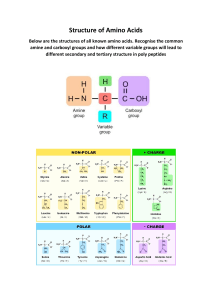
Chapter 1: Chemical Basis of Life Four Major Biomolecules I. II. III. IV. Amino Acids: contain an amino group and a carboxylic group and an side chain/R group Under physiological conditions, ionized to NH3+ and COO Ex. Alanine and Asparagine Carbohydrates: (monosaccharides/sugars) Formula = (CH2O)n Ex. Glucose Nucleotides: five-carbon sugar, nitrogen-containing ring, and one or more phosphate groups. Ex. Adenosine Triphosphate (ATP) Lipids: poorly soluble in water due to its bulky hydrocarbon-like structure. Ex. Palmitate and Cholesterol 3 Major Biological Polymers I. II. III. Proteins (Polypeptides): Polymers of amino acids. Amino acids linked via peptide bonds. Can perform tasks such as mediating chemical and provide structural support. Nucleic Acids (Polynucleotides): polymers of nucleotides; RNA Primary Role: carriers of genetic information, contained in sequence of nucleotide residues rather than their 3D RNA: nucleotides – adenine, cytosine, guanine, and uracil DNA: nucleotides – adenine, cytosine, guanine, and (ACGT) reactions DNA and their shape. (ACGU) thymine Polysaccharides: contain only one or a few different type of monosaccharide residue Limited potential for carrying genetic information in the sequence of their residues or for adopting a large variety of shapes and mediating chemical reactions Serve as fuel-storage molecules and provide structural support. Linked via glycosidic bonds Energy and Metabolism 1st Law of Thermodynamics: Energy cannot be created or destroyed. Free Energy (Gibbs Free Energy, G, units: J*mol-1) o Enthalpy (H): heat content of the system o Entropy (S): measure of how the energy is dispersed within that system; measure of the system’s disorder/randomness. G is less than zero for a spontaneous process. For a process to occur, the overall change in free energy (G) must be negative. o A reaction with G<0 (negative) has a decrease in enthalpy and an increase in entropy is spontaneous at all temperatures. o A nonspontaneous or endergonic reaction has a free energy change greater than zero; the reverse reaction is spontaneous. Thermodynamic spontaneity does NOT indicate how fast a reaction will occur, but if it WILL* *Dependent on molecules, temperature, or presence of catalyst. At equilibrium o A reaction with a decrease in enthalpy and an increase in entropy is spontaneous at all temperatures. nd 2 law: energy tends to spread out o Enthalpy increasing while entropy decreasing is NOT possible o If enthalpy and entropy both increase or both decrease during a reaction, the value of G then depends on the temperature which governs whether T S term is greater or less than the H term Life is thermodynamically possible In order to exist, life must be thermodynamically spontaneous. o Many of the cells metabolic reactions have free energy changes that are less than zero but some reactions do not. o The nonspontaneous reactions are reactions are able to proceed in vivo (in a living organism) because they occur in concert with other reactions that are thermodynamically favorable. Reduction, the gain of electrons is accomplished by the addition of hydrogen or the removal of oxygen. The plant or animal that eats can then break down the monosaccharide to use it as fuel to power other metabolic activities. In the process the carbon is oxidized it loses electrons through the addition of oxygen or the removal of hydrogen and ultimately becomes CO2. The oxidation of carbon is thermodynamically favorable so it can be coupled to energy requiring processes such as the synthesis of building blocks and their polymerization to form macromolecules Mostly all metabolic processes occur with the aid of catalysts called enzymes which most are proteins An organism with its high level of organization represents a state of low energy relative to its surroundings. (high organization/stability= low energy) the organism can maintain this thermodynamically unfavorable state as long as it continually obtains free energy from its food when it exhausts food, G= 0 resulting in death The Origin and Evolution of Life the ability to replicate is one of the universal characteristics of living organisms cells will carry out a set of instructions but will evolve throughout time The prebiotic world Theories: o H2, H2O, NH3, and CH4 could have given rise to simple biomolecules such as amino acids, when sparked by lighting. o HCN, HCOH, and phosphate could have been converted to nucleotides o Charles Darwin “warm little pond theory” Similar molecular building blocks could have accumulated and formed larger structures, particularly in shallow waters where evaporation would have had a concentrating effect. The first biological building blocks would have had to polymerize by gaining the capacity for self-replication. Complement- mirror image copy Origins of modern cells o Replicating molecules chances of decreasing depend on natural selection o This favors a replicator that was chemically stable and had a ready supply of building blocks and free energy for making copies of itself CO2 + H2O (CH2O) + O2 The first cells were able to fix CO2 that is convert it to reduced organic compounds using free energy released in the oxidation of readily available inorganic compounds such as H2S or Fe2+. Later on photosynthetic organisms similar to present day cyanobacteria used the suns energy to fix CO2 Prokaryotes: small unicellular organisms that lack a discrete nucleus and usually contain no internal membrane systems. o This group is composed of two groups Archaea and eubacteria Eukaryotes: cells usually larger than prokaryotic cells and contain a nucleus and other membrane bounded cellular compartments. May be unicellular or multicellular. o This group called the eukarya includes microscopic organisms as well as familiar macroscopic plants and animals o Eukaryotic cells exhibit characteristics exhibit characteristics of both bacteria and archaea o Eukaryotic cells also contain organelles that are almost certainly the descendants of free living cells The number of sequence differences between two groups of organisms indicates how long ago they diverged from a common ancestor. Species with similar sequences have a longer shared evolutionary history than species with dissimilar sequences Chapter 2: Aqueous Chemistry Water Molecules form Hydrogen Bonds Water is 60% by mass in humans Characteristics of water o Tetrahedral Bent Geometry o Polar; Partial negative and positive charges in oxygen and hydrogen o In Ice forms lattices, bonds with 4 H2O molecules o In water each bond is short, molecules flicker o Many biomolecules have atoms (hydroxyl, carbonyl, amine, amide) that can Hydrogen Bond with water. Water is highly cohesive o Accounts for its high surface tension o Explains why water remains a liquid Ionic Interactions (Electrostatic Forces in water) Intermediate in strength o Hydrogen Bonds In DNA In Proteins o Van-der-waals forces: between polar molecules o London-dispersion: between non-polar molecules Why do some drugs contain fluorine? Introduction of fluorine allowed alteration to drug’s chemical/biological properties without significantly altering the shape. Fluorine can take place of hydrogen o High electronegativity, behaves more like Oxygen Transforms C-H group to an electron-withdrawing polar C-F group, decreasing basicity. o Fewer positive charges allow a drug to pass more easily through the membrane o C-F bond can participate in hydrogen bonding/dipole-dipole interactions Better binding = more effective at lower concentrations and fewer side effects. Water dissolves many compounds Water has a high dielectric constant o Solvent’s ability to diminish the electrostatic attractions between dissolved ions o Higher dielectric constant, less able the ions to associate with each other. The hydrophobic effect: Why is it thermodynamically unfavorable to dissolve a hydrophobic substance in water? Enthalpy is needed to break the hydrogen bonds in water to create a hole for non-polar molecules to fit. o The free energy barrier in solvation is more dependent on entropy o When a hydrophobic molecule is hydrated, a layer of water molecules that cannot participate in hydrogen bonding and align themselves so the polar ends are not oriented toward the nonpolar solute. Loss of entropy in the system Non-polar molecules clump/aggregate in water because they are driven out by unfavorable entropy cost of individual hydration Hydrophobic Effect: exclusion of nonpolar substances from an aqueous solution o Do not experience any additional attractive force Amphiphilic molecules experience both hydrophilic interactions and the hydrophobic effect Amphiphilic molecules: have both hydrophobic and hydrophilic portions Ex. Nonpolar hydrocarbon tail and a polar carboxylate head Polar groups of amphiphilic molecules are oriented towards the polar solvent and are hydrated, nonpolar groups aggregate due to the hydrophobic effect to form a spherical micelle. Can form a sheet to form bilayers Hydrophobic Core of lipid bilayer is a barrier to diffusion Lipid bilayers close up to form a vesicle o Traps a volume of aqueous solution, polar solutes remain bc they cannot pass through the hydrophobic interior of the bilayer (too high of energetic cost) o Bilayers can prevent diffusion of high concentrated substances to lower concentration (movement down a concentration gradient, increase of entropy) Sweat, Exercise and Sport Drinks Heat due to metabolic activity can be lost by the vaporization of water, which has a significant cooling effect. o Increase in skin temperature triggers activity of sweat glands to secrete a solution of Na+, K+, and Cl Exercise at high temperatures can cause fluid loss (up to 2L per hour) o Fluid loss >2% of body’s weight can impair cardiovascular function Fluid intake should match losses due to sweat; rate of intake should be equal with rate of sweating o For activities less than 90 minutes, water is ok Sports Drinks have high carbohydrate content, but only advantageous during prolonged sustained activity (ex. marathon) Normal Diets usually contain enough Na+ and Cl- to match losses in sweat Acid-Base Chemistry Equilibrium constant for Ionization of water o 2H2 O ⇌ H+ + OH− o Kw (ionization constant for water) = [H+][OH-] = 10-14 Has no units Defining pH o Acidic Solutions: [H+] > 10-7 o Basic Solutions: [H+] < 10-7 A pK value describes an acid’s tendency to ionize Ka, Acid Dissociation Constant = [𝐶𝐻3𝐶𝑂𝑂−][𝐻+] [𝐶𝐻3𝐶𝑂𝑂𝐻] pK = -log (Ka) Larger an acid’s Ka, the smaller its pK and the greater its strength Henderson-Hasselbach equation: relates pH of a solution to the pK of an acid and the concenctration of the acid (HA) and its conjugate Base (A-) pH = pK + log [Base] [Acid] pK and pH convey protonation state o At pH < 3.5, amino acids are protonated Net Positive Charge (+1) COOH and NH3+ o At 3.5 < pH < 9.0, carboxyl is deprotonated Charge is neutral (0) Zwitter Ions o At pH > 9.0, both groups are deprotonated Net negative charge (-1) COO- and NH2 Rule of thumb: pH = pK +/- 1 o When [A] = 10[HA] pH = pK + log (10) = pK + 1 o When [HA] = 10 [A-] pH = pK log (0.1) = pK – 1 Buffers When HCl is added to a weak acid in equilibrium with its conjugate base, pH does not change dramatically because some added protons combine with the conjugate to reform the acid and will not contribute to increase [H+] Effective buffering capacity: generally within one pH unit of its (pK = +/- pH) Chapter 4 Protein Structure Protein: consists of one or more polypeptides (chains of polymerized amino acids) Amino Acid: small molecule with an amino (NH3+), a carboxylate (COO-), and R-group o R group (side chain): determines biological reactivity 20 Amino Acids Hydrophobic Amino Acids o Alanine (Ala, A) o Valine (Val, V) o Phenylalanine (Phe, F) o Tryptophan (Trp, W) o o o o Leucine (Leu, L) Isoleucine (Ile, I) Methionine (Met, M) Proline (Pro, P) Polar Amino Acids o Serine (Ser, S) o Threonine (Thr, T) o Tyrosine (Tyr, Y) o Cysteine (Cys, C) o o o o Asparagine (Asn, N) Glutamine (Gln, Q) Histidine (His, H) Glycine (Gly, G) Charged Amino Acids o Positive/Basic Histidine (His, H) Arginine (Arg, R) Lysine (Lys, K) o Negative/Acid Aspartate (Asp, D) Glutamate (Glu, E) (Refer to flashcards and/or sheet for structures and more information) Amino Acids are linked via condensation reaction (water molecule is eliminated) o Peptide bonds: linkage of two amino acids to form a polypeptide o Residues: amino acids within the peptide o Can be broken/hydrolyzed via exo or endopeptidases The chain read from the N-terminus to the C-terminus o N-terminus has free amino group o C-terminus has free carboxyl gr Sample Calculation. Estimate net charge of the polypeptide chain: Ala-Arg-Val-His-Asp-Gln 1. At pH = 7.4 2. At pH = 5.0 Sample problem. Draw the structure of the peptide Gly-Lys in its fully protonated forms. Which ionic form predominates at (a) pH = 1, (b) pH = 7, (c) pH = 13 Four Levels of Protein Structure I. II. III. IV. Primary Structure: sequence of amino acid residues Secondary Structure: localized conformation of the polypeptide backbone Tertiary Structure: 3-D structure of an entire polypeptide, with all its side chains Quaternary Structure: spatial arrangement of polypeptide chains in a protein with multiple subunits Primary Structure: sequence of amino acid residues Secondary Structure: localized conformation of the polypeptide backbone Rearrangement of atoms attached in backbone of protein forms repeating structural units that are maintained by H-bonds contributed to atoms in the main backbone Restricted rotation of peptide bonds due to the two resonance forms of the peptide bond o Groups involved in peptide bonds are strongly polar. o Rotation around C-C and N-C determines secondary structures of proteins The polypeptide chain folds to satisfy as many of the hydrogen-bonding requirements as possible. o Must adopt a conformation that minimizes steric strain Two kinds of secondary structures I. -Helix: twisted backbone conformation II. -Sheet: contains multiple polypeptide strands These are both regular secondary structures: component residues have backbone conformations that are the same from one residue to the next -Helix: exhibits a twisted backbone conformation Rod-like tightly coiled structure H-bonds form between carbonyl oxygen and amino hydrogen H-bonds are parallel to the main axis 3.6 residues per turn -Pleated Sheets: extended chains of polypeptide sheets arranged in two ways Parallel -Pleated Sheets: chains run in the same direction Anti-Parallel -Pleated Sheets: chains run in opposite directions Single sheet may have 2 to more than 12 polypeptide strands Irregular secondary structures (loops and coils) o Loops that link strands or helices have residues with irregular secondary structures o The polypeptide DOES NOT adopt a defined secondary structure (doesn’t mean disordered) o In every protein, elements of the secondary structure are linked together by polypeptide loops. Proteins can have a combination of secondary structures o Combination: usually the tertiary structure Tertiary Protein Structure: 3D shape of entire polypeptide and spatial arrangement of all its side chains Overall folding of the peptide backbone, includes its regular and irregular secondary structures o X-Ray Crystallography: determined protein structure (ex. myoglobin) o Globular Proteins Types of interactions that maintain tertiary structures I. Hydrophobic Interactions II. Hydrogen Bonding III. Disulfide Bridges IV. Ionic bonds Proteins have hydrophobic cores Globular proteins have at least 2 layers of secondary structures; protein has a definite surface and core regions o Protein has a hydrophilic surface and a hydrophobic core o The greater a residue’s hydrophobicity, the more likely it is in the protein interior. Ion Pair: when a charged residue is in the protein interior, it is located near another residue with an opposite charge for them to interact electrostatically Domain: polypeptide segment that has folded into a single structural unit with a hydrophobic core A two domain protein Protein structure stabilization via Hydrophobic Effect The largest force governing protein structure: Hydrophobic Effect o Driven by the increase in entropy of surrounding water molecules o Hydrophobic side chains in interior of protein Stabilizes the folded polypeptide backbone Hydrogen bonding (by itself) is NOT a major determinant of protein stability o Helps by fine-tuning the folded conformation that is stabilized by the hydrophobic effect Cross-links help stabilize proteins I. Ion Pairs II. Disulfide bonds III. Inorganic ions (Zinc Fingers) Ion Pairs: form between oppositely charged side chains or N- and C- terminal groups Strong electrostatic interaction But doesn’t contribute much to protein stability; favorable free energy of electrostatic interaction is offset by loss of entropy when side chains are fixed. Disulfide Bonds: form within and between polypeptide chains Not essential for stabilizing proteins More plentiful in extracellular (oxidizing) environments, prevent protein unfolding under relatively harsh extracellular conditions Zinc Fingers: structures of 20-60 residues with one or two Zn2+ ions o Common in DNA-binding proteins o Ideal ion for stabilizing proteins; can interact with ligands (S,N, or O) provided by amino acids and only has one oxidation state Protein Folding: begins with formation of secondary structures In cell, a newly synthesized polypeptides folds as soon as it emerges from the ribosome; part of the polypeptide may adopt its mature tertiary structure before entire chain is synthesized o Denatured: chemically unfolded Addition of highly soluble substances (ex. salts or urea) that interfere with the hydrophobic effect causing unfolding of the proteins o Renatured: refolding Protein renaturation is NOT random; approaches the native structure (most stable tertiary structure) through alternate pathways Small elements of secondary structure form first then come together due to the hydrophobic effect to produce a mass with a hydrophobic core; small rearrangements yield the native structure The information required for protein folding is all contained in the amino acid sequence. Molecular chaperones are smaller proteins that assist in protein folding. Proteins undergo processing before reaching mature forms Removal of amino acid residues Covalent attachment of a group (lipid, carbohydrate, or phosphate) which has a discrete biological function and helps stabilize the folded conformation of a protein Association with metal ions/specific organic molecules Proteins can adopt more than one conformation Minor movement bending/stretching of individual bonds is required for biological function Intrinsically Unstructured proteins (protein segments): highly flexible extended segments rich in hydrophilic amino acids The flexibility of a protein’s conformation has two stable alternatives in dynamic equilibrium that can tip between different conformations o Change in cellular conditions (pH or oxidation state) o Presence of a binding partner Protein Misfolding and Disease Degradation of a protein to its component amino acids is a mechanism in which cells can deal with misfolded proteins. Cystic fibrosis: mutated for of the CFTR protein folds incorrectly and never reaches cellular destination Diseases resulting from aggregated misfolded proteins (fibrous deposits - long, insoluble fibers) are deadliest when in the brain o Ex. Diseases characterized by fibrous deposits: Alzheimer’s, Parkinson’s and transmissible spongiform encephalopathies; lead to neurological abnormalitiy and loss of neurons o Amyloid deposits: aggregated proteins Alzheimer’s Disease (neurodegenerative): accompanied by intracellular tangles and extracellular plaques Tau: fibrous protein material inside cell involved in assembly of microtubules Excess amyloid- linked to Alzheimer’s disease Early stages of amyloid- misfolding and aggregation are toxic to neurons; ultimate cause Parkinson’s disease: neurons in a portion of the brain accumulate -synuclein protein -synuclein: forms alpha helices upon binding to other molecules; contributes to propensity to form amloid deposits Mutations in the gene for alpha-synuclein lead to increased expression or promote self-aggregation. Transmissible Spongiform Encephalopathies (TSE) (or Mad Cow Disease) Infectious agent is a protein called the prion Scrapie form of prion (PrPSC) Quaternary Structure: spatial arrangement of polypeptide chains in a protein with multiple subunits Two or more subunits oriented in 3D space o Due to noncovalent interactions Subunits vs. Domains o Domains: single chain that can form a local 3D structure; folded polypeptide segments Independent structural unit with a hydrophobic core o Subunit: two or more separate chains in a protein that orient in 3D space to give quaternary structures Domain Subunits Subunits can be identical are named homodimers, homotetramers, etc. o Nonidentical chains have a prefix of hetero There are similar forces that hold subunits together in tertiary structures o Hydrogen bonds; ion pairs; and disulfide bonds These interactions give an opportunity for subunits to influence each other’s behavior/cooperativity; regulates functions not possible in single-subunit proteins Advantages of multisubunit protein structure: o Construct extremely large proteins through incremental addition of small building blocks o Decreases the impact of errors in transcription and translation Tools and Techniques for Analyzing Protein Structure Chromatography: takes advantage of a polypeptides unique properties Proteins or other solutes pass through the column at different rates, depending on how they react with the stationary phase (polar; porous matrix). Size-Exclusion Chromatography (gel filtration chromatography) o Stationary phase has tiny beads with pores of characteristic size. o Larger proteins are excluded and pass through the column faster than smaller proteins. Ion-Exchange Chromatography o Exploiting a protein’s net charge at a particular pH. o Solid phase can have positively charged diethylaminoethyl (DEAE) or negatively charged carboxymethyl groups (CM) Negatively charged proteins bind to DEAE Postively charged proteins pass through column o Anionic groups become protonated Isoelectric point (pI): pH at which a protein carries no net charge = ½ (pK1 + pK2) Affinity chromatography: separation via the basis of high affinity of the protein of interest for specific chemical groups o Takes advantage of unique ability to interact with another molecule o Ex. Isolation of Concanavalin A, which binds to glucose Column has glucose to bind to ConA High-performance liquid chromatography (HPLC): carried out in closed columns under high pressure Standard approach to sequencing proteins I. Protein sample is purified II. Chains are separated III. Large polypeptides are broken into smaller pieces for individual sequencing IV. Sequencing (Edman’s Degradation): N-terminus is chemically derivatized, cleaved off and identified V. Reconstruct sequence; different set of fragments that overlap first is generated Mass spectrometry: more efficient; measures the sizes of peptide fragments o Determines the sequence of amino acids in a polypeptide Protein structures determined by X-ray crystallography, electron crystallography and NMR spectroscopy X-Ray crystallography: performed on samples of proteins that have been crystallized o X-ray’s bombard and electrons in crystals scatter the x-rays to produce a diffraction pattern o Trace the polypeptide backbone and discern general shapes o Crystallized proteins retain some ability to move Electron Crystallography: electron beams probe the protein’s structure o Proteins do not have to be crystallized o 3D structures can be reconstructed Nuclear Magnetic Resonance (NMR) spectroscopy: takes advantage of atomic nuclei (hydrogen) to resonate in an applied magnetic field o Spectrum consists of peaks that are analyzed to reveal distances between two H atoms close or covalently connected to other atoms in space o Constructs a 3D model of the protein Chapter 5: Protein Function Proteins have various functions: I. Transport: myoglobin transports oxygen throughout muscles; hemoglobin transports oxygen in blood II. Structural: actin forms microfilaments in cells; tubulin dimers constitute microtubules; keratin filamines in animal hair; collagen is a major protein in connective tissue III. Motor function: myosin interacts with actin to facilitate muscular movement IV. Catalysis V. Immunity VI. Regulation of Gene Expression Why focus on myoglobin and hemoglobin? o Oxygen transport is critical for life o Hemoglobin mutation can lead to disease o Characteristic of oxygen binding to myoglobin and hemoglobin are highly observed Myoglobin: transports oxygen throughout muscles Classical globular, tetrameric protein that lacks structure entirely Has an iron-containing porphyrin derivative known as heme which is a prosthetic group o Prosthetic group: an organic compound that allows a protein to carry out a function that a polypeptide cannot perform alone Heme is a porphyrin that chelates oxygen transpor Myoglobin transports oxygen via Iron (Fe) in heme Oxygen binding to myoglobin depends on oxygen concentration Y= 𝐵𝑜𝑢𝑛𝑑 𝑀𝑏 𝑇𝑜𝑡𝑎𝑙 𝑀𝑏 = [𝑀𝑏𝑂2] [𝑀𝑏]+[𝑀𝑏𝑂2] (fractional saturation) Kd (p50) = [O2] at which myoglobin is half saturated Kd pO2, partial pressure of O2 (in torr) The amount of O2 bound to myoglobin (Y) is a function of the oxygen concentration (pO2) and the affinity of Mb for O2 (K) O2 binding to the heme group of myoglobin exhibits a hyperbolic curve Myoglobin and hemoglobin are related by evolution Hemoglobin is a heterotetramer which have two alpha and two beta chains, called globin. o Globins are homologous proteins that evolved from a common evolutionary origin/ancestor through genetic mutation Hemoglobin and myoglobin have similar secondary and tertiary structures Have a heme group in a hydrophobic pocket Have His F8 that ligands the Fe2+ (Ferrous form) – reduced form that binds oxygen Have His E7 that forms hydrogen bonds with oxygen Primary structures not similar (~18% similarity in primary sequence) Oxygen binds cooperatively to Hemoglobin Myoglobin O2 binding = Hyperbola Hemoglobin O2 binding = Sigmoid * Sigmoidal data indicates cooperativity Hemoglobin has a lower affinity for O2 Hemoglobin binding is sigmoidal because at low [O2], hemoglobin is reluctant to bind to the first O2, but as the concentration (pO2) increases, O2 binding sharply increases Indicates that the four heme groups in Hb are not independent and communicate cooperatively Cooperative binding: binding of one molecule of O2 to hemoglobin causes a conformational change to hemoglobin which enhances the ability to bind more O2 molecules o Exhibits allosteric interactions o Has more than one binding site Key physiological function: hemoglobin’s low oxygen affinity and cooperative binding behavior In lungs, the pO2 is 100 torr – hemoglobin is 95% saturated with O2 In tissues, pO2 is 20-40 torr – hemoglobin is 55% saturation; Hb affinity decreases o Here, O2 released by hemoglobin is readily taken up by myoglobin in muscle cells, since myoglobin’s affinity for O2 is much higher at relatively low oxygen concentration. (from lecture) At 15 kPa (lungs) Hb 95% saturation - low ability to deliver/donate O2 at high [ ] At 5 kPa (tissues) Hb 75% saturation – high ability to donate O2 (low affinity) o Myoglobin highly saturated in muscle cells At 1-2 kPa (exercise) Hb 10% saturation – easier for Hb to deliver O2; myoglobin is source of O2 Erythropoietin (EPO) boosts red blood cell production Synthesized by kidneys; individuals with kidney disease are deficient in EPO and develop anemia o Treatment w recombinant EPO can increase red blood cell production Can be abused by endurance sport athletes (like Lance Armstrong….smh) Conformational shift explains myoglobin’s cooperative behavior Shift in conformation between oxy and deoxy states involves rotation of one unit o Oxygen binding decreases size of the central cavity There are two quaternary structures of hemoglobin: I. Deoxyhemoglobin (T, tense conformation): hemoglobin without any bound O2 Heme is dome shaped (nonplanar) in deoxygenated Hb; has five ligands Reluctant to bind the first O2 molecule because the T conformation is unfavorable for O2 binding II. Oxyhemoglobin (R, relaxed conformation): hemoglobin with bound O2 Has 6 ligands The following O2 molecules bind with higher affinity due to the R conformation, favorable for O2 binding. Allosteric proteins: proteins with multiple binding sites o Binding of a small molecule (ligand) to one site alters the ligand-binding affinity of other sites pH effects on Hemoglobin Affinity At higher pH – hemoglobin saturation is 75% At lower pH – hemoglobin saturation is 50% (decreased affinity) Bohr Effect: reduction of hemoglobin’s oxygen-binding affinity when the pH decreases Groups become more acidic and release H+ when O2 binds to protein In vivo, tissues release CO2 which enters RBC’s and is converted into bicarbonate (HCO3-) In lungs, high oxygen concentration promotes O2 binding to hemoglobin causing release of protons that combine with bicarbonate to reform CO2 as it’s breathed out Effect of 2,3 BPG on oxygen binding *BPG decreases hemoglobin’s O2 affinity Red blood cells use a mechanism with 2,3 Biphosphogycerate BPG binds in central cavity of a deoxygenated (T) hemoglobin Five negative charges in BPG interact with positively charged groups in deoxyhemoglobin Presence of BPG stabilizes the deoxy conformation of hemoglobin; would bind O2 too tight without Fetal hemoglobin has a higher O2 affinity than adult hemoglobin, which helps O2 transfer for maternal circulation across the placenta to the fetus. Chapter 6 How Enzymes Work What is an enzyme? There are three ways to increase the rate of a chemical reaction: 1. Increase the temperature (adding energy in the form of heat) 2. Increasing the concentrations of the reactants 3. Addition of a catalyst Living systems use enzymes Ribozymes are catalysts made of RNA (exception) Chymotrypsin: a digestive protein that is synthesized in the pancreas and secreted into the small intestine to break down dietary protein Hydrolysis of polypeptides take place in a cleft between the two domains near the side chains of three residues (His, Asp, Ser) called the active site. Catalyzes hydrolysis of peptide bonds at 190 per second o Rate enhancement of 108 to 1012. Acts under mild conditions Has a broad range of substrate specificity o Catalyzes hydrolysis of peptide, amide, and ester bonds following Phe, Trp, and Try. o P-nitrophenylacetate (an ester) is readily hydrolyzed by chymotrypsin. Enzymes have high reaction specificity. Most are highly specific for their reactants (substrates) and products. Function groups in the active site of an enzyme can distinguish its substrates. The activities of enzymes are regulated in order to respond to changing conditions of follow genetically determined developmental programs. Enzymes are usually named after the reaction they catalyzed All biochemical reactions involve the addition of some substance to another, or its removal, or the rearrangement of that substance. Enzyme Classification I. Oxidoreductases II. Transferases III. Hydrolases IV. Lyases V. Isomerases VI. Ligases = Oxidation-reduction reactions = Transfer of functional groups = Hydrolysis reactions = Group elimination to form double bonds = Isomerization reactions = Bond formation coupled with ATP hydrolysis Isozymes: multiple enzymes catalyzing the same reaction How do enzymes differ from nonbiological catalysts? Explain why an enzyme’s common name may not reveal its biological role. Chemistry of Catalysis In a biochemical reactions, reactants must come together and undergo electronic rearrangement to form products. Free energy of Activation (G++): energyrequiring step of the reaction/energy barrier Transition state: point of highest energy; intermediate between reactants and products. Process of breaking old bonds and forming new bonds. The height of the activation energy barrier determines the rate of reaction. The higher the barrier, the less likely and slower the reaction will occur. A catalyst provides a reaction pathway with a lower activation energy barrier A catalyst decreases the activation energy barrier (G++) by interacting with the reacting molecules so they are more likely to assume the transition state. Doesn’t alter the net free energy change; only provides a pathway from reactants to products through a transition state that has lower free energy. Overall: An enzyme lowers the height of the activation energy barrier (G++) by lowering the energy of the transition state. Enzymes use chemical catalytic mechanisms Cofactors: a small organic molecule/metal ion that is required for catalytic activity Coenzymes: cofactors that are organic molecules, derived from vitamins Prosthetic group: a tightly bound coenzyme that remains in the active site between reactions. There are three basic chemical catalytic mechanisms used by enzymes: I. Acid-Base Catalysis: a proton is transferred between the enzyme and substrate Acid Catalysis: donation of proton from catalyst – H+ transfer from acid by enzyme lowers free energy of transition state Amino acids in A-B Catalysis Base Catalysis: H+ is abstracted by a base to lower the free energy of the transition state II. Covalent Catalysis (Nucleophilic Catalysis): a covalent bond forms between the catalyst and the substrate during formation of the transition state An electron rich group (nucleophile) in the enzyme active site forms a covalent adduct with a substrate. Undergoes a two-part reaction process, so there is two energy barriers. Ser, Tyr, Cys, Lys, and His are often good covalent catalysts Ex. Catalysis of acetoacetate to acetone via a primary amine (RNH2) to form an imine (Schiff base) III. Metal Ion Catalysis: A zinc ion stabilizes a developing negative on the transition state o Metal ions participate in enzymatic reactions by mediating oxidation-reduction reactions o Or by promoting the reactivity of other groups in the enzyme’s active site through electrostatic effects. A protein-bound metal ion can also interact directly with the reacting substrate. o Coordinates covalent bonds that hold the substrate in proper orientation The catalytic triad of chymotrypsin promotes peptide bond hydrolysis Chymotrypsin uses both acid-base catalysis and covalent catalysis. Dependent on three active site residues: His 57, Asp 102, Ser 195 – Catalytic Triad Identified using chemical labeling Chymotrypsin is a serine protease. Ser 195 is essential for catalysis. The substrates scissile bond (bond cleaved by hydrolysis) is near Ser 195 when substrate binds to the enzyme. Catalytic Mechanism of Chymotrypsin *just know examples* I. Peptide substrate enters chymotrypsin active site; scissile bond is close to oxygen of Ser 195 o Removal of Ser hydroxyl proton by His 57 (base catalyst) o Resulting nucleophilic oxygen (covalent catalyst) attacks carbonyl on substrate II. Tetrahedral intermediate decomposes His 57 is now an acid catalyst, donates proton to nitrogen of scissile bond, cleaving the bond Asp 102 stabilizes His 57 through H-bonding Energetically favorable because electron flow is easy III. Portion of protein is gone; other part is covalently attached to Ser 195 C-term of cleaved peptide leaves N-term of the substrate (acyl group) linked to enzyme Newly exposed N-terminus IV. Water enters active site – donates proton to His 57 His 57 is now a base catalyst Hydroxyl group attacks carbonyl of substrate V. Second tetrahedral intermediate (His 57 – now acid catalyst) Donates proton to Ser oxygen Causes intermediate to collapse VI. Enzyme is regenerated; rest of protein is released N-terminal portion of the original substrate has a new Cterminus and diffuses away Regenerates the enzyme Unique properties of Enzyme Catalyst Catalytic activity depends on transition state stabilization, proximity effects, induced fit, and electrostatic catalysis Enzymes stabilize the transition state by binding tightly to the transition state. Noncovalent interactions form and release free energy, lowering G++ Lock and Key Model Emil Fisher: catalytic residues must be precisely aligned in the active site Linus Pauling: an enzyme increases the reaction rate by binding tightly to the reaction state (not to substrates) Substrates strained towards structures of products Stabilization (tight binding) of transition state occurs in addition to A-B, covalent, or metal ion catalysis. Accomplished through close complementarity in shape and charge between active site and transition state Induced Fit Model (Daniel Koshland) When binding to substrates, enzymes undergo pronounced conformational changes to almost fully enclose substrates. Transition State Model Not only looks at the active site that recognizes and binds the substrate but the substrate to active it and go through the transition state The active site orients the substrate in a way to activate toward reaction When the substrate is held in the active site, it takes characteristics of the transition state orients The catalyst enhances the formation and stabilization of the highly energetic transition state. Proximity and Orientation Effects Enzymes increase reaction rates by bringing reactive groups into close proximity to increase frequency of collisions When substrates bind to enzyme, they are brought into correct orientation by freezing out the translational and rotational motions Induced Fit Ex. The binding of hexokinase to glucose causes a conformational change in the enzyme to fit the substrate better. Electrostatic catalysis : Non-aqueous active site allows more powerful electrostatic interactions between the enzyme and substrate Water molecules are sequestered in the active site, allowing an enzyme to eliminate the energy barrier imposed by the solvent molecules. (Reactions proceed quickly when no solvent molecules interfere) Not all serine proteases are related by evolution The three active sites are identical; proteins diverged from common ancestor and retained overall structure and catalytic mechanism Chymotrypsin, trypsin, and elastase have very similar structures but different substrate specificity Specificity due to the specificity pocket: cavity on the enzyme surface at the active site that accommodates residue on the N-terminus side of scissile bond Chymotrypsin is activated by proteolysis Proteases are limited by the action of protease inhibitors Zymogens (inactive precursors) are synthesized by proteases that can be activated later when needed Chymotrypsinogen: inactive precursor of chymotrypsin Synthesized by the pancreas; along with zymogens of trypsin (trypsinogen), elastase (proelastase) They are all activated via proteolysis after secreted into the small intestine Autoactivation: (example) trypsin activation causes activation of trypsinogen Reading Assignment: Blood Coagulation requires a cascade of proteases Loss of blood due to severe trauma can be halted by formation of clots (aggregated platelets) and a mesh of fibrin protein Conversion of fibrinogen into fibrin polymers is the last step in coagulation – series of proteolytic reactions Thrombin (enzyme) cleaves fibrinogen to fibrin, specifically the peptide bonds following Arg residues o Circulates as an active precursor (zymogen) called prothrombin Factor Xa catalyzes specific hydrolysis of prothrombin to generate prothrombin through a series of activated reactions – Enzyme Cascade Coagulation reactions have an amplifying effect – each protease is a catalyst for the activation of another catalyst. Protease Inhibitor limit protease activity Inhibitors pose as protease substrates but are not completely hydrolyzed. Chapter 7: Enzyme Kinetics and Inhibition Enzyme Kinetics: fully describe enzyme activity by applying mathematical tools to quantify and enzyme’s catalytic power and its substrate affinity as well as its response to inhibitors The progress of any reaction is expressed as velocity (v). o The rate of disappearance of the substrate (S) or rate of appearance of product (P) Enzyme-Substrate (ES) Complex E + S ES E + P Has a hyperbolic curve that suggests that an enzyme physically combines with a substrate At low substrate concentration, the enzyme quickly converts all substrate to products o As more substrate is added, the enzyme becomes saturated with substrate Only for simple enzymes Rate equations describe chemical processes Unimolecular Reactions A B Involve only a single reactant v = k[A] k = sec-1 Is a first order reaction: rate depends on concentration of one reactant. Bimolecular (Second Order Reaction) A + B C Involve two reactions v = k[A][B] k = M-1s-1 Velocity of a second order reaction is proportional to the product of the two reactant concentrations Michaelis-Menten equation is a rate equation for an enzyme-catalyzed reaction Vmax: maximum reaction velocity; when [S] is very high, all the enzyme is in its ES form and approaches maximum activity o Vmax = k2[E]T Equation: 𝑣0 = 𝑉𝑚𝑎𝑥 [𝑆] 𝐾𝑚 +[𝑆] Km is the substrate concentration at which the reaction velocity is half-maximal o Indicates how efficiently an enzyme selects a substrate to convert to product. o Measure of an enzyme’s affinity for a substrate o Km = Substrate Affinity The catalytic constant describes how quickly an enzyme can act Catalytic Constant (kcat): rate constant of reaction when the enzyme is saturated with substrate o Also known as the turnover number: number of catalytic cycles that each active site undergoes per unit of time Number of substrate molecules transformed to product molecules by a single enzyme in a given period of time. Is a first-order rate constant; units = s-1 Kcat/Km indicates catalytic efficienc Enzyme effectiveness depends on how avidly in binds substrate and how rapidly it converts them to products At low concentrates of substrate ([S] < Km), o Very little Enzyme-Substrate complex (ES) forms and [E] = [E]T Kcat/Km (units = M-1s-1; second order reaction) o The value, more than either Km or Kcat alone, represents the enzyme’s overall ability to convert subsrate to product. An enzyme reaches catalytic perfection when its overall rate is diffusion controlled. o Diffusion-controlled limit: maximum rate at which two freely diffusing molecules can collide with each other in aqueous solution Km and Vmax are experimentally determined To meet assumptions of the Michaelis-Menten model, the concentration of the substrate must be greater than the concentration of the enzyme Lineweaver-Burk Plot: linear transformation of the velocity versus substrate curve 1 𝐾𝑚 1 1 =( ) + 𝑣0 𝑉𝑚𝑎𝑥 [𝑆] 𝑉𝑚𝑎𝑥 A plot of 1/v0 versus 1/[S] gives a straight line with Slope = Km/Vmax Y intercept on the 1/vo axis = 1/Vmax Not all enzymes fit the Michaelis-Menten Model 1. Multisubstrate Reactions Most bisubstrate reactions are redox reactions or transferase reactions X (oxidized) + Y (reduced) X (reduced) + Y (oxidized) Ex. Transferase reaction catalyzed by transketolase Transforms a 6C sugar and 3C sugar to a 4C sugar and 5C sugar. Vmax is the max reaction velocity when both substrates are present at concentrations that saturate their binding sites on the enzyme. Random Mechanism: substrates in a bisubstrate reaction that bind in any order but end up in the active site at the ame time Ordered mechanism: enzymes which one substrate must bind before the other Ping-pong mechanism: one substrate binds and one product is released before the other substrate binds and the second product is released. 2. Multistep Reactions An enzyme-catalyzed reaction may contain many steps. The meaning of Kcat is the same for single-step reactions. Ex. Multistep transketolase reaction 3. Nonhyperbolic Reactions Oligomeric enzymes with multiple active sites do not obey Michaelis-Menten rate equations Allosteric Enzymes: presence of a substrate at one active site can affect the catalytic activity of other active sites Cooperative Behavior: occurs when enzyme subunits are structurally inked to each other so that a substrateinduced conformational change in one subunit causes conformational change. Allosteric/Regulatory Enzymes: composed of two or more sub-units that undergo conformational change upon binding of a ligand For both catalysis AND regulation display sigmoidal curve Exhibit cooperativity Binding at one active site can affect the catalytic activity of other site Enzyme Inhibition 2 classes of inhibitors: irreversible and reversible Irreversible inhibitor: “Suicide Inhibitor” forms covalent bond or very strong non-covalent interaction with the enzyme. Enter the enzyme’s active site and begins a reaction, but cause an incomplete reaction and get stuck in the active site. Reversible Inhibition: results when a substance binds reversibly (noncovalently) to an enzyme to alter its catalytic properties Can easily combine with and dissociate from the enzyme and render it inactive E+I Can effect an enzyme’s Km, Kcat, or both. There are 3 common types I. Competitive II. Noncompetitive III. Uncompetitive EI Competitive inhibition: the inhibitor directly competes with a substrate for binding to the enzyme’s active site. INCREASE Km DOES NOT AFFECT Vmax Can happen simultaneously Transition State Analogs: compounds that mimic the transition state to take advantage of the active site; make better inhibition (competitive inhibition) Noncompetitive inhibitors: do not compete towards the active site; binds to a site on the enzyme other than the active site to elicit a conformational change that affects structure/chemical properties of the active site Affect Vmax Does NOT affect Km Mixed Inhibition: inhibitor binding to the enzyme alters its conformation in a way that affects both Vmax and Km Affects both Vmax and Km Less Affinity (Higher Kmax) Uncompetitive Inhibition: binds to the enzyme-substrate complex; inhibitor can bind after one substrate has bound to prevent the reaction from continuing and yielding product. Km decreases as Vmax decreases Chapter 11 – Carbohydrates What are carbohydrates? Carbohydrates (sugars or saccharides) have the generic formula (CH2O)n : n is greater or equal to 3. They are formed from CO2 and H2O. Roles include: o Energy in diet – fuel molecules; glucose and fructose (immediate use), starch and glycogen (chemical storage forms for future use) o Mediating intercellular communication – in combination w proteins and lipids on cell surfaces; informational markers for molecular recognition Cell-cell recognition and signal transduction in glycoproteins and glycolipids o Structural support (cell walls) – scaffolding for bacteria and plant cell wall; connective tissue; exoskeletons o Components of nucleic acids – structural role as ribose and deoxyribose; and are polar sites for catalytic processes by RNA ribozymes. Classification of carbohydrates by length Monosaccharides – are simple sugars Small polymers of sugars include: Disaccharides: two sugars bound together Trisaccharides: three sugars bound together Oligosaccharides: “several” sugars bound together Polysaccharides – large polymers of sugars Monosaccharides – simple sugars Glyceraldehyde (Aldose) and dihydroxyacetone (Ketose) are the simplest three carbon sugars. Monosaccharides can also be classified by the number of carbon atoms. Triose: 3 carbons Tetrose: four carbons Pentose: five carbons Etc. Most carbohydrates are chiral compounds Glucose is chiral glyceraldehyde has exhibited mirror symmetry (2) called enantiomers Left (L)/Right (D) Enantiomers D sugars: asymmetric C farthest from carbonyl group (OH on right) L sugars: chiral carbon farthest from the carbonyl group has (OH on the left) Epimers - carbohydrates that differ in configuration at one of these carbons L-amino acids can distinguish D and L prefixes Cyclization generate 𝛼 and 𝛽 anomers 𝛼 anomer- hydroxyl group lies in opposite side of the ring from the CH2OH group of the chiral carbon that determines (D) or (L) config. 𝛽 anomer- hydroxyl group points up due to hydroxyl group lies in same side of the ring from the CH2OH group of the chiral carbon that determines (D) or (L) config. Not interchangeable Enantiomers- mirror images of eachother Epimers- pair of stereoisomers Anomers in an aqueous solution freely interconvert between 𝛼 ↔ 𝛽* *unless hydroxyl group attached to the anomeric carbon is linked to another molecule Glucose gets chair conformation and is made of = 64% 𝛽 anomer + 36% 𝛼 anomer Hexoses and pentoses undergo cyclization DON’T form planar structures Monosaccharides can be derivatized in many different ways Anomeric carbon can undergo oxidation, reducing substances such as Cu(II) →Cu(I) Benedict’s reagent can distinguish reducing sugar (free monosaccharide) CH3OH + C6H12O6 (reducing sugar) → Non-reducing sugar Methyl group can end up 𝛼 or 𝛽 position Glycosidic bond- links the anomeric carbon to the other group Glycoside- sugar linked to another molecule Ribose − − − − 𝑟𝑖𝑏𝑜𝑛𝑢𝑐𝑙𝑒𝑜𝑡𝑖𝑑𝑒 𝑟𝑒𝑑𝑢𝑐𝑡𝑎𝑠𝑒 → 2’ Deoxyribose One metabolically essential carbohydrate-modifying reaction is the one catalyzed by ribonucleotide reductase (reduces 2’ –OH group of ribose to convert a ribonucleotide to a deoxyribonucleotide for DNA synthesis) 11-2 Polysaccharides monosaccharides can are hooked together linked by glycosidic bonds Only one anomeric carbon involved in the glycosidic bond (C1) each -OH group can participate in a condensation rxn which permits different bonding arrangements and allows for branching Glycans can be sequenced using mass spectrometry Fault: inability to distinguish isomers 3D structural normally analyzed through NMR Lactose and sucrose are the most common disaccharide Disaccharide- 2 monosaccharides linked by glycosidic bond Lactose ● major fuel source for newborn mammals ● broken down by 𝜷 − 𝒈𝒂𝒍𝒂𝒄𝒕𝒐𝒔𝒊𝒅𝒂𝒆 (𝒍𝒂𝒄𝒕𝒂𝒔𝒆) ● adults make little lactase, therefore inefficient in digestion disaccharide Galactose + Glucose → Lactose Sucrose Glucose + Fructose → Sucrose (𝛼 (1 → 2)) ❏ most abundant disaccharide ❏ major form of newly synthesized carbohydrates Starch and Glycogen are Fuel Storage Starch 𝛼 (1 → 4) 𝛼 (1 → 6) Amylose: a linear and helical polymer of glucose Amylopectin: a branched polymer of glucose Amylopectin 𝜶 (𝟏 → 𝟔) ● plants primary metabolic fuel ● highly branched, easy to add/remove glucose ● compact particle due to 𝛼 𝑙𝑖𝑛𝑘𝑒𝑑 𝑐ℎ𝑎𝑖𝑛𝑠 𝑐𝑢𝑟𝑣𝑒 𝑖𝑛𝑡𝑜 ℎ𝑒𝑙𝑖𝑐𝑒𝑠 Cellulose and Chitin Provide Structural Support Cellulose ● most abundant polysaccharide ● residues linked 𝛽 (1 → 4) ● extended fibers for rigidity and strength ex. cell walls 11-1 A Biochemistry Note: Cellulosic Biofuel wood + agricultural waste → biofuels Cellulose ● cellulose is never in pure form ● Synthesized by organisms that produce cellulases (capable of hydrolyzing 𝛽 (1 → 4)bonds ● cellulose normally consist of hemicellulose, pectin, lignin. biofuel ex. ethanol Hemicellulose approx. 500 - 3,000 residues ● heteropolymer (most abundant being xylose) ● forms a network filled with pectin Pectin ● highly hydrophilic due to -OH groups ● holds H2O ● gel-like characteristics Lignin NOT a polysaccharide ● heterogeneous ● aromatic compound lead to difficulty in characterization ● hydrophobic, few -OH groups present ● covalently links to hemicellulose ex. strength in cell walls Bioconversion All components in wood is a large amount of stored energy into other fuels ● separate the polysaccharide from lignin ● dependent on living organisms or where enzymes were derived from then ● carbohydrate polymers are accessible to hydrolytic enzymes (resulting to mix of monosaccharides) Glucose + Fungi ( Yeast ) → ethanol Through fermentation Bioengineering organism to carry out polysaccharide hydrolysis and convert those monosaccharides to ethanol Ethanol ● stable ● storable ● transportable Chitin 𝛽 (1 → 4) 𝑙𝑖𝑛𝑘𝑒𝑑 𝑟𝑒𝑠𝑖𝑑𝑢𝑒𝑠 𝑎𝑟𝑒 𝑔𝑙𝑢𝑐𝑜𝑠𝑒 𝑑𝑒𝑟𝑖𝑣𝑎𝑡𝑖𝑣𝑒𝑠 ex. N-acetylglucosamine Bacterial Polysaccharides from a biofilm Prokaryotes ● do not synthesize cellulosic cell walls ● store fuel as starch or glycogen ● produce extracellular polysaccharides (acts as protective matrix for growth) Biofilm (an extracellular hydrated polysaccharide) ● attach to surface ● protects from desiccation or from being washed ● harbors community of embedded bacteria Difficult to characterize biofilm extracellular composition ❖ houses mix of species ❖ proportion of polysaccharide composition depends on many environmental factors 11-3 Glycoproteins N-linked oligosaccharides ● undergo processing attached to glycoproteins linked to an Asn side chain ● undergo processing- N-glycosylation begins while a protein is being synthesized by a ribosome associated with the rough ER. When the newly synthesized protein leaves the ER and traverses the Golgi apparatus, glycosidases and glycosyltransferases act on it protein is being synthesized by a ribosome associated with rough ER as translocated into ER lumen oligosaccharide chain of 14 residues is attached to an Asn residue (N-linked) leaves ER → Golgi Apparatus → glycosidases or glycosyltransferases glycosidases- remove various monosaccharide residues glycosyltransferases- add new monosaccharide residues O-linked oligosaccharides ● built in Golgi Apparatus one glycosyltransferases ● tend to be large ● do NOT go undergo processing by glycosidases ● major component of mucus provides protective layer for respiratory and digestive tracts ● undergo processing attached to glycoproteins linked to Ser or Thr Purpose of Oligosaccharide Groups occupy the protein surface ● hydrophilic ● conformationally flexible ● stabilize protein structure ● intracellular addressing system (deliver newly synthesized proteins to proper cellular location) ● recognition and attachment points (ex. blood type) The ABO Blood Group System Best known- ABO Blood Group System oligosaccharides attached to sphingolipids and proteins on cells ● 15 different carbohydrate configurations on blood group systems Type A oligosaccharides has terminal N-acetylated galactose group ● develop antibodies recognize and crosslink red blood cells bearing Type-B oligo. ● can receive Type-O Type B oligosaccharides has terminal galactose group ● develop antibodies recognize and crosslink red blood cells bearing Type-A oligo. ● can receive Type-O Type O oligosaccharides has none of these terminal groups (mutation- lacking final residue) ● develop antibodies recognize and crosslink red blood cells bearing Type-A or Type-B oligo. ● if A, B, or AB blood fusion is done antibodies will react with transfused cells causing lysis, clumping, or blocked vessels AB ● DO NOT develop antibodies recognize and crosslink red blood cells bearing Type-A or Type-B oligo. Proteoglycans contain long glycosaminoglycan chains Proteoglycans- protein chain is an attachment site for enormous O-linked polysaccharide called glycosaminoglycans Glycosaminoglycans repeating disaccharides amino sugar N-acetylated and uronic acid (sugar w/ carboxylate group) repeating glycosaminoglycans = chondroitin sulfate Roles: ● in variably on the extracellular side of the plasma membrane ● attract water (occupy the spaces between the cells and extracellular matrix ex. collagen fibrils) Proteoglycans-transmembrane proteins lipid linked extracellular proteoglycan and glycosaminoglycan chains not connected to protein scaffold play a role in connective tissue Mechanical Pressure Squeezes H2O from glycosaminoglycans which allow accommodation to body movements ● brings negative charged sulfate and carboxylate group of polysaccharide close together ● “shock absorption qualities” spring back repulsion between anionic group is relieved and water drawn back into the molecule Bacterial cell-walls- Peptidoglycan Peptidoglycan- network of crosslinked carbohydrate chains and peptides (make bacteria cell wall) ● gives shape ( surrounds plasma membrane) ● repeating 𝛽 (1 → 4) linked disaccharides ● antibiotics ex. penicillin family block formation of peptide cross-links (killing bacteria) Staphylococcus aureus ➢ glycans chains are perpendicular to cell surface ➢ honeycomb like Box 11-C Antibiotics ● inhibits synthesis of DNA, RNA, or proteins Common role: disruption of cell wall synthesis Penicillin (𝛽 − 𝑙𝑎𝑐𝑡𝑎𝑚 𝑎𝑛𝑡𝑖𝑏𝑖𝑜𝑡𝑖𝑐) ● first antibiotic used clinically ● inhibits enzyme that cross-link peptidoglycan, this weakening cell wall through disruption of osmotic pressure MRSA- Bacterias resistant to 𝛽 − 𝑙𝑎𝑐𝑡𝑎𝑚 produce enzyme that cleaves amide bond of 𝛽 − 𝑙𝑎𝑐𝑡𝑎𝑚 𝑟𝑖𝑛𝑔 Chapter 12 – Metabolism and Bioenergetics ATP – the cell’s energy and is a nucleotide whose reactions have a vital role in metabolism. Catabolism: break down of molecules to release free energy Fats, carbohydrates, and proteins are broken down to CO2, H2O, and NH3 Production of ATP, NADH, FADH2, and NADPH Anabolism: building up of molecules by using the free energy released from catabolism Construction of large complex biomolecules from smaller precursor molecules Energy is supplied via ATP, NADH, or NADPH Food and Fuel Humans are heterotrophs, therefore the food digested and absorbed is a source of metabolic energy and material for growth and activity. Cells take up products of digestion. The human diet consists of four biomolecules, where digestion reduces these to smaller molecules: Proteins Amino Acids Nucleic Acids Nucleotides Polysaccharides (Carbs) Monosaccharides Fats (Trialglycerols) Fatty Acids *not technically polymers Cells take up products of digestion Digestion is extracellular, in the mouth, stomach and small intestine and catalyzed by hydrolytic enzymes. Amylase: hydrolyzes and breaks down starch; consists of linear polymers of amylose and branch polymers called amylopectin. Protease (Trypsin, chymotrypsin, elastase): secreted in the stomach and pancreas which degrade proteins into small peptides and amino acids. Lipase: synthesized in the pancreas and secreted into the small intestine; catalyze the release of fatty acids from triacylglycerols. Monomers are stored as polymers Triacylglycerols (Large globules) are the polymer form of fatty acids in adipocytes. Long-term storage of nutrients. Liver and Muscle tissue use monosaccharides to synthesize glycogen (the storage polymer of glucose) Short-term carbohydrates, lasts about 12 hours as a source of energy. There is high concentration in liver and muscles. Glycogen is highly branched, but compact. Therefore, several glycogen molecules can clump to form granules. Branching allows for quick addition or removal of glucose residues. Biological implication of glycogen branching: Each branch in a glycogen molecule has a non-reducing end that allows faster mobilization of glucose when it is in metabolic demand. In conditions of high glucose concentration, it is more efficient to build glycogen due to the multiple non-reducing ends for expand/building of glycogen for storage. Amino acids can be used to build peptides Because proteins are not dedicated storage molecules for amino acids, excess amino acids cannot be saved for later. Instead, excess amino acids are broken down and are converted into carbohydrates (stored as glycogen) or converted into acetyl units (Acetyl CoA) for conversion into fats. During starvation, proteins are catabolized to supply the body’s energy needs. Amino acids and glucose are required for nucleotide synthesis. Asp, Gln, and Gly supply carbon and nitrogen atoms to build bases. Fuels are mobilized as needed Metabolic fuels are molecules that are broken down by processes that make free energy available for cell activity. These fuels include: amino acids, monosaccharides and fatty acids. When fuel molecule supplies are exhausted, the body must mobilize its stored resources by converting its polysaccharide and triacylglycerol storage molecules (sometimes protein) to their monomeric units. Most body tissues, and primarily the CNS, prefer to use glucose as the primary metabolic fuel. Therefore, the liver mobilizes glucose by breaking down glycogen. Glycogen breakdown occurs via phosphorolysis (degradation of glycogen) Phosphate breaks the bonds between glucose residues and is catalyzed by glycogen phosphorylase, releasing residues from the ends of a glycogen polymer. The phosphate from Glucose-1-phosphate is removed before glucose is released from the liver into circulation. When glucose runs low, adipose tissue begins to mobilize its fat stores. Lipase hydrolyzes triacylglycerols in order for fatty acid release into the bloodstream. These fatty acids are not water-soluble and can bind to circulating proteins. The body cannot budget burning fatty acids however, the heart uses fatty acids as its primary fuel. Cellular proteins are continuously degraded therefore, there are two major mechanisms for protein degradation: I. Degradation via the Lysosome (an organelle that contains proteases and hydrolytic enzymes) o Breaks down proteins enclosed in a membranous vesicle. o Membrane proteins and extracellular proteins are taken up by endocytosis and are degraded II. Degradation via Proteasome (barrel-shaped structure where its inner chamber contains multiple active sites to carry out peptide bond hydrolysis) o Proteins can only enter the proteasome after is has been covalently tagged with ubiquitin. o Links C-terminus to a lys side chain (at least four ubiquitins) Metabolic Pathways Metabolic pathways can be considered as: a series of intermediates or metabolites; a set of enzymes that catalyze the reactions by which metabolites are interconverted; an energy-producing/requiring phenomenon; or a dynamic process that can be turned up or down. Major metabolic pathways share few common intermediates A handful of metabolites appear as precursors or products in pathways that lead to all other types of biomolecules. Glycolysis: the pathway that degrades glucose (6-carbon sugar) which is phosphorylated and split in half, yielding two molecules of glyceraldehyde-3phosphate. This is converted into pyruvate (3-carbon molecule), which is decarboxylated yielding acetyl-CoA (two carbon acetyl groups are linked to coenzyme A, CoA) Glyceraldehyde-3-phosphate, pyruvate, and acetyl-CoA are the key players in metabolism. Fatty acid breakdown yields acetyl-CoA Pyruvate can undergo amino-group transfer to yield alanine. Metabolic pathways include oxidation-reduction reactions. Catabolism of amino acids, monosaccharides, and fatty acids = oxidation of carbon atoms Anabolism of amino acids, monosaccharides, and fatty acids = reduction of carbon atoms Carbon in methane is highly reduced; Carbon in CO2 is highly oxidized. Fatty acids have Methylene carbons and Carbohydrates (CH2O) have carbons that undergo oxidation. Oxidation of metabolic fuels causes transfer of electrons to enzyme cofactors/coenzymes - nicotinamide adenine dinucleotide (NAD+) or nicotinamide adenine dinucleotide phosphate (NADP+) NAD+ = catabolic reactions NADP+ = anabolic reactions Ubiquinone: a lipid-soluble electron carrier in which a membrane-associated enzyme transfers electrons from a substrate to the electron carrier. Ubiquinone can take up one OR two electrons (NAD+ is strictly a two-electron carrier) Reduced ubiquinol can diffuse through membrane and donate its electrons. The citric acid cycle generates considerable amounts of reduced cofactors that are recycled through oxidative phosphorylation. Reoxidation of NADH and QH2 and production of ATP requires reduction of O2 and H2O Basically: NAD+ and ubiquinone collect electrons and free energy from reduced fuel molecules and are ultimately transferred to O2; this free energy is in the form of ATP. Overview of metabolism: Monomers are formed. Intermediates with two or three carbons are formed. Carbons are fully oxidized to CO2. Electron carriers gain electrons and are recycled via electron loss. ATP and H2O are produced. (from book) Metabolic pathways are all connected Pathway activity is regulated Not every cell carries out every pathway Each cell has a unique metabolic repertoire Organisms may be metabolically interdependent. Humans metabolism depends on vitamins Vitamins are the building blocks for coenzymes, which are needed by enzymes to complete reactivity. Niacin (Vitamin B3) – precursor for Nicotinamide Adenine Dinucleotide (NAD+) Riboflavin (Vitamin B2) – precursor for FAD (in oxidation reduction) Vitamin C – Precursor for hydroxylation reactions Fat Soluble Vitamins: A, D, E, K (Cannot have in excess) Vitamin C: a cofactor for prolyl hydroxylase and lysyl hydroxylase, which are responsible for hydroxylation of proline and lysine amino acids in collagen. Hydroxyproline and hydroxylysine stabilize collagen by cross-linking the propeptides in collagen. Deficiency (Scurvy): causes fragile capillary walls, easy bleeding of gums, loosened teeth, and bone-joint diseases Chapter 13 Glucose Metabolism Biochemical Roles of Glucose Source of metabolic energy Precursor for the synthesis of structural polysaccharides (cellulose), disaccharides (sucrose and lactose) and monosaccharides (galactose and fructose) All cells of the body utilize glucose for fuel Erythrocytes depends solely on glucose Brain cells are dependent on a constant supply of glucose. Glycolysis: ten-step conversion of 6-carbon glucose to a 3-carbon pyruvate. Role: universal pathway for extraction of energy from carbohydrates o Aerobic organisms: initial phase to prepare glucose for additional energy production o Anaerobic organisms: ATP and NADH is the only significant energy available from carb metabolism Does NOT require molecular oxygen Occurs in the cytoplasm Hexose (6C Glucose) is split into 2 molecules of -ketoacid pyruvate (3C) o Production of a small amount of energy o Net: 2 ATP, 2 NADH Glucose + 2 NAD+ + 2 ADP + 2 Pi 2Pyruvate + 2NADH + 2ATP Phosphorylation of two molecules of ADP to produce 2 ATP Reduction of two molecules of NAD Steps 1-5: Energy Investement Steps 6-10: Energy Payoff Energy Investment Phase: Requires the investment of free energy in the form of two ATP molecules Step 1: Hexokinase Reaction Hexokinase transfers a phosphoryl group from ATP to the C6 OH group of glucose to form glucose-6phosphate Kinase: enzyme that phosphorylates molecules Irreversible reaction (Negative standard free energy change) o Prevents glucose from backing out of glycolysis ATP is invested; ATP hydrolysis drives the reaction Step 2: Phosphoglucose Isomerase Reaction Isomerization reaction; glucose-6 phosphate is converted into fructose-6-phosphate Conversion of a hexose to a pentose Reaction is near-equilibrium; freely reversible Step 3: Phosphofructokinase Reaction Consumes 2nd ATP molecule via phosphorylation of fructose-6-phosphate to yield fructose 1,6-bisphosphate Primary Control/Regulation point of glycolysis Irreversible Reaction Slowest reaction in glycolysis Phosphofructokinase response to allosteric effectors ADP binds to the enzyme, causing a conformational change that promotes F-6-P binding. o ADP concentration in the cell is a good indicator for the need of ATP. Phosphoenolpyruvate (a product of step 9) binds to PFK, causing a conformation that destabilizes F-6-P binding, inhibiting catalytic activity. o PFK acts as a feedback inhibitor to slow the pathway by decreasing the rate of reaction when there is plenty of PEP and ATP. Fructose-2,6-bisphosphate: most potent activator of phosphofructokinase in mammals. Synthesized from F-6-P by phosphofructokinase-2 Phosphofructokinase-2 is hormonally stimulated when there is high glucose concentration in the blood. The increase in F-2,6-bP activates PFK to increase the flux of glucose in glycolysis. PFK reaction is rate-determining reaction, which operates far from equilibrium and has a large negative free change of energy and is irreversible under metabolic conditions. Rate of reaction is altered by allosteric effectors, but NOT by fluctuations of concentration of substrates and products. Step 4: Aldolase Converts fructose-1,6-bisphosphate to two three carbon molecules, Dihydroxyacetone phosphate and glyceraldehyde-3-phosphate. Reverse of an Aldol condensation reaction catalyzed by aldolase. Two catalytically important residues: Lys Residue – forms Schiff base (imine) with substrate Ionized Tyr residue – acts as a base catalyst Products of the aldolase reaction are both phosphorylated, but only glyceraldehyde-3-phosphate proceeds through the pathway Step 5: Triose Phosphate Isomerase Dihydroxyacetone phosphate is converted into glyceraldehyde-3-phosphate by triose phosphate isomerase. 2 molecules of glyceraldehyde-2-phosphate proceed through glycolysis Triose phosphate isomerase is a “catalytically perfect” enzyme – rate is limited only by the rate at which its substrates can diffuse to its active site. Catalytic mechanism involves low-barrier hydrogen bonds Catalytic power of triose phosphate isomerase depends on protein loop that closes over active site Energy payoff phase: 4 ATP molecules are produced; net gain of 2 ATP; involve three-carbon intermediates Step 6: Glyceraldehyde-3-phosphate Dehydrogenase Glyceraldehyde-3-phosphate is both oxidized and phosphorylated into 1,3bisphosphoglycerate. Addition of inorganic phosphate to the substrate (rather than phosphorylation via ATP) – group transfer reaction. Oxidation-Reduction reaction o NAD+ is reduced to NADH o Aldehyde group of GA-3-P is oxidized Glyceraldehyde-3-phosphate dehydrogenase catalyzes the removal of an Hatom. This reaction is inhibited by arsenate (AsO43-), which competes with Pi (PO43-) for the Cys residue in the active-site. Step 7: Phosphoglycerate Kinase 1,3-Bisphosphoglycerate donates a phosphoryl group to ADP to produce 3-phosphoglycerate and ATP catalyzed by phosphoglycerate kinase. Reaction occurs twice: 2 ATP have been recouped Kinase transfers phosphoryl group Substrate-Level Phosphorylation Step 8: Phosphoglycerate mutase 3-phosphoglycerate is converted to 2-phosphoglycerate. Isomerization of 3-PG to 2-PH requires an enzyme active site that contains a phosphorylated His residue. o The phosphor-His transfers phosphoryl group to 3-phosphoglycerate to generate 2,3bisphosphoglycerate, which gives a phosphoryl group back to the enzyme leaving 2phosphoglycerate and phospho-His Step 9: Enolase 2-Phosphoglycerate is converted to phosphoenolpyruvate and H2O via dehydration reaction Enzyme active site includes an Mg2+ ion that coordinates with the OH group at C3, making a better leaving group. Fluoride ion and Pi forms a complex with Mg2+, which can inhibit the enzyme. Step 10: Pyruvate Kinase Converts phosphoenolpyruvate to pyruvate and transfers a phosphoryl group to ADP to produce ATP. Reaction occurs in two parts: 1. ADP attacks phosphoryl group of PEP to form ATP and enolpyruvate 2. Tautomerization of enolpyruvate to pyruvate; highly exergonic Substrate-Level Phosphorylation Substrate-level phosphorylation vs. Oxidative Phosphorylation Substrate-level: process of forming ATP by phosphoryl transfer from a reactive intermediate Oxidative: process for making ATP from ADP by electron transfer linked to respiration. Regulatory enzymes of Glycolysis I. II. III. Hexokinase: catalyzes irreversible reaction and subject to inhibition by glucose-6-phosphate (product) Phosphofructokinase a. Inhibition via phosphoenolpyruvate and activation via ADP – Bacteria b. Activation via Fructose-2,6-bisphosphate Pyruvate Kinase Feed-forward Activation Fructose-1,6-bisphosphate activates pyruvate kinase at an allosteric site. Pyruvate Metabolism Pyruvate can be further broken down to either: Acetyl-CoA Used to synthesize compounds such as oxaloacetate Pyruvate can metabolize depending on the organism, type of cell, and intracellular conditions of the cell. In oxygen-plentiful conditions: pyruvate is completely oxidized to CO2 and electrons are ultimately transferred to O2 In oxygen-limited conditions: other molecules serve as electron acceptors to regenerate NAD + Fermentation: extraction of energy from carbohydrates and other organic substrates without using oxygen as an electron acceptor a. Lactate Fermentation: anaerobic microorganism and in muscle cells under limiting oxygen conditions b. Ethanol fermentation: yeast and other microogranisms. Yeast Fermentation Sugars are transformed to pyruvate via glycolysis Then, pyruvate decarboxylase removes carboxylate group on pyruvate to produce acetaldehyde Alcohol dehydrogenase then reduces acetaldehyde to ethanol. Alcohol metabolism The liver can metabolize ethanol, which is readily absorbed in the GI tract and transported by the blood stream. Alcohol dehydrogenase converts ethanol to acetaldehyde Then, acetaldehyde is converted to acetate via acetaldehyde hydrogenase. Both require NAD+ as a cofactors. Ethanol is considered mildly toxic. Toxicity of acetaldehyde and acetate in liver and brain tissue. Ethanol can induce vasodilation – flushing; warming and reddening of skin due to increased blood flow Heart and respiration rate is decreased Kidneys increase excretion of water – ethanol interferes with ability of hypothalamus to sense osmotic pressue Stimulation of signaling from neurotransmitters that function as ligand-gated ion channels – inhibits neuronal signaling. Hangover is a result of production of acetaldehyde and acetate. Production results in NAD+ consumption – lowers NAD+:NADH ratio Insufficient NAD+ diminished ATP production via glycolysis (NAD+ required for glyceraldehyde-3phosphate dehydrogenase) Shortage of liver NAD+ slows fatty acid breakdown and promotes fatty acid synthesis Further catabolism of pyruvate is initiated via decarboxylation to form a two-carbon acetyl group linked with coenzyme A Acetyl-CoA is a substrate for the citric acid cycle. Pyruvate is a precursor for several molecules. The two-carbon unit Acetyl-CoA (derived from pyruvate) can be used for fatty acid synthesis. Pyruvate is also a precursor for oxaloacetate – an intermediate in Amino Acid synthesis, gluconeogenesis, and the TCA cycle o Oxaloacetate is synthesized by pyruvate carboxylase o Pyruvate carboxylase has a biotin prosthetic group (carries CO2) – covalently linked to a Lys residue Gluconeogenesis The liver can synthesize glucose from non-carbohydrate precursors via gluconeogenesis. Kidneys can perform gluconeogenesis (limited) when glycogen supply in liver is exhausted. It is the reverse of glycolysis: conversion of two molecules of pyruvate to one molecule of glucose. Gluconeogenesis bypasses pyruvate kinase, phosphofructokinase, and hexokinase. Instead four enzymes are used in this pathway: o Hexokinase (in glycolysis) Glucose-6-Phosphatase (in gluconeogenesis) o Phosphofructokinase (in glycolysis) Fructose bisphosphatase o Pyruvate kinase (in glycolysis) Phosphoenolpyruvate carboxykinase & Pyruvate carboxylase Pyruvate cannot be directly converted to phosphoenolpyruvate (pyruvate kinase catalyzes irreversible reaction – step 10 in glycolysis) 1. Pyruvate is first carboxylated by pyruvate carboxylase to yield oxaloacetate (this utilizes ATP. 2. Oxaloacetate is then decarboxylated via phosphoenolpyruvate carboxykinase to form phosphoenolpyruvate. Pyruvate carboxylase consumes ATP; phosphoenolpyruvate carboxykinase consumes GTP Cleavage of phosphoanhydride bonds is required to supply free energy to convert. Alanine and Glutamine) are the main sources of precursors for gluconeogenesis because they can be converted to oxaloacetate and then to phosphoenolpyruvate. During starvation, proteins are broken down and are used to produce glucose to fuel the CNS. Two molecules of phosphoenolpyruvate are converted to one molecule of fructose-1,6-bisphosphate in a series of six reversible reactions. The final three reactions of gluconeogenesis require two enzymes. 1. First step undoes phosphofructokinase – fructose bisphosphatase hydrolyzes C1 phosphate of fructose1,6-bisphosphate to yield fructose-6-phosphate 2. Phosphoglucose isomerase produces glucose-6-phosphate (reverse of step 2) 3. Glucose- 6-phosphatase catalyzes hydrolytic reaction that yields glucose and inorganic phosphate (Pi) Regulation of gluconeogenesis – Fructose Bisphophosphatase Step Simultaneous occurrence of glycolysis and gluconeogenesis is energy wasting – there is regulation of opposing pathways. Combination of these metabolic reactions is a futile cycle (have no useful result) – provides a mean of finetuning the output of a metabolic pathway. Major regulatory point: interconversion of fructose-6-phosphate and fructose-1,6-bisphosphate When concentration of fructose-2,6-bisphoshphate is high: Glycolysis is stimulated Gluconeogenesis is inhibited. Fructose-2,6-bisphosphatase modulates activity of both fructokinase and fructose bisphosphatase – as activity of one enzyme increases, the other decreases – dual regulatory effect. Glycogen Synthesis and Degradation Glucose produced by gluconeogenesis is stored in the liver and other tissues as glycogen; glucose units can be removed from glycogen via phosphorolysis. Glycogen degradation is thermodynamically spontaneous. Glycogen synthesis requires input of free energy. Glycogen synthesis consumes free energy of UTP Glycogen is composed of monomers of glucose-1-phosphate, which is isomerized by phosphoglucomutase Glucose-1-phosphate is activated by the reaction of UTP to form UDP-glucose This is a reversible phosphoanhydride exchange. Glycogen synthase: transfers a glucose unit to the C4-OH group at the end of a glycogen branch – extends the linear (14)-linked residue Glycogenolysis Glycogen is phosphorolyzed to yield glucose-1-phosphate. In liver – phosphoglucomutase converts glucose-1-phosphate to glucose-6-phosphate, then glucose-6phosphatase hydrolyzes the release of free glucose from glucose-6-phosphate Linear chains are broken down via phosphorolysis. Branched chains are broken down via hydrolysis Glucose-6- phosphate can enter glycolysis at step 2 – one less ATP is consumed = higher net ATP Pentose Phosphate Pathway (PPP): converts glucose-6-phosphate to ribose-5-phosphate Is an oxidative pathway Generates NADPH *Starting point = glucose-6-phosphate 1. Glucose-6-phosphate dehydrogenase catalyzes irreversible transfer of hydride ion from glucose-6-phosphate to NADP+ to form a 6phosphoglucono-lactone and NADPH Deficiency in G6P-D causes decreased production of NADPH, interfering with normal oxidation-reduction processes – makes cells more susceptible to oxidative damage o More resistant to malaria 2. 6-phosphogluconolactonase hydrolyzes the lactone intermediate to 6-phosphogluconate 3. 6-phosphogluconate dehydrogenase oxidatively decarboxylates 6phosphogluconate to ribulose-5-phosphate Two molecules of NADP are produced 4. Ribulose-5-phosphate isomerase isomerizes ribulose-5-phosphate to ribose-5-phosphate Ribose-5-phosphate is a precursor for the ribose unit in nucleotides The PPP produces ribose and NADPH (reducing agent) for the reduction of ribose to deoxyribose via ribonucleotide reductase. The need for NADPH can be greater in other reactions – excess carbons are recycled into intermediates for glycolysis for degradation into pyruvate or for use in gluconeogenesis. Enzymes such as transketolase and transaldolase – transfer 2/3C units among intermediates to produce three to seven carbon sugars. Ex. Transform 5C-Ribulose into 6C-units (fructose-6-phosphate) and 3C-unites (glyceraldehyde-3phosphate) Chapter 14 – The Citric Acid Cycle Beri-Beri Disease: Nutritional disease caused by the deficiency of thiamine (Vitamin B1) The activated form of B1 is thiamine pyrophosphate (TPP), which is a coenzyme that assists enzymes involved in decarboxylation. Vitamin B1 (Thiamine) is a precursor for thiamine pyrophosphate, a cofactor used by the pyruvate dehydrogenase complex (PDH) It is also a co-factor for -ketoglutarate dehydrogenase And is also found in a variety of foods such as meat and grains. Decarboxylation reactions occur as pyruvate is further oxidized in the TCA, therefore TPP-B1/Thiamine is a required cofactor for decarboxylase. 3C pyruvate to a 2C acetyl with loss of CO2 2C acetyl + 4C Oxaloacetate = 6C 6C to 5C with loss of CO2 5C to 4C with loss of CO2 Preliminary Phase Bridge between glycolysis and aerobic metabolism Central and Universal pathway of aerobic metabolism The citric acid is central metabolic pathway whose starting material is 2C-acetyl units derived from aminoacids, monosaccharides, and fatty acids. These are oxidized to waste product CO2, with the reduction of cofactors: NAD+ and ubiquinone (Q) Pyruvate is the starting point of the citric acid cycle (end product of glycolysis) Mitrochondria: the powerhouse of the cell Has an inner matrix, where the electron transport chain is located Has an intermembrane space The citric acid cycle takes place in the mitochondria of eukaryotes The Pyruvate Dehydrogenase Complex Reaction : A multi-enzyme complex used to make chemical transformations economical, efficient, rapid, and prevents unwanted side reactions – catalyzes the decarboxylation of pyruvate, which is oxidized into 3 CO2 (1st CO2 is released when pyruvate is decarboxylated to an acetyl unit; the 2nd and 3rd are products of the citric acid cycle) The PDH complex is a metabolon It is located in the mitochondria Pyruvate translocase: is a permease that functions by exchanging pyruvate and hydroxide ions, in order to balance the electrical charge on each side of the membrane The pyruvate dehydrogenase complex directs conversion of pyruvate to acetyl CoA – this is the bridge between glycolysis and aerobic metabolism Pyruvate (3C) Acetyl CoA (2C) + NADH + CO2 Cytosol Mitochondria The oxidation of pyruvate requires 3 enzymes (E1, E2, E3), 5 coenymes, and 5 distinct reactions. The chemical transformation of pyruvate involves: Decarboxylation (Loss of CO2) Oxidation of the keto-group of C2 to a carboxyl group Activation by linkage through a thioester bond to coenzyme A NAD+ NADH + H+ PDC Pyruvate CO2 CoenzymeA Acetyl CoA The Pyruvate Dehydrogenase Complex happen in 5 Steps (*don’t have to know, FYI only) 1. E1 decarboxylates pyruvate; CO2 is released. 2. A leftover hydroxyethyl group is transferred to E2. 3. An acetyl group is transferred to CoA. 4. E2 is restored. 5. E3 is restored and NADH is produced. Coenzyme A (CoA) has Vitamin B5 – they help the body convert food (carbohydrates) into fuel (glucose), which is used to produce energy. These B vitamins are referred to as B complex vitamins, which also help the body use fats and proteins. The energy stored in the thioester bond drives the thermodynamically unfavorable reactions. Use of CoASH has an acetyl (TCA) and acyl carrier (fatty acid metabolism) o Fatty Acids + CoASH Fatty AcylCoA o Dehydrogenases is Metabolism via different Co-Enzymes If alcohol to aldehyde Alcohol Dehydrogenase NADH + Aldehyde If single bond to DB bond Dehydrogenase DB bond + FADH2 The Citric Acid Cycle (Eight Reactions) The citric acid cycle is a central and universal pathway of aerobic metabolism Also known as the TCA and Krebs cycle Occurs in the mitochondrial matrix It is considered amphibolic – it is both catabolic and anabolic Purposes: Degradation of acetyl-CoA (from pyruvate and other sources) to CO2 Supplying precursors for biosynthesis of amino acids, porphyrins, and purine and pyrimidine bases for nucleotides Production of energy-rich molecules: GTP, NADH, and QH2 Overview Acetyl-CoA is derived from pyruvate and is a product of amino acid catabolism (carbon skeletons of amino acids are broken down to pyruvate) Acetyl-CoA is a direct product of degradation of certain amino acids and of fatty acids Acetyl Co-A enters the TCA cycle for further oxidation. Process is highly exergonic and free energy is conserved via GTP and reduced cofactors. For each acetyl group, two molecules of fully oxidized CO2 are produced – loss of 4 pairs of electrons. These electrons are transferred to 3NAD+ and ubiquinone to produce 3 NADH and 1 QH2. Acetyl-CoA + GDP + Pi + 3 NAD+ + Q 2 CO2 + CoA + GTP + 3 NADH + QH2 The TCA is a high-energy-generating cycle 1 NADH = 2.5 ATP 1 QH2 = 1.5 ATP Reaction 1: Citrate Synthase Adds an acetyl group to oxaloacetate – the acetyl group of Acetyl-CoA condenses with 4C-Oxaloacetate to produce Citrate Citrate synthase is a dimer that undergoes conformational change upon binding of oxaloacetate, which allows the binding of Acetyl-CoA Can synthesize a C-C bond without a metal ion cofactor The first reaction intermediate is stabilized by formation of low-barrier hydrogen bonds The coenzyme A released can be reused by the PDH or be used in the TCA to synthesize succinyl-CoA. The free energy of the reaction is highly exergonic, citrate synthase has a free energy equivalent to breaking the thioester bond of Acetyl-CoA Reaction 2: Aconitase Catalyzes the reversible isomerization of citrate to isocitrate The reaction intermediate is Aconitate. Citrate is a symmetrical molecule – but only 2 carboxymethyl arms undergo dehydration and rehydration, they are no longer identical when bound to an asymmetrical enzyme Reaction 3: Isocitrate dehydrogenase Releases the first CO2 – is the oxidative carboxylation of isocitrate to -ketoglutarate The substrate is oxidized accompanied by the reduction of NAD+ to NADH The carboxylate group that is to the ketone function (meaning that is two carbons away from the ketone) is eliminated as CO2 o An Mn2+ ion in the active site helps stabilize the negative charges of the intermediates. The CO2 molecules generated (along with CO2 in the next reaction and CO2 produced by decarboxylation of pyruvate) are diffused out of the cell and carried in the bloodstream to the lungs. o Carbons are oxidized; NAD+ is reduced. Energy released from the cleavage of the thioester bond There is a push via a 3 alcohol group Reaction 4: -ketoglutarate dehydrogenase Releases the 2nd CO2 – catalyzes an oxidative decarboxylation reaction, transferring the remaining 4-carbon fragment to CoA -ketoglutarate dehydrogenase is a multienzyme complex that resembles the PDH complex in structure/mechanism; E3 is the same enzyme in both complexes Free energy of oxidation ketoglutarate is conserved in formation of the thioester succinyl-CoA Fates of carbons in the Citric Acid Cycle Two carbon atoms are lost as CO2 – catalyzed by isocitrate dehydrogenase (step 3) and -ketoglutarate dehydrogenase (step 4) are not the same carbons that entered the cycle as Acetyl-CoA. The acetyl carbons become part of oxaloacetate and are lost in the reactions Reaction 5: Succinyl-CoA synthetase Succinyl-CoA Also called succinate thiokinase Catalyzes substrate-level phosphorylation Cleaves the high-energy thioester bond of succinyl CoA Reaction is couple to phosphorylation of GDP to GTP Succinate Reaction 6: Succinate Dehydrogenase Succinate is oxidized to fumarate – reversible dehydrogenation The oxidation-reduction reaction requires an FAD prosthetic group, which is reduced to FADH2 Regeneration of the enzyme requires re-oxidation of FADH2. Enzyme is embedded into the inner mitochondrial membrane, therefore is reoxidized by the lipid-soluble electron carrier, ubiquinone (Q) – acquires 2 electrons to be QH2 (ubquinol) Reaction 7: Fumarase (Also fumarate hydratase) Catalyzes the reversible hydration of a double bond to convert fumarate to malate Reaction 8: Malate Dehydrogenase The last reaction is the regeneration of oxaloacetate from malate via an NAD+-dependent oxidation reaction Energy-generation of the Citric Acid Cycle The entire pathway catalytically disposes carbon atoms derived from amino acids, carbohydrates, and fatty acids. Each NADH yields approximately 2.5 ATP Each QH2 (of FADH) yields approximately 1.5 ATP Every acetyl unit that enters the TCA cycle can generate a total of 10 ATP equivalents. A muscle operating anaerobically produces only 2 ATP per glucose; a muscle operating aerobically produces 32 ATP equivalents per glucose – Pasteur effect (rate of glucose consumption by yeast decreased when cells were shifted from aerobic to anaerobic conditions) Summary of The Citric Acid Cycle 1. Acetate (C2) enters as Acetyl-CoA and 2 carbons leave the cycle as CO2 2. 3 NAD+ are reduced to NADH by dehydrogenase catalyzed reactions 3. 1 FAD is reduced to FADH2 Regulation of the Citric Acid Cycle at three irreversible steps The flux through the TCA is regulated at three metabolically irreversible steps 1. Citrate synthase reaction (Reaction 1) 2. Isocitrate dehydrogenase reaction (Reaction 3) 3. -ketoglutarate dehydrogenase (Reaction 4) Citrate synthase Inhibited by citrate Citrate also inhibits PFK (in glycolysis) which decreases the supply of Acetyl-CoA Inhibited by NADH Isocitrate dehydrogenase Inhibited by NADH Activated by ADP and Ca2+ -ketoglutarate dehydrogenase Inhibited by NADH and Succinyl-CoA Activated by Ca2+ The intermediates produced by the citric acid cycle are biosynthetic precursors for other molecules. Ex. -ketoglutarate can undergo reductive amination by glutamate dehydrogenase to produce glutamate Glutamate is also the precursor for glutamine, arginine and proline. Any of the TCA intermediates can be converted to oxaloacetate, which can serve as gluconeogenic precursors. Anaplerotic reactions replenish TCA intermediates Pyruvate + CO2 + ATP + H2O Oxaloacetate + ADP + Pi Citrate and pyruvate can cross the mitochondrial membrane via specific transport proteins This allows carbon atoms from mitochondrial Acetyl-CoA to be transferred to the cytosol for fatty-acid/cholesterol synthesis Intermediates diverted to other pathways can be replenished Oxaloacetate can be replenished by pyruvate and amino acids -ketoglutarate can be replenished by amino acids Succinyl-CoA can be replenished by amino acids and odd-chain fatty acids. o Acetyl-CoA activates pyruvate carboxylase so that more oxaloacetate is produced when there is low TCA activity and accumulation of Acetyl-CoA There is normally low oxaloacetate concentration because the malate dehydrogenase reaction is thermodynamically unfavorable o The replenished oxaloacetate is converted to citrate, isocitrate, -ketoglutarate, etc. The TCA cycle is a catalyst, therefore increasing concentrations of its components increases the flux through the pathway. Degradation of odd-numbered fatty-acids yields succinyl-CoA Degradation of amino acids can also produce some amino acids, -ketoglutarate, succinyl-CoA, fumarate and oxaloacetate – transaminations During vigorous exercise, the concentration of TCA intermediates increases 3 to 4-fold in a few minutes to help increase the energy-generating activity of the TCA cycle. There is also an increased activity of the TCA regulatory enzymes (citrate synthase, isocitrate dehydrogenase, and -ketoglutarate dehydrogenase) which increases the flux 100-fold Some glutamate can be converted to -ketoglutarate to boost activity of the TCA cycle Chapter 15 – Oxidative Phosphorylation Oxidation of reduced compounds (NADH, FADH2/QH2) in the ETC coupled to ATP synthesis. Oxidation is spontaneous and energy released is conserved in a proton gradient. Protons are pumped from the matrix to the intermembrane space. Oxidative phosphorylation is a more indirect process in which free energy is converted to (or conserved as) a transmembrane gradient of proons that is then used to drive ATP syntehses. Electrons removed from nutrients are transferred by dehydrogenases to the cofactors, NAD+ and FAD. Reduction potential indicates a substances tendency to accept electrons. Standard Reduction potential (’): affinity of a substance (such as Ubiq.) for electrons) The greater the value of ’, the greater the tendency of the oxidized form of the substance to accept electrons and become reduced Electrons flow spontaneously from substances with low to high reduction potential (from negative to a more positive reduction potential) Ex. The ’ for NAD+ (-.315V) is lower than in ubiquinone (0.045V), therefore NADH will tend to transfer its electrons to ubiquinone (NADH is oxidized, Q is reduced) Increase reduction potential = increase electron affinity The larger the difference in reduction potential the change in free energy = more spontaneous Mitochondrial Electron Transport Electrons are shuttle from NADH to O2 in a multistep process that offers several opportunities to conserve the free energy of oxidation Outer membrane- very porous, permits the transmembrane diffusion of substances with masses up to about 10 Kd Composition of the matrix differs from the intermembrane space Ionic composition of the intermembrane space is equivalent to the cytosol due to the presence of the porins in the outer membrane The relatively impermeable inner mitochondrial membrane encloses the proteinrich matrix – prevents the transmembrane movements of ions and small molecules. Individual mitochondria can move around the cell and undergo fusion and fission Transport of cytosolic NADH – Malate Aspartate and Glycerol-3-Phosphate Shuttle Much of the cell’s NADH and QH2 is generated by the TCA and fatty acid oxidation, which transfer electrons to the protein complexes of the respiratory ETC, such as Q, Complex I, Complex II, and Cyt C. However, the NADH produced by glycolysis and other oxidative processes in the cytosol cannot directly reach the mitochondria (respiratory chain) There is not a transport protein present to ferry NADH across the inner mitochondrial membrane (where ETC takes place) – instead “Reducing equivalents” are imported into the matrix: Malate-Asparatate Shuttle Mitochondria also need a mechanism to export ATP and to import ADP and Pi Adenine nucleotide translocase exports a ATP and imports ADP, binding one or the other and changing its conformation to release the bound nucleotide on the other side of the membrane Pi is imported from the cytosol in symport with H+ The protein complexes that carry out ETC and ATP synthesis are oriented in the inner mitochondrial membrane so that they can bind the NADH ATP translocase protein imports ADP, exports ATP Symport protein permits simultaneous movement of Pi and H+ Complex I transfers electrons from NADH to Q Complex I is the largest of the ETC (NADH Dehydrogenase) 45 different subunits, L-shaped protein with numerous transmembrane helices and a peripheral arm ETC takes place in the peripheral arm, which includes several prosthetic groups that undergo reduction as they receive electron to become oxidized as they give up their electrons to the next group Redox centers- prosthetic groups that undergo reduction and oxidation within the complex (E* between NAD and Q) Allows to form a chain where electrons travel a path of increasing reduction potential Two electrons donated by NADH are picked up by Flavin mononucleotide (FMN) FMN then transfers the electrons one at a time to Fe-S complex (9 total) Electrons travel between several Fe-S clusters (1 e carrier) before reaching ubiquinone Ubiquinone is a two-electron carrier, but it accepts one electron at a time from an Fe-S donor As electrons are transferred from NADH to Q, Complex I transfers 4 H+ from the matrix to the intermembrane space Each proton passes from one side of the membrane to the other via a proton wire Proton wire - series of hydrogen-bonded protein groups plus water molecules that form a chain through which a proton can be relayed Proton taken up from the matrix are not the same ones that are released into the intermembrane space Complex II – Other oxidation reactions contribute to the QH2 pool The reduced quinone product of Complex I joins a pool of reduced quinones which are augmented by the activity of other Redox reactions Complex II (Succinate dehydrogenase) Embedded in the inner membrane – not soluble in the mitochondrial matrix Produces QH2 in the TCA cycle Complex II is more like a tributary – does not undertake proton translocation and therefore does not directly contribute the free energy of redox reaction towards ATP synthesis (a) Another source of QH2 is fatty acid oxidation via the oxidation of a C-C bond in a fatty acid catalyzed by acylCoA dehydrogenase (b) Electrons from cytosolic NADH can also enter the Q-pool through cytosolic/mitochondrial glycerol-3phosphate dehydrogenase – shuttles electrons from NADH to Q (c) Bypasses complex I FADH2 goes from C2 to Q, only makes 1.5 ATP because only 6 H+ are pumped into the intermembrane space Cystolic NADH can start from C1 (heat and liver), which yields 2.5 ATP because 10 H+ are pumped into the intermembrane space Cystolic NADH can also go straight to Q (skeletal and brain), where it would only yield 1.5 ATP because 6 H+ are pumped into the inner membrane space (mitochondrial G-3-P dehydrogenase) G-3-P dehydrogenase is embedded in the inner membrane C1 = 4 H+, C3 = 4 H+, C4 = 2 H+ Complex III (Q:cytochrome c oxidoreductase/cytochrome bc1) Ubiquinol (QH2) is reoxidized by Complex III Transfers electrons to the peripheral membrane protein cytochrome C Complex III contains 2 cytochromes (b and c1) Functional Core: cytochrome b, c1, and iron-sulfur protein (Rieske protein) Cytochrome c transfers one electron at a time from C3 to C4 Flow of electrons through Complex III is complicated – two electrons donated by ubiquinol must split up to travel through a series of one-electron carriers that include cyt c1 and cyt b C3 has two active sites where Quinone cofactors undergo reduction and oxidation Q cycle Net Result Two electrons from QH2 reduce two molecules of cytochrome C Four protons are translocated to the intermembrane space: 2 from QH2 in the first round of the Q cycle and two from QH2 in the second round Complex IV (Cytochrome c oxidase) Cytochrome c ferries electrons between Complex III & IV Is soluble in the intermembrane space Complex IV is the last enzyme to deal with the electrons derived from the oxidation of metabolic fuels Four electrons delivered by cytochrome C are consumed in the reduction of molecular oxygen to water Relays four additional protons from the matrix to the IM space – two protons for every pair of electrons The production of water and the proton relays both deplete the matrix proton concentration and contribute to the formation of a proton gradient across the inner mitochondrial membrane. Free Radicals and aging Partial reduction of O2 by Complex IV or the other complexes can produce superoxide free radical O2 Can chemically alter nearby molecules – most of the damage is felt by the mitochondria, its lipids, proteins and DNA are susceptible to oxidation as superoxide steals and electron. As damage accumulates, mitochondria become less efficient to go through apoptosis Oxidative damage mediated by O2- and other free radicals is responsible for the degeneration of tissues that occurs with aging and is associated with Parkinson’s Disease and Alzheimer’s Disease All cells are equipped with antioxidant mechanisms Superoxide dismutase converts a superoxide to a less toxic product, peroxide Ascorbate and alpha-tocopherol may protect cells form oxidative damage by scavenging free radicals People that exercise regularly do not exhibit oxidative damage Weekend warriors are more susceptible to oxidative damages Chemiosmosis E from the H+ concentration gradient is used to make ATP via ATP synthase Energy is conserved in proton gradient during ETC Intermembrane space has a higher H+ concentration (lower pH) Matrix has a lower H+ concentration (higher pH) For a gradient to be created, ETC required For ATP synthase to function, need H+ gradient Proton wires: where H+ travel from matrix to intermembrane space (endergonic) overall spontaneous Chemiosmotic Theory: proton translocating activity of ETC complexes in the inner mitochondrial membrane generates a proton gradient across the membrane Protons cannot diffuse back into the matrix because the inner membrane is impermeable to ions Proton motive force- the imbalance of protons represents a source of free energy; this can drive the activity of ATP synthase The imbalance of protons, a nonequilibrium state, has an associated free E the force that would restore the system to equilibrium) Proton gradient has 2 components: reflecting the difference in the concentration of the chemical species and the difference in electrical charge of the positively charged protons = electrochemical gradient Matrix is more negative and intermembrane is more positive 10 protons translocated from the matrix to the intermembrane space is 200 kJ; therefore, going back it would be -200 kJ, which is enough to drive the phosphorylation of several molecules of ADP ATP synthase F1 (matrix) component has 3 alpha and 3 beta subunits surrounding a central shaft; only the beta serves catalytic role; catalytic subunit F0 (integrated in membrane) includes and A and two b subunits that extent upward to interact with the F1 component and a ring of c subunits; channel for H+ Mitochondrial ATP synthase has 8 c subunits Proton transport through ATP synthase involves the rotation of the c ring pass the stationary a subunit 3H+ needed to make one ATP, full rotation makes 3 ATP A c subunit can take a proton from the intermembrane space A slight rotation of the c ring brings another c subunit into position so that it can release its bound proton into the matrix All six subunits of F1 can bind adenine nucleotides, only beta subunits have catalytic activity The alpha/beta hexamer change their conformations as the y subunit rotates (like a shaft driven by the c ring “rotor”) The binding change mechanism explains how ATP is made ATP synthase uses mechanical energy to form a chemical bond (converts mechanical to chemical energy) The interaction between the y subunit and the alpha/beta hexamer explains this energy transduction Rotation-induced conformational change drives unfavorable ATP synthesis reaction Binding change mechanism- rotation-driven conformational changes alter the affinity of each catalytic beta subunit for an adenine nucleotide At any moment, each catalytic site has a different conformation (and binding affinity), referred to as the open, loose, or tight state 1. The substrates ADP and Pi bind to a beta subunit in the loose state 2. The substrates are converted to ATP as rotation of the y subunit causes the beta subunit to shift to the tight conformation 3. The product ATP is released after the next rotation, when the beta subunit shifts to the open conformation Because the three beta subunits of ATP synthase act cooperatively, they all change their conformation simultaneously as the y subunit turns A full rotation of 360 degrees is required to restore the enzyme to its initial state, but each rotation of 120 degrees results in the release of ATP from one of the three active sites In the absence of a proton gradient, no ATP is synthesized because there is no free E to drive the rotation of the y subunit P:O ration describes the stoichiometry of oxidative phosphorylation Chemical energy is transduced to a proton motive force, then to the mechanical movement of a rotary engine, and finally back to chemical energy in the form of ATP In oxidative phosphorylation the regulated set would be the reaction catalyzed by complex 4 (cytochrome c oxidase) Oxidative phosphorylation is regulated primarily by the availability of reduced co-factors NADH and FADH2 (QH2) Ch. 8 Lipids Lipids: are defined primarily by the absence of functional groups Non-polymeric Lack the ability to form hydrogen bonds Bulk of the structure is hydrocarbon like Several classifications: o Fatty Acids o Triacylglycerols o Glycerophospholipids o Sphingolipids o Isoprenoids: Cholesterol and Terpenes Functions of lipids: 1. Energy Metabolism: long term storage energy - non-polar lipids 2. Components of biological membranes – polar lipids with nitrogen and phosphate 3. Precursor of hormones – steroid class represented by cholesterol 4. Light absorbing pigment, electron carrier, signal molecules Fatty acids: simplest lipids that are long-chain carboxylic acids Can contain up to 24 carbon atoms Most common fatty acids are even numbered (C16 and C18) – Ex. palmitate and stereate Considered to be amphiphiles o Polar head group – carboxylic acid o Nonpolar tail – hydrocarbon chain Can be saturated with hydrogens, unsaturated, or polyunsaturated In unsaturated fatty acids, double bond usually has the cis configuration Free-Fatty acids in biological systems – they are usually esterified to glycerol Fats and oils in animals/plants are Triacylglycerols (acyl groups of three fatty acids are esterified to the three hydroxyl groups of glycerol via a condensation reactions) Triacylglycerols can aggregate in large globules. Serves as a storage depot for fatty acids that can be broken down metabolic energy. Tendency to aggregate means that cells can store a large amount interfering with other cellular activities in the aqueous cytosol. to release without it Humans cannot make any double bonds past carbon 9; other organisms can produce Omega-3-Fatty Acids Omega-3-Fatty Acid: has a double bond starting three carbons from the methyl end. Essential Fatty Acids cannot be synthesized by humans and must be supplied in the diet o Ex. EPA (C-20) and DHA (C-22) in cold-water fish; -linoleic acids in plants Linoleic and linolenic fatty acids are precursors for longer chain Omega3-Fatty Acids o Diets without fish oil acquire -linoleic acid from plants and lengthen the chain from the carboxylic end Why are essential fatty acids and omega-3-fatty acids important? Omega-3-fatty acids are essential for normal human growth EPA linked to decreased risk of cardiovascular disease – omega-3 compete with omega-6 for enzymes that convert fatty acids to signaling molecules o Omega-6 derivatives are stronger triggers of inflammation – atheroscelerosis o The relative amounts of omega-3 and omega-6 are more important than the absolute amount of omega-3 DHA (C-22): is abundant in the brain and retina and its concentration decreases with age. o Protects neural tissues from damage following a stroke, but cannot reverse cognitive decline associated with Alzheimer’s disease. Glycerophospholipids: contain a glycerol backbone with fatty acyl groups esterified at position 1 and 2, and a phosphate head group at position 3. Major lipids of the biological membrane Considered amphipathic: hydrophobic tails attached to polar/charged head groups. Phospholipases are enzymes that hydrolyzes glycerophospholipids to release acyl chains Phospholipids are cleaved at specific sites For degrading lipids Act as signaling molecules inside/between cells Sphingolipids: amphipathic membrane lipid molecules that use sphingosine as a backbone Sphingomyelins – have phosphocholine or phosphoethanolamine head groups In a sphingolipid, a second fatty acyl group is attached via an amide bond to the serine nitrogen Glycolipids are sphingolipids with one or more carbohydrate groups Cerebroside – monosaccharide head group Ganglioside – oligosaccharide head group Cholesterol: 27 carbon, four-ring molecule found in biological membranes. Important component in biological membranes Metabolic precursor of steroid hormones (such as estrogen and testosterone) Has an amphiphilic character It is an isoprenoid (terpenoids) – made out of a 5-carbon unit, isoprene Can be found in animal tissues Reactive Portion Ubiquitin is an isoprenoid derivative (functions in electron transport) Phytosterols: found in plants; main consituents in plant membrane structure Ex. stigmasterol, -sitosterol, campesterol Constituent in plant membrane structure Are found to decrease the level of LDL in the blood -sitosterol is found to be against enlargement of the prostate – may inhibit prostate cancer cells Clinical Connection Lipid Vitamins A, D, E, and K (isoprenoid derivatives) I. Vitamin A (Retinol) – derived from (orange pigment in carrots) Role in light reception in the eye; retinol is oxidized to retinal (an aldehyde) which functions as a light receptor’ Light causes retinal to isomerize, causing an impulse through the optic nerve Retinoic acid acts like a hormone by stimulating tissue repair II. Vitamin D (Steroid Derivative) Vitamin D2: derived from plants Vitamin D3: derived from endogenously produced cholesterol UV light is required for the formation of Vitamin D2 and D3 Enzymes in the liver and kidney convert Vitamin D to its active form, stimulating calcium absorption in the intestine o High [Ca2+] in bloodstream promotes Ca2+ deposition in bones and teeth III. Vitamin E (-Tocopherol): hydrophobic molecule incorporated into cell membranes Reacts with free radicals generated during oxidative reactions Stems from activity as a regulatory molecule that suppresses free-radical formation by inhibiting production or activation of oxidative enzymes Binds to biological membranes IV. Vitamin K (Phylloquinone): participates in enzymatic carboxylation of Glu residues in proteins involved in blood coagulation Vitamin K deficiency: prevents Glu carboxylation, inhibiting normal function of coagulating proteins causing excessive bleeding Half the daily uptake of Vitamin K is supplied by intestinal bacteria Excess vitamin A,D,E,K accumulate in fatty tissues. Excess vitamin D: kidney stones and abnormal calcification of soft tissues High levels of vitamin A: produce symptoms such as birth defects Terpenes Waxes (produced by plants on surfaces of leaves and fruits) In humans, derivative of Arachidonate (C-20): signaling molecules that regulate blood pressure, pain and inflammation. All molecules derived from isoprene. Provides odors and colors in plants. Capsaicin: compound found in chili peppers Hydrophobicity explains why it cannot be washed away with water. Pain Reliever: activates receptors on neurons that sense pain and heat and overwhelms the receptor, preventing neurons from receiving pain signals. Chapter 17 – Lipid Metabolism Atherosclerosis slow progressive disease that begins with the accumulation of lipids in the walls of large blood vessels (1/2 of US deaths) Trapped lipids initiate inflammation by triggering the production of chemical signals that attract leukocytes and macrophages – cells engorge themselves by taking up accumulated lipids, recruiting more macrophages leading to continued inflammation Damaged vessel wall forms a plaque with a core cholesterol, cholesteryl esters and dead macrophages that are surrounded by smooth muscle cells that can undergo calcification More energy from fats; cardiac muscle use 95% energy from fatty acids There is a higher energy of fatty acids because there are more reduced; more oxidation; more ATP Bile salts emulsify fatty acids (increase SA); more exposed to pancreatic lipases; break up micelles The products of emulsification are shuttled to brush border in bile micelles and then absorbed by enterocytes; bile reabsorbed by small intestine and transported to the liver Repackaging happens in intestinal mucosa Repackaged in lipoproteins Chylomicrons transport dietary TAG Lipoprotein lipases- breakdown lipoproteins TAGs reabsorbed by muscle or adipocytes What is the source of lipids that accumulate in vessel walls? They are deposited by the lipoprotein, LDL. Lipoproteins are made of phospholipids, can travel through bloodstream because of polar heads and proteins Lipids are in the core The density of lipoproteins are dependent on protein content and also differ in diameter o Chylomicrons are the largest (least dense) o HDL are the smallest (densest) Lipoproteins are the primary form of circulating lipid Dietary lipids travel from the intestine to other tissues as chylomicrons via the lymphatic system The main function of chylomicrons is to transport dietary TAG to adipose tissue and cholesterol to the liver The liver repackages the cholesterol and other lipids (TAG, phospholipids, cholesteryl esters) into VLDL VLDL give up TAG to the tissues, becoming smaller, denser, and richer in cholesterol and cholesteryl esters IDL becomes LDL LDL is considered bad because it has more TAG and cholesterol compared to HDL (good cholesterol) High concentration of circulating LDL measured as serum cholesterol; major factor in atherosclerosis Atherosclerosis less likely to occur in individuals who consume low-cholesterol diets and who have high levels of HDL Primary function of HDL is to transport body’s excess cholesterol back to the liver (Liver packages lipids) Counteracts the atherogenic tendencies of LDL HDL scrapes cholesterol from tissue to liver; signal to liver Cholesterol is negative feedback inhibitor of cholesterol synthesis Serum albumin- 50% of protein in blood stream, transports fatty acids, hormones Fatty Acid oxidation Lipoproteins carry TAGs to tissues where hydrolysis releases their fatty acids from the glycerol backbone The hydrolysis of TAG occurs extracellularly, catalyzed by lipoprotein lipase TAG that are stored in adipose tissue are mobilized by an intra-cellular hormone sensitive lipase Mobilized fatty acids travel through bloodstream bound to albumin (66 kD protein that accounts for half the serum protein, bind metal ions and hormones, serves as an allpurpose transport protein) The concentration of free fatty acids in the body is very low because these molecules are detergents and can disrupt cell membranes High glucose = insulin To be oxidatively degraded, a fatty acid must be activated Low glucose = glucagon, and epinephrine TAG activation requires 2 ATP equivalents, so we have to subtract 2 to achieve ne Activated fatty acids are acylated to CoA TAG activation occurs in the cytosol Reaction is driven by ATP hydrolysis The reaction is almost in equilibrium; to make it spontaneous, phospohanhydride bond is broken, releases E Catalyzed by Acyl-CoA synthetase Most cells contain a set of Acyl-CoA synthetases specific for fatty acids that are short, medium long, very long Acyl-CoA requires a transporter into mitochondria, carnitine acyl transferase (inhibited by malonylCoA) Cytosol into the mitochondria Beta Oxidation Beta oxidation is a spiral pathway that has 4 enzymatic steps; Each saturated fatty acid produces 1 NADH, 1 FADH2 (QH2), 1 Acetyl-CoA, an Acyl-CoA that is 2 C shorter Each Acetyl-CoA produced goes into the TCA cycle where 3 NADH, 1 FADH2, 1 ATP are produced Beta oxidation feeds directly into the ETC to produce ATP (NADH, FADH2 go into the ETC) Total ATP produced is 14 ATP from one round of beta oxidation (12 ATP net because 2 consumed in activated in cytosol) Rounds of beta oxidation: #C / 2 – 1 Beta oxidation occurs on the beta C, third carbon; the two carbons receding the beta C are removed during each round The steps of a beta oxidation round: Oxidation (produce FADH2) Hydration (enoyl-CoA hydratase) Oxidation (produce NADH) (Dehydrogenation) Cleavage (2 C at a time) The first step creates a trans 2, 3 double bond Enoyl-CoA hydratase requires cis 2, 3 double bond to be activated; unsaturated fatty acids have to go through extra steps to achieve this configuration; which is why they are less fattening Degradation of unsaturated fatty acids requires isomerization and reduction Common fatty acids (oleate and linoleate) contain cis double bonds – obstacle to enzymes that catalyze B-oxidation Linoleate has an acyl-CoA that has 3,4 double bond – cis enoyl-CoA Enoyl-CoA hydratase (2nd step of B-oxidation) recognizes only trans. Enoyl-CoA isomerase converts cis-3,4 to trans-2,3 double bond for B-oxidation to continue Isomerization skips step 1 (1 less FADH2) Reduction consumes NADPH = -1 NADH 4 total ATP less than saturated fatty acids per round of beta oxidation This is why unsaturated fatty acids are not as fattening, because they do not produce as much ATP Compounds with double bonds are more oxidized than saturated compounds, so less energy is released in converting them to CO2 The enoyl-CoA isomerase bypasses the QH2 producing acyl dehydrogenase step (1.5 fewer ATP) NADPH dependent reductase consumes 2.5 consumes 2.5 ATP Oxidation of odd-chain fatty acids yields propionyl-CoA Most fatty acids are even numbered Carbon chains because synthesizing fatty acids occurs by adding 2 C at a time (opposite of beta oxidation) The final round of beta oxidation of odd numbered fatty acids yields the three carbon compound propionyl-CoA rather than Acetyl-CoA The complete catabolism of carbons derived from propionyl-CoA requires that succinyl-CoA be converted to pyruvate and then to Acetyl-CoA, which enters the TCA cycle Propionyl-CoA Acetyl CoA Methylmalonyl-CoA mutase, which catalyzes step three is an unusual enzyme because it mediates a rearrangement of carbon atoms Requires a prosthetic group derived from B12 (cobalamin) Some fatty acid oxidation occurs in the peroxisomes Peroxisomes are organelles that are bound by a single membrane Peroxisome produce H2O2; which is broken down to water and O2 Longer fatty acids are oxidized > C16 and branched fatty acids as well in peroxisomes In plants, all fatty acid oxidation occurs in peroxisomes and glyoxysomes Electrons removed from Acyl-CoA are transferred from FADH2 to H2O2 instead of ubiquinone FADH2 does not form ATP Fatty acid oxidation is different than in peroxisomes Peroxisome catalase breaks down hydrogen peroxide The peroxisome serves as a chain shortening system because it has low affinity for short chains and high specificity for long chains – peroxisomes break down fatty acids unrecognized by mitochondrial enzymes Peroxisomes also degrade branched fatty acids (such as phytanate, which is from chlorophyll) Phytanate must be degraded by the peroxisome because because the methyl group at C3 prevents dehydrogenation by 3—hydroxyacyl-CoA dehydrogenase (step three of standard beta oxidation) Deficiency in phytanate-degrading enzymes results in Refsum’s disease, a degenerative neural disorder characterized by the accumulation of phytanate in the tissues Fatty Acid Synthesis Fatty acyl groups are built and degraded two carbons at a time The pathways for fatty acid synthesis and degradation must differ for thermodynamic reasons Fatty oxidation is a thermodynamically favorable process; reversing the steps of the pathway would not be energetically favorable Beta oxidation takes place in the mitochondrial matrix, while fatty acid synthesis takes place in the cytosol In beta oxidation, the acyl group is attached to coenzyme A In fatty acid synthesis, the acyl chain is bound by an acyl-carrier protein ACP Beta oxidation requires two ATP to activate the acyl group Fatty acid synthesis consumes one ATP and 2 NADPH per round (Addition of 2C) B7 (biotin) is the precursor of Acyl-Carrier protein and CoA NADPH is the source of reducing power Acyl chains are carried by: CoA in fatty acid oxidation Acyl-carrier protein in fatty acid synthesis Acetyl-CoA carboxylase catalyzes the first step of fatty acid synthesis Acetyl-CoA is the starting material for fatty acid synthesis (generated in mitochondria by the PDH) Acetyl-CoA cannot exit the cytosol for biosynthetic reactions Transport of acetyl groups to the cytosol involves the Citrate Transport System ATP is consumed in the ATP-citrate lyase reaction to drive formation of the thioester bond The first step of fatty acid synthesis is the carboxylation of acetyl-CoA (carried out by Acetyl-CoA carboxylase) Acetyl-CoA carboxylase is the regulatory enzyme (rate-controlling) CO2 (as bicarbonate) is “activated” by its attachment to a biotin (B7) prosthetic group (Uses 1 ATP) Malonyl-CoA is the donor of the 2C acetyl units that are used to build a fatty acid Biotin is the carrier of CO2 which is added to Acetyl-CoA to make malonyl-CoA We use the 3C malonyl-CoA because we need the other C for CO2 (released; E released) Fatty acid synthase catalyzes seven reactions The protein carrying out fatty acid synthesis is multifunctional enzyme made of two identical polypeptides Fatty acid synthase has six active sites to carry out seven discrete reaction In plants and bacteria, the reaction is catalyzed by separate polypeptides; same chem. Reactions 1 & 2 are transacetylation reactions – prime/load the enzyme w the reactants for the condensation reaction The condensation reaction combines Malonyl-ACP and Acetyl-Cys to make Acetoacetyl-ACP 1 ATP inputted to make malonyl-CoA; condensation reaction is energized form E released from CO2 release Malonyl-CoA is a two-carbon donor (CO2 released = E released) Growth of the acyl chain (like chain shortening in beta oxidation) occurs at the thioester end of the molecule The NADH required for reduction steps (4 and 6) of FAS is supplied by the Pentose Phosphate Pathway The synthesis of palmitate requires production of 7 malonyl-CoA at cost of 7 ATP Consumes 14 NADPH (= 35 ATP) – total cost of 42 ATP Packaging several enzyme activities into one multifunctional protein like mammalian fatty acid synthase allows the enzymes to be synthesized and controlled in a coordinated fashion The product of one reaction can quickly diffuse to the next acid site Other enzymes elongate and desaturate newly synthesized fatty acids Sphingolipids that contain C22 and C24 fatty acyl groups are generated by enzymes known as elongases They extend the C16 fatty acid produced by fatty acid synthase These reactions occur in the endoplasmic reticulum or mitochondria In ER: malonyl-CoA is the acetyl-group donor and are chemically similar to those of fatty acid synthase In mitochondria: fatty acids are elongated by reaction that closely resemble the reversal of beta-ox, but use NADPH Desaturases introduce double bonds into saturated fatty acids; take place in the ER Electrons removed in dehydrogenation of fatty acid are transferred to O2 to produce H2O Most common unsaturated fatty acids in animals are palmitoleate (C16) and oleate (C18); both with one cis double bond at the 9,10 position Trans fatty acids are relatively rare in plants and animals Elongation can follow desaturation (and vice versa) – animals can synthesize a variety of fatty acids , HOWEVER, mammals cannot introduce double bonds at positions beyond C9 Cannot synthesize fatty acids such as linoleate and linolenate (are precursors of arachidonate (C20) and other lipids with specialized biological activities) Mammals must obtain linoleate and linolenate from their diet Essential fatty acids are abundant in fish and plant oils Omega-3 fatty acids- unsaturated fatty acids with a double bond three carbons from the end, may have health benefits (deficiency from very-lowfat diet may cause slow growth/wound healing) Fats, Diet and Heat Disease (Biochemistry note) How do dietary lipids influence serum lipid levels? Diets rich in saturated fats have increased blood cholesterol (LDL) Diets rich in unsaturated vegetable oils replaced the saturated fats have the opposite effects Production of semisolid margarine from liquid plant oils (TAGs containing unsaturated fatty acids) often includes a hydrogenation step to chemically saturate the carbons of the fatty acyl chains In the hydrogenation process, some of the original cis double bonds are converted to trans Trans fatty acids have a tendency to increase LDL levels and decrease HDL levels Scientists still do not fully understand how the consumption of specific fatty acids (saturated or unsaturated, cis or trans) influences lipoprotein metabolism A consequence of low-fat diets is that individuals consume relatively more carbs Regulation of Fatty Acid Synthesis Under conditions of high metabolic fuel, the products of carb and amino acid catabolism are directed toward fatty acid synthesis (resulting fatty acids are stored as TAGs) Rate of FAS is regulated by Acetyl-CoA carboxylase, which catalyzes the first step of FAS Inhibited by palmitoyl-CoA Activated by citrate Is subject to allosteric regulation by hormone simulated phosphorylation and dephosphorylation High Acyl-CoA and citrate activates FAS Citrate activates FAS Malonyl-CoA is critical for preventing the wasteful simultaneous activity of fatty acid synthesis and fatty acid oxidation Is the source of acetyl groups that are incorporated into fatty acids Also blocks beta-oxidation by inhibiting carnitine acyltransferase (involved in shuttling acyl groups from the cytosol into the mitochondria) Consequently, when fatty acid synthesis is under way, no acyl groups are transported into the mitochondria for oxidation FAS inhibited by fatty acid Clinical Connection: inhibitors of FAS Triclosan was believed to act as a general microbicide (microbicide are effective because it is difficult for bacteria to evolve specific resistance mechanism) Triclosan operates more like an antibiotic with a specific biochemical target; enoyl-ACP reductase, which catalyzes step 6 of FAS In the active site, one of the phenyl rings of triclosan, whose structure mimics the structure of the reaction intermediate, stacks on top of the nicotinamide ring of the NADH cofactor Triclosan also binds through van der Waals interactions and hydrogen bonds with amino acid residues in the active site Isoniazid is used against M. tuberculosis Isoniazid is oxidized and the reaction product combines with NAD+ to generate a compound that inhibits one of the cell’s enoyl-ACP reductases (the target enzyme is specific for extremely long-chain fatty acids Some fungal species are susceptible to cerulenin, which inhibits its 3-ketoacyl-ACP synthase (step 3 of FAS) by blocking the reaction of malonyl-ACP (the condensation step) Cerulenin is also effective against M. tuberculosis, inhibiting the production of long-chain fatty acids required for cell-wall synthesis The drug contains a reactive epoxide group; reacts irreversibly with the enzyme’s active-site Cys residue forming a C2-S covalent bond; the hydrocarbon tail occupies the site that would normally accommodate the growing fatty acyl chain Acetyl-CoA can be converted to ketone bodies During a fast (when glucose is unavailable) tissues depend on fatty acids released from stored triacylglycerols, but the brain does not burn fatty acids (pass poorly through blood-brain barrier) Liver produces ketone bodies to supplement gluconeogenesis Acetoacetate and 3-hydroxybutyrate (ketone bodies) are synthesized from Acetyl-CoA in liver mitochondria by the process, Ketogenesis. Because it uses fatty acid-derived acetyl groups, it helps spare amino acids that would be diverted to gluconeogenesis Ketone body synthesis occurs in the liver mitochondria Liver does not use ketone bodies, however they donate synthesized ketone bodies, missing 3-ketoacyl-CoA transferase Assembly of ketone bodies is somewhat reminiscent of the synthesis of fatty acids or the oxidation of fatty acids in two-carbon steps Ketone bodies are transported in the bloodstream without specialized lipoproteins because they are small and water-soluble – easily pass through CNS High ketogenic activity (such as in diabetes), ketone bodies are produced faster than they are consumed. Excess acetoacetate breaks down to acetone - sweet breath is characteristic of acetone Ketone bodies are acidic, leading to a drop in blood pH – ketoacidosis Ketone bodies produced by the liver are used by other tissues as metabolic fuels after being converted back to Acetyl-CoA The liver itself cannot catabolize ketone bodies because it lacks 3-ketoacyl-CoA transferase Synthesis of Other Lipids TAGs and phospholipids are built from Acyl-CoA groups TAGS are synthesized by attaching fatty acyl groups to a glycerol backbone derived from phosphorylated glycerol or from glycolytic intermediates Synthetase requires ATP Synthase does not require ATP Ex. Dihydroxyacetone phosphate Fatty acyl groups are activated to CoA thioesters in an ATP-dep. manner catalyzed by Acyl-CoA synthetase (which also activates fatty acids for oxidation) Acyl transferase (that adds fatty acids to the glycerol backbone) are not highly specific, but human TAGs usually contain palmitate at C1 and unsaturated oleate at C2 The TAG biosynthetic pathway also provides the precursors for glycerophospholipids These amphipathic phospholipids are synthesized form phosphatidate (phosphorylated diacylglycerol) or diacylglycerol by pathways that include an activating step in which cytidine triphosphate (CTP) is cleaved (in some cases, the phospholipid head group is activates; in other cases, the lipid tail portion is activated) CTP is required for energy in order to activate the reactant The fundamental similarity between phospholipid and triacylglycerol is the glycerol backbone Phosphatidylserine is synthesized from phophatidylethanolamine by a head group exchange reaction in which serine displaces the ethanolamine head group Phosphatidylinositol synthesis: the diacylglycerol component is activated, rather than the head group, so the inositol head group adds to CDP diacylglycerol Glycerophospholipids (and sphingolipids) are components of cellular membranes New membranes are formed by inserting protein and lipids into preexisting membranes, mainly in the ER Newly synthesized membrane components reach final cellular destinations via vesicles that budd of the ER May undergo remodeling through the action of phospholipase and acyltransferases that remove and reattach different fatty acyl groups Cholesterol Synthesis begins with Acetyl-CoA Cholesterol molecules are built form two-carbon acetyl units The first steps of cholesterol synthesis resemble those of ketogenesis However, ketone bodies are synthesized in the mitochondria (and only in the liver), cholesterol is synthesized in the cytosol The reactions of cholesterol synthesis and ketogenesis diverge after the production of HMG-CoA In ketogenesis, HMG-CoA is cleaved to produce acetoacetate In cholesterol synthesis, the thioester of HMG-COA is reduced to alcohol, releasing mevalonate (6C) Mevalonate is converted to the five-carbon isopentyl pyrophosphate, an isoprene derivative. The isoprenoid derivative is the precursor of cholesterol As well as other isoprenoids such as ubiquinone, the farnesyl group that is attached to some lipid-linked membrane proteins, and pigments such as beta-carotene In cholesterol synthesis, six isoprene units condense to form squalene Cyclization of squalene leads to a structure with four rings, resembling cholesterol A total of 21 reactions are required to convert squalene to cholesterol (NADH or NADPH is required for several steps) The rate-determining step of cholesterol synthesis and the major control point is the conversion of HMG-CoA to mevalonate by HMG-CoA reductase (one of the most highly regulated enzymes known) Synthetic inhibitors known as statins bind extremely tightly to HMG-CoA reductase All the statins have and HMG-like group that acts as a competitive inhibitor of HMG-CoA binding to the enzyme The rigid hydrophobic groups also prevent the enzyme from forming a structure that would accommodate the pantothenate moiety of CoA Physiological effects of statins are to lower serum cholesterol levels by blocking mevalonate synthesis Cells must obtain cholesterol from circulating lipoproteins, but since mevalonate is also the precursor of other isoprenoids such as ubiquinone, the long-term use of statins can have negative side effects Cholesterol affects membrane fluidity Cholesterol packaged in lipoprotein Cholesterol can be used in several ways I. II. III. IV. V. It can be incorporated into a cells membrane It may be acylated to forma cholesteryl ester for storage or, in liver, for packaging in VLDL It is a precursor of steroid hormones such as testosterone and estrogen in the appropriate tissues It is a precursor of bile acids such as cholate Bile salts remove cholesterol; serve as route for cholesterol exodus Bile acids aid digestion by acting as detergents to solubilize dietary fats and make them more susceptible to lipases. Are mostly reabsorbed and recycled through liver for reuse, but some are excreted from the body – the only route for cholesterol disposal The cholesterol contained in bile will occasionally aggregate into lumps in the gallbladder forming gallstones Cells can synthesize cholesterol as well as obtain it from circulating DLD When LDL proteins dock with LDL receptor on the cell surface, the lipoprotein-receptor complex undergoes endocytosis – the lipoprotein is degraded and cholesterol enters the cytosol Familial hypercholesterolemia: genetic defect in the LDL receptor – the cells of homozygotes are unable to take up LDL, the concentration of serum cholesterol is about 3 times higher than normal High-density lipoproteins (HDL) are essential for removing excess cholesterol from cells The efflux of cholesterol requires the close juxtaposition of the cell membrane and an HDL particle as well as specific cell-surface proteins Such as the ABC transporter that acts as a flippase to move cholesterol from the cytosolic leaflet to the extracellular leaflet, from which it can diffuse into the HDL particle o Defect in the gene for the transporter cause Tangier disease which is characterized by accumulating of cholesterol in tissues and a high risk of heart attack Because cells do not break down cholesterol and because the accumulation of cholesterol is potentially toxic, the body must coordinate cholesterol synthesis and transport among tissues Cholesterol shuts down its own synthesis by inhibiting the synthesis of enzymes such as HMG-CoA reductase Cellular cholesterol also represses transcription of the gene for the LDL receptor Cholesterol metabolism in many cells is characterized by a balance between influx and efflux o Fatty acid metabolism has two opposing pathways of synthesis and degradation that operate in balance to meet the cell’s needs Chapter 18 – Nitrogen Metabolism Incorporation of ammonium ion into biological molecules 80% of air is Nitrogen (N2) and must be fixed (Fixed Nitrogen): Nitrite, Nitrate, and Ammonia Nitrogenase: enzyme that carries out reduction of N2 into NH3 (Nitrogen Fixation) Is a metalloprotein containing Fe-S clusters and Fe-Mo Cofactor (Molybdenum, Iron, Sulfur) Ammonia exists in protonated form (NH4+) Nitrogen fixing bacteria such as marine cyanobacteria and bacteria colonize root nodules of legumes Nitrogen Fixation – conversion of N2 to NH4+ using nitrogenase Consumes a lot of ATP Ferredoxin, a strong reducing agent donates 8 electrons Nitrate/Nitrite reductase catalyzes nitrate to nitrite to ammonia Bacteria produce nitrate via nitrification Other organisms convert nitrate to N2 via denitrification Ammonia assimilation via Glutamine Synthetase and Glutamate Synthase Glutamate and glutamine are at higher concentrations than other amino acids – carriers of amino groups Glutamine synthetase: produces glutamine from glutamate (Synthetase = uses ATP) Glutamate synthase: produces glutamate from glutamine (Synthase = does not use ATP) Glutamine synthetase: glutamate is phosphorylated via ATP, then ammonia reacts with the intermediate and displaces Pi to produce Glutamine Glutamate synthase: nitrogen is assimilated into an intermediate from the TCA cycle to produce Glu The source of nitrogen in Glutamate is Glutamine Glutamine is deaminated to produce a second Glutamate The combined action of the two enzymes assimilates fixed nitrogen (NH4+) into -ketoglutarate to produce Glutamate Mammals lack glutamate synthase, but [Glu] are relatively high due to other reactions Transamination: moves amino groups between compounds – is a reversible reaction Transaminase (or aminotransferase) catalyzes the transfer of an amino group to a -ketoacid o Use a prosthetic cofactor, Pyridoxal-5’-phosphate (PLP) – a prosthetic group that the amino group is transiently attached to o PLP is derived from pyridoxine (Vitamin B6) o Amino group transfer occurs through a Lys Residue (Schiff base linkage) Amino Acid Biosynthesis Amino Acids are synthesized from intermediates of glycolysis and the citric acid cycle Ultimate sources of essential amino acids (must be obtained from food) are from plants and microorganisms Nonessential Amino Acids (can be synthesized): Ala, Arg, Asn, Asp, Cys, Glu, Gln, Gly, Pro, Ser, Tyr Essential Amino Acids (must be obtained): His, Ile, Leu, Lys, Met, Phe, Thr, Trp, Val Several amino acids are easily synthesized from common intermediates Alanine produced from Pyruvate Aspartate produced from Oxaloacetate Glutamate from -Ketoglutarate Tetrahydrofolate: a carrier of one-carbon units in several reactions of amino acids and nucleotide metabolism. Mammals cannot synthesize folate and must be obtained from diet o Requirement for folate increases during first few weeks of pregnancy for development of fetal nervous system; prevents neural tube defects such as spina bifida Serine is derived from 3-phosphoglycerate (3PG, from glycolysis) and can give rise to Glycine via serine hydroxymethyl transferase Amino acids with sulfur, branched chains, or aromatic groups are more difficult to synthesize : Amino acids include Met, Cys, Tyr, Trp, His, Ile Ser Cysteine (Bacteria) (Humans) High levels of homocysteine in blood is associated with homocystinuria (excess homocysteine is excrete) Individuals develop atherosclerosis in children, homocysteine directly damages blood vessel walls Apartate is a precursor for Methionine, Threonine and Lysine Branched chain AA (Valine, Leucine and Isoleucine) are synthesized by pathways with pyruvate as a starting substrate and requires a step catalyzed by transaminase and glutamate as a substrate The synthesis for the Aromatic Amino Acids (Phenylalanine, Tyrosine, and Tyrptophan) begin with condensation of Phosphoenolpyruvate (3-C, glycolysis) and Erythose-4-phosphate (4-C, PPP) The 7 carbon reaction produce cyclizes and is modified to become chorismate Animals do no synthesize chorismate pathway is a target for agents that inhibit plant metabolism without affecting animals Phenylalanine and Tyrosine are derived from chorismate via diverging pathways In humans, tyrosine is produced by hydroxylating phenylalanine (Tyr is not an essential amino acid) Tryptophan Synthase: catalyzes final two steps of Trp synthesis Active sites in adjacent subunits are connected by a tunnel that accommodates indole Channeling: movement of reactant between two active sites – increases rate of metabolic process by preventing loss of intermediates Histidine is not formed from carbohydrate metabolites, rather is formed by ATP providing one nitrogen and one carbon; and glutamate and glutamine donating two N and remaining 5 carbons from 5-phosphoribosyl pyrophosphate (PRPP) Box 18-B – Glyphosate, the most popular herbicide Glycine phosphonate (glyphosphate) competes with PEP in pathway leading to chorismate Plants cannot manufacture aromatic AA without chorismate – glyphosate is an herbicide Glyphosate not directly absorbed to plant binds tight to soil particles and is broken down by bacteria Has less potential to contaminate water supply For effectiveness: glyphosate must enter plant tissues, therefore is packaged in a surfactant (amphiphilic) that helps penetrate waxy coatings of leaves Farmers plant genetically engineered glyphosate-resistant crops (corn, soybean, cotton) that express a bacterial version of the enzyme that uses PEP but not inhibited by glyphosate Amino acids are precursors of some signaling molecules Amino acids can be precursors to neurotransmitters Common neurotransmitters: glycine, glutamate, and -aminobutyrate (GABA, a glutamate derivate) Catecholamines: amino acid neurotransmitters that resemble catechol Ex. Tyrosine gives rise to dopamine, norepinephrine, and epinephrine Tryptophan (a catecholamine) is precursor of serotonin, which is a precursor to melatonin Box 18-C Nitric Oxide Arginine is a precursor for the signaling molecule, free-radical nitric oxide (NO) – relaxation factor that elicits vasodilation NO is synthesized by nitric oxide synthase (has cofactors FMN, FAD, tetrahydrobiopterin, & a heme group) At low concentrations, it induces blood vessel dilation At high concentrations (along with oxygen radicals), kills pathogens Cannot be stockpiled for later release; diffuses into cells – no cell-surface receptor or degradative enzyme Is produced when and where it is needed Nucleotide Biosynthesis Nucleotides are synthesized from precursors that include amino acids *Why don’t humans require purine and pyrimidines in their diet? Purine nucleotide synthesis Purines nucleotides (AdenineMP and GuanosineMP) are synthesized by building purine base onto ribose-5phosphate First step: Ribose-5-phosphate into 5-phosphoribosyl pyrophosphate (also precursor to histidine) Next ten steps require Glu, Gly, Asp, bicarbonate and a formyl from tetrahydrofolate to produce a nucleotide product: Inosine Monophosphate (IMP) IMP is the substrate for the pathway that yield AMP and GMP GTP participates in AMP synthesis: amino group from aspartate is transferred to purine ATP participates in GMP synthesis: glutamate is source of amino group High concentrations of ATP GMP Production High concentrations of GTP AMP Production *Regulated via Feedback inhibition Pyrimidine nucleotide synthesis *What are the requirements for pyrimidine nucleotide synthesis? Pyrimidine nucleotides (UracilTP and CytosineTP) are synthesized as a base that attaches to 5-phosphoribsyl pyrophosphate Six-step pathway yields Uridine Monophosphate (UMP) requires Glu, Asp, and bicarbonate UMP is phosphorylated into UDP and then to UTP, then CTP synthase catalyzes amination of UTP to CTP (glutamine is the donor) Regulated via Feedback Inhibition by UMP, UDP and UTP ATP activates the enzyme Ribonucleotide reductase: converts ribonucleotides to deoxyribonucleotides RNA has catalytic activity due to the OH on Carbon 2 o Ribose: OH on C2 o Deoxyribose: H on C2 – is more stable than ribose; DNA carries genetic information Each of the four nucleoside triphosphates (NTPs) are converted to their diphosphate (NDP) form, ribonucleotide reductase replaces the 2’ OH group, the resulting deoxynucleoside diphosphate (dNP) is phosphorylated to produce the corresponding triphosphate (dNTP) Thyrimidine nucleotides are produced by methylation *Explain the importance of thymidylate synthase and dihydrofolate reductase reactions Ribonucleotide reductase, followed by kinase phosphorylation generates dATP, dCTP, dGTP, and dUTP dUTP is not used for DNA synthesis it is rapidly converted to thymine nucleotides Thymine nucleotides help prevent accidental incorporation of uracil into DNA dUTP is hydrolyzed to dUMP, then thymidylate synthase adds methyl to dUMP to produce dTMP using methylene-tetrahydrofolate as one-carbon donor Main source of methylene-tetrahydrofolate: serine hydroxymethyltransferase reaction (serine to glycine) Thymidylate synthase oxidizes tetrahydrofolate to dihydrofolate Dihydrofolate reductase (NADPH-dependent enzyme) then regenerates tetrahydrofolate cofactor dTMP is then phosphorylated to dTTP – substrate for DNA polymerase In cancer cells that undergo rapid cell division, thymidylate synthase and dihydrofolate reductase are highly active Compounds that inhibit these reactions are anti-cancer agents Methoxetrate (an antifolate) is a competitive inhibitor of dihydrofolate reductase because they compete for binding o In presence of methoxetrate, Cancer cells cannot regenerate tetrahydrofolate required for dTMP production and cell dies Nucleotide degradation produces uric acid or amino acids Nucleotides from food or from synthesis by cells can be broken down to release ribose groups and purine/pyrimidine that can: Be further catabolized and excreted (purines) Or be used as metabolic fuel (pyrimidines) At some points in degradation pathways, intermediates can be redirected towards synthesis of new nucleotides by salvage pathways Degradation of nucleoside phosphate Begins with dephosphorylation to produces a nucleoside Phosphorylase breaks glycosidic bond between the base and ribose in a nucleoside The phosphorylated ribose can be catabolized or salvaged via conversion to 5-PRPP for synthesis of another nucleotide – fate of base depends on whether purine or pyrimidine: I. Purine base catabolism: purines are eventually converted into Uric Acid Other organisms may further catabolize urate to generate Urea or Ammonia May require deamination and oxidation – depends if base was adenine, guanine or hypoxanthine Generates a waste produce that is excreted from the body II. Pyrimidine base catabolism: cytosine, thymine, and uracil undergo deamination and reduction to open the pyrimidine ring and further catabolism of these produces the amino acids: -alanine (from cytosine and uracil) or -aminoisobutyrate (from thymine) Contributes to the pool of metabolites for anabolic and catabolic processes Amino Acid Catabolism Amino acids are metabolic fuels that can be broken down to release free energy. Amino acids are major fuel for cells in the lining of the small intestine, which absorb dietary amino acids and break down available Glu, Asp, and Gln supply The liver catabolize AA’s originating from the diet and from normal turnover of intracellular proteins. o During periods of unavailable dietary AA’s (fasting), amino acids are mobilized from breakdown of muscle tissue – 40% of total protein in the body. o AA’s undergo transamination and carbon skeletons enter metabolic pathways (TCA cycle) o However, there is not enough oxygen in liver for complete oxidation to CO2, therefore, amino acids are partially oxidized to substrates for gluconeogenesis or ketogenesis Classification of amino acids in terms of catabolism Glucogenic: giving rise to gluconeogenic precursors (TCA intermediates) Ketogenic: giving rise to Acetyl-coA; used for ketogenesis or fatty acid synthesis Some amino acids are converted to gluconeogenic substrates via transamination Alanine to Pyruvate Aspartate to Oxaloacetate Glutamate to -Ketoglutarate Cysteine can be converted into pyruvate that releases ammonia and sulfur Threonine is both glucogenic and ketogenic because it is broken down into acetyl-CoA and glycine Acetyl-CoA is a precursor for ketone bodies Glycine is potentially glucogenic if it is first converted to serine vie serine hydromethyl transferase o Glycine Cleavage System: major route for glycine disposal (multienzyme complex) Degradation for Branched-chain AA’s (Val, Leu, Ile) and Aromatic AA’s (Phe, Tyr, and Trp) are more complicated Ile yields succinyl-CoA and acetyl-CoA Leu yields acetyl-Aoa and acetoacetate (ketone body) Lysine yields Acetyl-CoA and acetoacetate (different pathway from branched AA’s) Met yields succinyl-CoA Phe, Tyr and Trp yield acetoacetate (Ketone Body) Box 18-D Inborn Errors of Metabolism Archibald Garrod: recognized link between genes and disease – studied alcaptonuria (urine turns black upon exposure to air due to homogentisate, a product of tyrosine catabolism – homogentisate is excreted because there is a missing or defective homogentisate dioxygenase enzyme) Phenylketonuria (PKU) results from deficiency of phenylalanine hydrolase Phenyalanine cannot be broken down but can undergo transamination that results in an accumulation of phenylpyruvate (-keto acid derivative) that is excreted in the urine Nitrogen Disposal: The Urea Cycle Excess supply of AA’s for cell’s immediate needs for protein synthesis or AA-consuming pathways causes carbon skeletons to break down and the nitrogen disposed of All AA’s (except lys) can be deaminated but they are not eliminated from the body. Approximately 80% of the body’s excess nitrogen is excreted in the form of Urea *How are amino groups for amino acids incorporated into urea? Glutamate supplies nitrogen to the urea cycle Glutamate is deaminated to regenerate -ketoglutarate and release NH4+ via glutamate dehydrogenase Many transaminases use -ketoglutarate as amino group acceptor – glutamate is abundant Only known enzyme that can use NAD+ or NADP+ as a cofactor Major route for feeding AA-derived amino groups into urea cycle Is subject to allosteric activation/inhibition The starting substrate for the urea cycle is an activated molecule produced from the condensation of bicarbonate and ammonia via carbamoyl phosphate synthetase The Urea Cycle The carbamoyl phosphate synthetase and glutamate dehydrogenase reaction are combined with the reactions of the Urea cycle are combined Overall effect: transaminated AA’s donate amino groups via glutamate and aspartate to urea synthesis Amino acids can be disposed via two routes: 1. Carb 2. Glu N-acetylglutamate is an activator of carbamoyl phosphate synthetase Rate of urea production is controlled by this enzyme and is allosterically synthesized by N-acetylglutamate (produced from glutamate and acetyl-CoA) Bacteria, fungi, and other organisms use urease to break down urea. Page Left Blank