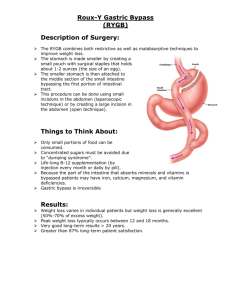
Mustansiriyah College of Medicine Digestive System Physiology Objectives after studying this chapter, you should be able to . . . 1. Describe the gross and microscopic anatomy and the basic functions of the digestive system. 2. Describe the composition and functions of saliva, and explain how salivation is regulated. 3. Describe the mechanisms of chewing and swallowing. 4. Explain how gastric secretion and stomach motility are regulated. 5. Explain the mechanism of vomiting. 6. Describe the role of the gallbladder and state the role of bile in digestion and describe how its entry into the small intestine is regulated. 7. Describe how entry of pancreatic juice into the small intestine is regulated and state the role of pancreatic juice in digestion. 8. List the major functions of the large and small intestine. 9. Describe the regulation of defecation. List the enzymes involved in chemical digestion; name the foodstuffs on which they act. 10. List the end products of protein, fat, carbohydrate, and nucleic acid digestion. 11. Describe the process of absorption of breakdown products of foodstuffs that occurs in the small intestine. 12. Describe the main functions of liver. The digestive system prepares food for use by cells through five basic activities: 1. Ingestion or the taking of food into the body. 2. Peristalsis or the physical movement or pushing of food along the digestive tract. 3. Digestion or the breakdown of food by both mechanical and chemical mechanisms. 4. Absorption or the passage of digested food from the digestive tract into the cardiovascular and lymphatic systems for distribution to the body’s cells. 5. Defecation or the elimination from the body of those substances that are indigestible and cannot be absorbed. 1 Mustansiriyah College of Medicine The primary function of the alimentary tract is to break down food and to provide the body with a continual supply of water, electrolytes and nutrients. In order to achieve these functions, the gastrointestinal tract (GIT) must perform the following processes: ingestion, movement, digestion, absorption, secretion and defecation. The adult GIT is a tube running through the body from mouth to anus. Anatomical features: The organizations of the structures that make up the wall of the gastrointestinal tract (GIT) from the posterior pharynx to the anus generally are as follow from the outer surface inward (figure 5.1): Figure 5.1: Histological cross section of the digestive tract. 1. Serosa which continues onto the mesentery, which contains the nerves, lymphatics, and blood vessels supplying the tract. 2. Longitudinal muscle. 3. Myenteric (Auerbach’s) p1exus which controls mainly the GIT movement. 4. Circular muscle which causes a decrease in the diameter of the lumen of the GI tract when it contracts. 5. Submucous (Meissner’s) plexus which is important in controlling secretion and blood flow and also subserves many sensory functions, receiving signals from chemoreceptors in the gut epithelium and from stretch receptors in the gut wall. 6. Mucosa and submucosa which consists of epithelium and subepithelial connective tissue and are specialized for secretion and absorption. The enteric nervous system (ENS): The GIT has an intrinsic nervous system of its own called the enteric nervous system which controls most GI functions, especially GIT movements and secretion. The enteric nervous system is composed of two layers of neurons and connecting fibers, the outer layer is called the myenteric (Auerbach’s) plexus, the inner layer is called the submucous (Meissner’s) plexus. The degree of activity of this enteric nervous system can strongly be altered by extrinsic (autonomic) nervous system, i.e. parasympathetic and sympathetic nervous systems. Both systems send signals to GIT from the brain and spinal cord to modulate the activity of the enteric nervous system. The parasympathetic nerve fibers: Stimulation of the parasympathetic nerves fibers releases 2 Mustansiriyah College of Medicine acetylcholine and causes in general: An increase in the activity of most GIT functions. Relaxation of sphincters (except the lower esophageal sphincter, which they stimulate). The parasympathetic supply to the gut is divided into: Cranial division: Is mediated almost entirely through the vagus (X cranial nerve). Vagus nerves innervate esophagus, stomach, pancreas, gallbladder and first half of the large intestine, and little innervations to the small intestine Sacral divisions: The sacral divisions originate in S2, 3, 4 sacral segments of the spinal cord, and pass through the pelvic nerves to the distal half of the large intestine. These fibers function especially in the defecation reflex. Because the parasympathetic nervous system plays a dominant role in the digestive process, it is often referred to as the “rest and digest”division. The sympathetic nerve fibers: The sympathetic nerve endings secrete norepinephrine. In general, stimulation of the sympathetic nervous system: Inhibits activity in the GIT Contraction of sphincters, causing effects essentially opposite to those of the parasympathetic system. The sympathetic nerve fibers to the GIT originate in the spinal cord between the segments T- 8 and L 2. The preganglionic fibers after leaving the cord pass through the sympathetic chains to outlying ganglia, such as the celiac, hypogastric, and mesenteric ganglia. Here, the postganglionic neuron bodies are located, and postganglionic fibers spread from them along with the blood vessels to all parts of the gut, terminating principally on neurons of the enteric nervous system. Reflexes of GIT are either occur entirely within the enteric nervous system (short reflex arc), or the reflex arc is originated from the gut and to the CNS (spinal cord or brain stem) and then back to the gut (long reflex arc) (figure 5.2). Signals transmitted through these reflexes can cause reflex excitation or inhibition of intestinal movements or secretion. In addition to its anatomic complexity, the ENS utilizes many different neurotransmitters, including acetylcholine, adenosine triphosphate (ATP), nitric oxide, and numerous peptides. Hirschsprung’s disease is a congenital absence of the myenteric plexus, usually involving a portion of the distal colon. The pathologic non-ganglionic section of large bowel lacks peristalsis and undergoes continuous spasm, leading to a functional obstruction. The normally innervated proximal bowel dilates as a result of the obstruction and can lead to the most feared complication of Hirschsprung’s disease, toxic megacolon. Types of GIT smooth muscle contractions: The smooth muscle of GIT is almost exclusively unitary smooth muscle while that of pharynx, upper one third of the esophagus, and external anal sphincter are striated muscles. Although the upper esophageal sphincter surrounds the upper part of the esophagus consists of skeletal muscle, but is not under conscious or voluntary control. There are two main types of smooth muscle contractions: [A] Phasic (rhythmical) contractions [B] Tonic contractions [A] Phasic (rhythmical) contractions that occur in the esophagus, stomach, and intestine. They include 3 Mustansiriyah College of Medicine peristaltic (figure 5.3 A), migrating Motor Complex (MMC), segmentation and haustration contractions (figure 5.3 B). In peristalsis, contraction of a small section of proximal muscle is followed immediately by relaxation of the muscle just distal to it. The resulting wavelike motion moves food along the GIT in a proximal to distal direction. Peristaltic wave could be of two types: [1] Primary peristaltic wave which is a continuation of the peristaltic wave that begins proximally and mediated by vagovagal reflexes, long reflex arc. [2] Secondary peristaltic wave which is generated by the enteric nervous system (short reflex pathway), initiated from the distension of the viscus by the retained food if the primary peristaltic wave fails to move all the food that has entered the viscus. MMC is a distinct pattern of electromechanical activity observed in gastrointestinal smooth muscle during the periods between meals, and it is interrupted by feeding. It is thought to serve a "housekeeping" role and sweep residual undigested material through the digestive tube. The cycle recurs every 1.5 to 2 hours. These motor complexes trigger peristaltic waves, which facilitate transportation of indigestible substances such as bone, fiber, and foreign bodies from the stomach, through the small intestine, past the ileocecal sphincter, and into the colon. An intact enteric nervous system is essential for propagation of migrating motor complexes along the bowel. The MMC is thought to be partially regulated by motilin (by a direct action on smooth muscle and by activation of excitatory enteric nerves and it is released during the interdigestive period). MMC is initiated in the stomach as a response to vagal stimulation. Control of the MMC is complex. MMC with an antral origin can be induced in humans through intravenous administration of motilin, erythromycin or ghrelin, whereas administration of serotonin or somatostatin induces MMC with duodenal origin. The role of the vagus nerve in control of the MMC seems to be restricted to the stomach, as vagotomy abolishes the motor activity in the stomach, but leaves the periodic activity in the small bowel intact. In segmentation contraction, nonadjacent segments of alimentary tract organs, especially small intestine, contract and relax, moving food forward then backward, resulting in food mixing and slow food propulsion. Segmentation contraction is the primary motility pattern of the digestive period in humans, and is defined as irregular and uncoordinated contraction of the circular muscle layer. This pattern, which develops in response to intestinal wall distention, is determined by activation of preprogrammed neural circuits within the myenteric plexus. The combined contractions of the circular and longitudinal smooth muscle cause the unstimulated portion in large intestine to bulge outward into baglike sacs called haustration. Therefore, the fecal material in the large intestine is squeezed, moving back and forth along the colon in a manner similar to that for the segmentation contractions in the small intestine. Distention-induced contraction of gastrointestinal smooth muscle develops as the result of long (vago-vagal) and local (enteric nerves) reflexes. The importance of long versus local reflex pathways varies along the gut. Secondary esophageal peristalsis, intestinal segmentation, and migrating motor complexes are unaffected by vagotomy, whereas primary peristaltic wave and proximal stomach peristalsis is decreased but not abolished by vagotomy. [B] Tonic contractions that occur in the lower esophageal sphincter, pyloric sphincter, ileocecal sphincter, and internal anal sphincter. Tonic contraction is continuous, occasionally increases or decreases in intensity. Electrical slow wave activity in the gastrointestinal tract appears to originate from specialized 4 Mustansiriyah College of Medicine pacemaker cells (interstitial cells of Cajal) located between the longitudinal and circular muscle layers and within the submucosal and myenteric plexuses. These pacemaker cells input directly onto the smooth muscle and receive input from enteric nerves. Figure 5.2: Arrangement of reflexes of the digestive system. Table 5A: Differences Between the Myenteric and Submucosal Plexuses Myenteric Plexus Submucosal Plexus It is concerned mainly with controlling muscle activity It is mainly concerned with controlling function along the length of the gut with specific effects: within the inner wall of each minute segment (i) Increased “tone" of the gut wall of the intestine such as: (ii) Increased intensity of the rhythmical contractions (i) local intestinal secretion (iii) Slightly increased rate of the rhythm of (ii) local absorption contraction (iii) local contraction of the submucosal muscle (iv) Increased velocity of conduction of excitatory waves along the gut wall, causing more rapid movement of the gut peristaltic waves Composed of excitatory as well as inhibitory neurons Inhibitory neurons are useful for inhibiting some of the intestinal sphincter muscles such as pyloric sphincter and the sphincter of the ileocecal valve Composed mainly of excitatory neurons Different Types of Neurotransmitters Secreted by Enteric Neurons 1. Acetylcholine 2. Norepinephrine 3. Adenosine triphosphate 4. Serotonin 5. Dopamine 6. Cholecystokinin 7. Substance P 8. Vasoactive intestinal polypeptide 9. Somatostatin 10. Leu-enkephalin 5 Mustansiriyah College of Medicine 11. Met-enkephalin 12. Bombesin The specific functions of many of these are not known well enough to justify discussion here, other than to point out the following. Acetylcholine most often excites gastrointestinal activity. Norepinephrine and epinephrine almost always inhibit gastrointestinal activity. Figure 5.3: Types of GIT smooth muscle contractions. Paralytic ileus is a temporary cessation of gut motility that is most commonly caused by abdominal surgery. Other common causes appendicitis, hypokalemia, and narcotics. Signs and symptoms of paralytic ileus include nausea and vomiting, abdominal distension, and absent bowel sounds. 6 Mustansiriyah College of Medicine Mastication (chewing): Is the process by which food taken into the mouth is chewed by the teeth. The chewing reflex (figure 5.3A) is controlled by nuclei in the medulla and cerebral cortex. Most of the muscles of chewing are innervated by the motor branch of the 5th cranial nerve (trigeminal N). Voluntary contraction of the muscles of chewing is in response to the presence of a bolus of food in the mouth compresses the bolus against the linings of the mouth and push the food to come in contact with the buccal receptors. The contact of food with buccal receptors causes reflex inhibition of the muscles of mastication, which allows the lower jaw to drop. The drop in turn initiates a stretch reflex of the jaw muscles (the masseter, medial pterygoid, and temporalis Figure 5.3A: The chewing reflex. muscles) that leads to rebound contraction. This automatically raises the jaw to cause closure of the teeth, but again, it also compresses the bolus against the linings of the mouth and pushes the food to come in contact with the buccal receptors, which inhibit the jaw muscles once again and allowing the jaw to drop and rebound another time, and this is repeated again and again. Chewing is important because: [1] Chewing breaks down the indigestible cellulose membranes around the nutrient portions of food before the food can be utilized. [2] Chewing increases the surface area of food so the rate of digestion of food by the digestive enzymes is increased. [3] Chewing grinds the food to a very fine particulate consistency prevents excoriation of the GIT and increases the ease with which food is swallowed and emptied from the stomach into the small intestine and thence into all succeeding segments of the gut. [4] Chewing mixes the food with salivary gland secretions which initiates the process of starch digestion (by salivary amylase) and to much less extent of lipid digestion (by lingual lipase) and to lubricate and soften the bolus of food, making it easier to swallow. [5] Chewing brings food into contact with taste receptors and releases odors that stimulate the olfactory receptors. The sensations generated by these receptors increase the pleasure of eating and initiate gastric secretions. 7 Mustansiriyah College of Medicine Salivary glands: The daily normal secretion of saliva is between 500—1500 ml. Saliva is secreted from the glands of salivation, the parotids (25%), submandibular glands (70%), and sublingual glands (5%). In addition, there are many small buccal glands. In comparison to plasma, saliva is: Hypotonic. Contains higher concentration of K ions and bicarbonate ions and lower concentrations of Na and Cl ions. Saliva has a pH 6.0 - 7.4. The structure of each gland is similar to a bunch of grapes (figure 5.4). The acinus is lined with acinar cells and secretes initial saliva which is a plasma-like solution (isotonic) containing amylase and/or mucin. A branching duct system is lined with columnar epithelial cell, which modify the initial saliva by: Active absorption of Na+ (3Na+-2K+ Figure 5.4: Secretion of saliva and electrolytes. ATPase), Passive reabsorption of Cl- ions (due to electrical gradient) in exchange to HCO3- ions which are secreted actively (Cl- - HCO3- secondary active counter-transport). Saliva is an important route of iodide excretion; its concentration in saliva is 20–100 times that in plasma. It is saturated with calcium ions; calcium salts are laid down as plaque on the teeth. Therefore, the composition of saliva varies with the salivary flow rate: [1] At the lowest flow rates, saliva has the lowest osmolarity (about 100 mOsm/L) and lowest Na+, and Clconcentrations, but has the highest K+ and HCO3- ion concentration. [2] At the highest flow rates, saliva is most like the initial secretion from the acinus and its composition is closet to that of plasma. Saliva consists of two major types of protein secretions: 1. Serous secretion (watery saliva) containing ptyalin (α-amylase which is an enzyme for digesting starches) and electrolytes. 2. Mucous secretion containing mucin (glycoprotein) for lubricating purposes. The parotid glands secrete entirely the serous type. The submandibular and sublingual glands secrete both the serous type and mucus. The buccal glands secrete only mucus. Control of salivary secretion: Salivary glands are controlled by salivatory nuclei which are located at the juncture of the medulla and pons. 8 Mustansiriyah College of Medicine • Mainly through parasympathetic fibers through 7th cranial nerve (Facial N) and 9th cranial nerve (Glossopharyngeal N) abundant watery saliva rich in enzymes. • And to less extent by sympathetic nervous signals originates from the superior cervical ganglia and then travels along blood vessels to the salivary glands scanty thick saliva rich in mucus. This is why the mouth may feel sticky or dry under conditions of stress. Under basal conditions, saliva is almost entirely of the mucus type and is secreted all the time except during sleep Figure 5.4A: Control of salivary secretion. when the secretion becomes very little. Unlike the autonomic nervous system elsewhere in the body, sympathetic and parasympathetic responses in the salivary glands are not antagonistic. Salivatory nuclei are excited or inhibited by (figure 5.4A): [A] Signals from the mouth: Taste (especially the sour taste excitatory, bitter taste inhibitory) and tactile stimuli (especially smooth objects excitatory) from the tongue and other areas of the mouth. [B] Signals from higher centers of CNS: Salivation can also be stimulated or inhibited by impulses arriving in the salivatory nuclei from higher centers of CNS. For instance, when a person smells or sees favorite foods, salivation is greater than when disliked food is smelled or seen. [C] Signals from GIT: Salivation also occurs in response to reflex originating in the stomach and upper intestine particularly when very irritating foods are swallowed or when a person is nauseated. The swallowed saliva may help to remove the irritating factor in the GIT by diluting or neutralizing the irritant substances. Saliva production is decreased (via inhibition of the parasympathetic nervous system) by sleep, dehydration, fear, and anticholinergic drugs (such as atropine). Functions of saliva: [A] Protection of the mouth by: Cooling hot foods, By maintaining healthy oral tissues. The mouth is loaded with pathogenic bacteria that can easily destroy tissues and can also cause dental caries. However, saliva helps prevent the harmful effects of bacteria by: Continuous washing away the pathogenic bacteria as well as the food particles that provide the bacteria with metabolic support. 9 Mustansiriyah College of Medicine Saliva contains many factors that can kill bacteria such as thiocyanate ions, proteolytic enzymes, and antibodies (IgA). [B] Digestion of starch by α-amylase (which is ultimately inactivated by the low pH of the stomach), and fat by lingual lipase. [C] Lubrication of food and making swallowing easier, and moisten the mouth, facilitating speech. It also neutralizes any gastric acid that refluxes from stomach into the lower esophagus (heartburn). Xerostomia is defined as dry mouth resulting from reduced or absent saliva flow. Xerostomia is not a disease, but it may be a symptom of various medical conditions, a side effect of a radiation to the head and neck, or a side effect of a wide variety of medications. Xerostomia is a common complaint affecting approximately 20 percent of the elderly. Swallowing (deglutition): The voluntary initiation of swallowing takes place in higher brain centers and starts with oral stage. From here on, the process of swallowing becomes entirely or almost entirely automatic and controlled by swallowing (or deglutition) center which is located at the medulla and the pons. The afferent (sensory) fibers to this center are: Trigeminal (5th cranial nerves), Glossopharyngeal (9th cranial nerves), The efferent (motor) nerve fibers from the swallowing center are conducted through: Trigeminal (5th cranial nerves), Glossopharyngeal (9th cranial nerves), Vagus (10th cranial nerves), Hypoglossal (12th cranial nerves), Few of the superior cervical nerves to the pharynx and upper esophagus Swallowing can be divided into the following stages: Oral stage. Pharyngeal stage. Esophageal stage. Relaxation of lower esophageal sphincter. [A] Oral (voluntary) stage of swallowing: Food is voluntarily squeezed posteriorly in the mouth to the oropharynx by pressure of the tongue upward and backward against the hard palate. From here on, the process of swallowing becomes entirely or almost entirely automatic and cannot be stopped. [B] Pharyngeal (involuntary) stage: When the bolus of food is pushed backward in the mouth, it stimulates swallowing receptor areas around the opening of the pharynx afferent impulses pass to Swallowing center to initiate a series of automatic pharyngeal muscular contractions and to inhibit the respiratory center of the medulla during swallowing, halting respiration at any point in its cycle to allow swallowing to proceed. The sequence of events of the pharyngeal stage of the swallowing is as follows: 1. The soft palate is pulled upward to close the posterior nares preventing reflux of food into the nasal cavities. 2. The palatopharyngeal folds on either side of the pharynx are pulled medial ward to approximate each other. In this way these folds form sagital slit through which the food must pass into the posterior pharynx. This slit performs a selective action, allowing food that has been masticated properly to pass with 10 Mustansiriyah College of Medicine ease while impede the passage of large objects. 3. The vocal cords of the larynx are strongly approximated, and the hyoid bone and larynx are pulled upward and anteriorly by the neck muscles causing epiglottis to swing backward over the superior opening of the larynx. All these effects (especially the approximation of vocal cords) prevent passage of food into the trachea. 4. The upward movement of the larynx also stretches the opening of the esophagus. At the same time, the upper 3 to 4 cm of the esophagus, an area called the upper esophageal sphincter (or pharyngeoesophageal sphincter or the cricopharyngeal muscle), relaxes, thus allowing food to move easily and freely from the posterior pharynx into the upper esophagus. This sphincter between swallows, remains tonically and strongly contracted (by the continual firing of the vagal fibers), thereby prevents air from going into the esophagus during respiration. 5. At the same time that the larynx is raised and the upper esophageal sphincter is relaxed, the superior constrictor muscle of the pharynx contracts, giving rise to a rapid peristaltic wave passing downward over the pharyngeal muscles and into the esophagus, which also propels the food into the esophagus. Swallowing becomes a great concern for the elderly since strokes and Alzheimer's disease can interfere with the autonomic nervous system. As you can gather from the information written above, there are multiple areas of the central nervous system which, if affected by a stroke, could disrupt the ability to swallow. This is especially true for strokes of the medulla, as this is a relatively small area that contains multiple structures that are critical in carrying out the swallowing reflex. In fact, people with medullary strokes frequently require temporary or permanent feeding tube placement. [C] Esophageal stage of swallowing: The esophagus functions to conduct food from the pharynx to the stomach. The movement of food down the esophagus is an active process (primary esophageal peristalsis integrated at swallowing center) that does not depend on gravity for its normal function. The upper esophageal sphincter and lower esophageal sphincter are closed in normal resting state. Closure of the upper esophageal sphincter prevents air from being drawn from the mouth into the esophagus during inspiration. Closure of the lower esophageal sphincter prevents gastric contents to move back from the stomach to the esophagus. Because most of the esophagus is contained within the thoracic cavity, the pressure in the lumen of the resting esophagus is approximately equal to intra-thoracic pressure (atmospheric or slightly below atmospheric) when the upper esophageal sphincter and lower esophageal sphincter are closed (normal resting state). During pulmonary inspiration, the intra-esophageal pressure will be sub-atmospheric. The term secondary esophageal peristalsis describes esophageal peristalsis and lower esophageal relaxation associated with distension or irritation of the smooth muscle portion of the esophageal body. The event is limited to the smooth muscle component of the esophagus and is the result of activation of enteric nerves. Initiation of secondary peristalsis does not involve extrinsic neural reflexes and, thus, is not accompanied by the oral-pharyngeal phase of swallowing. [D] Relaxation of lower esophageal sphincter: The lower esophageal sphincter is not anatomically separate identifiable muscles. The fundus of the stomach and lower esophageal sphincter extending about 2-5 cm above its junction with the stomach both relax during a swallow while the bolus of food is still higher in the esophagus. This phenomenon is called receptive relaxation. Receptive relaxation is vagally mediated, Nitric oxide is the neurotransmitter thought to mediate receptive relaxation at the smooth muscle cell allowing food to pass to the stomach. 11 Mustansiriyah College of Medicine Reflux of the acidic contents of the stomach up to the esophagus damages the esophageal mucosa. This is prevented by: [1] The muscles of the lower esophageal sphincter normally remains tonically contracted due to tonic activity of the vagal fibers innervating the esophagus (in contrast to the mid- and upper portions of the esophagus which normally remain completely relaxed). Vagal efferent nerve fibers control contraction and relaxation of the LES. Vagal excitatory fibers synapse on post-ganglionic, enteric motor neurons that release acetylcholine within the musculature of the LES. Vagal inhibitory fibers synapse on enteric inhibitory motor neurons that release nitric oxide (NO). These neurons may also utilize ATP and VIP as inhibitory transmitters, but NO is the primary inhibitory neurotransmitter. Sympathetic neurons also provide excitatory input to the LES via excitatory motor neurons. [2] Another factor that prevents reflux is the oblique entrance of the esophagus to the stomach which acts like a valve-like mechanism of that portion of the esophagus that lies immediately beneath the diaphragm. Greatly increased intra-abdominal pressure caves the esophagus inward at this point at the same time that the abdominal pressure also increases the intragastric pressure, preventing the high pressure in the stomach from forcing stomach contents into the esophagus. Esophageal reflux: Reflux of stomach acid to the esophagus causes esophageal pain (heartburn) and may lead to esophagitis. An increase in the intraabdominal pressure (by the ingestion of a very large meal, production of intestinal gas, pregnancy, an abdominal mass such as a tumor, bending at the waist, straining against a closed glottis as in defecation) will facilitate gastro-esophageal reflux if the LES is not contracted or there is distortion of the valve-like mechanism. Esophageal reflux may occur if: [A] If the intragastric pressure raises high enough to force the lower esophageal sphincter open. [B] If the lower esophageal sphincter is unable to maintain its normal tone. [C] If the lower esophageal sphincter is forced through the diaphragm and into the thoracic cavity as in hiatal hernia, in which the oblique entrance of the esophagus to the stomach is distorted, and consequently the valve-like mechanism is impaired. Belching (eructation): Following a heavy meal or the ingestion of large amounts of gas (e.g., from carbonated beverages), the gas bubble that is usually in the fundus of the stomach is displaced to the cardia. When lower esophageal sphincter relaxes during the swallowing process, gas enters the esophagus and is regurgitated. Dysphagia: Difficulty in swallowing. Persons with dysphagia usually report choking, coughing, or an abnormal sensation of food sticking in the back of the throat or upper chest when they swallow. If swallowing is painful, it is referred to as odynophagia. Dysphagia can result from altered nerve function or from disorders such as: Narrowing of the esophagus. 12 Mustansiriyah College of Medicine Lesions of the central nervous system (CNS), such as a stroke, often involve the cranial nerves that control swallowing. Strictures and cancer of the esophagus and strictures resulting from scarring can reduce the size of the esophageal lumen and make swallowing difficult. Achalasia: In which the lower esophageal sphincter fails to relax; food that has been swallowed has difficulty passing into the stomach, and the esophagus above the lower esophageal sphincter becomes enlarged. One or several meals may lodge in the esophagus and pass slowly into the stomach. There is danger of aspiration of esophageal contents into the lungs when the person lies down. The stomach: The stomach can be divided into a. the fundus, b. the body, and c. the antrum. Physiologically the fundus functions mainly as part of the body. The lining epithelium of the stomach mucosa is a simple columnar epithelium composed entirely of mucous cells. They produce a cloudy, protective two-layer coat of alkaline mucus in which the surface layer consists of viscous, insoluble mucus that traps a layer of bicarbonate-rich fluid beneath it. This otherwise smooth lining is dotted with millions of deep gastric pits, which lead into tubular gastric glands that produce the stomach secretion called gastric juice. The functions of the stomach: 1. Storage: Is mediated by the process of receptive relaxation of the stomach until the food can be delivered to the small intestine at the proper time. Normally, when food enters the stomach a vago-vagal reflex greatly reduces the tone in the muscular wall of the body of the stomach (active relaxation of smooth muscles, vagally mediated, Nitric oxide is the neurotransmitter thought to mediate receptive relaxation at the smooth muscle cell) so that the wall can bulge progressively outward accommodating greater and greater quantities of food up to a limit of about 1 liter. Stomach accommodation depends exclusively upon an intact vago-vagal reflex. If vagal innervation is interrupted, then intra-gastric pressure increases. This is a potential cause of vomiting due to the inability of the proximal stomach smooth muscle to undergo receptive relaxation. 2. Mixing and propelling of food: When the stomach is filled, weak peristaltic and segmentation (mixing) contractions move toward the antrum along the stomach wall, propelling and mixing food with gastric juice until it forms a semifluid mixture called chyme. This is due to distension of the stomach which elicit vagovagal reflex. 3. Emptying: Slow emptying of chyme from the stomach into the small intestine at a rate suitable for proper digestion and absorption by the small intestine. The degree of constriction of the pyloric sphincter and the intensity of antral peristaltic wave (mediated by myenteric reflexes) can be varied according to signals both, from the stomach and from duodenum. The antral peristaltic waves provide a pumping action and are frequently called the pyloric pump. The stomach is a poor absorptive area of the gastrointestinal tract because it lacks the typical villus type of absorptive membrane, and also because the junctions between the epithelial cells are tight junctions. Only a few highly lipid-soluble substances, such as alcohol and some drugs like aspirin can be absorbed in small quantities. Regulation of gastric emptying (pyloric pump): Gastric emptying is a key control point in the gastrointestinal tract to ensure the orderly delivery of nutrients in a form that can be digested and to give appropriate signals of fullness (satiety). Gastroparesis (“weak stomach”) is a common complication of poorly controlled diabetes mellitus and significantly slows gastric emptying. The rate at which the stomach empties is regulated by signals both from the stomach and duodenum. These signals will be discussed later in association with the factors that affect gastric secretion. Gastric emptying takes about 3 13 Mustansiriyah College of Medicine hours and very closely regulated so that nutrient absorption is maximized and H+ in the duodenum has time to be neutralized. Gastric secretion Gastric secretions aid in the breakdown of food into small particles and continue the process of digestion which had begun by the salivary enzymes. About 2 L / day of gastric secretions are produced. The stomach mucosa contains two main types of gastric glands (figure 5.5): [A] Gastric glands which are located in the fundus and the body of the stomach. They contain three types of secretory cells: 1. Mucus secreting cells which secretes mucus. 2. Parietal (oxyntic) cells which secrete intrinsic factor and HCl. 3. Peptic (chief) cells which secrete pepsinogen, the precursor for the proteolytic enzyme pepsin. 4. Enteroendocrine cells (or Figure 5.5: Histological cross section of gastric mucosa. enterochromaffin-like cells, ECL cell) release a variety of chemical messengers directly into the interstitial fluid of the mucosa of the stomach. Some of these are histamine, serotonin, and somatostatin. [B] Pyloric glands which are located in the antral and pyloric regions of the stomach. They contain: G cells and some mucous cells, G cells are responsible for the release of the hormone gastrin. D cells, which release somatostatin, a hormone that inhibits the release of gastrin. Gastric HCl secretion: HCl is secreted into the parietal cell canaliculi by the following steps (figure 5.6): [1] CO2 diffuses from blood to inside the parietal cells. [2] Within the parietal cells, carbonic acid is formed from the reaction: CO 2 + H2O H2CO3. The formation of H2CO3 from CO2 is catalyzed by the enzyme carbonic anhydrase. [3] HCO3- diffuses back into the plasma in exchange for Cl-, thus providing Cl- for the initial step in the secretory process. As HCO3- is added to the venous blood, the pH of the blood drained from the stomach increases (alkaline tide). The active transport process is begun by the transport of Cl - ion into the canaliculi that open to the lumen of the stomach. [4] The H+ that is supplied by the dissociation of carbonic acid into H+ and HCO3- within the parietal cells is exchanged for K+ by the H+-K+-ATPase pump (proton pump). [5] Chloride ions diffuse with the charged H+. [6] Water enters the canaliculi down the osmotic gradient created by the movement of HCl into the canaliculi. Most of the HCl that is secreted into the stomach is neutralized and reabsorbed within the small intestine. However, if the gastric contents are lost before they enter the small intestine as in case of 14 Mustansiriyah College of Medicine vomiting, sever alkalosis may ensue. The pH of the parietal cell secretion can be as low as 0.8 (or almost 4 million times as great as the H+ concentration of plasma). Parietal cells bear receptors for three potent stimulators of acid secretion, reflecting a triad of neural, paracrine and endocrine control: Acetylcholine (muscarinic type receptor) Gastrin Histamine (H2 type receptor) A variety of substances are capable of reducing gastric acid secretion including: Prostaglandin E2 (PGE2), Several peptides hormones, including Secretin, Gastric inhibitory polypeptide (GIP), Glucagon and, Somatostatin. GIP (also known as the glucose-dependent insulinotropic peptide) released from duodenal and jejunal mucosa in response to the presence of chyme especially by hyperosmolarity of glucose in the duodenum and inhibits gastric gastrin release and stimulate the release of insulin from pancreas, and inhibit the GI motility and secretion of acid. The amount of insulin secreted is greater when glucose is administered orally than intravenously. It is the only gastrointestinal hormone released by all three major foodstuffs (fats, proteins, and carbohydrates). PGE2, secretin and somatostatin may be physiologic regulators. Somatostatin inhibits secretion of gastrin and histamine, and appears to have a direct inhibitory effect on the parietal cell. 15 Mustansiriyah College of Medicine Figure 5.6: Steps of gastric HCl secretion. The H+-K+-ATPase pump can be inhibited by the drug omeprazole, which is now used for the treatment of duodenal and gastric ulcers (peptic ulcers). Inhibition of pump activity leads to a prolonged increase in gastric pH and the removal of the inhibitory effect of low pH (<3.0) on gastrin release. The hypergastrinemia that develops may actually increase the number of parietal cells as a result of the trophic effects of the hormone. In addition, two other drugs can also be used for treatment of peptic ulcer and these are cimetidin and ranitidine. They accomplish this by two mechanisms: (1) Histamine released by ECL cells in the stomach is blocked from binding on parietal cell H2 receptors, which stimulate acid secretion; (2) therefore, other substances that promote acid secretion (such as gastrin and acetylcholine) have a reduced effect on parietal cells when the H2 receptors are blocked. Mild injury to the mucosal barrier results in increased mucus secretion and surface desquamation followed by regeneration. A more serious injury breaks mucosal barrier and exposes the mucosal surface, forming an ulcer, and produces bleeding. Breaks of mucosal barrier and exposure of the mucosal surface to damage occurs due to highly concentrated HCl, 10% ethanol, salicylic acid, or acetylsalicylic acid (aspirin). The damaged mucosa liberates histamine, which in turn increases acid secretion and produces increased capillary permeability and vasodilatation. The latter two effects lead to edema. In addition, the exposure of mucosal capillaries to the digestive process leads to bleeding. Erosive gastritis can occur as a result of chronic use of non-steroidal anti-inflammatory drugs (NSAIDs). The mechanism by which NSAIDS cause gastritis involves the inhibition of prostaglandin synthesis 16 Mustansiriyah College of Medicine in the stomach. Prostaglandins normally maintain the physicochemical barrier on the gastroduodenal mucosal surface by stimulating the secretion of mucus and bicarbonate. Loss of the protective mucus and bicarbonate barrier renders the gastric mucosa susceptible to damage by the acidic environment. The functions of HCl: [1] It participates in the breakdown of protein: The high acidity of the chyme alters the protein molecules, thereby changing protein structure so as to break up connective tissue and cells in the ingested food. [2] It hinders the growth of pathogenic bacteria: HCl kills most of the bacteria that enter along with food. This process is not 100% effective, and some bacteria survive to take up residence and multiply in the intestinal tract. [3] It activates pepsinogens and provides an optimal pH for the action of pepsin: Several different types of pepsinogens are secreted by the peptic cells of the gastric glands and all perform the same function. When the pepsinogens are secreted, they have no digestive activity. However, as soon as they come in contact with HCl and especially when they come in contact with previously formed pepsin plus the HCl, they are immediately activated to form active pepsin which is an active proteolytic enzyme in a highly acid medium. Gastric mucus secretion: The surface of the stomach mucosa between glands has a continuous layer of mucous cells that secrete large quantities of a viscid and alkaline mucus that coats the mucosa with a mucous gel layer (the gastric mucosal barrier) often more than 1 mm thick, thus providing a major shell of protection for the stomach wall against acidity as well as contributing to lubrication of food transport. Even the slightest irritation of the mucosa directly stimulates the mucous cells to secrete copious quantities of this thick, viscid mucus. This in turn forms a gastric barrier that prevents Figure 5.7: The components of gastric mucosal barrier. digestion of the gastric wall and also greatly reduces the absorption of most substances by the gastric mucosa. If this barrier is damaged by toxic substances, such as occurs with excessive use of aspirin, and other nonsteroidal anti-inflammatory drugs, or alcohol, the secreted acid does leak down an electrochemical gradient into the mucosa, causing stomach mucosal damage. The components of gastric mucosal barrier enable the stomach to contain acid without injuring itself are (figure 5.7): The luminal membranes of the gastric mucosal cells are impermeable to H+ so that HCI cannot penetrate into the cells. The cells are joined by tight junctions that prevent HCI from penetrating between them. A mucus coating over the gastric mucosa serves as a physical barrier to acid penetration. The HCO3 rich mucus also serves as a chemical barrier that neutralizes acid in the vicinity of the mucosa. Even when luminal pH is 2, the mucus pH is 7. 17 Mustansiriyah College of Medicine Gastric hormonal secretion: Gastrin, main gastrointestinal hormone, is secreted by antral G cells and has the following effects: It stimulates the parietal cells to secret HCl. It stimulates the chief cells to secret pepsinogen. It stimulates the enterochromaffin-like cells (ECL cell, or enteroendorine cells) to release histamine. It enhances muscle contractions of pyloric pump (minor effect) and has a slight constrictor effect on the gastroesophageal sphincter. It stimulates contraction of intestinal muscle, relaxes ileocecal valve, and stimulates mass movements. Other hormones such as histamine, serotonin, and somatostatin release directly into the interstitial fluid of the mucosa of the stomach and act as chemical messengers. Gastric intrinsic factor secretion: A substance called intrinsic factor is secreted by the oxyntic cells, and it is essential for absorption of vitamin B12 in the ileum. If the acid producing cells of the stomach are destroyed as it occurs in chronic gastritis, the person develops achlorhydria and pernicious anaemia because of failure of maturation of the RBC in the absence of vitamins B12. Although intrinsic factor is secreted by the oxyntic cells of the gastric mucosa, binding to cobalamin occurs in the duodenum. The vitamin B12-intrinsic factor complex is propelled along the small intestine to the terminal ileum, where specific transporters located on the enterocyte microvilli bind the vitamin B12-intrinsic factor complex. Binding requires calcium and is optimal at pH 6.6. Absorption is an active transport process. Phases of gastric motility and secretion: Cephalic phase Gastric phase Intestinal phase 1. The cephalic phase: It occurs even before the food enters the stomach (figure 5.8A). It results from the sight, smell, thought, tactile sensation in the mouth, taste of food, or emotional state. Neurogenic signals causing the cephalic phase of secretion can originate in the cerebral cortex or in the appetite centers of the amygdala or hypothalamus and transmitted through the vagi to the stomach (insulin induced hypoglycemia also stimulates the vagus nerve). Psychic states have marked effects on gastric secretion and motility. Anger and hostility were associated with hypersecretion of the gastric mucosa and enhance gastric motility while fear and depression decrease gastric secretion and inhibit gastric motility. Almost 30% of the gastric secretions released during a meal occur as a result of cephalically induced vagal stimulation (figure 5.8). During this phase, the non-distended stomach secretes hormone Ghrelin from cells in the stomach. Ghrelin the "hunger hormone” is a peptide hormone produced by cells in the gastrointestinal tract which functions as a neuropeptide in the central nervous system. It acts on hypothalamic brain cells both to increase hunger, 18 Mustansiriyah College of Medicine and to increase gastric acid secretion and gastrointestinal motility to prepare the body for food intake. When the stomach is empty, ghrelin is secreted. When the stomach is stretched, secretion stops. Ghrelin is also a stimulator of the growth hormone secretagogue receptor in the anterior pituitary, a receptor that mediates growth hormone release. 2. The gastric phase: 60% of gastric secretions are associated with this phase. It starts once the food enters the stomach (see figure 5.8 and 5.8A): [A] The stomach excitatory signals: Distension of the antrum by food through myenteric reflexes, increase in the activity of the pyloric pump and at the same time inhibits the pyloric sphincter. The presence of the protein (small peptides and amino acids or other substance like caffeine) in the diet (the presence of protein digestion products in the diet is the most important determinant of acid secretion during the gastric phase) a reflex stimulation of G cell of mucosa to release gastrin. Consequently increase in gastric secretion and motility of pyloric pump as shown in figure 5.8. Therefore, carbohydrates empty fastest, followed by protein. Fats take longest to empty. decreasing in gastric [H+] (i.e. high pH) [B] The gastric inhibitory signals: The most potent inhibitor of HCl secretion and pyloric pump activity is the drop of pH (increasing in gastric [H+]) of the gastric content below 2-3. This inhibition is mediated by two mechanisms: A low pH directly inhibits G cells and gastrin secretion, consequently inhibits HCl secretion and pyloric pump activity (figure 5.8). Plasma gastrin levels are related inversely to acid secretory capacity because of a feedback mechanism by which antral acidification inhibits gastrin release. Thus, plasma gastrin levels are reduced in persons with duodenal ulcers. A low pH stimulates antral D cells to release somatostatin, which inhibit secretion of parietal cells, ECL cells, and G cells (not shown in figure 5.8), thus turning off the HCl-secreting cells and their most potent stimulatory pathway. 19 Mustansiriyah College of Medicine Facts about Gastric emptying: a. The rate of gastric emptying is fastest when the stomach contents are isotonic. If the stomach contents are hypertonic Figure 5.8: Cells involved in gastric or hypotonic, gastric emptying is slowed. b. Fat inhibits gastric emptying (i.e., increases gastric emptying secretion during cephalic and gastric phases. time) by stimulating the release of CCK. + c. H in the duodenum inhibits gastric emptying via direct neural reflexes. H+ receptors in the duodenum relay information to the gastric smooth muscle via interneurons in the GI plexuses. d. The initial rate of the gastric emptying varies directly with the volume of the meal ingested. Another phase of acid secretion is known as the basal state which occurs in the times between meals (interdigestive phase). The level of acid secretion during these times is regulated by body weight, individual, number of parietal cells, and time of day. Acid secretion is lowest in the morning before awakening and highest at night. The stomach secretes only few milliliters of gastric juice per hour during the interdigestive period (between meals) when little or no digestion is occurring anywhere in the gut which is almost entirely of non-oxyntic type i.e., it is composed mainly of mucus containing very little pepsin and almost no acid. However, strong emotional stimuli frequently increase the interdigestive secretion to 50 ml or more of highly peptic and highly acidic gastric juice per hour in very much the same manner that the cephalic phase of the gastric secretion excites secretion at the onset of a meal. This increase of secretion during the presence of emotional stimuli is believed to be one of the factors in the development of peptic ulcers. 20 Mustansiriyah College of Medicine 3. The intestinal phase: Intestinal receptors monitor the composition of chyme and elicit feedback mechanisms that regulate gastric acid secretion and gastric emptying. Absence of feedback leads to an increased presence of excitatory mediators of gastric function. 10% of gastric secretion is associated with the intestinal phase. This is achieved through hormones and nervous reflexes. The rate at which the stomach empties is regulated by signals from both the stomach and the duodenum. However, the duodenum provides by far the more potent of the signals, controlling the emptying of chyme into the duodenum at a rate no greater than the rate at which the chime can be digested and absorbed in the small intestine. These feedback inhibitory mechanisms work together to slow the rate of emptying when (1) too much chyme is already in the small intestine or (2) the chyme is excessively acidic, contains too much unprocessed protein or fat, is hypotonic or hypertonic, or is irritating. [A] The intestinal excitatory signals: 1. Nervous Feedback: Slight stretching the wall of the upper portion of the small intestine, particularly in the duodenum by food. 2. Hormonal Feedback: The presence of small amounts of food (especially protein digestion products) stimulates small amount Figure 5.9: The role of the intestinal inhibitory signals on of intestinal gastrin to be released gastric secretion and pyloric pump. 21 Mustansiriyah College of Medicine from the duodenal mucosa. This can slightly increase the activity of pyloric pump and gastric juice secretion (through the same mechanism mentioned before). [B] The intestinal inhibitory signals: 1. Nervous Feedback: Duodenal wall distension or irritation of duodenal mucosa provokes inhibitory reflex effect from the duodenum to the stomach. When food enters the duodenum, multiple nervous reflexes are initiated from the duodenal wall receptors. These reflexes are mediated through the enteric nervous system, or through long, and short nervous reflexes These signals depress the gastric juice secretion and at the same time decrease pyloric pump activity and usually increase pyloric tone to prevent the flow of chyme from exceeding the ability of the intestine to handle it. Osmolarity of the chyme: Iso-osmotic gastric contents empty faster than hyper or hypo-osmotic contents due to feedback inhibition produced by duodenal wall chemoreceptors (hyper more inhibitory than hypo). pH of chyme: Small intestinal chyme of < 3.5-4 stimulates the duodenal wall chemoreceptos and will activate reflexes that ultimately inhibits pyloric pump through. Presence of fats and protein digestion products in the duodenum. 2. Hormonal Feedback: The chyme coming from the stomach is normally hypertonic, acidic, and contains high concentrations of fat, protein, and carbohydrates. The intestinal inhibitory signals are: Fats entering the duodenum stimulate inhibitory hormone CCK which inhibits the pyloric pump. CCK is the most potent inhibitor of pyloric pump. Acidic chyme in the duodenum stimulates inhibitory hormone secretin which inhibits the pyloric pump. Fat, protein, and carbohydrates in the duodenum stimulate inhibitory hormones Gastric inhibitory peptide (GIP) that has a general but weak effect of decreasing gastrointestinal motility. CCK and secretin hormones are collectively called enterogastrones (figure 5.9). Inflammation of the duodenum may lead to increased acid output, hypocalcemia, and microcytic anemia. Increased basal and maximal acid outputs may result from excessive stimulation of the parietal cell (e.g., hypergastrinemia) or reduced inhibitory feedback (i.e., reduced effect of enterogastrone and the enterogastric reflex). The latter may occur when the proximal small intestine is inflamed. In summary: In humans, gastric acid secretion by the parietal cell occurs in response to (figure 5.8A): Excitatory neural (acetylcholine), hormonal (gastrin), and paracrine (histamine) stimuli. Inhibitory feedback regulation of acid output also involves neural (enterogastric reflex), hormonal (enterogastrone), and paracrine (somatostatin) influences. Obviously, these feedback signals allow chyme to enter the duodenum only as rapidly as it can be processed by small intestine. For example, whenever the pH of the chyme in the duodenum falls below 5.5 22 Mustansiriyah College of Medicine – 4.0, the duodenal inhibitory signals are immediately elicited, thus reducing or even blocking further release of acidic stomach content into the duodenum until the duodenal chyme can be neutralized by pancreatic and other secretion. Another example, hypo- or hypertonic fluids will elicit the duodenal inhibitory signals to prevent too rapid flow of non-isotonic fluids into the small intestine, thereby preventing marked changes in the body fluid osmolality. This feedback inhibition of the gastric glands plays an important role in protecting the stomach against excessively acid secretion, which would readily cause peptic ulceration. In addition to this protective effect, the feedback mechanism is also important in maintaining optimal pH for function of the peptic enzymes in the digestive process because whenever the pH rises above 2.5 - 3.5 gastrin begins to be secreted again and more acid is secreted. NB: Speed of delivery of nutrients from stomach to small intestine is as follow: CHO > protein > fat. Peptic ulcer (gastric and duodenal ulcers): Basically an ulcer forms when the capacity of the gastric mucosa to protect itself is overwhelmed by the existing level of gastric juice production. Gastric ulcers are frequently due to reduced defensive capacity, whereas, duodenal ulcers are frequently the result of increased exposure of the duodenal mucosa to gastric juice. Factors that may play a role in maintaining mucosal resistance are adequate blood flow to the gastric mucosa, the production of mucus, cellular 23 Mustansiriyah College of Medicine renewal, chemical factors (i.e. gastrin, prostaglandins, etc). Prostaglandins have been shown to dramatically increase the resistance of cells to toxic conditions. NSAIDs (nonsteroidal anti-inflammatory drugs) work by inhibiting prostaglandin synthesis. Chronic use of NSAID is a common cause of gastric ulceration. Recent studies have shown that the major acquired factor responsible for both gastric and duodenal ulcers is the bacterium Helicobacter pylori. H. pylori infections have high urease activity and metabolize urea into NH4+ which neutralizes the pH in the immediate area of the bacterial cells. This may be the major cause of H. pylori cytotoxicity because NH4+ directly damages epithelial cells and weakens the mucosal barrier. Treatment of ulcers with H2-receptor blockers or omeprazole in combination is very effective and has essentially replaced surgical and other forms of treatment. However, the ulcer frequently returns when treatment is discontinued unless the patient is also treated with oral antibiotics. In addition to its effect by inhibiting prostaglandin synthesis, Aspirin, is nonionized and lipid soluble in acid solutions, rapidly diffuses across this lipid layer, increasing mucosal permeability and damaging epithelial cells. Gastric irritation and gastric bleeding caused by gastric irritation occur in a significant number of persons who take aspirin on a regular basis. Alcohol, which also is lipid soluble, disrupts the mucosal barrier; when aspirin and alcohol are taken in combination, as they often are, there is increased risk of gastric irritation. Bile acids also attack the lipid components of the mucosal barrier and afford the potential for gastric irritation when there is reflux of duodenal contents into the stomach. The dumping syndrome: Is a condition that can develop after surgery to remove all or part of your stomach or after surgery to bypass your stomach to help you lose weight. It occurs because food moves from stomach into small bowel too quickly. Rapid emptying of the stomach can lead to low pH, elevated tonicity in the duodenum, incomplete digestion of food by the pancreas, liver, and duodenal mucosa and in incompletely emulsified fats in the duodenum. The increased rate of travel of food coupled with the reduced rate of digestion can ultimately result in incomplete digestion and absorption, particularly of fat and protein. The unabsorbed food retains osmotically water, which in conjunction with the increased rate of intestinal transport, can lead to diarrhea and steatorrhea (fat in the feces). The by-products of the bacterial metabolism of undigested food in the terminal ileum and colon can cause intestinal cramping, gas, and bloating. Weakness, dizziness, and sweating, are due in part to hypoglycemia, underly the presentation of dumping syndrome. Another cause of the symptoms is rapid entry of a hypertonic meal into the intestine, which promotes the movement of an abundance of water into the gut, producing significant hypovolemia and hypotension. Gastric digestion and absorption: Carbohydrate digestion in the stomach depends on the action of salivary amylase, which remains active until halted by the low pH in the stomach. Protein digestion occurs in the stomach by about 10% and mediated by HCl and later by gastric pepsin. Fat digestion is minimal in the stomach due to restriction of gastric lipase activity. Very little absorption of nutrients takes place in the stomach. The only substances absorbed to any appreciable extent are highly lipid-soluble substances (e.g., the non-ionized triglycerides of acetic, propionic, and butyric acids). Aspirin at gastric pH is non-ionized and fat soluble, after absorption, it ionizes intracellularly, damaging mucosal cells and ultimately producing bleeding. Ethanol is rapidly absorbed in proportion to its concentration. Water moves in both directions across the mucosa. Water-soluble substances including Na+, K+, glucose, and amino acids, are very poorly absorbed. Vomiting (emesis): It is a forceful expulsion of contents of the stomach and upper intestinal tract through the mouth. Vomiting is a reflex coordinated by the region in the brain stem medulla known as the vomiting 24 Mustansiriyah College of Medicine center. From here, motor impulses that cause the actual vomiting are transmitted from the vomiting center through parasympathetic vagal and sympathetic nerves to the lower tract and through motor spinal nerves to the diaphragm and abdominal muscles. Stimuli that elicit vomiting include (figure 5.10): Tactile (touch) stimulation of the back of the throat, which is one of the most potent stimuli. Irritation or distension of the stomach and duodenum. Stimulation of higher cortical centers by memory, feel, anticipation, sensory input (pain, smell, sight), elevated intracranial pressure (thus, vomiting after a head injury is considered a bad sign; it suggests swelling or bleeding within the cranial cavity). Rotation or acceleration of the head producing dizziness, such as in motion sickness. Body chemistry disturbances and chemical agents, including drugs or noxious substances that initiate vomiting (that is, emetics) either by acting in the upper parts of the gastrointestinal tract or by stimulating a specialized chemoreceptor trigger zone (CTZ, or postrema) next to the vomiting center in the brain. Activation of this zone triggers the vomiting reflex. For example, chemotherapeutic agents used in treating cancer often cause vomiting by acting on the chemoreceptor trigger zone. The importance of vomiting is to remove the ingested toxic substances before they can be absorbed. Moreover, the nausea that usually accompanies vomiting may have the importance in conditioning the individual to avoid the future ingestion of food containing the same toxic substances. Vomiting begins with: [1] Vomiting is usually preceded by increased salivation, sweating increased heart rate, pallor, and feeling of nausea, all are characteristic of a general discharge of the sympathetic nervous system in response to stress. Vomiting is often proceeded by retching (is the reverse movement (peristalsis) of the stomach and esophagus without vomiting). [2] Deep inspiration, closure of the glottis, and elevation of the soft palate. [3] The abdominal and thoracic muscles contract, raising the abdominal pressure which is transmitted to the contents of the stomach. In addition, the upper portion of small intestine contracts (retro peristalsis) and tends to force some of the intestinal contents back into the stomach. Thus some bile may be present in the vomitus. [4] The lower esophageal sphincter relaxes, and the high abdominal pressure forces the contents of the stomach into the esophagus. Excessive vomiting can lead to large losses of fluid and salts which normally would be absorbed in the small intestine. This can result in severe dehydration, upset the salt balance of the body and produce circulatory problems due to decrease in plasma volume. The loss of acid results in a low of body acidity. Figure 5.10: Main neurotransmitters involved in vomiting center regulation. CTZ = chemoreceptor trigger zone 25 Mustansiriyah College of Medicine Small intestine: The small intestine is the major site of digestion and absorption of carbohydrates, proteins, and fats in the GIT. The small intestine has three parts: the duodenum, the jejunum, and the ileum. Although the small intestine is approximately 5 m long, it has an absorptive area of over 250 m 2. Its large surface area is created by numerous folds of the intestinal mucosa (valvulae conniventes or plica circulares), by densely packed finger-like projections of the mucosa called villi, which line the entire mucosal surface, and by microvilli, which protrude from the surface of the intestinal epithelial cells (enterocytes). The microvilli give the intestinal mucosa its characteristics brush border appearance (figure 5.11). The life span of a typical epithelial cell varies from two to three days in the esophagus to six days in the large intestine. The divisions of epithelial stem cells continuously renew the lining of the entire digestive tract. These divisions normally keep pace with the rates of cell destruction and loss at epithelial surfaces. This high rate of cell division explains why radiation and anticancer drugs that inhibit mitosis have drastic effects on the digestive tract. Lost epithelial cells are no longer replaced. The cumulative damage to the epithelial lining quickly leads to problems in absorbing nutrients. In addition, the exposure of the lamina propria to digestive enzymes can cause internal bleeding and other serious problems. Though peristalsis in the small intestine is normally very weak, intense irritation of the intestinal mucosa, as occur in some sever case of infectious diarrhea, can cause both very powerful and rapid peristalsis called the peristaltic rush. This is initiated mainly by vagovagal nervous reflexes to the brain stem and back again to the gut. The powerful peristaltic contractions sweep the contents of the intestine into the colon and thereby relieving the small intestine of either irritative chyme or excessive distension. Function of the ileocecal valve: The principal function of the ileocecal valve is Figure 5.11: Histological cross section of small intestinal mucosa. 26 Mustansiriyah College of Medicine to prevent backflow of fecal content from the colon into the small intestine. The wall of the ileum for several centimeters immediately preceding the ileocecal valve has a thickened muscular coat called ileocecal sphincter which normally remains mildly constricted. Following meal, gastroileal reflex intensifies peristalsis, and the hormone, gastrin, which is liberated from the stomach mucosa in response to food in the stomach, has a relaxant effect on the ileocecal sphincter, both of them cause an increase emptying. On the other hand, when the cecum is distended, the degree of contraction of the ileocecal sphincter is intensified while ileal peristalsis is inhibited, which greatly delays emptying of additional chyme from the ileum. Also, any irritant in the cecum delays emptying. These reflexes from the cecum to the ileocecal sphincter and ileum are mediated both by way of the enteric nervous system and through vagovagal reflexes. Pancreatic secretions: Approximately 1200 to 1500 ml of clear pancreatic juice is produced daily (about pH 8). Pancreatic juice is secreted mainly in response to the presence of chyme in the upper portions of the small intestine and the characteristics of the pancreatic juice are determined to some extent by the types of food in the chyme. Pancreatic juice contains enzymes for digesting all three major types of food proteins, carbohydrates, and fats. It also contains large quantities of bicarbonate ions which play an important role in neutralizing the acid chyme emptied by the stomach into the duodenum. Pancreatic juice is isotonic and with the same Na+ and K+ concentrations and much lower Cl- concentration than plasma. The digestive enzymes of the pancreas are: The digestive enzymes for proteins (proteolytic enzymes) which are trypsin, chymotrypsin, carboxypolypeptidase, ribonuclease, and deoxyribonuclease. By far the most abundant of these is trypsin (figure 5.13). The digestive enzyme for carbohydrates is pancreatic amylase which hydrolyzes starches, glycogen and most other carbohydrates except cellulose to form disaccharides and a few trisaccharides. The digestive enzyme for fat which is pancreatic lipase that is capable of hydrolyzing neutral fat into fatty acids and monoglycerides, cholesterol esterase which cause hydrolysis of cholesterol esters, and phospholipase which splits fatty acids from phospholipids. Pancreas protects itself from autodigestion by proteolytic enzymes by the following ways: [1] When synthesized in the pancreatic cells, the proteolytic enzymes are in the inactive forms (inactive zymogens), trypsinogen, chymotrypsinogen, and procarboxypolypeptidase, which are all enzymatically inactive (figure 5.13). Figure 5.13: Mechanisms of activation of pancreatic proteolytic enzymes. 27 Mustansiriyah College of Medicine [2] These become activated only by trypsin. [3] Trypsin is activated only after it is secreted into the intestinal tract. Trypsinogen is activated by an enzyme called enteropeptidase (enterokinase) which is secreted by the intestinal mucosa when chyme comes in contact with the mucosa. Also, trypsinogen can be autocatalytically activated by trypsin that is already been formed. Chymotrypsinogen is activated by trypsin to form chymotrypsin, and procarboxypolypeptidase is activated in a similar manner. It is important that the proteolytic enzymes of the pancreatic juice not become activated until they have been secreted into the intestine which otherwise would digest the pancreas itself. [4] The same cells that secrete the proteolytic enzymes into the acini of the pancreas secrete simultaneously another substance called trypsin inhibitor. This substance is stored in the cytoplasm of the glandular cells surrounding the enzyme granules and it prevents activation of trypsin both inside the secretory cells and in the acini and ducts of the pancreas. However, when the pancreas becomes severely damaged or when a duct becomes blocked, large quantities of pancreatic secretion become pooled in the damaged areas of the pancreas. Under these conditions, the effect of trypsin inhibitor is sometimes overwhelmed, in which case the pancreatic secretions rapidly become activated and digest the pancreas giving rise to the condition called acute pancreatitis. The enzymes of the pancreatic juice are secreted entirely by the acini of the pancreatic glands. On the other hand, two other important components of the pancreatic juice, bicarbonate ions and water, are secreted in large amounts mainly by the epithelial cells of the ductules leading from the acini (figure 5.14). The alkaline ions or the pancreatic juice serves to neutralize acid emptied into the duodenum from the stomach. The secretion of bicarbonate ions by the pancreas is an active process. The mechanism of bicarbonate secretion is similar to the process of HCI secretion by the stomach, except that the events on the two sides of the cells are reversed. Because the amount of bicarbonate secreted by the pancreas is reflexly regulated by the amount of acid entering the duodenum from the stomach, the amount of bicarbonate secreted by the pancreas is normally about equal to the amount of acid secreted by the stomach. This also means that the bicarbonate released into the blood by the stomach is normally equal to the amount of acid released into the blood by the pancreas. When the alkaline blood leaving the stomach mixes with the acidified blood from the pancreas, the overall result is no change in the acidity of the blood returning to the heart. However, a loss of large quantities of bicarbonate ions from the intestinal tract during periods of prolonged diarrhea leads to a net accumulation of acid in the blood, just as, loss of acid from the stomach by vomiting leads to a net alkalinization of the blood. Figure 5.14: Mechanism of pancreatic ductule cell secretion of electrolytes. 28 Mustansiriyah College of Medicine Regulation of pancreatic secretion: Pancreatic secretion is regulated by both nervous and hormonal mechanisms. However, hormonal regulation is by far the more important. Like gastric secretion, pancreatic secretion is divided into the following three phases: [1] Cephalic phase: The thought, sight, smell, or taste of food produces the cephalic phase of pancreatic secretion via vagal stimulation. It is scanty and rich in enzymes. [2] Gastric phase: Pancreatic secretion is slightly enhanced during the gastric phase by: [A] Distention of the antrum and the body of the stomach, which initiates a vagovagal reflex resulting in a low volume of pancreatic secretion containing both bicarbonate ions and enzymes. [B] Food breakdown digestion products (primarily amino acids and peptides) can stimulate pancreatic secretions because of their ability to cause the G cells of the antrum to release gastrin. Gastrin produces a low-volume, high-enzyme pancreatic secretion. [3] Intestinal phase: The major stimulants for pancreatic secretion are the hormone CCK and secretin. They are released from endocrine cells in the duodenum and jejunum during the intestinal phase of pancreatic secretion in response to the entrance of chyme into the small intestine. CCK, in addition to its effect on the gallbladder, is a potent stimulant of pancreatic enzyme secretion. Protein and fat are the major stimuli for CCK secretion. CCK induces secretion of enzyme-rich pancreatic juice (figure 5.15). Secretin stimulates the HCO3- secretion. Low pH (< 4.5), caused by the presence of gastric acid (HCl) in the intestine, is a potent stimulus for the release of secretin. Secretin causes secretion of HCO3−-rich pancreatic juice (figure 5.16). Secretin is very important for two reasons: [A] Neutralization of the acidic chyme: Secretin is released from the mucosa of small intestine when the pH of the duodenal contents falls below 4.5 and its release increases very greatly as the pH falls to 3.0. This immediately causes large quantities of pancreatic juice containing abundant amounts of sodium bicarbonate to be secreted. In this way, the acid contents emptied into the duodenum from the stomach become neutralized, and peptic activity of the gastric juice is immediately blocked. Since the mucosa of the small intestine cannot withstand the digestive action of acid gastric juice. This is a highly important and even Figure 5.15: The control of pancreatic bicarbonate and digestive enzyme secretions. 29 Mustansiriyah College of Medicine essential protective mechanism against the development of duodenal ulcers. [B] Provides a proper alkaline medium for the action of pancreatic enzymes. Because CCK and secretin are potentiators of each other’s action, small concentrations of CCK and secretin together can produce significant amounts of pancreatic bicarbonate ions and enzymes secretions, while either one alone would have little or no effect. Pancreatitis and Pancreatic Cancer: Pancreatitis is an inflammation of the pancreas that occurs quite commonly. Pancreatitis involves the release of pancreatic enzymes within the pancreas itself, which digest pancreatic tissue. It can result from alcoholism, the use of certain drugs, pancreatic duct blockage, cystic fibrosis, viral infection, or pancreatic cancer. Symptoms can range from mild abdominal pain to systemic shock and coma. Cancer of the pancreas can obstruct the pancreatic and common hepatic ducts, resulting in jaundice. Pancreatic cancer may not be detected until the tumor has become fairly large, and it can become so large as to block off the pyloric region of the stomach. Bile secretion: Bile is a yellow-green, alkaline solution containing bile salts, bile pigments, cholesterol, triglycerides, phospholipids (lecithin and others), and a variety of electrolytes. Bile is required for the digestion and absorption of fats and for the excretion of water-insoluble substances such as cholesterol (the only route), bilirubin, and some drugs. Bile is formed by liver epithelium (hepatocytes) and by epithelial cells lining the bile ducts (ductal cells). Between 250 - 1200 ml of bile is formed daily which is normally stored in 30 - 60 ml volume of gallbladder after being concentrated as much as fivefold up to 18 fold by continual absorption of water, sodium, chloride, and most other small electrolytes through the mucosa of gallbladder by an active transport of sodium through the gallbladder epithelium. The concentrated bile constituents are: Bile salts (11%). Bile pigments (1%). Lipids (cholesterol 0.5%, lecithin 3%). Water and electrolytes (84%). The bile electrolyte concentrations are similar to those of plasma except more HCO3- and less Cl- (pH 7.59.0). [1] The bile salts: The normal rate of bile salt synthesis is 0.2 – 0.4 g/day. The precursor of bile salts is cholesterol which is converted by liver to cholic acid or chenodeoxycholic acid. These acids then combined principally with amino acid glycine and to a lesser extent with taurine to form glyco- and tauro- conjugated bile acids. These bile acids form sodium and potassium salts in the alkaline hepatic bile. Bile salts have the propensity to form micelles, which are small spherical globules composed of 20 - 40 molecules of bile salt. Each bile salt molecule is composed of a sterol nucleus which is highly fat soluble (in which lipids are dissolved), and a polar group that is highly water soluble. This aggregation causes the polar groups to project outward to cover the surface of micelle (figure 5.16). Since these polar groups are electrically charged, they allow the entire micelle globule to become dissolved in the water of the digestive fluids. These bile salts have two important actions in the intestinal tract: [A] Break the fat globules into minute sizes: Bile salts have a detergent action (or emulsification action) on the fat particles in the food which decreases the surface tension of the particles and allows the agitation in the intestinal tract to break the fat globules into minute sizes (figure 5.17). 30 Mustansiriyah College of Medicine [B] Making lipid droplets highly water soluble: Bile salts help in the absorption of lipids and other fat-soluble substances from the intestinal tract by forming minute complexes with micelles and become highly water soluble. The lipids are ferried in this form to the mucosa where they are then absorbed. Without the presence of bile salts Figure 5.16: Micelle globule. in the intestinal tract, up to 40% of the lipids are lost into the stool. Also, when fats are not absorbed adequately, the fat-soluble vitamins A, D, K, and E are not absorbed satisfactorily. Approximately 94% of the bile salts are reabsorbed by an active transport process (Na-bile salt cotransport) through the intestinal mucosa in the distal ileum. They enter the portal blood and pass to the liver. On reaching the liver the bile salts are reabsorbed almost totally on the first passage through the venous sinusoids into the hepatic cells and then resecreted into the bile. The small quantities of bile salts lost into the feces are replaced by new amounts formed continually by the liver cells. The recirculation of the bile salts is called the enterohepatic circulation. Removal of the terminal ileum can lead to diarrhea and steathorrea. Bile salts are necessary for adequate digestion and absorption of fat. In the absence of the terminal ileum there will be an increase in the amounts of bile acids and fatty acids delivered to the colon. Fats and bile salts in the colon increase the water content of the feces by promoting the influx (secretion) of water into the lumen of the colon. [2] The bile pigments (mainly bilirubin and biliverdin) which are responsible for the golden yellow color of bile and they are formed as a major end product of Hb degradation. Bilirubin is fat soluble, and therefore, highly soluble in all cell membranes and is also very toxic. Therefore, its excretion in the bile is one of the very important functions of the liver. The bilirubin released into the plasma is immediately combined very strongly with the plasma albumin and is transported in this combination throughout the blood and interstitial fluids. This type of bilirubin is called “unconjugated bilirubin” or “indirect bilirubin”. This type of bilirubin cannot be excreted by the kidneys because it is insoluble in water and is combined with a protein albumin. Because it is fat soluble, however, it can enter the brain or the subcutaneous adipose tissues, especially if high serum level exist. In premature infant, lack of subcutaneous fat exposes them to greater danger of unconjugated bilirubin entering the nervous system where it deposited in the basal ganglia leading to a condition called kernicterus. It should be realized, however, that only unconjugated bilirubin which is not bound to serum albumin is free to enter the central nervous system and other tissues, and the risks of hyperbilirubinaemia are therefore influenced by the bilirubin binding capacity of the serum albumin. Therefore, kernicterus consequent upon hyperbilirubinaemia will only develop when the serum Figure 5.17: Arrangement of micelles over the surface of lipid droplet. 31 Mustansiriyah College of Medicine albumin is unable to bind further unconjugated bilirubin. Within hours, the unconjugated bilirubin is absorbed through the hepatic cell membrane. In this process being released from the plasma albumin but almost instantly is combined with one of two proteins (called Y and Z proteins) inside the hepatic cells that trap the bilirubin inside the cells. Conjugated bilirubin is water-soluble and so readily excreted in the urine or bile, and as it is not soluble in fat, it is not toxic to the nervous system. A small portion of the conjugated bilirubin formed by the hepatic cells returns to the plasma. This causes a small portion of the bilirubin in the extracelluler fluids always to be of the conjugated type. Once in the intestine, about one half of the bilirubin is converted by bacterial action into the substance urobilinogen which is responsible for the brown color of stool. Some of the urobilinogen is reabsorbed through the intestinal mucosa into the blood. Most of this is re-excreted by the liver back into the gut (enterohepatic circulation), but about 5% is excreted by the kidneys into the urine. After exposure to air in the urine, the urobilinogen becomes oxidized to urobilin, or in the faces it becomes altered and oxidized to form stercobilin. Jaundice: It means a yellowish tint to the body tissues, including yellowness of the skin and also the mucous membrane and the deep tissues. The usual cause of jaundice is large quantities of bilirubin in the extracellular fluids either conjugated or unconjugated bilirubin. Jaundice may result from: [1] Excess production of bilirubin caused by excessive RBC destruction as in haemolytic anaemia. [2] Inability of hepatocytes to conjugate the plasma free bilirubin as in parenchymal liver diseases. [3] Obstruction of the bile ducts or liver cells preventing the secretion of conjugated bilirubin. [3] Cholesterol, phospholipids (primarily lecithin), and fatty acids. Cholesterol is almost insoluble in water, but the bile salts and lecithin in bile combine physically with cholesterol to form ultramicroscopic micelles that are soluble in water. Regulation of gallbladder emptying (contraction): Two basic conditions are necessary for the gallbladder to empty, the sphincter of Oddi must relax to allow bile to flow from the common bile duct into the duodenum, and the gallbladder itself must contract to provide the force required to move the bile along the common bile duct. After a meal both these effects take place in the following manner: [1] the fat and also partially digested protein in the food entering the small intestine cause the release of the hormone CCK from the mucosa of the upper region of small intestine which in turn is absorbed into the blood and on passing to the gallbladder, causes contraction of the gallbladder muscle. This provides the pressure that forces bile toward the duodenum. CCK is the major stimulus for gallbladder contraction and sphincter of Oddi relaxation. [2] Vagal stimulation associated with the cephalic phase of gastric secretion or via a vagovagal reflex during the gastric phase of digestion, causing contraction of the gallbladder. Control of biliary secretion from the liver cells: The quantity of bile secreted by the liver is highly dependent on: [1] The availability of bile salts: The greater the quantity of bile salts in the enterohepatic circulation, the greater the rate of bile secretion. [2] Secretin also increases bile secretion. However, this increase in secretion represents mainly secretion of a bicarbonate rich watery solution by the epithelial cells of the liver ductules and ducts and not increased secretion of bile acids by the liver parenchymal cells. The bicarbonate in turn passes into the 32 Mustansiriyah College of Medicine small intestine and joins the bicarbonate from the pancreas in neutralizing the acid from the stomach. Because bile salts are required for proper digestion and absorption of fats, any condition that disrupts the enterohepatic circulation as in ileal resection or small intestinal diseases leads to a decreased bile acid pool and malabsorption of fat and fat-soluble vitamins. The clinical manifestations of such conditions are steatorrhoea and nutrition deficiency. An increase in fecal losses of bile salts results in watery diarrhea, since bile salts inhibit water and Na+ absorption in the colon. Bile, and not the gallbladder, is essential to digestion. After removal of the gallbladder, bile empties slowly but continuously into the intestine, allowing digestion of fats sufficient to maintain good health and nutrition. Only high-fat meals need to be avoided. Gallstones: Cholesterol, secreted by the liver, may precipitate in the gallbladder to produce gallstones. Cholesterol is not soluble in water and is ordinarily kept in solution by bile salts. Gallstones can form when there is excess cholesterol in the bile due to a high-cholesterol diet or when cholesterol is overly concentrated in the gallbladder. Occasionally, a gallstone passes out of the gallbladder and enters the cystic duct, blocking the release of bile. Such a condition interferes with normal digestion, and the gallstone often must be removed surgically. If the gallstone moves far enough down the duct, it can also block the pancreatic duct, resulting in pancreatitis. Regulatory hormones in the GIT: Gastrin: It is released from G cells of the antral mucosa. The functions of gastrin are: 1. ↑ Gastric H+ secretion, 2. Stimulates growth of gastric mucosa. 3. Increases antral muscle mobility and promotes stomach contractions. 4. Strengthens antral contractions against the pylorus, and relaxes the pyloric sphincter, which increases the rate of gastric emptying. 5. Plays a role in the relaxation of the ileocecal valve. 6. Induces pancreatic secretions and gallbladder emptying. 7. May impact lower esophageal sphincter (LES) tone, causing it to contract, although pentagastrin, rather than endogenous gastrin, may be the cause. 8. Gastrin contributes to the gastrocolic reflex. Its release is stimulated by: A. The antral distension. B. The presence of protein digesting products in the antrum (the most potent are phenylalanine and tryptophan), coffee, and alcohol. C. Vagal stimulation via GRP (bombesin). Its release is inhibited by: A. High H+ concentration in the stomach (pH < 3). B. Somatostatin. CCK: It is released from I cells of the mucosa of Figure 5.17A: Sites of GI hormone release. 33 Mustansiriyah College of Medicine duodenum and jejunum. The functions of gastrin are: 1. Stimulates contraction of gallbladder and relaxation of sphincter of Oddi. 2. ↑ Pancreatic enzyme and HCO3– secretion. 3. ↑ Growth of exocrine pancreas/gallbladder. 4. Inhibits gastric emptying 5. It stimulates gallbladder contraction, and intestinal motility, while inhibits gastric emptying. Its release is stimulated by: A. The presence of fat digesting products in the small intestine (triglycerides do not stimulate the release of CCK because they cannot cross intestinal cell membranes). B. The presence of protein digesting products in the small intestine. Secretin: It is released from S cells of the mucosa of the duodenum and jejunum. 1. Inhibits gastric emptying. 2. ↑ Pancreatic HCO3–, secretion, 3. ↑ Biliary HCO3– secretion, 4. ↓ Gastric H+ secretion Its release is stimulated by the H+ in the lumen of the duodenum. Motilin: It stimulates migrating motor complex. Glucagon-Like Peptide I: Glucagon-like peptide 1 belongs to a family of hormones called the incretins, so-called because they enhance the secretion of insulin. Cells found in the lining of the small intestine (called L-cells) are the major source of glucagon-like peptide 1. Food is the main stimulus of glucagon-like peptide 1 release, with increased hormone levels detectable after 10 minutes of starting to eat and remaining raised in the blood circulation for several hours after that. The hormone somatostatin holds back the production of glucagon-like peptide 1. 1. Glucagon-like peptide 1 increases the feeling of fullness during and between meals by acting on appetite centers in the brain and by slowing the emptying of the stomach. 2. It slows gastric emptying. 3. Enhances the secretion of insulin Glucose-dependent insulinotropic peptide (GIP) also called Gastric inhibitory polypeptide (GIP): It is released from duodenum and jejunum. The stimulus for secretion is the fatty acids and amino acids and the presence of oral glucose load. Thus, oral glucose is more effective than intravenous glucose in causing insulin release and, therefore, glucose utilization. GIP is the only GI hormone that is released in response to the three main food groups, i.e. fat, protein, and carbohydrate. 1. ↑ Insulin secretion 2. ↓ Gastric H+ secretion 3. Inhibits the gastric and GI motility Somatostatin: is secreted by cells throughout the GI tract in response to H+ in the lumen. Its secretion is inhibited by vagal stimulation. Somatostatin is also released by beta-adrenergic agents. This observation might explain the inhibition of gastric secretion by anger or excitement. The release of somatostatin is inhibited by acetylcholine. That makes sense because vagal activity begins even before the food has been ingested. At that stage, gastric secretion needs to be initiated without a braking influence. 34 Mustansiriyah College of Medicine 1. Inhibits the release of all GI hormones. 2. Inhibits gastric H+ secretion. Histamine: is secreted by mast cells of the gastric mucosa. It increases gastric H+ secretion directly and by potentiating the effects of gastrin and vagal stimulation. Neurocrines: Are synthesized in neurons of the GI tract, moved by axonal transport down the axon, and released by action potentials in the nerves. Neurocrines then diffuse across the synaptic cleft to a target cell. The GI neurocrines are vasoactive intestinal peptide (VIP), GRP (bombesin), and enkephalins. A. VIP: contains 28 amino acids and is homologous to secretin. It is released from neurons in the mucosa and smooth muscle of the GI tract. 1. Produces relaxation of GI smooth muscle, including the lower esophageal sphincter. 2. Stimulates pancreatic HCO3– secretion and inhibits gastric H+ secretion. In these actions, it resembles secretin. B. GRP (bombesin): is released from vagus nerves that innervate the G cells. It stimulates gastrin release from G cells. C. Enkephalins (met-enkephalin and leu-enkephalin) are secreted from nerves in the mucosa and smooth muscle of the GI tract. 1. Stimulate contraction of GI smooth muscle, particularly the lower esophageal, pyloric, and ileocecal sphincters. 2. Inhibit intestinal secretion of fluid and electrolytes. This action forms the basis for the usefulness of opiates in the treatment of diarrhea. 35 Mustansiriyah College of Medicine Digestion & Absorption: Digestion of carbohydrates: Only three major sources of carbohydrates exist in the normal human diet. These are sucrose (disaccharide) also known as cane sugar or table sugar, lactose (disaccharide) in milk, and starches (polysaccharides) present in almost all non animal foods especially in the grains. Other carbohydrates ingested to a slight extent are glycogen, alcohol, lactic acid, pyruvic acid, pectins, dextrins, and others. Digestion of carbohydrates in mouth: Because the food remains only for short time in the mouth, only 3 -5% of all starches that have been eaten will have become hydrolyzed (digested). Enzyme α-amylase (ptyalin) hydrolyzes starches into the disaccharide maltose and other small polymers of glucose. Starches (polysaccharide) Maltose (disaccharide) + Glucose Digestion of carbohydrates in the stomach: In the stomach the action of salivary ptyalin can continue for as long as an hour after the food has entered the stomach until the contents of the fundus are mixed with the stomach secretions. Then the activity of the salivary amylase is blocked by the acid of the gastric secretions. By this time about 30 - 40% of the starches will have been hydrolyzed mainly to maltose. Pancreatic digestion of carbohydrates in the small intestine: Pancreatic secretion contains large quantities of amylase that continue splitting starches into maltose and other small polymers of glucose. By now, the starches are almost totally converted into maltose and other very small glucose polymers before they have passed beyond the jejunum. The small intestine surface digestion and hydrolysis of disaccharides and small glucose polymers: The brush border of epithelial cells lining the small intestine contain enzymes lactase, sucrase, and maltase, which are capable of splitting the disaccharides lactose, sucrose, and maltose respectively, into their constituent monosaccharides when the disaccharides come in contact with this border. The digested products, the monosaccharides, are then immediately absorbed into the portal blood. Lactose (disaccharide) Galactose + Glucose. Sucrose (disaccharide) Fructose + Glucose. Maltose (disaccharide) Glucose + Glucose. Absorption of carbohydrates: All carbohydrates are absorbed in the form of monosaccharides, only a small fraction of a percent being absorbed as disaccharides and almost none as larger carbohydrate compounds. [1] Glucose and galactose are transported from the small intestinal lumen into the cells by Na+-glucose or Na+-galactose secondary active cotransport in the luminal brush border of the membrane. When the intracellular glucose and galactose concentration increased to a higher level than normal, they will diffuse by facilitated diffusion through the basolateral membrane of the epithelial cell into the extracellular fluid and then to the blood. The Na+-K+ pump in the basolateral membrane keeps the intracellular Na + concentration low, thus maintaining the Na+ gradient across the luminal membrane. [2] Fructose is transported to small intestinal enterocyte by facilitated diffusion. Within the enterocytes, it is mainly converted into glucose inside the epithelial cell before entering the portal blood. Deficiency of one or more of these disaccharidase leads to diarrhoea, bloating, and flatulence after ingestion of sugar. The diarrhea is due to the increased number of osmotically active oligosaccharides molecules that remain in the intestinal lumen causing the volume of the intestinal contents to increase. The bloating and flatulence are due to the production of gas (CO 2 and H2) from disaccharide residues which are metabolized by intestinal flora in the lower small intestine and colon. Lactose intolerance is the most common cause of carbohydrate malabsorption. It results 36 Mustansiriyah College of Medicine from the inability of the small intestinal cells to produce lactase. In infants, the diarrhea-produced dehydration can be life threatening. This disorder is primarily hereditary, affecting 5-15% of Europians and 80-90% of Asians and Africans. Digestion of fat: By far the most common fats of the diet are the neutral fats (triglycerides). In the usual diet there are also small quantities of phospholipids, cholesterol, and cholesterol ester. Although the serous glands of the tongue secrete lingual lipase, very little, if any, lipid digestion occurs in the mouth or stomach. Essentially all fat digestion occurs in the small intestine as follows: [1] Emulsification of fat by bile acids: The first step in fat digestion is to break the fat globules into small sizes so that the water soluble digestive enzymes can act on the globule surfaces. This process is called emulsification of the fat and it is achieved under the influence of bile salts and aided by lecithin in the bile. Both decrease the interfacial tension of the fat globules which result in breaking up these globules into many minute particles when they are agitated. This results in an increase in surface area of fat globules as much as 1000 fold. [2] Digestion of fats by pancreatic lipases: It is by far the most important enzymes (lipases, cholesterol estrase, and phospholipase A2) for the digestion of fat. However, the epithelial cells of the small intestine also contain a minute quantity of lipase known as enteric lipase. Both of these act alike to cause hydrolysis of fat. Most of the triglycerides of the diet are spilt into free fatty acids and monoglycerides. However, small portions are not digested at all or remain in the diglyceride state. [3] Transportation of fats to the brush border of the epithelial cells: During triglyceride digestion, as rapidly as the monoglycerides and free fatty acids are formed they become dissolved in the fatty portion of the micelles. The bile salt micelles act as a transport medium to carry the monoglycerides and the free fatty acids to the brush border of the epithelial cells, which would otherwise be relatively insoluble to the brush border of the epithelial cells. There, the monoglycerides and free fatty acids are absorbed. On delivery of these substances to the brush border, the bile salts are again released back into the chyme to be used again and again for this ferrying process. However, few of the monoglycerides are further digested into glycerol and fatty acids by an epithelial cell, lipase. Both the cholesterol esters and phospholipids are hydrolyzed by lipases (cholesterol esterase and phospholipase A2) in the pancreatic secretion that free the fatty acids. The bile salt micelles play identical role in ferrying free cholesterol and phospholipids as that with monoglycerides and free fatty acids. Indeed, this role of the bile salt micelles is absolutely essential to absorption of cholesterol and no cholesterol absorption can take place without bile salts. Some of the triglycerides can be digested and absorbed even in the absence of bile salts. This is because short- and medium chain triglycerides are more water soluble and they diffuse through water layer adjacent to the brush border of the enterocytes and are absorbed intact. Once inside the cell, they are hydrolyzed by esterase. MCTs are not utilized for re-synthesis of triglycerides and therefore are not packaged into chylomicrons. Absorption of fats (figure 5.18): As micelles come in contact with the brush border microvilli, both the monoglycerides and the fatty acids immediately diffuse through the epithelial membrane, because they are equally as soluble in this membranes as in the micelles. As these fat products leave the micelles and are absorbed across the epithelial cell membranes, the micelles can pick up more monoglycerides and free fatty acids, which have been produced from digestion of other triglyceride molecules in the fat emulsion. Bile 37 Mustansiriyah College of Medicine salts continuously repeat their fat-solubilizing function down the length of the small intestine until all fat is absorbed. Then the bile salts themselves are reabsorbed in the terminal ileum by a Na+dependent active transport process. Although Figure 5.18: Absorption of fats. triglycerides, like monoglycerides and fatty acids, are highly soluble in the lipid membrane of the epithelial cell, however, only small quantities of these are normally absorbed because the bile acid micelles will not dissolve either triglycerides or diglycerides and therefore will not ferry them to the epithelial membrane. After entering the epithelial cell, the fatty acids and monoglycerides, are taken up by the smooth endoplasmic reticulum, and here they are recombined to form new triglycerides. Once formed, the triglycerides aggregate within the endoplasmic reticulum into globules along with absorbed and the newly synthesized cholesterol and phospholipids. The phospholipids arrange themselves in these globules with the fatty portions of phospholipids toward the center and the polar portions located on the surface. This provides an electrically charged surface that makes these globules miscible (water soluble) with the fluids of the cell. In addition, small amounts of β-lipoprotein, also synthesized by the endoplasmic reticulum, coat part of the surface of each globule. In this form the globule diffuse to the side of the epithelial cell and is excreted by the process of cellular exocytosis into the space between the cells, from these it passes into the lymph in the central lacteal of the villus. These globules are then called chylomicrons. Chylomicrons propelled along with the lymph upward through the thoracic duct to be emptied into the great veins of the neck. Small quantities of short chain fatty acid are absorbed directly into the portal blood rather than being converted into triglyceride and absorbed into the lymphatics. This is because these short chain fatty acids are more water soluble and are not reconverted into triglycerides by the endoplasmic reticulum. This allows direct diffusion of these fatty acids from the epithelial cells into the capillary blood of the villus. Lipid malabsorption is much more common than carbohydrate or protein malabsorption. It is usually results from one of the following two conditions: [1] the pancreas does not secrete sufficient quantities of lipase (as in pancreatitis). [2] The liver does not secrete sufficient quantities of bile (as in liver parenchymal diseases). Digestion of proteins: The dietary proteins are derived almost entirely from meats and vegetables. These proteins in turn are formed of long chain of amino acids bound together by peptide linkages. Digestion of proteins in the stomach: For pepsin to cause any digestive action on protein, the stomach juices must be acidic (pH 2 – 3). Pepsin digestion represents 10—30% of the total protein digestion. This splitting of proteins is a process of hydrolysis occurring at the peptide linkages between the amino acids. Digestion of proteins by pancreatic secretions: Most protein digestion occurs principally in the small intestine under the influence of the proteolytic enzymes of the pancreatic secretion. When the partial 38 Mustansiriyah College of Medicine breakdown proteins leave the stomach, they are attacked by the pancreatic enzymes. Trypsin and chymotrypsin can split protein molecules into small polypeptides, carboxypolypeptidase then leaves individual amino acids from the carboxyl ends of the polypeptides. Digestion of peptides by epithelial peptidases of small intestine: The brush border of the small intestine contains several different enzymes for hydrolyzing the final peptides linkages of the remaining dipeptides and other small polypeptides as they come in contact with the epithelium of the villi. The enzymes responsible are aminopolypeptidase and several dipeptidase. When the food has been properly masticated and is not eaten in too large a quantity at any one time, about 98% of all the proteins finally become either amino acids or dipeptides that can be absorbed into the blood. Absorption of amino acids: [1] Most proteins are absorbed in the form of amino acids, dipeptides and even tripeptides by Na+amino acid cotransport system in the brush border of the epithelial cell. Once inside the enterocytes, intracellular peptidases digest some of the polypeptides to amino acid. The amino acids are then transported from cell to blood by facilitated diffusion. [2] Larger molecules, especially in newborn infant, are engulfed by the plasma membrane of the epithelial cell (endocytosis), move through the cytoplasm, and are released on the opposite side of the cell by the reverse process (exocytosis). The capacity to absorb intact proteins is greater in a newborn infant and antibodies secreted in the mother’s milk may be absorbed by the infant in this manner, providing a short term passive immunity until the child begins to produce its own’ antibodies. Essentially no free amino acids can be found in the intestine during digestion because they are absorbed as rapidly as they are formed. Inadequate absorption of proteins due to lack of trypsin is a common consequence of pancreatic diseases. Absorption of Na+, Cl- and K+ ions: Electrolytes and H2O may cross intestinal epithelial cells by either cellular or paracellular (between cells) routes. Tight junctions attach the epithelial cells to one another at the luminal membrane. The permeability of the tight junctions varies with the type of epithelium. A “tight” (impermeable) epithelium is the colon. “Leaky” (permeable) epithelia are the small intestine and gallbladder. The major ions that are Figure 5.19: Absorption of NaCl, K, HCO3-, absorbed in the small intestine and proximal colon are Na+, and water from small intestine and K+, and Cl-, (figure 5.19): This this achieved primarily by the + + proximal colon. Na -K -ATPase pumps located at the basolateral membrane of the enterocytes. This pump creates low intracellular concentration of Na +. This will leads to: [1] Na+ enters the enterocyte in three ways: By passive diffusion down its electrochemical gradient. 39 Mustansiriyah College of Medicine By Na+- Cl- co-transport system (major mechanism). By a Na+-glucose, Na+-amino acid, Na+-(di- or tri-) peptide or Na+- water-soluble vitamins (except B12 and folic acid) co-transport system (Na+ enhances glucose absorption in the Glucose-saline solutions). By Na+-H+ counter-transport [2] Cl- enters the enterocyte through: Passive diffusion down its electrochemical gradient established by the active transport of Na +. Na+-Cl- co-transport (major mechanism). Cl--HCO3- counter-transport. Once inside the enterocyte, Na+ is transported across the basolateral membrane by a Na+-K+- ATPase active transport system and Cl- follows. In the small intestine, Na+–glucose cotransport, Na+–amino acid cotransport, and Na+–H+ exchange mechanisms are most important. In the colon, passive diffusion via Na+ channels is most important. The Na+ channels and Na+-K+ATPase of the colon are similar to those in the renal distal tubule and are stimulated by aldosterone (figure 5.19A). When a person becomes dehydrated, large amounts of aldosterone almost always are secreted by the cortices of the adrenal glands. Within 1 to 3 hours this aldosterone causes increased activation of the enzyme and transport mechanisms for all aspects of sodium absorption by the intestinal epithelium. And the increased sodium absorption in turn causes secondary increases in absorption of chloride ions, water, and some other substances. This effect of aldosterone is especially important in the colon because it allows virtually no loss of sodium chloride in the feces and also little water loss. Thus, the function of aldosterone in the intestinal tract is the same as that achieved by aldosterone in the renal tubules, which also serves to conserve sodium chloride and water in the Figure 5.19A: Colonic body when a person becomes dehydrated. However, in doing so, it reabsorption of Na and causes significant amounts of K+ to be lost from the body. secretion of K ions. [3] Absorption and secretion of potassium ions: The daily dietary K+ intake of approximately 100 mmol is excreted predominantly by the distal tubules of the kidney. About 10% of the ingested K+ is excreted via the intestine. K is reabsorbed across the intestinal mucosa through Simple diffusion. In the small intestine, about 85% of ingested K moves to interstitial space by simple diffusion mainly via paracellular route (i.e. through leaky tight junctions of the mucosa of the small intestine). As water is absorbed from the lumen, rising potassium levels in chyme create a concentration gradient for its absorption. Anything that interferes with water absorption (resulting in diarrhea) not only reduces potassium absorption but also “pulls” K+ from the interstitial space into the intestinal lumen. In large intestine, K+ is actively secreted in the colon by a mechanism similar to that for K+ secretion in the renal distal tubule. As in the renal distal tubule, K+ secretion in the colon is stimulated by aldosterone. Because of the “tight” nature of the tight junctions that connect cells in the colon, a relatively large potential difference exists between the luminal (negative) and basal (positive) surfaces of the absorptive cells. This electrical difference favors the net secretion of K+ into the lumen through paracellular route. 40 Mustansiriyah College of Medicine ■ In diarrhea, K+ secretion by the colon is increased because of a flow rate–dependent mechanism similar to that in the renal distal tubule. In diarrhea, there is less opportunity for [K+] in colon to rise up to significant level to act as chemical gradient force against the active and passive secretion of K. Therefore, excessive loss of K+ in diarrheal fluid causes hypokalemia. Absorption of water: Water is transported across the small intestinal membrane entirely by the process of osmotic diffusion which obeys the usual laws of osmosis, or through paracellular route between the enterocytes. Most of the water in the gastrointestinal tract is absorbed in the jejunum, while the duodenum serving primarily as the site of osmotic equilibration of chyme. Most of the remaining water and electrolytes are absorbed in the proximal half of the colon, giving this portion the name absorbing colon whereas the distal colon functions principally for storage and is therefore called the storage colon. In contrast to the small intestine, the colon has a limited capacity to absorb water (approximately 3 to 6 L per day). This is because the highly impermeable nature of the tight junctions connecting the colonic epithelial cells. Absorption and secretion of bicarbonate ions in the intestine: Absorption of bicarbonate from the small intestine occurs primarily in the jejunum. Often large quantities of bicarbonate ions reabsorbed from the upper small intestine in form of carbon dioxide which is readily absorbed into the blood and subsequently expired through the lungs. This is important because large amounts of bicarbonate ions have been secreted into the duodenum in both pancreatic secretion and bile. However, the mucosa of the large intestine actively secretes bicarbonate ions while it simultaneously actively absorbs equal amounts of chloride ions in an exchange transport process (Cl-HCO3- counter-transport). The bicarbonate helps neutralizing the acidic end products of bacterial action in the colon. Intestinal Ca2+ absorption (figure 5.20): Ca2+ is reabsorbed in the duodenum and upper jejunum. About 30– 80% of dietary Ca2+ typically is absorbed (figure 5.20) and is exactly controlled in relation to the need of the body for Ca2+. [A] About one third of the net Ca2+ uptake occurs passively via the paracellular route across the tight junctions. [B] About two third of the net Ca2+ uptake occurs actively through regulated Ca2+ transport pathways in the enterocytes and is stimulated by vitamin D as follows: ■ Ca2+ enters the enterocytes due to electrochemical gradient via the Ca2+ channels. ■ to prevent an increase in intracellular free Ca2+ concentration, Ca2+ binds to the protein calbindin within the cytoplasm, the synthesis of which is totally dependent on vitamin D. ■ Extrusion of Ca2+ from the cells occurs by primary active transport (Ca2+-ATPase) and secondary active transport (3Na+/Ca2+ exchange), against a large electrochemical Figure 5.20: Regulation of intestinal Ca2+ absorption. 41 Mustansiriyah College of Medicine gradient. Vitamin D increases synthesis of calbindin and both apical and basolateral transporters. It is inhibited by phosphates and oxalates because these anions form insoluble salts with Ca2+ in the intestine. Fe++ ions are also actively absorbed from the small intestine which is regulated according to the body need. K, Mg, phosphate, and other ions can also actively absorb through the mucosa. The vitamin B12-intrinsic factor complex is propelled along the small intestine to the terminal ileum, where specific transporters located on the enterocyte microvilli bind the vitamin B12intrinsic factor complex. Binding requires calcium and is optimal at pH 6.6. Absorption is an active transport process. Figure 5.21: Intestinal NaCl and water secretion. Intestinal secretions: [A] Fluid secretion: Both the small intestine and the large intestine secrete fluid from the crypt cells. Secretion is necessary for lubrication, as evidenced by the high incidence of obstruction of the gut when secretion is impaired in diseases such as cystic fibrosis. The mechanism of intestinal fluid secretion is clinically important because it is activated by a number of bacterial enterotoxins that cause secretory diarrhea. The key step in the mechanism of fluid secretion from the crypt enterocytes is opening of the Cl− channels in the luminal cell membrane. There are two types of Cl− channels present (figure 5.21) 1. cAMP-activated Cl− channels, for which the enteric nervous system (ENS) neurotransmitter VIP is an important stimulator. Na+ secretion follows passively via the paracellular pathway, and water secretion occurs by osmosis. 2. Ca2+-activated Cl- channels, for which acetylcholine from ENS neurons and serotonin from the enterochromaffin are both stimulators. Figure 5.21A summarizes the intestinal absorption and secretion. Enterotoxic Escherichia coli (traveler’s diarrhea due to E coli) and Vibrio cholera (cholera due to V cholera) both produce enterotoxins. The toxin from V. cholerae (cholera toxin) causes an irreversible increase in cAMP levels (not cGMP levels) in the enterocytes located in the crypts of Lieberkühn of the small intestine. This increase in cAMP causes an irreversible opening of chloride channels on the luminal membrane. Movement of chloride ions into the gut lumen causes a secondary movement of sodium ions to maintain electrical neutrality. Water follows the osmotic gradient created by sodium and chloride, causing a tremendous increase in fluid loss into the gut lumen. Severe diarrhea follows. 42 Mustansiriyah College of Medicine Loss of gastric juice results in hypokalemic, metabolic alkalosis. Excessive loss of fluid from the gastrointestinal tract can lead to dehydration and, depending upon the origin of the fluid loss, electrolyte and acid-base disturbances. The hydrogen ion and potassium ion concentration of gastric juice exceeds that of the plasma. As a result, excess fluid loss leads to metabolic alkalosis accompanied by hypokalemia. Because the pancreas, liver, ileum, and colon secrete bicarbonate, excessive loss leads to metabolic acidosis. In addition, the colon secretes potassium. Thus, the acidosis is accompanied by hypokalemia. The small intestinal glands normally secrete 1 to 2 L of intestinal juice daily. Normally, intestinal juice is slightly alkaline (7.4–7.8), and isotonic with blood plasma. Intestinal juice is largely water but also contains some mucus, which is secreted both by the duodenal glands and by goblet cells of the mucosa. The major stimulus for intestinal juice production comes from hypertonic or acidic Figure 5.21A: Summary of intestinal absorption and chyme, distension or irritation of the intestinal secretion. mucosa. Whenever a segment of large intestine becomes intensely irritated or infected, the mucosa then secretes large amounts of water and electrolytes in addition to the normal viscid solution of alkaline mucus. This acts to dilute the irritating factors and to cause rapid movement of the feces toward the anus. The usual result is diarrhoea with loss of large quantities of water and electrolytes but also earlier recovery from the disease than would otherwise occur. [B] Mucus: Mucus in small and large intestine contains bicarbonate ions to protect the mucosa of small intestine from acidic chyme and the mucosa of large intestine from the acidic end products of bacterial action in the colon. Small intestinal mucus is secreted by Brunner’s glands, which are located within the duodenum, and by Goblet cells, which are located along the length of intestinal epithelium. The mucus is secreted also in response to: Parasympathetic vagal stimulation, and to, Decrease pH of the duodenum to 4 or less GIT hormones especially secretin. Brunner’s glands are inhibited by sympathetic stimulation which may lead to peptic ulceration. In the large intestine the rate of secretion of mucus is regulated principally by: Direct tactile stimulation of the mucus cells on the surface of the mucosa and by, Local nervous reflexes to the mucous cells in the crypts of Lieberkuhn. 43 Mustansiriyah College of Medicine Parasympathetic innervation to the distal two thirds of large intestine also causes marked increase in secretion of mucus. This occurs along with an increase in motility. Therefore, during extreme parasympathetic stimulation, often caused by emotional disturbances, so much mucus may be secreted into the large intestine that the person has a bowel movement of ropy mucus often as every 30 min; the mucus contains little or no fecal material. Mucus in the large intestine protects the wall against excoriation, but in addition, it provides the adherent medium for holding fecal matter together. Furthermore, it protects the intestinal wall from the great amount of bacterial activity that takes place inside the feces, plus the alkalinity of the secretion (pH of 8.0 caused by large amounts of sodium bicarbonate) also provides a barrier to keep acids formed deep in the feces from attacking the intestinal wall. The large intestine: The contractile activity of the large intestine serves two functions: [1] It enhances the efficiency of water and electrolyte absorption. [2] It promotes the excretion of fecal material remaining in the colon. Movement of the colon: There are three types of movements in the colon and these are: [1] Mixing movements (segmentation, and haustration). [2] Peristalsis. [3] Mass movements: A mass movement is a modified type of peristalsis characterized by the following sequence of events: 1. A constrictive ring occurs at a distended or irritated point in the colon, usually in the transverse colon, 2. Then rapidly thereafter, the 20 or more centimeters of colon distal to the constriction contract almost as a unit, forcing the fecal material in this segment en masse down the colon. During this process, the haustration disappear completely. 3. After the contraction is completed, then relaxation occurs before another mass movement takes place, this time perhaps farther along the colon. When the mass movements have forced the feces into the rectum, the desire for defecation is felt. When the mass movements have forced the feces into the rectum, the desire for defecation is felt. The appearance of mass movements is facilitated by: Gastrocolic and duodenocolic reflexes as a result of distension of stomach and duodenum respectively. These reflexes are transmitted mainly through extrinsic nerves and only weakly through myenteric plexus. This explains the appearance of mass movement and urge to defecate after meals. They are greatly suppressed when the extrinsic autonomic nerves to the colon have been removed. Irritation of the colon can also initiate intense mass movement. Intense stimulation of the parasympathetic nervous system. Overdistension of a segment of the colon. Defecation reflexes: The average transit time from caecum to pelvic colon is about 12 hours but passage from the pelvic colon to the anus may take days. When a mass movement forces feces into the rectum, the desire (urge) for defecation is normally initiated, including reflex contraction of the rectum and relaxation of the anal sphincters. However, defecation is prevented by tonic constriction of 1. The internal anal sphincter which is a circular mass of smooth muscle that lies immediately inside the anus and supplied by sympathetic nerve fiber which is excitatory and by parasympathetic fibers which is inhibitory. 2. The external anal sphincter which is composed of striated voluntary muscle that surrounds the internal sphincter and also extends distal to it. The external sphincter is controlled by nerve fibers in the pudendal 44 Mustansiriyah College of Medicine nerve, which is part of the somatic nervous system and therefore is under voluntary conscious control. When the feces enter the rectum, distension of the rectal wall stimulates stretch receptors and initiates defecation reflex through parasympathetic nerve fibers and involves three processes (figure 5.22): 1. The sensory fibers pass through the pelvic nerves to the sacral segments of the spinal cord. 2. The spinal cord returns signals by way of the parasympathetic fibers in the pelvic nerves to the enteric nervous system and produces an effective and strong peristaltic wave in the sigmoid and rectum forcing feces toward anus. 3. At the same time, parasympathetic fibers relax the internal anal sphincter to allow the passage of feces. 4. Both the internal and external anal sphincters must relax for feces to be eliminated. However, the reflexes just mentioned open the internal sphincter but close the external sphincter. The actual release of feces requires a conscious effort to open the external sphincter. If circumstances Figure 5.22: Defecation reflex. are unfavorable for defecation, voluntary tightening of the external anal sphincter can prevent defecation despite the defecation reflex. If defecation is delayed, the distended rectal wall gradually relaxes, and the urge to defecate subsides until the next mass movement propels more feces into the rectum, once again distending the rectum and triggering the defecation reflex. If circumstances are favorable for defecation, voluntary relaxation of external anal sphincter can override the contraction directed by somatic motor neuron and consequently defecation occurs. When defecation does occur, it is usually assisted by voluntary straining movements, such as taking a deep breath, closure of the glottis, and contraction of the abdominal muscles (straining) to force the fecal contents of the colon downward, at the same time causing the pelvic floor to extend downward to pull outward on the anus to evacuate the feces. If the external sphincter is kept contracted, the defecation will not occur, and the defecation reflexes die out after a few minutes, and they remain quiescent for several hours or until additional amounts of feces enter the rectum. 45 Mustansiriyah College of Medicine Distention of the stomach by food initiates contraction of the rectum and frequently a desire to defecate. This response is called the gastrocolic reflex, which is mediated by parasympathetic fibers, although there is some evidence that it is due to an action of CCK and gastrin on the colon and is not neurally mediated. Because of the response, defecation after meals is the rule in children. In adults, habit and cultural factors play a large role in determining when defecation occurs. Emotional factors strongly influence large intestinal motility via the extrinsic autonomic nervous system. Irritable bowel syndrome may occur during periods of stress and may result in constipation (increased segmentation contraction) or diarrhea (decreased segmentation contractions). Bacterial action in the colon: Numerous bacteria especially colon bacilli, are present in the absorbing colon. The substances formed as a result of bacterial activity are vitamin K, vitamin B 12, thiamin, riboflavin, and various gases that contribute to flatus in the colon especially carbon dioxide, hydrogen gas, N2, NH4+ and methane. One of the actions of colonic bacteria is to convert NH3 to NH 4+. Thus, a total colectomy would increase blood ammonia levels, which would be exacerbated in a person with liver cirrhosis. Gas within the colon is primarily derived from Fermentation of undigested oligosaccharides by bacteria. Vitamin K is especially important, for the amount of this vitamin in the ingested food is normally insufficient to maintain adequate blood coagulation. The colonic gas, or flatus, is produced in large intestine. The gas is produced chiefly through the breakdown of undigested carbohydrates that reach the colon. The amount of gas varies markedly from one person to another and is influenced by diet; for example, ingestion of large amounts of beans, which contain indigestible carbohydrates in their hulls, will increase gas formation by intestinal bacteria. All gases except N2 diffuse readily through the intestinal mucosa. Diffusion of gas from the blood to the intestinal lumen is responsible for the N2 present in intestinal gas and is influenced by the atmospheric pressure. The feces normally are composed of dead bacteria (bacteria such as Escherichia coli make up about a third of faecal weight), fat (derived from fat formed by bacteria and fat in the sloughed epithelial cell), inorganic matter, protein, undigested roughage of the food and dried constituents of digestive juices such as bile pigment and sloughed epithelial cells. The brown color of feces is caused by stercobilin and urobilin, which are derivatives of bilirubin. The odor is caused by the products of bacterial action which vary from one person to another depending on each person’s colonic bacterial flora and on the type of food eaten. The liver: The total blood flow through the liver is about 1450 ml/min. It has double blood supply from hepatic artery (20%) and portal vein (80%). The portal vein drains the digestive tract, the spleen, pancreas and gallbladder. The liver lobule is constructed around a central vein that empties into the hepatic veins and thence into the vena cave (figure 5.23). The venous sinusoids are lined by two types of cells [1] typical endothelial cells and [2] large Kupffer cells which are tissue macrophages capable of phagocytizing bacteria and other foreign matter in the blood. These cells can remove 99% of bacteria in the portal venous blood before they can pass all the way through the liver sinusoids. The endothelial lining of the venous sinusoids has extremely large pores, therefore, the plasma substances and even the proteins can move freely from the blood through the endothelium to the space below it and become in direct contact with hepatic cells. Because of the pores in the endothelium of the hepatic sinusoids, the plasma proteins can pass to the interstitial spaces. Therefore the lymph draining from the liver usually has a protein concentration of about 6 gm/dl, which is only slightly less than the protein concentration of plasma. Also, the extreme permeability of the liver sinusoids allows large quantities of lymph to form. Therefore, about one half of all the lymph formed in the body under resting conditions arises in the liver. 46 Mustansiriyah College of Medicine Pressure and resistance in hepatic vessels: The pressure in the hepatic vein leading from the liver into the vena cava is about 0 mm Hg, whereas the pressure in the portal vein leading into the liver is about 9 mm Hg. This means that the resistance to blood flow through the liver sinusoids is normally low. However, in some pathological conditions such as liver cirrhosis, the hepatic vascular resistance may increase markedly because many vascular channels Figure 5.23: The basic histological structure of a liver lobule. through the liver are severely restricted by fibrotic constriction of the sinusoids or even complete blockage or destruction. An increase in pressure in the veins draining the liver dams blood in the liver sinusoids and thereby causes the entire liver to swell markedly and congested. The most, common cause of hepatic congestion is cardiac failure. When the hepatic venous pressure rises only 3-7 mm Hg above normal, excessive amounts of fluid begin to transudate into the lymph and also to leak through the outer surface of the liver capsule directly into the abdominal cavity. This fluid is almost pure plasma, containing 80 - 90% as much protein as normal plasma which may be so large that lead to ascitis (free fluid in the abdominal cavity) especially when the hepatic venous pressure is between 10-15 mm Hg. Functions of the liver [1] Carbohydrate metabolism Storage of glycogen: The liver is especially important for maintaining a normal blood glucose concentration. For instance, storage of glycogen allows the liver to remove excess glucose from the blood, store it, and then return it to the blood when the blood glucose concentration begins to fall too low. This is called the glucose buffer function of the liver. Circulating adrenaline and glucagon mobilize this liver glycogen and convert it to blood glucose. Only liver has the glucose-6-phosphatase necessary for this breakdown. Conversion of monosaccharides galactose and fructose to glucose. Gluconeogenesis. When the glucose concentration begins to fall below normal, large amounts of amino acids are converted into glucose, thereby contributing still another means for maintaining relatively normal blood glucose. Formation of many important chemical compounds from the intermediate products of carbohydrate metabolism. [2] Fat metabolism Bile salts produced by the liver are essential for the digestion and absorption of fat. The fat-soluble vitamins are stored in the liver such as vitamins A and D. In addition, water-soluble 47 Mustansiriyah College of Medicine vitamins such as B12 are also stored in the liver. The depot fat is mobilized when required and is converted to the ketone bodies (acetoacetic acid, βhydroxybutyric acid and acetone) by the liver. These ketone bodies are metabolized by other tissues with production of heat and energy. Ketosis, which is an excess of ketone bodies in the blood, does not occur in liver failure. The liver is the site for the synthesis of large quantities of cholesterol and phospholipids. The liver is the site for conversion of large quantities of carbohydrates and proteins to fat. [3] Protein metabolism The liver is the main organ for deamination of surplus amino acids with the production of ammonia. In liver failure, the amino acids in the blood are increased and they appear in the urine. The liver is the only organ that can convert ammonia to urea. . Ammonia is a toxic gas which is difficult to be excreted out while urea is less toxic and easier excreted. In the absence of this function of the liver to form urea, the plasma ammonia concentration rises rapidly and results in hepatic coma and death. Among the most important functions of the liver is its ability to synthesize certain amino acids and also to synthesize other important chemical compounds from amino acids. For instance the non-essential amino acids can all be synthesized in the liver. [4] Blood The liver plays an important role in the formation and destruction of RBC. The liver is a site of formation of RBC in fetal life. It stores the factors principal for the normal maturation of RBC such as iron and B 12. It removes from the blood the bilirubin formed when the RBC are broken down and excretes this bilirubin down the bile duct into the duodenum, as a bile pigments. The liver manufactures the plasma proteins with the exception of some of the globulin fraction (which is the antibodies formed mainly by plasma cells in the lymph tissue of the body). The plasma albumin exceeds the plasma globulin and the normal albumin/globulin ratio is 1.7/1. In liver failure the plasma protein pattern is altered and the globulin fraction may exceed the albumin fraction. The blood clotting factors such as prothrombin and fibrinogen, etc.., are made in the liver. In liver diseases, blood clotting may be impaired leading to bleeding from the skin and mucous membranes. [5] Vitamin Mineral Storage: Vitamin A, Vitamin D, Vitamin E, Vitamin K, vitamin B12, and Iron. [6] Poisons: Many normal physiological substances as well as drugs are modified by the liver in some way before being excreted by the kidneys. This is called detoxication. Some substances, such as the short-acting barbiturates, are completely destroyed by the liver. Other, substances are conjugated with glucuronic acid, glycine, sulphate or acetate. 48