662 -0,4906 -2,7481 0,3925 2,3712 *SzD 5%= 0,017 (2004); 0,241 (2005); R2(2004)= 0,98; R2(2005)= 0,81 The highest general combining ability values for grain yield was found in the case of lines UDL 1 and UDL 6 (Table 36.). The highest specific combining ability (SCA) value for grain yield was found by UDH 3, 10 hybrids. The result of the variation analysis proves to be a significant correlation between the values of SCA and GCA and the value of yields in diallel system, respectively. As the value of the regression coefficient is always higher than 0,8, the correlation seems to be significant. We estimated genetic similarity/distance (GD) in four maize inbreed lines; classified the lines according to their GD and GS; determined hybrid performance based on GD and heterosis for yield ability in 4x4 full diallel system. We can summarize the following: • The application of mutation breeding method is of great help in widening the range of usable genotypes in maize breeding. • With the help of diallel analysis, the lines having the highest general and specific combining abilities (GCA, SCA) could be selected. • The dendogram based on genetic similarities (GS) and genetic distance (GD) and morphological description separated four inbred lines into well-defined groups. Morphological description just with AFLP and zein analysis showed reliable results. • The analysis of the results indicated that lines UDL 4, UDL 5 and UDL 6 were most closely related to other, while being genetically distant from line UDL 1. The highest GCA value for grain yield was found in the case of line UDL 1. On the basis of the specific combining ability values, two excellent hybrid combination were selected (UDH3, UDH10). 111 Created by XMLmind XSL-FO Converter. 16. PLANT IDENTIFICATION FOR SELECTION 112 Created by XMLmind XSL-FO Converter.