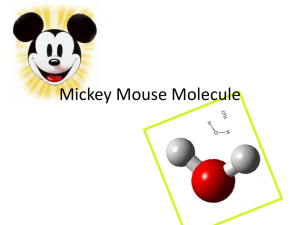
Collection on atomic electron configurations and orbitals
advertisement

AST 376/381 Scalo: Planets and Life Readings on Biochemical Bonding This document contains three exerpts having to do with chemical bonding, with an emphasis on simple biomolecules, taken from cell biology or biochemistry texts. The three selections overlap, but with different-enough presentations that you should read all three, at least up to the “optional” stop points that are marked. The fonts are different among the three selections, to help you tell them apart if you are reading on a computer monitor (that is why the font size is large), or if your hard copy blows away and randomizes. If you decide to print them, you should probably reduce the 16 or 14 pt fonts to 12 pt. Besides acquainting you with the most important chemical bond mechanisms, the reading should also be used to gain a familiarity with biology background that you are probably lacking (I’m assuming the average student has had no more biology than a distant memory in high school, although I realize there are one or two exceptions; these readings should still be accessible to upper division students--later in the semester I will ask you to look at (but not read!) some more “serious” papers and parts of books. Remember--most of any problem you have with this reading is only a terminology problem: There is almost no theoretical underpinning to anything that is described; biology is primarily a phenomenological (nice way to say “not much quantitative theory”) science, although that is rapidly changing as more and more physicists (and even a few researchers in planetary sciences and astrobiology) try to take over biophysics and make it boring and so technical that no one can understand it except themselves. Just (~ halfway) kidding. Start with the general survey of bonding, including water, from the greatest of cell biology books, Alberts, Molecular Biology of the Cell. This presentation also will give you a bit of an introduction to some of the biomolecules we will be discussing. This exerpt is followed by a little more advanced reading, sec. 1.3 from Berg et al., Biochemistry. Notice the emphasis put on the weak noncovalent bonds--keep a list going. Finally, a section covering mostly the same material, from a book by Lodish et al., Biochemistry. Depending on your background, you will probably encounter unfamiliar terms. When you do, take advantage of Wikipedia or some other internet resource to gradually close up terminological holes in your background. Be careful! Note: If you want to link to the full text of this book, go to the free NCBI/NIH bookshelf at: http://www.ncbi.nlm.nih.gov/entrez/query.fcgi?db=Books The problem is that you cannot view the figures at the same time as the text, or easily navigate the text. Here is the best procedure I have found. Click on the book of your choice (see below). Then click on the table of contents to see the full contents. Type the name of the chapter or subsection whose text or images (will be listed to the left) you want in the box to the upper right corner. You can also search by key word--try typing DNA and stand back! These are not the most recent editions, but trust me, I have compared with many other sources in these fields, and these are all excellent and do not suffer from their publication date except in a few places if you were a specialist. These books are meant for beginning biology majors (for example, they are a step less detailed than biochemistry textbooks), so later chapters are almost certainly too detailed to be of interest to you. I have already made Word versions of the first few chapters of most of them (and many more), besides stealing some of their great illustrations for lecture use, so my advice is to just read what I assign, beginning with this chapter. Other books you can access here, some of which I have already copied into Word documents like this one (and will put at the course web site or print and pass out in class when the time arrives), are: Berg, J. et al. Biochemistry, 2002, 5e (Sec. 1.3 is the third reading here) Cooper, G. M. The Cell: A molecular approach 2000, 2e. Great book--I will have you read a section from this book on biomacromolecules after you have had a chance to digest the current reading). Alberts, B. et al. Molecular Biology of the Cell, 4e (First reading here is from ch.2) Lodish, H. et al., Molecular Cell Biology--1999 (Part of chapter 2 copied in here) Brown, T. A. Genomes, 2e Griffiths, A. et al. Modern Genetic Analysis 1999. An important way in which biology textbooks are, out of necessity, superior to physics and astronomy books, is to have great graphics and refined organization and writing style, in order to compensate for having no explanations for the subject matter. You will find, as the deceased baseball player Yogi Berra said “You can learn a lot by just looking,” so take advantage of the pictures--I have copied all of the ones that go with the present reading into this document, and there are links to a few tables of “panels” that I couldn’t include easily. You may also find that a trip to half-price books will score you a $10-$15 copy of one of these $100-$200 books in pretty good condition. That is how I started. Besides the books below, I will be providing links to free access textbooks and the best tutorials in all the areas we discuss. One way you can become more comfortable with a new field of knowledge is to get a pile of books on the subject and just look through it to see how it is organized. Today we can do that on the internet, except that there is a huge danger of wasting much time with sites that are low on content, something like the material you see at most NASA or university or even course websites, meant for the public (except more press releases at NASA), or high-school teaching aide, not as an upper-level teaching tool. I have already weeded through 100s of these, but feel free to try if you want. I concluded that I would know much more had I settled on a high-level sight and then actually concentrated on the material instead of spending so much time collecting materials fo this class, especially the 1000s of images I accumulated. So try to leave the searching to me unless there is some special topic that you are interested in that we are not covering. Now on to biochemical bonding... From B. Alberts et al., Molecular Biology of the Cell (online at NIH bookshelf) The Outermost Electrons Determine How Atoms Interact To understand how atoms bond together to form the molecules that make up living organisms, we have to pay special attention to their electrons. Protons and neutrons are welded tightly to one another in the nucleus and change partners only under extreme conditions—during radioactive decay, for example, or in the interior of the sun or of a nuclear reactor. In living tissues, it is only the electrons of an atom that undergo rearrangements. They form the exterior of an atom and specify the rules of chemistry by which atoms combine to form molecules. Electrons are in continuous motion around the nucleus (really? [JMS] , but motions on this submicroscopic scale obey different laws from those we are familiar with in everyday life. These laws dictate that electrons in an atom can exist only in certain discrete states, called orbitals, and that there is a strict limit to the number of electrons that can be accommodated in an orbital of a given type—a so-called electron shell. The electrons closest on average to the positive nucleus are attracted most strongly to it and occupy the innermost, most tightly bound shell. This shell can hold a maximum of two electrons. The second shell is farther away from the nucleus, and its electrons are less tightly bound. This second shell can hold up to eight electrons. The third shell contains electrons that are even less tightly bound; it can also hold up to eight electrons. The fourth and fifth shells can hold 18 electrons each. Atoms with more than four shells are very rare in biological molecules. The electron arrangement of an atom is most stable when all the electrons are in the most tightly bound states that are possible for them—that is, when they occupy the innermost shells. Therefore, with certain exceptions in the larger atoms, the electrons of an atom fill the orbitals in order—the first shell before the second, the second before the third, and so on. An atom whose outermost shell is entirely filled with electrons is especially stable and therefore chemically unreactive. Examples are helium with 2 electrons, neon with 2 + 8, and argon with 2 + 8 + 8; these are all inert gases. Hydrogen, by contrast, with only one electron and therefore only a half-filled shell, is highly reactive. Likewise, the other atoms found in living tissues all have incomplete outer electron shells and are therefore able to donate, accept, or share electrons with each other to form both molecules and ions (Figure 2-4). QuickTime™ and a TIFF (Uncompressed) decompressor are needed to see this picture. Figure 2-4. Filled and unfilled electron shells in some common elements. All the elements commonly found in living organisms have unfilled outermost shells (red) and can thus participate in chemical reactions with other atoms. For comparison, some elements that have only filled shells (yellow) are shown; these are chemically unreactive. Because an unfilled electron shell is less stable than a filled one, atoms with incomplete outer shells have a strong tendency to interact with other atoms in a way that causes them to either gain or lose enough electrons to achieve a completed outermost shell. This electron exchange can be achieved either by transferring electrons from one atom to another or by sharing electrons between two atoms. These two strategies generate two types of chemical bonds between atoms: an ionic bond is formed when electrons are donated by one atom to another, whereas a covalent bond is formed when two atoms share a pair of electrons (Figure 2-5). QuickTime™ and a TIFF (Uncompressed) decompressor are needed to see this picture. Figure 2-5. Comparison of covalent and ionic bonds. Atoms can attain a more stable arrangement of electrons in their outermost shell by interacting with one another. An ionic bond is formed when electrons are transferred from one atom to the other. A covalent bond is formed when electrons are shared between atoms. The two cases shown represent extremes; often, covalent bonds form with a partial transfer (unequal sharing of electrons), resulting in a polar covalent bond (see Figure 2-43). Often, the pair of electrons is shared unequally, with a partial transfer between the atoms; this intermediate strategy results in a polar covalent bond, as we shall discuss later. An H atom, which needs only one more electron to fill its shell, generally acquires it by electron sharing, forming one covalent bond with another atom; in many cases this bond is polar. The other most common elements in living cells—C, N, and O, with an incomplete second shell, and P and S, with an incomplete third shell (see Figure 2-4)—generally share electrons and achieve a filled outer shell of eight electrons by forming several covalent bonds. The number of electrons that an atom must acquire or lose (either by sharing or by transfer) to attain a filled outer shell is known as its valence. The crucial role of the outer electron shell in determining the chemical properties of an element means that, when the elements are listed in order of their atomic number, there is a periodic recurrence of elements with similar properties: an element with, say, an incomplete second shell containing one electron will behave in much the same way as an element that has filled its second shell and has an incomplete third shell containing one electron. The metals, for example, have incomplete outer shells with just one or a few electrons, whereas, as we have just seen, the inert gases have full outer shells. Ionic Bonds Form by the Gain and Loss of Electrons Ionic bonds are most likely to be formed by atoms that have just one or two electrons in addition to a filled outer shell or are just one or two electrons short of acquiring a filled outer shell. They can often attain a completely filled outer electron shell more easily by transferring electrons to or from another atom than by sharing electrons. For example, from Figure 2-4 we see that a sodium (Na) atom, with atomic number 11, can strip itself down to a filled shell by giving up the single electron external to its second shell. By contrast, a chlorine (Cl) atom, with atomic number 17, can complete its outer shell by gaining just one electron. Consequently, if a Na atom encounters a Cl atom, an electron can jump from the Na to the Cl, leaving both atoms with filled outer shells. The offspring of this marriage between sodium, a soft and intensely reactive metal, and chlorine, a toxic green gas, is table salt (NaCl). When an electron jumps from Na to Cl, both atoms become electrically charged ions. The Na atom that lost an electron now has one less electron than it has protons in its nucleus; it therefore has a single positive charge (Na+). The Cl atom that gained an electron now has one more electron than it has protons and has a single negative charge (Cl-). Positive ions are called cations, and negative ions, anions. Ions can be further classified according to how many electrons are lost or gained. Thus sodium and potassium (K) have one electron to lose and form cations with a single positive charge (Na+ and K+), whereas magnesium and calcium have two electrons to lose and form cations with two positive charges (Mg2+ and Ca2+). Because of their opposite charges, Na+ and Cl- are attracted to each other and are thereby held together in an ionic bond. A salt crystal contains astronomical numbers of Na+ and Cl- (about 2 × 1019 ions of each type in a crystal 1 mm across) packed together in a precise three-dimensional array with their opposite charges exactly balanced (Figure 2-6). QuickTime™ and a TIFF (Uncompressed) decompressor are needed to see this picture. Figure 2-6. Sodium chloride: an example of ionic bond formation. (A) An atom of sodium (Na) reacts with an atom of chlorine (Cl). Electrons of each atom are shown schematically in their different energy levels; electrons in the chemically reactive (incompletely filled) shells are red. The reaction takes place with transfer of a single electron from sodium to chlorine, forming two electrically charged atoms, or ions, each with complete sets of electrons in their outermost levels. The two ions with opposite charge are held together by electrostatic attraction. (B) The product of the reaction between sodium and chlorine, crystalline sodium chloride, consists of sodium and chloride ions packed closely together in a regular array in which the charges are exactly balanced. (C) Color photograph of crystals of sodium chloride. Substances such as NaCl, which are held together solely by ionic bonds, are generally called salts rather than molecules. Ionic bonds are just one of several types of noncovalent bonds that can exist between atoms, and we shall meet other examples. Because of a favorable interaction between water molecules and ions, ionic bonds are greatly weakened by water; thus many salts (including NaCl) are highly soluble in water—dissociating into individual ions (such as Na+ and Cl-), each surrounded by a group of water molecules. In contrast, covalent bond strengths are not affected in this way. Covalent Bonds Form by the Sharing of Electrons All the characteristics of a cell depend on the molecules it contains. A molecule is defined as a cluster of atoms held together by covalent bonds; here electrons are shared between atoms to complete the outer shells, rather than being transferred between them. In the simplest possible molecule—a molecule of hydrogen (H2)—two H atoms, each with a single electron, share two electrons, which is the number required to fill the first shell. These shared electrons form a cloud of negative charge that is densest between the two positively charged nuclei and helps to hold them together, in opposition to the mutual repulsion between like charges that would otherwise force them apart. The attractive and repulsive forces are in balance when the nuclei are separated by a characteristic distance, called the bond length. A further crucial property of any bond—covalent or noncovalent— is its strength. Bond strength is measured by the amount of energy that must be supplied to break that bond. This is often expressed in units of kilocalories per mole (kcal/mole), where a kilocalorie is the amount of energy needed to raise the temperature of one liter of water by one degree centigrade. Thus if 1 kilocalorie must be supplied to break 6 × 1023 bonds of a specific type (that is, 1 mole of these bonds), then the strength of that bond is 1 kcal/mole. An equivalent, widely used measure of energy is the kilojoule, which is equal to 0.239 kilocalories. To get an idea of what bond strengths mean, it is helpful to compare them with the average energies of the impacts that molecules are constantly undergoing from collisions with other molecules in their environment (their thermal, or heat, energy), as well as with other sources of biological energy such as light and glucose oxidation (Figure 2-7). QuickTime™ and a TIFF (Uncompressed) decompressor are needed to see this picture. Figure 2-7. Some energies important for cells. Note that these energies are compared on a logarithmic scale. Typical covalent bonds are stronger than the thermal energies by a factor of 100, so they are resistant to being pulled apart by thermal motions and are normally broken only during specific chemical reactions with other atoms and molecules. The making and breaking of covalent bonds are violent events, and in living cells they are carefully controlled by highly specific catalysts, called enzymes. Noncovalent bonds as a rule are much weaker; we shall see later that they are important in the cell in the many situations where molecules have to associate and dissociate readily to carry out their functions. Whereas an H atom can form only a single covalent bond, the other common atoms that form covalent bonds in cells—O, N, S, and P, as well as the all-important C atom—can form more than one. The outermost shell of these atoms, as we have seen, can accommodate up to eight electrons, and they form covalent bonds with as many other atoms as necessary to reach this number. Oxygen, with six electrons in its outer shell, is most stable when it acquires an extra two electrons by sharing with other atoms and therefore forms up to two covalent bonds. Nitrogen, with five outer electrons, forms a maximum of three covalent bonds, while carbon, with four outer electrons, forms up to four covalent bonds—thus sharing four pairs of electrons (see Figure 2-4). When one atom forms covalent bonds with several others, these multiple bonds have definite orientations in space relative to one another, reflecting the orientations of the orbits of the shared electrons. Covalent bonds between multiple atoms are therefore characterized by specific bond angles as well as bond lengths and bond energies (Figure 2-8). QuickTime™ and a TIFF (Uncompressed) decompressor are needed to see this picture. Figure 2-8. The geometry of covalent bonds. (A) The spatial arrangement of the covalent bonds that can be formed by oxygen, nitrogen, and carbon. (B) Molecules formed from these atoms have a precise three-dimensional structure, as shown here by ball and stick models for water and propane. A structure can be specified by the bond angles and bond lengths for each covalent linkage. The four covalent bonds that can form around a carbon atom, for example, are arranged as if pointing to the four corners of a regular tetrahedron. The precise orientation of covalent bonds forms the basis for the three-dimensional geometry of organic molecules. There Are Different Types of Covalent Bonds Most covalent bonds involve the sharing of two electrons, one donated by each participating atom; these are called single bonds. Some covalent bonds, however, involve the sharing of more than one pair of electrons. Four electrons can be shared, for example, two coming from each participating atom; such a bond is called a double bond. Double bonds are shorter and stronger than single bonds and have a characteristic effect on the three-dimensional geometry of molecules containing them. A single covalent bond between two atoms generally allows the rotation of one part of a molecule relative to the other around the bond axis. A double bond prevents such rotation, producing a more rigid and less flexible arrangement of atoms (Figure 2-9 and Panel 2-1, pp. 111–112). QuickTime™ and a TIFF (Uncomp resse d) de com press or are nee ded to s ee this picture. Figure 2-9. Carbon-carbon double bonds and single bonds compared. (A) The ethane molecule, with a single covalent bond between the two carbon atoms, illustrates the tetrahedral arrangement of single covalent bonds formed by carbon. One of the CH3 groups joined by the covalent bond can rotate relative to the other around the bond axis. (B) The double bond between the two carbon atoms in a molecule of ethene (ethylene) alters the bond geometry of the carbon atoms and brings all the atoms into the same plane (blue); the double bond prevents the rotation of one CH2 group relative to the other. Some molecules share electrons between three or more atoms, producing bonds that have a hybrid character intermediate between single and double bonds. The highly stable benzene molecule, for example, comprises a ring of six carbon atoms in which the bonding electrons are evenly distributed (although usually depicted as an alternating sequence of single and double bonds, as shown in Panel 2-1). When the atoms joined by a single covalent bond belong to different elements, the two atoms usually attract the shared electrons to different degrees. Compared with a C atom, for example, O and N atoms attract electrons relatively strongly, whereas an H atom attracts electrons more weakly. By definition, a polar structure (in the electrical sense) is one with positive charge concentrated toward one end (the positive pole) and negative charge concentrated toward the other (the negative pole). Covalent bonds in which the electrons are shared unequally in this way are therefore known as polar covalent bonds (Figure 2-10). Quick Time™a nd a TIFF ( Unco mpre ssed ) dec ompr esso r ar e nee ded to see this pictur e. Figure 2-10. Polar and nonpolar covalent bonds. The electron distributions in the polar water molecule (H2O) and the nonpolar oxygen molecule (O2) are compared (δ+, partial positive charge; δ-, partial negative charge). For example, the covalent bond between oxygen and hydrogen, O-H, or between nitrogen and hydrogen, -N-H, is polar, whereas that between carbon and hydrogen, -C-H, has the electrons attracted much more equally by both atoms and is relatively nonpolar. Polar covalent bonds are extremely important in biology because they create permanent dipoles that allow molecules to interact through electrical forces. Any large molecule with many polar groups will have a pattern of partial positive and negative charges on its surface. When such a molecule encounters a second molecule with a complementary set of charges, the two molecules will be attracted to each other by permanent dipole interactions that resemble (but are weaker than) the ionic bonds discussed previously for NaCl. An Atom Often Behaves as if It Has a Fixed Radius When a covalent bond forms between two atoms, the sharing of electrons brings the nuclei of these atoms unusually close together. But most of the atoms that are rapidly jostling each other in cells are located in separate molecules. What happens when two such atoms touch? For simplicity and clarity, atoms and molecules are usually represented in a highly schematic way—either as a line drawing of the structural formula or as a ball and stick model. However, a more accurate representation can be obtained through the use of so-called space-filling models. Here a solid envelope is used to represent the radius of the electron cloud at which strong repulsive forces prevent a closer approach of any second, non-bonded atom—the so-called van der Waals radius for an atom. This is possible because the amount of repulsion increases very steeply as two such atoms approach each other closely. At slightly greater distances, any two atoms will experience a weak attractive force, known as a van der Waals attraction. As a result, there is a distance at which repulsive and attractive forces precisely balance to produce an energy minimum in each atom's interaction with an atom of a second, non-bonded element (Figure 2-11). QuickTime™ and a TIFF (Uncompressed) decompressor are needed to see this picture. Figure 2-11. The balance of van der Waals forces between two atoms. As the nuclei of two atoms approach each other, they initially show a weak bonding interaction due to their fluctuating electric charges. However, the same atoms will strongly repel each other if they are brought too close together. The balance of these van der Waals attractive and repulsive forces occurs at the indicated energy minimum. Depending on the intended purpose, we shall represent small molecules either as line drawings, ball and stick models, or space filling models throughout this book. For comparison, the water molecule is represented in all three ways in Figure 2-12. QuickTime™ and a TIFF (Uncompressed) decompressor are needed to see this picture. Figure 2-12. Three representations of a water molecule. (A) The usual line drawing of the structural formula, in which each atom is indicated by its standard symbol, and each line represents a covalent bond joining two atoms. (B) A ball and stick model, in which atoms are represented by spheres of arbitrary diameter, connected by sticks representing covalent bonds. Unlike (A), bond angles are accurately represented in this type of model (see also Figure 2-8). (C) A space-filling model, in which both bond geometry and van der Waals radii are accurately represented. When dealing with very large molecules, such as proteins, we shall often need to further simplify the representation used (see, for example, Panel 3-2, pp. 138–139). Water Is the Most Abundant Substance in Cells Water accounts for about 70% of a cell's weight, and most intracellular reactions occur in an aqueous environment. Life on Earth began in the ocean, and the conditions in that primeval environment put a permanent stamp on the chemistry of living things. Life therefore hinges on the properties of water. In each water molecule (H2O) the two H atoms are linked to the O atom by covalent bonds (see Figure 2-12). The two bonds are highly polar because the O is strongly attractive for electrons, whereas the H is only weakly attractive. Consequently, there is an unequal distribution of electrons in a water molecule, with a preponderance of positive charge on the two H atoms and of negative charge on the O (see Figure 2-10). When a positively charged region of one water molecule (that is, one of its H atoms) comes close to a negatively charged region (that is, the O) of a second water molecule, the electrical attraction between them can result in a weak bond called a hydrogen bond. These bonds are much weaker than covalent bonds and are easily broken by the random thermal motions due to the heat energy of the molecules, so each bond lasts only an exceedingly short time. But the combined effect of many weak bonds is far from trivial. Each water molecule can form hydrogen bonds through its two H atoms to two other water molecules, producing a network in which hydrogen bonds are being continually broken and formed (Panel 22, pp. 112–113). It is only because of the hydrogen bonds that link water molecules together that water is a liquid at room temperature, with a high boiling point and high surface tension— rather than a gas. Molecules, such as alcohols, that contain polar bonds and that can form hydrogen bonds with water dissolve readily in water. As mentioned previously, molecules carrying plus or minus charges (ions) likewise interact favorably with water. Such molecules are termed hydrophilic, meaning that they are water-loving. A large proportion of the molecules in the aqueous environment of a cell necessarily fall into this category, including sugars, DNA, RNA, and a majority of proteins. Hydrophobic (water-hating) molecules, by contrast, are uncharged and form few or no hydrogen bonds, and so do not dissolve in water. Hydrocarbons are an important example (see Panel 2-1, pp. 110–111). In these molecules the H atoms are covalently linked to C atoms by a largely nonpolar bond. Because the H atoms have almost no net positive charge, they cannot form effective hydrogen bonds to other molecules. This makes the hydrocarbon as a whole hydrophobic—a property that is exploited in cells, whose membranes are constructed from molecules that have long hydrocarbon tails, as we shall see in Chapter 10. Some Polar Molecules Form Acids and Bases in Water One of the simplest kinds of chemical reaction, and one that has profound significance in cells, takes place when a molecule possessing a highly polar covalent bond between a hydrogen and a second atom dissolves in water. The hydrogen atom in such a molecule has largely given up its electron to the companion atom and so exists as an almost naked positively charged hydrogen nucleus—in other words, a proton (H + ). When the polar molecule becomes surrounded by water molecules, the proton is attracted to the partial negative charge on the O atom of an adjacent water molecule and can dissociate from its original partner to associate instead with the oxygen atoms of the water molecule to generate a hydronium ion (H 3 QuickTime™ and a TIFF (Uncompressed) decompressor are needed to see this picture. O (Figure 2-13A). +) Figure 2-13. Acids in water. (A) The reaction that takes place when a molecule of acetic acid dissolves in water. (B) Water molecules are continuously exchanging protons with each other to form hydronium and hydroxyl ions. These ions in turn rapidly recombine to form water molecules. The reverse reaction also takes place very readily, so one has to imagine an equilibrium state in which billions of protons are constantly flitting to and fro from one molecule in the solution to another. Substances that release protons to form H3O+ when they dissolve in water are termed acids. The higher the concentration of H3O+, the more acidic the solution. H3O+ is present even in pure water, at a concentration of 10-7 M, as a result of the movement of protons from one water molecule to another (Figure 2-13B). By tradition, the H3O+ concentration is usually referred to as the H+ concentration, even though most H+ in an aqueous solution is present as H3O+. To avoid the use of unwieldy numbers, the concentration of H+ is expressed using a logarithmic scale called the pH scale, as illustrated in Panel 2-2 (pp. 112–113). Pure water has a pH of 7.0. Because the proton of a hydronium ion can be passed readily to many types of molecules in cells, altering their character, the concentration of H3O+ inside a cell (the acidity) must be closely regulated. Molecules that can give up protons will do so more readily if the concentration of H3O+ in solution is low and will tend to receive them back if the concentration in solution is high. The opposite of an acid is a base. Just as the defining property of an acid is that it donates protons to a water molecule so as to raise the concentration of H3O+ ions, the defining property of a base is that it raises the concentration of hydroxyl (OH-) ions—which are formed by removal of a proton from a water molecule. Thus sodium hydroxide (NaOH) is basic (the term alkaline is also used) because it dissociates in aqueous solution to form Na+ ions and OHions. Another class of bases, especially important in living cells, are those that contain NH2 groups. These groups can generate OHby taking a proton from water: -NH2 + H2O → -NH3+ + OH-. Because an OH- ion combines with a H3O+ ion to form two water molecules, an increase in the OH- concentration forces a decrease in the concentration of H3O+, and vice versa. A pure solution of water contains an equally low concentration (10-7 M) of both ions; it is neither acidic nor basic and is therefore said to be neutral with a pH of 7.0. The inside of cells is kept close to neutrality. [Stop here if you do not have time to finish--I would prefer that you read all three selections up to the places marked like this, and leave the somewhat more difficult or detailed material that follows. In that case skip to the selection by Berg et al.] [I included this material on noncovalent interactions although we are concentrating on covalent bonding before we turn to what will be our main target, the noncovalent bonds. So read it if you are eager.] Four Types of Noncovalent Interactions Help Bring Molecules Together in Cells In aqueous solutions, covalent bonds are 10 to 100 times stronger than the other attractive forces between atoms, allowing their connections to define the boundaries of one molecule from another. But much of biology depends on the specific binding of different molecules to each other. This binding is mediated by a group of noncovalent attractions that are individually quite weak, but whose bond energies can sum to create an effective force between two separate molecules. We have already introduced three of these noncovalent forces: ionic bonds, hydrogen bonds and van der Waals attractions. In Table 2-2, the strengths of these three types of bonds are compared to that of a typical covalent bond, both in the presence and the absence of water. Because of their fundamental importance in all biological systems, we shall summarize their properties here. Ionic bonds. These are purely electrostatic attractions between oppositely charged atoms. As we saw for NaCl, these forces are quite strong in the absence of water. However, the polar water molecules cluster around both fully charged ions and polar molecules that contain permanent dipoles (Figure 214). on water molecules orient to reduce the affinity of oppositely charged ions or polar groups for each other. QuickTime™ and a TIFF (Uncompressed) decompressor are needed to see this picture. Figure 2-14. How the dipoles This greatly reduces the potential attractiveness of these charged species for each other (see Table 2-2). Hydrogen bonds. The structure of a typical hydrogen bond is QuickTime™ and a TIFF (Uncompressed) decompressor are needed to see this picture. illustrated in Figure 2-15. Figure 2-15. Hydrogen bonds. (A) Ball- and-stick model of a typical hydrogen bond. The distance between the hydrogen and the oxygen atom here is less than the sum of their van der Waals radii, indicating a partial sharing of electrons. (B) The most common hydrogen bonds in cells. This bond represents a special form of polar interaction in which an electropositive hydrogen atom is partially shared by two electronegative atoms. Its hydrogen can be viewed as a proton that has partially dissociated from a donor atom, allowing it to be shared by a second acceptor atom. Unlike a typical electrostatic interaction, this bond is highly directional—being strongest when a straight line can be drawn between all three of the involved atoms. As already discussed, water weakens these bonds by forming competing hydrogen-bond interactions with the involved molecules (see Table 2-2). van der Waals attractions. The electron cloud around any nonpolar atom will fluctuate, producing a flickering dipole. Such dipoles will transiently induce an oppositely polarized flickering dipole in a nearby atom. This interaction generates an attraction between atoms that is very weak. But since many atoms can be simultaneously in contact when two surfaces fit closely, the net result is often significant. These so-called van der Waals attractions are not weakened by water (see Table 2-2). The fourth effect that can play an important part in bringing molecules together in water is a hydrophobic force. This force is caused by a pushing of nonpolar surfaces out of the hydrogenbonded water network, where they would physically interfere with the highly favorable interactions between water molecules. Because bringing two nonpolar surfaces together reduces their contact with water, the force is a rather nonspecific one. Nevertheless, we shall see in Chapter 3 that hydrophobic forces are central to the proper folding of protein molecules. Panel 2-3 provides an overview of the four types of interactions just described. (You should be able to link to it from this document.) QuickTime™ and a TIFF (Uncompressed) decompressor are needed to see this picture. Figure 2-16. How two macro-molecules with complementary surfaces can bind tightly to one another through noncovalent interactions. In this schematic illustration, plus and minus are used to mark chemical groups that can form attractive interactions when paired. And Figure 2-16 above illustrates, in a schematic way, how many such interactions can sum to hold together the matching surfaces of two macromolecules, even though each interaction by itself would be much too weak to be effective. [Chap. 2 of Alberts continues with “A cell is formed from carbon compounds, which introduces a little organic chemistry before going into the major classes of macromolecules: Save for another reading collection, and keep this close to bonding.] _____________________________________________ NEXT section of this document: A parallel but slightly more extended discussion of “Chemical bonds in biochemistry,” Sec. 1.3 in Berg et al., Biochemistry. Note the emphasis on the various weak noncovalent forces and the roles they play. Keep thinking--what sort of structures or phenomena could they give rise to in nonbiological mesoscale macromolecular assemblies? (Note: This is NOT a detailed treatment of each kind of molecule--nearly each topic, like protein folding, is treated in separate chapters later in the book.) Berg et al. Biochemistry 1.3. Chemical Bonds in Biochemistry [I have included the several pages discussing the noncovalent interactions which are of primary interest to us because it is a short and painless introduction. I have omitted the (longer) sections on this topic from the other two reading selections. By the time you read this we should have covered the interactions from a physics point of view, so this will be complementary.] The essence of biological processes—the basis of the uniformity of living systems—is in its most fundamental sense molecular interactions; in other words, the chemistry that takes place between molecules. Biochemistry is the chemistry that takes place within living systems. To truly understand biochemistry, we need to understand chemical bonding. We review here the types of chemical bonds that are important for biochemicals and their transformations. The strongest bonds that are present in biochemicals are covalent bonds, such as the bonds that hold the atoms together within the individual bases shown in Figure 1.3. Figure 1.3. Watson-Crick Base Pairs. Adenine pairs with thymine (A-T), and guanine with cytosine (G- C). The dashed lines represent hydrogen bonds. A covalent bond is formed by the sharing of a pair of electrons between adjacent atoms. A typical carbon-carbon (C-C) covalent bond has a bond length of 1.54 Å and bond energy of 85 kcal mol-1 (356 kJ mol-1). Because this energy is relatively high, considerable energy must be expended to break covalent bonds. More than one electron pair can be shared between two atoms to form a multiple covalent bond. For example, three of the bases in Figure 1.4 QuickTime™ and a TIFF (Uncompressed) decompressor are needed to see this picture. Figure 1.4. Base-Pairing in DNA. The base-pairs A-T (blue) and C-G (red) are shown overlaid. The Watson-Crick base-pairs have the same overall size and shape, allowing them to fit neatly within the double helix. include carbon-oxygen (C=O) double bonds. These bonds are even stronger than C-C single bonds, with energies near 175 kcal mol-1 (732 kJ mol-1). For some molecules, more than one pattern of covalent bonding can be written. For example, benzene can be written in two equivalent ways called resonance structures. Benzene's true structure is a composite of its two resonance structures. A molecule that can be written as several resonance structures of approximately equal energies has greater stability than does a molecule without multiple resonance structures. Thus, because of its resonance structures, benzene is unusually stable. Chemical reactions entail the breaking and forming of covalent Quic kTime™ and a bonds. The flow of electrons in TIFF (Unc ompres sed) dec ompres sor are needed to see this pic ture. the course of a reaction can be depicted by curved arrows, a method of representation called “arrow pushing.” Each arrow represents an electron pair. QuickTime™ and a TIFF (Uncompressed) decompressor are needed to see this picture. 1.3.1. Reversible Interactions of Biomolecules Are Mediated by Three Kinds of Noncovalent Bonds Readily reversible, noncovalent molecular interactions are key steps in the dance of life. Such weak, noncovalent forces play essential roles in the faithful replication of DNA, the folding of proteins into intricate three-dimensional forms, the specific recognition of substrates by enzymes, and the detection of molecular signals. Indeed, all biological structures and processes depend on the interplay of noncovalent interactions as well as covalent ones. The three fundamental noncovalent bonds are electrostatic interactions, hydrogen bonds, and van der Waals interactions. They differ in geometry, strength, and specificity. Furthermore, these bonds are greatly affected in different ways by the presence of water. Let us consider the characteristics of each: 1. Electrostatic interactions. An electrostatic interaction depends on the electric charges on atoms. The energy of an electrostatic interaction is given by Coulomb's law: QuickTi me™ and a TIFF ( Uncompressed) decompressor are needed to see thi s pi ctur e. QuickTime™ and a TIFF (Uncompressed) decompressor are needed to see this picture. where E is the energy, q1 and q2 are the charges on the two atoms (in units of the electronic charge), r is the distance between the two atoms (in angstroms), D is the dielectric constant (which accounts for the effects of the intervening medium), and k is a proportionality constant (k = 332, to give energies in units of kilocalories per mole, or 1389, for energies in kilojoules per mole). Thus, the electrostatic interaction between two atoms bearing single opposite charges separated by 3 Å in water (which has a dielectric constant of 80) has an energy of 1.4 kcal mol-1 (5.9 kJ mol-1). 2. Hydrogen bonds. Hydrogen bonds are relatively weak interactions, which nonetheless are crucial for biological macromolecules such as DNA and proteins. These interactions are also responsible for many of the properties of water that make it such a special solvent. The hydrogen atom in a hydrogen bond is partly shared between two relatively electronegative atoms such as nitrogen or oxygen. The hydrogenbond donor is the group that includes both the atom to which the hydrogen is more tightly linked and the hydrogen atom itself, whereas the hydrogen-bond acceptor is the atom less tightly linked to the hydrogen atom (Figure QuickTime™ and a TIFF (Uncompress ed) dec ompres sor are needed to s ee this pic ture. 1.9). Figure 1.9. Hydrogen Bonds that Include Nitrogen and Oxygen Atoms. The positions of the partial charges (δ+ and δ-) are shown. Hydrogen bonds are fundamentally electrostatic interactions. The relatively electronegative atom to which the hydrogen atom is covalently bonded pulls electron density away from the hydrogen atom so that it develops a partial positive charge (δ+). Thus, it can interact with an atom having a partial negative charge (δ-) through an electrostatic interaction. QuickTime™ and a TIFF (Uncompress ed) dec ompres sor are needed to s ee this pic ture. Hydrogen bonds are much weaker than covalent bonds. They have energies of 1–3 kcal mol-1 (4–13 kJ mol-1) compared with approximately 100 kcal mol-1 (418 kJ mol-1) for a carbon-hydrogen covalent bond. Hydrogen bonds are also somewhat longer than are covalent bonds; their bond distances (measured from the hydrogen atom) range from 1.5 to 2.6 Å; hence, distances ranging from 2.4 to 3.5 Å separate the two nonhydrogen atoms in a hydrogen bond. The strongest hydrogen bonds have a tendency to be approximately straight, such that the hydrogen-bond donor, the hydrogen atom, and the hydrogen-bond acceptor lie along a straight line. 3. van der Waals interactions. The basis of a van der Waals interaction is that the distribution of electronic charge around an atom changes with time. At any instant, the charge distribution is not perfectly symmetric. This transient asymmetry in the electronic charge around an atom acts through electrostatic interactions to induce a complementary asymmetry in the electron distribution around its neighboring atoms. The resulting attraction between two atoms increases as they come closer to each other, until they are separated by the van der Waals contact distance QuickTime™ and a TIFF (Uncompressed) decompressor are needed to see this picture. (Figure 1.10). Figure 1.10. Energy of a van der Waals Interaction as Two Atoms Approach One Another. The energy is most favorable at the van der Waals contact distance. The energy rises rapidly owing to electron- electron repulsion as the atoms move closer together than this distance. At a shorter distance, very strong repulsive forces become dominant because the outer electron clouds overlap. Energies associated with van der Waals interactions are quite small; typical interactions contribute from 0.5 to 1.0 kcal mol-1 (from 2 to 4 kJ mol-1) per atom pair. When the surfaces of two large molecules come together, however, a large number of atoms are in van der Waals contact, and the net effect, summed over many atom pairs, can be substantial. 1.3.2. The Properties of Water Affect the Bonding Abilities of Biomolecules Weak interactions are the key means by which molecules interact with one another—enzymes with their substrates, hormones with their receptors, antibodies with their antigens. The strength and specificity of weak interactions are highly dependent on the medium in which they take place, and the majority of biological interactions take place in water. Two properties of water are especially important biologically: QuickTi me™ and a TIFF ( Uncompressed) decompressor are needed to see thi s pi ctur e. 1. Water is a polar molecule. The water molecule is bent, not linear, and so the distribution of charge is asymmetric. The oxygen nucleus draws electrons away from the hydrogen nuclei, which leaves the region around the hydrogen nuclei with a net positive charge. The water molecule is thus an electrically polar structure. 2. Water is highly cohesive. Water molecules interact strongly with one another through hydrogen bonds. These interactions are apparent in the structure of ice (Figure 1.11). QuickTime™ and a TIFF (Uncompressed) decompressor are needed to see this picture. Figure 1.11. Structure of Ice. Hydrogen bonds (shown as dashed lines) are formed between water molecules. Networks of hydrogen bonds hold the structure together; simi-lar interactions link molecules in liquid water and account for the cohesion of liquid water, although, in the liquid state, some of the hydrogen bonds are broken. The highly cohesive nature of water dramatically affects the interactions between molecules in aqueous solution. What is the effect of the properties of water on the weak interactions discussed in Section 1.3.1? The polarity and hydrogen-bonding capability of water make it a highly interacting molecule. Water is an excellent solvent for polar molecules. The reason is that water greatly weakens electrostatic forces and hydrogen bonding between polar molecules by competing for their attractions. For example, consider the effect of water on hydrogen bonding between a carbonyl group and the NH group of an amide. QuickTime™ and a TIFF (Uncompressed) decompressor are needed to see this picture. A hydrogen atom of water can replace the amide hydrogen atom as a hydrogen-bond donor, whereas the oxygen atom of water can replace the carbonyl oxygen atom as a hydrogen-bond acceptor. Hence, a strong hydrogen bond between a CO group and an NH group forms only if water is excluded. The dielectric constant of water is 80, so water diminishes the strength of electrostatic attractions by a factor of 80 compared with the strength of those same interactions in a vacuum. The dielectric constant of water is unusually high because of its polarity and capacity to form oriented solvent shells around ions. These oriented solvent shells produce electric fields of their own, which oppose the fields produced by the ions. Consequently, the presence of water markedly weakens electrostatic interactions between ions. The existence of life on Earth depends critically on the capacity of water to dissolve a remarkable array of polar molecules that serve as fuels, building blocks, catalysts, and information carriers. High concentrations of these polar molecules can coexist in water, where they are free to diffuse and interact with one another. However, the excellence of water as a solvent poses a problem, because it also weakens interactions between polar molecules. The presence of water-free microenvironments within biological systems largely circumvents this problem. We will see many examples of these specially constructed niches in protein molecules. Moreover, the presence of water with its polar nature permits another kind of weak interaction to take place, one that drives the folding of proteins (Section 1.3.4) and the formation of cell boundaries (Section 12.4). The essence of these interactions, like that of all interactions in biochemistry, is energy. To understand much of biochemistry—bond formation, molecular structure, enzyme catalysis—we need to understand energy. Thermodynamics provides a valuable tool for approaching this topic. We will revisit this topic in more detail when we consider enzymes (Chapter 8) and the basic concepts of metabolism (Chapter 14). [Stop here if you do not have time to finish--I would prefer that you read all three selections up to the places marked like this, and leave the somewhat more difficult or detailed material that follows. In that case skip to the selection by Lodish et al. ________________________________________ [I included this material on thermodynamical view of bond formation in case you are interested or if you have never had thermodynamics, as an opportunity to see this point of view. However sec. 1.3.3 and 1.3.4 below (on protein folding) are optional reading, and you won’t be tested on it and no assigned homework will depend on it.] 1.3.3. Entropy and the Laws of Thermodynamics The highly structured, organized nature of living organisms is apparent and astonishing. This organization extends from the organismal through the cellular to the molecular level. Indeed, biological processes can seem magical in that the well-ordered structures and patterns emerge from the chaotic and disordered world of inanimate objects. However, the organization visible in a cell or a molecule arises from biological events that are subject to the same physical laws that govern all processes—in particular, the laws of thermodynamics. How can we understand the creation of order out of chaos? We begin by noting that the laws of thermodynamics make a distinction between a system and its surroundings. A system is defined as the matter within a defined region of space. The matter in the rest of the universe is called the surroundings. The First Law of Thermodynamics states that the total energy of a system and its surroundings is constant. In other words, the energy content of the universe is constant; energy can be neither created nor destroyed. Energy can take different forms, however. Heat, for example, is one form of energy. Heat is a manifestation of the kinetic energy associated with the random motion of molecules. Alternatively, energy can be present as potential energy, referring to the ability of energy to be released on the occurrence of some process. Consider, for example, a ball held at the top of a tower. The ball has considerable potential energy because, when it is released, the ball will develop kinetic energy associated with its motion as it falls. Within chemical systems, potential energy is related to the likelihood that atoms can react with one another. For instance, a mixture of gasoline and oxygen has much potential energy because these molecules may react to form carbon dioxide and release energy as heat. The First Law requires that any energy released in the formation of chemical bonds be used to break other bonds, be released as heat, or be stored in some other form. Another important thermodynamic concept is that of entropy. Entropy is a measure of the level of randomness or disorder in a system. The Second Law of Thermodynamics states that the total entropy of a system and its surroundings always increases for a spontaneous process. At first glance, this law appears to contradict much common experience, particularly about biological systems. Many biological processes, such as the generation of a well-defined structure such as a leaf from carbon dioxide gas and other nutrients, clearly increase the level of order and hence decrease entropy. Entropy may be decreased locally in the formation of such ordered structures only if the entropy of other parts of the universe is increased by an equal or greater amount. An example may help clarify the application of the laws of thermodynamics to a chemical system. Consider a container with 2 moles of hydrogen gas on one side of a divider and 1 mole of oxygen gas on the other (Figure 1.12). QuickTime™ and a TIFF (Uncompressed) decompressor are needed to see this picture. Figure 1.12. From Order to Disorder. The spontaneous mixing of gases is driven by an increase in entropy. If the divider is removed, the gases will intermingle spontaneously to form a uniform mixture. The process of mixing increases entropy as an ordered arrangement is replaced by a randomly distributed mixture. Other processes within this system can decrease the entropy locally while increasing the entropy of the universe. A spark applied to the mixture initiates a chemical reaction in which hydrogen and oxygen combine to form water: QuickTi me™ and a TIFF (Uncompressed) decompressor are needed to see this picture. If the temperature of the system is held constant, the entropy of the system decreases because 3 moles of two differing reactants have been combined to form 2 moles of a single product. The gas now consists of a uniform set of indistinguishable molecules. However, the reaction releases a significant amount of heat into the surroundings, and this heat will increase the entropy of the surrounding molecules by increasing their random movement. The entropy increase in the surroundings is enough to allow water to form spontaneously from hydrogen and oxygen (Figure 1.13). QuickTime™ and a TIFF (Uncompressed) decompressor are needed to see this picture. Figure 1.13. Entropy Changes. When hydrogen and oxygen combine to form water, the entropy of the system is reduced, but the entropy of the universe is increased owing to the release of heat to the surroundings. The change in the entropy of the surroundings will be proportional to the amount of heat transferred from the system and inversely proportional to the temperature of the surroundings, because an input of heat leads to a greater increase in entropy at lower temperatures than at higher temperatures. In biological systems, T [in kelvin (K), absolute temperature] is assumed to be constant. If we define the heat content of a system as enthalpy (H), then we can express the relation linking the entropy (S) of the surroundings to the transferred heat and temperature as a simple equation: QuickTime™ and a TIFF (Uncompressed) decompressor are needed to see this picture. The total entropy change is given by the expression QuickTime™ and a TIFF (Uncompressed) decompressor are needed to see this picture. Substituting equation 1 into equation 2 yields QuickTime™ and a TIFF (Uncompressed) decompressor are needed to see this picture. Multiplying by -T gives QuickTime™ and a TIFF (Uncompressed) decompressor are needed to see this picture. The function -TΔS has units of energy and is referred to as free energy or Gibbs free energy, after Josiah Willard Gibbs, who developed this function in 1878: QuickTime™ and a TIFF (Uncompressed) decompressor are needed to see this picture. The free-energy change, ΔG, will be used throughout this book to describe the energetics of biochemical reactions. Recall that the Second Law of Thermodynamics states that, for a reaction to be spontaneous, the entropy of the universe must increase. Examination of equation 3 shows that the total entropy will increase if and only if QuickTime™ and a TIFF (Uncompressed) decompressor are needed to see this picture. Rearranging gives TΔSsystem > ΔH, or entropy will increase if and only if QuickTime™ and a TIFF (Uncompressed) decompressor are needed to see this picture. w In other words, the free-energy change must be negative for a reaction to be spontaneous. A negative free-energy change occurs with an increase in the overall entropy of the universe. Thus, we need to consider only one term, the free energy of the system, to decide whether a reaction can occur spontaneously; any effects of the changes within the system on the rest of the universe are automatically taken into account. 1.3.4. Protein Folding Can Be Understood in Terms of Free-Energy Changes The problem of protein folding illustrates the utility of the concept of free energy. ConsIder a system consisting of a solution of unfolded protein molecules in aqueous solution (Figure 1.14). QuickTime™ and a TIFF (Uncompressed) decompressor are needed to see this picture. Figure 1.14. Protein Folding. Protein folding entails the transition from a disordered mixture of unfolded molecules to a relatively uniform solution of folded protein molecules. Each unfolded protein molecule can adopt a unique conformation, so the system is quite disordered and the entropy of the collection of molecules is relatively high. Yet, protein folding proceeds spontaneously under appropriate conditions. Thus, entropy must be increasing elsewhere in the system or in the surroundings. How can we reconcile the apparent contradiction that proteins spontaneously assume an ordered structure, and yet entropy increases? The entropy decrease in the system on folding is not as large as it appears to be, because of the properties of water. Molecules in aqueous solution interact with water molecules through the formation of hydrogen and ionic interactions. However, some molecules (termed nonpolar molecules) cannot participate in hydrogen or ionic interactions. The interactions of nonpolar molecules with water are not as favorable as are interactions between the water molecules themselves. The water molecules in contact with these nonpolar surfaces form “cages” around the nonpolar molecule, becoming more well ordered (and, hence, lower in entropy) than water molecules free in solution. As two such nonpolar molecules come together, some of the water molecules are released, and so they can interact freely with bulk water (Figure 1.15). QuickTime™ and a TIFF (Uncompressed) decompressor are needed to see this picture. Figure 1.15. The Hydrophobic Effect. The aggregation of nonpolar groups in water leads to an increase in entropy owing to the release of water molecules into bulk water. Hence, nonpolar molecules have a tendency to aggregate in water because the entropy of the water is increased through the release of water molecules. This phenomenon, termed the hydrophobic effect, helps promote many biochemical processes. How does the hydrophobic effect favor protein folding? Some of the amino acids that make up proteins have nonpolar groups. These nonpolar amino acids have a strong tendency to associate with one another inside the interior of the folded protein. The increased entropy of water resulting from the interaction of these hydrophobic amino acids helps to compensate for the entropy losses inherent in the folding process. Hydrophobic interactions are not the only means of stabilizing protein structure. Many weak bonds, including hydrogen bonds and van der Waals interactions, are formed in the protein-folding process, and heat is released into the surroundings as a consequence. Although these interactionws replace interactions with water that take place in the unfolded protein, the net result is the release of heat to the surroundings and thus a negative (favorable) change in enthalpy for the system. The folding process can occur when the combination of the entropy associated with the hydrophobic effect and the enthalpy change associated with hydrogen bonds and van der Waals interactions makes the overall free energy negative. Lodish, Molecular Cell Biology Ch. 2 Chemical Foundations 2.1. Covalent Bonds [I have left out sec. 2.2, a great discussion of noncovalent bonds; we will read that separately.] Covalent bonds, which hold the atoms within an individual molecule together, are formed by the sharing of electrons in the outer atomic orbitals. The distribution of shared as well as unshared electrons in outer orbitals is a major determinant of the three-dimensional shape and chemical reactivity of molecules. For instance, as we learn in Chapter 3, the shape of proteins is crucial to their function and their interactions with small molecules. In this section, we discuss important properties of covalent bonds and describe the structure of carbohydrates to illustrate how the geometry of bonds determines the shape of small biological molecules. Each Atom Can Make a Defined Number of Covalent Bonds Electrons move around the nucleus of an atom in clouds called orbitals, which lie in a series of concentric shells, or energy levels; electrons in outer shells have more energy than those in inner shells. Each shell has a maximum number of electrons that it can hold. Electrons fill the innermost shells of an atom first; then the outer shells. The energy level of an atom is lowest when all of its orbitals are filled, and an atom’s reactivity depends on how many electrons it needs to complete its outermost orbital. In most cases, in order to fill the outermost orbital, the electrons within it form covalent bonds with other atoms. A covalent bond thus holds two atoms close together because electrons in their outermost orbitals are shared by both atoms. Most of the molecules in living systems contain only six different atoms: hydrogen, carbon, nitrogen, phosphorus, oxygen, and sulfur. The outermost orbital of each atom has a characteristic number of electrons: QuickTime™ and a TIFF (Uncompressed) decompressor are needed to see this picture. These atoms readily form covalent bonds with other atoms and rarely exist as isolated entities. As a rule, each type of atom forms a characteristic number of covalent bonds with other atoms. For example, a hydrogen atom, with one electron in its outer shell, forms only one bond, such that its outermost orbital becomes filled with two electrons. A carbon atom has four electrons in its outermost orbitals; it usually forms four bonds, as in methane (CH4), in order to fill its outermost orbital with eight electrons. The single bonds in methane that connect the carbon atom with each hydrogen atom contain two shared electrons, one donated from the C and the other from the H, and the outer (s) orbital of each H atom is filled by the two shared electrons: QuickTime™ and a TIFF (Uncompressed) decompressor are needed to see this picture. Nitrogen and phosphorus each have five electrons in their outer shells, which can hold up to eight electrons. Nitrogen atoms can form up to four covalent bonds. In ammonia (NH3), the nitrogen atom forms three covalent bonds; one pair of electrons around the atom (the two dots on the right) are in an orbital not involved in a covalent bond: QuickTime™ and a TIFF (Uncompress ed) dec ompres sor are needed to s ee this pic ture. In the ammonium ion (NH4+), the nitrogen atom forms four covalent bonds, again filling the outermost orbital with eight electrons: Quick Time™a nd a TIFF ( Unco mpre ssed ) dec ompr esso r ar e nee ded to see this pictur e. Phosphorus can form up to five covalent bonds, as in phosphoric acid (H3PO4). The H3PO4 molecule is actually a “resonance hybrid,” a structure between the two forms shown below in which nonbonding electrons are shown as pairs of dots: QuickTime™ and a TIFF (Uncompressed) decompressor are needed to see this picture. In the resonance hybrid on the right, one of the electrons from the P=O double bond has accumulated around the O atom, giving it a net negative charge and leaving the P atom with a net positive charge. The resonance hybrid on the left, in which the P atom forms the maximum five covalent bonds, has no charged atoms. Esters of phosphoric acid form the backbone of nucleic acids, as discussed in Chapter 4; phosphates also play key roles in cellular energetics (Chapter 16) and in the regulation of cell function (Chapters 13 and 20). The difference between the bonding patterns of nitrogen and phosphorus is primarily due to the relative sizes of the two atoms: the smaller nitrogen atom has only enough space to accommodate four bonding pairs of electrons around it without creating destructive repulsions between them, whereas the larger sphere of the phosphorus atom allows more electron pairs to be arranged around it without the pairs being too close together. Both oxygen and sulfur contain six electrons in their outermost orbitals. However, an atom of oxygen usually forms only two covalent bonds, as in molecular oxygen, O2: QuickTime™ and a TIFF (Uncompress ed) dec ompres sor are needed to s ee this pic ture. Primarily because its outermost orbital is larger than that of oxygen, sulfur can form as few as two covalent bonds, as in hydrogen sulfide (H2S), or as many as six, as in sulfur trioxide (SO3) or sulfuric acid (H2SO4): QuickTime™ and a TIFF (Uncompressed) decompressor are needed to see this picture. Esters of sulfuric acid are important constituents of the proteoglycans that compose part of the extracellular matrix surrounding most animal cells (Chapter 22). The Making or Breaking of Covalent Bonds Involves Large Energy Changes Covalent bonds tend to be very stable because the energies required to break or rearrange them are much greater than the thermal energy available at room temperature (25 °C) or body temperature (37 °C). For example, the thermal energy at 25 °C is less than 1 kilocalorie per mole (kcal/mol), whereas the energy required to breakQuickTime™ a C—C bond in ethane is about 83 kcal/mol: and a TIFF (Uncompressed) decompressor are needed to see this picture. where ΔH represents the difference in the total energy of all of the bonds (the enthalpy) in the reactants and in the products.*The positive value indicates that an input of energy is needed to cause the reaction, and that the products contain more energy than the reactants. The high energy needed for breakage of the ethane bond means that at room temperature (25 °C) well under 1 in 1012 ethane molecules exists as a pair of ·CH3 radicals. The covalent bonds in biological molecules have ΔH values similar to that of the C—C bond in ethane (Table 2-1). Table 2-1. The Energy Required to Break Some Important Covalent Bonds Found in Biological Molecules* Type of Bond SINGLE BOND O—H H—H P—O C—H C—O C—C S—H C—N C—S N—O S—S Energy (kcal/mol) 110 104 100 99 84 83 81 70 62 53 51 Type of Bond Energy (kcal/mol) DOUBLE BOND C=O C=N C=C P=O 170 147 146 120 TRIPLE BOND C≡O 195 * Note that double and triple bonds are stronger than single bonds. * A calorie is defined as the amount of thermal energy required to heat 1 cm3 of water by 1 °C from 14 °C to 15 °C. Many biochemistry textbooks use the joule (J), but the two units can be interconverted quite readily (1 cal = 4.184 J). The energy changes in chemical reactions, such as the making or breaking of chemical bonds, are measured in kilocalories per mole in this book (1 kcal = 1000 cal). One mole of any substance is the amount that contains 6.02 × 1023 items of that substance, which is known as Avogadro’s number. Thus, one can speak of a mole of photons, or 6.02 × 1023 photons. The weight of a mole of a substance in grams (g) is the same as its molecular weight. For example, the molecular weight of water is 18, so the weight of 1 mole of water, or 6.02 × 1023 water molecules, is 18 g. Covalent Bonds Have Characteristic Geometries When two or more atoms form covalent bonds with another central atom, these bonds are oriented at precise angles to one another. The angles are determined by the mutual repulsion of the outer electron orbitals of the central atom. These bond angles give each molecule its characteristic shape (Figure 2-2). QuickTime™ and a TIFF (Uncompressed) decompressor are needed to see this picture. In methane, for example, the central carbon atom is bonded to four hydrogen atoms, whose positions define the four points of a tetrahedron, so that the angle between any two bonds is 109.5°. Like methane, the ammonium ion also has a tetrahedral shape. In these molecules, each bond is a single bond, a single pair of electrons shared between two atoms. When two atoms share two pairs of electrons — for example, when a carbon atom is linked to only three other atoms — the bond is a double bond: Qu ic kTi me™ a nd a TIFF (Unc om pres se d) de co mp re ss or are n ee de d to s ee th is pi ctu re . In this case, the carbon atom and all three atoms linked to it lie in the same plane (Figure 2-3). QuickTime™ and a TIFF (Uncompressed) decompressor are needed to see this picture. Atoms connected by a double bond cannot rotate freely about the bond axis, while those in a single bond generally can. The rigid planarity imposed by double bonds has enormous significance for the shape of large biological molecules such as proteins and nucleic acids. (In triple bonds, two atoms share six electrons. These are rare in biological molecules.) [Conversely, atoms connected by a single bond can rotate freely and allow the molecules to adopt a variety of shapes which are called conformational isomers. This will become an important theme as we approach more complex molecules.] All outer electron orbitals, whether or not they are involved in covalent bond formation, contribute to the properties of a molecule, in particular to its shape. For example, the outer shell of the oxygen atom in a water molecule has two pairs of nonbonding electrons; the two pairs of electrons in the H—O bonds and the two pairs of nonbonding electrons form an almost perfect tetrahedron. However, the orbitals of the nonbonding electrons have a high electron density and thus tend to repel each other, compressing the angle between the covalent H—O—H bonds to 104.5° rather than the 109.5° in a tetrahedron (see Figure 2-2). [More on molecular shape provided separately.] Electrons Are Shared Unequally in Polar Covalent Bonds In a covalent bond, one or more pairs of electrons are shared between two atoms. In certain cases, the bonded atoms exert different attractions for the electrons of the bond, resulting in unequal sharing of the electrons. The power of an atom in a molecule to attract electrons to itself, called electronegativity, is measured on a scale from 4.0 (for fluorine, the most electronegative atom) to a hypothetical zero (Figure 2-4). QuickTime™ and a TIFF (Uncompressed) decompressor are needed to see this picture. Figure 2-4. Electronegativity values of main-group elements in the periodic table. Atoms located to the upper right tend to have high electronegativity, fluorine being the most electronegative. Elements with low electronegativity values, such as the metals lithium, sodium, and potassium, are often called electropositive. The electronegativities of several atoms abundant in biological molecules differ enough that they form polar covalent bonds (e.g., O—H, N—H) or ionic bonds (e.g., Na+Cl−). Because the inert gases (He, Ne, etc.) have complete outer shells of electrons, they neither attract nor donate electrons, rarely form covalent bonds, and have no electronegativity values. Knowing the electronegativity of two atoms allows us to predict whether a covalent bond can form between them; if the differences in electronegativity are considerable — as in sodium and chloride — an ionic bond, rather than a covalent bond, will form. This type of interaction is discussed in a later section. In a covalent bond in which the atoms either are identical or have the same electronegativity, the bonding electrons are shared equally. Such a bond is said to be nonpolar. This is the case for C—C and C—H bonds. However, if two atoms differ in electronegativity, the bond is said to be polar. One end of a polar bond has a partial negative charge (δ−), and the other end has a partial positive charge (δ+). In an O—H bond, for example, the oxygen atom, with an electronegativity of 3.4, attracts the bonded electrons more than does the hydrogen atom, which has an electronegativity of 2.2. As a result, the bonding electrons spend more time around the oxygen atom than around the hydrogen. Thus the O—H bond possesses an electric dipole, a positive charge separated from an equal but opposite negative charge. We can think of the oxygen atom of the O—H bond as having, on average, a charge of 25 percent of an electron, with the H atom having an equivalent positive charge. The dipole moment of the O—H bond is a function of the size of the positive or negative charge and the distance separating the charges. [Later we will see that dipole moments, whether permanent or induced or fluctuating around zero, play the major role in controlling the formation of the macromolecules and supramolecular assemblies or macromolecules that we label as grains, PAH assemblies, aerosols, nanoparticles, and biomolecules.] In a water molecule both hydrogen atoms are on the same side of the oxygen atom. As a result, the side of the molecule with the two H atoms has a slight net positive charge, whereas the other side has a slight net negative charge. Because of this separation of positive and negative charges, the entire molecule has a net dipole moment (Figure 2-5). Some molecules, such as the linear molecule CO2, have two polar bonds: QuickTime™ and a TIFF (Uncompressed) decompressor are needed to see this picture. Figure 2-5. The water molecule has two polar O—H bonds and a net dipole moment. The symbol δ represents a partial charge (a weaker charge than the one on an electron or a proton), and each of the polar H—O bonds has a dipole moment. The net dipole moment of the molecule is determined by the sizes and directions of the dipole moments of each of the bonds. Because the dipole moments of the two C=O bonds point in opposite directions, they cancel each other out, resulting in a molecule without a net dipole moment. NOTE: Water is so essential, along with carbon, that we will devote many pages to understanding its amazing behavior and interactions with other molecules. I thought this was completely overblown a year ago, but no longer. Asymmetric Carbon Atoms Are Present in Most Biological Molecules A carbon (or any other) atom bonded to four dissimilar atoms or groups is said to be asymmetric. The bonds formed by an asymmetric carbon atom can be arranged in three dimensional space in two different ways, producing molecules that are mirror images of each other. Such molecules are called optical isomers, or stereoisomers. One isomer is said to be right-handed and the other left-handed, a property called chirality. Most molecules in cells contain at least one asymmetric carbon atom, often called a chiral carbon atom. The different stereoisomers of a molecule usually have completely different biological activities. Amino Acids Except for glycine, all amino acids, the building blocks of the proteins, have one chiral carbon atom, called the α carbon, or Cα, which is bonded to four different atoms or groups of atoms. In the amino acid alanine, for instance, this carbon atom is bonded to — NH2, —COOH, —H, and —CH3 (Figure 2-6). QuickTime™ and a TIFF (Uncompressed) decompressor are needed to see this picture. Figure 2-6. Stereoisomers of the amino acid alanine. The asymmetric α carbon is black. Although the chemical properties of such optical isomers are identical, their biological activities are distinct. By convention, the two mirror-image structures are called the D (dextro) and the L (levo) isomers of the amino acid. The two isomers cannot be interconverted without breaking a chemical bond. With rare exceptions, only the L forms of amino acids are found in proteins. We discuss the properties of amino acids and the covalent peptide bond that links them into long chains in Chapter 3. Stop here if your time or interest is too limited. _____________________________________________ Carbohydrates The three-dimensional structures of carbohydrates provide another excellent example of the structural and biological importance of chiral carbon atoms, even in simple molecules. A carbohydrate is constructed of carbon (carbo-) plus hydrogen and oxygen (-hydrate, or water). The formula for the simplest carbohydrates — the monosaccharides, or simple sugars — is (CH2O)n, where n equals 3, 4, 5, 6, or 7. All monosaccharides contain hydroxyl (—OH) groups and either an aldehyde or a keto group: QuickTime™ and a TIFF (Uncompressed) decompressor are needed to see this picture. In the linear form of D-glucose (C6H12O6), the principal source of energy for most cells in higher organisms, carbon atoms 2, 3, 4, and 5 are asymmetric (Figure 2-7, top). If the hydrogen atom and the hydroxyl group attached to carbon atom 2 (C2) were interchanged, the resulting molecule would be a different sugar, D-mannose, and could not be converted to glucose without breaking and making covalent bonds. Enzymes can distinguish between this single point of difference. QuickTime™ and a TIFF (Uncompressed) decompressor are needed to see this picture. Figure 2-7. Three alternative configurations of D-glucose. The ring forms, shown as Haworth projections, are generated from the linear molecule by reaction of the aldehyde at carbon 1 with the hydroxyl on carbon 5 or carbon 4. D-Glucose can exist in three different forms: a linear structure and two different hemiacetal ring structures (see Figure 2-7). If the aldehyde group on carbon 1 reacts with the hydroxyl group on carbon 5, the resulting hemiacetal, D-glucopyranose, contains a sixmember ring. Similarly, condensation of the hydroxyl group on carbon 4 with the aldehyde group results in the formation of Dglucofuranose, a hemiacetal containing a five-member ring. Although all three forms of D-glucose exist in biological systems, the pyranose form is by far the most abundant. The planar depiction of the pyranose ring shown in Figure 2-7 is called a Haworth projection. When a linear molecule of D-glucose forms a pyranose ring, carbon 1 becomes asymmetric, so two stereoisomers (called anomers) of D-glucopyranose are possible. The hydroxyl group attached to carbon 1 “points” down (below the plane of projection) in α-D-glucopyranose, as shown in Figure 2-7, and points up (above the plane of projection) in the β anomer. In aqueous solution the α and β anomers readily interconvert spontaneously; at equilibrium there is about one-third α anomer and two-thirds β, with very little of the open-chain form. Because enzymes can distinguish between the α and β anomers of D-glucose, these forms have specific biological roles. Most biologically important sugars are six-carbon sugars, or hexoses, that are structurally related to D-glucose. Mannose, as noted, is identical with glucose except for the orientation of the substituents on carbon 2. In Haworth projections of the pyranose forms of glucose and mannose, the hydroxyl group on carbon 2 of glucose points downward, whereas that on mannose points upward (Figure 2-8). Similarly, galactose, another hexose, differs from glucose only in the orientation of the hydroxyl group on carbon 4. QuickTime™ and a TIFF (Uncompressed) decompressor are needed to see this picture. The Haworth projection is an oversimplification be-cause the actual pyranose ring is not planar. Rather, sugar molecules adopt a conformation in which each of the ring carbons is at the center of a tetrahedron, just like the carbon in methane (see Figure 2-2). The preferred conformation of pyranose structures is the chair (Figure 2-9). In this conformation, the bonds going from a ring carbon to nonring atoms may take two directions: axial (perpendicular to the ring) and equatorial (in the plane of the ring). QuickTime™ and a TIFF (Uncompressed) decompressor are needed to see this picture. Figure 2-9. Chair conformations of glucose, mannose, and galactose in their pyranose forms. The chair is the most stable conformation of a six-membered ring. (In an alternative form, called the boat, both carbon 1 and carbon 4 lie above the plane of the ring.) The four bonds at each of the ring carbon atoms are tetrahedral. As shown in the generalized pyranose ring at the top left, bonds that extend nearly perpendicular to the plane of the ring are said to be axial (a); those that extend nearly parallel to the ring are said to be equatorial (e). In α-D-glucopyranose, all the hydroxyl groups except the one bonded to carbon 1 are equatorial. In α-D-mannopyranose, the hydroxyl groups bonded to carbons 1 and 2 are axial. In α-D-galactopyranose, the hydroxyl groups bonded to carbons 1 and 4 are axial. Note that, as in Figure 2-8, the hydroxyl groups with orientations different from those in glucose are highlighted. The L isomers of sugars are virtually unknown in biological systems except for L-fucose. One of the unsolved mysteries of molecular evolution is why only D isomers of sugars and L isomers of amino acids were utilized, and not the chemically equivalent L sugars and D amino acids. α and β Glycosidic Bonds Link Monosaccharides In addition to the monosaccharides discussed above, two common disaccharides, lactose and sucrose, occur naturally (Figure 2-10). A disaccharide consists of two monosaccharides linked together by a C—O—C bridge called a glycosidic bond. The disaccharide lactose is the major sugar in milk; sucrose is a principal product of plant photosynthesis and is refined into common table sugar. In the formation of any glycosidic bond, the carbon 1 atom of one sugar molecule reacts with a hydroxyl group of another. As in the formation of most biopolymers, the linkage is accompanied by the loss of water. In principle, a large number of different glycosidic bonds can be formed between two sugar residues. Glucose could be bonded to fructose, for example, by any of the following linkages: α(1 → 1), α(1 → 2), α(1 → 3), α(1 → 4), α(1 → 6), β(1 → 1), β(1 → 2), β(1 → 3), β(1 → 4), or β(1 → 6), where α or β specifies the conformation at carbon 1 in glucose and the number following the arrow indicates the fructose carbon to which the glucose is bound. Only the α(1 → 2) linkage occurs in sucrose because of the specificity of the enzyme (the biological catalyst) for the linking reaction. Glycosidic linkages also join chains of monosaccharides into longer polymers, called polysaccharides, some of which function as reservoirs for glucose. The most common storage carbohydrate in animal cells is glycogen, a very long, highly branched polymer of glucose units linked together mainly by α(1 → 4) glycosidic bonds. As much as 10 percent by weight of the liver can be glycogen. The primary storage carbohydrate in plant cells, starch, also is a glucose polymer with α(1 → 4) linkages. It occurs in two forms, amylose, which is unbranched, and amylopectin, which has some branches. In contrast to glycogen and starch, some polysaccharides, such as cellulose, have structural and other nonstorage functions. An unbranched polymer of glucose linked together by β(1 → 4) glycosidic bonds, cellulose is the major constituent of plant cell walls and is the most abundant organic chemical on earth. Because of the different linkages between the glucose units, cellulose forms long rods, whereas glycogen and starch form coiled helices. Human digestive enzymes can hydrolyze α(1 → 4) glycosidic bonds, but not β(1 → 4) bonds, between glucose units; for this reason humans can digest starch but not cellulose. The synthesis and utilization of these polysaccharides are described in later chapters. SUMMARY Covalent bonds, which bind the atoms composing a molecule in a fixed orientation, consist of pairs of electrons shared by two atoms. Relatively high energies are required to break them (50 – 200 kcal/mol). In covalent bonds between unlike atoms that differ in electronegativity, the bonding electrons are distributed unequally. In such polar bonds, one end has a partial positive charge and the other end has a partial negative charge (see Figure 2-5). Most molecules in cells contain at least one chiral (asymmetric) carbon atom, which is bonded to four dissimilar atoms. Such molecules can exist as optical isomers, designated D and L, which have identical chemical properties but completely different biological activities. In biological systems, nearly all amino acids are L isomers and nearly all sugars are D isomers. Glucose and other hexoses can exist in three forms: an openchain linear structure, a six-member (pyranose) ring, and a five-member (furanose) ring (see Figure 2-7). In biological systems, the pyranose form of D- glucose predominates. The two possible stereoisomers of D-glucopyranose (the α and β anomers) differ in the orientation of the hydroxyl group attached to carbon 1. Glycosidic bonds link carbon 1 of one monosaccharide to a hydroxyl group on another sugar, leading to formation of disaccharides and polysaccharides. Many different glycosidic bonds are theoretically possible between two sugar residues, but the enzymes that make and break these bonds are specific for the α or β anomer of one sugar and a particular hydroxyl group on the other.