fgfgrc022814 - Texas A&M University
advertisement

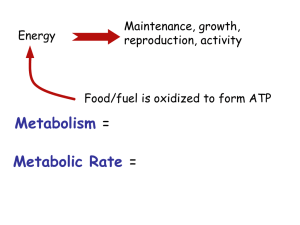
Forty Years of FGF: Regulator of Cellular & Metabolic Homeostasis Wallace McKeehan Center for Cancer & Stem Cell Biology F HSPG F PI3K AKT sos The Klothos Grb2sos Ras PKC Raf MEK ? ERK1/2 Gordon Sato, The Mangrove Man Gospodarowicz D, Jones KL, Sato G. 1974 Purification of a growth factor for ovarian cells from bovine pituitary glands. PNAS 71:2295 Gospodarowicz D. 1975 Purification of a fibroblast growth factor from bovine pituitary. JBC 250:2515 Maciag T, Mehlman T, Friesel R, Schreiber AB. 1984. Heparin binds endothelial cell growth factor, the principal endothelial cell mitogen in bovine brain. Science 31;225:932. Gospodarowicz D, Cheng J, Lui GM, Baird A, Böhlen P. 1984. Isolation of brain fibroblast growth factor by heparin-Sepharose affinity chromatography: identity with pituitary fibroblast growth factor. PNAS 81:6963. 1985-1986: Cloning of FGF1 and FGF2 1989-1993: Cloning and diversity of FGFR tyrosine kinases via splice variants 1993-2002: Structure of FGFR kinases and interaction with heparan sulfate Structure, Mechanism of Assembly & Signaling FGF1 FGF2 FGF3 FGF4 FGF5 of the FGFR Complex Continued Discovery Through Homology & Genome Sequencing: FGF6 FGF7 FGF11 FGF12 FGF8 FGF9 FGF16 FGF17 FGF18 FGF10 22 FGF Homologues 18 High Affinity FGFR Activators Four Transmembrane Tyrosine Kinase Receptors Numerous Splice Variants: FGFR1-4 FGF20 FGF22 Blue: Positive Charge Red: Negative Charge White: Neutral FGF19 FGF21 FGF23 F F Yayon…Ornitz, Cell 1991 Xu…McKeehan, JBC 1992 Kan…McKeehan, Science 1993 Pantoliano…Sisk Biochem. 1994 F Spivak-Kroizman…Schlessinger 1994 Venkataraman…Sasisekharan, PNAS 1996 Moy…Powers, Biochem. 1997 F McKeehan, Wang, Kan, PNAR 1998 Kan…McKeehan, JBC 1996 F DiGabriele…Hendrickson, Nature 1998 Plotnikov…Mohammadi, Cell 1999 Kan…McKeehan, JBC 1996 McKeehan, Wang, Kan, PNAR 1998 Schlessinger….Mohammadi, Cell 2000 Conformational Model of Control and Activation (Derepression) of the FGFR Complex Pre-existent unliganded symmetric complex of 2:2 FGFR-HS Conformational maintenance of dependence on FGF for kinase derepression by transphorylation by heparan sulfate in the ectodomain Pelligrini…..Blundell, Nature 2000 Conformational transmembrane communication between ecto and intracellular domains How Can FGF-dependent Change Outside Be Transmitted Across the Membrane? Next Generation: Are Intracellular Substrates/Adaptors Organized and Waiting for Conformational Derepression? F HSPG F PI3K AKT sos Grb2 sos Ras PKC Raf MEK ? ERK1/2 Role of Heparan Sulfate COOO COOOH CH2OX O Linker O OX O OX IdoA/GlcA NY GlcN FGF Core protein HS proteoglycan n 2-O-, 2-N-, 3-O- and 6-O-sulfates, 2-N-acetyl and epimerization HS FGFR 1. Matrix and Membrane: Reservoir, Stabilizer, FGF Delivery System 2. Integral Component of the FGFR Complex: Specificity for FGF Assembly of the oligomeric complex Negative restriction of the unliganded dimeric complex Stabilization of the derepressed, ligand-activated complex Is There Structural Specificity in Heparan Sulfate Beyond Variations in Charge Density? 7,8-S-OctaF7, which has anticoagulant activity and likely the motif containing a 3-O sulfate, specifically supports high affinity FGF7 binding and mitogenesis in cells expressing FGFR2IIIb. It failed to support high affinity FGF1 binding and mitogenesis. Purification of an undersulfated 7,8-S-octasaccharide mixture (7,8-S-OctaF7) by affinity chromatography with FGF7 Hypothetical deduced structure of 7,8-S-OctaF7. The x, y or z could be a sulfate, preferably 6-O-sulfate with the other two as hydrogens (8-S), or x, y and z may all be hydrogens (7-S). The Klotho Co-factors: single pass transmembrane proteins involved in endocrine FGF activities αKlotho N (1014aa) βKlotho N (1043aa) SS αKl1 αKl2 βKl1 βKl2 C TM C Similarities to Heparan Sulfate (HS): Independent binding to eFGFs or FGFR Direct participation in the FGFR complex with both FGF and FGFR Determinants of specificity for eFGFs Are HS and Klothos at play in the same FGFR complex? Do HS or Klotho co-factors alter quality of the FGFR signal? Does the protein core of HS or the intracellular domain of HS or Klothos play a role in signaling? Development and Adult Homeostasis Modes of Function: Autocrine, Paracrine, Endocrine Development: Autocrine and Paracrine Control: Changing, Short-lived Cell Autonomy, Transcription rate-limiting Adult Tissue Homeostasis: Largely Paracrine Control: Partition of FGF and FGFR between Cells, Activity, not transcription rate-limiting Autocrine is Pathological Intracrine (FGF, FGFR or FGF-FGFR)? Is there tissue and target cell specificity among the 18 FGFs and the 4 tyrosine kinase receptors and variants? Specificity set by: 1. Paracrine partition of FGF and the FGFR complex between cells. 2. Endocrine partition of FGF and FGFR complex between organs. 3. Cell-specific co-receptors HS and klothos. Do different FGFR isotypes have exhibit different signaling endpoints in the same context? Subversion and Autocrine Switch of Canonical Matrix-controlled Short Range Paracrine FGF Signaling Results in Pathologies Epithelium Loss of R2b Ectopic FGFR1 Autocrine Switch Cancer FGR2b FGF 7 FGF10 FGF9 FGFR3 Heparan sulfate Stroma Feng, Wang, Matsubara, Kan, McKeehan. Fibroblast growth factor receptor 2 limits and receptor 1 accelerates tumorigenicity of prostate epithelial cells. Cancer Res. 1997 Overlapping and Distinct TyrP Targets Between: FGFR2IIIb PTPN18 CDK2 (Tyr15) Emerin ZRF1 LAP2 SAP102 FGFR1IIIc FGFR SHP2 SHIP2 ERK2 IRS4 FRS2 PLCλ growth/tumor suppression, nuclear-cytosol interplay, cell structural maintenance RSK2 Fyn FAK ShcA P85α P85β Paxillin growth promotion, cell survival, adhesion, motility Luo et al. Novel phosphotyrosine targets of FGFR2IIIb signaling. Cell Signal 2009 2000: First implication of FGF signaling in regulating metabolic circuits: Cholesterol/bile acid homeostasis A5 4 Hepatocyte FGFR4 +/+ -/- 3 2 1 0 24 48 72 96 120144168 B 9080 +/+ -/- 70 60 50 40 30 20 10 0 0 24 48 72 96 120144168 Time after partial hepatectomy (hr) Hepatocyte Cyp7a Cholesterol to Bile Acids Yu, Wang, Kan…McKeehan. Elevated cholesterol metabolism and bile acid synthesis in mice lacking membrane tyrosine kinase receptor FGFR4. JBC 2000 Evolution of the Endocrine FGFS The FGF15/19-FGFR4 Axis: An enterohepatic regulator of cholesterol/bile acid homeostasis Early 2000: FGFR4 involved in cholesterol/bile acid metabolism. Yu et al. JBC 2000 Late 2000: Mutation in FGF23 involved in mineral metabolism. ADHR Consortium, Nat Genetics 2000 2002: FGF19 impacts metabolic rate and adiposity. Tomlinson et al., Endocrinol 2002 2005: FGF21 is a metabolic regulator Kharitonenkov et al., JCI 2005 Inagaki et al., Cell Metab. 2005 2005: Klothos impact metabolism & FGF signaling Steroids FGF21 effectors have expanded! Steroid receptors Paracrine FGFs FGF7,9,10, others heparan sulfates FGFR2IIIb FGFR3 Cellular compartmental homeostasis D.D. Moore, Science 2007 Hepatocyte FGFR4 Has Multiple Effects Beyond Hepatic Cholesterol/Bile Acid Metabolism 1. Limits extent of toxic liver injury and fibrosis 2. Modulates hepatic lipid and glucose metabolism 3. Supports fatty liver in obesity or starvation 4. Reported as both hepatoma promoter and hepatoma suppressor Which effects beyond bile acid metabolism occur directly within hepatocytes due to FGF19-FGFR4-bklotho signaling is unclear. Resident hepatocyte FGFR4 limits hepatocarcinogenesis while ectopic hepatocyte FGFR1 accelerates it Hepatoma cells Hepatocytes Huang, et al. Ectopic activity of fibroblast growth factor receptor 1 in hepatocytes accelerates hepatocarcinogenesis by driving proliferation and vascular endothelial growth factor-induced angiogenesis. Cancer Res 2006 Huang, et al. Resident hepatocyte fibroblast growth factor receptor 4 limits hepatocarcinogenesis. Mol Carcinog. 2008 FGFR4 Knockout Ectopic FGFR1 in Hepatocytes WT R1TG WT R1TG bklotho (KLB) is reduced in human & mouse hepatomas Relative Expression KLB 8 6 Normal Liver Hepatoma 4 2 Without comparable changes in FGFR4 0 -2 -4 -6 Human SIB-CleanEX DbGSE7307;2109 2.5 2 1.5 1 0.5 0 P=0.0018 Apoptosis (A550 x 100) Relative Expression of KLB -8 40 FGFR4-/-/FGFR4ecto FGFR4-/- 30 20 10 0 KLB construct 0 0.04 0.2 1 5 Restoration of KLB and FGFR4 to KLB- and FGFR4-deficient mouse hepatoma cells induces apoptosis Relative Expression KLB drops in liver after partial hepatectomy and recovers during restoration Day after partial hepatectomy Cell Number x 10-5 HEK293 8 6 293 KLB Constitutive(c) FGFR4 Induced(i) cKLB iR4 iR4 iR4 iR4 (R4) iR4 cKLB cKLB F19 cKLB F1 F19 cKLB + iFGFR4 iR4 iR1 iR1 iR1 cKLB cKLB cKLB cKLB F1 F19 F1 4 2 0 KLB partners with FGFR4 to inhibit cell population growth via apoptosis Apoptosis induced by the FGFR1/4-KLB pair is enhanced by either FGF1 or FGF19 FGF21 is specific for the FGFR1-KLB partner KLB can confer growth controlling, anti-tumorigenic pro-apoptotic activity on both FGFR4 and FGFR1 signaling complexes. This is in addition to the role of KLB in conferring high affinity of FGFR4 for endocrine FGF19 and FGFR1 for both FGF19 and FGF21. Direction of pro-apoptotic signaling is likely through KLB-dependent abrogation of anti-apoptotic AKT and mTOR pathways How might this KLB-dependent redirection occur? KLB-dependent abrogation of anti-apoptotic AKT and mTOR pathways? Canonical(c) FGF Paracrine/Autocrine Signaling Endocrine(e) FGF Signaling F F1 HSPG Local PKC Raf PI3K Grb2 sos Ras AKT PI3K AKT sos Distal HSPG Local Local eF F Klothos cF cF sos Grb2 sos Ras PKC MEK ? ERK1/2 Cellular Homeostasis (Development & Adult) Growth, migration, morphogenesis Promotion of Tumor Phenotype Raf MEK ? ERK1/2 Metabolic Homeostasis Inhibition of growth, pro-cell death Tumor Suppression Net KLB-directed anti-growth and anti-tumor effects is consistent with primary function of eFGFs in control of metabolic homeostasis. Targeting hepatic FGFR4 will have serious effects on metabolic homeostasis, particularly bile acid metabolism and may have tumor-promoting effects. Nicholes et al. A mouse model of hepatocellular carcinoma: ectopic expression of fibroblast growth factor 19 in skeletal muscle of transgenic mice. Am J Pathol 2002 Desnoyers et al. Targeting FGF19 inhibits tumor growth in colon cancer xenograft and FGF19 transgenic hepatocellular carcinoma models. Oncogene 2008 French et al. Targeting FGFR4 inhibits hepatocellular carcinoma in preclinical mouse models. PLoS One 2012 Mellor. Targeted inhibition of the FGF19-FGFR4 pathway in hepatocellular carcinoma; translational safety considerations. Liver Int. 2014 FGF21 In contrast to bile acid-controlled diurnal ileal FGF19, FGF21 is significant under conditions of metabolic extremes as starvation and obesity and other sources of organismic stress. FGF21 binds and activates the FGFR1-KLB complex, but not FGFR4KLB. FGFR1 is not expressed in hepatocytes, but is the major FGFR in adipocytes where KLB is also expressed. FGF19 binds and activates both FGFR4- and FGFR1-KLB complexes. [Yang et al. Differential specificity of endocrine FGF19 and FGF21 to FGFR1 and FGFR4 in complex with KLB. PLoS One 2012] •What is the role of adipocyte FGFR1 during metabolic stress conditions where FGF21 is significant? •Is adipocyte FGFR1 an additional target of FGF19 and the major or sole target of FGF21? •Does FGFR1 account for the beneficial effects of FGF21 and FGF19 on obesity and diabetes? Adipocyte-specific ablation of FGFR1 indicates that the adipocyte via FGFR1 is the (possibly the sole) FGFR target that accounts for metabolic effects of FGF21 and the extra-hepatic effects of FGF19 •Yang et al. Control of lipid metabolism by adipocyte FGFR1-mediated adipohepatic communication during hepatic stress. Nutr Metab (Lond). 2012 •Adams & Yang et al. The breadth of FGF21s metabolic actions are governed by FGFR1 in adipose tissue. Mol. Metab. 2012 •Foltz et al. Treating diabetes and obesity with an FGF21-mimetic antibody activating the βKlotho/FGFR1c receptor complex. Sci Transl Med. 2012 Under normal conditions, adipocyte FGFR1 deficiency causes an increase in transcriptional activity of hepatic lipogenic genes without effect on adipocyte genes. Under starvation conditions the FGFR1 deficiency indirectly causes an increase in hepatic steatosis concurrent with an increase in hepatic lipogenic genes without much effect on adipocyte gene expression. Under starvation conditions the adipocyte FGFR1 deficiency causes concurrent elevation of triglyceride and NEFA without effect on glucose or ketone bodies. This occurs concurrent with an increase in adipocyte lipase activity. FGF21/FGFR1-mediated adipo-hepatic communication Under starvation conditions that cause hepatic stress and steatosis, adipose FGFR1 concurrently imposes restrictions on adipocyte lipolysis and indirectly hepatic lipogenesis. This serves to attenuate extent of compensatory hepatic steatosis that often occurs during hepatic stress. Lipolysis and lipogenesis are normally tightly coupled to glucose and ketone body metabolism. Overall lipolysis and lipogenesis are tightly coupled and inversely related. Uncoupling these normally tightly linked domains may mete out and extend lipid reserves for neural fuels (glucose and ketone bodies) during metabolic extremes and other conditions causing hepatic stress. Adipocyte FGFR1-KLB is a target of FGF21 whose primary origin is hepatocytes and an additional target of FGF19 in addition to hepatocyte FGFR4. We speculate this mechanism may underlie the beneficial effects of both endocrine FGFs under both metabolic extremes of starvation and obesity as well as other sources of hepatic stress. [Yang et al. Control of lipid metabolism by adipocyte FGFR1-mediated adipohepatic communication during hepatic stress. Nutr Metab (Lond). 2012] Is adipocyte FGFR1 the sole direct mediator of beneficial effects of FGF21 in obesity/diabetes (metabolic stress)? Adipose FGFR1 deficiency abolishes weight loss, glucose and energy regulation by FGF21 in the obese Foltz et al. Treating diabetes and obesity with an FGF21-mimetic antibody activating the βKlotho/FGFR1c receptor complex. Sci Transl Med. 2012 What about FGF19’s effects in the adipocytes? Adipose FGFR1 deficiency also abolishes weight loss, glucose and energy regulation by FGF19 in the obese Thus FGF19 targets adipocytes via FGFR1 with essentially identical effects to FGF21 FGFR4/KLB: Hepatic Cholesterol to Bile Acids Hepatic Lipogenesis Ileal FGF19 Conclusion During normal feeding ileal FGF19 is likely the normal coordinator of adipo-hepatic communication in the lipid metabolism domain. FGF21 is a hepatokine reserved to instruct adipocytes under conditions of metabolic extremes and other stress conditions sensed by the liver. Effects of FGF19/FGF21 on glucose metabolism are largely an indirect consequence of direct regulation of lipid metabolism in both liver and adipocytes in both normal and extreme conditions. Dr. Chundong Yu Professor Biology Xiamen University US Public Health Service grants John S. Dunn Research Foundation Komen Breast Cancer Foundation Amgen & Eli Lilly Dr. Xinqiang Huang Regulus Therapeutics Dr. Yongde Luo Chaofeng Yang Assistant Professor Postdoctoral IBT Texas A&M UT Southwestern Dr. Fen Wang Professor Center Director IBT Texas A&M How could an extracellular co-factor (HS/klotho) alter diverse intracellular signaling endpoints & phenotypes? Transmembrane conformational transmission? SHP-2 GRB2 SNT1/FRS2 PI3K AKT sos •Few P-Tyr substrate sites like other TKR F F •Mostly SNT1/FRS2a multi-P adapter HSPG m-klotho FGF15/19 FGF21 FGF23 s-klotho •Activated pathways remarkably similar Grb2sos Ras PKC Raf MEK ? ERK1/2 βKlotho (KLB) partners with FGFR4 or FGFR1 to inhibit cell population growth Control cKLB iFGFR4 cKLB + iFGFR4 KLB- and FGFR4-dependent apoptotic cell death is induced by either FGF19 or FGF1 cKLB iFGFR4 9 9.5 cKLB iFGFR4 FGF19 (10) cKLB iFGFR4 FGF19 (102) cKLB iFGFR4 FGF19 (103) PI HEK293 cKLB iFGFR4 FGF19 (1) Apoptosis (%) 4.6 10.5 FGF1 (10) FGF1 (102) 23.8 FGF1 (103) PI FGF1 (1) 16.5 Annexin V Apoptosis (%) 4.5 9 10.5 13.4 Luo et al. Metabolic regulator betaKlotho interacts with fibroblast growth factor receptor 4 (FGFR4) to induce apoptosis and inhibit tumor cell proliferation 2010 JBC 21.4 14.5