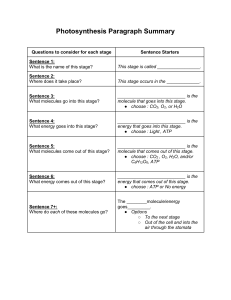
")
INSTRUCTIONAL MATERIAL FOR NUDI 20125: NUTRITIONAL BIOCHEMISTRY (Lecture and Laboratory) Compiled by: Reynaldo L. Magpantay, Jr., RND Department of Nutrition and Dietetics College of Science i TABLE OF CONTENTS Lecture Module Introduction Licensing and Attribution Unit 1. Chemical Foundation of Life Lesson 1: Water Lesson 2: pH, Buffers, Acids and Bases Lesson 3: Carbon Unit summary Unit Critical Thinking Questions Unit 2: Macromolecules Lesson 1: Carbohydrates Lesson 2: Lipids Lesson 3: Protein Lesson 4: Nucleic Acids Unit Summary Unit Test Unit 3: Enzymes Lesson 1: Enzymes Unit Summary Unit Critical Thinking Questions Unit Test Unit 4: Cellular Respiration Lesson 1: Overview of the Metabolic Reactions Lesson 2: Energy in the Living Systems Lesson 3: Glycolysis Lesson 4: Oxidation of Pyruvate and Krebs Cycle Lesson 5: Electron Transport Chain Lesson 6: Metabolism without Oxygen a. Lactic Acid Fermentation b. Alcoholic Fermentation c. Cori Cycle Lesson 7: Connections of the Carbohydrates, Protein and Lipids Pathway a. Glycogenesis b. Glycogenolysis Lesson 8: Gluconeogenesis Lesson 9: Lipid Metabolism a. Beta-Oxidation of Fatty Acids b. Ketogenesis c. Ketolysis d. Lipogenesis Lesson 10: Protein Metabolism a. Urea Cycle Unit Summary Unit Critical Thinking Questions Unit Test References PAGE i iii 1 1 6 9 16 17 18 19 27 35 44 52 54 57 57 66 67 68 69 70 75 80 85 90 96 96 97 98 99 100 102 107 110 110 111 113 115 119 122 123 126 ii Laboratory Special Paper Exercise 1 Part 1: Summer Must-Do: Ice Cream in a Bag Part 2: Cartesian Diver Experiment Exercise 2: Fruits Gone Bad? Discover Enzymatic Browning Exercise 3: Growing Yeast: Sugar Fermentation Exercise 4: Extraction of Fat Exercise 5: Denaturing Protein References 128 129 134 139 145 153 159 iii Welcome to Nutritional Biochemistry! It aims for you to explore and understand the structures, metabolisms, and interrelationship of nutrient biomolecules such as carbohydrates, protein, lipids, vitamins and minerals in the human body and how they relate to nutritional health. Isn’t amazing? Isn’t surprising? Isn’t it? After completing this course, you will be able to: 1. Describe the chemical structures that make up the components of living matter. 2. Understand the special properties of water and how the aqueous environment influences the behavior of biological macromolecules. 3. Identify the structural elements of proteins, the basic features of enzyme catalysis and regulation 4. Describe the synthesis of proteins, lipids, nucleic acids, and carbohydrates and their role in metabolic pathways Mode of Delivery: This course will be delivered remotely. Thus, you will be studying on your own. By default, our classes will be conducted asynchronously in my self-made website www.myyaya.info or www.ouryaya.blogspot.com Communication Plan: For those who have internet connection, please be reminded that we will do the following: • Synchronous Session I will hold 6 synchronous sessions (max of 3 hour each) with you, primarily for consultations, clarifications and monitoring and feedback. This be conducted on a whole class basis through Zoom (or any workable platform). • Course site I will post in the website (www.myyaya.info) any announcements or updates relevant to the course. You can find there a feedback form and I will try my best to get back to you within 48 hours. You will find some videos related to our topic. I highly recommend that you read the topic in this module before you watch the video. For those who do not have internet connection, please follow the sequence of this module. If you have concerns, you may reach me at 0966-442-3992 (GLOBE). You are also welcome to visit and watch the videos uploaded in the website ☺ Study Schedule: You will see a table below that provides the calendar of course activities. It shows the weekly topics and expected output from you. i LECTURE GUIDE WEEK TOPICS EXPECTED OUTPUT Week 1 Overview of the course PUP and CS Vision, Mission, and Objectives Course requirements None Week 2 Unit 1: Chemical Foundation of Life Answer the Critical Thinking Questions Week 3-4 Unit 2: Macromolecules Answer the Practice Tests and Unit Test Week 5 Unit 3: Enzymes Answer the Critical Thinking Question and Unit Test Week 6 Unit 4: Lessons 1 and 2 None Week 7 Unit 4: Lessons 3, 4 and 5 None Week 8 Unit 4: Lessons 6 and 7 None Week 9 Unit 4: Lesson 8 None Week 10 Unit 4: Lessons 9, 10 and Unit Summary Answer the Unit Test and Critical Thinking Questions GRADING SYSTEM: You will be graded according to the following: CRITERIA LECTURE CLASS Class Standing (70%) Critical Thinking Questions (Unit 1 3 4) and Practice Tests (Unit 4) Major Examinations (30%) Unit Test from Unit 2 and Unit 4 100% 1.0 1.25 1.5 1.75 2.0 2.25 2.50 2.75 3.0 5.0 INC W D TOTAL Excellent Very Good Good Satisfactory Pass Fail Incomplete Withdrawn Dropped 97-100 94-96 91-93 88-90 85-87 82-84 79-81 76-78 75 65-74 About the Instructor: My name is Reynaldo L. Magpantay, Jr, or you can just call me Sir Mags (short for Magpantay). This is our first time to do a remote learning. How I wish we can go back to normal and do face-to-face activities. I miss school! I am here now in Los Banos, Laguna eating Buco Pie. jk. I am not always online due to increasing workload from the University, so please bear with me. More info about me in the website. I made that! That’s why I am proud of myself HAHA! You may reach me at rlmagpantayjr@pup.edu.ph. Mags Magpantay on Facebook – Mon-Fri 8AM5PM only. GOODLUCK to all of us ☺ ii Licensing and Attribution This learning module was created as part of your undergraduate program in BS Nutrition and Dietetics at the Polytechnic University of the Philippines. The module could take from 10 class periods, depending on how much time you decide to spend on each part. This is a selfpaced module. It could be taught in consecutive classes, or spread out over an extended time (once a week, for example). Lecture videos are also available at www.myyaya.info Majority of the parts of this instructional material is derived from the https://openstax.org/books/biology-2e/pages/1-introduction licensed under a CC BY-NC-SA 4.0 license (https://creativecommons.org/licenses/by-nc-sa/4.0/) iii UNIT I. CHEMICAL FOUNDATION OF LIFE Lesson 1: Water By the end of this section, you will be able to do the following: 1. Describe the properties of water that are critical to maintaining life 2. Explain why water is an excellent solvent 3. Provide examples of water’s cohesive and adhesive properties 4. Discuss the role of acids, bases, and buffers in homeostasis Why do scientists spend time looking for water on other planets? Why is water so important? It is because water is essential to life as we know it. Water is one of the more abundant molecules and the one most critical to life on Earth. Water comprises approximately 60–70 percent of the human body. Without it, life as we know it simply would not exist. The polarity of the water molecule and its resulting hydrogen bonding make water a unique substance with special properties that are intimately tied to the processes of life. Life originally evolved in a watery environment, and most of an organism’s cellular chemistry and metabolism occur inside the watery contents of the cell’s cytoplasm. Special properties of water are its high heat capacity and heat of vaporization, its ability to dissolve polar molecules, its cohesive and adhesive properties, and its dissociation into ions that leads to generating pH. Understanding these characteristics of water helps to elucidate its importance in maintaining life. Water’s Polarity One of water’s important properties is that it is composed of polar molecules: the hydrogen and oxygen within water molecules (H2O) form polar covalent bonds. While there is no net charge to a water molecule, water's polarity creates a slightly positive charge on hydrogen and a slightly negative charge on oxygen, contributing to water’s properties of attraction. Water generates charges because oxygen is more electronegative than hydrogen, making it more likely that a shared electron would be near the oxygen nucleus than the hydrogen nucleus, thus generating the partial negative charge near the oxygen. As a result of water’s polarity, each water molecule attracts other water molecules because of the opposite charges between water molecules, forming hydrogen bonds. Water also attracts or is attracted to other polar molecules and ions. We call a polar substance that interacts readily with or dissolves in water hydrophilic (hydro- =“water”; philic =“loving”). In contrast, nonpolar molecules Figure 1.1. Oil and water do not mix. As this macro image of oil and water shows, oil does not dissolve in water but forms droplets instead. This is because it is a nonpolar compound. (credit: Gautam Dogra). 1 such as oils and fats do not interact well with water, as Figure 1.1 shows. A good example of this is vinegar and oil salad dressing (an acidic water solution). We call such nonpolar compos hydrophobic (hydro- = “water”; -phobic = “fearing”). Water’s States: Gas, Liquid, and Solid The formation of hydrogen bonds is an important quality of the liquid water that is crucial to life as we know it. As water molecules make hydrogen bonds with each other, water takes on some unique chemical characteristics compared to other liquids and, since living things have a high water content, understanding these chemical features is key to understanding life. In liquid water, hydrogen bonds constantly form and break as the water molecules slide past each other. The water molecules' motion (kinetic energy) causes the bonds to break due to the heat contained in the system. When the heat rises as water boils, the water molecules' higher kinetic energy causes the hydrogen bonds to break completely and allows water molecules to escape into the air as gas (steam or water vapor). Alternatively, when water temperature reduces and water freezes, the water molecules form a crystalline structure maintained by hydrogen bonding (there is not enough energy to break the hydrogen bonds) that makes ice less dense than liquid water, a phenomenon that we do not see when other liquids solidify. Water’s lower density in its solid form is due to the way hydrogen bonds orient as they freeze: the water molecules push farther apart compared to liquid water. With most other liquids, solidification when the temperature drops includes lowering kinetic energy between molecules, allowing them to pack even more tightly than in liquid form and giving the solid a greater density than the liquid. The lower density of ice, as Figure 1.2 depicts, an anomaly causes it to float at the surface of liquid water, such as in an iceberg or ice cubes in a glass of water. In lakes and ponds, ice will form on the water's surface creating an insulating barrier that protects the animals and plant life in the pond from freezing. Without this insulating ice layer, plants and animals living in the pond would freeze in the solid block of ice and could not survive. The expansion of ice relative to liquid water causes the detrimental effect of freezing on living organisms. The ice crystals that form upon freezing rupture the delicate membranes essential for living cells to function, irreversibly damaging them. Cells can only survive freezing if another liquid like glycerol temporarily replaces the water in them. Figure 1.2 Hydrogen bonding makes ice less dense than liquid water. The (a) lattice structure of ice makes it less dense than the liquid water's freely flowing molecules, enabling it to (b) float on water. (credit a: modification of work by Jane Whitney, image created using Visual Molecular Dynamics (VMD) software1; credit b: modification of work by Carlos Ponte) 2 Water’s High Heat Capacity Water has the highest specific heat capacity of any liquids. We define specific heat as the amount of heat one gram of a substance must absorb or lose to change its temperature by one degree Celsius. For water, this amount is one calorie. It therefore takes water a long time to heat and a long time to cool. In fact, water's specific heat capacity is about five times more than that of sand. This explains why the land cools faster than the sea. Due to its high heat capacity, warm blooded animals use water to more evenly disperse heat in their bodies: it acts in a similar manner to a car’s cooling system, transporting heat from warm places to cool places, causing the body to maintain a more even temperature. Water’s Heat of Vaporization Water also has a high heat of vaporization, the amount of energy required to change one gram of a liquid substance to a gas. A considerable amount of heat energy (586 kcal) is required to accomplish this change in water. This process occurs on the water's surface. As liquid water heats up, hydrogen bonding makes it difficult to separate the liquid water molecules from each other, which is required for it to enter its gaseous phase (steam). As a result, water acts as a heat sink or heat reservoir and requires much more heat to boil than other liquid such as ethanol (grain alcohol), whose hydrogen bonding with other ethanol molecules is weaker than water’s hydrogen bonding. Eventually, as water reaches its boiling point of 100° Celsius (212° Fahrenheit), the heat is able to break the hydrogen bonds between the water molecules, and the kinetic energy (motion) between the water molecules allows them to escape from the liquid as a gas. Even when below its boiling point, water’s individual molecules acquire enough energy from other water molecules such that some surface water molecules can escape and vaporize: we call this process evaporation. The fact that hydrogen bonds need to be broken for water to evaporate means that bonds use a substantial amount of energy in the process. As the water evaporates, energy is taken up by the process, cooling the environment where the evaporation is taking place. In many living organisms, including in humans, the evaporation of sweat, which is 90 percent water, allows the organism to cool so that it can maintain homeostasis of body temperature. Water’s Solvent Properties Since water is a polar molecule with slightly positive and slightly negative charges, ions and polar molecules can readily dissolve in it. Therefore, we refer to water as a solvent, a substance capable of dissolving other polar molecules and ionic compounds. The charges associated with these molecules will form hydrogen bonds with water, surrounding the particle with water molecules. We refer to this as a sphere of hydration, or a hydration shell, as Figure 1.3 illustrates and serves to keep the particles separated or dispersed in the water. When we add ionic compounds to water, the individual ions react with the water molecules' polar regions and their ionic bonds are disrupted in the process of dissociation. Dissociation occurs when atoms or groups of atoms break off from molecules and form ions. Consider table salt (NaCl, or sodium chloride): when we add NaCl crystals to water, the NaCl molecules dissociate into Na+ and Cl– ions, and spheres of hydration form around the ions, as Figure 1.3 illustrates. The partially negative charge of the water molecule’s oxygen surrounds the positively charged sodium ion. The hydrogen's partially positive charge on the water molecule surrounds the negatively charged chloride ion. 3 Figure 1.3 When we mix table salt (NaCl) in water, it forms spheres of hydration around the ions. Water’s Cohesive and Adhesive Properties Have you ever filled a glass of water to the very top and then slowly added a few more drops? Before it overflows, the water forms a dome-like shape above the rim of the glass. This water can stay above the glass because of the property of cohesion. In cohesion, water molecules are attracted to each other (because of hydrogen bonding), keeping the molecules together at the liquid-gas (water-air) interface, although there is no more room in the glass. Cohesion allows for surface tension, the capacity of a substance to withstand rupturing when placed under tension or stress. This is also why water forms droplets when on a dry surface rather than flattening by gravity. When we place a small scrap of paper onto a water droplet, the paper floats on top even though paper is denser (heavier) than the water. Cohesion and surface tension keep the water molecules' hydrogen bonds intact and support the item floating on the top. It’s even possible to “float” a needle on top of a glass of water if you place it gently without breaking the surface tension, as Figure 1.4 shows. Figure 1.4 A needle's weight pulls the surface downward. At the same time, the surface tension pulls it up, suspending it on the water's surface preventing it from sinking. Notice the indentation in the water around the needle. (credit: Cory Zanker) These cohesive forces are related to water’s property of adhesion, or the attraction between water molecules and other molecules. This attraction is sometimes stronger than water’s cohesive forces, especially when the water is exposed to charged surfaces such as those on the inside of thin glass tubes known as capillary tubes. We observe adhesion when water “climbs” up the tube placed in a glass of water: notice that the water appears to be higher on the tube's sides than in the middle. This is because the water molecules are attracted to the 4 capillary's charged glass walls more than they are to each other and therefore adhere to it. We call this type of adhesion capillary action, as Figure 1.5 illustrates. Figure 1.5 The adhesive forces exerted by the glass' internal surface exceeding the cohesive forces between the water molecules themselves causes capillary action in a glass tube. (credit: modification of work by Pearson-Scott Foresman) Why are cohesive and adhesive forces important for life? Cohesive and adhesive forces are important for transporting water from the roots to the leaves in plants. These forces create a “pull” on the water column. This pull results from the tendency of water molecules evaporating on the plant's surface to stay connected to water molecules below them, and so they are pulled along. Plants use this natural phenomenon to help transport water from their roots to their leaves. Without these properties of water, plants would be unable to receive the water and the dissolved minerals they require. In another example, insects such as the water strider, as Figure 1.6 shows, use the water's surface tension to stay afloat on the water's surface layer and even mate there. Figure 1.6 Water’s cohesive and adhesive properties allow this water strider (Gerris sp.) to stay afloat. (credit: Tim Vickers) 5 Lesson 2: pH, Buffers, Acids, and Bases The pH of a solution indicates its acidity or basicity. You may have used litmus or pH paper, filter paper treated with a natural water-soluble dye for use as a pH indicator, tests how much acid (acidity) or base (basicity) exists in a solution. You might have even used some to test whether the water in a swimming pool is properly treated. In both cases, the pH test measures hydrogen ions' concentration in a given solution. Hydrogen ions spontaneously generate in pure water by the dissociation (ionization) of a small percentage of water molecules into equal numbers of hydrogen (H+) ions and hydroxide (OH-) ions. While the hydroxide ions are kept in solution by their hydrogen bonding with other water molecules, the hydrogen ions, consisting of naked protons, immediately attract to unionized water molecules, forming hydronium ions (H3O+). Still, by convention, scientists refer to hydrogen ions and their concentration as if they were free in this state in liquid water. The concentration of hydrogen ions dissociating from pure water is 1 × 10 -7 moles H ions per liter of water. Moles (mol) are a way to express the amount of a substance (which can be atoms, molecules, ions, etc.). One mole represents the atomic weight of a substance, expressed in grams, which equals the amount of the substance containing as many units as there are atoms in 12 grams of 12C. Mathematically, one mole is equal to 6.02 × 10 23 particles of the substance. Therefore, 1 mole of water is equal to 6.02 × 10 23 water molecules. We calculate the pH as the negative of the base 10 logarithm of this concentration. The log10 of 1 × 10 -7 is 7.0, and the negative of this number (indicated by the “p” of “pH”) yields a pH of 7.0, which is also a neutral pH. The pH inside of human cells and blood are examples of two body areas where near-neutral pH is maintained. + Non-neutral pH readings result from dissolving acids or bases in water. Using the negative logarithm to generate positive integers, high concentrations of hydrogen ions yield a low pH number; whereas, low levels of hydrogen ions result in a high pH. An acid is a substance that increases hydrogen ions' (H +) concentration in a solution, usually by having one of its hydrogen atoms dissociate. A base provides either hydroxide ions (OH–) or other negatively charged ions that combine with hydrogen ions, reducing their concentration in the solution and thereby raising the pH. In cases where the base releases hydroxide ions, these ions bind to free hydrogen ions, generating new water molecules. The stronger the acid, the more readily it donates H +. For example, hydrochloric acid (HCl) completely dissociates into hydrogen and chloride ions and is highly acidic; whereas the acids in tomato juice or vinegar do not completely dissociate and are weak acids. Conversely, strong bases are those substances that readily donate OH – or take up hydrogen ions. Sodium hydroxide (NaOH) and many household cleaners are highly alkaline and give up OH – rapidly when we place them in water, thereby raising the pH. An example of a weak basic solution is seawater, which has a pH near 8.0. This is close enough to a neutral pH that marine organisms have adapted in order to live and thrive in a saline environment. 6 The pH scale is, as we previously mentioned, an inverse logarithm and ranges from 0 to 14 (Figure 1.7). Anything below 7.0 (ranging from 0.0 to 6.9) is acidic, and anything above 7.0 (from 7.1 to 14.0) is alkaline. Extremes in pH in either direction from 7.0 are usually inhospitable to life. The pH inside cells (6.8) and the pH in the blood (7.4) are both very close to neutral. However, the environment in the stomach is highly acidic, with a pH of 1 to 2. As a result, how do stomach cells survive in such an acidic environment? How do they homeostatically maintain the near neutral pH inside them? The answer is that they cannot do it and are constantly dying. The stomach constantly produces new cells to replace dead ones, which stomach acids digest. Scientists estimate that the human body completely replaces the stomach lining every seven to ten days. Figure 1.7 The pH scale measures hydrogen ions' (H+) concentration in a solution. (credit: Edward Stevens) How can organisms whose bodies require a near-neutral pH ingest acidic and basic substances (a human drinking orange juice, for example) and survive? Buffers are the key. Buffers readily absorb excess H + or OH–, keeping the body's pH carefully maintained in the narrow range required for survival. Maintaining a constant blood pH is critical to a person’s wellbeing. The buffer maintaining the pH of human blood involves carbonic acid (H 2CO3), bicarbonate ion (HCO3–), and carbon dioxide (CO2). When bicarbonate ions combine with free hydrogen ions and become carbonic acid, it removes hydrogen ions and moderates pH changes. Similarly, as Figure 1.8 shows, excess carbonic acid can convert to carbon dioxide gas which we exhale through the lungs. This prevents too many free hydrogen ions from building up in the blood and dangerously reducing the blood’s pH. Likewise, if too much OH– enters into the system, carbonic acid will combine with it to create bicarbonate, lowering the pH. Without this buffer system, the body’s pH would fluctuate enough to put survival in jeopardy. 7 Figure 1.8 This diagram shows the body’s buffering of blood pH levels. The blue arrows show the process of raising pH as more CO2 is made. The purple arrows indicate the reverse process: the lowering of pH as more bicarbonate is created. Other examples of buffers are antacids that some people use to combat excess stomach acid. Many of these over-the-counter medications work in the same way as blood buffers, usually with at least one ion capable of absorbing hydrogen and moderating pH, bringing relief to those who suffer “heartburn” after eating. Water's unique properties that contribute to this capacity to balance pH—as well as water’s other characteristics—are essential to sustaining life on Earth. 8 Lesson 3: Carbon By the end of this section, you will be able to do the following: 1. Explain why carbon is important for life 2. Describe the role of functional groups in biological molecules Many complex molecules called macromolecules, such as proteins, nucleic acids (RNA and DNA), carbohydrates, and lipids comprise cells. The macromolecules are a subset of organic molecules (any carbon-containing liquid, solid, or gas) that are especially important for life. The fundamental component for all of these macromolecules is carbon. The carbon atom has unique properties that allow it to form covalent bonds to as many as four different atoms, making this versatile element ideal to serve as the basic structural component, or “backbone,” of the macromolecules. Individual carbon atoms have an incomplete outermost electron shell. With an atomic number of 6 (six electrons and six protons), the first two electrons fill the inner shell, leaving four in the second shell. Therefore, carbon atoms can form up to four covalent bonds with other atoms to satisfy the octet rule. The methane molecule provides an example: it has the chemical formula CH4. Each of its four hydrogen atoms forms a single covalent bond with the carbon atom by sharing a pair of electrons. This results in a filled outermost shell. Hydrocarbons Hydrocarbons are organic molecules consisting entirely of carbon and hydrogen, such as methane (CH4) described above. We often use hydrocarbons in our daily lives as fuels—like the propane in a gas grill or the butane in a lighter. The many covalent bonds between the atoms in hydrocarbons store a great amount of energy, which releases when these molecules burn (oxidize). Methane, an excellent fuel, is the simplest hydrocarbon molecule, with a central carbon atom bonded to four different hydrogen atoms, as Figure 1.9 illustrates. The shape of its electron orbitals determines the shape of the methane molecule's geometry, where the atoms reside in three dimensions. The carbons and the four hydrogen atoms form a tetrahedron, with four triangular faces. For this reason, we describe methane as having tetrahedral geometry. Figure 1.9 Methane has a tetrahedral geometry, with each of the four hydrogen atoms spaced 109.5° apart. As the backbone of the large molecules of living things, hydrocarbons may exist as linear carbon chains, carbon rings, or combinations of both. Furthermore, individual carbon-tocarbon bonds may be single, double, or triple covalent bonds, and each type of bond affects the 9 molecule's geometry in a specific way. This three-dimensional shape or conformation of the large molecules of life (macromolecules) is critical to how they function. Hydrocarbon Chains Successive bonds between carbon atoms form hydrocarbon chains. These may be branched or unbranched. Furthermore, a molecule's different geometries of single, double, and triple covalent bonds alter the overall molecule's geometry as Figure 1.10 illustrates. The hydrocarbons ethane, ethene, and ethyne serve as examples of how different carbon-to-carbon bonds affect the molecule's geometry. The names of all three molecules start with the prefix “eth-,” which is the prefix for two carbon hydrocarbons. The suffixes “-ane,” “-ene,” and “-yne” refer to the presence of single, double, or triple carbon-carbon bonds, respectively. Thus, propane, propene, and propyne follow the same pattern with three carbon molecules, butane, butene, and butyne for four carbon molecules, and so on. Double and triple bonds change the molecule's geometry: single bonds allow rotation along the bond's axis; whereas, double bonds lead to a planar configuration and triple bonds to a linear one. These geometries have a significant impact on the shape a particular molecule can assume. Figure 1.10 When carbon forms single bonds with other atoms, the shape is tetrahedral. When two carbon atoms form a double bond, the shape is planar, or flat. Single bonds, like those in ethane, are able to rotate. Double bonds, like those in ethene, cannot rotate, so the atoms on either side are locked in place. Hydrocarbon Rings So far, the hydrocarbons we have discussed have been aliphatic hydrocarbons, which consist of linear chains of carbon atoms. Another type of hydrocarbon, aromatic hydrocarbons, consists of closed rings of carbon atoms with alternating single and double bonds. We find ring structures in aliphatic hydrocarbons, sometimes with the presence of double bonds, which we can see by comparing cyclohexane's structure to benzene in Figure 1.11. Examples of biological molecules that incorporate the benzene ring include some amino acids and cholesterol and its derivatives, including the hormones estrogen and testosterone. We also find the benzene ring in the herbicide 2,4-D. Benzene is a natural component of crude oil and has been classified as a carcinogen. Some hydrocarbons have both aliphatic and aromatic portions. Beta-carotene is an example of such a hydrocarbon. 10 Figure 1.11 Carbon can form five- and six-membered rings. Single or double bonds may connect the carbons in the ring, and nitrogen may be substituted for carbon. Isomers The three-dimensional placement of atoms and chemical bonds within organic molecules is central to understanding their chemistry. We call molecules that share the same chemical formula but differ in the placement (structure) of their atoms and/or chemical bonds isomers. Structural isomers (like butane and isobutane in Figure 1.12a) differ in the placement of their covalent bonds: both molecules have four carbons and ten hydrogens (C4H10), but the different atom arrangement within the molecules leads to differences in their chemical properties. For example, butane is suited for use as a fuel for cigarette lighters and torches; whereas, isobutane is suited for use as a refrigerant and a propellant in spray cans. Geometric isomers, alternatively have similar placements of their covalent bonds but differ in how these bonds are made to the surrounding atoms, especially in carbon-to-carbon double bonds. In the simple molecule butene (C 4H8), the two methyl groups (CH3) can be on either side of the double covalent bond central to the molecule, as Figure 1.12b illustrates. When the carbons are bound on the same side of the double bond, this is the cis configuration. If they are on opposite sides of the double bond, it is a trans configuration. In the trans configuration, the carbons form a more or less linear structure; whereas, the carbons in the cis configuration make a bend (change in direction) of the carbon backbone. 11 Figure 1.12 We call molecules that have the same number and type of atoms arranged differently isomers. (a) Structural isomers have a different covalent arrangement of atoms. (b) Geometric isomers have a different arrangement of atoms around a double bond. (c) Enantiomers are mirror images of each other. In triglycerides (fats and oils), long carbon chains known as fatty acids may contain double bonds, which can be in either the cis or trans configuration, as Figure 1.13 illustrates. Fats with at least one double bond between carbon atoms are unsaturated fats. When some of these bonds are in the cis configuration, the resulting bend in the chain's carbon backbone means that triglyceride molecules cannot pack tightly, so they remain liquid (oil) at room temperature. Alternatively, triglycerides with trans double bonds (popularly called trans fats), have relatively linear fatty acids that are able to pack tightly together at room temperature and form solid fats. In the human diet, trans fats are linked to an increased risk of cardiovascular disease, so many food manufacturers have reduced or eliminated their use in recent years. In contrast to unsaturated fats, we call triglycerides without double bonds between carbon atoms saturated fats, meaning that they contain all the hydrogen atoms available. Saturated fats are a solid at room temperature and usually of animal origin. Figure 1.13 These space-filling models show a cis (oleic acid) and a trans (eliadic acid) fatty acid. Notice the bend in the molecule caused by the cis configuration. 12 Enantiomers These are molecules that share the same chemical structure and chemical bonds but differ in the three-dimensional placement of atoms so that they are non-superimposable mirror images. Figure 1.14 shows an amino acid alanine example, where the two structures are nonsuperimposable. In nature, the L-forms of amino acids are predominant in proteins. Some D forms of amino acids are seen in the cell walls of bacteria and polypeptides in other organisms. Similarly, the D-form of glucose is the main product of photosynthesis and we rarely see the molecule's L-form in nature. Figure 1.14 D-alanine and L-alanine are examples of enantiomers or mirror images. L-forms of amino acids are predominant in proteins. 13 Functional Groups These are groups of atoms that occur within molecules and confer specific chemical properties to those molecules. We find them along the “carbon backbone” of macromolecules. Chains and/or rings of carbon atoms with the occasional substitution of an element such as nitrogen or oxygen form this carbon backbone. Molecules with other elements in their carbon backbone are substituted hydrocarbons. The functional groups in a macromolecule are usually attached to the carbon backbone at one or several different places along its chain and/or ring structure. Each of the four types of macromolecules—proteins, lipids, carbohydrates, and nucleic acids—has its own characteristic set of functional groups that contributes greatly to its differing chemical properties and its function in living organisms. A functional group can participate in specific chemical reactions. Figure 1.15 shows some of the important functional groups in biological molecules. They include: hydroxyl, methyl, carbonyl, carboxyl, amino, phosphate, and sulfhydryl. These groups play an important role in forming molecules like DNA, proteins, carbohydrates, and lipids. We usually classify functional groups as hydrophobic or hydrophilic depending on their charge or polarity characteristics. An example of a hydrophobic group is the nonpolar methyl molecule. Among the hydrophilic functional groups is the carboxyl group in amino acids, some amino acid side chains, and the fatty acids that form triglycerides and phospholipids. This carboxyl group ionizes to release hydrogen ions (H+) from the COOH group resulting in the negatively charged COO - group. This contributes to the hydrophilic nature of whatever molecule on which it is found. Other functional groups, such as the carbonyl group, have a partially negatively charged oxygen atom that may form hydrogen bonds with water molecules, again making the molecule more hydrophilic. 14 Figure 1.15 These functional groups are in many different biological molecules. R, also known as R-group, is an abbreviation for any group in which a carbon or hydrogen atom is attached to the rest of the molecule. Hydrogen bonds between functional groups (within the same molecule or between different molecules) are important to the function of many macromolecules and help them to fold properly into and maintain the appropriate shape for functioning. Hydrogen bonds are also involved in various recognition processes, such as DNA complementary base pairing and the binding of an enzyme to its substrate, as Figure 1.16 illustrates. Figure 1.16 Hydrogen bonds connect two strands of DNA together to create the double-helix structure. 15 UNIT SUMMARY Water Water has many properties that are critical to maintaining life. It is a polar molecule, allowing for forming hydrogen bonds. Hydrogen bonds allow ions and other polar molecules to dissolve in water. Therefore, water is an excellent solvent. The hydrogen bonds between water molecules cause the water to have a high heat capacity, meaning it takes considerable added heat to raise its temperature. As the temperature rises, the hydrogen bonds between water continually break and form anew. This allows for the overall temperature to remain stable, although energy is added to the system. Water also exhibits a high heat of vaporization, which is key to how organisms cool themselves by evaporating sweat. Water’s cohesive forces allow for the property of surface tension; whereas, we see its adhesive properties as water rises inside capillary tubes. The pH value is a measure of hydrogen ion concentration in a solution and is one of many chemical characteristics that is highly regulated in living organisms through homeostasis. Acids and bases can change pH values, but buffers tend to moderate the changes they cause. These properties of water are intimately connected to the biochemical and physical processes performed by living organisms, and life would be very different if these properties were altered, if it could exist at all. Carbon The unique properties of carbon make it a central part of biological molecules. Carbon binds to oxygen, hydrogen, and nitrogen covalently to form the many molecules important for cellular function. Carbon has four electrons in its outermost shell and can form four bonds. Carbon and hydrogen can form hydrocarbon chains or rings. Functional groups are groups of atoms that confer specific properties to hydrocarbon (or substituted hydrocarbon) chains or rings that define their overall chemical characteristics and function. 16 UNIT’S CRITICAL THINKING QUESTIONS: Write your answers in a clean whole yellow paper. Write your name, section and unit number. 1. Discuss how buffers help prevent drastic swings in pH. 2. Why can some insects walk on water? 3. What property of carbon makes it essential for organic life? 4. Compare and contrast saturated and unsaturated triglycerides. 17 UNIT II. MACROMOLECULES Introduction Think back to what you ate for lunch. Did any of your lunch items have a “Nutrition Facts” label on the back of them? If so, and if you had a look at the food's protein, carbohydrate, or fat content, you may already be familiar with several types of large biological molecules we’ll discuss here. If you’re wondering what something as weird-sounding as a “large biological molecule” is doing in your food, the answer is that it’s providing you with the building blocks you need to maintain your body – because your body is also made of large biological molecules! Just as you can be thought of as an assortment of atoms or a walking, talking bag of water(Opens in a new window), you can also be viewed as a collection of four major types of large biological molecules: carbohydrates (such as sugars), lipids (such as fats), proteins, and nucleic acids (such as DNA and RNA). That’s not to say that these are the only molecules in your body, but rather, that your most important large molecules can be divided into these groups. Together, the four groups of large biological molecules make up the majority of the dry weight of a cell. (Water, a small molecule, makes up the majority of the wet weight). Large biological molecules perform a wide range of jobs in an organism. Some carbohydrates store fuel for future energy needs, and some lipids are key structural components of cell membranes. Nucleic acids store and transfer hereditary information, much of which provides instructions for making proteins. Proteins themselves have perhaps the broadest range of functions: some provide structural support, but many are like little machines that carry out specific jobs in a cell, such as catalyzing metabolic reactions or receiving and transmitting signals. Carbohydrates, nucleic acids, and proteins are often found as long polymers in nature. Because of their polymeric nature and their large (sometimes huge!) size, they are classified as macromolecules, big (macro-) molecules made through the joining of smaller subunits. Lipids are not usually polymers and are smaller than the other three, so they are not considered macromolecules by some sources. However, many other sources use the term “macromolecule” more loosely, as a general name for the four types of large biological molecules. This is just a naming difference, so don’t get too hung up on it. Just remember that lipids are one of the four main types of large biological molecules, but that they don’t generally form polymers. 18 Lesson 1: Carbohydrates By the end of this section, you will be able to do the following: 1. Discuss the role of carbohydrates in cells and in the extracellular materials of animals and plants 2. Explain carbohydrate classifications 3. List common monosaccharides, disaccharides, and polysaccharides What’s in a potato? Besides water, which makes up most of the potato’s weight, there’s a little fat, a little protein…and a whole lot of carbohydrate (about 37 grams in a medium potato). Some of that carbohydrate is in the form of sugars. These provide the potato, and the person eating the potato, with a ready fuel source. A bit more of the potato's carbohydrate is in the form of fiber, including cellulose polymers that give structure to the potato’s cell walls. Most of the carbohydrate, though, is in the form of starch, long chains of linked glucose molecules that are a storage form of fuel. When you eat French fries, potato chips, or a baked potato with all the fixings, enzymes in your digestive tract get to work on the long glucose chains, breaking them down into smaller sugars that your cells can use. Carbohydrates make up a group of chemical compounds found in plant and animal cells. They have the empirical formula C nH2nOn, or (CH2O)n. An empirical formula tells the atomic composition of the compound, but nothing about structure, size, or what chemical bonds are present. Since this formula is essentially a combination of carbon and water, these materials are called “hydrates of carbon”, or carbohydrates for short. Carbohydrates are widely available and inexpensive, and are used as an energy source for our bodies and for cell structures. Food carbohydrates include the simple carbohydrates (sugars) and complex carbohydrates (starches and fiber). Before a big race, distance runners and cyclists eat foods containing complex carbohydrates (pasta, pizza, rice and bread) to give them sustained energy. Carbohydrates are divided into monosaccharides, disaccharides, and polysaccharides. Most of the oxygen atoms in monosaccharides are found in hydroxyl (OH) groups, but one of them is part of a carbonyl (C=O) group. The position of the carbonyl (C=O) group can be used to categorize the sugars: • • If the sugar has an aldehyde group, meaning that the carbonyl C is the last one in the chain, it is known as an aldose. If the carbonyl C is internal to the chain, so that there are other carbons on both sides of it, it forms a ketone group and the sugar is called a ketose. Sugars are also named according to their number of carbons: some of the most common types are trioses (three carbons), pentoses (five carbons), and hexoses (six carbons). 19 Figure 2.1. Different types of Carbohydrates depending of their number of carbons Glucose and its isomers The chemical formula for glucose is C6H12O6. In humans, glucose is an important source of energy. During cellular respiration, energy releases from glucose, and that energy helps make adenosine triphosphate (ATP). Plants synthesize glucose using carbon dioxide and water, and glucose in turn provides energy requirements for the plant. Humans and other animals that feed on plants often store excess glucose as catabolized (cell breakdown of larger molecules) starch. Galactose (part of lactose, or milk sugar) and fructose (found in sucrose, in fruit) are other common monosaccharides. Although glucose, galactose, and fructose all have the same chemical formula (C6H12O6), they differ structurally and chemically (and are isomers) because of the different arrangement of functional groups around the asymmetric carbon. All these monosaccharides have more than one asymmetric carbon (Figure 2.2). Figure 2.2 Glucose, galactose, and fructose are all hexoses. 20 Glucose and galactose are stereoisomers of each other: their atoms are bonded together in the same order, but they have a different 3D organization of atoms around one of their asymmetric carbons. You can see this in the diagram as a switch in the orientation of the hydroxyl (OH) group, marked in red. This small difference is enough for enzymes to tell glucose and galactose apart, picking just one of the sugars to take part in chemical reactions. Glucose, galactose, and fructose are isomeric monosaccharides (hexoses), meaning they have the same chemical formula but have slightly different structures. Glucose and galactose are aldoses, and fructose is a ketose. Ring forms of sugars of Monosaccharides You may have noticed that the sugars we’ve looked at so far are linear molecules (straight chains). That may seem odd because sugars are often drawn as rings. As it turns out both are correct: many five- and six-carbon sugars can exist either as a linear chain or in one or more ring-shaped forms. Monosaccharides can exist as a linear chain or as ring-shaped molecules. In aqueous solutions they are usually in ring forms (Figure 2.3). Glucose in a ring form can have two different hydroxyl group arrangements (OH) around the anomeric carbon (carbon 1 that becomes asymmetric in the ring formation process). If the hydroxyl group is below carbon number 1 in the sugar, it is in the alpha (α) position, and if it is above the plane, it is in the beta (β) position. Figure 2.3 Five and six carbon monosaccharides exist in equilibrium between linear and ring forms. When the ring forms, the side chain it closes on locks into an α or β position. Fructose and ribose also form rings, although they form five-membered rings as opposed to the six-membered ring of glucose. 21 Disaccharides Disaccharides (di- = “two”) form when two monosaccharides join together via a dehydration reaction, also known as a condensation reaction or dehydration synthesis. In this process, the hydroxyl group of one monosaccharide combines with the hydrogen of another, releasing a molecule of water and forming a covalent bond known as a glycosidic linkage. For instance, the diagram below shows glucose and fructose monomers combining via a dehydration reaction to form sucrose, a disaccharide we know as table sugar. (The reaction also releases a water molecule, not pictured.) In some cases, it’s important to know which carbons on the two sugar rings are connected by a glycosidic bond. Each carbon atom in a monosaccharide is given a number, starting with the terminal carbon closest to the carbonyl group (when the sugar is in its linear form). This numbering is shown for glucose and fructose, above. In a sucrose molecule, the 1 carbon of glucose is connected to the 2 carbon of fructose, so this bond is called a 1-2 glycosidic linkage. Common disaccharides include lactose, maltose, and sucrose. Lactose is a disaccharide consisting of glucose and galactose and is found naturally in milk. Many people can't digest lactose as adults, resulting in lactose intolerance (which you or your friends may be all too familiar with). Maltose, or malt sugar, is a disaccharide made up of two glucose molecules. The most common disaccharide is sucrose (table sugar), which is made of glucose and fructose (Figure 2.4). Figure 2.4 Common disaccharides include maltose (grain sugar), lactose (milk sugar), and sucrose (table sugar). 22 Polysaccharides A long chain of monosaccharides linked by glycosidic bonds is known as a polysaccharide (poly- = “many”). The chain may be branched or unbranched and may contain different types of monosaccharides. Plants store starch in the form of sugars. In plants, an amylose and amylopectic mixture (both glucose polymers) comprise these sugars. Plants are able to synthesize glucose, and they store the excess glucose, beyond their immediate energy needs, as starch in different plant parts, including roots and seeds. The starch in the seeds provides food for the embryo as it germinates and can also act as a food source for humans and animals. Enzymes break down the starch that humans consume. For example, an amylase present in saliva catalyzes, or breaks down this starch into smaller molecules, such as maltose and glucose. The cells can then absorb the glucose. Storage polysaccharides Starch is the stored form of sugars in plants and is made up of a mixture of two polysaccharides, amylose and amylopectin (both polymers of glucose). Plants are able to synthesize glucose using light energy gathered in photosynthesis, and the excess glucose, beyond the plant’s immediate energy needs, is stored as starch in different plant parts, including roots and seeds. The starch in the seeds provides food for the embryo as it germinates and can also serve as a food source for humans and animals, who will break it down into glucose monomers using digestive enzymes. In starch, the glucose monomers are in the α form (with the hydroxyl group of carbon 1 sticking down below the ring), and they are connected primarily by 14 glycosidic linkages (i.e., linkages in which carbon atoms 1 and 4 of the two monomers form a glycosidic bond). • • Figure 2.5 Amylose and amylopectin are two different starch forms. Unbranched glucose monomer chains comprise amylose by α 1-4 glycosidic linkages. Unbranched glucose monomer chains comprise amylopectin by α 1-4 and α 1-6 glycosidic linkages. Amylose consists entirely of unbranched chains of glucose monomers connected by 1-4 linkages. Amylopectin is a branched polysaccharide. Although most of its monomers are connected by 1-4 linkages, additional 1-6 linkages occur periodically and result in branch points. Because of the way the subunits are joined, the glucose chains in amylose and amylopectin typically have a helical structure. That’s great for plants, but what about us, humans? Glycogen is the storage form of glucose in humans and other vertebrates. Like starch, glycogen is a polymer of glucose monomers, and it is even more highly branched than amylopectin. 23 Glycogen is usually stored in liver and muscle cells. Whenever blood glucose levels decrease, glycogen is broken down via hydrolysis to release glucose monomers that cells can absorb and use. Structural polysaccharides Although energy storage is one important role for polysaccharides, they are also crucial for another purpose: providing structure. Cellulose, for example, is a major component of plant cell walls, which are rigid structures that enclose the cells (and help make lettuce and other veggies crunchy). Wood and paper are mostly made of cellulose, and cellulose itself is made up of unbranched chains of glucose monomers linked by 1-4 glycosidic bonds. Unlike amylose, cellulose is made of glucose monomers in their β form, and this gives it very different properties. As shown in the figure, every other glucose monomer in the chain is flipped over in relation to its neighbors, and this results in long, straight, non-helical chains of cellulose. These chains cluster together to form parallel bundles that are held together by hydrogen bonds between hydroxyl groups. This gives cellulose its rigidity and high tensile strength, which are important to plant cells. The β glycosidic linkages in cellulose can't be broken by human digestive enzymes, so humans are not able to digest cellulose. (That’s not to say that cellulose isn’t found in our diets, it just passes through us as undigested, insoluble fiber.) However, some herbivores, such as cows, koalas, buffalos, and horses, have specialized microbes that help them process cellulose. These microbes live in the digestive tract and break cellulose down into glucose monomers that can be used by the animal. Wood-chewing termites also break down cellulose with the help of microorganisms that live in their guts. Figure 2.6 In cellulose, glucose monomers are linked in unbranched chains by β 1-4 glycosidic linkages. Because of the way the glucose subunits are joined, every glucose monomer is flipped relative to the next one resulting in a linear, fibrous structure. Cellulose is specific to plants, but polysaccharides also play an important structural role in non-plant species. For instance, arthropods (such as insects and crustaceans) have a hard external skeleton, called the exoskeleton, which protects their softer internal body parts. This exoskeleton is made of the macromolecule chitin, which resembles cellulose but is made out of modified glucose units that bear a nitrogen-containing functional group. Chitin is also a major component of the cell walls of fungi, which are neither animals nor plants but form a kingdom of their own. 24 CAREER CONNECTION______________________________________________________________________ Registered Dietitian Obesity is a worldwide health concern, and many diseases such as diabetes and heart disease are becoming more prevalent because of obesity. This is one of the reasons why people increasingly seek out registered dietitians for advice. Registered dietitians help plan nutrition programs for individuals in various settings. They often work with patients in health care facilities, designing nutrition plans to treat and prevent diseases. For example, dietitians may teach a patient with diabetes how to manage blood sugar levels by eating the correct types and amounts of carbohydrates. Dietitians may also work in nursing homes, schools, and private practices. To become a registered dietitian, one needs to earn at least a bachelor’s degree in dietetics and nutrition. In addition, registered dietitians must complete a supervised internship program and pass a national exam. Those who pursue careers in dietetics take courses in nutrition, chemistry, biochemistry, biology, microbiology, and human physiology. Dietitians must become experts in the chemistry and physiology (biological functions) of food (proteins, carbohydrates, and fats). Benefits of Carbohydrates Are carbohydrates good for you? Some people believe that carbohydrates are bad and they should avoid them. Some diets completely forbid carbohydrate consumption, claiming that a low-carbohydrate diet helps people to lose weight faster. However, carbohydrates have been an important part of the human diet for thousands of years. Artifacts from ancient civilizations show the presence of wheat, rice, and corn in our ancestors’ storage areas. As part of a well-balanced diet, we should supplement carbohydrates with proteins, vitamins, and fats. Calorie-wise, a gram of carbohydrate provides 4.3 Kcal. For comparison, fats provide 9 Kcal/g, a less desirable ratio. Carbohydrates contain soluble and insoluble elements. The insoluble part, fiber, is mostly cellulose. Fiber has many uses. It promotes regular bowel movement by adding bulk, and it regulates the blood glucose consumption rate. Fiber also helps to remove excess cholesterol from the body. Fiber binds to the cholesterol in the small intestine, then attaches to the cholesterol and prevents the cholesterol particles from entering the bloodstream. Cholesterol then exits the body via the feces. Fiber-rich diets also have a protective role in reducing the occurrence of colon cancer. In addition, a meal containing whole grains and vegetables gives a feeling of fullness. As an immediate source of energy, glucose breaks down during the cellular respiration process, which produces ATP, the cell's energy currency. Without consuming carbohydrates, we reduce the availability of “instant energy”. Eliminating carbohydrates from the diet may be necessary for some people, but such a step may not be healthy for everyone. 25 PRACTICE TEST Write your answers in ¼ sheet of yellow paper. Write your name, section, unit number and lesson number. Multiple Choice. 1. Carbohydrates are able to undergo a reaction known as hydrolysis. Which of the following correctly describes hydrolysis? a. Monomers are linked together by the removal of one or more water molecules. b. Long carbohydrates are broken into monomers through the addition of water molecules between the subunits. c. Hydrogen atoms are added to the each end of the carbohydrate to protect it from degradation. d. Hydrogen atoms are removed from the end of a carbohydrate. 2. The structure of sucrose is shown below. What type of carbohydrate is sucrose? a. Trisaccharide b. Disaccharide c. Monosaccharide d. Polysaccharide 3. Which of the following statements is true of the carbohydrate glucose? Choose 1 answer: a. Glucose is a structural isomer of galactose. b. Glucose is a polysaccharide. c. Glucose is a pentose sugar. d. Glucose contains carbon, hydrogen, and oxygen atoms. 26 Lesson 2: Lipids By the end of this section, you will be able to do the following: 1. Describe the four major types of lipids 2. Explain the role of fats in storing energy 3. Differentiate between saturated and unsaturated fatty acids 4. Describe phospholipids and their role in cells 5. Define the basic structure of a steroid and some steroid functions 6. Explain how cholesterol helps maintain the plasma membrane's fluid nature We sometimes talk about fat as if it were a malevolent substance bent on our dietary destruction. In reality, fats are elegant little molecules, each one made of three long hydrocarbon tails attached to a little coathanger-like molecule called glycerol. Like the other large biological molecules, they play essential roles in the biology of humans and other organisms. (Also, many recent dietary studies see sugar as causing a lot more health problems than fat!) Fats are just one type of lipid, a category of molecules united by their inability to mix well with water. Lipids tend to be hydrophobic, nonpolar, and made up mostly of hydrocarbon chains, though there are some variations on this, which we'll explore below. The different varieties of lipids have different structures, and correspondingly diverse roles in organisms. For instance, lipids store energy, provide insulation, make up cell membranes, form water-repellent layers on leaves, and provide building blocks for hormones like testosterone. Here, we’ll look in greater detail at some of the most important types of lipids, including fats and oils, waxes, phospholipids, and steroids. Fats and oils A fat molecule consists of two kinds of parts: a glycerol backbone and three fatty acid tails. Glycerol is a small organic molecule with three hydroxyl (OH) groups, while a fatty acid consists of a long hydrocarbon chain attached to a carboxyl group. A typical fatty acid contains 12–18 carbons, though some may have as few as 4 or as many as 36. To make a fat molecule, the hydroxyl groups on the glycerol backbone react with the carboxyl groups of fatty acids in a dehydration synthesis reaction. This yields a fat molecule with three fatty acid tails bound to the glycerol backbone via ester linkages (linkages containing an oxygen atom next to a carbonyl, or C=O, group). Triglycerides may contain three identical fatty acid tails, or three different fatty acid tails (with different lengths or patterns of double bonds) (Figure 2.7). 27 Figure 2.7 Joining three fatty acids to a glycerol backbone in a dehydration reaction forms triacylglycerol. Three water molecules release in the process. Fat molecules are also called triacylglycerols, or, in bloodwork done by your doctor, triglycerides. In the human body, triglycerides are primarily stored in specialized fat cells, called adipocytes, which make up a tissue known as adipose tissue. While many fatty acids are found in fat molecules, some are also free in the body, and they are considered a type of lipid in their own right. Some fatty acids have common names that specify their origin. For example, palmitic acid, a saturated fatty acid, is derived from the palm tree. Arachidic acid is derived from Arachis hypogea, the scientific name for groundnuts or peanuts. Saturated and Unsaturated Fatty Acids As shown in the example above (Figure 2.7), the three fatty acid tails of a triglyceride need not be identical to each other. Fatty acid chains may differ in length, as well as in their degree of unsaturation. • • If there are only single bonds between neighboring carbons in the hydrocarbon chain, a fatty acid is said to be saturated. (The thing that fatty acids are saturated with is hydrogen; in a saturated fat, as many hydrogen atoms as possible are attached to the carbon skeleton.) When the hydrocarbon chain has a double bond, the fatty acid is said to be unsaturated, as it now has fewer hydrogens. If there is just one Figure 2.8 Saturated fatty acids have double bond in a fatty acid, it’s hydrocarbon chains connected by single bonds only. Unsaturated fatty acids have one or more monounsaturated, while if there are multiple double bonds. Each double bond may be in double bonds, it’s polyunsaturated. The double bonds in unsaturated fatty acids, like other types of double bonds, can exist in either a cis or a trans configuration. In the cis configuration, the two hydrogens associated with the bond are on the a cis or trans configuration. In the cis configuration, both hydrogens are on the same side of the hydrocarbon chain. In the trans configuration, the hydrogens are on opposite sides. A cis double bond causes a kink in the chain. 28 same side, while in a trans configuration, they are on opposite sides. A cis double bond generates a kink or bend in the fatty acid, a feature that has important consequences for the behavior of fats. Saturated fatty acids tails are straight, so fat molecules with fully saturated tails can pack tightly against one another. This tight packing results in fats that are solid at room temperature (have a relatively high melting point). For instance, most of the fat in butter is saturated fat. In contrast, cis-unsaturated fatty acid tails are bent due to the cis double bond. This makes it hard for fat molecules with one or more cis-unsaturated fatty acid tails to pack tightly. So, fats with unsaturated tails tend to be liquid at room temperature (have a relatively low melting point) – they are what we commonly call oils. For instance, olive oil is mostly made up of unsaturated fats. Trans fats At this point, you may be noticing that I’ve left something out: I didn’t say anything about unsaturated fats with trans double bonds in their fatty acid tails, or trans fats. Trans fats are rare in nature, but are readily produced in an industrial procedure called partial hydrogenation. In this process, hydrogen gas is passed through oils (made mostly of cis-unsaturated fats), converting some – but not all – of the double bonds to single bonds. The goal of partial hydrogenation is to give the oils some of the desirable properties of saturated fats, such as solidity at room temperature, but an unintended consequence is that some of the cis double bonds change configuration and become trans double bonds. Trans-unsaturated fatty acids can pack more tightly and are more likely to be solid at room temperature. Some types of shortening, for example, contain a high fraction of trans fats. Partial hydrogenation and trans fats might seem like a good way to get a butter-like substance at oil-like prices. Margarine, some types of peanut butter, and shortening are examples of artificially hydrogenated trans fats. Recent studies have shown that an increase in trans fats in the human diet may lead to higher levels of low-density lipoproteins (LDL), or “bad” cholesterol, which in turn may lead to plaque deposition in the arteries, resulting in heart disease. Many fast food restaurants have recently banned using trans fats, and food labels are required to display the trans fat content. Omega fatty acids Another class of fatty acids that deserves mention includes the omega-3 and omega-6 fatty acids. There are different types of omega-3 and omega-6 fatty acids, but all of them are made from two basic precursor forms: alpha-linolenic acid (ALA) for omega-3s and linoleic acid (LA) for omega-6s. The human body needs these molecules (and their derivatives), but can't synthesize either ALA or LA itself. Accordingly, ALA and LA are classified as essential fatty acids and must be obtained from a person’s diet. Some fish, such as salmon, and some seeds, such as chia and flax, are good sources of omega-3 fatty acids. Omega-3 and omega-6 fatty acids have at least two cis-unsaturated bonds, which gives them a curved shape. ALA, shown below, is quite bent but isn’t the most extreme example – 29 DHA, an omega-3 fatty acid made from ALA by the formation of additional double bonds, has six cis-unsaturated bonds and is curled up almost in a circle! Figure 2.9 Alpha-linolenic acid is an example of an omega3 fatty acid. It has three cis double bonds and, as a result, a curved shape. In the figure 2.9, the farthest carbon away from the carboxyl group is numbered as the omega (ω) carbon, and if the double bond is between the third and fourth carbon from that end, it is an omega-3 fatty acid. Nutritionally important because the body does not make them, omega-3 fatty acids include alpha-linoleic acid (ALA), eicosapentaenoic acid (EPA), and docosahexaenoic acid (DHA), all of which are polyunsaturated. Salmon, trout, and tuna are good sources of omega-3 fatty acids. Research indicates that omega-3 fatty acids reduce the risk of sudden death from heart attacks, lower triglycerides in the blood, decrease blood pressure, and prevent thrombosis by inhibiting blood clotting. They also reduce inflammation, and may help lower the risk of some cancers in animals. Like carbohydrates, fats have received considerable bad publicity. It is true that eating an excess of fried foods and other “fatty” foods leads to weight gain. However, fats do have important functions. Many vitamins are fat soluble, and fats serve as a long-term storage form of fatty acids: a source of energy. They also provide insulation for the body. Therefore, we should consume “healthy” fats in moderate amounts on a regular basis. Role of fats Fats have received a lot of bad publicity, and it’s true that eating large amounts of fried foods and other “fatty” foods can lead to weight gain and cause health problems. However, fats are essential to the body and have a number of important functions. For instance, many vitamins are fat-soluble, meaning that they must be associated with fat molecules in order to be effectively absorbed by the body. Fats also provide an efficient way to store energy over long time periods, since they contain over twice as much energy per gram as carbohydrates, and they additionally provide insulation for the body. Like all the other large biological molecules, fats in the right amounts are necessary to keep your body (and the bodies of other organisms) functioning correctly. 30 Waxes Waxes are another biologically important category of lipids. Wax covers the feathers of some aquatic birds and the leaf surfaces of some plants, where its hydrophobic (water-repelling) properties prevent water from sticking to, or soaking into, the surface. This is why water beads up on the leaves of many plants, and why birds don’t get soaked through when it rains (Figure 2.10). Long fatty acid chains esterified to long-chain alcohols comprise waxes. Figure 2.10 Lipids comprise waxy coverings on some leaves. (credit OpenStax Biology) Phospholipids What keeps the watery goo (cytosol) inside of your cells from spilling out? Cells are surrounded by a structure called the plasma membrane, which serves as a barrier between the inside of the cell and its surroundings. Phospholipids are major plasma membrane constituents that comprise cells' outermost layer. Like fats, they are comprised of fatty acid chains attached to a glycerol or sphingosine backbone. However, instead of three fatty acids attached as in triglycerides, there are two fatty acids forming diacylglycerol, and a modified phosphate group occupies the glycerol backbone's third carbon (Figure 2.11). A phosphate group alone attached to a diacylglycerol does not qualify as a phospholipid. It is phosphatidate (diacylglycerol 3-phosphate), the precursor of phospholipids. An alcohol modifies the phosphate group. Phosphatidylcholine and phosphatidylserine are two important phospholipids that are in plasma membranes. Figure 2.11 A phospholipid is a molecule with two fatty acids and a modified phosphate group attached to a glycerol backbone. Adding a charged or polar chemical group may modify the phosphate. 31 A phospholipid is an amphipathic molecule, meaning it has a hydrophobic and a hydrophilic part. The fatty acid chains are hydrophobic and cannot interact with water; whereas, the phosphate-containing group is hydrophilic and interacts with water (Figure 2.11). Figure 2.12 The phospholipid bilayer is the major component of all cellular membranes. The hydrophilic head groups of the phospholipids face the aqueous solution. The hydrophobic tails are sequestered in the middle of the bilayer. The head is the hydrophilic part, and the tail contains the hydrophobic fatty acids. In a membrane, a bilayer of phospholipids forms the structure's matrix, phospholipids' fatty acid tails face inside, away from water; whereas, the phosphate group faces the outside, aqueous side (Figure 2.12). Phospholipids are responsible for the plasma membrane's dynamic nature. If a drop of phospholipids is placed in water, it spontaneously forms a structure that scientists call a micelle, where the hydrophilic phosphate heads face the outside and the fatty acids face the structure's interior. Steroids Steroids are another class of lipid molecules, identifiable by their structure of four fused rings. Although they do not resemble the other lipids structurally, steroids are included in lipid category because they are also hydrophobic and insoluble in water. All steroids have four linked carbon rings and several of them, like cholesterol, also have a short tail (Figure 2.13). Many steroids also have an –OH functional group attached at a particular site, as shown for cholesterol below; such steroids are also classified as alcohols, and are thus called sterols. Cholesterol, the most common steroid, is mainly synthesized in the liver and is the precursor to many steroid hormones. These include the sex hormones testosterone and estradiol, which are secreted by the gonads (testes and ovaries). Cholesterol also serves as the starting material for other important molecules in the body, including Figure 2.13 Four fused hydrocarbon rings comprise steroids such as cholesterol and cortisol. 32 vitamin D and bile acids, which aid in the digestion and absorption of fats from dietary sources. It’s also a key component of cell membranes, altering their fluidity and dynamics. Of course, cholesterol is also found in the bloodstream, and blood levels of cholesterol are what we often hear about at the doctor’s office or in news reports. Cholesterol in the blood can have both protective effects (in its high-density, or HDL, form) and negative effects (in its low-density, or LDL, form) on cardiovascular health. 33 PRACTICE TEST Write your answers in ¼ sheet of yellow paper. Write your name, section, unit number and lesson number. Multiple Choice. 1. Both vegetable oil and butter are made up of fats. Why is butter a solid at room temperature, while vegetable oil is a liquid? a. Butter is a saturated fat and vegetable oil is an unsaturated fat. b. Butter is an unsaturated fat and vegetable oil is a saturated fat. c. Butter is a polar molecule and vegetable oil is a nonpolar molecule. d. Butter is a nonpolar molecule and vegetable oil is a polar molecule. 2. Which of the following is a characteristic of lipids? a. They are composed of nitrogenous chains. b. They are not soluble in water. c. They are either fats or oils. d. They are polar molecules. 3. How are triglycerides formed? a. Triglycerides are formed through dehydration synthesis, which results in the loss of water molecules. b. Triglycerides are formed through dehydration synthesis, which results in the addition of water molecules. c. Triglycerides are formed through hydrolysis, which results in the loss of water molecules. d. Triglycerides are formed through hydrolysis, which results in the production of water molecules. 4. Which of the following occurs when hydrogen is reacted with vegetable oil? a. The hydrogenated vegetable oil will contain fewer trans fats. b. The hydrogenated vegetable oil will become solid at room temperature. c. The hydrogenated vegetable oil will become polarized. d. The hydrogenated vegetable oil will become a saturated fat. 34 Lesson 3: Proteins By the end of this section, you will be able to do the following: 1. Describe the functions proteins perform in the cell and in tissues 2. Discuss the relationship between amino acids and proteins 3. Explain the four levels of protein organization 4. Describe the ways in which protein shape and function are linked Introduction We tend to think of protein as a mass noun: a homogeneous substance, something that your diet should contain in a certain proportion. But if you ever work in a molecular biology lab (say, for a summer internship), protein may start to look very different to you. How so? Well, you may see firsthand that protein isn’t just a single substance. Instead, there are lots and lots of different proteins in an organism, or even in a single cell. They come in every size, shape, and type you can imagine, and each one has a unique and specific job. Some are structural parts, giving cells shape or helping them move. Others act as signals, drifting between cells like messages in a bottle. Still others are metabolic enzymes, putting together or snapping apart biomolecules needed by the cell. And, odds are, one of these unique molecular players will become yours for the duration of your research! Proteins are among the most abundant organic molecules in living systems and are way more diverse in structure and function than other classes of macromolecules. A single cell can contain thousands of proteins, each with a unique function. Although their structures, like their functions, vary greatly, all proteins are made up of one or more chains of amino acids. In this article, we will look in more detail at the building blocks, structures, and roles of proteins. Types and functions of proteins Proteins can play a wide array of roles in a cell or organism. Here, we’ll touch on a few examples of common protein types that may be familiar to you, and that are important in the biology of many organisms (including us). Enzymes Enzymes act as catalysts in biochemical reactions, meaning that they speed the reactions up. Each enzyme recognizes one or more substrates, the molecules that serve as starting material for the reaction it catalyzes. Different enzymes participate in different types of reactions and may break down, link up, or rearrange their substrates. One example of an enzyme found in your body is salivary amylase, which breaks amylose (a kind of starch) down into smaller sugars. The amylose doesn’t taste very sweet, but the smaller sugars do. This is why starchy foods often taste sweeter if you chew them for longer: you’re giving salivary amylase time to get to work. 35 Hormones Hormones are long-distance chemical signals released by endocrine cells (like the cells of your pituitary gland). They control specific physiological processes, such as growth, development, metabolism, and reproduction. While some hormones are steroid-based (see the article on lipids), others are proteins. These protein-based hormones are commonly called peptide hormones. For example, insulin is an important peptide hormone that helps regulate blood glucose levels. When blood glucose rises (for instance, after you eat a meal), specialized cells in the pancreas release insulin. The insulin binds to cells in the liver and other parts of the body, causing them to take up the glucose. This process helps return blood sugar to its normal, resting level. Some additional types of proteins and their functions are listed in the table below: Protein types and functions Table 1. Different types and functions of protein. ROLE EXAMPLES FUNCTIONS Amylase, lipase, pepsin Break down nutrients in food into small pieces that can be readily absorbed Transport Hemoglobin Carry substances throughout the body in blood or lymph Structure Actin, tubulin, keratin Build different structures, like the cytoskeleton Hormone signaling Insulin, glucagon Coordinate the activity of different body systems Defense Antibodies Protect the body from foreign pathogens Contraction Myosin Carry out muscle contraction Storage Legume proteins, (albumin) Digestive enzyme egg storage white Provide food for the early development of the embryo or the seedling modified from OpenStax College, Biology. Proteins come in many different shapes and sizes. Some are globular (roughly spherical) in shape, whereas others form long, thin fibers. For example, the hemoglobin protein that carries oxygen in the blood is a globular protein, while collagen, found in our skin, is a fibrous protein. A protein’s shape is critical to its function, and many different types of chemical bonds may be important in maintaining this shape. Changes in temperature and pH, as well as the presence of certain chemicals, may disrupt a protein’s shape and cause it to lose functionality, a process known as denaturation. 36 Amino acids Amino acids are the monomers that make up proteins. Specifically, a protein is made up of one or more linear chains of amino acids, each of which is called a polypeptide. (We'll see where this name comes from a little further down the page.) There are 20 types of amino acids commonly found in proteins. Each amino acid has the same fundamental structure, which consists of a central carbon atom, or the alpha (α) carbon, bonded to an amino group (NH2), a carboxyl group (COOH), and to a hydrogen atom. Every amino acid also has another atom or group of atoms bonded to the central Figure 2.14 Amino acids have a central atom known as the R group (Figure 2.14). asymmetric carbon to which an amino group, a carboxyl group, a hydrogen atom, Scientists use the name "amino acid" because and a side chain (R group) are attached. these acids contain both amino group and carboxyl-acidgroup in their basic structure. As we mentioned, there are 20 common amino acids present in proteins. Nine of these are essential amino acids in humans because the human body cannot produce them and we obtain them from our diet. For each amino acid, the R group (or side chain) is different (Figure 2.15). Figure 2.15 There are 20 common amino acids commonly found in proteins, each with a different R group (variant group) that determines its chemical nature. The properties of the side chain determine an amino acid’s chemical behavior (that is, whether it is considered acidic, basic, polar, or nonpolar). For example, amino acids such as valine and leucine are nonpolar and hydrophobic, while amino acids like serine and glutamine have hydrophilic side chains and are polar. Some amino acids, such as lysine and arginine, have side chains that are positively charged at physiological pH and are considered basic amino 37 acids. (Histidine is sometimes put in this group too, although it is mostly deprotonated at physiological pH) Aspartate and glutamate, on the other hand, are negatively charged at physiological pH and are considered acidic. A few other amino acids have R groups with special properties, and these will prove to be important when we look at protein structure: • • Proline has an R group that’s linked back to its own amino group, forming a ring structure. This makes it an exception to the typical structure of an amino acid, since it no longer has the standard NH3+ amino group. If you think that ring structure looks a little awkward, you’re right: proline often causes bends or kinks in amino acid chains. Cysteine contains a thiol (-SH) group and can form covalent bonds with other cysteines. Just as some fatty acids are essential to a diet, some amino acids also are necessary. These essential amino acids in humans include isoleucine, leucine, and cysteine. Essential amino acids refer to those necessary to build proteins in the body, but not those that the body produces. Which amino acids are essential varies from organism to organism. The sequence and the number of amino acids ultimately determine the protein's shape, size, and function. A covalent bond, or peptide bond, attaches to each amino acid, which a dehydration reaction forms. One amino acid's carboxyl group and the incoming amino acid's amino group combine, releasing a water molecule. The resulting bond is the peptide bond (Figure 2.16). Figure 2.16 Peptide bond formation is a dehydration synthesis reaction. The carboxyl group of one amino acid is linked to the incoming amino acid's amino group. In the process, it releases a water molecule. The products that such linkages form are peptides. As more amino acids join to this growing chain, the resulting chain is a polypeptide. Each polypeptide has a free amino group at one end. This end the N terminal, or the amino terminal, and the other end has a free carboxyl group, also the C or carboxyl terminal. While the terms polypeptide and protein are sometimes used interchangeably, a polypeptide is technically a polymer of amino acids, whereas the term protein is used for a polypeptide or polypeptides that have combined together, often have bound non-peptide prosthetic groups, have a distinct shape, and have a unique function. After protein synthesis (translation), most proteins are modified. These are known as post-translational modifications. They may undergo cleavage, phosphorylation, or may require adding other chemical groups. Only after these modifications is the protein completely functional. Protein Structure Have you ever wondered why egg whites go from clear to opaque when you fry an egg? 38 Egg whites contain large amounts of proteins called albumins, and the albumins normally have a specific 3D shape, thanks to bonds formed between different amino acids in the protein. Heating causes these bonds to break and exposes hydrophobic (water-hating) amino acids usually kept on the inside of the protein. The hydrophobic amino acids, trying to get away from the water surrounding them in the egg white, will stick to one another, forming a protein network that gives the egg white structure while turning it white and opaque. Ta-da! Thank you, protein denaturation, for another delicious breakfast. As we mentioned in the last article on proteins and amino acids, the shape of a protein is very important to its function. To understand how a protein gets its final shape or conformation, we need to understand the four levels of protein structure: primary, secondary, tertiary, and quaternary. Primary structure The simplest level of protein structure, primary structure, is simply the sequence of amino acids in a polypeptide chain. For example, the hormone insulin has two polypeptide chains, A and B, shown in diagram below. (The insulin molecule shown here is cow insulin, although its structure is similar to that of human insulin.) Each chain has its own set of amino acids, assembled in a particular order. For instance, the sequence of the A chain starts with glycine at the N-terminus and ends with asparagine at the C-terminus, and is different from the sequence of the B chain (Figure 2.17). Figure 2.17 Bovine serum insulin is a protein hormone comprised of two peptide chains, A (21 amino acids long) and B (30 amino acids long). In each chain, three-letter abbreviations that represent the amino acids' names in the order they are present indicate primary structure. The amino acid cysteine (cys) has a sulfhydryl (SH) group as a side chain. Two sulfhydryl groups can react in the presence of oxygen to form a disulfide (S-S) bond. Two disulfide bonds connect the A and B chains together, and a third helps the A chain fold into the correct shape. Note that all disulfide bonds are the same length, but we have drawn them different sizes for clarity. The sequence of a protein is determined by the DNA of the gene that encodes the protein (or that encodes a portion of the protein, for multi-subunit proteins). A change in the gene's DNA sequence may lead to a change in the amino acid sequence of the protein. Even changing just one amino acid in a protein’s sequence can affect the protein’s overall structure and function. For instance, a single amino acid change is associated with sickle cell anemia, an inherited disease that affects red blood cells. In sickle cell anemia, one of the polypeptide chains that make up hemoglobin, the protein that carries oxygen in the blood, has a slight sequence 39 change. The glutamic acid that is normally the sixth amino acid of the hemoglobin β chain (one of two types of protein chains that make up hemoglobin) is replaced by a valine. This substitution is shown for a fragment of the β chain in the diagram below. Figure 2.18 The beta chain of hemoglobin is 147 residues in length, yet a single amino acid substitution leads to sickle cell anemia. In normal hemoglobin, the amino acid at position seven is glutamate. In sickle cell hemoglobin, a valine replaces glutamate. What is most remarkable to consider is that a hemoglobin molecule is made up of two α chains and two β chains, each consisting of about 150 amino acids, for a total of about 600 amino acids in the whole protein. The difference between a normal hemoglobin molecule and a sickle cell molecule is just 2 amino acids out of the approximately 600. A person whose body makes only sickle cell hemoglobin will suffer symptoms of sickle cell anemia. These occur because the glutamic acid-to-valine amino acid change makes the hemoglobin molecules assemble into long fibers. The fibers distort disc-shaped red blood cells into crescent shapes. Examples of “sickled” cells can be seen mixed with normal, disc-like cells in the blood sample below (Figure 2.19) Figure 2.19 In this blood smear, visualized at 535x magnification using bright field microscopy, sickle cells are crescent shaped, while normal cells are disc-shaped. (credit: modification of work by Ed Uthman; scale-bar data from Matt Russell) 40 The sickled cells get stuck as they try to pass through blood vessels. The stuck cells impair blood flow and can cause serious health problems for people with sickle cell anemia, including breathlessness, dizziness, headaches, and abdominal pain. Secondary structure The local folding of the polypeptide in some regions gives rise to the secondary structure of the protein. The most common are the α-helix and β-pleated sheet structures (Figure 2.20). Both structures are held in shape by hydrogen bonds. The hydrogen bonds form between the oxygen atom in the carbonyl group in one amino acid and another amino acid that is four amino acids farther along the chain. Every helical turn in an alpha helix has 3.6 amino acid residues. The polypeptide's R groups (the variant groups) protrude out from the α-helix chain. In the β- Figure 2.20 The α-helix and β-pleated sheet are secondary structures of proteins that form because of pleated sheet, hydrogen bonding between hydrogen bonding between carbonyl and amino atoms on the polypeptide chain's backbone groups in the peptide backbone. Certain amino acids form the "pleats". The R groups are attached have a propensity to form an α-helix, while others to the carbons and extend above and below have a propensity to form a β-pleated sheet. the pleat's folds. The pleated segments align parallel or antiparallel to each other, and hydrogen bonds form between the partially positive nitrogen atom in the amino group and the partially negative oxygen atom in the peptide backbone's carbonyl group. The α-helix and β-pleated sheet structures are in most globular and fibrous proteins and they play an important structural role. Tertiary Structure The overall three-dimensional structure of a polypeptide is called its tertiary structure. The tertiary structure is primarily due to interactions between the R groups of the amino acids that make up the protein (Figure 2.21). R group interactions that contribute to tertiary structure include hydrogen bonding, ionic bonding, dipole-dipole interactions, and London dispersion forces – basically, the whole gamut of non-covalent bonds. For example, R groups with like charges repel one another, while those with opposite charges can form an ionic bond. Similarly, polar R groups can form hydrogen bonds and other dipole-dipole interactions. Also important to tertiary structure are hydrophobic interactions, in which amino acids with nonpolar, hydrophobic R Figure 2.21 A variety of chemical interactions determine the proteins' tertiary structure. These include hydrophobic interactions, ionic bonding, hydrogen bonding, and disulfide linkages. 41 groups cluster together on the inside of the protein, leaving hydrophilic amino acids on the outside to interact with surrounding water molecules. Finally, there’s one special type of covalent bond that can contribute to tertiary structure: the disulfide bond. Disulfide bonds, covalent linkages between the sulfur-containing side chains of cysteines, are much stronger than the other types of bonds that contribute to tertiary structure. They act like molecular "safety pins," keeping parts of the polypeptide firmly attached to one another. Quaternary structure Many proteins are made up of a single polypeptide chain and have only three levels of structure (the ones we’ve just discussed). However, some proteins are made up of multiple polypeptide chains, also known as subunits. When these subunits come together, they give the protein its quaternary structure (Figure 2.22) We’ve already encountered one example of a protein with quaternary structure: hemoglobin. As mentioned earlier, hemoglobin carries oxygen in the blood and is made up of four subunits, two each of the α and β types. Another example is DNA polymerase, an enzyme that synthesizes new strands of DNA and is composed of ten subunits. In general, the same types of interactions that contribute to tertiary structure (mostly weak interactions, such as hydrogen bonding and London dispersion forces) also hold the subunits together to give quaternary structure. Figure 2.22 Observe the four levels of protein structure in these illustrations. (credit: modification of work by National Human Genome Research Institute) 42 Denaturation and protein folding Each protein has its own unique shape. If the temperature or pH of a protein's environment is changed, or if it is exposed to chemicals, these interactions may be disrupted, causing the protein to lose its three-dimensional structure and turn back into an unstructured string of amino acids. When a protein loses its higher-order structure, but not its primary sequence, it is said to be denatured. Denatured proteins are usually non-functional. For some proteins, denaturation can be reversed. Since the primary structure of the polypeptide is still intact (the amino acids haven’t split up), it may be able to re-fold into its functional form if it's returned to its normal environment. Other times, however, denaturation is permanent. One example of irreversible protein denaturation is when an egg is fried. The albumin protein in the liquid egg white becomes opaque and solid as it is denatured by the heat of the stove, and will not return to its original, raw-egg state even when cooled down. Researchers have found that some proteins can re-fold after denaturation even when they are alone in a test tube. Since these proteins can go from unstructured to folded all by themselves, their amino acid sequences must contain all the information needed for folding. However, not all proteins are able to pull off this trick, and how proteins normally fold in a cell appears to be more complicated. Many proteins don’t fold by themselves, but instead get assistance from chaperone proteins (chaperonins). 43 PRACTICE TEST Write your answers in ¼ sheet of yellow paper. Write your name, section, unit number and lesson number. Multiple Choice. 1. Which of the following is an example of protein denaturation? a. A protein binds with a substrate, lowering the activation energy of a reaction. b. Several amino acids are joined together via peptide bonds. c. A protein is exposed to extremely high heat, causing it to lose its secondary structure and be left with only its primary structure. d. Amino acids fold into repeating patterns due to hydrogen bonding of the peptide backbone. 2. Polypeptides are chains of amino acids linked together in a specific order. a. Peptide bonds b. Hydrogen bonds c. Glycosidic linkages d. Ester linkage 3. Which of the following are functions of proteins? Choose 2 answers. a. Cell signaling b. Energy storage c. Encoding genetic information d. Catalyzing chemical reactions 4. The conformation, or shape, of a protein determines its function. There are four orders of protein structure: primary, secondary, tertiary, and quaternary. Which of the following statements is accurate regarding these protein structures? a. Secondary structures are formed by multiple polypeptide chains. b. Interactions between the R groups in amino acids form tertiary structure. c. The two types of primary structure are α helices and β pleated sheets. d. Proteins in a quaternary structure consist of a simple polypeptide chain. 44 Lesson 4: Nucleic Acids By the end of this section, you will be able to do the following: 1. Describe nucleic acids' structure and define the two types of nucleic acids 2. Explain DNA's structure and role 3. Explain RNA's structure and roles Introduction Nucleic acids, and DNA in particular, are key macromolecules for the continuity of life. DNA bears the hereditary information that’s passed on from parents to children, providing instructions for how (and when) to make the many proteins needed to build and maintain functioning cells, tissues, and organisms. Here, we’ll just take a quick look at nucleic acids from the macromolecule perspective. Roles of DNA and RNA in cells Nucleic acids, macromolecules made out of units called nucleotides, come in two naturally occurring varieties: deoxyribonucleic acid (DNA) and ribonucleic acid (RNA). DNA is the genetic material found in living organisms, all the way from single-celled bacteria to multicellular mammals like you and me. Some viruses use RNA, not DNA, as their genetic material, but aren’t technically considered to be alive (since they cannot reproduce without help from a host). DNA in cells In eukaryotes, such as plants and animals, DNA is found in the nucleus, a specialized, membrane-bound vault in the cell, as well as in certain other types of organelles (such as mitochondria and the chloroplasts of plants). In prokaryotes, such as bacteria, the DNA is not enclosed in a membranous envelope, although it's located in a specialized cell region called the nucleoid. In eukaryotes, DNA is typically broken up into a number of very long, linear pieces called chromosomes, while in prokaryotes such as bacteria, chromosomes are much smaller and often circular (ring-shaped). A chromosome may contain tens of thousands of genes, each providing instructions on how to make a particular product needed by the cell. From DNA to RNA to proteins Many genes encode protein products, meaning that they specify the sequence of amino acids used to build a particular protein. Before this information can be used for protein synthesis, however, an RNA copy (transcript) of the gene must first be made. This type of RNA is called a messenger RNA (mRNA), as it serves as a messenger between DNA and the ribosomes, molecular machines that read mRNA sequences and use them to build proteins. This progression from DNA to RNA to protein is called the “central dogma” of molecular biology. Importantly, not all genes encode protein products. For instance, some genes specify ribosomal RNAs (rRNAs), which serve as structural components of ribosomes, or transfer RNAs (tRNAs), cloverleaf-shaped RNA molecules that bring amino acids to the ribosome for protein synthesis. Still other RNA molecules, such as tiny microRNAs (miRNAs), 45 act as regulators of other genes, and new types of non-protein-coding RNAs are being discovered all the time. Nucleotides DNA and RNA are polymers (in the case of DNA, often very long polymers), and are made up of monomers known as nucleotides. When these monomers combine, the resulting chain is called a polynucleotide (poly- = "many"). Each nucleotide is made up of three parts: a nitrogen-containing ring structure called a nitrogenous base, a five-carbon sugar, and at least one phosphate group. The sugar molecule has a central position in the nucleotide, with the base attached to one of its carbons and the phosphate group (or groups) attached to another. Let’s look at each part of a nucleotide in turn (Figure 2.23). Figure 2.23 Three components comprise a nucleotide: a nitrogenous base, a pentose sugar, and one or more phosphate groups. Nitrogenous bases The nitrogenous bases of nucleotides are organic (carbon-based) molecules made up of nitrogen-containing ring structures. Each nucleotide in DNA contains one of four possible nitrogenous bases: adenine (A), guanine (G) cytosine (C), and thymine (T). Adenine and guanine are purines, meaning that their structures contain two fused carbon-nitrogen rings. Cytosine and thymine, in contrast, are pyrimidines and have a single carbon-nitrogen ring. RNA nucleotides may also bear adenine, guanine and cytosine bases, but instead of thymine they have another pyrimidine base called uracil (U). As shown in the figure above, each base has a unique structure, with its own set of functional groups attached to the ring structure. 46 In molecular biology shorthand, the nitrogenous bases are often just referred to by their one-letter symbols, A, T, G, C, and U. DNA contains A, T, G, and C, while RNA contains A, U, G, and C (that is, U is swapped in for T). Sugars In addition to having slightly different sets of bases, DNA and RNA nucleotides also have slightly different sugars. The five-carbon sugar in DNA is called deoxyribose, while in RNA, the sugar is ribose. These two are very similar in structure, with just one difference: the second carbon of ribose bears a hydroxyl group, while the equivalent carbon of deoxyribose has a hydrogen instead. The carbon atoms of a nucleotide’s sugar molecule are numbered as 1′, 2′, 3′, 4′, and 5′ (1′ is read as “one prime”), as shown in the figure above. In a nucleotide, the sugar occupies a central position, with the base attached to its 1′ carbon and the phosphate group (or groups) attached to its 5′ carbon. Phosphate Nucleotides may have a single phosphate group, or a chain of up to three phosphate groups, attached to the 5’ carbon of the sugar. Some chemistry sources use the term “nucleotide” only for the single-phosphate case, but in molecular biology, the broader definition is generally accepted. In a cell, a nucleotide about to be added to the end of a polynucleotide chain will bear a series of three phosphate groups. When the nucleotide joins the growing DNA or RNA chain, it loses two phosphate groups. So, in a chain of DNA or RNA, each nucleotide has just one phosphate group. Polynucleotide chains A consequence of the structure of nucleotides is that a polynucleotide chain has directionality – that is, it has two ends that are different from each other. At the 5’ end, or beginning, of the chain, the 5’ phosphate group of the first nucleotide in the chain sticks out. At the other end, called the 3’ end, the 3’ hydroxyl of the last nucleotide added to the chain is exposed. DNA sequences are usually written in the 5' to 3' direction, meaning that the nucleotide at the 5' end comes first and the nucleotide at the 3' end comes last. As new nucleotides are added to a strand of DNA or RNA, the strand grows at its 3’ end, with the 5′ phosphate of an incoming nucleotide attaching to the hydroxyl group at the 3’ end of the chain. This makes a chain with each sugar joined to its neighbors by a set of bonds called a phosphodiester linkage. Properties of DNA Deoxyribonucleic acid, or DNA, chains are typically found in a double helix, a structure in which two matching (complementary) chains are stuck together, as shown in the Figure 2.24. The sugars and phosphates lie on the outside of the helix, forming the backbone of the DNA; this portion of the molecule is sometimes called the sugar-phosphate backbone. The nitrogenous bases extend into the interior, like the steps of a staircase, in pairs; the bases of a pair are bound to each other by hydrogen bonds. The two strands of the helix run in opposite directions, meaning that the 5′ end of one strand is paired 47 up with the 3′ end of its matching strand. (This is referred to as antiparallel orientation and is important for the copying of DNA.) So, can any two bases decide to get together and form a pair in the double helix? The answer is a definite no. Because of the sizes and functional groups of the bases, base pairing is highly specific: A can only pair with T, and G can only pair with C, as shown below. This means that the two strands of a DNA double helix have a very predictable relationship to each other. For instance, if you know that the sequence of one strand is 5’-AATTGGCC-3’, the complementary strand must have the sequence 3’-TTAACCGG-5’. This allows each base to match up with its partner: Figure 2.24 Native DNA is an antiparallel double helix. The phosphate backbone (indicated by the curvy lines) is on the outside, and the bases are on the inside. Each base from one strand interacts via hydrogen bonding with a base from the opposing strand. (credit: Jerome Walker/Dennis Myts) These two strands are complementary, with each base in one sticking to its partner on the other. The A-T pairs are connected by two hydrogen bonds, while the G-C pairs are connected by three hydrogen bonds. When two DNA sequences match in this way, such that they can stick to each other in an antiparallel fashion and form a helix, they are said to be complementary. Hydrogen bonding between complementary bases holds DNA strands together in a double helix of antiparallel strands. Thymine forms two hydrogen bonds with adenine, and guanine forms three hydrogen bonds with cytosine. (Credit: OpenStax Biology). Properties of RNA Ribonucleic acid (RNA), unlike DNA, is usually single-stranded. A nucleotide in an RNA chain will contain ribose (the five-carbon sugar), one of the four nitrogenous bases (A, U, G, or C), and a phosphate group. Here, we'll take a look at four major types of RNA: messenger RNA (mRNA), ribosomal RNA (rRNA), transfer RNA (tRNA), and regulatory RNAs. Messenger RNA (mRNA) 48 Messenger RNA (mRNA) is an intermediate between a protein-coding gene and its protein product. If a cell needs to make a particular protein, the gene encoding the protein will be turned “on,” meaning an RNA-polymerizing enzyme will come and make an RNA copy, or transcript, of the gene’s DNA sequence. The transcript carries the same information as the DNA sequence of its gene. However, in the RNA molecule, the base T is replaced with U. For instance, if a DNA coding strand has the sequence 5’-AATTGCGC-3’, the sequence of the corresponding RNA will be 5’-AAUUGCGC-3’. Once an mRNA has been produced, it will associate with a ribosome, a molecular machine that specializes in assembling proteins out of amino acids. The ribosome uses the information in the mRNA to make a protein of a specific sequence, “reading out” the mRNA’s nucleotides in groups of three (called codons) and adding a particular amino acid for each codon. Image of a ribosome (made of proteins and rRNA) bound to an mRNA, with tRNAs bringing amino acids to be added to the growing chain. The tRNA that binds, and thus the amino acid that's added, at a given moment is determined by the sequence of the mRNA that is being "read" at that time. (Credit: OpenStax Biology) Ribosomal RNA (rRNA) and transfer RNA (tRNA) Ribosomal RNA (rRNA) is a major component of ribosomes, where it helps mRNA bind in the right spot so its sequence information can be read out. Some rRNAs also act as enzymes, meaning that they help accelerate (catalyze) chemical reactions – in this case, the formation of bonds that link amino acids to form a protein. RNAs that act as enzymes are known as ribozymes. Transfer RNAs (tRNAs) are also involved in protein synthesis, but their job is to act as carriers – to bring amino acids to the ribosome, ensuring that the amino acid added to the chain is the one specified by the mRNA. Transfer RNAs consist of a single strand of RNA, but this strand has complementary segments that stick together to make double-stranded regions. This base-pairing creates a complex 3D structure important to the function of the molecule (Figure 2.25) 49 Figure 2.25 Structure of a tRNA. The overall molecule has a shape somewhat like an L. (Credit: Protein Data Bank) Regulatory RNA (miRNAs and siRNAs) Some types of non-coding RNAs (RNAs that do not encode proteins) help regulate the expression of other genes. Such RNAs may be called regulatory RNAs. For example, microRNAs (miRNAs) and small interfering RNAs siRNAs are small regulatory RNA molecules about 22 nucleotides long. They bind to specific mRNA molecules (with partly or fully complementary sequences) and reduce their stability or interfere with their translation, providing a way for the cell to decrease or fine-tune levels of these mRNAs. These are just some examples out of many types of noncoding and regulatory RNAs. Scientists are still discovering new varieties of noncoding RNA. Table 2. Summary of Features of DNA and RNA DNA RNA Function Repository of genetic information Involved in protein synthesis and gene regulation; carrier of genetic information in some viruses Sugar Deoxyribose Ribose Structure Double helix Usually single-stranded Bases C, T, A, G C, U, A, G modified from OpenStax Biology. 50 Practice Test Write your answers in ¼ sheet of yellow paper. Write your name, section, unit number and lesson number. Multiple Choice. 1. In DNA, which base pairing is a Watson-Crick base pairing? a. Guanine and cytosine b. Thymine and guanine c. Uracil and thymine d. Cytosine and adenine 2. DNA replication produces a new strand of DNA that is complementary to the parent strand. The parent strand's sequence is 5' GGCATCGAG 3' What is the sequence of the complementary DNA strand? a. 3′ CCGTAGCTC 5′ b. 5′ GGCATCGAG 3′ c. 5′ GAGCTACGG 3′ d. 5′ CTCGATGCC 3′ 51 UNIT SUMMARY Carbohydrates Carbohydrates are a group of macromolecules that are a vital energy source for the cell and provide structural support to plant cells, fungi, and all of the arthropods that include lobsters, crabs, shrimp, insects, and spiders. Scientists classify carbohydrates as monosaccharides, disaccharides, and polysaccharides depending on the number of monomers in the molecule. Monosaccharides are linked by glycosidic bonds that form as a result of dehydration reactions, forming disaccharides and polysaccharides with eliminating a water molecule for each bond formed. Glucose, galactose, and fructose are common monosaccharides; whereas, common disaccharides include lactose, maltose, and sucrose. Starch and glycogen, examples of polysaccharides, are the storage forms of glucose in plants and animals, respectively. The long polysaccharide chains may be branched or unbranched. Cellulose is an example of an unbranched polysaccharide; whereas, amylopectin, a constituent of starch, is a highly branched molecule. Glucose storage, in the form of polymers like starch of glycogen, makes it slightly less accessible for metabolism; however, this prevents it from leaking out of the cell or creating a high osmotic pressure that could cause the cell to uptake excessive water. Lipids Lipids are a class of macromolecules that are nonpolar and hydrophobic in nature. Major types include fats and oils, waxes, phospholipids, and steroids. Fats are a stored form of energy and are also known as triacylglycerols or triglycerides. Fats are comprised of fatty acids and either glycerol or sphingosine. Fatty acids may be unsaturated or saturated, depending on the presence or absence of double bonds in the hydrocarbon chain. If only single bonds are present, they are saturated fatty acids. Unsaturated fatty acids may have one or more double bonds in the hydrocarbon chain. Phospholipids comprise the membrane's matrix. They have a glycerol or sphingosine backbone to which two fatty acid chains and a phosphate-containing group are attached. Steroids are another class of lipids. Their basic structure has four fused carbon rings. Cholesterol is a type of steroid and is an important constituent of the plasma membrane, where it helps to maintain the membrane's fluid nature. It is also the precursor of steroid hormones such as testosterone. Proteins Proteins are a class of macromolecules that perform a diverse range of functions for the cell. They help in metabolism by acting as enzymes, carriers, or hormones, and provide structural support. The building blocks of proteins (monomers) are amino acids. Each amino acid has a central carbon that bonds to an amino group, a carboxyl group, a hydrogen atom, and an R group or side chain. There are 20 commonly occurring amino acids, each of which differs in the R group. A peptide bond links each amino acid to its neighbors. A long amino acid chain is a polypeptide. Proteins are organized at four levels: primary, secondary, tertiary, and (optional) quaternary. The primary structure is the amino acids' unique sequence. The polypeptide's local folding to form structures such as the α-helix and β-pleated sheet constitutes the secondary structure. The overall three-dimensional structure is the tertiary structure. When two or more 52 polypeptides combine to form the complete protein structure, the configuration is the protein's quaternary structure. Protein shape and function are intricately linked. Any change in shape caused by changes in temperature or pH may lead to protein denaturation and a loss in function. Nucleic Acids Nucleic acids are molecules comprised of nucleotides that direct cellular activities such as cell division and protein synthesis. Pentose sugar, a nitrogenous base, and a phosphate group comprise each nucleotide. There are two types of nucleic acids: DNA and RNA. DNA carries the cell's genetic blueprint and passes it on from parents to offspring (in the form of chromosomes). It has a double-helical structure with the two strands running in opposite directions, connected by hydrogen bonds, and complementary to each other. RNA is a singlestranded polymer composed of linked nucleotides made up of a pentose sugar (ribose), a nitrogenous base, and a phosphate group. RNA is involved in protein synthesis and its regulation. Messenger RNA (mRNA) copies from the DNA, exports itself from the nucleus to the cytoplasm, and contains information for constructing proteins. Ribosomal RNA (rRNA) is a part of the ribosomes at the site of protein synthesis; whereas, transfer RNA (tRNA) carries the amino acid to the site of protein synthesis. The microRNA regulates using mRNA for protein synthesis. 53 54 55 56 UNIT III. ENZYMES By the end of this section, you will be able to do the following: 1. Describe the role of enzymes in metabolic pathways 2. Explain how enzymes function as molecular catalysts 3. Discuss enzyme regulation by various factors Enzymes and activation energy A substance that helps a chemical reaction to occur is a catalyst, and the special molecules that catalyze biochemical reactions are enzymes. Almost all enzymes are proteins, comprised of amino acid chains, and they perform the critical task of lowering the activation energies of chemical reactions inside the cell. Enzymes do this by binding to the reactant molecules, and holding them in such a way as to make the chemical bond-breaking and bondforming processes take place more readily. It is important to remember that enzymes do not change the reaction's ∆G. In other words, they do not change whether a reaction is exergonic (spontaneous) or endergonic. This is because they do not change the reactants' or products' free energy. They only reduce the activation energy required to reach the transition state (Figure 3.1). Figure 3.1 Enzymes lower the reaction's activation energy but do not change the reaction's free energy. Enzyme Active Site and Substrate Specificity To catalyze a reaction, an enzyme will grab on (bind) to one or more reactant molecules. These molecules are the enzyme's substrates. In some reactions, one substrate is broken down into multiple products. In others, two substrates come together to create one larger molecule or to swap pieces. In fact, whatever type of biological reaction you can think of, there is probably an enzyme to speed it up! The part of the enzyme where the substrate binds is called the active site (since that’s where the catalytic “action” happens). 57 Since enzymes are proteins, there is a unique combination of amino acid residues (also side chains, or R groups) within the active site. Different properties characterize each residue. These can be large or small, weakly acidic or basic, hydrophilic or hydrophobic, positively or negatively charged, or neutral. The unique combination of amino acid residues, their positions, sequences, structures, and properties, creates a very specific chemical environment within the active site. This specific environment is suited to bind, albeit briefly, to a specific chemical substrate (or substrates). Due to this jigsaw puzzle-like match between an enzyme and its substrates (which adapts to find the best fit between the transition state and the active site), enzymes are known for their specificity. The “best fit” results from the shape and the amino acid functional group’s attraction to the substrate. There is a specifically matched enzyme for each substrate and, thus, for each chemical reaction; however, there is flexibility as well. The fact that active sites are so perfectly suited to provide specific environmental conditions also means that they are subject to local environmental influences. It is true that increasing the environmental temperature generally increases reaction rates, enzyme-catalyzed or otherwise. However, increasing or decreasing the temperature outside of an optimal range can affect chemical bonds within the active site in such a way that they are less well suited to bind substrates. High temperatures will eventually cause enzymes, like other biological molecules, to denature, a process that changes the substance's natural properties. Likewise, the local environment's pH can also affect enzyme function. Active site amino acid residues have their own acidic or basic properties that are optimal for catalysis. These residues are sensitive to changes in pH that can impair the way substrate molecules bind. Enzymes are suited to function best within a certain pH range, and, as with temperature, extreme environmental pH values (acidic or basic) can cause enzymes to denature. Induced Fit and Enzyme Function For many years, scientists thought that enzyme-substrate binding took place in a simple “lock-and-key” fashion. This model asserted that the enzyme and substrate fit together perfectly in one instantaneous step. However, current research supports a more refined view scientists’ call induced fit (Figure 3.2). This model expands upon the lock-and-key model by describing a more dynamic interaction between enzyme and substrate. As the enzyme and substrate come together, their interaction causes a mild shift in the enzyme’s structure that confirms an ideal binding arrangement between the enzyme and the substrate's transition state. This ideal binding maximizes the enzyme’s ability to catalyze its reaction. Figure 3.2 According to the induced-fit model, both enzyme and substrate undergo dynamic conformational changes upon binding. The enzyme contorts the substrate into its transition state, thereby increasing the reaction's rate. 58 Metabolism Control through Enzyme Regulation It would seem ideal to have a scenario in which all the encoded enzymes in an organism’s genome existed in abundant supply and functioned optimally under all cellular conditions, in all cells, at all times. In reality, this is far from the case. A variety of mechanisms ensure that this does not happen. Cellular needs and conditions vary from cell to cell, and change within individual cells over time. The required enzymes and energetic demands of stomach cells are different from those of fat storage cells, skin cells, blood cells, and nerve cells. Furthermore, a digestive cell works much harder to process and break down nutrients during the time that closely follows a meal compared with many hours after a meal. As these cellular demands and conditions vary, so do the amounts and functionality of different enzymes. Since the rates of biochemical reactions are controlled by activation energy, and enzymes lower and determine activation energies for chemical reactions, the relative amounts and functioning of the variety of enzymes within a cell ultimately determine which reactions will proceed and at which rates. This determination is tightly controlled. In certain cellular environments, environmental factors like pH and temperature partly control enzyme activity. There are other mechanisms through which cells control enzyme activity and determine the rates at which various biochemical reactions will occur. Molecular Regulation of Enzymes Enzymes can be regulated in ways that either promote or reduce their activity. There are many different kinds of molecules that inhibit or promote enzyme function, and various mechanisms exist for doing so. For example, in some cases of enzyme inhibition, an inhibitor molecule is similar enough to a substrate that it can bind to the active site and simply block the substrate from binding. When this happens, the enzyme is inhibited through competitive inhibition, because an inhibitor molecule competes with the substrate for active site binding (Figure 3.3). Alternatively, in noncompetitive inhibition, an inhibitor molecule binds to the enzyme at an allosteric site, a binding site away from the active site, and still manages to block substrate binding to the active site. Figure 3.3 Competitive and noncompetitive inhibition affect the reaction's rate differently. Competitive inhibitors affect the initial rate but do not affect the maximal rate; whereas, noncompetitive inhibitors affect the maximal rate. 59 Some inhibitor molecules bind to enzymes in a location where their binding induces a conformational change that reduces the enzyme's affinity for its substrate. This type of inhibition is an allosteric inhibition (Figure 3.4). More than one polypeptide comprise most allosterically regulated enzymes, meaning that they have more than one protein subunit. When an allosteric inhibitor binds to an enzyme, all active sites on the protein subunits change slightly such that they bind their substrates with less efficiency. There are allosteric activators as well as inhibitors. Allosteric activators bind to locations on an enzyme away from the active site, inducing a conformational change that increases the affinity of the enzyme’s active site(s) for its substrate(s). Figure 3.4 Allosteric inhibitors modify the enzyme's active site so that substrate binding is reduced or prevented. In contrast, allosteric activators modify the enzyme's active site so that the affinity for the substrate increases. Many enzymes don’t work optimally, or even at all, unless bound to other specific non-protein helper molecules, either temporarily through ionic or hydrogen bonds or permanently through stronger covalent bonds. Two types of helper molecules are cofactors and coenzymes. Binding to these molecules promotes optimal conformation and function for their respective enzymes. Cofactors are inorganic ions such as iron (Fe++) and magnesium (Mg++). One example of an enzyme that requires a metal ion as a cofactor is the enzyme that builds DNA molecules, DNA polymerase, which requires a bound zinc ion (Zn++) to function. Coenzymes are organic helper molecules, with a basic atomic structure comprised of carbon and hydrogen, which are required for enzyme action. The most common sources of coenzymes are dietary vitamins (Figure 3.5). Some vitamins are precursors to coenzymes and others act directly as coenzymes. Vitamin C is a coenzyme for multiple enzymes that take part in Figure 3.5 Vitamins are important coenzymes or precursors of coenzymes, and are required for enzymes to function properly. Multivitamin capsules usually contain mixtures of all the vitamins at different percentages. 60 building the important connective tissue component, collagen. An important step in breaking down glucose to yield energy is catalysis by a multi-enzyme complex scientists call pyruvate dehydrogenase. Pyruvate dehydrogenase is a complex of several enzymes that actually requires one cofactor (a magnesium ion) and five different organic coenzymes to catalyze its specific chemical reaction. Therefore, enzyme function is, in part, regulated by an abundance of various cofactors and coenzymes, which the diets of most organisms supply. Enzyme Compartmentalization In eukaryotic cells, molecules such as enzymes are usually compartmentalized into different organelles. This allows for yet another level of regulation of enzyme activity. Enzymes required only for certain cellular processes are sometimes housed separately along with their substrates, allowing for more efficient chemical reactions. Examples of this sort of enzyme regulation based on location and proximity include the enzymes involved in the latter stages of cellular respiration, which take place exclusively in the mitochondria, and the enzymes involved in digesting cellular debris and foreign materials, located within lysosomes. Feedback Inhibition in Metabolic Pathways Molecules can regulate enzyme function in many ways. However, a major question remains: What are these molecules and from where do they come? Some are cofactors and coenzymes, ions, and organic molecules, as you have learned. What other molecules in the cell provide enzymatic regulation, such as allosteric modulation, and competitive and noncompetitive inhibition? The answer is that a wide variety of molecules can perform these roles. Some include pharmaceutical and non-pharmaceutical drugs, toxins, and poisons from the environment. Perhaps the most relevant sources of enzyme regulatory molecules, with respect to cellular metabolism, are cellular metabolic reaction products themselves. In a most efficient and elegant way, cells have evolved to use their own reactions' products for feedback inhibition of enzyme activity. Feedback inhibition involves using a reaction product to regulate its own further production (Figure 3.6). The cell responds to the abundance of specific products by slowing down production during anabolic or catabolic reactions. Such reaction products may inhibit the enzymes that catalyzed their production through the mechanisms that we described above. Figure 3.6 Metabolic pathways are a series of reactions that multiple enzymes catalyze. Feedback inhibition, where the pathway's end product inhibits an upstream step, is an important regulatory mechanism in cells. 61 Producing both amino acids and nucleotides is controlled through feedback inhibition. Additionally, ATP is an allosteric regulator of some of the enzymes involved in sugar's catabolic breakdown, the process that produces ATP. In this way, when ATP is abundant, the cell can prevent its further production. Remember that ATP is an unstable molecule that can spontaneously dissociate into ADP. If too much ATP were present in a cell, much of it would go to waste. Alternatively, ADP serves as a positive allosteric regulator (an allosteric activator) for some of the same enzymes that ATP inhibits. Thus, when relative ADP levels are high compared to ATP, the cell is triggered to produce more ATP through sugar catabolism. Enzymes in Clinical Diagnosis Plasma enzymes can be classified into two major groups: 1. Those, relatively, small group of enzymes secreted into the plasma by certain organs (i.e. Enzymes those have function in plasma) For example: - the liver secretes zymogens of the enzymes involved in blood coagulation. 2. Those large enzyme species released from cells during normal cell turnover. These enzymes are normally intracellular and have no physiologic function in the plasma. In healthy individuals the levels of these enzymes are fairly constant and represent steady state in which the rate of release from cells into the plasma is balanced by an equal rate or removal from the plasma. Many diseases that cause tissue damage result in an increased release of intracellular enzymes into the plasma. The activities of many of these enzymes are routinely 20 determined for diagnostic purposes in diseases of the heart, liver, skeletal muscle, and other tissues. The level of specific enzyme activity in the plasma frequently correlates with the extent of tissue damage. Thus, the degree of elevation of a particular enzyme activity in plasma is often useful in evaluating the diagnosis and prognosis for the patient. Measurement of enzymes concentration of mostly the latter type in plasma gives valuable informatio0n about disease involving tissues of their origin. 1. Lipase: It is an enzyme catalyzing the hydrolysis of fats. It is secreted by pancreas and Liver. The plasma lipase level may be low in liver disease, Vitamin A deficiency, some malignancies, and diabetes mellitus. It may be elevated in acute pancreatitis and pancreatic carcinoma. 2. α- Amylase: α- amylase is the enzyme concerned with the breakdown of dietary starch and glycogen to maltose. It is present in pancreatic juice and saliva as well as in liver fallopian tubes and muscles. The enzyme is excreted in the Urine. The main use of amylase estimations is in the diagnosis of acute pancreatitis. The plasma amylase level may be low in liver disease and increased in high intestinal obstruction, mumps, acute pancreatitis and diabetes. 3. Trypsin: Trypsin is secreted by pancreas. Elevated levels of trypsin in plasma occur during acute pancreatic disease. 4. Alkaline phosphates (ALP): The alkaline phosphates are a group of enzymes, which hydrolyze phosphate esters at an alkaline pH. They are found in bone, liver, kidney, intestinal wall, lactating mammary gland and placenta. In bone the enzyme is found in osteoblasts and is probably important for normal bone function. The level of these enzymes may be increased in 62 rickets and osteomalacia, hyperparathyroidism, paget's disease of bone, obstructive jaundice, and metastatic carcinoma. Serum alkaline phosphatase levels may be increase in congestive heart failure result of injury to the liver. 5. Acid Phosphatase (ACP) Acid phosphatases catalyzing the hydrolysis of various phosphate esters at acidic pH is found in the prostate, liver, red cells, platelets and bone. It may be elevated in metastatic prostatic carcinoma. 6. Transaminases: Two transaminases are of clinical interest. 1. Aspartate Transaminase, AST (Glutamate oxaloacetate transaminase, GOT) catalyzes the transfer of the amino group of aspartic acid to α- ketoglutarate forming glutamate and oxaloacetate. AST or GOT is widely distributed, with high concentration, in the heart, liver, skeletal muscle, kidney and erythrocytes, and damage to any of these tissues may cause raised levels. 2. Alanine transaminase, ALT (Glutamate pyruvate transaminase, GPT) Transfer the amino group of alanine to α- ketoglutarate, forming glutamate and pyruvate. It is present in high concentration in liver and to a lesser extent in skeletal muscle, kidney and heart. Serum levels of glutamate- pyruvate transaminase (SGOT) and Glutamateoxaloacetatetransaminase (SGOT) are useful in the diagnosis of liver parenchymal damage and myocardial damage respectively. In liver damage, both enzymes are increased, but SGPT increases more. In myocardial infarction SGOT is increased with little or no increase in SGPT. 7. Lactate Dehydrogenase (LDH): It catalyzes the reversible interconversion of lactate and pyruvate. It is widely distributed with high concentrations in the heart, skeletal muscle, liver, kidney, brain and erythrocytes. The enzyme is increased in plasma in myocardial infarction, acute leukemias, and generalized carcinomatosis and in acute hepatitis. Estimation of it isoenzymes is more useful in clinical diagnosis to differentiate hepatic disease and myocardial infarction. 8. Creatine kinase (CK) or creatine phosphokinase (CPK) CK (CPK) is found in heart muscle brain and skeletal muscle. Measurement of serum creatine phosphokinase activity is of value in the diagnosis of disorders affecting skeletal and cardiac muscle. The level of CPK in plasma highly increased in myocardial infarction. Factors Affecting Enzymes Activity Physical and chemical factors are affecting the enzyme activity. These include: 1. Temperature 2. pH 3. Substrate/enzyme concentration Temperature Starting from low temperature as the temperature increases to certain degree the activity of the enzyme increases because the temperature increase the total energy of the chemical 63 system. There is an optimal temperature at which the reaction is most rapid (maximum). Above this optimal temperature, the reaction rate decreases sharply, mainly due to denaturation of the enzyme by heat. The temperature at which an enzyme shows maximum activity is known as the optimum temperature for the enzyme. For most body enzymes the optimum temperature is around 37OC, which is body temperature. Figure 3.7 Temperature influence on rate of reaction (Image source: By Thomas Shafee CC BY 4.0, via Wikimedia Commons) Effect of pH The concentration of H+ affects reaction velocity in several ways. First, the catalytic process usually requires that the enzyme and substrate have specific chemical groups in an ionized or unionized sate in order to interact. For example, catalytic activity may require that an amino-group of the enzyme be in the protonated form (-NH3+). At alkaline pH, this group is deprotonated and the rate of reaction therefore declines. Extreme pH can also lead to denaturation of the enzyme, because the structure of the catalytically active protein molecule depends on the ionic character of the amino acid chains. The pH at which maximum enzyme activity is achieved is different for every enzymes. For example, pepsin, a digestive enzyme in the stomach, has maximum action at pH 2, whereas other enzymes, designed to work at neutral pH, are denatured by such an acidic environment. Figure 3.8 pH influence on rate of reaction (Image source: www.toppr.com) 64 Concentration of substrate The activity of enzymes is influenced by increase in substrate concentration. An increase in the substrate concentration increases the enzyme activity until a maximum point is reached. Further increase in substrate concentration does not increase rate of reaction. This condition shows that as concentration of substrate is increased, the substrate molecule combine with all available enzyme molecules at their active site until no more active sites are available (The active Sites become saturated). At this state, the enzyme is obtained it maximum rate (V max). Figure 3.9 Substrate concentration influence on rate of reaction (Image source: www.philpoteducation.com) 65 UNIT SUMMARY Enzymes Enzymes are chemical catalysts that accelerate chemical reactions at physiological temperatures by lowering their activation energy. Enzymes are usually proteins consisting of one or more polypeptide chains. Enzymes have an active site that provides a unique chemical environment, comprised of certain amino acid R groups (residues). This unique environment is perfectly suited to convert particular chemical reactants for that enzyme, scientists call substrates, into unstable intermediates that they call transition states. Enzymes and substrates bind with an induced fit, which means that enzymes undergo slight conformational adjustments upon substrate contact, leading to full, optimal binding. Enzymes bind to substrates and catalyze reactions in four different ways: bringing substrates together in an optimal orientation, compromising the bond structures of substrates so that bonds can break down more easily, providing optimal environmental conditions for a reaction to occur, or participating directly in their chemical reaction by forming transient covalent bonds with the substrates. Enzyme action must be regulated so that in a given cell at a given time, the desired reactions catalyze and the undesired reactions are not. Enzymes are regulated by cellular conditions, such as temperature and pH. They are also regulated through their location within a cell, sometimes compartmentalized so that they can only catalyze reactions under certain circumstances. Enzyme inhibition and activation via other molecules are other important ways that enzymes are regulated. Inhibitors can act competitively, noncompetitively, or allosterically. Noncompetitive inhibitors are usually allosteric. Activators can also enhance enzyme function allosterically. The most common method by which cells regulate the enzymes in metabolic pathways is through feedback inhibition. During feedback inhibition, metabolic pathway products serve as inhibitors (usually allosteric) of one or more of the enzymes (usually the first committed enzyme of the pathway) involved in the pathway that produces them. 66 UNIT CRITICAL THINKING QUESTIONS Write your answer in a ¼ sheet of yellow paper. Write your name, section, unit number and lesson number. 1. If no activation energy were required to break down sucrose (table sugar), would you be able to store it in a sugar bowl? 67 68 UNIT IV. CELLULAR RESPIRATION Introduction The electrical energy plant in Figure 4.1 converts energy from one form to another form that can be more easily used. This type of generating plant starts with underground thermal energy (heat) and transforms it into electrical energy that will be transported to homes and factories. Like a generating plant, plants and animals also must take in energy from the environment and convert it into a form that their cells can use. Mass and its stored energy enter an organism’s body in one form and are converted into another form that can fuel the organism’s life functions. In the process of photosynthesis, plants and other photosynthetic producers take in energy in the form of light (solar energy) and convert it into chemical energy in the form of glucose, which stores this energy in its chemical bonds. Then, a series of metabolic pathways, collectively called cellular respiration, extracts the energy from the bonds in glucose and converts it into a form that all living things can use. Figure 4.1 This geothermal energy plant transforms thermal energy from deep in the ground into electrical energy, which can be easily used. (credit: U.S. Department of Defense) 69 Lesson 1: Overview of the Metabolic Reactions By the end of this section, you will be able to: 1. Describe the process by which polymers are broken down into monomers 2. Describe the process by which monomers are combined into polymers 3. Discuss the role of ATP in metabolism 4. Describe the hormones that regulate anabolic and catabolic reactions Metabolic processes are constantly taking place in the body. Metabolism is the sum of all of the chemical reactions that are involved in catabolism and anabolism. The reactions governing the breakdown of food to obtain energy are called catabolic reactions. Conversely, anabolic reactions use the energy produced by catabolic reactions to synthesize larger molecules from smaller ones, such as when the body forms proteins by stringing together amino acids. Both sets of reactions are critical to maintaining life. Because catabolic reactions produce energy and anabolic reactions use energy, ideally, energy usage would balance the energy produced. If the net energy change is positive (catabolic reactions release more energy than the anabolic reactions use), then the body stores the excess energy by building fat molecules for long-term storage. On the other hand, if the net energy change is negative (catabolic reactions release less energy than anabolic reactions use), the body uses stored energy to compensate for the deficiency of energy released by catabolism. Catabolic Reactions Catabolic reactions break down large organic molecules into smaller molecules, releasing the energy contained in the chemical bonds. These energy releases (conversions) are not 100 percent efficient. The amount of energy released is less than the total amount contained in the molecule. Approximately 40 percent of energy yielded from catabolic reactions is directly transferred to the high-energy molecule adenosine triphosphate (ATP). ATP, the energy currency of cells, can be used immediately to power molecular machines that support cell, tissue, and organ function. This includes building new tissue and repairing damaged tissue. ATP can also be stored to fulfill future energy demands. The remaining 60 percent of the energy released from catabolic reactions is given off as heat, which tissues and body fluids absorb. Structurally, ATP molecules consist of an adenine, a ribose, and three phosphate groups (Figure 4.2). The chemical bond between the second and third phosphate groups, termed a high-energy bond, represents the greatest source of energy in a cell. It is the first bond that catabolic enzymes break when cells require energy to do work. The products of this reaction are a molecule of adenosine diphosphate (ADP) and a lone phosphate group (P i). ATP, ADP, and Pi are constantly being cycled through reactions that build ATP and store energy, and reactions that break down ATP and release energy. 70 Figure 4.2. Structure of ATP Molecule. Adenosine triphosphate (ATP) is the energy molecule of the cell. During catabolic reactions, ATP is created and energy is stored until needed during anabolic reactions (Credit: opentextbc.ca). The energy from ATP drives all bodily functions, such as contracting muscles, maintaining the electrical potential of nerve cells, and absorbing food in the gastrointestinal tract. The metabolic reactions that produce ATP come from various sources (Figure 4.3). Figure 4.3. Sources of ATP. During catabolic reactions, proteins are broken down into amino acids, lipids are broken down into fatty acids, and polysaccharides are broken down into monosaccharides. These building blocks are then used for the synthesis of molecules in anabolic reactions (Credit: opentextbc.ca) Of the four major macromolecular groups (carbohydrates, lipids, proteins, and nucleic acids) that are processed by digestion, carbohydrates are considered the most common source of energy to fuel the body. They take the form of either complex carbohydrates, polysaccharides like starch and glycogen, or simple sugars (monosaccharides) like glucose and fructose. Sugar catabolism breaks polysaccharides down into their individual monosaccharides. Among the monosaccharides, glucose is the most common fuel for ATP production in cells, and as such, 71 there are a number of endocrine control mechanisms to regulate glucose concentration in the bloodstream. Excess glucose is either stored as an energy reserve in the liver and skeletal muscles as the complex polymer glycogen, or it is converted into fat (triglyceride) in adipose cells (adipocytes). Among the lipids (fats), triglycerides are most often used for energy via a metabolic process called β-oxidation. About one-half of excess fat is stored in adipocytes that accumulate in the subcutaneous tissue under the skin, whereas the rest is stored in adipocytes in other tissues and organs. Proteins, which are polymers, can be broken down into their monomers, individual amino acids. Amino acids can be used as building blocks of new proteins or broken down further for the production of ATP. When one is chronically starving, this use of amino acids for energy production can lead to a wasting away of the body, as more and more proteins are broken down. Nucleic acids are present in most of the foods you eat. During digestion, nucleic acids including DNA and various RNAs are broken down into their constituent nucleotides. These nucleotides are readily absorbed and transported throughout the body to be used by individual cells during nucleic acid metabolism. Anabolic Reactions In contrast to catabolic reactions, anabolic reactions involve the joining of smaller molecules into larger ones. Anabolic reactions combine monosaccharides to form polysaccharides, fatty acids to form triglycerides, amino acids to form proteins, and nucleotides to form nucleic acids. These processes require energy in the form of ATP molecules generated by catabolic reactions. Anabolic reactions, also called biosynthesis reactions, create new molecules that form new cells and tissues, and revitalize organs. Hormonal Regulation of Metabolism Catabolic and anabolic hormones in the body help regulate metabolic processes. Catabolic hormones stimulate the breakdown of molecules and the production of energy. These include cortisol, glucagon, adrenaline/epinephrine, and cytokines. All of these hormones are mobilized at specific times to meet the needs of the body. Anabolic hormones are required for the synthesis of molecules and include growth hormone, insulin-like growth factor, insulin, testosterone, and estrogen. Table 4.1 summarizes the function of each of the catabolic hormones and Table 4.2 summarizes the functions of the anabolic hormones. Table 4.1. Catabolic Hormones and their functions Hormone Function Released from the adrenal gland in response to stress; its main role is to Cortisol increase blood glucose levels by gluconeogenesis (breaking down fats and proteins) Released from alpha cells in the pancreas either when starving or when the body needs to generate additional energy; it stimulates the Glucagon breakdown of glycogen in the liver to increase blood glucose levels; its effect is the opposite of insulin; glucagon and insulin are a part of a negative-feedback system that stabilizes blood glucose levels 72 Table 4.1. Catabolic Hormones and their functions Hormone Function Released in response to the activation of the sympathetic nervous system; increases heart rate and heart contractility, constricts blood Adrenaline/epinephrine vessels, is a bronchodilator that opens (dilates) the bronchi of the lungs to increase air volume in the lungs, and stimulates gluconeogenesis Table 4.2. Catabolic Hormones and their functions Hormone Function Growth hormone Synthesized and released from the pituitary gland; stimulates the growth (GH) of cells, tissues, and bones Insulin-like Stimulates the growth of muscle and bone while also inhibiting cell death growth factor (apoptosis) (IGF) Produced by the beta cells of the pancreas; plays an essential role in carbohydrate and fat metabolism, controls blood glucose levels, and promotes the uptake of glucose into body cells; causes cells in muscle, Insulin adipose tissue, and liver to take up glucose from the blood and store it in the liver and muscle as glucagon; its effect is the opposite of glucagon; glucagon and insulin are a part of a negative-feedback system that stabilizes blood glucose levels Produced by the testes in males and the ovaries in females; stimulates Testosterone an increase in muscle mass and strength as well as the growth and strengthening of bone Produced primarily by the ovaries, it is also produced by the liver and Estrogen adrenal glands; its anabolic functions include increasing metabolism and fat deposition Metabolic Processes: Cushing Syndrome and Addison’s disease As might be expected for a fundamental physiological process like metabolism, errors or malfunctions in metabolic processing lead to a pathophysiology or—if uncorrected—a disease state. Metabolic diseases are most commonly the result of malfunctioning proteins or enzymes that are critical to one or more metabolic pathways. Protein or enzyme malfunction can be the consequence of a genetic alteration or mutation. However, normally functioning proteins and enzymes can also have deleterious effects if their availability is not appropriately matched with metabolic need. For example, excessive production of the hormone cortisol (see Table 1) gives rise to Cushing syndrome. Clinically, Cushing syndrome is characterized by rapid weight gain, especially in the trunk and face region, depression, and anxiety. It is worth mentioning that tumors of the pituitary that produce adrenocorticotropic hormone (ACTH), which subsequently stimulates the adrenal cortex to release excessive cortisol, produce similar effects. This indirect mechanism of cortisol overproduction is referred to as Cushing disease. Patients with Cushing syndrome can exhibit high blood glucose levels and are at an increased risk of becoming obese. They also show slow growth, accumulation of fat between the shoulders, weak muscles, bone pain (because cortisol causes proteins to be broken down to make glucose via gluconeogenesis), and fatigue. Other symptoms include excessive sweating 73 (hyperhidrosis), capillary dilation, and thinning of the skin, which can lead to easy bruising. The treatments for Cushing syndrome are all focused on reducing excessive cortisol levels. Depending on the cause of the excess, treatment may be as simple as discontinuing the use of cortisol ointments. In cases of tumors, surgery is often used to remove the offending tumor. Where surgery is inappropriate, radiation therapy can be used to reduce the size of a tumor or ablate portions of the adrenal cortex. Finally, medications are available that can help to regulate the amounts of cortisol. Insufficient cortisol production is equally problematic. Adrenal insufficiency, or Addison’s disease, is characterized by the reduced production of cortisol from the adrenal gland. It can result from malfunction of the adrenal glands—they do not produce enough cortisol—or it can be a consequence of decreased ACTH availability from the pituitary. Patients with Addison’s disease may have low blood pressure, paleness, extreme weakness, fatigue, slow or sluggish movements, lightheadedness, and salt cravings due to the loss of sodium and high blood potassium levels (hyperkalemia). Victims also may suffer from loss of appetite, chronic diarrhea, vomiting, mouth lesions, and patchy skin color. Diagnosis typically involves blood tests and imaging tests of the adrenal and pituitary glands. Treatment involves cortisol replacement therapy, which usually must be continued for life. 74 Lesson 2: Energy in Living Systems By the end of this section, you will be able to do the following: 1. Discuss the importance of electrons in the transfer of energy in living systems 2. Explain how ATP is used by cells as an energy source Energy production within a cell involves many coordinated chemical pathways. Most of these pathways are combinations of oxidation and reduction reactions, which occur at the same time. An oxidation reaction strips an electron from an atom in a compound, and the addition of this electron to another compound is a reduction reaction. Because oxidation and reduction usually occur together, these pairs of reactions are called oxidation reduction reactions, or redox reactions. Electrons and Energy The removal of an electron from a molecule (oxidizing it), results in a decrease in potential energy in the oxidized compound. The electron (sometimes as part of a hydrogen atom) does not remain unbonded, however, in the cytoplasm of a cell. Rather, the electron is shifted to a second compound, reducing the second compound. The shift of an electron from one compound to another removes some potential energy from the first compound (the oxidized compound) and increases the potential energy of the second compound (the reduced compound). The transfer of electrons between molecules is important because most of the energy stored in atoms and used to fuel cell functions is in the form of high-energy electrons. The transfer of energy in the form of high-energy electrons allows the cell to transfer and use energy in an incremental fashion—in small packages rather than in a single, destructive burst. This chapter focuses on the extraction of energy from food; you will see that as you track the path of the transfers, you are tracking the path of electrons moving through metabolic pathways. Electron Carriers In living systems, a small class of compounds functions as electron shuttles: they bind and carry high-energy electrons between compounds in biochemical pathways. The principal electron carriers we will consider are derived from the B vitamin group and are derivatives of nucleotides. These compounds can be easily reduced (that is, they accept electrons) or oxidized (they lose electrons). Nicotinamide adenine dinucleotide (NAD) (Figure 4.4) is derived from vitamin B3, niacin. NAD+ is the oxidized form of the molecule; NADH is the reduced form of the molecule after it has accepted two electrons and a proton (which together are the equivalent of a hydrogen atom with an extra electron). Note that if a compound has an “H” on it, it is generally reduced (e.g., NADH is the reduced form of NAD). NAD+ can accept electrons from an organic molecule according to the general equation: When electrons are added to a compound, it is reduced. A compound that reduces another is called a reducing agent. In the above equation, RH is a reducing agent, and NAD+ is reduced to NADH. When electrons are removed from a compound, it is oxidized. A compound 75 that oxidizes another is called an oxidizing agent. In the above equation, NAD+ is an oxidizing agent, and RH is oxidized to R. Figure 4.4 The oxidized form of the electron carrier (NAD+) is shown on the left, and the reduced form (NADH) is shown on the right. The nitrogenous base in NADH has one more hydrogen ion and two more electrons than in NAD+. This illustration shows the molecular structure of NAD+ and NADH. Both compounds are composed of an adenine nucleotide and a nicotinamide nucleotide, which bond together to form a dinucleotide. The nicotinamide nucleotide is at the 5 prime end, and the adenine nucleotide is at the 3 prime end. Nicotinamide is a nitrogenous base, meaning it has nitrogen in a sixmembered carbon ring. In NADH, one extra hydrogen is associated with this ring, which is not found in NAD+. Similarly, flavin adenine dinucleotide (FAD+) is derived from vitamin B2, also called riboflavin. Its reduced form is FADH 2. A second variation of NAD, NADP, contains an extra phosphate group. Both NAD+ and FAD+ are extensively used in energy extraction from sugars, and NADP plays an important role in anabolic reactions and photosynthesis in plants. ATP Structure and Function At the heart of ATP is a molecule of adenosine monophosphate (AMP), which is composed of an adenine molecule bonded to a ribose molecule and to a single phosphate group (Figure 4.5). Ribose is a five-carbon sugar found in RNA, and AMP is one of the nucleotides in RNA. The addition of a second phosphate group to this core molecule results in the formation of adenosine diphosphate (ADP); the addition of a third phosphate group forms adenosine triphosphate (ATP). 76 Figure 4.5 ATP (adenosine triphosphate) has three phosphate groups that can be removed by hydrolysis (addition of H2O) to form ADP (adenosine diphosphate) or AMP (adenosine monophosphate). The negative charges on the phosphate group naturally repel each other, requiring energy to bond them together and releasing energy when these bonds are broken. This illustration shows the molecular structure of ATP. This molecule is an adenine nucleotide with a string of three phosphate groups attached to it. The phosphate groups are named alpha, beta, and gamma in order of increasing distance from the ribose sugar to which they are attached. The addition of a phosphate group to a molecule requires energy. Phosphate groups are negatively charged and thus repel one another when they are arranged in series, as they are in ADP and ATP. This repulsion makes the ADP and ATP molecules inherently unstable. The release of one or two phosphate groups from ATP, a process called dephosphorylation, releases energy. Energy from ATP Hydrolysis is the process of breaking complex macromolecules apart. During hydrolysis, water is split, or lysed, and the resulting hydrogen atom (H +) and a hydroxyl group (OH -), or hydroxide, are added to the larger molecule. The hydrolysis of ATP produces ADP, together with an inorganic phosphate ion (Pi), and the release of free energy. To carry out life processes, ATP is continuously broken down into ADP, and like a rechargeable battery, ADP is continuously regenerated into ATP by the reattachment of a third phosphate group. Water, which was broken down into its hydrogen atom and hydroxyl group (hydroxide) during ATP hydrolysis, is regenerated when a third phosphate is added to the ADP molecule, reforming ATP. Obviously, energy must be infused into the system to regenerate ATP. Where does this energy come from? In nearly every living thing on Earth, the energy comes from the metabolism of glucose, fructose, or galactose, all isomers with the chemical formula C 6H12O6 but different molecular configurations. In this way, ATP is a direct link between the limited set of exergonic pathways of glucose catabolism and the multitude of endergonic pathways that power living cells. Phosphorylation Recall that, in some chemical reactions, enzymes may bind to several substrates that react with each other on the enzyme, forming an intermediate complex. An intermediate complex is a temporary structure, and it allows one of the substrates (such as ATP) and reactants to more readily react with each other; in reactions involving ATP, ATP is one of the substrates and ADP is a product. During an endergonic chemical reaction, ATP forms an intermediate complex with the substrate and enzyme in the reaction. This intermediate complex allows the ATP to transfer its third phosphate group, with its energy, to the substrate, a process 77 called phosphorylation. Phosphorylation refers to the addition of the phosphate (~P). This is illustrated by the following generic reaction, in which A and B represent two different substrates: When the intermediate complex breaks apart, the energy is used to modify the substrate and convert it into a product of the reaction. The ADP molecule and a free phosphate ion are released into the medium and are available for recycling through cell metabolism. Substrate Phosphorylation ATP is generated through two mechanisms during the breakdown of glucose. A few ATP molecules are generated (that is, regenerated from ADP) as a direct result of the chemical reactions that occur in the catabolic pathways. A phosphate group is removed from an intermediate reactant in the pathway, and the free energy of the reaction is used to add the third phosphate to an available ADP molecule, producing ATP (Figure 4.6). This very direct method of phosphorylation is called substrate-level phosphorylation. Figure 4.6 In phosphorylation reactions, the gamma (third) phosphate of ATP is attached to a protein. This illustration shows a substrate-level phosphorylation reaction in which the gamma phosphate of ATP is attached to a protein. Oxidative Phosphorylation Most of the ATP generated during glucose catabolism, however, is derived from a much more complex process, chemiosmosis, which takes place in mitochondria (Figure 4.7) within a eukaryotic cell or the plasma membrane of a prokaryotic cell. Chemiosmosis, a process of ATP production in cellular metabolism, is used to generate 90 percent of the ATP made during glucose catabolism and is also the method used in the light reactions of photosynthesis to harness the energy of sunlight. The production of ATP using the process of chemiosmosis is called oxidative phosphorylation because of the involvement of oxygen in the process. 78 Figure 4.7 In eukaryotes, oxidative phosphorylation takes place in mitochondria. In prokaryotes, this process takes place in the plasma membrane. (Credit: modification of work by Mariana Ruiz Villareal) This illustration shows the structure of a mitochondrion, which has an outer membrane and an inner membrane. The inner membrane has many folds, called cristae. The space between the outer membrane and the inner membrane is called the intermembrane space, and the central space of the mitochondrion is called the matrix. ATP synthase enzymes and the electron transport chain are located in the inner membrane. 79 Lesson 3: Glycolysis By the end of this unit, you will be able to do the following: 1. Describe the overall result in terms of molecules produced during the chemical breakdown of glucose by glycolysis 2. Compare the output of glycolysis in terms of ATP molecules and NADH molecules produced Nearly all of the energy used by living cells comes to them in the bonds of the sugar glucose. Glycolysis is the first step in the breakdown of glucose to extract energy for cellular metabolism. In fact, nearly all living organisms carry out glycolysis as part of their metabolism. The process does not use oxygen directly and therefore is termed anaerobic. Glycolysis takes place in the cytoplasm of both prokaryotic and eukaryotic cells. Glucose enters heterotrophic cells in two ways. One method is through secondary active transport in which the transport takes place against the glucose concentration gradient. The other mechanism uses a group of integral proteins called GLUT proteins, also known as glucose transporter proteins. These transporters assist in the facilitated diffusion of glucose. Glycolysis begins with the six-carbon ring-shaped structure of a single glucose molecule and ends with two molecules of a three-carbon sugar called pyruvate. Glycolysis consists of two distinct phases. The first part of the glycolysis pathway traps the glucose molecule in the cell and uses energy to modify it so that the six-carbon sugar molecule can be split evenly into the two three-carbon molecules. The second part of glycolysis extracts energy from the molecules and stores it in the form of ATP and NADH—remember: this is the reduced form of NAD. Each reaction in glycolysis is catalyzed by its own enzyme. The most important enzyme for regulation of glycolysis is phosphofructokinase, which catalyzes formation of the unstable, two-phosphate sugar molecule, fructose-1,6-bisphosphate. Phosphofructokinase speeds up or slows down glycolysis in response to the energy needs of the cell. First Half of Glycolysis (Energy-Requiring Steps) Step 1: The first step in glycolysis is catalyzed by hexokinase, an enzyme with broad specificity that catalyzes the phosphorylation of six-carbon sugars. Hexokinase phosphorylates glucose using ATP as the source of the phosphate, producing glucose-6-phosphate, a more reactive form of glucose. This reaction prevents the phosphorylated glucose molecule from continuing to interact with the GLUT proteins, and it can no longer leave the cell because the negatively charged phosphate will not allow it to cross the hydrophobic interior of the plasma membrane. 80 Step 2. In the second step of glycolysis, an isomerase converts glucose-6-phosphate into one of its isomers, fructose-6-phosphate (this isomer has a phosphate attached at the location of the sixth carbon of the ring). An isomerase is an enzyme that catalyzes the conversion of a molecule into one of its isomers. (This change from phosphoglucose to phosphofructose allows the eventual split of the sugar into two three-carbon molecules.) Step 3. The third step is the phosphorylation of fructose-6-phosphate, catalyzed by the enzyme phosphofructokinase. A second ATP molecule donates a high-energy phosphate to fructose-6phosphate, producing fructose-1,6-bisphosphate. In this pathway, phosphofructokinase is a rate-limiting enzyme. It is active when the concentration of ADP is high; it is less active when ADP levels are low and the concentration of ATP is high. Thus, if there is “sufficient” ATP in the system, the pathway slows down. This is a type of end product inhibition, since ATP is the end product of glucose catabolism. Step 4. The newly added high-energy phosphates further destabilize fructose-1,6-bisphosphate. The fourth step in glycolysis employs an enzyme, aldolase, to cleave fructose-1,6-bisphosphate into two three-carbon isomers: dihydroxyacetone phosphate (DHAP) and glyceraldehyde-3phosphate (GAP). 81 Step 5. In the fifth step, an isomerase transforms the dihydroxyacetone-phosphate into its isomer, glyceraldehyde-3-phosphate. Thus, the pathway will continue with two molecules of a glyceraldehyde-3-phosphate. At this point in the pathway, there is a net investment of energy from two ATP molecules in the breakdown of one glucose molecule. Second Half of Glycolysis (Energy-Releasing Steps) So far, glycolysis has cost the cell two ATP molecules and produced two small, threecarbon sugar molecules. Both of these molecules will proceed through the second half of the pathway, and sufficient energy will be extracted to pay back the two ATP molecules used as an initial investment and produce a profit for the cell of two additional ATP molecules and two even higher-energy NADH molecules. 2 2 Step 6. The sixth step in glycolysis oxidizes the 2 units of sugar (glyceraldehyde-3-phosphate), extracting high-energy electrons, which are picked up by the electron carrier NAD+, producing NADH. The sugar is then phosphorylated by the addition of a second phosphate group, producing 1,3-bisphosphoglycerate. Note that the second phosphate group does not require another ATP molecule. CAVEAT: Here again is a potential limiting factor for this pathway. The continuation of the reaction depends upon the availability of the oxidized form of the electron carrier, NAD +. Thus, NADH must be continuously oxidized back into NAD+ in order to keep this step going. If NAD + is not available, the second half of glycolysis slows down or stops. If oxygen is available in the system, the NADH will be oxidized readily, though indirectly, and the high-energy electrons from the hydrogen released in this process will be used to produce ATP. In an environment without oxygen, an alternate pathway (fermentation) can provide the oxidation of NADH to NAD +. 82 2 2 Step 7. In the seventh step, catalyzed by phosphoglycerate kinase (an enzyme named for the reverse reaction), two units of 1,3-bisphosphoglycerate donates a high-energy phosphate to ADP, forming one molecule of ATP. (This is an example of substrate-level phosphorylation). A carbonyl group (a functional group composed of a carbon atom double-bonded to an oxygen atom: C=O) on the 1,3-bisphosphoglycerate is oxidized to a carboxyl group, and 3phosphoglycerate is formed. 2 2 Step 8. In the eighth step, the remaining phosphate group in 3-phosphoglycerate moves from the third carbon to the second carbon, producing 2-phosphoglycerate (an isomer of 3phosphoglycerate). The enzyme catalyzing this step is a mutase (isomerase). 2 2 Step 9. Enolase catalyzes the ninth step. This enzyme causes 2-phosphoglycerate to lose water from its structure; this is a dehydration reaction, resulting in the formation of a double bond that increases the potential energy in the remaining phosphate bond and produces phosphoenolpyruvate (PEP). PEP is an unstable molecule, poised to lose its phosphate group in the final step of glycolysis. 83 Step 10. The last step in glycolysis is catalyzed by the enzyme pyruvate kinase (the enzyme in this case is named for the reverse reaction of pyruvate’s conversion into PEP) and results in the production of a second ATP molecule by substrate-level phosphorylation and the compound pyruvic acid (or its salt form, pyruvate). Many enzymes in enzymatic pathways are named for the reverse reactions, since the enzyme can catalyze both forward and reverse reactions. 84 Lesson 4: Oxidation of Pyruvate and the Citric Acid Cycle By the end of this section, you will be able to do the following: 1. Explain how a circular pathway, such as the citric acid cycle, fundamentally differs from a linear biochemical pathway, such as glycolysis 2. Describe how pyruvate, the product of glycolysis, is prepared for entry into the citric acid cycle If oxygen is available, aerobic respiration will go forward. In eukaryotic cells, the pyruvate molecules produced at the end of glycolysis are transported into the mitochondria, which are the sites of cellular respiration. There, pyruvate is transformed into an acetyl group that will be picked up and activated by a carrier compound called coenzyme A (CoA). The resulting compound is called acetyl CoA. CoA is derived from vitamin B5, pantothenic acid. Acetyl CoA can be used in a variety of ways by the cell, but its major function is to deliver the acetyl group derived from pyruvate to the next stage of the pathway in glucose catabolism. Breakdown of Pyruvate In order for pyruvate, the product of glycolysis, to enter the next pathway, it must undergo several changes. The conversion is a three-step process. Figure 4.8 Upon entering the mitochondrial matrix, a multienzyme complex converts pyruvate into acetyl CoA. In the process, carbon dioxide is released, and one molecule of NADH is formed. Step 1. A carboxyl group is removed from pyruvate, releasing a molecule of carbon dioxide into the surrounding medium. This reaction creates a two-carbon hydroxyethyl group bound to the enzyme (pyruvate dehydrogenase). We should note that this is the first of the six carbons from the original glucose molecule to be removed. (This step proceeds twice because there are two pyruvate molecules produced at the end of glycolysis for every molecule of glucose metabolized anaerobically; thus, two of the six carbons will have been removed at the end of both steps.) Step 2. The hydroxyethyl group is oxidized to an acetyl group, and the electrons are picked up by NAD+, forming NADH. The high-energy electrons from NADH will be used later to generate ATP. Step 3. The enzyme-bound acetyl group is transferred to CoA, producing a molecule of acetyl CoA. 85 Note that during the second stage of glucose metabolism, whenever a carbon atom is removed, it is bound to two oxygen atoms, producing carbon dioxide, one of the major end products of cellular respiration. Note that during the second stage of glucose metabolism, whenever a carbon atom is removed, it is bound to two oxygen atoms, producing carbon dioxide, one of the major end products of cellular respiration. If we consider the two pyruvates that enter from glycolysis (for each glucose molecule), we can summarize pyruvate oxidation as follows: 1. Two molecules of pyruvate are converted into two molecules of acetyl CoA. 2. Two carbons are released as carbon dioxide—out of the six originally present in glucose. 3. 2 NADH are generated from NAD +. Why make acetyl CoA? Acetyl CoA serves as fuel for the citric acid cycle in the next stage of cellular respiration. The addition of CoA helps activate the acetyl group, preparing it to undergo the necessary reactions to enter the citric acid cycle. Citric Acid Cycle In the presence of oxygen, acetyl CoA delivers its acetyl (2C) group to a four-carbon molecule, oxaloacetate, to form citrate, a six-carbon molecule with three carboxyl groups; this pathway will harvest the remainder of the extractable energy from what began as a glucose molecule and release the remaining four CO2 molecules. This single pathway is called by different names: the citric acid cycle (for the first intermediate formed—citric acid, or citrate— when acetate joins to the oxaloacetate), the TCA cycle (because citric acid or citrate and isocitrate are tricarboxylic acids), and the Krebs cycle, after Hans Krebs, who first identified the steps in the pathway in the 1930s in pigeon flight muscles. Like the conversion of pyruvate to acetyl CoA, the citric acid cycle takes place in the matrix of mitochondria. Almost all of the enzymes of the citric acid cycle are soluble, with the single exception of the enzyme succinate dehydrogenase, which is embedded in the inner membrane of the mitochondrion. Unlike glycolysis, the citric acid cycle is a closed loop: the last part of the pathway regenerates the compound used in the first step. The eight steps of the cycle are a series of redox, dehydration, hydration, and decarboxylation reactions that produce two carbon dioxide molecules, one GTP/ATP, and the reduced carriers NADH and FADH 2. This is considered an aerobic pathway because the NADH and FADH2 produced must transfer their electrons to the next pathway in the system, which will use oxygen. If this transfer does not occur, the oxidation steps of the citric acid cycle also do not occur. Note that the citric acid cycle produces very little ATP directly and does not directly consume oxygen. 86 Step 1. Prior to the first step, a transitional phase occurs during which pyruvic acid is converted to acetyl CoA. Then, the first step of the cycle begins: This condensation step combines the two-carbon acetyl group with a four-carbon oxaloacetate molecule to form a six-carbon molecule of citrate with the help of enzyme synthase. CoA is bound to a sulfhydryl group (-SH) and diffuses away to eventually combine with another acetyl group. This step is irreversible because it is highly exergonic. The rate of this reaction is controlled by negative feedback and the amount of ATP available. If ATP levels increase, the rate of this reaction decreases. If ATP is in short supply, the rate increases. Step 2. In step two, citrate loses one water molecule and gains another as citrate is converted into its isomer, isocitrate. Enzyme involved is called isomerase. Step 3. In step three, isocitrate is oxidized, producing a five-carbon molecule, α-ketoglutarate, along with a molecule of CO2 and two electrons, which reduce NAD + to NADH. This step is also 87 regulated by negative feedback from ATP and NADH and a positive effect of ADP. The enzyme catalyzing this step is isocitrate dehydrogenase. Step 4. Steps three and four are both oxidation and decarboxylation steps, which as we have seen, release electrons that reduce NAD + to NADH and release carboxyl groups that form CO2 molecules. Alpha-ketoglutarate is the product of step three, and a succinyl group is the product of step four. CoA binds with the succinyl group to form succinyl CoA. The enzyme, αketoglutarate dehydrogenase, which catalyzes step four is regulated by feedback inhibition of ATP, succinyl CoA, and NADH. Step 5. In step five, a phosphate group is substituted for coenzyme A, and a high-energy bond is formed. This energy is used in substrate-level phosphorylation (during the conversion of the succinyl group to succinate) to form either guanine triphosphate (GTP) or ATP. There are two forms of the enzyme, called isoenzymes, for this step, depending upon the type of animal tissue in which they are found. One form is found in tissues that use large amounts of ATP, such as heart and skeletal muscle. This form produces ATP. The second form of the enzyme is found in tissues that have a high number of anabolic pathways, such as liver. This form produces GTP. GTP is energetically equivalent to ATP; however, its use is more restricted. In particular, protein synthesis primarily uses GTP. 88 Step 6. Step six is a dehydration process that converts succinate into fumarate. Two hydrogen atoms are transferred to FAD, reducing it to FADH 2. (Note: the energy contained in the electrons of these hydrogens is insufficient to reduce NAD + but adequate to reduce FAD.) Unlike NADH, this carrier remains attached to the enzyme and transfers the electrons to the electron transport chain directly. This process is made possible by the localization of the enzyme (dehydrogenase) catalyzing this step inside the inner membrane of the mitochondrion. Step 7. Water is added by hydrolysis to fumarate during step seven, and malate is produced. The last step in the citric acid cycle regenerates oxaloacetate by oxidizing malate. Another molecule of NADH is then produced in the process. Products of the citric acid cycle Let’s take a step back and do some accounting, tracing the fate of the carbons that enter the citric acid cycle and counting the reduced electron carriers – NADH and FADH2 and ATP are produced. 1. Two cycles for each glucose molecule 2. Two CO2 released per cycle 3. 3 NADH are produces per cycle 4. 1 FADH2 is produced per cycle 5. 1 ATP or GTP is produced per cycle 89 Lesson 5: Electron Transport Chain By the end of this section, you will be able to do the following: 1. Describe how electrons move through the electron transport chain and explain what happens to their energy levels during this process 2. Explain how a proton (H+) gradient is established and maintained by the electron transport chain You have just read about two pathways in glucose catabolism—glycolysis and the citric acid cycle—that generate ATP. Most of the ATP generated during the aerobic catabolism of glucose, however, is not generated directly from these pathways. Instead, it is derived from a process that begins by moving electrons through a series of electron carriers that undergo redox reactions. This process causes hydrogen ions to accumulate within the intermembranous space. Therefore, a concentration gradient forms in which hydrogen ions diffuse out of the intermembranous space into the mitochondrial matrix by passing through ATP synthase. The current of hydrogen ions powers the catalytic action of ATP synthase, which phosphorylates ADP, producing ATP. Electron Transport Chain The electron transport chain (Figure X.X) is the last component of aerobic respiration and is the only part of glucose metabolism that uses atmospheric oxygen. Oxygen continuously diffuses into plant tissues (typically through stomata), as well as into fungi and bacteria; however, in animals, oxygen enters the body through a variety of respiratory systems. Electron transport is a series of redox reactions that resembles a relay race or bucket brigade in that electrons are passed rapidly from one component to the next, to the endpoint of the chain where the electrons reduce molecular oxygen and, along with associated protons, produces water. There are four complexes composed of proteins, labeled I through IV in Figure 4.9, and the aggregation of these four complexes, together with associated mobile, accessory electron carriers, is called the electron transport chain. The electron transport chain is present with multiple copies in the inner mitochondrial membrane of eukaryotes and within the plasma membrane of prokaryotes. Figure 4.9 The electron transport chain is a series of electron transporters embedded in the inner mitochondrial membrane that shuttles electrons from NADH and FADH2 to molecular oxygen. In the process, protons are pumped from the mitochondrial matrix to the intermembrane space, and oxygen is reduced to form water. 90 Complex I First, two electrons are carried to the first complex via NADH. This complex, labeled I below, is composed of flavin mononucleotide (FMN) and an iron-sulfur (Fe-S)-containing protein. FMN, which is derived from vitamin B2 (also called riboflavin), is one of several prosthetic groups or cofactors in the electron transport chain. A prosthetic group is a nonprotein molecule required for the activity of a protein. Prosthetic groups are organic or inorganic, non-peptide molecules bound to a protein that facilitate its function. Prosthetic groups include coenzymes, which are the prosthetic groups of enzymes. The enzyme in complex I is NADH dehydrogenase and is a very large protein, containing 45 amino acid chains. Complex I can pump four hydrogen ions across the membrane from the matrix into the intermembrane space, and it is in this way that the hydrogen ion gradient is established and maintained between the two compartments separated by the inner mitochondrial membrane. Q and Complex II Complex II (below) directly receives FADH2—which does not pass through complex I. The compound connecting the first and second complexes to the third is ubiquinone B. The Q molecule is lipid soluble and freely moves through the hydrophobic core of the membrane. Once it is reduced (QH2), ubiquinone delivers its electrons to the next complex in the electron transport chain. Q receives the electrons derived from NADH from complex I, and the electrons derived from FADH2 from complex II. This enzyme and FADH 2 form a small complex that delivers electrons directly to the electron transport chain, bypassing the first complex. Since these electrons bypass and thus do not energize the proton pump in the first complex, fewer ATP molecules are made from the FADH 2 electrons. The number of ATP molecules ultimately obtained is directly proportional to the number of protons pumped across the inner mitochondrial membrane. 91 FADH2 FAD Complex III The third complex (below) is composed of cytochrome b—another Fe-S protein, a Rieske center (2Fe-2S center), and cytochrome c proteins. This complex is also called cytochrome oxidoreductase. Cytochrome proteins have a prosthetic group of heme. The heme molecule is similar to the heme in hemoglobin, but it carries electrons, not oxygen. As a result, the iron ion at its core is reduced and oxidized as it passes the electrons, fluctuating between different oxidation states: Fe ++ (reduced) and Fe+++ (oxidized). The heme molecules in the cytochromes have slightly different characteristics due to the effects of the different proteins binding to them, giving slightly different characteristics to each complex. Complex III pumps protons through the membrane and passes its electrons to cytochrome c for transport to the fourth complex of proteins and enzymes. (Cytochrome c receives electrons from Q; however, whereas Q carries pairs of electrons, cytochrome c can accept only one at a time.) Complex IV The fourth complex (below) is composed of cytochrome proteins c, a, and a 3. This complex contains two heme groups (one in each of the two cytochromes, a, and a3) and three copper ions (a pair of Cu A and one CuB in cytochrome a3). The cytochromes hold an oxygen molecule very tightly between the iron and copper ions until the oxygen is completely reduced by the gain of two electrons. The reduced oxygen then picks up two hydrogen ions from the surrounding medium to make water (H2O). The removal of the hydrogen ions from the system contributes to the ion gradient that forms the foundation for the process of chemiosmosis. 92 Chemiosmosis In chemiosmosis, the free energy from the series of redox reactions just described is used to pump hydrogen ions (protons) across the mitochondrial membrane. The uneven distribution of H+ ions across the membrane establishes both concentration and electrical gradients (thus, an electrochemical gradient), owing to the hydrogen ions’ positive charge and their aggregation on one side of the membrane. If the membrane were continuously open to simple diffusion by the hydrogen ions, the ions would tend to diffuse back across into the matrix, driven by the concentrations producing their electrochemical gradient. Recall that many ions cannot diffuse through the nonpolar regions of phospholipid membranes without the aid of ion channels. Similarly, hydrogen ions in the matrix space can only pass through the inner mitochondrial membrane by an integral membrane protein called ATP synthase (Figure 4.10). This complex protein acts as a tiny generator, turned by the force of the hydrogen ions diffusing through it, down their electrochemical gradient. The turning of parts of this molecular machine facilitates the addition of a phosphate to ADP, forming ATP, using the potential energy of the hydrogen ion gradient. Figure 4.10 ATP synthase is a complex, molecular machine that uses a proton (H+) gradient to form ATP from ADP and inorganic phosphate (Pi). (Credit: modification of work by Klaus Hoffmeier) 93 Chemiosmosis (Figure 4.11) is used to generate 90 percent of the ATP made during aerobic glucose catabolism; it is also the method used in the light reactions of photosynthesis to harness the energy of sunlight in the process of photophosphorylation. Recall that the production of ATP using the process of chemiosmosis in mitochondria is called oxidative phosphorylation. The overall result of these reactions is the production of ATP from the energy of the electrons removed from hydrogen atoms. These atoms were originally part of a glucose molecule. At the end of the pathway, the electrons are used to reduce an oxygen molecule to oxygen ions. The extra electrons on the oxygen attract hydrogen ions (protons) from the surrounding medium, and water is formed. Thus, oxygen is the final electron acceptor in the electron transport chain. Figure 4.11 In oxidative phosphorylation, the pH gradient formed by the electron transport chain is used by ATP synthase to form ATP. ATP Yield The number of ATP molecules generated from the catabolism of glucose varies. For example, the number of hydrogen ions that the electron transport chain complexes can pump through the membrane varies between species. Another source of variance stems from the shuttle of electrons across the membranes of the mitochondria. (The NADH generated from glycolysis cannot easily enter mitochondria.) Thus, electrons are picked up on the inside of mitochondria by either NAD+ or FAD+. As you have learned earlier, these FAD+ molecules can transport fewer ions; consequently, fewer ATP molecules are generated when FAD+ acts as a carrier. NAD+ is used as the electron transporter in the liver and FAD+ acts in the brain. Another factor that affects the yield of ATP molecules generated from glucose is the fact that intermediate compounds in these pathways are also used for other purposes. Glucose catabolism connects with the pathways that build or break down all other biochemical compounds in cells, and the result is somewhat messier than the ideal situations described thus far. For example, sugars other than glucose are fed into the glycolytic pathway for energy extraction. In addition, the five-carbon sugars that form nucleic acids are made from intermediates in glycolysis. Certain nonessential amino acids can be made from intermediates of both glycolysis and the citric acid cycle. Lipids, such as cholesterol and triglycerides, are also made from intermediates in these pathways, and both amino acids and triglycerides are broken down for energy through these pathways. Overall, in living systems, these pathways of glucose 94 catabolism extract about 34 percent of the energy contained in glucose, with the remainder being released as heat. For this class, we will use the following values: 1. For every unit of FADH2, it will yield 1.5 ATP. 2. For every unit of NADH, it will yield 2.5 ATP. Hence, these are the over-all products of cellular respiration (aerobic): Glycolysis: 2 pyruvate, 2 NADH, 2 ATP Oxidative Decarboxylation: 2 NADH Krebs Cycle: 6 NADH, 2 FADH, 2 ATP Total ATP acquired for every molecule of glucose: 10 NADH x 2.5 ATP = 25 ATP 2 FADH x 1.5 ATP = 3 ATP 4 ATP = 4 ATP 32 ATP 95 Lesson 6: Metabolism without Oxygen By the end of this section, you will be able to do the following: 1. Discuss the fundamental difference between anaerobic cellular respiration and fermentation 2. Describe the type of fermentation that readily occurs in animal cells and the conditions that initiate that fermentation In aerobic respiration, the final electron acceptor is an oxygen molecule, O 2. If aerobic respiration occurs, then ATP will be produced using the energy of high-energy electrons carried by NADH or FADH2 to the electron transport chain. If aerobic respiration does not occur, NADH must be reoxidized to NAD + for reuse as an electron carrier for the glycolytic pathway to continue. How is this done? Some living systems use an organic molecule as the final electron acceptor. Processes that use an organic molecule to regenerate NAD + from NADH are collectively referred to as fermentation. In contrast, some living systems use an inorganic molecule as a final electron acceptor. Both methods are called anaerobic cellular respiration, in which organisms convert energy for their use in the absence of oxygen. Anaerobic Cellular Respiration Certain prokaryotes, including some species in the domains Bacteria and Archaea, use anaerobic respiration. For example, a group of archaeans called methanogens reduces carbon dioxide to methane to oxidize NADH. These microorganisms are found in soil and in the digestive tracts of ruminants, such as cows and sheep. Similarly, sulfate-reducing bacteria, most of which are anaerobic, reduce sulfate to hydrogen sulfide to regenerate NAD+ from NADH. Lactic Acid Fermentation The fermentation method used by animals and certain bacteria, such as those in yogurt, is lactic acid fermentation (Figure X.14). This type of fermentation is used routinely in mammalian red blood cells, which do not have mitochondria, and in skeletal muscle that has an insufficient oxygen supply to allow aerobic respiration to continue (that is, in muscles used to the point of fatigue). In muscles, lactic acid accumulation must be removed by the blood circulation, and when the lactic acid loses a hydrogen, the resulting lactate is brought to the liver for further metabolism. The chemical reactions of lactic acid fermentation are the following: The enzyme used in this reaction is lactate dehydrogenase (LDH). The reaction can proceed in either direction, but the reaction from left to right is inhibited by acidic conditions. Such lactic acid accumulation was once believed to cause muscle stiffness, fatigue, and soreness, although more recent research disputes this hypothesis. Once the lactic acid has been removed from the muscle and circulated to the liver, it can be reconverted into pyruvic acid and further catabolized for energy. 96 Figure 4.12 Lactic acid fermentation is common in muscle cells that have run out of oxygen. Clinical Importance • Tissues that function under hypoxic conditions will produce lactic acid from glucose oxidation. Produces local acidosis. If lactate production is more it can produce metabolic acidosis. • Vigorously contracting skeletal muscle will produce lactic acid. Alcohol Fermentation Another familiar fermentation process is alcohol fermentation, which produces ethanol. The first chemical reaction of alcohol fermentation is the following (CO2 does not participate in the second reaction): The first reaction is catalyzed by pyruvate decarboxylase, a cytoplasmic enzyme, with a coenzyme of thiamine pyrophosphate (TPP, derived from vitamin B 1 and also called thiamine). A carboxyl group is removed from pyruvic acid, releasing carbon dioxide as a gas. The loss of carbon dioxide reduces the size of the molecule by one carbon, producing acetaldehyde. The second reaction is catalyzed by alcohol dehydrogenase to oxidize NADH to NAD+ and reduce acetaldehyde to ethanol. The fermentation of pyruvic acid by yeast produces the ethanol found in alcoholic beverages. Ethanol tolerance of yeast is variable, ranging from about 5 percent to 21 percent, depending on the yeast strain and environmental conditions. Other Types of Fermentation Other fermentation methods take place in bacteria. We should note that many prokaryotes are facultative anaerobic. This means that they can switch between aerobic respiration and fermentation, depending on the availability of free oxygen. Certain prokaryotes, such as Clostridia, are obligate anaerobes. Obligate anaerobes live and grow in the absence of molecular oxygen. Oxygen is a poison to these microorganisms and kills them on exposure. We should also note that all forms of fermentation, except lactic acid fermentation, produce gas. The production of particular types of gas is used as an indicator of the fermentation of specific 97 carbohydrates, which plays a role in the laboratory identification of the bacteria. Various methods of fermentation are used by assorted organisms to ensure an adequate supply of NAD+ for the sixth step in glycolysis. Without these pathways, this step would not occur, and ATP could not be harvested from the breakdown of glucose. Cori Cycle With respect to energy, the liver and muscles act complementarily. The liver is the major organ in the body for the synthesis of glucose. Muscles are major users of ATP. Actively exercising muscles generate lactate as a result of running glycolysis faster than the blood can deliver oxygen during periods of heavy exercise. As a consequence, the muscles go anaerobic and produce lactate. This lactate is of no use to muscle cells, so they dump it into the blood. Lactate travels in the blood to the liver, which takes it up and reoxidizes it back to pyruvate, catalyzed by the enzyme lactate dehydrogenase. Pyruvate in the liver is then converted to glucose by gluconeogenesis (to be discussed in Lesson 8). The glucose thus made by the liver is dumped into the bloodstream where it is taken up by muscles and used for energy, completing a very important intercellular pathway known as the Cori cycle. Figure 4.13: The Cori Cycle (Credit: bio.libretexts.org) 98 Lesson 7: Connections of Carbohydrate, Protein, and Lipid Metabolic Pathways By the end of this section, you will be able to do the following: 1. Discuss the ways in which carbohydrate metabolic pathways, glycolysis, and the citric acid cycle interrelate with protein and lipid metabolic pathways 2. Explain why metabolic pathways are not considered closed systems You have learned about the catabolism of glucose, which provides energy to living cells. But living things consume organic compounds other than glucose for food. How does a chicken sandwich end up as ATP in your cells? This happens because all of the catabolic pathways for carbohydrates, proteins, and lipids eventually connect into glycolysis and the citric acid cycle pathways (see Figure 4.14). Metabolic pathways should be thought of as porous and interconnecting—that is, substances enter from other pathways, and intermediates leave for other pathways. These pathways are not closed systems! Many of the substrates, intermediates, and products in a particular pathway are reactants in other pathways. Figure 4.14 Glycogen from the liver and muscles, as well as other carbohydrates, hydrolyzed into glucose-1-phosphate, together with fats and proteins, can feed into the catabolic pathways for carbohydrates. Connections of Other Sugars to Glucose Metabolism Glycogen, a polymer of glucose, is an energy storage molecule in animals. When there is adequate ATP present, excess glucose is stored as glycogen in both liver and muscle cells. The glycogen will be hydrolyzed into glucose 1-phosphate monomers (G-1-P) if blood sugar levels drop. The presence of glycogen as a source of glucose allows ATP to be produced for a longer period of time during exercise. Glycogen is broken down into glucose-1-phosphate (G-1P) and converted into glucose-6-phosphate (G-6-P) in both muscle and liver cells, and this product enters the glycolytic pathway. Sucrose is a disaccharide with a molecule of glucose and a molecule of fructose bonded together with a glycosidic linkage. Fructose is one of the three “dietary” monosaccharides, along with glucose and galactose (part of the milk sugar dissacharide lactose), which are absorbed directly into the bloodstream during digestion. The catabolism of both fructose and galactose produces the same number of ATP molecules as glucose. 99 Glycogenesis Glycogen is a polymer of glucose residues linked by α-(1,4)- and α-(1,6)-glycosidic bonds. Stores of readily available glucose, to supply the tissues of the body with an oxidizable energy source, are found as glycogen, solely in the liver. Although the liver is the only tissue that can release glucose from glycogen to the blood, other tissues also synthesize glycogen and release glucose from glycogen but this glucose is used for direct cellular energy needs. Two primary tissues that store glucose as glycogen, as a store of energy, are skeletal muscle and the brain, principally astrocytes, however, the kidney, heart, and adipose tissue also store glucose as glycogen. The glucose in muscle and brain glycogen, and other non-hepatic tissues, is not available to other tissues, because of the presence of hexokinase which has a very high affinity for glucose, thereby rapidly phosphorylating any glucose as well as due to the lack of glucose-6-phosphatase. Figure 4.15. Glycogen Structure. Highly simplified section of a glycogen granule depicting glucose monomers as colored balls. The blue balls represent glucose linked by α1,4 glycosidic bonds. The red balls represent glucose at branch points where there are both α1,4 and α1,6 glycosidic bonds. The orange balls represent the reducing ends of the polymeric chains of α1,4-linked glucoses. The area in the box is expanded to show the actual structure of the glucose monomers in both α-1,4- and α-1,6 glycosidic linkages (Credit: http://themedicalbiochemistrypage.org/) The liver is a so-called "altruistic" organ, which releases glucose into the blood to meet tissue need. Glucose released from muscle glycogen stores is used on site to provide energy for muscle contraction. Like glycolysis and gluconeogenesis, glycogenolysis and glycogenesis are NOT reversals of each other. It is the hormone insulin that triggers glycogenesis. This particular hormone has a huge impact on the metabolism of glucose in the liver cells. It stimulates glycogenesis and at the same time inhibit the breakdown of glycogen into glucose (glycogenolysis). These are the steps in Glycogenesis: 1. The first step in glycogen synthesis is glucose activation to glucose-6-phosphate, in the reaction catalyzed by glucokinase in the liver and hexokinase in the muscle and the other organs and tissues. 100 2. When glycogen synthesis is occurring phosphoglucomutase, the same enzyme that acts also during glycogenolysis catalyzing the conversion of glucose 1-phosphate to glucose-6phosphate, shifts the phosphate group from C6 to C1 (therefore the enzyme catalyzes a reversible reaction). 3. The next step is UDP-glucose production from glucose-1-phosphate and UTP in the reaction catalyzed by UDP-glucose pyrophosphorylase; the reaction, reversible becomes irreversible thanks to the rapid hydrolysis of pyrophosphate to inorganic phosphate in the reaction catalyzed by pyrophosphatase. 4. Then glycogen synthase transfers the activated glucose to 4’-OH group of a glucose residue (a nonreducing ends) present in the molecule catalyzing the formation of an α-(1,4) glycosidic bond and therefore extending the chain by one glucose unit. 5. The branches are inserted in the reaction catalyzed by the branching enzyme, also called amylo-α-(1,4)→α-(1,6)-transglucosidase (EC 2.4.1.18), that catalyzes the transfer en bloc of an oligosaccharide of six to seven glucose units from a nonreducing termini of a newly elongated chain of at least eleven units to another chain forming a new α-(1,6) glycosidic bond. The new branches are introduced at least at four glucose residues from an adjacent branch point. 6. Then, glycogen synthase may add further glucose residues to the new branch. 101 Figure 4.16. Glycogenesis pathway (Credit: laboratoryinfo.com and tuscany-diet.net) Glycogenolysis Glycogenolysis is defined as metabolism of glycogen polymers occurred during fasting. The glycogen is broken down in the liver, kidney or muscles into glucose or to glucose-6phosphate for use in glycolysis pathway. Here are the steps in Glycogenolysis: 1. Phosphorolysis/Shoterning of chains Glycogen is a branched polymer of glucose units in chains are linked by α-1,4-glycosidic bonds with a branch point created by α-1,6-glycosidic bonds at approximately every 10 residues of glucose. The key enzyme for glycogenolysis, glycogen phosphorylase, will cleave the α-1,4glycosidic bonds of the terminal glucose residues at the non-reducing end of glycogen (i.e. the end of the glycogen molecule with a free 4-OH group (Figure 4.17) until only four glucosyl units remain on each chain before a branch point. Figure 4.17 Structure of Glycogen (Credit: teaching.ncl.ac.uk (Berg, J.M., Tymoczko, J.L and Stryer, L.)) Orthophosphate (Pi), an inorganic phosphate, cleaves the the glycosidic bond between C1 of the terminal residue and C4 of the adjacent glycogen molecule via phosphorolysis to yield glucose 1-phosphate. The α configuration at C1 is retained even after the glycogen 102 phosphorylase cleaves the bond between the C1 carbon atom and the glycosidic oxygen atom (refer to Figure 4.19). Figure 4.18 Equation for phosphorolysis(Credit: teaching.ncl.ac.uk (Berg, J.M., Tymoczko, J.L and Stryer, L.)) Figure 4.19 Phosphorolysis by an orthophosphate (inorganic phosphate) (Credit: teaching.ncl.ac.uk (Berg, J.M., Tymoczko, J.L and Stryer, L.)) 2. Debranching/Removal of branches Glycogen phosphorylase can only carry out the glycogen breakdown process by itself until a limited extent before encountering an obstacle. When phophorylase reaches a terminal residue four residues away from a branch point (i.e. after release of six glucose molecules), it will stop cleaving and the α-1,6 linkages are not susceptible to cleavage by phosphorylase. The branches of the glycogen molecule are removed by the debranching enzyme, a single bifunctional protein with two enzymic activities. The debranching enzyme can act as a transferase as well as an α-1,6-glucosidase to aid the continued degradation by phosphorylase. A block of three glycosyl residues from one outer branch was shifted by the transferase. The remaining single glucose molecule has a α-1,6-glycosidic bond joined to the glycogen molecule. The debranching enzyme, which act as α-1,6-glucosidase will cleave the linkage and results in the release of a free glucose molecule. The glycolytic enzyme, hexokinase, will phosphorylate this free glucose molecule. Thus, the net result is linear structures which can be continually degraded by glycogen phosphorylase. 103 Figure 4.20 Glycogen remodelling in glycogenolysis (Step 1 and step 2) (Credit: teaching.ncl.ac.uk (Berg, J.M., Tymoczko, J.L and Stryer, L.)) 3. Recovery As in the glycolysis pathway, phosphoglucomutase is used to convert glucose 1phosphate formed in the cleavage of glycogen into glucose 6-phosphate to enter the metabolic mainstream. 4. Release This process will only occur in liver. In contrast with glucose, the phosphorylated glucose produce in the glycogen breakdown is not readily to be transported out of the cell. The liver contains a hydrolytic enzyme, glucose 6-phosphatase, located on the luminal side of smooth endoplasmic reticulum, which convert the glucose 6-phosphate into glucose by cleaving the phosphoryl group. Muscle cells, lack the enzyme that can make glucose from glucose 6-phosphate. Skeletal muscle, in fasted state, will metabolise glucose 6-phosphate to either pyruvate (aerobic conditions) or lactate (anaerobic conditions). Effect of Hormone on Glycogenolysis Metabolic effects of glucagon on glycogen breakdown – Primary target of glucagon is liver as glucagon receptors are not found on skeletal muscle. In fasted state, glucagon will be produced in order to increase the plasma concentration of glucose. Under the influence of glucagon, enzymes that break down glycogen become more active but the enzymes for glycogen synthesis (glycogenesis) will become less active or inhibited. Stimulation of glycogen breakdown by epinephrine/adrenaline – An epinephrine/adrenaline molecule binds to a β-adrenergic receptor on the plasma membrane of a liver or muscle cell. The G protein neighbouring the receptor is activated and it in turn activate stimulate the adenylyl cyclase. Adenylyl cyclase will then generate cAMP from ATP. The 104 increase of cAMP in the cytosol will activates the protein kinase A (PKA). PKA then increases the phosphorylation of the enzyme phosphorylase kinase. Glycogen phosphorylase will then be converted from phosphorylase b, the less active form, to phosphorylase a, the more active form. The rate of glycogen breakdown will increase significantly. Connections of Proteins to Glucose Metabolism Proteins are hydrolyzed by a variety of enzymes in cells. Most of the time, the amino acids are recycled into the synthesis of new proteins. If there are excess amino acids, however, or if the body is in a state of starvation, some amino acids will be shunted into the pathways of glucose catabolism (Figure 4.21). It is very important to note that each amino acid must have its amino group removed prior to entry into these pathways. The amino group is converted into ammonia. In mammals, the liver synthesizes urea from two ammonia molecules and a carbon dioxide molecule. Thus, urea is the principal waste product in mammals, produced from the nitrogen originating in amino acids, and it leaves the body in urine. It should be noted that amino acids can be synthesized from the intermediates and reactants in the cellular respiration cycle. Figure 4.21 The carbon skeletons of certain amino acids (indicated in boxes) derived from proteins can feed into the citric acid cycle. (credit: modification of work by Mikael Häggström) . Connections of Lipid and Glucose Metabolisms The lipids connected to the glucose pathway include cholesterol and triglycerides. Cholesterol is a lipid that contributes to cell membrane flexibility and is a precursor of steroid hormones. The synthesis of cholesterol starts with acetyl groups and proceeds in only one direction. The process cannot be reversed. Triglycerides—made from the bonding of glycerol and three fatty acids—are a form of long-term energy storage in animals. Animals can make most of the fatty acids they need. 105 Triglycerides can be both made and broken down through parts of the glucose catabolism pathways. Glycerol can be phosphorylated to glycerol-3-phosphate, which continues through glycolysis. Fatty acids are catabolized in a process called beta-oxidation, which takes place in the matrix of the mitochondria and converts their fatty acid chains into two-carbon units of acetyl groups. The acetyl groups are picked up by CoA to form acetyl CoA that proceeds into the citric acid cycle. 106 Lesson 8: Gluconeogenesis Gluconeogenesis is the synthesis of new glucose molecules from pyruvate, lactate, glycerol, or the amino acids alanine or glutamine. This process takes place primarily in the liver during periods of low glucose, that is, under conditions of fasting, starvation, and low carbohydrate diets. So, the question can be raised as to why the body would create something it has just spent a fair amount of effort to break down? Certain key organs, including the brain, can use only glucose as an energy source; therefore, it is essential that the body maintain a minimum blood glucose concentration. When the blood glucose concentration falls below that certain point, new glucose is synthesized by the liver to raise the blood concentration to normal. Gluconeogenesis is not simply the reverse of glycolysis. There are some important differences (Figure 4.22). Pyruvate is a common starting material for gluconeogenesis. First, the pyruvate is converted into oxaloacetate. Oxaloacetate then serves as a substrate for the enzyme phosphoenolpyruvate carboxykinase (PEPCK), which transforms oxaloacetate into phosphoenolpyruvate (PEP). From this step, gluconeogenesis is nearly the reverse of glycolysis. PEP is converted back into 2-phosphoglycerate, which is converted into 3phosphoglycerate. Then, 3-phosphoglycerate is converted into 1,3 bisphosphoglycerate and then into glyceraldehyde-3-phosphate. Two molecules of glyceraldehyde-3-phosphate then combine to form fructose-1-6-bisphosphate, which is converted into fructose 6-phosphate and then into glucose-6-phosphate. Finally, a series of reactions generates glucose itself. In gluconeogenesis (as compared to glycolysis), the enzyme hexokinase is replaced by glucose-6phosphatase, and the enzyme phosphofructokinase-1 is replaced by fructose-1,6bisphosphatase. This helps the cell to regulate glycolysis and gluconeogenesis independently of each other. As will be discussed as part of lipolysis, fats can be broken down into glycerol, which can be phosphorylated to form dihydroxyacetone phosphate or DHAP. DHAP can either enter the glycolytic pathway or be used by the liver as a substrate for gluconeogenesis. 107 Figure 4.22. Gluconeogenesis. Gluconeogenesis is the synthesis of glucose from pyruvate, lactate, glycerol, alanine, or glutamate (Credit: opentextbc.ca) Body’s Metabolic Rate The human body’s metabolic rate decreases nearly 2 percent per decade after age 30. Changes in body composition, including reduced lean muscle mass, are mostly responsible for this decrease. The most dramatic loss of muscle mass, and consequential decline in metabolic rate, occurs between 50 and 70 years of age. Loss of muscle mass is the equivalent of reduced strength, which tends to inhibit seniors from engaging in sufficient physical activity. This results in a positive-feedback system where the reduced physical activity leads to even more muscle loss, further reducing metabolism. There are several things that can be done to help prevent general declines in metabolism and to fight back against the cyclic nature of these declines. These include eating breakfast, eating small meals frequently, consuming plenty of lean protein, drinking water to 108 remain hydrated, exercising (including strength training), and getting enough sleep. These measures can help keep energy levels from dropping and curb the urge for increased calorie consumption from excessive snacking. While these strategies are not guaranteed to maintain metabolism, they do help prevent muscle loss and may increase energy levels. Some experts also suggest avoiding sugar, which can lead to excess fat storage. Spicy foods and green tea might also be beneficial. Because stress activates cortisol release, and cortisol slows metabolism, avoiding stress, or at least practicing relaxation techniques, can also help. 109 Lesson 8: Lipid Metabolism By the end of this section, you will be able to: 1. Explain how energy can be derived from fat 2. Explain the purpose and process of ketogenesis 3. Describe the process of ketone body oxidation 4. Explain the purpose and the process of lipogenesis Beta Oxidation To obtain energy from fat, triglycerides must first be broken down by hydrolysis into their two principal components, fatty acids and glycerol. This process, called lipolysis, takes place in the cytoplasm. The resulting fatty acids are oxidized by β-oxidation into acetyl CoA, which is used by the Krebs cycle. The glycerol that is released from triglycerides after lipolysis directly enters the glycolysis pathway as DHAP. Because one triglyceride molecule yields three fatty acid molecules with as much as 16 or more carbons in each one, fat molecules yield more energy than carbohydrates and are an important source of energy for the human body. Triglycerides yield more than twice the energy per unit mass when compared to carbohydrates and proteins. Therefore, when glucose levels are low, triglycerides can be converted into acetyl CoA molecules and used to generate ATP through aerobic respiration. The breakdown of fatty acids, called fatty acid oxidation or beta (β)-oxidation, begins in the cytoplasm, where fatty acids are converted into fatty acyl CoA molecules. This fatty acyl CoA combines with carnitine to create a fatty acyl carnitine molecule, which helps to transport the fatty acid across the mitochondrial membrane. Once inside the mitochondrial matrix, the fatty acyl carnitine molecule is converted back into fatty acyl CoA and then into acetyl CoA (Figure 4.23). The newly formed acetyl CoA enters the Krebs cycle and is used to produce ATP in the same way as acetyl CoA derived from pyruvate. 110 Figure 4.23. Breakdown of Fatty Acids. During fatty acid oxidation, triglycerides can be broken down into acetyl CoA molecules and used for energy when glucose levels are low (Credit: www.abcam.com) ATP synthesis Acetyl-CoA generated by the beta-oxidation pathway enters the mitochondrial TCA cycle, where is further oxidized to generate NADH and FADH 2. The NADH and FADH2 produced by both beta oxidation and the TCA cycle are used by the mitochondrial electron transport chain to produce ATP. Complete oxidation of one palmitate molecule (fatty acid containing 16 carbons) generates 129 ATP molecules. Ketogenesis If excessive acetyl CoA is created from the oxidation of fatty acids and the Krebs cycle is overloaded and cannot handle it, the acetyl CoA is diverted to create ketone bodies. These ketone bodies can serve as a fuel source if glucose levels are too low in the body. Ketones serve as fuel in times of prolonged starvation or when patients suffer from uncontrolled diabetes and cannot utilize most of the circulating glucose. In both cases, fat stores are liberated to generate energy through the Krebs cycle and will generate ketone bodies when too much acetyl CoA accumulates. In this ketone synthesis reaction, excess acetyl CoA is converted into hydroxymethylglutaryl CoA (HMG CoA). HMG CoA is a precursor of cholesterol and is an intermediate that is subsequently converted into β-hydroxybutyrate, the primary ketone body in the blood (Figure 4.24). 111 Figure 4.24 Ketogenesis Excess acetyl CoA is diverted from the Krebs cycle to the ketogenesis pathway. This reaction occurs in the mitochondria of liver cells. The result is the production of β-hydroxybutyrate, the primary ketone body found in the blood. Ketone Body Oxidation Organs that have classically been thought to be dependent solely on glucose, such as the brain, can actually use ketones as an alternative energy source. This keeps the brain functioning when glucose is limited. When ketones are produced faster than they can be used, they can be broken down into CO2 and acetone. The acetone is removed by exhalation. One symptom of ketogenesis is that the patient’s breath smells sweet like alcohol. This effect provides one way of telling if a diabetic is properly controlling the disease. The carbon dioxide produced can acidify the blood, leading to diabetic ketoacidosis, a dangerous condition in diabetics. Ketones oxidize to produce energy for the brain. beta (β)-hydroxybutyrate is oxidized to acetoacetate and NADH is released. An HS-CoA molecule is added to acetoacetate, forming acetoacetyl CoA. The carbon within the acetoacetyl CoA that is not bonded to the CoA then detaches, splitting the molecule in two. This carbon then attaches to another free HS-CoA, resulting in two acetyl CoA molecules. These two acetyl CoA molecules are then processed through the Krebs cycle to generate energy (Figure 4.25). Figure 4.25 Ketone Oxidation When glucose is limited, ketone bodies can be oxidized to produce acetyl CoA to be used in the Krebs cycle to generate energy. 112 Lipogenesis When glucose levels are plentiful, the excess acetyl CoA generated by glycolysis can be converted into fatty acids, triglycerides, cholesterol, steroids, and bile salts. This process, called lipogenesis, creates lipids (fat) from the acetyl CoA and takes place in the cytoplasm of adipocytes (fat cells) and hepatocytes (liver cells). When you eat more glucose or carbohydrates than your body needs, your system uses acetyl CoA to turn the excess into fat. Although there are several metabolic sources of acetyl CoA, it is most commonly derived from glycolysis. Acetyl CoA availability is significant, because it initiates lipogenesis. Lipogenesis begins with acetyl CoA and advances by the subsequent addition of two carbon atoms from another acetyl CoA; this process is repeated until fatty acids are the appropriate length. Because this is a bond-creating anabolic process, ATP is consumed. However, the creation of triglycerides and lipids is an efficient way of storing the energy available in carbohydrates. Triglycerides and lipids, high-energy molecules, are stored in adipose tissue until they are needed. Although lipogenesis occurs in the cytoplasm, the necessary acetyl CoA is created in the mitochondria and cannot be transported across the mitochondrial membrane. To solve this problem, pyruvate is converted into both oxaloacetate and acetyl CoA. Two different enzymes are required for these conversions. Oxaloacetate forms via the action of pyruvate carboxylase, whereas the action of pyruvate dehydrogenase creates acetyl CoA. Oxaloacetate and acetyl CoA combine to form citrate, which can cross the mitochondrial membrane and enter the cytoplasm. In the cytoplasm, citrate is converted back into oxaloacetate and acetyl CoA. Oxaloacetate is converted into malate and then into pyruvate. Pyruvate crosses back across the mitochondrial membrane to wait for the next cycle of lipogenesis. The acetyl CoA is converted into malonyl CoA that is used to synthesize fatty acids. Figure 4.26 summarizes the pathways of lipid metabolism. 113 Figure 4.26 Lipid Metabolism Lipids may follow one of several pathways during metabolism. Glycerol and fatty acids follow different pathways. 114 Lesson 10. Protein Metabolism By the end of this section, you will be able to: 1. Explain how the urea cycle prevents toxic concentrations of nitrogen 2. Differentiate between glucogenic and ketogenic amino acids 3. Explain how protein can be used for energy Urea Cycle The urea cycle is a set of biochemical reactions that produces urea from ammonium ions in order to prevent a toxic level of ammonium in the body. It occurs primarily in the liver and, to a lesser extent, in the kidney. Prior to the urea cycle, ammonium ions are produced from the breakdown of amino acids. In these reactions, an amine group, or ammonium ion, from the amino acid is exchanged with a keto group on another molecule. This transamination event creates a molecule that is necessary for the Krebs cycle and an ammonium ion that enters into the urea cycle to be eliminated. In the urea cycle, ammonium is combined with CO 2, resulting in urea and water. The urea is eliminated through the kidneys in the urine (Figure 4.27). Let us make an in-depth study of the urea cycle. Here are the steps of Urea cycle: 1. Formation of Carbamoyl Phosphate: Condensation of ammonium ion with bicarbonate ion resulting in the formation of carbamoyl phosphate by the help of the enzyme carbamoyl phosphate synthase-I present in the liver mitochondria. It requires Mg2+ and a dicarboxylic acid i.e. N-acetyl glutamate. This step requires 2 ATPs. 2. Synthesis of Citrulline: Carbamoyl phosphate formed in the first step combines with ornithine resulting in the synthesis of citrulline aided by the enzyme citrulline synthase or ornithine transcarbamoylase. Citrulline is easily permeable to the mitochondrial membrane and hence it diffuses into the cytosol. 3. Synthesis of Argininosuccinate: In the cytosol, citrulline combines with the amino acid aspartate forming argininosuccinate catalysed by the enzyme argininosuccinate synthase. It requires ATP which is hydrolysed to AMP resulting in utilization of two high energy bonds. Mg 2+ acts as cofactor. 4. Cleavage of Argininosuccinate: The enzyme argininosuccinase acts reversibly to cleave arginino-succinate into Arginine and fumarate. Fumarate enters the TCA cycle (the linkage between TCA and urea cycle is known as Krebs bi-cycle). 5. Cleavage of Arginine: 115 Arginine is lysed into ornithine and urea under the influence of the enzyme arginase. Hence arginine is known as semi-essential amino acid i.e. though it is synthesized in the body it is not available for protein synthesis. Ornithine is regenerated in this step and the urea cycle completes by the formation of urea. Ornithine and lysine are potent inhibitors of the enzyme arginase. Arginase is also present in testis, renal tubules, mammary gland and skin in minute quantities. The intermediate amino acids formed in the urea cycle i.e. ornithine, citrulline and argininosuccinate are known as non-protein amino acids. The overall equation of urea formation is: NH4 + CO2 + Asparate + 3ATP → Urea + Fumarate + 2ADP + 2Pi + AMP + PPi The urea cycle brings two amino groups and HCO3 together to form urea. Thus toxic, insoluble ammonia is converted into non-toxic, water soluble, excretable urea. Hence, urea cycle disposes two waste products i.e. NH 4 and HCO3. This fact suggests that urea cycle participates in the regulation of blood pH, which depends on the HCO 3/H2CO3. Though 3 ATPs are utilized, the ultimate cost of making a molecule of urea is 4 ATPs (one ATP is converted into AMP). The rate limiting steps of urea cycle are 1, 2, & 5. Figure 4.27 Urea Cycle Nitrogen is transaminated, creating ammonia and intermediates of the Krebs cycle. Ammonia is processed in the urea cycle to produce urea that is eliminated through the kidneys. 116 Amino acids can also be used as a source of energy, especially in times of starvation. Because the processing of amino acids results in the creation of metabolic intermediates, including pyruvate, acetyl CoA, acetoacyl CoA, oxaloacetate, and α-ketoglutarate, amino acids can serve as a source of energy production through the Krebs cycle (Figure 4.28). Figure 4.29 summarizes the pathways of catabolism and anabolism for carbohydrates, lipids, and proteins. Figure 4.28 Energy from Amino Acids Amino acids can be broken down into precursors for glycolysis or the Krebs cycle. Amino acids (in bold) can enter the cycle through more than one pathway. 117 Figure 4.29 Catabolic and Anabolic Pathways Nutrients follow a complex pathway from ingestion through anabolism and catabolism to energy production. 118 UNIT SUMMARY Overview of Metabolic Reactions Metabolism is the sum of all catabolic (break down) and anabolic (synthesis) reactions in the body. The metabolic rate measures the amount of energy used to maintain life. An organism must ingest a sufficient amount of food to maintain its metabolic rate if the organism is to stay alive for very long. Catabolic reactions break down larger molecules, such as carbohydrates, lipids, and proteins from ingested food, into their constituent smaller parts. They also include the breakdown of ATP, which releases the energy needed for metabolic processes in all cells throughout the body. Anabolic reactions, or biosynthetic reactions, synthesize larger molecules from smaller constituent parts, using ATP as the energy source for these reactions. Anabolic reactions build bone, muscle mass, and new proteins, fats, and nucleic acids. Errors in metabolism alter the processing of carbohydrates, lipids, proteins, and nucleic acids, and can result in a number of disease states. Energy in Living Systems ATP functions as the energy currency for cells. It allows the cell to store energy briefly and transport it within the cell to support endergonic chemical reactions. The structure of ATP is that of an RNA nucleotide with three phosphates attached. As ATP is used for energy, a phosphate group or two are detached, and either ADP or AMP is produced. Energy derived from glucose catabolism is used to convert ADP into ATP. When ATP is used in a reaction, the third phosphate is temporarily attached to a substrate in a process called phosphorylation. The two processes of ATP regeneration that are used in conjunction with glucose catabolism are substrate-level phosphorylation and oxidative phosphorylation through the process of chemiosmosis. Glycolysis Glycolysis is the first pathway within the cytoplasm used in the breakdown of glucose to extract energy. It was probably one of the earliest metabolic pathways to evolve and is used by nearly all of the organisms on Earth. Glycolysis consists of two parts: The first part prepares the six-carbon ring of glucose for cleavage into two three-carbon sugars. ATP is invested in the process during this half to energize the separation. The second half of glycolysis extracts ATP and high-energy electrons from hydrogen atoms and attaches them to NAD +. Two ATP molecules are invested in the first half and four ATP molecules are formed by substrate phosphorylation during the second half. This produces a net gain of two ATP and two NADH molecules for the cell. Oxidation of Pyruvate and the Citric Acid Cycle In the presence of oxygen, pyruvate is transformed into an acetyl group attached to a carrier molecule of coenzyme A. The resulting acetyl CoA can enter several pathways, but most often, the acetyl group is delivered to the citric acid cycle for further catabolism. During the conversion of pyruvate into the acetyl group, a molecule of carbon dioxide and two high-energy electrons are removed. The carbon dioxide accounts for two (conversion of two pyruvate 119 molecules) of the six carbons of the original glucose molecule. The electrons are picked up by NAD+, and the NADH carries the electrons to a later pathway for ATP production. At this point, the glucose molecule that originally entered cellular respiration has been completely oxidized. Chemical potential energy stored within the glucose molecule has been transferred to electron carriers or has been used to synthesize a few ATPs. The citric acid cycle is a series of redox and decarboxylation reactions that removes high-energy electrons and carbon dioxide. The electrons, temporarily stored in molecules of NADH and FADH2, are used to generate ATP in a subsequent pathway. One molecule of either GTP or ATP is produced by substrate-level phosphorylation on each turn of the cycle. There is no comparison of the cyclic pathway with a linear one. Oxidative Phosphorylation The electron transport chain is the portion of aerobic respiration that uses free oxygen as the final electron acceptor of the electrons removed from the intermediate compounds in glucose catabolism. The electron transport chain is composed of four large, multiprotein complexes embedded in the inner mitochondrial membrane and two small diffusible electron carriers shuttling electrons between them. The electrons are passed through a series of redox reactions, with a small amount of free energy used at three points to transport hydrogen ions across a membrane. This process contributes to the gradient used in chemiosmosis. The electrons passing through the electron transport chain gradually lose energy. High-energy electrons donated to the chain by either NADH or FADH 2 complete the chain, as low-energy electrons reduce oxygen molecules and form water. The end products of the electron transport chain are water and ATP. A number of intermediate compounds of the citric acid cycle can be diverted into the anabolism of other biochemical molecules, such as non-essential amino acids, sugars, and lipids. These same molecules can serve as energy sources for the glucose pathways. Metabolism without Oxygen If NADH cannot be oxidized through aerobic respiration, another electron acceptor is used. Most organisms will use some form of fermentation to accomplish the regeneration of NAD+, ensuring the continuation of glycolysis. The regeneration of NAD+ in fermentation is not accompanied by ATP production; therefore, the potential of NADH to produce ATP using an electron transport chain is not utilized. Connections of Carbohydrate, Protein, and Lipid Metabolic Pathways The breakdown and synthesis of carbohydrates, proteins, and lipids connect with the pathways of glucose catabolism. The simple sugars are galactose, fructose, glycogen, and pentose. These are catabolized during glycolysis. The amino acids from proteins connect with glucose catabolism through pyruvate, acetyl CoA, and components of the citric acid cycle. Cholesterol synthesis starts with acetyl groups, and the components of triglycerides come from glycerol-3-phosphate from glycolysis and acetyl groups produced in the mitochondria from pyruvate. Lipid Metabolism Lipids are available to the body from three sources. They can be ingested in the diet, stored in the adipose tissue of the body, or synthesized in the liver. Fats ingested in the diet are 120 digested in the small intestine. The triglycerides are broken down into monoglycerides and free fatty acids, then imported across the intestinal mucosa. Once across, the triglycerides are resynthesized and transported to the liver or adipose tissue. Fatty acids are oxidized through fatty acid or β-oxidation into two-carbon acetyl CoA molecules, which can then enter the Krebs cycle to generate ATP. If excess acetyl CoA is created and overloads the capacity of the Krebs cycle, the acetyl CoA can be used to synthesize ketone bodies. When glucose is limited, ketone bodies can be oxidized and used for fuel. Excess acetyl CoA generated from excess glucose or carbohydrate ingestion can be used for fatty acid synthesis or lipogenesis. Acetyl CoA is used to create lipids, triglycerides, steroid hormones, cholesterol, and bile salts. Lipolysis is the breakdown of triglycerides into glycerol and fatty acids, making them easier for the body to process. Protein Metabolism Digestion of proteins begins in the stomach, where HCl and pepsin begin the process of breaking down proteins into their constituent amino acids. As the chyme enters the small intestine, it mixes with bicarbonate and digestive enzymes. The bicarbonate neutralizes the acidic HCl, and the digestive enzymes break down the proteins into smaller peptides and amino acids. Digestive hormones secretin and CCK are released from the small intestine to aid in digestive processes, and digestive proenzymes are released from the pancreas (trypsinogen and chymotrypsinogen). Enterokinase, an enzyme located in the wall of the small intestine, activates trypsin, which in turn activates chymotrypsin. These enzymes liberate the individual amino acids that are then transported via sodium-amino acid transporters across the intestinal wall into the cell. The amino acids are then transported into the bloodstream for dispersal to the liver and cells throughout the body to be used to create new proteins. When in excess, the amino acids are processed and stored as glucose or ketones. The nitrogen waste that is liberated in this process is converted to urea in the urea acid cycle and eliminated in the urine. In times of starvation, amino acids can be used as an energy source and processed through the Krebs cycle. 121 UNIT CRITICAL THINKING QUESTIONS Write your answers in a whole sheet of clean yellow paper. Write your name, section, unit number. 1. Nearly all organisms on Earth carry out some form of glycolysis. How does this fact support or not support the assertion that glycolysis is one of the oldest metabolic pathways? 2. What is the primary difference between a circular pathway and a linear pathway? 3. What is the primary difference between fermentation and anaerobic respiration? 4. How does citrate from the citric acid cycle affect glycolysis? 5. Why might negative feedback mechanisms be more common than positive feedback mechanisms in living cells? 122 123 124 References Biochemistry Module. Carbohydrate Metabolism. https://nios.ac.in/media/documents/dmlt/Biochemistry/Lesson-03.pdf Khan Academy. Biomolecules. https://www.khanacademy.org/test-prep/mcat/biomolecules BC campus. Anatomy and Physiology. Open Textbooks. Rice University. https://openstax.org/books/biology-2e/pages/1-introduction 125 Welcome to the Nutritional Biochemistry laboratory! Laboratory! Here, you will do laboratory activities in your home! Isn’t amazing? Isn’t surprising? Isn’t it? Don’t worry, these activities are super easy and you will enjoy doing these thing! After complete this course, you will be able to: 1. Apply some of the theories you learned in the lecture. 2. See firsthand the changes in food due to enzymes. 3. See how lipids are incorporated in our food. 4. Know how yeast works. LABORATORY GUIDE WEEK TOPICS EXPECTED OUTPUT Week 1 Introduction to Special paper Special paper Week 2 Exercise 1 Part 1: Summer Must-Do: Ice Cream in a Bag Part 2: Cartesian Diver Experiment Worksheet Worksheet Week 4 Exercise 2: Fruits Gone Bad? Discover Enzymatic Browning Worksheet Week 6 Exercise 3: Growing Yeast: Sugar Fermentation Worksheet Week 8 Exercise 4: Extraction of Fat Worksheet Week 10 Exercise 5: Denaturing Protein Worksheet GRADING SYSTEM: The student will be graded according to the following: CRITERIA LECTURE CLASS Class Standing (70%) Worksheets for Exercise1-5 Major Examination (30%) Concept paper 100% 126 SPECIAL PAPER “Biochemistry as part of my life” Question: How is biochemistry used in everyday life? (Short bond paper) (Can be printed or handwritten) Here are the guidelines for printed format: 1. Title Page On the title page include the title (Times New Roman (TNR), size 15, bold, centered, all capital letters), your name (TNR, size 12, centered, capital the first letter only) and the date (mm/dd/yyyy, TNR, size 12). **you can change the title if you want 2. Body – should cover all the main topics in the lecture (water; acid and bases; enzymes; macromolecules; cellular respirations and metabolism) a. Stay focused. Keep your discussion focused on your topic.. Don't let tangents or extraneous material get in the way of a concise, coherent discussion. A well-focused paper is crucial in getting your message across to your reader. b. Organize your points. Keeping your points organized makes it easier for the reader to follow along and make sense of your review. Start each paragraph with a topic sentence that relates back to your topic/experience. 3. Conclusions A good conclusion should illustrate the key connections between your major points and your topic/experience—what is the significance of your paper in a larger context? Make some conclusions—where have you arrived as a result of writing this paper? Be careful not to present any new information in the conclusion section. **If you opt to do this handwritten, you have the freedom to how you write this paper but please make it readable. You can include drawings to showcase your talent. 127 WATER AND LIFE EXERCISE 1 Summer Must-Do: Ice Cream in a Bag Cartesian Diver Experiment At the end of this exercise, you will witness some of the properties of water and know the respective use of those properties. Do you ever wonder why scientists spend time looking for water on other planets? It is because water is essential to life; even minute traces of it on another planet can indicate that life could or did exist on that planet. Water Is Polar The hydrogen and oxygen atoms within water molecules form polar covalent bonds. The shared electrons spend more time associated with the oxygen atom than they do with hydrogen atoms. There is no overall charge to a water molecule, but there is a slight positive charge on each hydrogen atom and a slight negative charge on the oxygen atom. Because of these charges, the slightly positive hydrogen atoms repel each other and form the unique shape seen in Figure 1. Each water molecule attracts other water molecules because of the positive and negative charges in the different parts of the molecule. Water also attracts other polar molecules (such as sugars), forming hydrogen bonds. When a substance readily forms hydrogen bonds with water, it can dissolve in water and is referred to as hydrophilic (“water-loving”). Hydrogen bonds are not readily formed with nonpolar substances like oils and fats. These nonpolar compounds are hydrophobic (“water-fearing”) and will not dissolve in water. Diagram showing hydrogen bonds formed between adjacent water molecules. In a single water molecule, two hydrogen atoms are bound to an oxygen atom with a covalent bond. Adjacent water molecules are bound together with weak hydrogen bonds, where hydrogen atoms of water molecules bind together with the oxygen atom of another water molecule. Figure 1. Hydrogen bonds form between slightly positive (δ+) and slightly negative (δ–) charges of polar covalent molecules, such as water. Water Stabilizes Temperature The hydrogen bonds in water allow it to absorb and release heat energy more slowly than many other substances. Temperature is a measure of the motion (kinetic energy) of molecules. As the motion increases, energy is higher and thus temperature is higher. Water absorbs a great deal of energy before its temperature rises. Increased energy disrupts the hydrogen bonds between water molecules. Because these bonds can be created and disrupted rapidly, water absorbs an increase in energy and temperature changes only minimally. This means that water moderates temperature changes within organisms and in their environments. As energy input continues, the balance between hydrogen-bond formation and destruction swings toward the destruction side. More bonds are broken than are formed. This process 128 results in the release of individual water molecules at the surface of the liquid (such as a body of water, the leaves of a plant, or the skin of an organism) in a process called evaporation. Evaporation of sweat, which is 90 percent water, allows for cooling of an organism, because breaking hydrogen bonds requires an input of energy and takes heat away from the body. Conversely, as molecular motion decreases and temperatures drop, less energy is present to break the hydrogen bonds between water molecules. These bonds remain intact and begin to form a rigid, lattice-like structure (e.g., ice) (Figure 2a). When frozen, ice is less dense than liquid water (the molecules are farther apart). This means that ice floats on the surface of a body of water (Figure 2b). In lakes, ponds, and oceans, ice will form on the surface of the water, creating an insulating barrier to protect the animal and plant life beneath from freezing in the water. If this did not happen, plants and animals living in water would freeze in a block of ice and could not move freely, making life in cold temperatures difficult or impossible. Figure 2. (a) The lattice structure of ice makes it less dense than the freely flowing molecules of liquid water. Ice’s lower density enables it to (b) float on water. (credit a: modification of work by Jane Whitney; credit b: modification of work by Carlos Ponte) Water Is an Excellent Solvent Because water is polar, with slight positive and negative charges, ionic compounds and polar molecules can readily dissolve in it. Water is, therefore, what is referred to as a solvent—a substance capable of dissolving another substance. The charged particles will form hydrogen bonds with a surrounding layer of water molecules. This is referred to as a sphere of hydration and serves to keep the particles separated or dispersed in the water. In the case of table salt (NaCl) mixed in water (Figure 3), the sodium and chloride ions separate, or dissociate, in the water, and spheres of hydration are formed around the ions. Figure 3. When table salt (NaCl) is mixed in water, spheres of hydration form around the ions. A positively charged sodium ion is surrounded by the partially negative charges of oxygen atoms in water molecules. A negatively charged chloride ion is surrounded by the partially positive charges of hydrogen atoms in water molecules. These spheres of hydration are 129 also referred to as hydration shells. The polarity of the water molecule makes it an effective solvent and is important in its many roles in living systems. Water Is Cohesive Have you ever filled up a glass of water to the very top and then slowly added a few more drops? Before it overflows, the water actually forms a dome-like shape above the rim of the glass. This water can stay above the glass because of the property of cohesion. In cohesion, water molecules are attracted to each other (because of hydrogen bonding), keeping the molecules together at the liquid-air (gas) interface, although there is no more room in the glass. Cohesion gives rise to surface tension, the capacity of a substance to withstand rupture when placed under tension or stress. When you drop a small scrap of paper onto a droplet of water, the paper floats on top of the water droplet, although the object is denser (heavier) than the water. This occurs because of the surface tension that is created by the water molecules. Cohesion and surface tension keep the water molecules intact and the item floating on the top. It is even possible to “float” a steel needle on top of a glass of water if you place it gently, without breaking the surface tension (Figure 4). These cohesive forces are also related to the water’s property of adhesion, or the attraction between water molecules and other molecules. This is observed when water “climbs” up a straw placed in a glass of water. You will notice that the water appears to be higher on the sides of the straw than in the middle. This is because the water molecules are attracted to the straw and therefore adhere to it. Cohesive and adhesive forces are important for sustaining life. For example, because of these forces, water can flow up from the roots to the tops of plants to feed the plant. Figure 4. The weight of a needle on top of water pulls the surface tension downward; at the same time, the surface tension of the water is pulling it up, suspending the needle on the surface of the water and keeping it from sinking. Notice the indentation in the water around the needle. (credit: Cory Zanker) 130 Part 1. Summer Must-Do: Ice Cream in a Bag Materials Needed: Big plastic bag (or ziplock) Small plastic bag (or ziplock 1 tetrabrick all-purpose cream, room temperature Ice tubes or ice cubes 1/2 cup rock salt 1/2 tsp vanilla extract (optional) 1 Tablespoon sugar 1. Fill the big plastic bag with ice (about half way). 2. Pour in 1/2 cup salt into the big plastic bag along with the ice. (Note: You will NOT be eating the salt.) 3. To the smaller bag, add cream, sugar, vanilla. Press out as much air as possible. Close the bag tightly. 4. Place smaller bag in larger bag. Adjust the ice so that it surrounds smaller bag. Press out air. Close the bag. 5. Toss, shake, jiggle, smush, and dance! 6. After about 15 minutes, check the ice cream. Shake more if needed until desired consistency is achieved. 7. Observe the consistency of the ice cream. Part 2. Cartesian Diver Experiment Materials needed: Water Packet of ketchup or soy sauce (sachet of condiments usually seen in fast food chains) Bottle with an airtight cap 1. Fill the bottle completely with water. 2. Put the packet of ketchup into the bottle. You may have to squeeze the packet a little bit to fit it into the bottle's opening. Be careful not to break open the packet. 3. Top the bottle with water again before closing the cap. 4. Now close the cap carefully. Try not to spill any water. 5. Here is the fun part. Squeeze the bottle and see the ketchup packet sink to the bottom. 131 RESULTS Name: _________________________________________________________ Section:_____________________________________ Please write your answers in the provided space. 1. How does salt affect ice? Explain the science behind it. ____________________________________________________________________________ ____________________________________________________________________________ ____________________________________________________________________________ ____________________________________________________________________________ ____________________________________________________ 2. Why does a ketchup packet float in water? Discuss the principles of buoyancy and density. ____________________________________________________________________________ ____________________________________________________________________________ ____________________________________________________________________________ ____________________________________________________________________________ ____________________________________________________ 132 ENZYMES EXERCISE 2 Fruits Gone Bad? Discover Enzymatic Browning At the end of this experiment, you will know a thing or two ways to prevent the browning of fruits. Browning of foods can be either non-enzymatic (caramelization or maillard reaction) or enzymatic. Non-Enzymatic Browning Caramelization is a non-enzymatic reaction that occurs when carbohydrates or sugars in food are heated. It is the process of removal of water from a sugar followed by isomerization and polymerization steps. The Maillard reaction is a chemical reaction between an amino acid and a reducing sugar, usually requiring the addition of heat. Like caramelization, it is a form of non-enzymatic browning. The reactive carbonyl group of the sugar interacts with the nucleophilic amino group of the amino acid, and interesting but poorly characterized odor and flavor molecules result. This process accelerates in an alkaline environment because the amino groups do not neutralize. This reaction is the basis of the flavoring industry, since the type of amino acid determines the resulting flavor. In the process, hundreds of different flavor compounds are created. These compounds in turn break down to form yet more new flavour compounds, and so on. Each type of food has a very distinctive set of flavor compounds that are formed during the Maillard reaction. It is these same compounds that flavor scientists have used over the years to create artificial flavors. Figure 1. Non-enzymatic browning -- the result of the Maillard reaction and Caramelization of carrots. Enzymatic Browing Enzymatic browning takes place when the enzyme polyphenol oxidase or other enzymes catalyze the oxidation of phenols in the fruit to form compounds called quinones. The quinones can then polymerize to form melanins, which cause the brown pigments. Enzymatic browning of fruits and vegetables is usually not desirable and creates heavy economic losses for growers. Enzymatic browning is beneficial for: 1. Developing flavor in tea (here the reaction is incorrectly called fermentation) 2. Developing color and flavor in dried fruit such as figs and raisins. Enzymatic browning is detrimental to: 1. Fresh fruit and vegetables, in particular apples and potatoes Seafood such as shrimp 133 Control of browning Enzymatic browning is usually controlled with chemicals, or by destroying the responsible chemicals with heat. Blanching to destroy the enzymes is commonly used to preserve color in vegetables. Lemon juice and other acids are used to preserve color in fruit, particularly apples, by lowering the pH and removing the copper site necessary for the enzyme to function. Have you ever wondered why apple slices turn brown once you cut them, or why a yellow banana gets dark spots over time? Both of these phenomena have the same cause: enzymatic browning triggered by an enzyme called polyphenol oxidase (PPO). In this activity you will find out how this enzyme works by turning a banana from yellow to brown in just a matter of seconds. Then you will explore how you can keep your apple slices looking fresh! Tons of fruits and vegetables are produced, processed and shipped on a daily basis so that we can buy them fresh. Many of these pieces of produce, however, never make it into stores. This is because some fruits and vegetables such as apricots, mushrooms, lettuce and pears degrade over time because of enzymatic browning. You can see this for yourself if you observe a banana for several days or weeks. Would you buy a brown banana? Enzymatic browning is one of the largest causes of quality loss in fruits and vegetables—even though it does not make the food harmful to eat. So what exactly happens during enzymatic browning? The process occurs over several steps. The enzyme responsible for the browning is called polyphenol oxidase (or PPO). In the presence of oxygen the PPO enzyme changes substances known as phenolic compounds (through a process of oxidation) into different compounds called quinones. The quinones then react with other compounds to form melanin. Melanin is the same dark brown pigment that colors hair, skin and the irises of our eyes. It also turns fruit and vegetables brown. This reaction, however, usually does not happen within fresh fruits and vegetables because the PPO and the phenolic compounds are separated in produce plant cells. The enzymatic browning process is only triggered when PPO, phenolic compounds and oxygen come in contact with one another. This is exactly what happens when a fruit is cut, falls or is knocked around too much. When fruit tissue is damaged because of heat, cold, age or mechanical stress, its cells break open and the phenolic compounds and the enzyme are released and mix with oxygen in the air. As a result the damaged tissue turns brown almost immediately. In this activity you will see this browning process for yourself and investigate how this reaction can be avoided. So grab some fruit and get started! Materials Needed: Part 1. Banana 1 piece Banana (Yellow with no brown spots) Pot Water 1. Fill a pot with water. 2. Place the pot on the stove and heat the water until boiling. 3. Take the banana and look closely at its peel. What color is it? Do you see any brown spots? Record on the answer sheet on page 11. 4. Carefully dip the bottom third of the banana into the boiling water for 30 seconds. What happens to the banana when you submerge it in hot water?. Record on the answer sheet on page 10. 5. After the 30 seconds remove the banana from the boiling water and observe it for another three minutes. What do you notice? Does the banana look different after a while? How?. Record on the answer sheet on page 10 6. When the banana has cooled down, peel the banana. Look at the fruit that was inside the peel. Did you expect the banana to look like that? Record on the answer sheet on page 11. 134 Part 2. Apple 1 piece Apple Milk powder Vinegar Calamansi juice or Lime juice or Lemon juice Cutting board Knife 1. Cut two slices from the apple on a cutting board. Place each slice onto its side. How do they look? Record on the answer sheet on page 10. 2. Poke one of the apple slices with a fork several times. Then observe both slices for 15 to 20 minutes. How do the apple slices change over time? Do you notice a difference between the two slices? If yes, can you explain why? Record on the answer sheet on page 12. 3. Cut five more slices from the apple and place each slice on its side. Immediately after cutting, sprinkle milk on top of the first slice, vinegar on the second slice, lemon juice on the third slice and water on the fourth slice. Keep the last slice as is. Then poke each apple slice several times with a fork. What do you think these liquids will do to the apple? Record on the answer sheet on page 11. 4. Observe all five apple slices for another 15 to 20 minutes. How are the apple slices different after 15 to 20 minutes? What did each liquid do to the apple slice? Can you explain your results? Record on the answer sheet on page 12. 135 RESULTS Name: _________________________________________________________ Section:_____________________________________ Please write your answers in the provided space. BANANA: 1. What color is it? Do you see any brown spots? ____________________________________________________________________________ ____________________________________________________________________________ 2. What happens to the banana when you submerge it in hot water? 3. What do you notice? Does the banana look different after a while? How? 4. Did you expect the banana to look like that? ____________________________________________________________________________ ____________________________________________________________________________ 136 APPLE: 1. How do they look? ____________________________________________________________________________ ____________________________________________________________________________ 2. How do the apple slices change over time? Do you notice a difference between the two slices? If yes, can you explain why? 3. What do you think these liquids will do to the apple? 4. How are the apple slices different after 15 to 20 minutes? What did each liquid do to the apple slice? Can you explain your results? 137 CARBOHYDRATES EXERCISE 3 Growing Yeast: Sugar Fermentation At the end of this experiment, you will be able to explain how yeast works. Carbohydrates make up a group of chemical compounds found in plant and animal cells. They have the empirical formula CnH 2nOn, or (CH2O)n. An empirical formula tells the atomic composition of the compound, but nothing about structure, size, or what chemical bonds are present. Since this formula is essentially a combination of carbon and water, these materials are called “hydrates of carbon”, or carbohydrates for short. Carbohydrates are the primary products of plant photosynthesis. The simplified lightdriven reaction of photosynthesis results in the formation of a carbohydrate: nH 2O + nCO2 → (CH2O)n- + nO2. This type of carbohydrate is found in the structures of plants and is used in the reverse reaction of photosynthesis (respiration) or is consumed as fuel by plants and animals. Carbohydrates are found in a wide array of both healthy and unhealthy foods—bread, beans, milk, popcorn, potatoes, cookies, spaghetti, soft drinks, corn, and cherry pie. They also come in a variety of forms. The most common and abundant forms are sugars, fibers, and starches. Foods high in carbohydrates are an important part of a healthy diet. Carbohydrates provide the body with glucose, which is converted to energy used to support bodily functions and physical activity. But carbohydrate quality is important; some types of carbohydrate-rich foods are better than others: • The healthiest sources of carbohydrates—unprocessed or minimally processed whole grains, vegetables, fruits and beans—promote good health by delivering vitamins, minerals, fiber, and a host of important phytonutrients. • Unhealthier sources of carbohydrates include white bread, pastries, sodas, and other highly processed or refined foods. These items contain easily digested carbohydrates that may contribute to weight gain, interfere with weight loss, and promote diabetes and heart disease. Carbohydrates are divided into monosaccharides, disaccharides, and polysaccharides. As shown in the following molecular model structures, carbohydrates may be found as hexagon (6-sided, see Figure 1A) and pentagon (5-sided, see Figure 1B) shaped rings. Monosaccharides Monosaccharides are single-molecule sugars (the prefix “mono” means one) that form the basic units of carbohydrates. They usually consist of three to seven carbon atoms with attached hydroxyl (OH) groups in specific stereochemical configurations. The carbons of carbohydrates are traditionally numbered starting with the carbon of the carbonyl end of the chain (the carbonyl group is the carbon double-bonded to oxygen). The number of carbons in the molecule generally categorizes monosaccharides. For example, three-carbon carbohydrate molecules are called trioses, five-carbon molecules are called pentoses, and six-carbon molecules are called hexoses. Ribose and 2-deoxyribose are pentoses, and both have a crucial role in reproduction as polymers known as ribonucleic acid (RNA) and deoxyribonucleic acid (DNA). One of the most important monosaccharides is glucose (dextrose). This molecule is the primary source of chemical energy for living systems. Plants and animals alike use this molecule for energy to carry out cellular processes. Mammals produce peptide hormones (insulin and glucagon) that regulate blood glucose levels, and a disease of high blood glucose is called diabetes. Other hexoses include fructose (found in fruit juices) and galactose. 138 Different structures are possible for the same monosaccharide. Although glucose and fructose are identical in chemical composition (C 6H12O6), they are very different in structure (see molecular models). Such materials are called isomers. Isomers in general have very different physical properties based on their structure. Figure 1. Examples of carbohydrates. (A). Glucose, a six-membered ring monosaccharide. (B). Fructose, a five-membered ring monosaccharide. (C). Sucrose, a disaccharide containing glucose and fructose. (D). Molecular representation of starch illustrating the alpha-glycosidic linkages joining monosaccharides to form the polysaccharide structure. Disaccharides Disaccharides are two monosaccharide sugar molecules that are chemically joined by a glycosidic linkage (- O -) to form a “double sugar” (the prefix “di” means two). When two monosaccharide molecules react to form a glycosidic bond (linkage), a water molecule is generated in the process through a chemical reaction known as condensation. Therefore, condensation is a reaction where water is removed and a polymer is formed. The most wellknown disaccharide found in nature is sucrose, which is also called cane sugar, beet sugar, or table sugar (see Figure 1(C)). Sucrose is a disaccharide of glucose and fructose. Lactose or milk sugar is a disaccharide of glucose and galactose and is found in milk. Maltose is a disaccharide composed of two glucose units. Disaccharides can easily be hydrolyzed (the reverse of condensation) to become monosaccharides, especially in the presence of enzymes (such as the digestive enzymes in our intestines) or alkaline catalysts. Invert sugar is created from the hydrolysis of sucrose into glucose and fructose. Bees use enzymes to create invert sugar to make honey. Taffy and other invert sugar type candies are made from sucrose using heat and alkaline baking soda. Disaccharides are classified as oligosaccharides (the prefix “oligo” means few or little). This group includes carbohydrates with 2 to 20 saccharide units joined together. Carbohydrates containing more than 20 units are classified as polysaccharides. Polysaccharides Polysaccharides (the prefix “poly” means many) are formed when many single sugars are joined together chemically. Carbohydrates were one of the original molecules that led to the discovery of what we call polymers. Polysaccharides include starch, glycogen (storage starch in animals), cellulose (found in the cell walls of plants), and DNA. Starch is the predominant storage molecule in plants and provides the majority of the food calories consumed by people worldwide. Most starch granules are composed of a mixture of two polymers: a linear polysaccharide called amylose and a branched-chain polysaccharide called amylopectin. Amylopectin chains branch approximately every 20- 25 saccharide units. Amylopectin is the more common form of starch found in plants. Animals store energy in the muscles and liver as glycogen. This molecule is more highly branched than amylopectin. For 139 longer-term storage, animals convert the food calories from carbohydrates to fat. In the human and animals, fats are stored in specific parts of the body called adipose tissue. Cellulose is the main structural component of plant cell walls and is the most abundant carbohydrate on earth. Cellulose serves as a source of dietary fiber since, as explained below; humans do not have the intestinal enzymes necessary to digest it. Starch and cellulose are both homopolymers (“homo” means same) of glucose. The glucose molecules in the polymer are linked through glycosidic covalent bonds. There are two different stereochemical configurations of glycosidic bonds—an alpha linkage and a beta linkage. The only difference between the alpha and beta linkages is the orientation of the linked carbon atoms. Therefore, glucose polymers can exist in two different structures, with either alpha or beta linkages between the glucose residues. Starch contains alpha linkages (see Figure 1(D)) and cellulose contains beta linkages. Because of this difference, cornstarch has very different physical properties compared to those for cotton and wood. Salivary amylase only recognizes and catalyzes the breakdown of alpha glycosidic bonds and not beta bonds. This is why most mammals can digest starch but not cellulose (grasses, plant stems, and leaves). Yeast is most commonly used in the kitchen to make dough rise. Have you ever watched pizza crust or a loaf of bread swell in the oven? Yeast makes the dough expand. But what is yeast exactly and how does it work? Yeast strains are actually made up of living eukaryotic microbes, meaning that they contain cells with nuclei. Being classified as fungi (the same kingdom as mushrooms), yeast is more closely related to you than plants! In this experiment we will be watching yeast come to life as it breaks down sugar, also known as sucrose, through a process called fermentation. Let’s explore how this happens and why! Materials: 4 350mLplastic bottles (used mineral water bottle 350mL) 4 small balloons Permanent marker 2 cups warm water (not boiling, not room temperature) 12 teaspoon white sugar 4 teaspoon yeast 1. Label your plastic bottles 1, 2, 3 and 4. 2. Fill each bottle with ½ cup of warm water. (Note: Do not put boiled water; the plastic bottle will be deformed.) 3. In bottle 1, mix 1 teaspoon yeast only. 4. In bottle 2, mix 1 teaspoon yeast and 2 teaspoon white sugar. 5. In bottle 3, mix 1 teaspoon yeast and 4 teaspoon white sugar. 6. In bottle 4, mix 1 teaspoon yeast and 6 teaspoon white sugar. 7. Give a little bit of shake. Make sure that you shake all the bottles in the same way. 8. Place the balloons on the mouth of the bottles. 9. Observe what will happen on the balloon after 15 minutes. 140 RESULTS Name: _________________________________________________________ Section:_____________________________________ Please write your answers in the provided space. 1. Why do you think the reactions in each bottle differed from one another? What is the science behind it. ____________________________________________________________________________ ____________________________________________________________________________ ____________________________________________________________________________ ____________________________________________________________________________ ____________________________________________________ 2. Kindly draw what you have observed. 141 Cryptic Carbohydrates Name: _________________________________________________________ Section:_____________________________________ Fill in the blank spaces with the appropriate terms to complete the sentences. Solve the hidden message by entering the boxed letters in the spaces at the bottom of the page. Please write your answers in the provided space. 1. are identical in chemical composition but differ structurally. 2. is a polymer of glucose and serves as a source of dietary fiber for humans. 3. are an inexpensive and widely available source of energy for our bodies. 4. is a disaccharide found in cow’s milk 5. is a disaccharide composed of glucose and fructose. 6. is a starch that has gelling properties and is used in making jams and preserves. 7. Glucose is a . 8. The reaction is a nonenzymatic browning reaction that occurs when foods are roasted or baked. 9. bonds chemically join two or more monosaccharide molecules. 10. Carbohydrates are the primary products of plant . HIDDEN MESSAGE: A polysaccharide called carrageenan is a seaweed extract. Carrageenan is used as a stabilizer in what popular frozen dessert product? 142 Cool Carbs Name: _________________________________________________________ Section:_____________________________________ Find and encircle the words listed below in the word search. After all the words are found, the letters that are not used reveal a hidden message at the bottom of this sheet. Please write your answers in the provided space. HIDDEN MESSAGE: We should __ __ __ __ __ __ __ __ __ __ __ __ __ __ __ __ __ __ __ __ __ __ __ __ __ __ __ __ __ __ __ __ __ __ __ __ __ __ __ __. 143 LIPIDS EXERCISE 4 Extraction of Fat In this experiment, you will be extracting and examining the fat in selected food items. Lipids include fats, oils, waxes, cholesterol, other sterols, and most steroids. In the body, fat serves as a source of energy, a thermal insulator and cushion around organs, and an important cellular component. The fat-soluble vitamins are A, D, E, and K. You are probably most familiar with the nutritional aspects of dietary fats and oils. Since fats have 2.25 times the energy content of carbohydrates and proteins, most people try to limit their intake of dietary fat to avoid becoming overweight. The food industry has a big market for low-fat and non-fat foods. Just take a look around your local grocery stores! Lipids are classified as organic compounds that are soluble (dissolvable) in organic solvents, but only sparingly soluble in water. Lipids are biologically important for making barriers (membranes of animal cells), which control the flow of water and other materials into a cell. Fats and oils make up 95% of food lipids and phospholipids, and sterols make up the other 5%. Traditionally, fats were considered to be solid at room temperature, and oils were considered to be liquid. However, this designation is often used to distinguish between fats and oils from animals and plants, respectively. Animal fats are found in meats (beef, chicken, lamb, pork, and veal), milk products, eggs, and seafood (fish oil). Plant (vegetable) oils come from nuts (peanuts), olives, and seeds (soybean, canola, safflower, and corn). We use lipids for flavor (butter and olive oil), to cook foods (oils and shortening), to increase the palatability of foods by improving the texture or “mouthfeel” (cakes, creamy ice cream), and in food processing (emulsifiers). Fatty acids are generally long, straight chains of carbon atoms with hydrogen atoms attached (hydrocarbons) with a carboxylic acid group (COOH) at one end and a methyl group (CH3) at the other end. These long, straight chains combine with the glycerol molecule (see Figure 1(A)) to form lipids (glycerol lipids). Figure 1. Examples of lipids. (A) Glycerol molecule is the backbone of a glycerol lipid. The triacylglycerol contains three fatty acids attached at the oxygen atoms of glycerol. (B) Configuration of a cis double bond. (C) Configuration of a trans double bond. (D) Linoleic acid is an essential fatty acid containing two double bonds. It is needed for growth and health. (E) Stearic acid is a saturated fatty acid found in foods from animal and plant sources. (F) Milk fat triacylglycerol molecule illustrating the ester bonds between fatty acids and glycerol. Most naturally occurring fatty acids contain an even number of carbon atoms. The 18carbon fatty acids are the most abundant in our food supply; examples are linoleic acid (an omega-6 fatty acid) found in corn oil and linolenic acid (an omega-3 fatty acid) found in flaxseed oil. Linoleic and linolenic acids are considered essential fatty acids because they are needed for normal physiological functions and our body cannot make them. We need to get these fatty 144 acids from food sources. These fatty acids are found in the vegetable oils used in several different food products. Structure of Lipids Most of the carbon−carbon bonds in fats are single bonds, which allow the carbons to freely rotate, making the attached groups chemically identical. However, the number of unsaturated bonds (double bonds) may vary from one to many in the hydrocarbon part of the fatty acid. Since double bonds do not allow free rotation between the attached carbons, any attached chemical groups are fixed in their respective positions. There are two possible orientations for groups attached to the carbons in a double bond. If they are on the same side of the double bond (close together), they are in the cis conformation. The opposite of the cis conformation is the trans conformation, where the residues at ends of the double bond are farther apart. Double bonds in natural vegetable oils and in animal fats are mostly in the cis conformation (see Figures 1B and 1C); however, a few exceptions are known where the trans conformation is present. The presence of the double bonds is responsible for the liquid properties of native vegetable oil. Because the cis double bonds are “kinked”, they disrupt the physical interactions between fatty acid molecules, preventing them from packing together tightly to form crystals (see Figure 1D, structure of linoleic acid). This disruption keeps the fatty acid molecules from associating with each other, resulting in a liquid structure. If the double bonds are removed by adding hydrogen (hydrogenation), the kinks are removed, allowing the fatty acid molecules to more easily associate with each other (see Figure 1E, structure of stearic acid). The result is crystallization (solid fat) at room temperature. Depending on how the various fatty acid chains associate, the crystalline structure of the solid fat can have different appearances, such as a smooth, shiny solid or a rough, puffy solid. These crystalline forms also have different lightreflection characteristics and physical hardness. This difference in physical properties is used when making shortening, which is crystallized into a very white, soft crystalline form at the factory. However, upon melting and re-solidification, it becomes more translucent and grayer, due to the formation of a different crystal structure. Nomenclature for Fats If all the bonds are single, the fatty acid molecule is saturated, because the maximum number of hydrogen atoms is associated with the carbon atoms. Some examples are tallow (beef fat), lard (pork fat), and butter (milk fat). If there is a double bond among the carbon atoms, the fatty acid molecule is unsaturated. Examples of unsaturated fats are canola oil, corn oil, cottonseed oil, and soybean oil. If there are multiple double bonds (two or more), it is called polyunsaturated. You may recall seeing the saturated, unsaturated, and polyunsaturated terms with respect to nutritional aspects of oils. Corn and soybean oils are some of the most important food sources of polyunsaturated fatty acids in our food supply. Shown below are the shorthand notation used to describe some important food sources of 18-carbon (C18) fatty acids. • C18:0 is a fully saturated 18-carbon fatty acid called stearic acid. • C18:1 has one double bond, between carbons 9-10, (18:1n9) counting from the COOH end, and is called oleic acid. • C18:2 has two double bonds, between carbons 9-10 and 12-13, counting from the COOH end, and is called linoleic acid (9, 12-octadecadienoic acid or 18:2n6). • C18:3 has three double bonds at carbons 9-10, 12-13, and 15-16, counting from the COOH end, and is called linolenic acid (9,12,15-octadecatrienoic acid or 18:3n3). The number of fatty acids joined to the glycerol molecule also plays a part in how the molecule is named. If only one fatty acid is connected, the general name for the molecule is a monoacylglycerol. If two are joined, the molecule is called a diacylglycerol, and if three are joined, a triacylglycerol. The bond between the fatty acid and the glycerol also has a special name. It is called an ester bond (see Figure 1F). The carboxyl end (COOH) of the fatty acid molecule attaches to one of the -OH groups of the glycerol molecule. Because of this combination, an -OH group and -H are left, which combine to form a water molecule. 145 Since triacylglycerols have three fatty acids, you can get mixed-fatty-acid triacylglycerols, in which there are different fatty acids on each of the glycerol bonds. Naturally occurring soybean oil is a mixed triacylglycerol, containing saturated, monounsaturated, and polyunsaturated fatty acids. Soybean oil contains more monounsaturated and polyunsaturated fatty acids than saturated fatty acids. Surfactant is a short term for surface-active agent. Polar lipids, like lecithin in soybean oil, serve as specialized surfactants known as emulsifiers. By interacting with water on one end of the molecule and repelling water on the other end, emulsifiers keep fat globules dispersed in water or water droplets dispersed in fat. Lipid surfactants are important to our own cellular functions, as well as useful in stabilizing specific food products. Lecithin is a phospholipid, which functions as a surfactant. Lecithin and other phospholipid emulsifiers are found in food from animal and plant sources. The food sources of lecithin are eggs, milk, cheese, and soybean oil. These chemical properties of lecithin are used in the food industry to prevent fats from separating out of chocolate, mayonnaise, peanut butter, and salad dressings. The fats that you see in raw beef, chicken, and pork are known as visible fats. These fats are in plain view and are solid at room temperature. Vegetable oils are also visible fats. The fats that are in snack foods, cookies, desserts, and candy are known as invisible fats. Although you cannot see them, they can add extra calories to your diet. 146 Objectives: In this experiment, we will be extracting and examining the fat in chocolate, potato chips, and sunflower seeds. In chocolate, sugar and cocoa are dispersed in a crystallized fat matrix. To keep the fat from separating out of the chocolate, an emulsifier called lecithin is used. The fat in the potato chip is mostly on the surface of the chip from the frying process. The fat in the sunflower seed is in the seed itself. The cooking oils that we use come primarily from nuts and seeds. Examples of these fat sources are corn, soybean, and peanut oils. Materials Needed: Chocolate (example flat tops, curly tops, nips or any chocolate) Sunflower seeds or sesame seeds or raw peanut (without skin) (whichever is available or accessible) Potato chips (any brand) Acetone (use for removing nail polish) Foil Bowls Small plastic container Part 1. Extraction of Fat from Chocolate Chips 1. Weigh out 5 grams of chocolate chips. Crush the chocolate between two sheets of foil with a hammer (or any hard material in your kitchen) 2. Label the bowls that you are using to put the food in, one each for chocolate chips, potato chips, and sunflower seeds. 3. Place the crushed chocolate in the designated bowl (bowl labeled with “chocolate”). 4. Add 10 milliliters (2 teaspoons) of acetone to the crushed chocolate chips in the beaker. 5. Swirl for 1 minute in a well-ventilated area. 6. Transfer the acetone into a plastic container, making sure the chocolate remains in the bowl. 7. Add 10 milliliters (2 teaspoon) of acetone to the chocolate again and repeat steps 5 and 6. 8. Allow the acetone in the saucer to dry overnight in a well-ventilated area to visualize the lipid that was extracted. 9. Allow the bowl with the chocolate to dry overnight. Part 2. Extraction of Fat from Potato Chips 1. Weigh out 5 grams of potato chips. Break into small pieces (size of a 5 cent) with your fingers. 2. Repeat steps 2−9 in Part 1. Part 3. Extraction of Fat from Sunflower Seeds 1. Weigh out 5 grams of sunflower seeds (or sesame seeds or peanut). Crush (almost ground texture) the seeds between two pieces of foil with a hammer. 2. Repeat steps 2−9 in Part 1. 147 RESULTS Name: _________________________________________________________ Section:_____________________________________ Please write your answers in the provided space. DESCRIPTION OF FATS FOOD SAMPLE COLOR TEXTURE ODOR VISCOCITY Chocolate Potato Chips Sunflower seeds Questions: 1. Rank from most to least the percentage of lipid extracted from all three foods. Look at the Nutrition Facts label on the packages of all three foods and rank them. Did your ranking agree with the ranking of the product labels? ____________________________________________________________________________ ____________________________________________________________________________ ____________________________________________________________________________ ____________________________________________________________________________ _________________________________________________ 2. Determine which lipids contained saturated and unsaturated fatty acids in this experiment, based on your descriptions of the fats in the plastic container. ____________________________________________________________________________ ____________________________________________________________________________ ____________________________________________________________________________ ____________________________________________________________________________ ___________________________________________________ 3. Why are fats needed in the body? ____________________________________________________________________________ ____________________________________________________________________________ ____________________________________________________________________________ 148 ____________________________________________________________________________ ___________________________________________________ 4. Why are fats a concern when eaten in excess amounts? ____________________________________________________________________________ ____________________________________________________________________________ ____________________________________________________________________________ ____________________________________________________________________________ ___________________________________________________ 149 Freaky Fats Name: _________________________________________________________ Section:_____________________________________ Fill in the blank spaces with the appropriate terms to complete the sentences. Solve the hidden message by entering the boxed letters in the spaces at the bottom of the page. 1. are long chains of carbon and hydrogen atoms that combine with glycerol molecules to form a lipid. 2. extracted from olives, canola, and corn are plant lipids and are liquid at room temperature. 3. is a phospholipid emulsifier found in eggs and soybean oil. 4. are found between the fatty acids and the glycerol molecules in triacylglycerols. 5. is a crystalline form of a solid fat. 6. are used to prevent the cocoa butter from separating out of chocolate bars. 7. fatty acids have multiple double bonds. 8. molecules form the backbone of attachment for fatty acid molecules. 9. fat is saturated because it contains all single bonds and is solid at room temperature. 10. oils like corn and soybean are important food sources of polyunsaturated fatty acids in our food supply. HIDDEN MESSAGE: In the United State, this once popular oil is no longer used by the food industry as an ingredient because it was reported to increase blood cholesterol level. It was used primarily in cookies, cakes, and snack foods. 150 Fats Name: _________________________________________________________ Section:_____________________________________ Find the words listed below in the word search. After all the words are found, the letters that are not used reveal a hidden message at the bottom of this sheet. HIDDEN MESSAGE: We should __ __ __ __ __ __ __ __ __ __, __ __ __ __ __ __ __ __ __ __ __ __ __ __ __ __ __ __ __ __ __ __ __ __ __ __ __ __ __ __ __ __ __. 151 PROTEIN EXERCISE 5 Denaturing Proteins At the end of the experiment, you will be able to identify what chemical has the ability to denature proteins in food. Body builders and football players eat a lot of protein (eggs, cheese, and meat) to build muscle mass. You have probably seen protein-enriched drinks and protein-enriched foods (power bars) at the supermarket. Proteins are the most complex and important group of molecules because they possess diverse functionality to support life. Every cell that makes up plants and animals requires proteins for structure and function. Your body and plants also have enzymes. These specialized proteins catalyze chemical reactions that are necessary for metabolism and cell reproduction. Your muscles are made from a variety of proteins, and these proteins allow your muscles to contract, facilitating movement. Other types of proteins in your body are the peptide hormones; insulin and glucagon are two common examples. Proteins are complex polymers composed of amino acids. Amino acids contain carbon, hydrogen, nitrogen, and sometimes sulfur and serve as the monomers for making peptides and proteins. Amino acids have a basic structure that includes an amino group (NH 2) and a carboxyl group (COOH) attached to a carbon atom (see Figure 1(A)). This carbon atom also has a side chain (an “R” group). This side chain can be as simple as an -H or a -CH3, or even a benzene group. The R groups on an amino acid are analogous to an athlete's clothing and sports equipment. By changing clothing or equipment, an athlete can become more effective as a soccer, football, or baseball player. Although this person is still an athlete, the change can make the athlete more effective in a particular activity or function. The same is true with amino acids. They are still amino acids regardless of the attached R group, but different R groups produce different functions and different properties. Figure 1. Proteins. (A) Lysine is an essential amino acid (notice the blue nitrogen atom and the carboxyl group). (B) Glycylglycine is a dipeptide containing two glycine molecules connected by a peptide bond. (C) Aspartame is a dipeptide containing aspartic acid and phenylalanine. There are twenty amino acids found in the body. Eight of these amino acids are essential for adults and children, and nine are essential for infants. Essential means that we cannot synthesize them in adequate quantities for growth and repair of our bodies, and therefore, must be included in the diet. Amino acids are linked together by a peptide bond in which the carboxyl carbon of one amino acid forms a covalent bond with the amino nitrogen of the other amino acid (see Figure 1(B)). Short chains of amino acids are called peptides. Longer chains of amino acids are called polypeptides. Although the term polypeptides should include proteins, chains with less than 152 100 amino acid residues are considered to be polypeptides, while those with 100 or more amino acid residues are considered to be proteins. Many of the major hormones in the body are peptides. These hormones can influence enzyme action, metabolism, and physiology. Insulin, which is given to a person with a specific type of diabetes, is an example of a peptide hormone. Certain antibiotics and a few anti-tumor agents are also peptides. The artificial sweetener aspartame (Equal®) is a dipeptide composed of aspartic acid and phenylalanine with a methyl group attached at the carboxyl terminal group (L-aspartyl-L-phenylalanine methyl ester) (see Figure 1(C)). The sequence of amino acid residues in a polypeptide chain is critical for biological function. For example, a genetic disease (mutation of a single base pair in DNA) called sickle cell anemia is caused by the substitution of one amino acid (glutamate) with another (valine) in a structural protein called beta-globulin, which is a part of hemoglobin in red blood cells. Hence, a single structural change resulted in a dramatic alteration in physiological function. The ability of an enzyme to catalyze a particular reaction depends on its specific shape. It’s a lot like a key and lock if the key is broken or in a different shape, it won’t open the lock. The receptor sites on cell surfaces must be in a specific shape for polypeptide hormones to interact with the cell. With twenty different amino acids and each polypeptide consisting of hundreds of amino acids, it is no wonder that proteins play such a variety of roles in the human body. Chemistry of Proteins The protein backbone is formed from the peptide bonds created from the amino and carboxyl groups of each monomer that repeat the pattern -N-C-C- or C-C-N-. The number and sequence of amino acids in a polypeptide chain is referred to as the primary structure of a protein. The free amino group and carboxyl group on opposite ends of a polypeptide chain allow proteins to act as pH buffers (resist changes in pH) inside the cell. The amino group (NH 2) accepts a proton and becomes (NH 3+), and the carboxyl group (COOH) donates a proton and becomes dissociated (COO-). As noted previously, each amino acid residue in the polymer may have a different side chain or chemical group attached to it, such as hydroxyl (OH), amino (NH 2), aromatic ring (conjugate rings such as the phenol ring in phenylalanine), sulfhydryl (SH), carboxyl (COOH), or various alkyl (CHn). This variety of side chain groups on the polymer backbone gives proteins remarkable chemical and physical properties. For example, carboxylate groups can function as carboxylic acids (COO-), or amino groups can behave as bases (NH 3+). This allows protein polymers to be multifunctional molecules, with both acidic and basic behavior at the same time! Additionally, the presence of hydroxyls, carboxylates, sulfhydryls, and amino groups allows hydrogen bonding, and the alkyl groups provide hydrophobic interactions, both within the protein polymer itself and between separate protein molecules. In the case of macromolecules, such as proteins, the polymeric structure of the macromolecule allows it to simultaneously carry many different charges (on different amino acid residues). However, unlike the small single molecules, the amino acid residues are constrained by linear peptide linkages and thus cannot move freely to randomly associate with other charged molecules. Assuming that charged residues will seek to bond with the nearest convenient counter ion, it is most likely that oppositely charged amino acid residues located at different points within a single protein chain will bond. These structural differences result in the folding of proteins into a three-dimensional structure, which is, in part, responsible for their functional properties as biocatalysts, structural materials, muscles, and chemical receptors. Proteins can be shaped as long flat sheets or in globular spheres. This leads to the names fibrous or globular for protein shapes. Most enzymes are globular proteins. In standard acid−base chemistry, students learn that molecules carry electrostatic charges based on the type of atoms that make up a molecule and the environment of the molecule. Given that opposite charges attract, cationic and anionic atoms can combine to form covalent bonds, in which electrons are shared between atomic orbitals, or form ionic bonds, in which only electrostatic attractions exist. In solution with smaller molecules, such as HCl (an acid) or NaOH (a base), protein molecules can freely move around and associate with each other on a more-or-less random basis. 153 Protein polymers extend the simple acid−base charged chemical species concepts to explain how biological systems have greater levels of complexity and can utilize simple, monomeric chemical structures (like amino acids) to create exquisitely complex biological structures like antibodies, muscle, and skin. Protein polymers have physical structure, even when dissolved in liquids. The charged and hydrophobic residues within a protein tend to associate, causing the protein to fold up. When you unfold the protein molecule (called denaturation), its charged residues can reassociate with other charged molecules (precipitation or coagulation). Protein precipitation is widely used to recover recombinant protein products, enzymes, or in the production of many common foods. Cheeses and soybean tofu are examples of coagulated protein food products. Food Uses of Proteins Proteins also serve important roles in the processing of food products. They are used for their thickening, gelling, emulsifying, and water-binding properties in meats (sausages), bakery products, cheese, desserts, and salad dressings. Proteins are used for their cohesive and adhesive properties in sausage making, pasta, and baked goods. Egg proteins are used for their foaming properties in desserts, cakes, and whipped toppings. Milk, egg, and cereal proteins are used as fat and flavor binders in low-fat bakery products. Proteins are used for texture and palatability in bakery products (breads, cakes, crackers, and pizza crust) and sausages. Materials needed: 5 pieces eggs 1 Big pot 5 pieces small glass bottles Salt Baking soda Vinegar Calamansi Juice (lemon or lime) Spoon 1. Fill half of the big pot with water . Heat to boil. 2. Label the glass bottles #1-5. 3. Separate 5 eggs, placing the egg white into the glass bottles. Discard the egg yolk. 4. Place glass bottle #1 in the boiling water and allow to “cook” till egg turns white. 5. Add 2 teaspoon salt to glass bottle #2 and stir. (Do not put in the boiling water) 6. Add 2 teaspoon baking soda to glass bottle #3 and stir. (Do not put in the boiling water) 7. Add 2 teaspoon vinegar to glass bottle #4 and stir. (Do not put in the boiling water) 8. Add 2 teaspoon calamansi juice to glass bottle #5 and stir. (Do not put in the boiling water) 9. Record observations. 154 RESULTS Name: _________________________________________________________ Section:_____________________________________ Please write your answers in the provided space. GLASS BOTTLES TREATMENT 1 Heat 2 Salt 3 Baking Soda 4 Vinegar 5 Calamansi Juice (lime or lemon) OBSERVATIONS Questions: 1. Which method appeared to have the most dramatic denaturing affect on egg albumin? Why do you think this method had a greater affect? ____________________________________________________________________________ ____________________________________________________________________________ ____________________________________________________________________________ ____________________________________________________________________________ 2. Of the methods you tested, which would be more likely to be used in the food industry? ____________________________________________________________________________ ____________________________________________________________________________ ____________________________________________________________________________ ____________________________________________________________________________ 155 Powerful Proteins Name: _________________________________________________________ Section:_____________________________________ Fill in the blank spaces with the appropriate terms to complete the sentences. Solve the hidden message by entering the boxed letters in the spaces at the bottom of the page. Please write your answers in the provided space. 1. 2. are short chains of amino acids. of amino acids make up protein molecules. 3. are the building blocks of proteins. 4. can influence enzyme action, metabolism, and physiology. 5. is a proteolytic enzyme that is used to make cheese. 6. is a milk protein. 7. involves the hydrolysis of proteins to amino acids. 8. is a protein enzyme that breaks down starch in the mouth. 9. is the transformation of a liquid into a soft semi-solid or solid mass. 10. proteins are used for their foaming properties in desserts, cakes, and whipped toppings. HIDDEN MESSAGE: Elementary school children frequently use this casein-based adhesive that was introduced by Borden® over 50 years ago. Hint: There is a picture of Elsie the cow on the container. 156 Puzzling Proteins Name: _________________________________________________________ Section:_____________________________________ Find the words listed below in the word search. After all the words are found, the letters that are not used reveal a hidden message at the bottom of this sheet. Please write your answers in the provided space. HIDDEN MESSAGE: You should 157 References Growing Yeast: Sugar Fermentation. https://www.education.com/download-pdf/sciencefair/65029/ http://cnx.org/contents/b3c1e1d2-839c-42b0-a314-e119a8aafbdd@9.25. http://www.math.unl.edu/~jump/Center1/Labs/DenaturingProteins.pdf https://www.rookieparenting.com/how-do-fish-dive-and-rise-diving-ketchup-cartesian-diverexperiment/ https://www.scientificamerican.com/article/fruits-gone-bad-discover-enzymatic-browning/ https://www.whatdowedoallday.com/ice-cream-in-a-bag/ Institute of Food Technologists, IFT Experiments in Food Science Series School of Public Health. Carbohydrates: Quality matters. https://www.hsph.harvard.edu/nutritionsource/carbohydrates/ 158