Food web analysis with ecopath/ecosim
advertisement

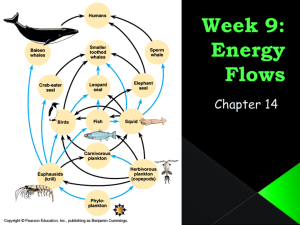
Introduction to multispecies modeling using Ecopath with Ecosim Software for examining impacts of trophic interactions and harvesting on multispecies biomass patterns in Ecosystems Approaches to multispecies/ecosystem modeling • Static community “types” modeling in relation to habitat factors • Empirical assessments of linkage “strength” (bottom up vs top down effects, etc) • Single species models with mortality rate crosslinkages (MSVPA,MSFOR) • Biomass/numbers trophic interaction models (Ecopath/Ecosim) • Agent-based models with arbitrary components and linkages (ATLANTIS) • Complex, spatial multispecies IBMs Multispecies fisheries issues • Almost all fisheries are “multispecies” in sense that gear takes more than one species • Usual to classify multispecies fisheries interactions as: – “Technical”—fishing targeted at one or more species usually takes others as bycatch – “Ecological”—fishery appropriates production that could otherwise go to higher trophic level species, but frees up production of species at lower trophic levels Trophic level Species can be positioned in food webs by mean trophic level of food consumed 4 Piscivores 3 Planktivores and benthivores 2 Invertebrates • Trophic level pattern creates possibility of “trophic cascades”: decrease at 4 causes increase at 3 causes decrease at 2, etc. • But web structure weakens such effects Mixed technical and trophic interactions Fishery 1 Trophic level 4 Fishery 2 3 Fishery 3 2 • Fishery 1 creates value tradeoffs among species at TL 4, and increase in surplus production at TL 3 • Fisheries 2, 3 may “rob” surplus production from Fishery 1 • Fishery 2 may deplete both the piscivore(s) and their prey Trophic level Trophic ontogeny creates risk of “cultivation-depensation” effects 4 Piscivores 3 Planktivores and benthivores 2 Invertebrates • Piscivores typically increase in trophic level as they grow • High abundance of adult piscivores causes reduction in abundance of lower TL species that compete with and prey upon piscivore juveniles • Reduction in adult piscivores can lead to reduced survival of its juveniles, lower equilibrium dominated by TL 2-3 species (examples: bass-bluegill, cod-herring) Example fisheries questions involving food web interactions • Does the Atlantic menhaden fishery impact production of striped bass? • Have Bering Sea fisheries caused collapse of Stellar sea lions? • Are seals and sea lions causing collapse of Pacific salmon populations? • Will bycatch reduction devices in Gulf of Mexico shrimp fisheries result in decreased shrimp production due to increases in shrimp-eating fish that have been fished down by trawling? • Is food production limiting to rainbow trout in Grand Canyon, and do rainbow trout have strong negative impact on native fish recruitment? Ecopath with Ecosim (EwE) is the only general modeling software now available for dealing with such questions • Can be parameterized with relatively simple data • Can represent complex food web structure, multiple fisheries with complex targeting and bycatch impacts • Incorporates “foraging arena” behavioral effects in trophic interaction predictions • Allows representation of trophic ontogeny and cultivation-depensation risks EwE main components and data entry/analysis steps • ECOPATH—Enter basic biomass and trophic interaction information, output is set of biomasses Bi and “flows” Qij from biomass pools i to j. • ECOSIM—Predict biomass (and age structure) changes over time starting from Ecopath base values • ECOSPACE—Predict biomass changes over spatial grid of habitat cells over time, starting with ECOSIM parameter estimates Think about biomasses Bi and flows Qij between them as a web Catch • Qij is a rate (biomass per year) • Total consumption by a predator is a sum of Qs, eg Q24+Q34 • Prey mortality rate on any link is Mij=Qij/Bi • Fishing rate is Fi=Ci/Bi C5 B5 B4 Q34 Q35 B3 Q24 B2 Q13 Q12 B1 ECOPATH MODEL DEVELOPMENT • Define a set of biomass “pools” or functional groups that are of policy interest • Enter estimates of three of the following four quantities for each group: – – – – Bi-biomass per area PBi- annual production per biomass QBi-annual consumption per biomass EEi-proportion of production explained by modeled predation and harvest • Enter estimates of DCij, proportions of species i in diets of species j (food web structure) Ecopath “solution” • Given 3 of the 4 basic inputs for each group i, and the diet composition matrix DCij, Ecopath solves for the fourth quantity for each i, and for the trophic flows Qij • Qij=QBj x Bj x DCij (Total consumption by j x DCij) • • (Consumption of j Catch of j) EEj (T otalMortalityof j BA of j) EEj is a critical measure of “mass balance”; EE>1.0 is physically impossible and indicates some error in the inputs used to calculate it Defining biomass pools or “groups” • There is no “right” way to do this; more groups (finer splitting) does not imply “better” model • Start simple, examine effect of adding complexity • Two approaches to defining pools – “Partition” whole ecosystem biomass – “Selection” of groups of particular interest (Fulton: selection approach is “best”) Estimating production PB • Definition of P: ΔB = Production - Mortality • This means Production= ΔB + Mortality • If the initial B’s are assumed to be near an “equilibrium” (B’s changing slowly), ΔB=0 and Production=Mortality • If we express production and mortality as specific rates x biomasses, this means PBxB=ZB, i.e. PB=Z where Z is an estimate of total instantaneous mortality rate of B • Over complex multispecies/age “groups”, need to estimate PB as a weighted average of PB components PBa, each weighted by its Ba/B. Estimating Consumption QB • Gastric evacuation: QB=(gut wt)x(guts/yr) • Bioenergetics: backcalculate QB from observed weight changes and estimated efficiencies and metabolic losses • Temming method assuming vonB growth – Q=(dW/dt+3KW)/e – K=vonBertalanffy growth K – e=0.5-0.6 -QB=sum over ages of Qage x Nage, divided by biomass Ecotrophic Efficiency EE i M ij Fi j PB i Note here that PBi is usually an input (total mortality rate+biomass accumulation rate), while Mij=Qij/Bi and Fi=Catchi/Bi are calculated from other inputs PB,QB,DC What to do when your model does not “balance”, i.e. EEi>1.0 implies inconsistency in input values • Increase estimated biomass Bi • Increase estimated PBi (total mortality rate Zi) • Reduce DCij for one or more predators j • Reduce QBj for one or more predators j • Make BAj less negative Other ECOPATH inputs for more complex models • • • • Immigration and emigration rates Biomass accumulation rates (dB/dt) Detritus “fate” (partitioning of available B) Partitioning of catch into landings, discards for multiple fishing fleets—>Fik, k=fleet • Multistanza population dynamics Nearshore rockfish 4+ 1+ 0 Log Numbers at age Multistanza, age-structured life history representation in Ecosim and Ecospace Cascading bottleneck effects Weight at age Shift from density dependent mortality to density dependent growth Age (months) Each stanza (range of ages) can be assigned distinctive: 1) Total mortality rate Z, varying with stanza-specific predation rates 2) Prey and habitat preferences (diet composition, distribution) 3) Behavioral tactics: respond to changes in food availability by changing growth rate and/or activity and associated predation risk 4) Vulnerability to fishing and bycatch Simulating time dynamics: ECOSIM • Simple biomass pools simulated over time by integrating the rate equation dBi/dt=ei∑Qji-∑Qij-Fi-Mi where changes in Qij are predicted with the foraging arena equation Qij=aijvijBiBj/(2vij+aijBj) • Multistanza populations simulated with agestructured equations for numbers N, body weights W at age: Ni,a+1,t+1=Ni,a,te-Zi Wi,a+1,t+1=ki1aQi/Bi+ki2aWi,a,t • These equations are all solved on one-month time steps Estimating trophic interaction rate parameters for Qij predictions • The a’s and v’s of Qij=aijvijBiBj/(2vij+aijBj) are estimated in two steps: • First, assume that the maximum possible consumption rate vijBi is a multiple κij of the Ecopath base rate MijBi: vij= κijMij (user must specify κij) • Next, calculate rate of effective search aij by solving the foraging arena equation for aij with Qij set to Ecopath base value and vij set to κijMij. • This approach forces the dynamic model to predict the Ecopath base rates whenever all the Bi are at Ecopath base values. Estimation of the critical κij vulnerability multipliers • These determine basic model stability and diversity properties • Κij >1.0, and depends on initial predator biomass Bj relative to “natural” maximum as well as on foraging arena behaviors. • For species j that have been depressed severely by fishing (low Ecopath Bj), must set κij very high (>10) to predict recovery • Very low κij (near 1.0) often work best for links involving juvenile predators • Can sometimes estimate by fitting to time series data • Play same role as recruitment compensation ratios, but in reverse (low κij implies high compensation ratio); often difficult to estimate for same reasons (lack of contrast in historical data) Time forcing in Ecosim • The basic rate equation dBi/dt=ei∑Qji-∑Qij-Fi-Mi is parameterized so as to predict no change (dB/dt=0) unless Ecopath biomass accumulation rates are nonzero. • Dynamic change can be caused by forcing variation over time in 1. Fishing mortality rates Fi 2. Primary production rates PBk 3. Trophic interaction parameters a,v • Patterns of forcing variation can be imposed by: 1. Sketching changes in the user interface, or 2. Using Excel .csv time series data files Dynamic response in B’s is dominated by assumptions about the vulnerability multipliers κij • High κij values (e.g. 10) imply weaker limitation of predation effects through foraging arena behaviors, stronger interaction effects • Low κij imply less variation in Mij with changes in predator abundance, and also stronger “compensatory” changes in predator per-capita consumption rates ∑Qij/Bj Adding realism to a,v predictions in the foraging arena equation • Foraging time adjustment: a’s and v’s can be varied so as to simulate changes in feeding time aimed at keeping food intake and growth constant (causes destabilizing type II response) • Risk-sensitive foraging (indirect trait mediated effects): foraging time can be made inversely related to predation risk. • Handling time limitations: type II effects caused by high handling times per prey • Prey switching: rates of effective search aij can be made to vary positively with prey biomasses Bi to simulate processes like search image formation and choice of alternative foraging modes by predators • Trophic mediation: effects of other Bs on vulnerability Main ECOSIM capabilities • Simple time scenarios with sketched forcing patterns • Fishing efforts varied dynamically with bioeconomic response submodel • Fitting to time series data in .CSV files by varying κij (“stock assessment” methods) • Optimization of fishing efforts by fleet and over time using search algorithms • Stochastic simulations with randomly varying ECOPATH parameters (sensitivity analysis) Evaluation of spatial policies: ECOSPACE • Consists of a map grid of Ecosim models linked through dispersal/migration rates and movement of fishing effort • Each map grid cell has user-defined habitat type, each group uses one or more types (has high predation rate, low feeding rate in habitat types that are not suitable) • Mainly useful for evaluation of spatial closure (MPA) policies; NOT a next step or improvement on Ecosim for examining dynamic change The Ecospace basemap • Map layers include habitat type, relative primary productivity, MPA type, relative fishing cost • Can generate from global bathymetry and primary productivity database, by web link to UBC Sea Around Us Project server • More often, simply define map area, resolution and enter habitat type data using the Ecospace user interface manually or using copy/paste from data in Excel csv files Key Ecospace inputs for each biomass group • Which habitats does it use? • How much does it move, and how much does it suffer when in unsuitable cells? • What is its seasonal migration pattern? • How do its movement rates respond to local variations in predation risk and food availability? Ecospace solution options • • Ecospace time predictions involve solving very large system of differential/difference equations, m x n x p of them (m=rows, n=columns, p=groups), plus spatial effort predictions Three solution options: 1. Simple biomass equations (fast) 2. Biomass equations+aggregate age-structure dynamics for multistanza populations (slower) 3. IBM packet solutions for multistanza populations (much slower) (Avoid option 1 for models with multistanza groups) Linking Ecospace with hydrodynamic models • Ecospace can read and use some spatial and temporal outputs from physical models • These outputs include – Advection fields (surface current patterns) – Total nutrient concentration fields – Salinity fields • NB: the idea here is that physics affects biology but not the reverse, so can first run a physical model then force the biology calculations with its results