chapter_22
advertisement

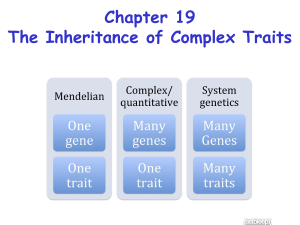
Chapter 22 - Quantitative genetics:
•
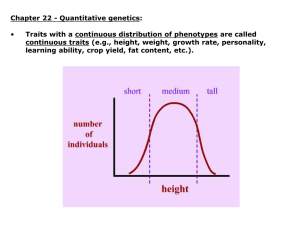
Traits with a continuous distribution of phenotypes are called
continuous traits (e.g., height, weight, growth rate, personality,
learning ability, crop yield, fat content, etc.).
Continuous traits arise from effects of:
•
Multiple loci
•
Pleiotropy (one gene has many effects)
•
Epistasis
•
Variable expressivity and penetrance
•
Environment (produces a range of phenotypes)
Examples of different distributions:
Lognormal
Normal
Exponential
Continuous traits:
•
Francis Galton and Karl Pearson (late 1800s):
Recognized that continuous traits are statistically correlated
between parents and offspring, but could not determine how
transmission occurs.
•
Wilhelm Johannsen (1903):
Demonstrated that bean seed weight is partly heritable and partly
environmental.
•
Sir Ronald Fisher:
First to demonstrate mathematically that Mendelian models of allele
segregation apply to multiple genetic loci.
Types of questions studied in quantitative genetics:
•
How do genetics and the environment affect a trait?
•
Which and how many genes produce a set of phenotypes for a trait;
where in the genome are they located?
•
Do some genes play a major role, whereas other genes modify or
play a small role?
•
Do alleles interact to produce additive effects?
•
How does selection affect the trait; does it affect other traits?
•
What types of mating and selection produce desired phenotypes?
Types of data collected and analyses to consider:
•
Sample size/randomization
•
Type of distribution (e.g., normal distribution)
•
Mean, variance, and standard deviation
•
Correlation/regression
•
Analysis of variance (ANOVA)
Quantitative trait loci (QTLs):
QTLs =specific genomic segments correlated with continuous
phenotypic trait variation.
Perform Genome Wide Association Study (GWAS)
1.
Cross inbred lines with different phenotypes (homozygotes for
different alleles at most loci) to produce heterozygotes.
2.
Self F1 or cross to parental lines to increase phenotypic variation
and segregation of traits.
3.
Analyze F2 with physical markers (microsatellites) that correlate
with phenotypic variation.
4.
Create a linkage map.
5.
Calculate components of phenotypic variance (VP) due to genetic
effects (VG) and components due to environment effects (VE).
VP = VG + VE + 2COVG,E + VG x E
Components of genetic variance:
1.
Additive genetic variance (VA): effects of alleles at two or more loci
contribute to phenotype. F1 will will appear intermediate to the
parental phenotypes for repeated test crosses.
2.
Dominance variance (VD): effects of alleles are not strictly additive;
must consider how alleles interact in the heterozygote. F1 will
resemble one of the parental phenotypes.
3.
Interaction variance (VI): accounts for epistatic interactions
between two or more loci. F1 phenotype is unpredictable.
VG = VA + V D + V I
VP = (VA + VD + VI) + VE + 2COVG,E + VG x E
Calculate Lod Score:
1.
Lod = log of the ratio odds that two loci (or a locus and a trait) are
linked with a recombination factor (q) greater than 0 and less than
0.5.
2.
z = log10 {Prob(data|q)/Prob(data|0.5)}
3.
Lod score of +3.00 (odds of 1000:1) or greater is regarded as
acceptable evidence for linkage.
Figure 1. Graph of multipoint lod scores assuming heterogenity
The peak multipoint lod score of 3.85 is located between DXS1200 and DXS297.
Nature Genetics 20, 175 - 179 (1998)
Evidence for a prostate cancer susceptibility locus on the X chromosome.
Jianfeng Xu et al.
Timmerman-Vaughan et al. 2004 - Linkage mapping of QTLs for seed yield, yield components
and developmental traits in pea (Pisum sativum L.)
Chromosome map of human QTLs for plasma concentrations of HDL-C, LDL-C and triglyceride
levels. 2007. The Jackson Laboratory
Genome Scan
•
Search for islands of genetic differentiation in otherwise
undifferentiated genetic background.
•
Alternative method for searching for genes underlying functionally
important traits.
•
Does not require crossing experiment, but rather perform genome
scan (e.g., next-generation sequencing) for two populations that
differ in a single environmental variable subject to strong selection.
•
Works best for two populations that are in migration-selection
balance equilibrium experiencing strong divergent selection and high
gene flow.
•
Utilize measures such as Fst
Alternate interpretations of outliers.
Via S Phil. Trans. R. Soc. B 2012;367:451-460
©2012 by The Royal Society
Relationship between the maximum FST that can be maintained by DH at equilibrium and the
map distance from a selected gene, for two intensities of divergent selection (modified from
[27], for a population with Ne = 1000 and m = 0.001).
Via S Phil. Trans. R. Soc. B 2012;367:451-460
©2012 by The Royal Society
Probable DH regions in threespine stickleback.
Via S Phil. Trans. R. Soc. B 2012;367:451-460
©2012 by The Royal Society
Broad- and Narrow-Sense Heritability:
1.
Broad-sense heritability
= hB2 = VG/VP
2.
Narrow-sense heritability
= hN2 = VA/VP
*Broad-sense heritability measures proportion of phenotypic variance
among individuals in a population that results from genetic
differences.
*Narrow-sense heritability measures proportion of phenotypic variance
that results from additive genetic variance.
*Narrow sense heritability is what can be used to predict resemblance
between offspring and parents.
*Heritability is a measure of variance and is only meaningful for
characteristics of a population (not the individual).
Example showing how to calculate narrow-sense heritability using
parent-offspring regression:
Example showing response to selection in artificial experiment:
h2 = R/S