Lesson 7 Hydrophilic signalling

Hydrophilic signal molecules
Hydrophilic signal molecules such as peptide hormones and neurotransmitters .
These cannot pass through a cell membrane and must activate the surface receptor proteins
Once activated the surface receptor protein generates an intracellular response
This process is called signal transduction
The mechanism by which hydrophilic extra-cellular molecules such as peptide hormones generate an intracellular response
Endocrine cell
Target cell
Receptor
Bloodstream
Hormones , such as peptide hormones , are produced in endocrine glands , secreted into the bloodstream and carried throughout the body.
These signal molecules only produce a response in target molecules with the appropriate surface receptor.
This type of cell signalling is used by the body to coordinate the bodies metabolism and causes relatively slow, long lasting changes. Important peptide hormone include insulin and glucagon
A second group of hydrophilic signalling molecules are neurotransmitters
An electrical signal is passed along a nerve and on reaching the terminal point stimulates the release of neurotransmitter signalling molecules.
These diffuse across the gap between nerves known as a synapse and lock onto receptors found on the surface of the nearest nerve generating an electrical impulse
Neurotransmitters produce a fast acting , short lived response between nerves
The mechanism by which hydrophilic extra-cellular molecules such as neurotransmitters generate an intracellular response
Neuron consists of dendron + cell body + axon
Dendron Cell body Axon
Target cell
Direction of nerve impulse
Receptor
Synapse showing movement of neurotransmitter molecules
When activated by signals from the surroundings, or other nerve cells, the neuron sends electrical impulses along its axon at speeds of up to 100 meters/second. On reaching the axon terminal, the intracellular electrical signals are converted to an extra-cellular form: each electrical impulse stimulates the terminal to secrete a pulse of chemical signal called a neurotransmitter. Neurotransmitters diffuse across the narrow gap, known as a synapse, and bind to receptors on the surface of the target cell
In both cases above the extracellular signal molecule binds to cell surface receptors as the hydrophilic molecules cannot cross the lipid bilayer. These receptors act as transducers which convert the signal on the outside of the cell to an intracellular signal
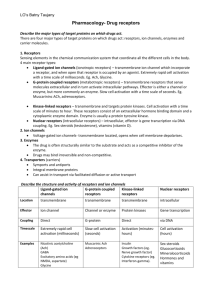
There are three main classes of cell-surface receptors
Three types of signal transduction mechanism:
ion channels enzyme-linked G-protein-linked
(a) (b) (c)
S
+
+
+ +
+
+ +
+ +
+
+
+
+
+
+
+
+
+
+
+
+
+
+
+
+
+ +
+
+
closed open inactive active
R
G
Transport ions rapidly across membranes. Very important in
Muscles and nerves
Extracellular signal binds to inactive form and activates the enzyme function at the cytosol side
G
T response
G-linked protein actives
G-protein which in turn starts sequence of intracellular events
Class1 - Ion-channel receptors.
These are found on the surface of muscles and nerves and tranduce a signal in the form of a neurotransmitter into an electrical voltage.
Class 2 - G-protein-linked receptors . This is the largest group.
G-protein-linked receptors activate a G-protein which sets off a chain of events within the cell. These are found in all cells
Class 3 - Enzyme linked receptors . An enzyme linked receptor binds an extracellular signal molecule switching on an enzyme activity, usually a kinase . on the other side of the membrane. This kinase activity causes the phosphorylation of other intracellular proteins. These are found in all cells
G-protein-linked receptors
(a) (b) signal
AC
GP
GDP
The peptide hormone glucagon sets off a chain of reactions as follows:-
• Glucagon molecule binds to G-linked protein other
• Inactive G-Protein is switched on by addition of phosphate to GDP effects
• Activated G protein binds to enzyme adenylate cyclase (AC)
• Enzyme AC breaks down ATP to cyclic AMP
• Cyclic AMP causes intracellular effect e.g. breakdown of glycogen or fats or activates gene regulatory proteins which switch on genes
AC
GP
GTP
ATP cAMP