(一)-- 从DNA到RNA
advertisement
-- 从DNA到RNA")
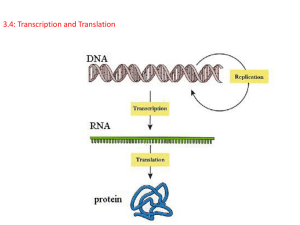
第三讲 生物信息的传递 (上)从DNA到RNA Crick的中心法则(central dogma) DNA 1957年 transcription DNA Reverse 1970年 transcription RNA translation Protein RNA Protein From DNA to Protein DNA序列是遗传信息的 贮存者,它通过自主复 制得到永存,并通过转 录生成信使RNA,翻译 生成蛋白质的过程来控 制生命现象。 基因表达包括转录(transcription)和 翻译(translation)两个阶段。 转录是指拷贝出一条与DNA链序列完全相 同(除了T→U之外)的RNA单链的过程,是 基因表达的核心步骤。 翻译是指以新生的mRNA为模板,把核苷酸 三联遗传密码子翻译成氨基酸序列、合成多 肽链的过程,是基因表达的最终目的。 2006 年度诺贝尔医学奖: 发现了RNA干扰现象 Andrew Z. Fire Craig C. Mello 斯坦福医学院病理学 和遗传学教授 马萨诸塞州医学院分子 医学教授 2006 年度诺贝尔化学奖: 描述真核细胞的转录 Roger D. Kornberg 斯坦福大学医学院医学教授 首位在分子基础上展示真核转录过程是如何运行的。 他制作了详细的检晶仪图片,形容了真核细胞转录的整个运传 情况。我们在他的图片中可以看到新的RNA反转录酶是如何演变 的,和数个在转录过程中必需的其它分子的作用。 RNA主要以单链形式存在于生物体内 Secondary structure of RNA hairpin bulge loop RNA chains fold back on themselves to form local regions of double helix. The double helical structure of RNA resembles the A-form structure of DNA The minor groove is wide and shallow, but offers little sequence-specific information. The major groove is so narrow and deep that it is not very accessible to amino acid side chains from interacting proteins. Thus RNA structure is less well suited for sequence-specific interactions with proteins. Pseudoknot Pseudoknots are complex structure resulted from base pairing of discontiguous RNA segments G:U base pair Non-Watson-Crick G:U base pairs represent additional regular base pairing in RNA, which enriched the capacity for self-complementarity 生物体内拥有三类主要RNA: 1、编码特定蛋白质序列的mRNA; 2、能特异性解读mRNA 中的遗传信息并 将其转化成相应氨基酸后加入多肽链中 的tRNA; 3、直接参与核糖体中蛋白质合成的rRNA。 Functions of RNAs Functions in protein synthesis a. mRNA: as the intermediate between the gene and the proteinsynthesizing machinery. b. tRNA: as an adaptor between the codons in the mRNA and amino acids. c. rRNA: play a structural role, as in the case of the RNA components of the ribosome. As genetic material Serving as a template for its own replication in certain viruses RNA as catalysts (ribozyme) Some RNAs (including one of the structural RNAs of the ribosome) are enzymes that catalyze essential reactions in the cell. RNA is a regulatory molecule Small non-coding RNA which through sequence complementarity binds to, and interferes with the translation of certain mRNAs. Replication: synthesis of two DNA strands using both parental DNA strands as templates. Duplication of a DNA molecule 1 DNA molecule 2 DNA molecules •Transcription: synthesis of one RNA molecule using one of the two DNA strands as a template. Transcription vs. Replication Transcription is chemically and enzymatically very similar to DNA replication. 15 Some important differences: 1.RNA is made of ribonucleotides. 2.RNA polymerase catalyzes the reaction, which does not require a primer (de novo synthesis). 3.The RNA product does not remain base-paired to the template DNA strand. 4.Less accurate (error rate: 10-4). 16 Schematic illustration of transcription Transcription bubble The length of the bubble is ~12-14 bp, and the length of RNA-DNA hybrid within it is ~8-9 bp. DNA-mRNA-the encoded peptide (sense strand) (antisense strand) 编码链(coding strand): 与mRNA序列相同的那条DNA链, 或 称有意义链(sense strand). 模板链(template strand): 根据碱基互补原则指导mRNA合 成的DNA链, 或称反义链(antisense strand)。 coding strand* positive strand Crick strand sense strand 3’ 5’ GTAC 3’ CATG GUAC3’ 5’ antisense strand 5’ Watson strand new RNA strand negative strand template strand* A transcription unit is a sequence of DNA transcribed into a single RNA, starting at the promoter and ending at the terminator. Key terms related to transcription RNA polymerases are enzymes that synthesize RNA using a DNA template (formally described as DNAdependent RNA polymerases). Promoter is a region of DNA where RNA polymerase binds to initiate transcription. Startpoint (Startsite) refers to the position on DNA corresponding to the first base incorporated into RNA. Terminator is a sequence of DNA that causes RNA polymerase to terminate transcription. Transcription unit is the distance between sites of initiation and termination by RNA polymerase. Upstream identifies sequences proceeding in the opposite direction from expression; for example, the bacterial promoter is upstream of the transcription unit, the initiation codon is upstream of the coding region. Downstream identifies sequences proceeding farther in the direction of expression; for example, the coding region is downstream of the initiation codon. Primary transcript is the original unmodified RNA product corresponding to a transcription unit. How does RNA polymerase find promoters on DNA? ( how do proteins distinguish their specific binding sites in DNA from other sequences?) • How do regulatory proteins interact with RNA polymerase to activate or to repress specific steps in the initiation, elongation, or termination of transcription? RNA的转录 (Transcription) RNA合成的特点 1. RNA 是以5’3’方向合成的,它的序列是 与DNA编码链(意义链)相同。 2. RNA 的合成是以反义链(模板链)为模板。 3. 同在DNA中一样,形成磷酸二脂键 ( Phosphodiester bonds)。 4. 必需的成分: RNA polymerase, rNTPs, transcription factors, promoter & terminator/template 转录的基本过程 1、模板识别 (Template Recognition) 2、转录起始 (Initiation) 3、转录的延伸 (Elongation) 4、转录的终止 (Termination) 模板识别 1、RNA聚合酶与启动子DNA双链相互 作用并与之相结合的过程。 2、转录起始前,启动子附近的DNA双 链分开形成转录泡以促使底物核糖核苷 酸与模板DNA的碱基配对。 转录起始 转录起始就是RNA链上第一个核苷 酸键的产生。 1.在起始位点合成RNA链:第一个核苷酸 键的产生,该位点被称为position +1 。 2. 转录起始后直到形成9个核苷酸短链是通 过启动子阶段。 3. 通过启动子的时间代表一个启动子的强 弱:时间越短,转录起始的频率也越高。 一般情况下,转录起始复合物可以进入两 条不同的反应途径: •合成并释放2-9个核苷酸的短RNA转录物,即 所谓的流产式起始; •一旦RNA聚合酶成功地合成9个以上核苷酸,尽 快释放σ亚基,RNA聚合酶离开启动子区,转 录就进入正常的延伸阶段。转录起始复合物通 过上游启动子区并生成由核心酶、DNA和新生 RNA所组成的转录延伸复合物。 Transcription initiation involves 3 defined steps 封闭复合物(closed complex) 开放复合物(open complex) 三元复合物(ternary complex ) 封闭复合物(closed complex) •The initial binding of polymerase to a promoter. •DNA remains double stranded. •The enzyme is bound to one face of the helix. 开放复合物(open complex) The DNA strand separate over a distance of ~14 bp (-11 to +3 ) around the start site (+1 site) Transcription bubble forms 32 三元复合物(ternary complex ) The enzyme escapes from the promoter The transition to the elongation phase Stable ternary complex =DNA +RNA + enzyme RNA合成的起始 RNA聚合酶全酶+启动子 DNA处于双链状态 聚合酶全酶所结合的DNA序 列中有一小段双链被解开 RNA聚合酶、DNA和新生RNA 转录的延伸 RNA 聚 合 酶 离 开 启 动 子,沿DNA链移动并使 新生RNA链不断伸长的 过程就是转录的延伸。 • 共价地向生长RNA链的3’端添加核糖核苷酸 • RNA聚合酶是以5’ 3’ 方向来延长RNA链 • RNA聚合酶本身沿着反义链以3’ 5’方向移动 • 大肠杆菌RNA聚合酶的活性一般为每秒 50-90个核苷酸。 •随着RNA聚合酶的移动,DNA双螺旋持续 解开,暴露出新的单链DNA模板, 新生RNA 链的3’末端不断延伸,在解链区形成RNADNA杂合物。 Proofreading by RNA polymerase Pyrohosphorolytic (焦磷酸键解)editing: the enzyme catalyzes the removal of an incorrectly inserted ribonucleotide by reincorporation of PPi. Hydrolytic (水解)editing: the enzyme backtracks by one or more nucleotides and removes the error-containing sequence. This is stimulated by Gre factor, a elongation stimulation factor. 转录的终止 合成的终止: 当RNA链延伸到转录终止位 点时,RNA polymerase 和RNA链均从 DNA模板上释放出来。 Terminator: 通常含有自我互补区域(selfcomplementary regions) ,在RNA产物 中可以形成stem-loop 或hairpin结构。 Two types of terminators in E. coli •不依赖于ρ因子的终止 intrinsic terminator (内在) •依赖于ρ因子的终止 Rho非依赖型终止子的序列 由两个序列原件组成: •一段短的反向重复序列 (大约20个核苷酸); •其后是一段大约8个A:T 碱基对的序列。 由它转录出mRNA可形 成茎环结构,可阻止 RNA pol的前进。 不依赖于ρ因子 的终止子 (内在 终止子 ) Weakest base pairing: A:U make the dissociation easier 依赖于ρ因子的终止 提纯的RNA聚合酶并不能识别特异性的转录终 止信号,而加入大肠杆菌ρ因子后该聚合酶就 能在DNA模板上准确地终止转录。 • 只含有自我互补区域,可形成茎环结构,但在茎中的 G.C含量少,茎环易打开。 • 其终止需要 ρ 因子的参与。 •ρ因子与ssRNA的特定位点结合(C丰富,G缺乏 )。 • ρ通过催化NTP的水解促使新生RNA链从三元转录复 合物中解离。 ρ因子 ρ因子是一个相对分子 质量为2.0×105的六聚 体蛋白。 其功能是作为RNA pol的一种辅助因子, 当其浓度为RNA pol 浓度的10%时在体外 可发挥最高的活性。 ρ因子参与的RNA合成终止模式 “穷追”(hot pursuit)模型 ρ结合到RNA链终止子上游的 某一点 ρ因子结合以后延着RNA向3′ 端移动,跟踪聚合酶 ρ追上在终止位点暂停的RNA 聚合酶 终止---三元复合物解体 常识数据 •转录生成mRNA的速度大约是每分钟 2500个核苷酸(14个密码子/秒),与翻译 速度(15aa/秒)基本相等,但比DNA复制 的速度要慢得多(800bp /秒) 。 •基因开始表达→mRNA的间隔约为2.5分 钟,而再过半分钟就能在细胞内测到相应 的蛋白质。 转录机器的主要成分 1. RNA聚合酶 (RNA polymerase) 2. 转录复合物 RNA聚合酶 (RNA polymerase) RNA聚合酶是转录过程中最关键的酶。 1. 主要以双链DNA为模板,以4种核苷三磷酸作为 活性前体。 4. 需要Mg2+/Mn2+为辅助因子。 它不需要任何引物。 以5’ 3’ 方向合成RNA链。 5. 缺乏3’ 5’ 外切酶活性。 6. 是一个含有多个亚单位(multi-subunit)的酶。 2. 3. RNA聚合酶需执行的功能 识别DNA双链上的起始子; (2) 使DNA变性在启动子处解旋成单链; (3) 通过阅读启动子序列,RNA pol 确 定它自己的转录方向和模板链。 (4) 最后当它达到终止子时,通过识别停 止转录。 (1) 原核和真核生物的RNA聚合酶虽然 都能催化RNA的合成,但在其分子 组成、种类和生化特性上各有特色。 原核生物(E.coli)的RNA聚合酶 2 1 1 1 1 alpha (α) subunit, beta (β) subunit, beta prime (β’) subunit, omega (ω) subunit, sigma (σ) subunit 155 KD 11 KD 70 KD 151 KD 参与转录延伸 聚合酶全酶 相对分子量:4.65×105 36.5 KD 36.5 KD Core enzyme Holoenzyme 只与转录的 起始有关 原核生物(E.coli)的RNA聚合酶 • E. coli 只有一个 DNA-directed RNA聚合酶 , 来合成所有类型的RNA。 • 是细胞中最大的酶之一。 • 由5种subunits 组成 聚合酶全酶(holoenzyme), 包括 2 α, 1 β, 1β’, 1 ω 以及1 σ subunits 。 • 形状象一个圆筒状通道,可以直接与16 bp DNA结合。整个聚合酶可结合60 bp DNA。 • RNA 合成速率: 40 nt /秒, 37oC 。 E. coli RNA polymerase :αsubunit •是核心酶中的两个相同的亚单位 •由rpoA 基因编码 •与核心酶的组装有关 •参与RNA聚合酶和部分调节因子的相互作用 E. coli RNA polymerase :β &β’ subunit •β 和β’分别由 rpoB 和 rpoC 基因编码。 •由β和β’亚基组成了聚合酶的催化中心。 •它们在序列上与真核生物RNA聚合酶的两 个大亚基有同源性。 •β亚基能与模板DNA、新生RNA链及核苷 酸底物相结合。 rpoB 和 rpoC 基因的突变 会影响转录所有的阶段。 The β and β’ subunit of RNA polymerase have a channel for the DNA template. E. coli RNA polymerase : σfactor 负责模板链的选择和转录的起始: •与σ因子的结合使RNA聚合酶从核心酶转变为聚 合酶全酶。 •是启动子识别的关键的酶,不仅增加聚合酶对启 动子的亲和力(提高103倍),还可降低它对非专一 位点的亲和力(降低104倍),使酶底复合物的半衰 期小于1s。 •在细胞中对σ因子量的需求少于聚合酶中其它亚 单位。 The conformational change in σ at open complex formation 在某些细菌中含有识别不同启动子 的σ因子,以适应不同生长发育阶 段的要求,控制不同基因转录的起 始。 大肠杆菌中的σ因子能识别并与 启动子区的特异性序列相结合 因子 基因 功能 -35区 间隔 (bp) -10区 σ70 rpoD 广泛 TTGAC A 16-18 TATAAT σ32 rpoH 热休克 TCTCNC CCTTG AA 13-15 CCCCAT NTA σ54 rpoN 氮代谢 CTGGN A 6 TTGCA • σ70 当与核心酶结合时会改变其结构,以释放其DNA-结合区域. •σ70 与 –35 和 –10 序列均结合。 T3 和 T7 噬菌体的RNA 聚合酶是由一条小的多 肽链组成, 相对分子量小于1×105; 在37ºC下,其转录速度为 200 nt/秒; 它们只能识别不同于E. coli 启动子的自身的启动 子。 RNA polymerase differs from organism to organism 真核生物的RNA聚合酶 •有3类RNA聚合酶; •结构比大肠杆菌RNA聚合酶复杂; •在细胞核中的位置不同; •负责转录的基因不同,对α-鹅膏蕈碱的敏感 性也不同。 •真核生物RNA聚合酶一般有8-14个亚基所组成, 相对分子质量超过5×105。 真核细胞中三类RNA聚合酶特性比较 酶 细胞内 定位 转录产 物 相对活 性 对α-鹅膏 蕈碱的敏 感程度 RNA聚合 酶I 核仁 rRNA 50%70% 不敏感 RNA聚合 酶II 核质 hnRNA*, snRNA, mRNA 20%40% 敏感 RNA聚合 酶III 核质 tRNA 5SRNA 约 10% 存在物种 特异性 (28S,18S, 5.8S) 某些涉及 RNA加工的 snRNA *hnRNA: heterogeneous muclearRNA,核内不均一RNA, RNA的前体 转录的抑制剂 抑制剂 靶酶 抑制作用 利福霉素 细菌全酶 链霉溶菌素 细菌核心酶 放射线素D 真核PolⅠ α-鹅膏蕈 真核PolⅡ 和β亚基结合,抑制 起始 和β亚基结合,抑制 起始 和DNA结合,阻止 延伸 和RNA PolⅡ结合 RNA合成抑制剂主要分两类: 1. 模板结合抑制剂 2. 聚合酶抑制剂 在动、植物及昆虫的细胞中,RNA pol Ⅱ的活性可被低浓度的α-鹅膏蕈碱所 抑制。但却不抑制pol Ⅰ。 Pol Ⅲ对α-鹅膏蕈的反应,不同的生 物有所差异。在动物细胞中高浓度的 α-鹅膏蕈可抑制转录,在昆虫中不受 抑制。 真核生物RNA聚合酶的主要特征 •聚合酶中有两个相对分子质量超 过1×105的大亚基; •同种生物3类聚合酶有“共享”小 亚基的倾向,即有几个小亚基是其 中3类或2类聚合酶所共有的。 真核生物RNA聚合酶的亚基 RNA聚合酶I RNA聚合酶II RNA聚合酶III RPA 1 RPB 1 (’) RPC 1 RPA 2 RPB 2 ( ) RPC 2 RPC 5 RPB 3(αI) RPC 5 RPC 9 RPB 11 (αII) RPC 9 RPB 6 RP 6 (ω) RPB 6 其它9个亚基 其它7个亚基 其它11个亚基 注:亚基按照分子量由大到小的顺序排列。 真核生物RNA聚合酶一般有8-16个亚基所组成, 相对分 子质量超过5×105。 真核生物线粒体和叶绿体中存在 不同的RNA聚合酶 线粒体中RNA聚合酶: 只有一条多肽链,相对分子量小于7 X 104,是已知最 小的RNA聚合酶之一; 与T7噬菌体RNA聚合酶有同源性。 叶绿体中RNA聚合酶: 比较大; 结构上与细菌中的聚合酶相似,由多个亚基组成,部分 亚基由叶绿体基因编码。 线粒体和叶绿体RNA聚合酶活性不受α-鹅膏蕈 碱所抑制。 a a eukaryotic RPB3 “Crab claw” shape of RNAP ’ prokaryotic The same color indicate the homologous of the two enzymes w RPB2 Active center cleft RPB1 RPB11 RPB6 模板DNA进入转录复合体的路径 (A)俯瞰图。双螺旋DNA用倾 斜的圆筒表示,TF II转录因子的 结合位点用虚圈表示; (B)后视图。DNA从钳子 型结构和墙体或平滑结构的 中间穿越。 There are various channels allowing DNA, RNA and ribonucleotides (rNTPs) into and out of the enzyme’s active center cleft RNA polymerase/transcription and DNA polymerase/replication RNA pol DNA pol Template dsDNA dsDNA Require primer No Yes Initiation promoter origin elongation 40 nt/ sec 900 bp/sec Exonuclease activity terminator No Yes Synthesized RNA Template DNA 转录机器的主要成分 1. RNA聚合酶 (RNA polymerase) 2. 转录复合物 转录复合物 封闭复合物 RNA聚合酶全酶+启动子 DNA处于双链状态 开放复合物 聚合酶全酶所结合的DNA序 列中有一小段双链被解开 三元复合物 RNA聚合酶、DNA和新生RNA RNA合成起始 除RNA聚合酶之外,真核生 物转录起始过程中至少还需要 7种辅助因子参与。 真核生物RNA聚合酶II所形成的转录起始复合物 蛋白质 亚基数 亚基的分子量 (ku) 功 能 RNA聚合酶II 12 10-220 催化RNA的生物合成 TBP 1 38 与启动子上的TATA区相结合 TFIIA 3 12,19,35 使TBP及TFIIB与启动子的结合比 较稳定 TFIIB 1 35 与TBP相结合,吸引RNA聚合酶和 TFIIF到启动区上 TFIID 12 15-250 与各种调控因子相互作用 TFIIE 2 34,57 吸引TFIIH,有ATP酶及解链酶活 性 TFIIF 2 30,74 结合RNA聚合酶II并在TFIIB帮助 下阻止聚合酶与非特异性DNA序列 相结合 TFIIH 12 35-89 在启动子区解开DNA双链,使 RNA聚合酶II磷酸化,接纳核苷酸 切除修复体系 前起始复合物 (preinitiation transcription complex, PIC) 真核生物RNA聚合酶不能直接识别基因 的启动子区,需要一些被称为转录调控 因子的辅助蛋白质按特定顺序结合于启 动子上,RNA聚合酶才能与之相结合并 形成复杂的前起始复合物(PIC)以保证有 效地起始转录。 Assembly of the pre-initiation complex in presence of mediator, nucleosome modifiers and remodelers, and transcriptional activators Mediator consists of many subunits, some conserved from yeast to human. Only subunit Srb4 is essential for transcription of essentially all Pol II genes in vivo. PNAS 2007 vol. 104 no. 32 12955–12961 Trends in Microbiology, Vol 16(3), 2008, Pages 126-134 转录单元(transcription unit) 转录起点是指与新生RNA链第一个核苷酸相对应 DNA链上的碱基,通常为嘌呤。 把起点5’末端的序列称为上游(upstream), 把其3’末端的序列称为下游(downstream)。 转录起始位点 (Transcription start site) • 在原核中90%为嘌呤,A或G; • 位置固定 ; • 通常在起始核苷酸的两侧为C 和 T (i.e. CGT or CAT) 启动子区的基本结构 启动子是一段位于结构基因5’端上游区 的保守的DNA序列,能活化RNA聚合酶, 使之与模板DNA准确地相结合并具有转 录起始的特异性。 启动子与转录起始 大肠杆菌RNA聚合酶与启动子的相互作用主 要包括: • 启动子区的识别; • 酶与启动子的结合; • σ因子的结合与解离。 原核生物不同基因的启动子 -10区(Pribnow区)的发现 1975年,Pribnow和Schaller将RNA 聚合酶全酶与模板DNA结合后,用 DNaseI降解DNA,得到41~44个核 苷酸对的DNA片段。 序列分析发现,在被保护区内有一 个由5个核苷酸组成的保守序列, 是聚合酶结合位点,称为Pribnow 区,其中央大约位于起点上游10bp 处,所以又称为–10区。 -10序列 (Pribnow框盒) (1) (2) (3) 其保守序列为TATAAT,位于-10bp左右,其中 3′端的“T”十分保守。 A.T较丰富,易于解链。 它和转录起始位点I一般相距5bp。 功能: RNA pol紧密结合; 形成开放启动复合体; 使RNA pol定向转录。 如果把Pribnow区从TATAAT变成AATAAT就会使 该启动子发生下降突变(down mutation); 如果增加Pribnow区的共同序列,将乳糖操纵子的 启动子中的TATGTT变成TATATT,就会提高启动 子的效率,称为上升突变(up mutation)。 提纯被保护的片段后却发现, RNA聚合酶并不能重新结 合或并不能选择正确的起始 点,表明在保护区外可能还 存在与RNA聚合酶对启动 子的识别有关的序列。 ? -35区的发现 科学家又从噬菌体的左、右启动子PL 及PR 和SV40启动子的–35 bp附近找到了 另一段共同序列:TTGACA。 -35序列 (Sextama盒) 其保守序列为TTGACA, 与-10序列相隔16-19bp。 功能: (1)为RNA pol的识别位点。 (2)RNA Pol的核心酶只能起到和模板结合和催 化的功能,并不能识别-35序列,只有σ亚基才 能识别-35序列,为转录选择模板链。 100个E.coli的不同启动子的序列测定结果 –35区 –10区 ……T85T83G81A61C69A52……T89A89T50A65A100…… 大部分启动子都存在这两段共同序列,即位于– 10 bp处的TATA区和–35 bp处的TTGACA区。 它们是RNA聚合酶与启动子的结合位点,能与σ因 子相互识别而具有很高的亲和力。 -10区和-35区的最佳距离 在原核生物中, -10区和-35区的最佳距离大 约是16~19 bp。 过大或过小都会降低转录活性。 这可能是因为RNA Pol本身的大小和空间结构有 关。 Some stronger bacterial promoter Confers additional specificity UP-element is an additional DNA elements that increases polymerase binding by providing the additional interaction site for RNA polymerase 92 promoter 40-80bp 5’ 1 -50 -40 -30 TTGACA AACTGT Recognition site -20 2 3 -10 1 gene10 TATAATPu ATATTAPy transcribed initiation site Binding site Pribnow box (1)结构典型,都含有识别(R),结合(B)和起始(I)三个位点; (2)序列保守,如-35序列,-10序列结构都十分保守; (3)位置和距离都比较恒定; (4)直接和多聚酶相结合; (5)常和操纵子相邻; (6)都在其控制基因的5′端; (7)决定转录的启动和方向。 原核生物启动子 的共同的特点 真核生物启动子的结构特点 真核生物RNA聚合酶II所识别的启动子区 Hogness等发现类似Pribnow区的Hogness区, 在转录起始点上游–25~–30 bp处,保守序列为 TATAAA,也称TATA区。 在起始位点上游–70~–78 bp处还有另一段共同 序列CCAAT,称为CAAT区(CAAT box)。 真核基因的启动子 1. 核心元件 TATA box: –25~–30 bp区 启始子(initiator,Inr) :转录起始位点附近 2. 上游启动子元件(UPE) CAAT box :–70~–80区CCAAT序列 GC box: –80~–110含有GCCACACCC 或GGGCGGG序列。 上游启动子元件 (upstream promoter element,UPE): 将TATA区上游的保守序列称为上游启动 子元件或称上游激活序列(upstream activating sequence,UAS)。 真核生物启动子对转录的影响 • TATA区--使转录精确地起始: 如果除去TATA区或进行碱基突变,转录产物下降 的相对值不如CAAT区或GC区突变后明显,但发 现所获得的RNA产物起始点不固定。 • CAAT区和GC区主要控制转录起 始频率: 基本不参与起始位点的确定。 研究SV40晚期基因启动子发现,上游激活区的存在与否,对该 启动子的生物活性有着根本性的影响。若将该基因5' 上游–21~–47 核苷酸序列切除,基因完全不表达。 远端调控区 增强子(enhancer) 1981年由Benerji, Rusconi小组和Chambom 等发现的,又称远上游序列(far upstream sequence)。 已在SV40的转录单元上发现其转录起始位点 上游约200bp处有两段72bp长的重复序列, 它们不是启动子的一部分,但能增强或促进 转录的起始,因此,称这种能强化转录起始 的序列为增强子或强化子(enhancer)。 增强子(Enhancer ) 增强子很可能通过影响染色质DNA-蛋白质结构或改变 超螺旋的密度而改变模板的整体结构,从而使得RNA聚 合酶更容易与模板DNA相结合,起始基因转录。 An enhancer can activate a promoter from upstream or downstream locations, and its sequence can be inverted relative to the promoter. 增强子特点 ① 具有远距离效应。 常在上游-200bp处,但可增强远处启动子的转录,即使相距十几Kb也能发挥 其作用; ② 无方向性。 无论在靶基因的上游,下游或内部都可发挥增强转录的作用; ③ 顺式调节。 只调节位于同一染色体上的靶基因,而对其它染色体上的基因无作用; ④ 无物种和基因的特异性, 可以接到异源基因上发挥作用; ⑤ 具有组织的特异性。 SV40的增强子在3T3细胞中比多瘤病毒的增强子要弱,但在HeLa细胞中SV40 的增强子比多瘤病毒的要强5倍。增强子的效应需特定的蛋白质因子参与。 ⑥ 有相位性。 其作用和DNA的构象有关。 ⑦ 有的增强子可以对外部信号产生反应。 如热体克基因在高温下才表达。编码重金属蛋白的金属硫蛋白基因在镉和锌存 在下才表达。某些增强子可以被固醇类激素所激活。 真核生物的启动子特点 promoter 5’ -50 GC box CAAT box -40 -30 -20 -10 1 10 gene TATA box Hogness box (1)有多种元件:TATA框,GC框,CATT框,OCT等; (2)结构不恒定。有的有多种框盒,如组蛋白H2B;有的只有TATA框和 GC框,如SV40早期转录蛋白; (3)它们的位置、序列、距离和方向都不完全相同; (4)有的有远距离的调控元件存在,如增强子; (5)这些元件常常起到控制转录效率和选择起始位点的作用; (6)不直接和RNA pol结合。转录时先和其它转录激活因子相结合,再和 聚合酶结合。 原核与真核生物mRNA的特征比较 原核生物中: • mRNA的转录和翻译发生在同一个细胞空间, • 这两个过程几乎是同步进行的 。 真核细胞中: 真核细胞mRNA的合成和功能表达发生在不同 的空间和时间范畴内。 • mRNA以较大分子量的前体RNA出现在核内, •只有成熟的、相对分子质量明显变小并经化学 修饰的mRNA才能进入细胞质,参与蛋白质的 合成。 mRNA的组成: •编码区(coding region):从起始密码子AUG开 始经一连串编码氨基酸的密码子直至终止密码 子。 •5’端上游非编码区(5’UTR):位于AUG之前不 翻译的区域。 •3’端下游非编码区(3’UTR) :位于终止密码子 之后不翻译的区域。 原核生物mRNA的特征 半衰期短。 许多原核生物mRNA以多顺反子的形式存 在。 原核生物mRNA的5’端无帽子结构,3’端没 有或只有较短的多聚(A)结构。 原核生物mRNA的特征 1. 半衰期短 •原核生物中,mRNA的 转录和翻译是在同一个 细胞空间里同步进行的, 蛋白质合成往往在 mRNA刚开始转录时就 被引发了。 •大多数细菌mRNA在转 录开始1分钟后就开始降 解。mRNA降解的速度大 概只有转录或翻译速度的 一半。 原核生物mRNA的特征 2. 许多以多顺反子的形式存在: 原核细胞的mRNA(包括病毒)有时可以同 时编码几个多肽。 Prokaryotic mRNA (polycistrionic) 单顺反子mRNA (monocistronic mRNA): 只编码一个蛋白质的mRNA。 多顺反子mRNA(polycistronic mRNA): 编码多个蛋白质的mRNA。 原核生物mRNA的特征 3. 原核生物mRNA的5’端无帽子结构,3’ 端没有或只有较短的多聚(A)结构。 原核生物起始密码子AUG上游有一被称为Ribosome Binding Site (RBS)或SD序列(Shine –Dalgarno sequence)的保守区,因为该 序列与16S-rRNA 3’端反向互补,所以被认为在核糖体-mRNA的 结合过程中起作用。 原核生物mRNA的特征 4. 原核生物常以AUG(有时GUG,甚至 UUG)作为起始密码子; 真核生物几乎永远以AUG作为起始密 码子。 真核生物mRNA的特征 • 单顺反子形式存在。 • 5’端存在“帽子”结构。 • 绝大多数具有多聚(A)尾巴。 真核生物mRNA的结构模式 Eukaryotic mRNA (monocistrionic) “基因”的分子生物学定义是:产 生一条多肽链或功能RNA所必需的全 部核苷酸序列! A gene can be defined as following: The entire nucleic acid sequence that is necessary for the synthesis of a functional polypeptide or RNA molecule. 真核生物mRNA的特征 1. 真核生物mRNA的5’端存在“帽子” 结构。 真核生物基因转录一般从嘌呤起始,其 5’端大都经过修饰。 5’Capping 通过 5′→ 5′磷酸二酯键在原初mRNA的5’端倒扣 一个“G”。 在新生mRNA链达到50个核苷酸前,甚至可能在 RNA Pol II 离开转录起始位点之前,帽子结构就 已加到mRNA的第一个核苷酸上了。 5′末端加上鸟苷是由鸟苷转移酶催化的 。 帽子结构是GTP和原5’三磷酸腺苷(或鸟苷)缩 合反应的产物。 mRNA的帽子结构常常被甲基化。 真核生物mRNA的“帽子”结构 鸟苷酸-7甲基转移酶 2’-O-甲基转移酶 mRNA的帽子结构常常被甲基化 零类帽子(cap0):第一个甲基出现在所有真核细胞的 mRNA中(单细胞真核生物mRNA主要是这个结构), 由鸟苷酸-7甲基转移酶催化,称为零类帽子。 1类帽子(cap1):如在第二个核苷酸(原mRNA 5’第一位) 的2’-OH位上加另一个甲基,这步反应由2’-O-甲基转移 酶完成。一般把有这两个甲基的结构称为1类帽子。真核 生物中以这类帽子结构为主。 2类帽子(cap2): 在某些生物细胞内,mRNA链上的第三 个核苷酸的2’-OH位也可能被甲基化,因为这个反应只 以带有1类帽子的mRNA为底物,所以被称为2类帽子。 只占有帽mRNA总量的10%-15%以下。 帽子结构的功能 (1)有助于mRNA越过核膜,进入胞质; (2)保护5′不被核酶降解; (3)翻译时供IFⅢ(起始因子)和核糖体识别, 是翻译所必需的。 真核生物mRNA的特征 2. 绝大多数真核生物mRNA具有多聚 (A)尾巴。 •除组蛋白基因外,真核生物mRNA的3’末端都 有多聚(A)序列,其长度因mRNA种类不同而变 化,一般为40-200个左右。 •由多聚(A)聚合酶催化的; •它是在转录后加上的; •Poly(A)被特异的蛋白质PABP结合。 在高等生物中(酵母除外)在poly(A)上游11-30nt处 有一特殊序列AAUAAA,这一序列是高度保守的, 对于初级转录产物的准确切割及加多聚(A)是必需 的。 真核生物mRNA中的加多聚A反应 CPSF (cleavage and polyadenylation specificity factor) CstF (cleavage stimulation factor) 真核基因的3‘ 末端转录终止位点上游15~30bp处的保守序列 AAUAAA对于初级转录产物的准确切割及加多聚(A)是必需的 多聚腺苷化(polyadenylation) 反应要经过2个阶段 (1)首先将一个短的寡聚A序列(10nt)加到3’端, 此反应绝对依赖于AAUAAA序列,这是由 poly(A)聚合酶在特殊因子指导下完成的。 (2)寡聚A尾巴延伸到240nt的长度。 此反应并不需要AAUAAA序列,但需要一个识 别寡聚A并指导poly(A)聚合酶延伸的刺激因 子。 多聚(A)的功能 •是mRNA由细胞核进入细胞质所必需的形式; •它大大提高了mRNA在细胞质中的稳定性。 mRNA刚从细胞核进入细胞质时,其多聚(A)尾巴一般 比较长,随着mRNA在细胞质内逗留时间延长,多聚 (A)逐渐变短消失,mRNA进入降解过程。 •它可促进核糖体的有效循环。 Poly-A in the 3’ end promotes the efficient recycling of ribosomes Poly (A) - 尽管大部分真核mRNA有poly (A)尾巴,细 胞中仍有多大1/3没有poly (A)的mRNA,将 其称为Poly (A) – 约1/3的Poly (A) –mRNA编码了不同形式的 组蛋白。 Poly(A)+ RNA can be separated from other RNAs by fractionation on Sepharose-oligo(dT). 在动物细胞中,mRNA的 表达需转录、修饰、加 工、核质转运和翻译。 内含子的剪接、编辑及化学修饰 RNA中的内含子 真核基因大多是断裂的: •一个基因可由多个内含子和外显子间 隔排列而成。 •内含子在真核基因中所占的比例很高, 甚至超过99%。 非编码区 编码区 Exons (外显子): the coding sequences Introns (内含子) : the intervening sequences 部分人类基因中内含子序列所占的比重分析 基因 长度(kb) 内含子数量 内含子所占比重 (%) 胰岛素 1.4 2 67 ß-球蛋白 1.4 2 69 血清蛋白 18 13 89 胶原蛋白组分VII 31 117 71 VIII因子 186 25 95 萎缩性肌强直因子 2400 78 >99 剪接 (Splicing) RNA splicing: removal of introns and joining of exons. • It takes place in the nucleus before the mature mRNA can be exported to the cytoplasm. • Catalyzed by spliceosome (RNA +protein) mRNA的前体 (核不均一RNA, hnRNA) 5’加“帽”和 3’酶切加多聚腺苷酸 RNA的剪接 连续的可译框(ORF) (open reading frame) 通过核孔进入细胞质 蛋白质合成的模板 RNA splicing 可通过体外 凝胶电泳进行分析 RNA加工过程及其生理功能 加工过程 推测的生理功能 加帽子反应 mRNA从细胞核向细胞质运转, 翻译起始 加多聚A反应 转录终止,翻译起始和mRNA降解 RNA的剪接 从mRNA, tRNA和rRNA分子中切除 内含子 RNA的切割 从前体RNA中释放成熟tRNA和 rRNA分子 •真核基因平均含有8-10个内含子,前体分 子一般比成熟mRNA大4-10倍。 •不同生物细胞内含子的边界存在相似的核 苷酸序列。 •内含子的“功能”及其在生物进化中的地 位是一个引人注目的问题。 内含子剪接异常引起疾病 例如: 地中海贫血病人的珠蛋白基因中,大约有 1/4的核苷酸突变发生在内含子的5’或3’边界 保守序列上,或者直接干扰了前体mRNA的 正常剪接。 生物体内的各种内含子 内含子类型 细胞内定位 GU-AG 细胞核,前体mRNA (真核) AU-AC 细胞核,前体mRNA(真核) I类内含子 细胞核,前体rRNA(真核), 细胞器RNA,少数细菌RNA II类内含子 细胞器RNA,部分细菌RNA III类内含子 细胞器RNA 双内含子 细胞器RNA tRNA前体中的内含子 细胞核,tRNA前体 (真核) GU-AG或AU-AC分别代表了不同内含子的5’和3’边界序列。 断裂基因发现不久,Crick(1978年)就 提出了一系列发人深思的问题: (1)剪切作用若是通过酶来进行的,那么这 种剪接酶是怎样识别RNA上特定的位点? (2)剪切酶有没有特异性?对不同的RNA是 否需要不同的酶? (3)切除的RNA是以线状还是环状存在的? 切下的RNA的命运将如何? Chambon等分析比较了大量结构基因的内含 子切割位点,发现有2个特点: (1)内含子的两个末端并不存在同源或互补。 (2)连接点具有很短的保守序列,亦称边界序列。 100种内含子的5′端都是GU;3′端都是AG, 因此称为GU-AG法则(GU-AG rule),又称 为Chambon法则。 A 脊椎动物前体mRNA中常见内含子 剪接所必需的保守序列 两个剪接位点的序列是不同的: • 左边的剪接位点称供体(donor)位点, • 右边的剪接位点称受体(acceptor)位点。 GU-AG法则(GU-AG rule)不适用于 线粒体、叶绿体的内含子,也不适用于 酵母的tRNA基因 。 除了边界序列之外,外显子与内含子 交界处的序列,内含子内部的部分序 列都有可能参与内含子的剪接。 核mRNA结构特点 1. 边界序列:其边界序列是完全符合GU-AG法则 2. 分枝点序列:具有分枝点序列,位于内含子3’端上游1850nt处,序列为Py80NPy87Pu75APy95,其中A为百分之百 的保守,且具有2’-OH。 3. 内含子5′端有一保守序列(5′GUAAGUA3’)可以和U1 snRNA的5’端的保守序列3’CAUUUCAU5’互补。 核mRNA的剪接 转录产生的核内mRNA前体分子与蛋白质 结合,形成RNA和蛋白质组成的snRNP 复合物(ribonucleo-protein protein)。 随着RNA链的延伸,每个内含子5’和3’两 端的复合物成对联结,产生60S的颗粒— 剪接体(spliceosome),进行RNA前体 分子的剪接。 细胞核中的小分子RNA称为细胞核小RNA(small nuclear RNA, snRNA); 位于细胞质中的称为细胞质小RNA (small cytoplasmic RNA, scRNA)。 在自然状态下,它们以核糖核蛋白颗粒(SnRNP和 scRNP)的形式存在,俗称snurps和scyrps。 在核仁中也存在着一类小的RNA,称为核仁小RNA (small nucleolar RNA, snoRNA),它们在rRNA的加工中 起作用。 snRNA参与剪接过程,并与其它蛋白一起构成一个大 的颗粒复合体,称为剪接体(splicesome)。 Spliceosome (剪接体) Catalyzes pre-mRNA splicing in nucleus. The spliceosome comprises about 150 proteins (splicing factors etc.) and five snRNAs (U1, U2, U4, U5 and U6),and the pre-mRNA being assembled. The complexes of snRNA and proteins are called small nuclear ribonuclear proteins (snRNP, pronounces “snurps”). Three roles of snRNPs in splicing 1. Recognizing the 5’ splice site and the branch site. 2. Bringing those sites together. 3. Catalyzing (or helping to catalyze) the RNA cleavage. RNA-RNA, RNA-protein and proteinprotein interactions are all important during splicing. 真核生物mRNA前体中内含子剪接过程示意图 • 由U1 snRNA以碱基互补的方式识别mRNA前体5’剪接点,由结合在3’剪接点上 游富嘧啶区的U2AF(U2 auxiliary factor)识别3’剪接点并引导U2 snRNP与分支 点相结合,形成剪接前体(pre-spliceosome)。 • 剪接前体进一步与U4、U5、U6 snRNP三聚体相结合,形成剪接体。 The lariat (套索) is an intermediate in RNA splicing in which a circular structure with a tail is created by a 5 ’ -2 ’ bond. The branch site (分支点) is a short sequence just before the end of an intron at which the lariat intermediate is formed in splicing by joining the 5’ nucleotide of the intron to the 2 ’ position of an Adenosine. Biochemical steps of pre-mRNA splicing Step 1: a cut is made at the 5′splice site, separating the left exon and the right intron-exon molecule. Step 2: cutting at the 3′ splice site releases the free intron in lariat form, while the right exon is ligated (spliced) to the left exon. 哺乳动物细胞中mRNA前体上的snRNP是从5’向 下游“扫描”,选择在分支点富嘧啶区3’下游的 第一个AG作为剪接的3’受点。 AG前一位核苷酸可以影响剪接效率,一般说来, CAG=UAG>AAG>GAG。 如果mRNA前体上同时存在几个AG,可能发生 剪接竞争。 内含子的变位剪接 Alternative Splicing • 在高等真核生物个体发育或细胞分化过程中 可以有选择性地越过某些外显子或某个剪接点 进行变位剪接,产生出组织或发育阶段特异性 mRNA。 • 脊椎动物中大约有5%的基因能以这种方式进 行剪接,保证各同源蛋白质之间既具有大致相 同的结构或功能域,又具有特定的性质差异, 进而拓展了基因所携带的遗传信息。 Alternative splicing (可变剪接): some pre-mRNAs can be spliced in more than one way , generating alternative mRNAs. 60% of the human genes are spliced in this manner. Drosophila DSCAM gene can be spliced in 38,000 alternative ways Different ways of alternative splicing Self-splicing introns -I、II类内含子 带有这些内含子的RNA本身具有催化活性, 能进行内含子的自我剪接。 Autosplicing (Self-splicing,自我剪接) describes the ability of an intron to excise itself from an RNA by a catalytic action that depends only on the sequence of RNA in the intron. There are two classes of selfsplicing introns: group I self-splicing introns group II self-splicing introns. Self-splicing introns fold into a specific conformation catalyze the chemical reaction using metal ions as cofactors I类内含子的结构特点 1. 其边界序列为5’U-G 3’ 2. 内含子中具有中部核心结 构(Central core structure) I 类内含子常分布于低等真核生物的 细胞器中,如四膜虫、绒泡菌的 rRNA等 I类内含子的自我剪接过程 自由鸟苷酸的3’-OH作为 亲核基团攻击内含子5’端 的磷酸二酯键,从上游切 开RNA链。 上游外显子的自由3’OH作为亲核基团攻击内 含子3’位核苷酸上的磷 酸二酯键,使内含子被完 全切开。 上下游两个外显子通过新 的磷酸二酯键相连 Ⅱ类内含子 和I类内含子剪接不同; 和核mRNA 内含子的剪切有些相似,但也有不 同之处。 Ⅱ类内含子分布在: 酵母mtRNA, 细胞色素氧化酶的α亚基,β亚基, 玉米mitochondrial RNA (mtRNA),tRNA 真核mRNA前体中 Ⅱ类内含子的剪接无需鸟苷的辅助, 但需镁离子的存在。 II类内含子的自我剪接过程 分枝点腺苷酸的2’-OH作为亲核 基团攻击内含子5’端的磷酸二酯 键,内含子5′的边界序列上的G以 5′磷酸和分枝点A的2′-OH形成磷 酸二酯键,从而产生了套索 (lariat)结构。 上游外显子的3’-OH作为亲核基团 攻击内含子3’位核苷酸上的磷酸二 酯键,使套索结构完全解离。 上下游两个外显子通过新的磷酸 二酯键相连。 几种不同内含子剪接反应的区别 酵母tRNA I类内含子 Ⅱ类内含子 核mRNA前体 边界顺序 无 5’U↓-G↓3’ 5’↓GUAG↓3’ 5’ ↓GU-AG↓3’ 特殊顺序 C(茎上)- G(环上) 内部引导顺 序 分支点序列 分支点序列,5’ 外 显子顺序 二级结构 茎环构象 核心结构 5、6功能区 连接体 基因外的 成分 内切酶,连接 酶 GTP,镁离 子 镁离子 U1U2,U4,U5,U6 能量要求 ATP 不 不 ATP 中间型分 子 半分子tRNA 环状L-19 IVS 套索 套索 The chemistry of group II intron splicing and RNA intermediates produced are the same as that of the nuclear pre-mRNA. 三类剪接反应都是通 过两次连续的 转酯反应进行的 第一次转酯反应中,由一个 游离的2’-羟基提供,发动对5’ 外显子-内含子连接点的攻击。 第二次转酯反应中,已经释 放的外显子末端的游离3’-羟 基接着攻击3’内含子-外显子 的连接点。 RNA的编辑和化学修饰 •RNA的编辑(RNA editing)是某些RNA, 特别是mRNA的一种加工方式。 •它导致了DNA所编码的遗传信息的改变。 RNA的编辑(RNA editing) 编辑(editing)是指转录后的RNA在编码区发生 碱基的加入,丢失或转换等现象。 介导RNA编辑的两种机制: • 位点特异性脱氨基作用; • 引导RNA指导的尿嘧啶插入或删除。 A specifically targeted C residue within mRNA is converted into U by the deaminase (脱氨酶). The process occurs only in certain tissues or cell types and in a regulated manner. 哺乳动物载脂蛋白基因转录产物的编辑 肝中剪接的mRNA编码 含4563 aa的蛋白质 肠mRNA有UAA密码子在 第2153位密码子终止合成 •RNA的编辑虽然不是很普遍,但在 真核生物中却时有发生。 •RNA的编辑可能是充分发挥生理功能 所必须的。 哺乳动物中RNA编辑的实例 组织 靶标RNA 所改变 的 碱基 结果 肝脏,肠 载脂蛋白B C→U 谷氨酰胺密码子→终止子 肌肉 半乳糖苷酶 U→A 苯丙氨酸密码子→酪氨酸 睾丸,肿瘤 等 Wilms肿瘤基因 -1 U→C 亮氨酸密码子→脯氨酸 肿瘤 神经纤维瘤基因 -1 C→U 精氨酸密码子→终止子 脑 谷氨酸受体蛋白 A→I 多个谷氨酸密码子→精氨 酸 RNA编辑的另一种形式是尿苷酸的 缺失和添加。 研究发现,利什曼原虫属细胞色素b mRNA 中含有许多独立于核基因的尿嘧啶残基,而 特 异 性 插 入 这 些 残 基 的 信 息 来 自 指 导 RNA (guide RNA)。 指导RNA(guide gRNA) 1990年, L.Simpsom等在研究锥虫线粒体 mRNA时发现了一类新的小分子RNA: 可以和mRNA分子被编辑的部分发生非常规的 互补,G-U配对, 对mRNA前体分子的编辑起了指导作用,故称 其为指导RNA(guide gRNA)。 gRNAs Having three regions: anchor– directing the gRNAs to the region of mRNAs it will edit. editing region – determining where the Us will be inserted poly-U stretch 指导RNA和RNA的 编辑机制 指导RNA与被编辑区及其周围 部分核酸序列虽然有相当程度 的互补性,但该RNA上存在一 些未能配对的腺嘌呤,形成缺 口,为插入尿嘧啶提供了模板。 反应完成后,指导RNA从mRNA 上解离下来,而mRNA则被用做 翻译的模板。 RNA编辑的生物学意义 (1) 校正作用。 (2) 调控翻译。 通过编辑可以构建或去除起始密码子和终止 密码子。 (3) 扩充遗传信息。 除了RNA的编辑之外,有些RNA,特 别是前体rRNA和tRNA,还可能有位点 特异性的化学修饰。 mRNA Transport • Once processed, mRNA is packaged and exported from the nucleus into the cytoplasm for translation Movement from the nucleus to the cytoplasm is an active and carefully regulated process. The damaged, misprocessed and liberated introns are retained in the nucleus and degraded. • A typical mature mRNA carries a collection of proteins that identifies it as being ready for transport. • Export takes place through the nuclear pore complex. • Once in the cytoplasm, some proteins are discarded and are then imported back to the nucleus for another cycle of mRNA transport. Some proteins stay on the mRNA to facilitate translation. 核 酶(ribozyme) 核酶是指一类具有催化功能的RNA分子, 通过催化靶位点RNA链中磷酸二酯键的 断裂,特异性地剪切底物RNA分子,从 而阻断基因的表达。 ribozyme是核糖核酸和酶两个词的缩合 词。 核酶的锤头结构 • 该酶包括三个茎环区,其间有一个11-13个保守核苷酸 构成的催化中心。 • N代表该位置上可以为任意碱基, • 箭头代表切割部位, • H代表除了G以外的核苷酸。 • 在类病毒中,茎III的顶部并没有被环连接起来。 锤头进行自我切割 •类病毒和拟病毒形成 一种锤头状二级结构, 具有催化活性。 •当酶链被引入细胞时, 它可与底物链配对, 并将其切割。 核酶的生物学意义 核酶的发现使我们对RNA的重要功能又有了新的 认识。 核酶是继逆转录现象之后对中心法则的又一个重 要修正,说明RNA既是遗传物质又是酶。 核酶的发现为生命起源的研究提供了新思路,也 许曾经存在以RNA为基础的原始生命。照这么说, 蛋白质世界也可能(仅仅是可能)起源于RNA世 界! 思考题 1.什么是编码链?什么是模板链? 2.简述RNA转录的概念及其基本过程。 3.大肠杆菌的RNA聚合酶由哪些组成成分?各个亚基的作用如何? 4.什么是封闭复合物、开放复合物以及三元复合物? 5.简述σ因子的作用。 6.什么是Pribnow box?它的保守序列是什么? 7.什么是上升突变?什么是下降突变? 8.简述原核生物和真核生物mRNA的区别。 9.大肠杆菌的终止子有哪两大类?请分别介绍一下它们的结构特点。 10.真核生物的原始转录产物必须经过哪些加工才能成为成熟mRNA,以 用作蛋白质合成的的模板。 11.简述I、II类内含子的剪接特点。 12.什么是RNA编辑?其生物学意义是什么? 13.核酶具有哪些结构特点?其生物学意义是什么?