jcb25372-sup-0010-SupData-S1
advertisement

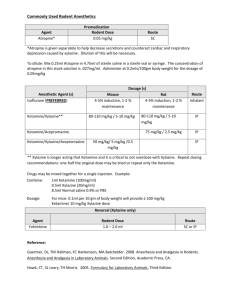
Supplemental Materials and Methods Cell growth curve- MDCK cells were seeded at a density of 2×104 per well in 12-well plates and cultured in regular DMEM. Cells were then serum-starved for 16 h, treated or not treated with ketamine (20 g/ml) and incubated at 37°C for 0, 12, 24, 36 and 48 h. After treatment, the cells were washed twice with ice-cold PBS and trypsinized with trypsin-EDTA (Gibco-BRL, Life Technologies, Grand Island, NY, USA). Cell suspensions were then mixed with trypan blue dye (Sigma-Aldrich, St. Louis, MO, USA) for cell counting using a hemocytometer under a microscope. The cell growth curves were calculated and established based on the numbers of viable cells using trypan blue exclusion. The trypan blue-stained cells were classified as damaged or dead cells. Supplemental Figure Legends Supplemental Fig. 1. Ketamine modulates EPC-related protein expression in MDCK cells. MDCK cells were treated with different concentrations of ketamine (0-20 μg/ml) for 24 h, and total cell lysates were collected for immunoblot analysis using antibodies against epithelial junction proteins, including ZO-1 (A), Occludin (B), and E-cadherin (C), as well as to detect the mesenchymal marker proteins N-cadherin (D), Fibronectin (E) and Vimentin (F). A set of representative immunoblots is shown in Figure 4B. The density of each band was measured using a densitometer with normalization to the loading control (α-tubulin). Quantification values were further calculated and expressed as the ratio to vehicle (mean±SD) for three independent 1 experiments. ** P<0.01 compared with the vehicle group. Supplemental Fig. 2. Ketamine induces morphological changes and promotes cell proliferation in MDCK cells. (A) Cell morphology of ketamine-treated MDCK cells was analyzed by phase-contrast microscopy. Control cells were treated with vehicle. MDCK cells were seeded at a density of 1×104 per well on 6-well plates and cultured in regular DMEM. Cells were then serum-starved for 16 h and treated or not treated with ketamine (20 μg/ml) in DMEM containing 0.5% FBS for 24, 48 and 72 h. Cell morphology was visualized on a phase-contrast microscope. In addition, serum was utilized as a positive control to show the epithelial phenotypic changes of MDCK cells. MDCK cells were serum-starved for 16 h and then exposed to 10, 20, 30, 40 and 50% serum for 24, 48, and 72 h. The results showed that 24 h after treatment, serum concentrations of 20% or higher induced morphological changes in MDCK cells, with reduced tightness of cell-cell contacts and roundish or spindle-like shapes. Forty-eight hours after serum exposure, serum concentrations of 20% or higher dramatically altered the MDCK phenotypes, leading to the loss of cell polarity and loose cell-cell contacts. These data indicate that the effects of serum on the phenotypic changes of MDCK cells are dose- and time-dependent. Interestingly, our results further showed that 72 h of treatment with 20 μg/ml ketamine also induced morphological changes in MDCK cells that were similar to the effects of serum on cell morphology. These results suggest that, like serum, ketamine can induce epithelial phenotypic changes in MDCK cells. Images are representative of at least three independent experiments. Bar, 100 μm. (B) Analysis of the effects of ketamine on the growth of MDCK cells. MDCK cells were seeded at a density of 2×104 per well in 12-well plates and cultured in regular DMEM. Cells were then serum-starved for 16 h and treated or not treated with ketamine (20 g/ml) for 0, 12, 24, 36 and 48 h. For cell number measurement, 2 cells were trypsinized and counted at each time-point using a trypan blue exclusion assay and a hemocytometer under a microscope. Each experiment was performed in triplicate. The data are presented as the mean±SD of three independent experiments. ** P<0.01 compared with the vehicle group at the corresponding time-points. The results showed that ketamine had no significant effect on MDCK cell proliferation before or at 24 h after the treatment and significantly induced cell proliferation after 36 h or longer treatment. The data indicate that ketamine exhibits an inductive effect on the proliferation of renal distal tubular cells. Supplemental Fig. 3. Ketamine induces the phosphorylation of FAK, Akt and GSK-3 and increases the protein levels of GSK-3downstream targets (-catenin and Cyclin D1) in MDCK cells. (A) Quantification of FAK, Akt and GSK-3β phosphorylation status in ketamine-treated MDCK cells from Figure 6A. Cells were treated with the indicated concentrations of ketamine (0-20 μg/ml) for 3 h or with ketamine (20 μg/ml) for the indicated times. Total cell lysates were collected for western blot analysis with anti-p-FAK, anti-FAK, anti-p-Akt, anti-Akt, anti-p-GSK-3, and anti-GSK-3β antibodies. A set of representative immunoblot images was shown in Figure 6A. Quantification of p-FAK, p-Akt, p-GSK-3β and their corresponding total protein levels was performed using a densitometer with normalization to their total protein levels. The levels of FAK, Akt and GSK-3β phosphorylation were further calculated and expressed as ratios with respect to vehicle. The results were calculated from three independent experiments and showed that ketamine significantly induced the phosphorylation of FAK, Akt and GSK-3β in a dose-response manner. Moreover, ketamine increased the phosphorylation of FAK, Akt and GSK-3β as soon as 3 h after treatment with 20 μg/ml ketamine and reached a plateau. (B) Immunoblot analysis of GSK-3 downstream targets in ketamine-treated MDCK cells. Cells were treated with 3 different concentrations of ketamine (0-20 μg/ml) for 24 h, and total cell lysates were collected for immunoblot analysis using antibodies against GSK-3 downstream targets including -catenin and Cyclin D1. The density of each band was measured using a densitometer and normalized to the loading control (α-tubulin). Quantification values were calculated and expressed as ratios relative to vehicle. All data are presented as the mean±SD of three independent experiments. * P<0.05; ** P<0.01 compared with the vehicle group. The results showed that ketamine induced β-catenin and Cyclin D1 protein levels in MDCK cells, indicating that ketamine can affect GSK-3β activity, leading to increased levels of β-catenin and Cyclin D1 protein. Supplemental Fig. 4. Ketamine induces GSK-3β phosphorylation and modulates EPC-related protein expression in MDCK cells. (A) Immunoblot analysis of Akt and GSK-3β phosphorylation levels, as well as c-Myc protein levels in ketamine-treated MDCK cells in the presence or absence of PI3K/Akt or GSK-3β inhibitor. MDCK cells were pretreated with 2.5 M of a PI3K/Akt inhibitor, LY294002, or 5 M of a GSK-3β inhibitor, 3F8, for 1 h and then treated or not treated with ketamine (20 μg/ml) for 3 h. Total cell lysates were collected for western blot analysis with anti-p-Akt, anti-Akt, anti-p-GSK-3, anti-GSK-3β, anti-c-Myc and anti-α-tubulin antibodies. The experiments were independently performed three times. Quantifications of p-Akt, p-GSK-3β, their corresponding proteins and c-Myc protein levels were performed using a densitometer. Akt and GSK-3β phosphorylation levels were further normalized to their total protein levels, while the levels of c-Myc were normalized to the loading control -tubulin. The results were further calculated and expressed as ratios relative to vehicle. The results showed that ketamine significantly induced the phosphorylation of Akt and GSK-3β, as well as the protein level of c-Myc in MDCK cells. Inhibition of PI3K/Akt signaling by LY294002 blocked the effect of ketamine on Akt and GSK-3β 4 phosphorylation and c-Myc expression. Moreover, GSK-3β inhibition by 3F8 increased c-Myc protein levels, similar to the results of ketamine treatment. A combination of ketamine and 3F8 had no additive effect on Akt and GSK-3β phosphorylation levels or on c-Myc expression. Taken together, these results indicate that ketamine can induce the activation of PI3K/Akt/GSK-3β signaling in MDCK cells. (B) Quantification of the immunoblot results from the examination of the effect of PI3K/Akt or GSK-3β inhibition on the levels of ketamine-modulated EPC-related proteins. MDCK cells were pretreated with 2.5 M of the PI3K/Akt inhibitor LY294002 or 5 M of the GSK-3β inhibitor 3F8 for 1 h and then treated or not treated with ketamine (20 μg/ml) for 24 h. Total cell lysates were collected for immunoblot analysis with anti-ZO-1, anti-Occludin, anti-E-cadherin and anti-Vimentin antibodies. A set of immunoblot images was shown in Figure 6B. The density of each band was measured using a densitometer with normalization to the loading control (α-tubulin). The quantification results were further statistically calculated and expressed as ratios relative to vehicle. All data are presented as the mean±SD of three independent experiments. ** P<0.01 compared with the vehicle group; ## P<0.01 compared with the ketamine group. The results showed that ketamine significantly reduced the levels of junctional proteins (ZO-1, Occludin and E-cadherin) and increased the level of the mesenchymal protein Vimentin. Inhibiting PI3K/Akt signaling with LY294002 suppressed the effects of ketamine on the EPC-related proteins. Interestingly, treatment with a selective GSK-3β inhibitor, 3F8, reduced ZO-1, Occludin and E-cadherin protein levels and enhanced Vimentin protein levels, similar to ketamine treatment. The 3F8 treatment had no additive effect on ketamine-suppressed ZO-1, Occludin and E-cadherin, or ketamine-induced Vimentin, suggesting a common mechanism of action through GSK-3β inactivation in ketamine signaling to induce EPCs in renal distal tubular cells. 5 Supplemental Fig. 5. Examination of the effects of the well-known GSK-3β inhibitor LiCl on Akt and GSK-3β phosphorylation levels, EPC-related protein levels, and TEER in MDCK cells. (A) Immunoblot analysis of the effects of the GSK-3β inhibitor LiCl on Akt and GSK-3β phosphorylation and protein levels in ketamine-treated MDCK cells. MDCK cells were pretreated with 10 mM LiCl for 1 h and then treated or not treated with ketamine (20 μg/ml) for 3 h. Ten millimolar NaCl was applied to MDCK cells as a negative control to exclude ionic strength or osmotic effects. Total cell lysates were collected for western blot analysis with anti-p-Akt, anti-Akt, anti-p-GSK-3 and anti-GSK-3β antibodies. The experiment was independently performed three times, and a representative blot is shown in the upper panels. p-Akt and p-GSK-3β levels were quantified using a densitometer with normalization to their corresponding total protein levels. The results were further calculated and expressed as ratios relative to vehicle and showed that LiCl increased GSK-3β phosphorylation levels with no effect on Akt. NaCl exhibited no significant effect on either Akt or GSK-3β, suggesting that the ionic strength or osmotic effect of LiCl does not play a significant role in the increased phosphorylation levels of GSK-3β. (B) Examination of the effects of the GSK-3β inhibitor LiCl on ZO-1, Occludin, E-cadherin and Vimentin protein levels in ketamine-treated renal distal tubular cells. MDCK cells were pretreated with 10 mM LiCl for 1 h and then treated or not treated with ketamine (20 μg/ml) for 24 h. Ten 6 millimolar NaCl was applied to treat MDCK cells as negative control to exclude ionic strength or osmotic effects. Total cell lysates were collected for immunoblot analysis with anti-ZO-1, anti-Occludin, anti-E-cadherin and anti-Vimentin antibodies. The density of each band was analyzed using a densitometer with normalization to the loading control (α-tubulin). The experiment was performed independently three times. Quantification values were further statistically calculated and expressed as ratios relative to vehicle. The results showed that GSK-3β inhibition with LiCl significantly reduced the levels of junctional proteins (ZO-1, Occludin and E-cadherin) and increased the level of the mesenchymal protein Vimentin, similar to the effects of ketamine on these proteins. Again, NaCl had no significant effect on these proteins, suggesting that the LiCl-altered levels of these EPC-related proteins were not due to ionic strength or osmotic effects. (C) Examination of the effects of LiCl on the TEER of polarized MDCK cells in the presence or absence of ketamine. A polarized MDCK monolayer on a filter was pretreated with 10 mM LiCl for 1 h and then treated or not treated with ketamine (20 µg/ml) on both the apical and basolateral surfaces for 24 h. TEER was then measured using a Millicell-ERS Volt-ohm Meter resistance apparatus. Ten millimolar NaCl was used to treat MDCK cells as negative control to exclude ionic strength or osmotic effects. TEER values were calculated statistically with normalization to vehicle cells. All data are presented as the mean±SD of three 7 independent experiments. ** P<0.01 compared with the vehicle group. The results showed that GSK-3β inhibition with LiCl exhibited a similar effect as ketamine in reducing the TEER values of polarized MDCK cells, and NaCl had no effect on the TEER values of the polarized cells. Taken together, these results indicate that the inhibition of GSK-3β by LiCl can promote the EPC of renal distal tubular cells by altering junctional proteins and increasing the permeability of polarized cells. LiCl-induced EPC is apparently not due to the ionic strength or osmotic effect of this compound. Supplemental Fig. 6. Ketamine induces proinflammatory cytokines production via PI3K/Akt/GSK-3β signaling in MDCK cells. MDCK cells were pretreated with 2.5 μM of the PI3K/Akt inhibitor LY294002 for 1 h and then treated or not treated with ketamine (20 μg/ml) for 24 h. Otherwise, cells were exposed to 5 μM of the GSK-3β inhibitor 3F8 for 24 h. The mRNA levels for TNF-, IL-6 (B) and TGF-1 (C) in response to individual treatment was measured using quantitative RT-PCR. The data were normalized to vehicle cells and are presented as the mean±SD from at least three independent experiments. ** P<0.01 compared with the vehicle group; ## P<0.01 compared with the ketamine group. 8