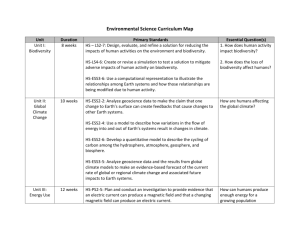
Models Ability to Predict Terrestrial Biodiversity
advertisement

Heather O’Keefe 905373807 Models Ability to Predict Terrestrial Biodiversity Currently, there are several models that are used to predict potential changes in terrestrial biota. Modeling is useful in predicting future responses in examples such as guiding conservation efforts, legislation, and even economies. “The science of predicting extinctions from global warming is only a few years old, and the best models are rife with uncertainties.”6 While models are based off of forcers such as climate change, land-use, and adaptation ability of species, the complex interactions between species and their environment make modeling difficult. Though modeling is difficult it is important because “ecosystems provide many goods and services that are crucial to human survival”7. Therefore, the reliability of these models to predict future changes in terrestrial biota is crucial. There are several different forms of models which take different approaches on predicting which then are based on several different forcers. Due to sheer variation of model types, this can create many different predictions on future outcomes on the same data set. One of the key forcers that drive many models is climate change. “Changes in species distribution linked to changes in climatic factors have been observed” 7 according to the IPCC. It is known that “climate change directly affects the functions of individual organisms (e.g., growth and behavior), modifies populations (e.g., size and age structure), and affects ecosystem structure and function (e.g., decomposition, nutrient cycling, water flows, and species composition and species interactions) and the distribution of ecosystems within landscapes; and indirectly through, for example, changes in ecosystem structure and function are assumed to be related to changes in various aspects of biodiversity” 7. This statement describes that climate change has several complex interactions between various forms of terrestrial biota. By understanding the impacts of climate change, it can consequently lead to a greater significance in the data of the model. Therefore, “models used to estimate global change impacts on biodiversity vary substantially in complexity and underlying hypotheses”6 which in turn creates different predictions on future responses of biodiversity. Models can be grouped into two different forms: generalized linear models and additive models. Generalized linear models “impose a theoretical response curve”1 and were proven to be “more easily transferable in space and time”1. While additive models “which produce data-driven response curves” have been proven to yield “more precise predictions in the regions where the models had been calibrated.”1 Yet in each case the models have been proven to have several flaws. One of these flaws is by imposing a general linear curve you assume that several parameters are held constant. Another flaw to be shown is that in additive model data it is generally only ‘reliable’ to the area in which the data was collected. Several different types of models for biodiversity include bioclimatic envelope, ecosystem, dynamic global vegetation (DGVMs), species-area, dose-response, and process-based models. Dose-response models are based on relationships from “experimental or observational data to estimate the impacts of drivers on biodiversity”6. An example would be the “effect of nitrogen deposition on mean species abundance”6. Process-based models “simulate processes such as population growth or mechanisms such as eco-physiological responses” 6. Dynamic global vegetation models (DGVMs) “are complex ecosystem models integrating processes such as photosynthesis, respiration, plant competition for resources, and biogeochemical cycles”6. DGVMs are popular in the sense that by having a larger source of vegetation available this then can support greater biodiversity and higher level organisms. Yet, this type of model is limiting due to the fact that it takes agriculture into account which has a result of creating a high rating for the model. But the flaw of agriculture is that it lacks the ability to support a high level of biodiversity which is a main positive point of being a DGVM. A different type of model is the bioclimatic envelope model also known as niche-based model which “use statistical relationships between current species distributions and environmental variables, such as temperature and precipitation, to project the future distribution of a species under climate change”6. Species-area models analyze the “empirical relationship between area and species number to estimate species committed to extinction after habitat loss”6. Hence, there are several different types of models which focus on different forcers in predicting potential future biodiversity trends. Even with the vast extent of different types of models, there are still various limitations for each model. One popular technique is to model climate change impacts on biodiversity because it is traceable. Yet there are several other forcers which differ depending on the types of model. “A wide range of climate scenarios and bioclimatic envelope modeling tools are readily available, but it is vital to develop models of other important drivers and their interactions. This will require the development of mechanistic models linking changes in land use, pollution levels, and biotic competition (e.g., invasive species) to population dynamics of individual species through changes in life-history parameters such as survival and dispersal, using techniques that are scalable across space and across species assemblages”6. Thus, the need for knowledge based on species complex reactions with its environment is still needed in order to create reliable models. Therefore, in order to create models with a greater source of credibility within the scientific community, "it is vital to develop modes of other important drivers and their interactions." 6 By not understanding these complex interactions it’s difficult to “assess how interactions between life history and disturbance regime mediate species extinction risk under climate change”6. Through previous indications that bioclimatic models have “insufficient treatment of key mechanisms, such as migration, biotic interactions, and interactions between drivers such as climate and land use”6 this then limits “the accuracy of future range projections”6. Therefore, each model cannot take into account all the interactions associated with biodiversity. Consequently no model in itself can be solely relied on to be an effective predictor of future changes. “Predictions of future distributions of species from bioclimatic models may fail because of uncertain predictions of local climate change, inaccurate estimates of the climatic tolerance of species, and unforeseen evolutionary changes in populations.”1 Therefore we as humans can never “predict the future with accuracy, but we need a strategy for using existing knowledge and bioclimatic modeling to improve understanding of the likely effects of future climate on biodiversity”1. Bioclimatic models are also known to “produce highly variable predictions of species-range shifts, and there is a poor correlation between a model’s ability to fit present and future distributions”1. During one experiment there was a result of nine different “bioclimatic models to predict the distributions of four South African plants under current and future climates. Predicted distribution changes varied from 92% loss to 322% gain for one species; similar variability was recorded for the other species”1. Thus there is not only variability based on the forcers chosen for the model, there is also the variability between the data collected between similar models. With this high variability between models of the same species, this makes forecasting trends difficult and also the ability to pass legislation nearly impossible. Even with limitations, some general characteristics of models can enable it to be more reliable in terms of measuring biodiversity. The use of “larger, more detailed data sets and morerefined models than previously available” often avoids “the problems often encountered in trying to scale up results from small local-scale studies”4. Thus, by having more details on a larger set of data it allows for ‘better’ averages which can be applicable in more than one region. As opposed to trying to use a small set of data which is then only ‘reliable’ in the region in which it was collected. This could be due to localization of the terrestrial biota to that region, or even the ability for a population to shift their natural ranges. It has also been shown that the “models with the best performance were the most recent and complex ones and fell into two groups: machinelearning programs that seek to obtain a stable selection of predictors from a larger range of alternatives, and community models that simultaneously analyze all species in relation to environmental parameters and then calibrate model coefficients for individual species”1. Therefore models which take into account what drives a species (predictors) that are both unique to that specie and have a stable relationship, have been shown to be effective. Using more complex systems with a larger set of variables and even individual variables depending on the species creates more accuracy then generalizations because it is more specific. While these models are better predictors due to the higher accuracy, the need for accurate models is crucial. There are many unknowns associated with models which are simply predictions. Even having all the information and greater specificity in the data, which in turn can create a more reliable prediction, but it does not necessarily create a reliable model. Bioclimatic models analyze all the current areas in which species have been, and look for areas which are similar to those areas previously recorded. This type of model is known to have high variation in results. One prediction mentioned earlier had results from a “92% range reduction to a 322% explansion”4. Other highly contrasting predictions have been obtained through other experiments such as this example where tropical biomes and “the physiological effects of elevated atmospheric CO2 concentrations and temperature on trees”4. Such studies indicated that an increase in “atmospheric CO2 affects photosynthesis rates and enhances net primary productivity—more so in tropical than in temperate regions—yet previous climatevegetation simulations did not take this into account”4. Therefore, between old and current models there is difference because environmental factors such as climate change were not accounted for. Yet it has been proven that “averaging the results of many climate-envelope models provides a more accurate prediction of where species can be found than any one model”6. In result with an increased amount of data the ability to create viable data increases. Yet “some experts have criticized the IPCC for implying that climate-envelope models are more precise than they actually are. “Simply presenting those numbers as factual, saying this is how many species will go extinct, is misleading,” says Richard Pearson, a postdoctoral researcher at the American Museum of Natural History in New York City.” By having professionals criticize the results, the reliability of bioclimatic models have begun to be controversial in the scientific community. This suggests that the ability of bioclimatic models to be used as projections of the future is unreliable. Some scientists have suggested that “to improve the performance of these models… researchers (should) include biological details about species, such as how quickly they disperse and how they interact with other species”6. Therefore while bioclimatic models are very popular and by using several different models together creates a ‘reliable’ prediction, there is still a vast amount of variability and skepticism associated with bioclimatic models. Ecosystem models are another form of model which looks at an ecosystem and how there may be potential shifts in that type of ecosystem. Thus this shifting of a “biome (i.e., the collection of ecosystems within a particular climatic zone with similar structure but differing species)”7 are generally “not well suited to projecting changes in regional biodiversity”7. This is because the ability to assume that “ecosystems or biomes will simply shift location while retaining their current composition, function, and structure”7 is naive. There are many complex relationships associated with each ecosystem, and the ability for one ecosystem to essentially shift all those current relationships to a new location is nearly impossible. The ability for many of the species to ‘range shift’ would be impossible. Just picking up one ecosystem and shifting it to possibly another continent would potentially change species migration patterns, altering the food web, and would take a long time because of the necessary type of vegetation could potentially grow less effectively in that new territory. This very idea of an “ecosystem movement approach assumes that ecosystems will migrate relatively intact to new locations that are close analogs to their current climate and environment.”7 This in itself is such a gross simplification because of the many complex interactions it does not address. Such interaction include “different climatic tolerance of the species involved, including within-species genetic variability, different longevities, different migration abilities, and the effects of invading species.”7 Therefore ecosystem models are not very effective in terms of modeling potential changes in biodiversity. Instead the ecosystem model “has the advantage that the well-demonstrated relationship between ecosystem range and existing climate can be used to project new ecosystem distributions under changed climate scenarios. As such, these models are useful for screening scenarios of climate change for potential significant effects.”7 Hence, models can be used to screen for other possible effects based on different factors. The ecosystem model may be a bad predictor for changes in biodiversity of that ecosystem but it could be useful in identifying individual species that may be in high risk of extinction in the future. Another useful type of modeling is hind-casting. Hind-casting consists of using historical data in models to predict future trends. Such models “are calibrated with current species-climate relationships and are then tested with reconstructed species distributions from the fossil record. This approach has been used to test whether climatic requirements of species remain stable over time”1.This type of model is useful because it has shown events that have already happened and therefore have a greater chance of happening again in the future. Yet, since hind-casting models do use historical data this could also be a disadvantage because “of the lack of agreement with extinction patterns in the recent and distant past”6. Another disadvantage is that by using historical data, models are “only feasible for a few species and regions for which a good fossil record is available”1. One hind-casting model analyzed the northern limit of boreal forests. These forests are “projected to move further north into the arctic tundra, whereas the southern limit will experience dieback, giving way to temperate conifer and mixed forest”6. These “projections are in qualitative agreement with the paleontological record, which indicates that climate change has resulted in large shifts in the distribution of vegetation types in the past”6. Thus, not only do hind-casting models predict future trends of a species based on what the species has done in the past, but it can analyze the responsiveness of the species to climate change. These reactions could be useful due to the fact that they could then be used to help model other similar species and their potential reactions in the future. Some of the keys aspects that many terrestrial biodiversity models take into account are a species’ adaptive capacity. Range shift is the ability of species to shift geographically to other areas that are similar to their current habitats. Range shifts are currently correlated with extinction rates, some examples being “migration rates [e.g., the highest projected extinction rates are 38% with unlimited migrations rates versus 58% with no migration in] and habitat specificity [e.g., the highest extinction rates are 7% for broad habitat specificity versus 43% for narrow habitat specificity]”6. Extinction rates of species are directly correlated with a species adaptive capacity. The less a species is able to adapt, the greater the probability of that species’ extinction. Extinction is currently associated with a species vulnerability (which includes exposure and sensitivity). This means that “we should expect to see species turnover, migrations, and novel communities”4. Land use also influences a species adaptive capacity. Without having anywhere to live, there is no hope for the species to survive, thus the analysis of fragmentation is currently a hot topic. Currently, “over 75% of the Earth’s terrestrial biomes now show evidence of alteration as a result of human residence and land use”4.Thus there is a great stress on studies that have focused on land fragmentation and climate change and their combined ability to effect species adaptive capacity. “First, the degree of land use and climate change explains a substantial fraction of the range of projected extinctions within studies [e.g., projected vertebrate extinctions are 11 to 34% for 0.8° to 1.7°C global warming versus 33 to 58% for >2.0°C warming in (7)], indicating that limitation of land-use change, especially in tropical and subtropical regions, and aggressive climate mitigation could substantially reduce extinction risks.”6 This shows that the linkage between the intensity of climate change and land use or fragmentation has been shown to have an impact on biodiversity. A solution for conserving biodiversity may lie with increased legislation around climate mitigation. Therefore the ability to control human impact on climate change through climate mitigation has the potential to decrease predicted extinction rates for terrestrial biodiversity models. Overall models themselves can only be predictors of what could be in terms of terrestrial biodiversity in the future. The ability of current models consequently is that they may be good predictors of determining future distributions and changes in biodiversity but are limited in many aspects. The relevance of data and how the model takes into account environmental stressors which are relevant to that particular species are some aspects that need to be taken into consideration. In other words, being able to model a species without information based off that species’ complex interactions within its habitat, and its role in its ecosystem with other species would not necessarily create a realistic prediction. Therefore the ability for models to identify complex interaction between ecosystems and even global change factors can skew data. This creates having ‘coarser’ models which will not take into account topography (also known as ‘microclimate buffering’) which may be necessary for some species survival. Hence the “complexity that we are faced with in trying to model and predict the possible consequences of future climate change”4 leaves many models unreliable. Is into a model creates an increased source of variability. Even taking a different model with similar data has been known to create different predictions. The lack of information that is needed in several different systems which models can be based off of also limits our ability to model as scientists and consequently creates a need for improved monitoring. Models are strewn with many inaccuracies yet still play an important role in today’s culture. Models are important for legislation, the creation of conservation, and potentially preventive strategies. For conservation, models can potentially predict species that will be at high risk for extinction. Therefore through the use of models, prevention strategies can be put in place to help conserve biodiversity and give that specie a greater chance to survive. We also must note that “the combination of climate change and habitat destruction, novel ecosystems are going to become increasingly common. Their conservation will require a whole new definition of what is ‘natural’. Since models give a rough idea of the potential impact of global warming, they may also offer guidance for planning reserves. But some scientists are concerned that policymakers will be expecting them to provide more precise estimates than they can deliver. Yet there is a need to create legislation on where to place conservations in order to limit human influence on the environment through fewer potential extinctions and a ‘healthier’ biosphere. This in turn could also be important for economical reasons. With more resources available, there could be a greater potential for greater gross domestic product (GDP) which in turn fuels the economy. With fewer available resources this in turn limits the ability for an increase in GDP which has the potential to lower the ability for economies to raise the standard of living. Therefore, models are important for guidance in preservation of biodiversity which affects economies. The economies in turn affect the overall quality of life which includes important topics such as literacy rates and life expectancy. Therefore models are of great importance to human society even with their deficiencies. Overall models are important in human society because they can enable us to create legislation for everyone’s betterment. Therefore while there are many inconsistencies associated with the science of modeling, the potential prediction can help guide decisions concerning conservation and preservation. Without such modeling efforts, there would be no data or guidance for legislators to have to provide reasons for conservation. This in turn would lead to no conservation for biota and most likely a decrease in biodiversity. This decrease in biodiversity would create numerous negative consequence including not only impact the species which may become extinct but humans ability to grow economically as well. With all of the impacts of these models the need for reliable models is therefore crucial. Thus in order for modeling to contain any substance the model will need to take into account complex interactions of the species within its environment which is being modeled. The model will also need to be large in order to have the greatest consistency and therefore application on a global scale. Therefore modeling may be an imprecise science, but its usefulness in creating future scenarios which can be used to prevent potential extinctions. Bibliography 1 Araujo, M. B. "ECOLOGY: How Does Climate Change Affect Biodiversity?" Science. (2006): xxxxx313.1396-397. 2 Dawson, Terence. "Beyond Predictions: Biodiversity Conservation in a Changing Climate." xxxxxScience 1 Apr. 2011: 53-58. 3 Pereira, Henrique M. "Scenarios for Global Biodiversity in the 21st Century." xxxxxScience 10 Dec. 2010: 1496-501. 4 Willis, Kathy. "Biodiversity and Climate Change." Science 326 (2009): 806-07. 5 Worm et all. "Impacts of Biodiversity Loss on Ocean Ecosystem Services." Science 13 Nov. xxxxx2006: 787-90. . 6 Zimmer, Carl. "Predicting Oblivion: Are Existing Models Up to the Task." Science 17 Aug. 2007: xxxxx892-93. 7 Climate Change and Biodiversity. Tech. no. 5. IPCC, 2002. Print.