Chapter 10 - The Electrical Conductivity of Tissues
advertisement

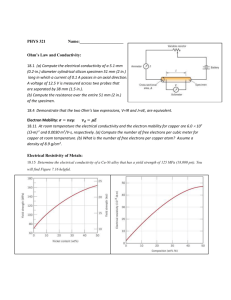
Roth, B. J. “The Electrical Conductivity of Tissues.” The Biomedical Engineering Handbook: Second Edition. Ed. Joseph D. Bronzino Boca Raton: CRC Press LLC, 2000 10 The Electrical Conductivity of Tissues Bradley J. Roth Oakland University 10.1 10.1 10.2 10.3 10.4 Introduction Cell Suspensions Fiber Suspensions Syncytia Introduction One of the most common problems in bioelectric theory is the calculation of the potential distribution, Φ (V), throughout a volume conductor. The calculation of Φ is important in impedance imaging, cardiac pacing and defibrillation, electrocardiogram and electroencephalogram analysis, and functional electrical stimulation. In bioelectric problems, Φ often changes slowly enough so that we can assume it to be quasistatic [Plonsey, 1969]; that is, we ignore capacitive and inductive effects and the finite speed of electromagnetic radiation. (For bioelectric phenomena, this approximation is usually valid for frequencies roughly below 100 kHz.) Under the quasistatic approximation, the continuity equation states that the divergence, ∇•, of the current density, J (A/m2), is equal to the applied or endogenous source of electrical current, S (A/m3): ∇ • J = S. (10.1) In regions where there are no such sources, S is zero. In these cases, the divergenceless of J is equivalent to the law of conservation of current that is often invoked when analyzing electrical circuits. Another fundamental property of a volume conductor is that the current density and the electric field, E (V/m), are related linearly by Ohm’s Law, J = g E, (10.2) where g is the electrical conductivity (S/m). Finally, the relationship between the electric field and the gradient, ∇, of the potential is E = –∇ Φ . (10.3) The purpose of this chapter is to characterize the electrical conductivity. This task is not easy, because g is generally a macroscopic parameter (an “effective conductivity”) that represents the electrical properties of the tissue averaged in space over many cells. The effective conductivity can be anisotropic, complex (containing real and imaginary parts), and can depend on both the temporal and spatial frequencies. © 2000 by CRC Press LLC Before beginning our discussion of tissue conductivity, let us consider one of the simplest volume conductors: saline. The electrical conductivity of saline arises from the motion of free ions in response to a steady electric field, and is on the order of 1 S/m. Besides conductivity, another property of saline is its electrical permittivity, ε (S s/m). Permittivity is related to the dielectric constant, κ (dimensionless), by ε = κ εo, where εo is the permittivity of free space, 8.854 × 10-12 S s/m. (The permittivity is also related to the electric susceptibility, χ, by ε = εo + χ.) Dielectric properties arise from a bound charge that is displaced by the electric field, creating a dipole. Dielectric behavior also arises if the applied electric field aligns molecular dipoles (such as the dipole moments of water molecules) that normally are oriented randomly. The DC dielectric constant of saline is similar to that of water (about κ = 80). The movement of free charge produces conductivity, whereas stationary dipoles produce permittivity. In steady state the distinction between the two is clear, but at higher frequencies the concepts merge. In such a case, we can combine the electrical properties into a complex conductivity, g′: g′ = g + iωε , (10.4) where ω (rad/s) is the angular frequency (ω = 2πf, where f is the frequency in Hz) and i is – 1 . The real part of g′ accounts for the movement of charge that is in phase with the electric field; the imaginary part accounts for out-of-phase motion. Both the real and the imaginary parts of the complex conductivity may depend on the frequency. For many bioelectric phenomena, the first term in Eq. (10.4) is much larger than the second, so the tissue can be represented as purely conductive [Plonsey, 1969]. (The imaginary part of the complex conductivity represents a capacitive effect, and therefore technically violates our assumption of quasistationarity. This violation is the only exception we make to our rule of a quasistatic potential.) 10.2 Cell Suspensions The earliest and simplest model describing the effective electrical conductivity of a biological tissue is a suspension of cells in a saline solution [Cole, 1968]. Let us consider a suspension of spherical cells, each of radius a (Fig. 10.1a). The saline surrounding the cells constitutes the interstitial space (conductivity σe), while the conducting fluid inside the cells constitutes the intracellular space (conductivity σi). (We FIGURE 10.1 (A) A schematic diagram of a suspension of spherical cells; the effective conductivity of the suspension is IL/VA. (B) An electric circuit equivalent of the effective conductivity of the suspension. © 2000 by CRC Press LLC shall follow Henriquez [1993] in denoting macroscopic effective conductivities by g and microscopic conductivities by σ.) The cell membrane separates the two spaces; a thin layer having conductivity per unit area Gm (S/m2) and capacitance per unit area Cm (F/m2). One additional parameter—the intracellular volume fraction, f (dimensionless)—indicates how tightly the cells pack together. The volume fraction can range from nearly zero (a dilute solution) to almost 1 (spherical cells cannot approach a volume fraction of 1, but tightly packed nonspherical cells can). For irregularly shaped cells, the cell “radius” can be difficult to define. In these cases, it is easier to specify the surface-to-volume ratio of the tissue (the ratio of the membrane surface area to tissue volume). For spherical cells, the surface-to-volume ratio is 3f/a. We can define operationally the effective conductivity, g, of the cell suspension by the following process (Fig. 10.1a): Place the cell suspension in a cylindrical tube of length L and cross-sectional area A (be sure L and A are large enough so the volume contains many cells). Apply a DC potential difference V across the two ends of the cylinder (so that the electric field has strength V/L) and measure the total current, I, passing through the suspension. The effective conductivity is IL/VA. Deriving an expression for the effective conductivity of a suspension of spheres in terms of microscopic parameters is an old and interesting problem in electromagnetic theory [Cole, 1968]. For DC fields, the effective conductivity, g, of a suspension of insulating spheres placed in a saline solution of conductivity σe is g= ( )σ . 2 1− f 2+f (10.5) e For most cells, Gm is small enough so that the membrane behaves essentially as an insulator at DC, in which case the assumption of insulating spheres is applicable. The net effect of the cells within the saline is to decrease the conductivity of the medium (the decrease can be substantial for tightly packed cells). The cell membrane has a capacitance of about 0.01 F/m2 (or, in traditional units, 1 µF/cm2), which introduces a frequency dependence into the electrical conductivity. The suspension of cells can be represented by the electrical circuit in Fig. 10.1b: Re is the effective resistance to current passing entirely through the interstitial space; Ri is the effective resistance to current passing into the intracellular space; and C is the effective membrane capacitance. (The membrane conductance is usually small enough so that it has little effect on the suspension behavior, regardless of the frequency.) At low frequencies, all of the current is restricted to the interstitial space, and the electrical conductivity is given approximately by Eq. (10.5). At large frequencies, C shunts current across the membrane, so that the effective conductivity of the tissue is again resistive: g= ( ) ( ) (2 + f ) σ + (1 − f ) σ 2 1 − f σ e + 1 + 2f σ i e σe . (10.6) i At intermediate frequencies, the effective conductivity has both real and imaginary parts, because the membrane capacitance contributes significantly to the effective conductivity. In these cases, Eq. (10.6) still holds if σi is replaced by σ i∗ , where σ∗i = σ i Ym a , σ i + Ym a with Ym = Gm + i ω C m . (10.7) Figure 10.2 shows the effective conductivity (magnitude and phase) as a function of frequency for a typical tissue. The increase in the phase at about 300 kHz is sometimes called the “beta dispersion”. © 2000 by CRC Press LLC FIGURE 10.2 The magnitude and phase of the effective conductivity as a function of frequency, for a suspension of spherical cells: f = 0.5; a = 20 µm; σe = 1 S/m; σi = 0.5 S/m; Gm = 0; and Cm = 0.01 F/m2. 10.3 Fiber Suspensions Some of the most interesting electrically active tissues, such as nerve and skeletal muscle, are better approximated as a suspension of fibers rather than as a suspension of spheres. This difference has profound implications because it introduces anisotropy: The effective electrical conductivity depends on direction. Henceforth, we must speak of the longitudinal effective conductivity parallel to the fibers, gL, and the transverse effective conductivity perpendicular to the fibers, gT. (In theory, the conductivity could be different in three directions; however, we consider only the case in which the electrical properties in the two directions perpendicular to the fibers are the same.) The conductivity is no longer a scalar quantity, but is a tensor instead, and therefore must be represented by a 3 × 3 symmetric matrix. If we choose our coordinate axes along the principle directions of this matrix (invariably, the directions parallel to and perpendicular to the fibers), then the off-diagonal terms of the matrix are zero. If, however, we choose our coordinate axes differently, or if the fibers curve so that the direction parallel to the fibers varies over space, we have to deal with tensor properties, including off-diagonal components. When the electric field is perpendicular to the fiber direction, a suspension of fibers is similar to the suspension of cells described above (in Fig. 10.1a, we must now imagine that the circles represent crosssections of cylindrical fibers, rather than cross-sections of spherical cells). The expression for the effective transverse conductivity of a suspension of cylindrical cells, of radius a and intracellular conductivity σi , placed in a saline solution of conductivity σe , with intracellular volume fraction f, is [Cole, 1968] © 2000 by CRC Press LLC FIGURE 10.3 An electrical circuit representing a one-dimensional nerve or muscle fiber: ri and re are the intracellular and extracellular resistances per unit length (Ω/m); rm is the membrane resistance times unit length (Ω m); and cm is the membrane capacitance per unit length (F/m). gT = (1 − f ) σ + (1 + f ) σ (1 + f ) σ + (1 − f ) σ e ∗ i e ∗ i σe , (10.8) where Eq. (10.7) defines σi∗. At DC (and assuming that Gm = 0) Eq. (10.8) reduces to gT = 1− f σe . 1+ f (10.9) When an electric field is parallel to the fiber direction, a new behavior arises that is fundamentally different from that observed for a suspension of spherical cells. Let us return for a moment to our operational definition of the effective conductivity. Surprisingly, the effective longitudinal conductivity of a suspension of fibers depends on the length L of the tissue sample used for the measurement. To understand this phenomenon, we must consider one-dimensional cable theory [Plonsey, 1969]. A single nerve or muscle fiber can be approximated by the circuit shown in Fig. 10.3. Adopting the traditional electrophysiology definitions, we denote the intracellular and extracellular resistances per unit length along the fiber by ri and re (Ω/m), the membrane resistance times unit length by rm (Ω m), and the capacitance per unit length by cm (F/m). The cable equation governs the transmembrane potential, Vm : λ2 ∂2 Vm ∂x 2 =τ ∂Vm + Vm , ∂t (10.10) where τ is the time constant, rmcm, and λ is the length constant, r m ⁄ ( r i + r e ) . For a truncated fiber of length L (m) with sealed ends, and with a steady-state current I (A) injected into the extracellular space at one end and removed at the other, the solution to the cable equation is Vm = I re λ ( ), cosh(L 2λ ) sinh x λ (10.11) where the origin of the x-axis is at the midpoint between electrodes. The extracellular potential, Ve , consists of two terms: One is proportional to x, and the other is re /(ri+re) times Vm . We can evaluate Ve at the two ends of the fiber to obtain the voltage drop between the electrodes, ∆Ve © 2000 by CRC Press LLC ∆Ve = ri re r I L + e 2λ tanh L 2λ . ri + re ri ( ) (10.12) Two limiting cases are of interest. If L is very large compared to λ , the extracellular voltage drop reduces to ∆Ve = ri re LI ri + re L >> λ . (10.13) The leading factor is the parallel combination of the intracellular and extracellular resistances. If, on the other hand, L is very small compared to λ , the extracellular voltage drop becomes ∆Ve = re L I L << λ . (10.14) In this case, the leading factor is the extracellular resistance alone. Physically, there is a redistribution of current into the intracellular space that occurs over a distance on the order of a length constant. If the tissue length is much longer than a length constant, the current is redistributed completely between the intracellular and extracellular spaces. If the tissue length is much smaller than a length constant, the current does not enter the fiber, but instead is restricted to the extracellular space. If either of these two conditions is met, then the effective conductivity (IL/A∆Ve , where A is the cross-sectional area of the tissue strand) is independent of L. However, if L is comparable to λ , the effective conductivity depends on the size of the sample. A complete model of the effective conductivity of a suspension of fibers must take this behavior into account. To avoid having the effective conductivity depend on the length of the sample used for the measurement, Roth et al. [1988] recast the expression for the effective longitudinal conductivity in terms of spatial frequency, k (rad/m). There are two advantages to this approach. First, the temporal and spatial behavior are both described using frequency analysis. Second, a parameter derived from a specific source geometry is not necessary: the spatial frequency dependence becomes a property of the tissue, not the source. The expression for the DC effective longitudinal conductivity is (1 − f ) σ + f σ e gL = 1+ f σi (1 − f )σ i (10.15) 1 e 1 1+ λk 2 To relate the effective longitudinal conductivity to Eqs. (10.13) and (10.14), note that 1/k plays the same role as L. If kλ <<1, gL reduces to (1-f)σe + fσi, which is equivalent to the parallel combination of resistances in Eq. (10.13). If kλ >>1, gL becomes (1-f)σe, implying that the current is restricted to the interstitial space, as in Eq. (10.14). Equation (10.15) can be generalized to all temporal frequencies by defining λ in terms of Ym instead of Gm [Roth et al., 1988]. Figure 10.4 shows the magnitude and phase of the longitudinal and transverse effective conductivities as functions of the temporal and spatial frequencies. The measurement of effective conductivities is complicated by the traditionally used electrode geometry. Most often, a uniform electric field is not (or cannot be) applied to the tissue. Typically, one uses a four-electrode technique [Steendijk et al., 1993], in which two electrodes inject current and two other electrodes measure the potential (Fig. 10.5). Gielen et al. [1984] used this method to measure the electrical properties of skeletal muscle and found that the effective conductivity depended on the interelectrode distance. Roth [1989] reanalyzed Gielen et al.’s data using the spatial frequency dependent model and © 2000 by CRC Press LLC FIGURE 10.4 The magnitude (a, c) and phase (b, d) of the effective longitudinal (a, b) and transverse (c, d) effective conductivities, calculated using the spatial, k, and temporal, ω, frequency model. The parameters used in this calculation are Gm = 1 S/m2; Cm = 0.01 F/m2; f = 0.9; a = 20 µm; σi = 0.55 S/m; and σe = 2 S/m. Source: Roth, B. J., Gielen, F. L. H., and Wikswo, J. P., Jr. 1988. Math. Biosci., 88:159-189. With permission. FIGURE 10.5 A schematic diagram of the four-electrode technique for measuring tissue conductivities. Current, I, is passed through the outer two electrodes, and the potential, V, is measured between the inner two. The interelectrode distance is s. found agreement with some of the more unexpected features of their data (Fig. 10.6). Table 10.1 contains typical values of skeletal muscle effective conductivities and microscopic tissue parameters. Table 10.2 lists nerve effective conductivities. © 2000 by CRC Press LLC FIGURE 10.6 The calculated (A) amplitude and (B) phase of gL (solid) and gT (dashed) as a function of frequency, for an interelectrode distance of 0.5 mm; (C) and (D) show the quantities for an interelectrode distance of 3.0 mm. Circles represent experimental data; gL (filled), gT (open). Source: Roth, B. J. 1989. Med. Biol. Eng. Comput., 27:491-495. With permission. TABLE 10.1 Skeletal Muscle Ref. gL gT Note Macroscopic Effective Conductivities (S/m) Gielen et al., 1984 0.35 0.20 0.52 0.70 0.67 Epstein and Foster, MBEC, 21:51 Rush et al., Circ Res, 12:40 0.086 0.092 0.076 0.32 0.040 10 Hz, IED = 3 mm 10 Hz, IED = 0.5 mm 20 Hz, IED = 17 mm 100 kHz, IED = 17 mm 0.1 sec pulse Microscopic Tissue Parameters Gielen et al., MBEC, 24:34 σi (S/m) σe (S/m) f Cm (F/m2) Gm (S/m2) 0.55 2.4 0.9 0.01 1.0 Note: (IED = interelectrode distance). TABLE 10.2 Nerve Ref. gL gT Note Macroscopic Effective Conductivities (S/m) Tasaki, J., Neurophysiol, 27:1199 Ranck & BeMent, Exp Neurol, 11:451 0.41 0.57 0.01 0.083 Toad sciatic nerve Cat dorsal column, 10 Hz Microscopic Tissue Parameters Roth & Altman, Med Biol Eng Comput, 30:103 σi (S/m) σe (S/m) f Cm (F/m2) Gm (S/m2) 0.64 1.54 0.35 1 0.44 Note: Volume fraction of myelin = 0.27. Gm is proportional to axon diameter; the above value is for an axon with outer diameter of 6.5 µm. © 2000 by CRC Press LLC FIGURE 10.7 A circuit representing a two-dimensional syncytium (i.e., a bidomain). The lower array of resistors represents the intracellular space, the upper array represents the interstitial space, and the parallel resistors and capacitors represent the membrane. 10.4 Syncytia Cardiac tissue is different from the other tissues we have discussed in that it is an electrical syncytium: the cells are coupled through intercellular junctions. To the extent that we can justify a continuum model and thereby ignore the discrete cellular structure of the tissue, we can use the bidomain model to describe the electrical properties of cardiac muscle [Henriquez, 1993]. The bidomain model is essentially a twoor three-dimensional cable model that takes into account the resistance of both the intracellular and the interstitial spaces (Fig. 10.7). Thus, the concept of current redistribution, discussed above in the context of the longitudinal effective conductivity of a suspension of fibers, now applies in all directions. Furthermore, cardiac muscle is markedly anisotropic. These properties make impedance measurements of cardiac muscle difficult to interpret [Plonsey and Barr, 1986]. The situation is complicated further because the intracellular space is more anisotropic than the interstitial space (in the jargon of bidomain modeling, this condition is known as “unequal anisotropy ratios”) [Henriquez, 1993]. Consequently, an expression for a single effective conductivity for cardiac muscle is difficult, if not impossible, to derive. In general, one must solve a pair of coupled partial differential equations simultaneously for the intracellular and interstitial potentials. For unequal anisotropy ratios, this solution is invariably found numerically. The bidomain model characterizes the electrical properties of the tissue by four effective conductivities: giL, giT , geL , and geT , where i and e denote the intracellular and interstitial spaces, and L and T denote the directions parallel to and perpendicular to the myocardial fibers. We can relate these parameters to the microscopic tissue properties by using an operational definition of an effective bidomain conductivity, similar to the operational definition given earlier. (The operational definition of a bidomain conductivity is most useful for thought experiments, since its actual implementation would be fraught with experimental difficulties.) To determine the interstitial conductivity, we first dissect a cylindrical tube of tissue of length L and cross-sectional area A (one must be sure that L and A are large enough so the volume contains many cells, and that the dissection damages a negligible amount of tissue). Next, we apply a drug to the tissue that makes the membrane essentially insulating (i.e., the length constant is much longer © 2000 by CRC Press LLC than L). Finally, we apply a DC potential difference, V, across the two ends of the cylinder and measure the total current, I, passing through the tissue. The effective interstitial conductivity is then IL/VA. This procedure could be performed twice, once with the fibers parallel to the long axis of the cylinder, and once with the fibers perpendicular to it. To determine the effective intracellular conductivities, the above procedure should be followed, except that the voltage difference must be applied to the intracellular space instead of to the interstitial space. Although the procedure would be extraordinarily difficult in practice, we can imagine two arrays of microelectrodes that impale the cells at both ends of the cylinder, and that maintain the potential at each end constant. Expressions have been derived for the effective bidomain conductivities in terms of the microscopic tissue parameters [Roth, 1988; Henriquez, 1993; Neu and Krassowska, 1993]. The effective conductivities in the direction parallel to the fibers are simplest. Imagine that the tissue is composed of long, straight fibers (like skeletal muscle) and that the intracellular space of these fibers occupies a fraction f of the tissue cross-sectional area. If the conductivity of the interstitial fluid is σe, then the effective interstitial conductivity parallel to the fibers, g eL, is simply geL = (1-f ) σe . (10.16) If we neglect the resistance of the gap junctions, we obtain a similar expression for the effective intracellular conductivity parallel to the fibers in terms of the myoplasmic conductivity, σi : g iL = f σ i . When the gap junctional resistance is not negligible compared to the myoplasmic resistance, the expression for g iL is more complicated: g iL = 1 f σi , πa 2 σ i 1+ bG (10.17) where G is the junctional conductance between two cells (S), b is the cell length (m), and a is the cell radius (m). The effective interstitial conductivity perpendicular to the fibers is identical with the DC transverse effective conductivity for skeletal muscle given in Eq. (10.9) g eT = 1− f σe . 1+ f (10.18) The effective intracellular conductivity perpendicular to the fibers is the most difficult to model, but a reasonable expression for g iT is g iT = 1 σi . b σi 1+ G (10.19) Table 10.3 contains measured values of the bidomain conductivities (see also Roth [1997]). Typical values of the microscopic tissue parameters are given in Table 10.3, although some are quite uncertain (particularly G). In our analysis of skeletal muscle, we accounted for the redistribution of current between the intracellular and interstitial spaces by the spatial and temporal frequency dependence of the effective conductivity. In the bidomain model, however, the current redistribution is accounted for by the pair of coupled equations that govern the intracellular and extracellular potentials. Thus, the conductivities are simpler for cardiac muscle than for skeletal muscle (no temporal or spatial frequency dependence); but for cardiac muscle, we must solve two equations for the intracellular and extracellular potentials instead of one. Roth and Gielen [1987] showed that, to a good approximation, the frequency-dependent conductivity model presented in Eq. (10.15) is equivalent to the bidomain model in the limit when g iT = 0. © 2000 by CRC Press LLC TABLE 10.3 Cardiac Muscle Ref. giL giT geL geT Macroscopic Effective Conductivities (Ventricular Muscle) (S/m) Clerc, J Physiol, 255:335 Roberts et al., Circ Res, 44:701 Roberts & Scher, Circ Res, 50:342 0.17 0.28 0.34 0.019 0.026 0.060 0.62 0.22 0.12 0.24 0.13 0.080 Microscopic Tissue Parameters Roth, Ann Biomed Eng, 16:609 Neu & Krassowska, 1993 σi (S/m) σe (S/m) f a (µm) b (µm) G (µS) 1 0.4 1 2 0.7 0.85 100 100 10 7.5 3 0.05 If the intercellular junctions contribute significantly to the intracellular resistance, the bidomain model will only approximate the tissue behavior [Neu and Krassowska, 1993]. For sufficiently large junctional resistance, the discrete cellular properties become important, and a continuum model no longer represents the tissue well. Interestingly, as the junctional resistance increases, cardiac tissue behaves less like a syncytium and more like a suspension of cells. Thus, we come full circle. We started by considering a suspension of cells, then examined suspensions of fibers, and finally generalized to syncytia. Yet, when the intercellular junctions in a syncytium are disrupted, we find ourselves again thinking of the tissue as a suspension of cells. Acknowledgments I thank Dr. Craig Henriquez for several suggestions and corrections, and Barry Bowman for carefully editing the manuscript. Defining Terms Anisotropic: Having different properties in different directions. Bidomain: A two- or three-dimensional cable model that takes into account the resistance of both the intracellular and the extracellular spaces. Cable theory: Representation of a cylindrical fiber as two parallel rows of resistors (one each for the intracellular and extracellular spaces) connected in a ladder network by a parallel combination of resistors and capacitors (the cell membrane). Conservation of current: A fundamental law of electrostatics, stating that there is no net current entering or leaving at any point in a volume conductor. Conductivity: A parameter (g) that measures how well a substance conducts electricity. The coefficient of proportionality between the electric field and the current density. The units of conductivity are siemens per meter (S/m). A siemens is an inverse ohm, sometimes called a “mho” in older literature. Interstitial space: The extracellular space between cells in a tissue. Ohm’s Law: A linear relation between the electric field and current density vectors. Permittivity: A parameter (ε) that measures the size of the dipole moment induced in a substance by an electric field. The units of permittivity are siemens second per meter (S s/m). Quasistatic: A potential distribution that changes slowly enough so that we can accurately describe it by the equations of electrostatics (capacitive, inductive, and propagation effects are ignored). Spatial frequency: A parameter governing how rapidly a function changes in space; k = 1/(2πs), where s is the wavelength of a sinusoidally varying function. Syncytium (pl., syncytia): A tissue in which the intracellular spaces of adjacent cells are coupled through intercellular channels, so that current can pass between any two intracellular points without crossing the cell membrane. Volume conductor: A three-dimensional region of space containing a material that passively conducts electrical current. © 2000 by CRC Press LLC References Cole, K.S. 1968. Membranes, Ions, and Impulses, University of California Press, Berkeley, CA. Gielen, F.L.H., Wallinga-de Jonge, W., and Boon, K.L. 1984. Electrical conductivity of skeletal muscle tissue: experimental results from different muscles in vivo. Med. Biol. Eng. Comput., 22:569-577. Henriquez, C.S. 1993. Simulating the electrical behavior of cardiac tissue using the bidomain model. Crit. Rev. Biomed. Eng., 21:1-77. Neu, J.C. and Krassowska, W. 1993. Homogenization of syncytial tissues. Crit. Rev. Biomed. Eng., 21:137-199. Plonsey, R. 1969. Bioelectric Phenomena, McGraw-Hill, New York. Plonsey, R. and Barr, R.C. 1986. A critique of impedance measurements in cardiac tissue. Ann. Biomed. Eng., 14:307-322. Roth, B. J., 1988. The electrical potential produced by a strand of cardiac muscle: A bidomain analysis. Ann. Biomed. Eng., 16:609-637. Roth, B.J. 1989. Interpretation of skeletal muscle four-electrode impedance measurements using spatial and temporal frequency-dependent conductivities. Med. Biol. Eng. Comput., 27:491-495. Roth, B.J. and Gielen, F.L.H. 1987. A comparison of two models for calculating the electrical potential in skeletal muscle. Ann. Biomed. Eng., 15:591-602. Roth, B. J. 1997. Electrical conductivity values used with the bidomain model of cardiac tissue. IEEE Trans. Biomed. Eng., 44:326-328. Roth, B.J., Gielen, F.L.H., and Wikswo, J.P., Jr. 1988. Spatial and temporal frequency-dependent conductivities in volume-conduction for skeletal muscle. Math. Biosci., 88: 159-189. Steendijk, P., Mur, G., van der Velde, E. T., and Baan, J. 1993. The four-electrode resistivity technique in anisotropic media: Theoretical analysis and application on myocardial tissue in vivo. IEEE Trans. Biomed. Eng., 40:1138-1148. Further Information Mathematics and Physics Jackson, J. D. Classical Electrodynamics, 2nd Ed., John Wiley & Sons, New York, 1975. (The classic graduatelevel physics text; a third edition is scheduled for 1999.) Purcell, E. M. Electricity and Magnetism, Berkeley Physics Course, Vol. 2, McGraw-Hill, New York, 1963. (An undergraduate physics text full of physical insight.) Schey, H. M. Div, Grad, Curl and All That, Norton, New York, 1973. (An accessible and useful introduction to vector calculus.) Bioelectric Phenomena and Tissue Models The texts by Cole and Plonsey, cited above, are classics in the field. Geddes, L.A. and Baker, L.E. 1967. The specific resistance of biologic material—a compendium of data for the biomedical engineer and physiologist. Med. Biol. Eng. Comput., 5:271-293. (Measured conductivity values for a wide variety of tissues.) Plonsey, R. and Barr, R. C. Bioelectricity, A Quantitative Approach. Plenum Press, New York, 1988. (An updated version of Plonsey’s Bioelectric Phenomena.) Polk, C. and Postow, E. (Eds.). CRC Handbook of Biological Effects of Electromagnetic Fields, CRC Press, Boca Raton, FL, 1986. Journals IEEE Transactions on Biomedical Engineering, Medical & Biological Engineering & Computing, Annals of Biomedical Engineering. © 2000 by CRC Press LLC