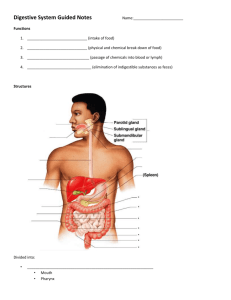
Absorption lecture transcript GAST41 • Nutrient Absorptio • The entire small intestine absorbs carbohydrates proteins and lipids. The major site for nutrient and uid absorption is the jejunum. • Before detailing the mechanism of nutrient digestion in carbs proteins and lipids and vitamins let us de ne the ve general patterns of nutrient digestion and absorption • As we have learned in GI lectures the mechanisms of digestion and absorption are interlinked • Lets examine the schematic diagra • Over here is the enterocyte the absorptive cell in the small intestine • Apical side of SI is endowed with microvilli, apical side of enterocyte faces the lumen • Tight junctions between the neighboring enterocytes • The interstitial spac • Basolateral surface of the epithelial cell faces i • Five patterns of digestion all written in the picture in the slid • In the case of glucose, it is a monomer a monosaccharide. In the lumen glucose as a monomer does not need any process of digestion. We will tackle how it enters the epithelial cell that is the enterocyte • Over here we have a protein a polymer and we see how a protein needs to be hydrolyzed into the lumen into amino acids before amino acids can enter the enterocyte to be absorbed by i • In the third model we see a disaccharide such as sucrose that needs to be hydrolyzed by a brush border enzyme into the two monosaccharides glucose and fructose. • We will see how these two monosaccharides enter the cell for absorptio • The 4th model: a tri-peptide for example enters the enterocyte and is hydrolyzed within it into three amino acids before being absorbed • Last mechanism o digestion applies to fats. We have a triacylglycerol molecule and we see how enzymes in the lumen break it into glycerol and fatty acids. The glycerol and fatty acids enter the enterocyte and the triacylcglycerol is resynthesized in the cytoplasm of the enterocyt n JB e t e 3 m fi t fi e fl n Page 1 of 9 • Digestion and absorption of carbohydrate • In panel A we see as an example of a carbohydrate a starch molecule. Panel A details how starch as a complex carb is digested in the lumen. We know that salivary alpha amylase and mainly pancreatic alpha amylase ensure complex carb digestion in the SI lumen • Salivary and pancreatic alpha amylases are endo-enzymes: these enzymes are able to digest the linear internal alpha-1,4 linkages between glucose residues. Cannot hydrolyze the terminal alpha 1,4 linkages between the two last sugars in the chain • Alpha amylase can't split the alpha 1,6 linkages at the branch point of amylopectin or the adjacent alpha 1,4 linkages • As a result linear glucose polymers are formed • starch digested in lumen not ready to be absorbed by epithelial cell. Need to be further hydrolyzed by speci c brush border enzymes on the apical side of the enterocyte • In panel B we talk about the digestion of oligosaccharides such as maltose, maltotriose, alpha-limit dextrins, and even lactose or even sucrose via these brush border enzymes • Over here as an example we have a brush border enzyme known as lactase it si able to break lactose into glucose and galactos • Once lactose is broken down into these two monosaccharides each monosaccharide is able to be transported across the epithelial cell via the SGLT1 secondary active transporter sodium -glucose symporter • SGLT1 is able to transport glucose or galactose via the electrochemical gradient of sodium which pushes sodium into the cell. • Another example is sucrose. • Sucrose a disaccharide consisting of glucose and fructose. You see in green the monomer fructose which is able to enter the cell membrane via facilitated transport by the transporter GLUT5 whereas glucose enters the enterocyte via SGLT • Sucrose should rst be hydrolyzed by sucrase a brush border enzyme into glucose and fructose • Brush border membrane does not only contain lactase • Also gluco-amylase hydrolyzes maltose or maltotriose • Sucrase isomaltase actually two enzymes sucrase and isomaltas 1 JB e e s fi fi Page 2 of 9 • Once these brush border enzymes have split the oligosaccharides and disaccharides into monosaccharides that are able to enter the enterocytes via either SGLT1 or GLUT • Not gonna talk about biochemistry and glucose phosphorylation into G6 • Just how glucose and fructose exit enterocyte into interstitial space by GLUT2 on the basolateral side • SGLT1 as a glucose transporter is a protein with 12 transmembrane domains and cytosolic amino and carboxyl termini. It has a sodium glucose stoichiometry of 2: • In panel B we see D-glucose • D galactose is identical to D galactose except H and OH at C4 are inverte • SGLT1 can only transport D-hexoses with a pyranose rin • The mechanism of digestion and digestion of proteins and oligopeptide • Let's explain the mechanisms of digestions and absorption of proteins and oligopeptide • Enterocyte • Notice that gastric peptidase and referring to pepsin and pancreatic peptidases (5 pancreatic enzymes: trypsin, chymotrypsin, elastase, carboxypeptidase A and carbozypeptidase B) can cleave proteins into oligopeptides and amino acid • These reactions don't only occur in the lumen of the small intestine but also can happen in the stomach. • Notice various peptidases at the brush border of enterocyte • Tetra peptidase tripeptidase, dipeptidas • These brush border enzymes break down the oligopeptides or dipeptides into individual amino acids which should be taken into the enterocyte via speci c transporter • PepT1: H+ oligopeptide cotransporter in this case a triaminoacid/ or a dipeptide is able to enter the enterocyte powered by the electrochemical gradient of H • Dipeptidase breaks down dipeptide into 2 amino acids each of which can enter the enterocyte via a transporter that is powered by the electrochemical gradient of Sodiu • If u have a dipeptide or tripeptide that entered the enterocyte there are further cytosolic peptidases that can break them down into amino acid • Cytosolic tripeptidase • Cytosolic dipeptidas • Not going to write what they do it's obviou 1 fi + d s P s s JB s g s e , e 5 s s m Page 3 of 9 • Each amino acid crosses to the interstitial space by facilitated transpor • Once an oligopeptide has been hydrolyzed in the lumen: in the schematic diagram a tripeptide • It was able to enter the enterocyte via PepT1 the H+/tripeptide cotransporte • Once it enters the cytosol tripeptidase hydrolyzes it into three amino acids that leave the cell via facilitated transpor • Pept1 uses H+ electrochemical gradient in order to restore the H+ gradient apical membrane of enterocyte carries Na/H exchanger NHX /NHE like in rena • The sodium that enters via Na/H exchanger its electrochemical gradient is restored by sodium / potassium pum Digestion and Absorption of Lipids Schematic diagram • You have on the diagram the bulk phase or what we call the intestinal lumen and of course you know it is alkaline. We call it the bulk or the water phas • Then you have the acid microclimate or the disequilibrium zone or what we also call the unstirred water layer. We call it acid microclimate because it is due to the presence of a sodium/proton exchanger on the apical surface of the enterocyte • Therefore you have the bulk water phase i.e. intestinal lumen alkaline and acid microclimate in close proximity to the apical surface of the enterocyte and then the apical membrane of the enterocyte • We have taken how emulsion droplets in the intestinal lumen are broken down into vesicles and how these vesicles are transformed into mixed micelles. • Once the emulsion droplet gives rise to the vesicle you will have pancreatic lipase as well as bile salt adsorbing on the surface of these vesicles. The addition of more bile salts creates mixed micelles. Within mixed micelles are fatty acid • Hydrophobic tails are inward. • Polar heads on the outer surface of the mixed micelle • The mixed micelles are the ones that carry the lipid product thought the acidic microclimate (acidic unstirred layer) of the surface of the enterocyt • Whether it is the monoacylglycerols or the fatty acids or the lipophospholipids or cholesterol can leave the mixed micelles and they all enter the acidic microclimate where their protonation is favored in acidic medium. • As an end result the lipid can enter the enterocyte by simple diffusio • Or incorporated in the enterocyte membrane by collision • A carrier protein known as FAT/CD36 can carry it in the cel r l t e s n e JB l . t p : Page 4 of 9 • These are the three mechanisms by which the lipid products carried by the mixed micelles can enter the enterocytes ◦ Simple diffusio ◦ Incorporation into membrane by collisio ◦ A carrier-mediated transpor • What happens upon the entry of the products of the mixed micelles into the enterocytes I am sure that you know from basic biochem that once in the enterocyte there is re-esteri cation of digestive lipids, and formation and secretion of chylomicrons. The enterocyte re- esteri es the digested lipid components and assembles them into chylomicrons. The enterocyte secretes chylomicrons into the lymphatic circulation during feeding a vLDL into lymphatic circulation during fasting Absorption of vitamins Absorption of Fat-soluble Vitamins A, D, E, & K • The intestinal absorption of these vitamins follows the pathways of lipid transport and absorptio • After ingestion fat soluble vitamins are released from their association with proteins in a meal by the acidity of the gastric juice and by proteolysis • Carboxyl ester hydrolases in the pancreatic juices and in mucosal brush border liberate free vitamins from their ester • In the proximal intestine, fat-soluble vitamins incorporate with other lipid product into emulsion droplets, vesicles, that bud off from emulsion droplets, and mixed micelles which ferry them into the enterocyte surface for uptak • By simple diffusion or through speci c transporter Absorption of Water-soluble vitamins: folate and B1 Dietary Folate Absorption • Medicinal folat • Its medicinal form is pteroyl mono glutamat • Folate present in food is pteroyl glutamate 7 (pteroylpolyglutamate • THF is cofactor in biochemical reactions involved in the transfer of 1-carbon groups • When we talk about medicinal folate you know that the recommended RDA is 200 micrograms for men and 180 micrograms for women. More than doubled in ) . JB e 2 s n : e fi s t : : fi n : fi e n ? Page 5 of 9 pregnant women. Folic acid supplementation during pregnancy reduces the risk of neural tube defect • We've also learned that THF is essential for synthesis of thiamine and purines critical components of DN • Let's learn about folate absorptio • This is the small intestine speci cally the jejunum (proximal small intestine • The intestinal absorption of dietary folate pteroylglutamate n(7) requires deconjugation by a brush border peptidase to pteroyl mono glutamate which then enters the enterocyte through a transporter • Here you go the carrie • The deconjugation is catalyzed by folate conjugate which is a zinc-activated exopeptidase present in the brush border membrane. It removes glutamate residues from dietary folate in a stepwise fashion before absorption of pteroyl mono glutamate • This stepwise hydrolysis of the polyglutamate chain of dietary folate is the ratelimiting step in folate digestion and absorptio • Remember, both folate de-conjugation and folate absorption occur only in the proximal small intestine, basically beginning of the jejunum and maximally active at a pH of 5 • Once inside the enterocyte, the pteroyl mono glutamate exits across the basolateral membrane through an unknown transporter. Within the enterocyte also, the enterocyte may reduce some of the pteroyl monoglutamte to dihydrofolate and then to THF and it can also methylate some of the THF to form N-5-methyl-TH Cobalamin (vitamin B12) absorptio Synthesized only by microorganisms and not by human cells. • The primary source of B12 in human is the ingestion of animal products such as meat, sh, shell sh, eggs, and to a limited extent, milk. Not present in vegetables or fruits, strict vegetarians are at risk of developing dietary cobalamin de ciency. • The load of cobalamin presented to the small intestine is derived about equally from two sources, rst diet and second the biliary secretions as a result of the enterohepatic secretion of cobalamin. • After its absorption, cobalamin is delivered throughout the body, excess secreted by liver into bile where it can once be reabsorbed by the small intestine and recirculated • Major steps in cobalamin absorption F ) fi JB n . : n n fi A r fi s fi . fi Page 6 of 9 • We have to say that steps 1-8 show the fate of dietary cobalamin • Steps 4-8 show the role of intrinsic factor • Bile carries cobalamin into the duodenum • Vitamin B12 binds to haptocorin in the stomach and then to intrinsic factor in the small intestine before it is endocytose by enterocytes in the ileum. • How is this done • First cobalamin is bound to proteins in a meal. The acid pH and the enzyme pepsin in the stomach release cobalamin from the dietary protein. The gastric glands secrete haptocorin which then binds to cobalami • Gastric parietal cells secrete intrinsic factor • Pancreas secretes bicarbonate and protease • Cobalamin is release after proteolytic degradation of haptocori • The intrinsic factor-cobalamin complex forms and ileal enterocytes absorb this complex • You should know that cobalamin is liberated within the enterocyte by mechanisms that have not yet been established. Within it, it binds to transcobalamin II, which is required for cobalamin exit from the enterocyte. Now we talk about the absorption of two minerals calcium and iro Absorption of Calciu A schematic diagram showing you the small intestine You have the lumen over here and enterocyt Before explaining the absorption of calcium, let me indicate to you and I know you didn't take endocrinology yet, that calcium absorption is regulated primarily by vitamin D. It occurs via passive paracellular diffusion throughout the small intestine. Can occur also via active transport only in the duodenum (active trans cellular transport Actively in duodenum, and paracellularly (passively) throughout the SI • SI absorbs calcium by these two mechanism. (Wow repeats a lot • The passive paracellular pathway predominates, and is not under control of vitamin D • Calcium enters the enterocyte across the apical membrane through a channel. Inside the enterocyte the calcium is buffered by binding proteins such as calbindin • Calcium and calbindin form a complex. • Even this complex can be taken by intracellular organelles such as th ER • Enterocyte extrudes calcium across the basolateral membrane through a calcium ATPase (pump) and through a sodium-calcium Na/Ca X (exchanger . . . ) n ) JB n n s e m ? ) . . Page 7 of 9 • The active form of vitamin D (1,25 dehydro) stimulates all the steps in trans cellular calcium absorptio Absorption of non-heme and heme iron in the duodenu • Non heme iron may be either ferric or ferrou • Ferric Fe3+ tends to form salt complexes with anions very easily. Ferric nonheme iron is not readily absorbed and not soluble at pH values higher than 3 • Ferrous iron Fe2+ does not complex easily and is soluble at pH values as high as 8 • Vitamin C forms soluble complexes with iron and reduces iron from the ferric to ferrous state, enhancing its absorption • Tannins, compounds in tea, form insoluble complexes with iron and lower its absorption. • Let me remind you that the absorption of non-heme iron is solely restricted to the duodenum • Mechanism of absorption • Step 1A • An enzyme called ferric reductase D-cytochrome B related to cytB is able to reduce non-heme ferric into ferrous iron. This is very important because only non heme iron in the ferrous state can enter the enterocyte. In fact, enterocyte in the duodenum can take up the non-heme iron across the apical membrane through a transmembrane transporter called the divalent metal transporter DMT1 • This bimetal transporter cotransports non-heme ferrous iron with H+ • Non heme ferrous iron within the cytoplasm binds to mobilferrin protein which transports the non-heme ferrous ion to basolateral membrane of the enterocyte which then translocates the ferrous iron across the basolateral membrane through a transporter called FP1 or ferroportin transporter 1 • Following the exit of non-heme ferrous ion from the enterocyte via the FP1 an ezyme called ferro-oxidase hephaestin oxidizes the ferrous ion to the ferric state that then binds to a plasma transferrin protein for carrying it into the blood. Once in the circulation, non-heme iron bound to transferrin is ultimately deposited in all body tissues, but it has a preference to be deposited in the liver into the reticuloendothelial system where it binds to apoferritin to form ferritin which is the major storage form of iron. • Smaller amounts of storage iron in insoluble forms called hemosiderin . . . JB . m . s . : n : . Page 8 of 9 e JB s e s n n : fl fl ! fl fl Page 9 of 9 fl fl • therefore the absorption of non-heme iron occurs almost exclusively as ferrous ion which crosses the duodenum enterocyte membrane through DMT1 driven by the H+ electrochemical gradient which is maintained by the NHE and NKATPas • Heme enters the enterocyte by an unknown mechanism • Inside the enterocyte hemeoxygenase releases ferric heme iron which is then reduced to ferrous iron • The cytoplasmic ferrous iron then binds to mobilferrin for exit across the enterocyte basolateral membrane • This heme ferrous exits the enterocyte most probably thorough the basolateral FP1 as well • The ferrooxidase activity of hephaestin converts ferrous to ferrous for transport into blood bound to transferrin Brie y uid absorption: water and electrolyte Fluid balance in GI tract In ow of secretions by Dr Nassar in secretion lectur SI major site for uid absorptio Colon major site for reabsorption of water and electrolyte • The major site for uid absorption is the SI and major reabsorption up to 95% of uid i.e. water and electrolyte reabsorption occurs in the colon itself. • Let's talk about electrolyte absorptio • The small intestine, mainly the jejunum, absorbs water, sodium, chloride, and potassium. Small intestine secretes bicarbonate. • The colon absorbs water, sodium and chloride. It secretes potassium and bicarbonate • A scheme showing you different mechanisms used by SI and colon to absorb sodium and chloride. Not responsible for each mechanism. Simply put them to illustrate to us how GI system is very and highly involved in the absorption of water and electrolytes • Another scheme to show you cellular mechanisms of chloride absorption in SI and colon • Not responsible for these mechanism • Should know what kind of ions absorbed by SI and colon and which ones are secreted in them. The END