ppt - Neurodynamics Lab
advertisement

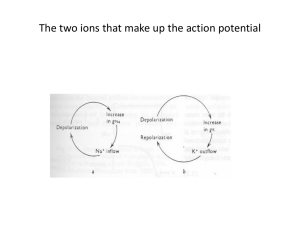
BME 6938 Neurodynamics Instructor: Dr Sachin S Talathi Cable Equation: Transient Solution ~ dV (X,T) ¶ 2V (X,T) + V (X,T) = i ext (X,T) 2 dT ¶X Green’s Function G(X,T) ~for infinite cable: solution of above equation for: i ext = (a /2)d (X).d (T) With initial condition: V(X,0) = (a /2)d (X) and Boundary condition: G(X) = C.Q(T)e æ X2ö -çT + 4T ÷ø è C =a p t General Solution to Cable Equation: t V (X) = ¥ ~ ò dT' ò dX'G(X - X';T - T') i -¥ -¥ Hint: Use the formula: ext (X - X';T - T') Graphical representation Two points worth noticing: 1.The potential is described as a Gaussian function centered at the site of current injection that broadens and shrinks in amplitude with time 2. Membrane potential measured from further location reaches its maximum value at later time Ralls Model-Equivalent cylinder Ralls Assumptions: 1. All membrane properties are the same 2. All terminal branches end with same boundry condition 3. All terminal branches end at same electrotonic distance from soma 4. At every branch point 3/2 rule is obeyed 5. Any dendritic input must be delivered proportionally to all branches at a given electrotonic distance Practical Situation: Choose L=average eletrotonic length of all dendritic branches and Also read the classic papers by Rall (posted on the web: Rall_1959, Rall_1962, Rall_1973) Synaptic Integration Model for current injection into neuron through synapse-alpha function Imp Points to Note: 1. Distal synaptic input produces measurable signals in the soma 2. Obvious fact, the measured EPSP at soma due to stimulation at distal input is smaller as compared to closer to soma 3. Rise time to peaks is progressively delayed for inputs at increasing distance from soma 4. Decay times of all the inputs is the same, ie. Potential cannot decay slower than membrane time constant We know now that dendrites are not passive, what is the current view on dendritic integration. Read the 2 review articles (Magee_2000, London_2005) posted on the website Nonlinear Membrane: dV( x,t) d ¶ 2V( x,t) CM + Iion (V( x,t),t) = + i ext ( x,t) 2 dt 4RA ¶x I M (V( x,t),t) = å I iion i Ions: Na+, K+, Ca2+, Cl- Re-visiting Goldman Equation: The idea of time dependent conductance zF V= Vm RT V Na + zF = VNa + RT Na+ reversal potential (Nernst Equilibrium Potential) Membrane voltage and time dependent ion channel conductance First order approximation The Gate Model HH proposed the gate model to provide a quantitative framework for determining the time and membrane potential dependent properties of ion channel conductance. The Assumptions in the Gate Model: Membrane comprise of aqueous pores through which the ions flow down their concentration gradient These pores contain voltage sensitive gates that close and open dependent on trans membrane potential The transition from closed to open state and vice-versa follow first order kinetics with rate constants: and I have posted the original paper by HH where they proposed to above model and the experiments they conducted to develop the now famous HH model for neuron membrane dynamics (HH_1952). I urge all of you to read this classic work; that led them to receive Nobel Prize in Medicine (The only Noble Prize so far that the field of computational neuroscience have produced) Kinetics of Gate Transition Let P represent the fraction of gates within the ion channel that are open at any given instant in time 1-P then represents the fraction that are closes at that instant in time If and are the rate constants we have Open P Closed 1-P Steady State: Steady state implies Multiple Gates: If a ion channel is comprised of multiple gates; then each and every gate must be open for the channel to conduct ion flow. The probability of gate opening then is given by: Gate Classification Activation Gate: P(t,V) increases with membrane depolarization Inactivation Gate: P(t,V) decreases with membrane depolarization The unknowns In order to use the gate model to describe channel dynamics of cell membrane HH had to determine the following 3 things Macro characteristics of channel type The number and type of gates on each channel The dependence of transition rate constants on membrane voltage V and The Experiments Two important factors permitted HH analysis as they set about to design experiments to find the unknowns Giant Squid Axon (Diameter approx 0.5 mm), allowed for the use of crude electronics of 1950’s (Squid axon’s utility for of nerve properties is credited to J.Z Young (1936) ) Development of feed back control device called the voltage clamp capable of holding the membrane potential to a desired value Before we look into the experiments; lets have a look at the model proposed by HH to describe the dynamics of squid axon cell membrane The HH model HH proposed the parallel conductance model wherein the membrane current is divided up into four separate contributions Current carried by sodium ions Current carried by potassium ions Current carried by other ions (mainly chloride and designated as leak currents) The capacitive current We have already seen this idea being utilized in GHK equations The Equivalent Circuit Current flows assumes Ohms Law Goal: Find and Example: Time and Voltage dependence of Potassium channel conductance GK (V,t) GNa (V,t) The Experiments Space Clamp: Eliminate axial dependence of membrane voltage Stimulate along the entire length of the axon Can be done using a pair of electrodes as shown Provides complete axial symmetry Result: Eliminate the axial component in The cable equation CM dV(x,t) + Iion (V(x,t),t) = i ext (x,t) dt Voltage Clamp: Eliminate Capacitive Current http://www.sinauer.com/neuroscience4e/animations3.1.html Example of Voltage Clamp Recording Clamped Voltage=20 mV The sum of parts Note the different time scale Series of Voltage Clamp Expt Selective blocking with pharmacological agents TTX: Tetrodotoxin; selectively blocks sodium channels TEA: Tetraethylamonium; selectively blocks potassium channels H-H experiments to test Ohms law Foundation of Cellular Neurophysiology, Johnston and Wu HH measurement of Na and K conductance Maximum conductance Gating variables Functional fitting to gate variable We see from last slide Na comprise of activation and inactivation K comprise of only activation term HH fit the the time dependent components of the conductance such that Activation gate Inactivation gate m,n and h are gate variables and follow first order kinetics of the gate model Gate model for m,n and h Activation: Inactivation: Determining and Determine and Use the following relationship Do empirical curve fitting to obtain Profiles of fitted transition functions Summary of HH experiments Determine the contributions to cell membrane current from constituent ionic components Determine whether Ohms law can be applied to determine conductances Determine time and voltage dependence of sodium and potassium conductances Use gate model to fit gating variables Use equations from gate model to determine the voltage dependent transition rates The complete HH model