A D
advertisement

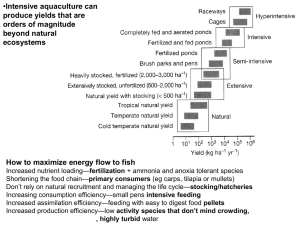
The food web Death and sedimentation Primary producers A1 A2 D Detritus and associated Microflora (bacteria/fungi) inedible Primary consumers H1 H2 detritivore herbivore P Productivity Biomass Secondary Productivity: Primary production supports a web of consumers—a simple example Defining some productivity terms A rate equation for biomass can be written as a mass balance bB/t Birth (production) term B Death (loss) term dB mB/t bB mB dt that is the net growth rate of biomass is the difference between the rate at which it is formed and lost. Productivi ty is the rate at which new biomass is formed. The birth term is called the productivi ty of the population , b is the specific production or growth rate and m is the specific death rate The productivity is a combination of the birth of new organisms and the growth of the organisms already present Similarly, the death process is a combination of death of organisms and weight loss by existing organisms. If the productivity (birth term) exceeds the death term the biomass is increasing, and if the death term is larger, the biomass is decreasing Biomass of a consumer dB 0 dt B t Time (t) dB 0 dt dB 0 dt dB bB mB b m B dt production minus losses or 1 dB bm B dt dR 0 Rotifer dt Biomass R dR 0 dt dR 0 dt t Time (t) dB and dt So if we measure B at any point in time and we can estimate the specific birth rate, we can then obtain the specific death rate by subtraction. 1 dB m b B dt A tiny organism like a rotifer is born at full size, so productivity amounts to measuring the rate at which new animals are born Rotifers carrying eggs •For a tiny consumer like a rotifer the birth rate is easy to estimate since the adult females carry their eggs around until they hatch •When they hatch they come out as full sized rotifers . •If we know the fraction of adults carrying egs and the average time it takes for eggs to hatch, we can calculate the birth rate. •Since the rotifers are born more or less full size, so there is no need to model or measure the growth of individuals. # eggs Egg Ratio E , # adults T avg time required for hatching (d) 1 hatching rate T E b d -1 T E 1 dB m T B dt dR 0 Rotifer dt Biomass R dR 0 dt dR 0 dt t Time (t) For a large organism like a fish, biomass production occurs mostly from individual growth. New born fish are so tiny that birth of individuals makes a negligible contribution to biomass production W4 W3 Wt 1 SGRt ln Wt Year classes W2 W1 The Wt represent the Weights of each year class •We can calculate the growth rate of biomass individual fish by weighing fish and determining their age and then seeing how much weight they gain each year. •The productivity of each age class is the Specific growth rate (SGR) of that age class times the total biomass of that age class in the population. This approach assumes that the size vs age relationship is relatively constant. W4 W3 SGRt ln P SGRt * Bt i Wt 1 Wt W2 W1 The Wtrepresent the Weights of each year class t Production sum of [specific growth rate times biomass present] for each of t age classes,and summed over all i species in the trophic category By this method the average SGR for the whole population can be calculated as the weighted average over all age classes T SGR wt SGRt t 1 SGRt ln where wt Wt 1 Wt Bt , the fraction of Biomass in the t ' th age class B However there is a problem with this approach…??? However there is a problem with this approach. By considering only the gain in weight across age classes, this method ignores weight gained and lost within the same year, eg Gonad tissue Adult fish usually convert a considerable portion of their body mass to gonads and release it during spawning every year. Thus it does not add to next year’s weight and would not be recorded as growth We can quite easily correct for this by factoring in gonad production (add GSI) SGRt GSI where GSI (Gonadosom atic index) mature gonad weight total weight We would of course only make this GSI correction on adult age classes. How can we tell how old a fish is? Scales of a chum salmon Measure distances from scale center to each annulus along a chosen 2+ 3+ axis distance to annulus LA total scale length LT 4+ This allows us to construct a growth curve based on length and age. LA Age yr Convert the growth curve based on length to weights using a length-weight plot for the species Many types of bony structures are commonly used to determine age of fish Scale Otolith Opercular bone These three structures are all from the same 3+ year old 30 cm cutthroat trout The specific death rates can also be estimated from the population structure. This time we assume that the age structure of the population is constant, and that the numbers of individuals of each age within a sample reflects the proportion of that age group in the population. Assume we have a sample of 215 pike from a population. N 4 20 N 3 30 Nt 1 mt ln Nt N 2 55 N 1 110 N 215 20 m3 ln 0.41 30 30 m 2 ln 0.61 55 55 m1 ln 0.69 110 t t The Nt / Nt represent the proportion of the population in each year class 150 The survivorship curve assuming stable age structure for the population looks like this * 100 110 Nt 1 mt ln Nt m1 =0.69 # 55 * 50 m2 =0.61 30 * 20 m3 =0.41 0 0 1 3 * 5 Age Summary The rate of change of biomass for a species is usually modelled as the difference between its growth and loss processes dB B SGR GSI B m GSI B SGR m dt Productivity term loss term If productivi ty exceeds losses, the population is growing in biomass otherwise the population is decreasing in biomass For a stable population the two terms are equal. Productivity at different levels in the food web 500 x (0.1)3 g/m /yr 2 500 x (0.1)2 g/m /yr 2 500 x 0.1 g/m2/yr NPP around 500 g/m /yr 2 Zooplankton Benthic & epiphytic invertebrates PhytoplanktonDiet shift Benthic & epiphytic algae plus detritus Trophic link Net productivity at level n = the rate of growth of biomass at that level = [SGR +GSI] * Biomass = NPP (TE) n-1 Ecological efficiency (n) for a consumer at the nth trophic level Pn n Pn 1 Ecological efficiency of zooplankton is usually around 10% of NPP in lakes Variability?? Fecal pellets Zooplankton such as Daphnia filter-feed using currents generated by their thoracic appendages. Fecal pellets sediment rapidly to the bottom 80-95% of energy is lost at each trophic step, much of it as feces Assimilation efficiency Herbivores depends on diet ≈100% for sugary nectar ≈40-80% for small phytoplankton and filamentous algae <20% for mud and detritus Carnivores 60-70% for aquatic insects 70-90% for meat Fecal pellets The undigested material in the zooplankton fecal pellets was not assimilated. Assimilation efficiency depends on the digestibility of the diet Cellulose, chitin, lignin or other undigestible material makes AE low Ingested energy ─ egested energy = assimilated energy Assimilation efficiency (AE, %)= assimilated energy/ingested energy x 100 Exploitation efficiency or Consumption Efficiency (EE) In EEn 100 Pn 1 Exploitation efficiency is the consumption rate at a given trophic level divided by the productivity of the trophic level it feeds on. Zooplankton will have low EE (CE) when phytoplankton are sedimenting rapidly to the bottom before they are being eaten. If EE(CE) is high then most of the sedimentation will be in the form of fecal pellets, which sink more rapidly than individual cells. Zooplankton fecal pellets are good food for benthic invertebrates If EE for herbivorous zooplankton is low then dead (sedimenting) phytoplankton will be readily available for detritivores (zoobenthos) Activity is energetically expensive and high Metabolic rate means low Production efficiency Gross PE Endotherms ≈5% or less ≈1% some birds Ectotherms ≈10-30% for fish ≈ 5-15% insects Otter swim about rapidly and spend large amounts of energy looking for fish to eat Assimilated energy ─ respiration ─ excretion = production (growth) Net Production efficiency (NPE, %)= growth/assimilation x 100 Gross PE (%)=[assim/ingest x growth/assim] x100=growth/ingest x 100 Pelagic fish like kokanee salmon expend a huge amount of energy actively searching for prey--they have high basal metabolic rates low conversion efficiencies The deepwater sculpin sits on the bottom and ambushes unsuspecting prey. They have very low basal metabolic rates and high conversion efficiencies If these two species were fed the same amount of food, the sculpin would grow more than twice as fast as the salmon Copepod dominated communites have lower trophic efficiency than cladoceran dominated communities—possible reasons? Filter-feeding by a calanoid copepod •Copepods are rapid swmmers and generate feeding currents as they swim •Copepods filter-feed by generating currents with their 1st antennae and their thoracic appendages.. •Water from small eddy currents around the mouthparts is drawn over the fine setae of the maxillae, where small algae are collected and moved to the mouth. http://www.ucmp.berkeley.edu/arthropoda/crustacea/images/copepoda03.jpg Energy budget for herbivorous zooplantkon NPP = rate of formation of phytoplankton biomass S = rate of production of uneaten algae, mostly inedible species (sedimentation) F= rate of production of fecal pellets (sedimentation) The Bioenergetic budget for an organism C= G + M*A+ SDA+U+F Metabolic costs include basal metabolism, activity costs and specific dynamic action (costs of digestion etc)+ excretion Zooplankton production is the rate at which biomass (energy) becomes available for consumption by zooplanktivores Energetic losses in the food chain Less than 1% of the incident light energy is captured by photosynthesis as NPP. Productivity declines by about 10-fold for each trophic level in the food chain. Most of the losses are are in the form of waste heat. Some energy from each trophic level winds up in the detrital pool, and some of this remains buried as sediment (or soil) organic matter (fossilized) Pn n Pn 1 In An Pn Pn EEn , AEn , PEn , GPEn Pn 1 In An In Show n EEn AEn PEn n EEn GPEn Pelagic fish like kokanee salmon expend a huge amount of energy actively searching for prey--they have high basal metabolic rates low conversion efficiencies The deepwater sculpin sits on the bottom and ambushes unsuspecting prey. They have very low basal metabolic rates and high conversion efficiencies If these two species were fed the same amount of food, the sculpin would grow more than twice as fast as the salmon The Bioenergetic budget for an organism C= G + M*A+ SDA+F+U A Activity multiplier 5 4 Br3+ Mg3+ 3 Mg2+ Me3+ 2 Bmt2+ 1 Wa2+ Me2+ Bmt3+ Wa3+ 0 2.3 2.4 2.5 2.6 2.7 2.8 Log LDH activity •The anaerobic capacity of fish muscles is closely linked to amount of energy spent on Activity •Lactate dehydrogenase (LDH) is an important enzyme for anerobic respiration, and anaerobic respiration generates bursts of power--but is very inefficient and builds up an oxygen debt. There is a trade-off between power (the rate of energy consumption) and efficiency. (abs units per mg prot) LDH activity LDH vs body size for perch 600 500 400 300 200 100 fish diet inverts zooplankton 0.1 1 10 100 1000 body size (g) In order to keep growing carnivorous fish usually need to switch to larger and larger prey If they do not, the activity costs escalate rapidly, and fish fail to grow (stunting) Thus trophic position usually increases as the fish matures. Trophic niches filled by yellow perch In the foodweb of an unimpacted lake Benthic & epiphytic invertebrates Zooplankton Phytoplankton Diet shift Trophic link Pelagic food chain Benthic & epiphytic algae plus detritus Trophic ecology of yellow perch In the foodweb metal impacted lake X bottleneck Diet shift Trophic link X Zooplankton X X Benthic & epiphytic invertebrates Phytoplankton Pelagic food chain Benthic & epiphytic algae plus detritus Classenia, a predatory stonefly, and some of the stream insect larvae that it preys on Stoneflies prefer prey that are near the energetically optimum size, when they are given the opportunity to select from a variety of sizes. Yields of piscivorous fish are well correlated with primary productivity but are several orders of magnitude lower than PP Rivers support more fish biomass than lakes for the same TP Level? Why? Fish community biomass kg ● ha-1 1000 Rivers 100 10 Lakes 1 1 10 100 Total Phosphorus µg ● 1000 L-1 Log B = 0.94+ 0.52 (± 0.09) Log TP -0.18 (± 0.05) L/R, RMS=0.27, R2=0.71 Comparing regulated vs unregulated rivers ? Fish community biomass kg ● ha-1 1000 100 Warta River, Poland Oldman River, Alberta 10 50 m, 0.03 ha seine 1 1 10 100 Total Phosphorus µg ● L-1 1000 •Intensive aquaculture can produce yields that are orders of magnitude beyond natural ecosystems How to maximize energy flow to fish Increased nutrient loading—fertilization + ammonia and anoxia tolerant species Shortening the food chain—primary consumers (eg carps, tilapia or mullets) Don’t rely on natural recruitment and managing the life cycle—stocking/hatcheries Increasing consumption efficiency—small pens intensive feeding Increased assimilation efficiency—feeding with easy to digest food pellets Increased production efficiency—low activity species that don’t mind crowding, , highly turbid water Lepeophtheirus salmonis Many aquaculture proponents argue that aquaculture reduces harvesting pressure on wild fisheries. Salmonid aquaculture not very trophically efficient, food pellets made from by-catch of wild species Major water quality issues—nutrient pollution from cages, anti-fouling paint, antibiotics, habitat destruction Transmit diseases to wild salmonids—bacteria, viruses, protozoans, fungi, “fish lice” –parasitic copepods and other Crustacea Genetic problems when domestic escapees compete with or interbreed with wild fish Argulus Summarizing concepts on Secondary production •The organic matter produced by primary producers (NPP) is used by a web of consumers •NPP is used directly by primary consumers (herbivores and detritivores), which are in turn consumed by carnivores. •Measurement of 2o Production is done by estimating the rate of growth of individuals and multiplying by the number of individuals per unit area in the cohort (age or size group). •The efficiency of secondary production ranges from 5-20% (Avg 10%) at each trophic level. •Efficiency depends on several factors--palatability, digestibility, energy requirements for feeding (activity costs)(eg homeotherms vs poikilotherms , other limiting factors eg water, and nutrient quality of food. •Trophic efficiency can be represented as the product of CE*AE*PE, each of which is dependent on one or more of the above factors. •The yields of many important fisheries depends on a combination of NPP, the length ofthe food chain leading to the fish being harvested, and the efficiency of each step. •Many of the species that we harvest or very high in the food chain, so a great deal of NPP is required to support them. Pn n Pn 1 In An Pn Pn EEn , AEn , PEn , GPEn Pn 1 In An In Show n EEn AEn PEn n EEn GPEn If the productivity of a phytoplankton population is 4000 k J (kilo Joules) /yr / m2, If sedimentation rate of dead cells to the substrate constitutes 1600 kJ/m2yr, and the phytoplankton population is dB/dt=0. If the rain of zooplankton fecal pellets to the bottom is 1400 kJ/m2/yr. What is the assimilation efficiency of the zooplankton trophic level (assume that they are all feeding on phytoplankton). 1. 2. 3. 4. 5. 0.42 or 42% 0.60 or 60% 0.35 or 35% 0.25 or 25% None of these What is the exploitation efficiency EE (or Consumption efficiency CE) of the zooplankton trophic level 1. 2. 3. 4. 5. 0.42 or 42% 0.60 or 60% 0.35 or 35% 0.25 or 25% None of these If the net production efficiency of the zooplankton trophic level is 0.40 (40%) what is the ecological efficiency () of the trophic level 1. 0.15 or 15% 2. 0.05 or 5% 3. 0.10 or 10% 4. 0.25 or 25% 5. None of these If the zooplanktivorous fish are consuming zooplankton at the rate of 400 kJ/yr/m2, their EE (CE) is 1. 0.40 or 40% 2. 0.60 or 60% 3. 1.00 or 100% 4. 0.25 or 25% 5. None of these If the zooplanktivorous fish have an assimilation efficiency of 0.70 (70%) and Net production efficiency (NPE) of 0.20 (20%), the productivity at this trophic level is 1. 40 kJ/yr/m2 2. 56 kJ/yr/m2 3. 100 kJ/yr/m2 4. 280 kJ/yr/m2 5. None of these If in another lake with similar zooplankton productivity the planktivore fish productivity was 2 X higher, a possible explanation for this would be 1. the AE of the fish in that lake was 2X as high 2. the NPE of the fish in that lake was 2X as high 3. the EE (CE) in that lake was 2X as high 4. the AE*NPE in that lake was 2X as high 5. both b and d are true 6. both b and c are true Residence time and turnover of energy by trophic levels The standing stock of energy in the plankton is low but it is turned over rapidly, because the organisms are small, grow rapidly and don’t live long Planktonic Herbivore (50mg) life span 1 month A H1 P Phytoplankton (0.01mg, life span, few days H2 Benthic Detritivore (0.1 g) life span 1yr Carnivorous fish (100g) life span 5-10 yr Turnover is slower at higher trophic levels, since larger organisms accumulate energy over a longer life span—longer residence time and slower turnover