Georgidis Hartebes Genetics
advertisement

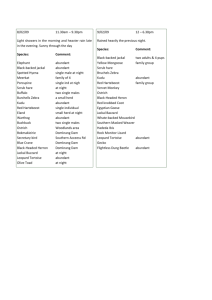
cokei796 H79-LAIK H8-NAIV H34-NAIR cokei747 H20-LAIK H13-RUMA H32-RUMA-s Laikipia Hartebeest: Exactly what are we attempting to conserve? H69-LAIK cokei748 swaynei10 swaynei9 swaynei13 swaynei7 swaynei8 swaynei45 swaynei5 swaynei6 RESULTS Nicholas Georgiadis and Collins Ouma Mpala Research Centre THANKS TO: Our collaborators were Drs. Al Roca, Olivier Hanotte, Nick Oguge, and Joel Ochieng. We thank Dr. Oystein Flagstad, Nasser Olwero, Tee Taylor, Robert Mills, Edward Parfet, Joseph Kioko, Dr. Richard Bagine, and KWS staff in Nakuru, Ruma and Nairobi NP’s. The project was supported by the Whitley Foundation, Lincoln Park Zoo, St. Louis Zoo, Mpala Wildlife Foundation, African Wildlife Foundation, Nancy and Lambeth Townsend, and Joan and Robert Weiss. RATIONALE Species’ geographical ranges are today so fragmented, it is often difficult to know whether remnant populations were once connected by dispersal and migration, or isolated by natural barriers. We need to know this information when action must be taken to conserve a species, for example, when individuals from one location are added to those in another. Fortunately, the required information can be reconstructed by analyzing patterns of genetic variation that is encoded in the DNA of individuals from different locations. We used this approach to examine the genetic makeup of hartebeest in Laikipia, asking, Exactly what, in the evolutionary sense, are we attempting to conserve? cokei794 H61-SERO H54-SERO H43-NAIR H7-NAIV cokei800 cokei786 H5-SERO H58-NAIV cokei797 We found only subtle genetic differences between hartebeest with Lelwel morphology (in Laikipia and Ruma NP) and those with Cokes morphology (in Naivasha, Meru NP, Nairobi NP, the Mara-Serengeti, and Ngorongoro), and the genetic transition between them appeared seamless. This was surprising because morphological differences between the two are striking, even over distances as short as 100 km (Fig. 1). H57-NAIV cokei35 cokei789 cokei783 H2-NGOR cokei798 H27-NGOR H12-NAIV cokei802 cokei792 cokei790 cokei803 cokei804 H9-NAIV cokei46 cokei806 cokei809 cokei807 cokei791 Previous analyses have suggested that different hartebeest forms in Africa diverged over the last two hundred thousand years when climate changes triggered continent-wide habitat shifts, ‘temporarily’ confining separate hartebeest populations to isolated savanna patches. Our results agree with this scenario, but further suggest that Cokes and Lelwel subsequently recontacted each other and interbred, presumably after the climate cycle reversed, and hartebeest spread as the savannas expanded once more. As a result, the populations in Laikipia, Ruma NP, and Meru NP are all hybrid, but each has a subtly unique genetic makeup (Table 1) and morphology. The former two have greater affinities with Lelwel, the latter, at least morphologically, with Cokes. Despite their morphological similarities, the Meru NP hartebeest were not closely aligned with Cokes in Nairobi NP and Naivasha in the genetic sense (Fig. 3). H3-SERO cokei795 cokei787 lelwel3 lelwel44 lelwel727 lelwel730 neumanni lelwel729 tora78 major57 lelwel72 tora33 swaynei68 lelwel32 lelwel42 lelwel73 lelwel4 major79 lelwel41 lelwel43 H18-LAIK H17-LAIK H76-LAIK-s lelwel74 H1-SERO cokei784 lelwel2 lelwel726 lelwel728 tora76 tora532 Naivasha Serengeti NP Nairobi Ngorongoro tora75 swaynei815 swaynei1 swaynei2 swaynei3 swaynei23 swaynei4 swaynei12 somali swaynei11 swaynei14 Laikipia Ruma NP HORN SHAPES AND SAMPLING LOCATIONS For a century hartebeest in this ecosystem have been known to be morphologically ‘different’ from hartebeest elsewhere (Fig. 1). They have been variously referred to as ‘Laikipia’, ‘Jackson’s’, ‘Kenya’, or ‘Lelwel’ hartebeest, but rarely with formal reference or definition. Many resemble true Lelwel hartebeest, with robust horns that stand upright then sweep backwards, but there is broad variation in horn shape and size. Lelwel hartebeest were once distributed between central Kenya and the Central African Republic, but persist today in highly fragmented remnants (Fig. 2). Hartebeest in Laikipia have long been considered to be hybrid between Lelwel and Cokes hartebeest, the common form in southern Kenya and northern Tanzania. major7 major732 Meru NP buselaphus major6 major58 Fig. 3. Cluster diagram of focal hartebeest populations based on genetic (microsatellite) similarities. Numbers represent % bootstrap values, an index of confidence in the robustness of each node. Nairobi Meru NP liecht757 liecht754 caama723 0.01 substitutions/site Fig. 4. Mitochondrial DNA genealogy of hartebeest, colour-coded by morphotype: cokei (yellow), lelwel (blue), swaynei (light green), tora (dark green), and major (red). Southern African hartebeest (lichtensteini and caama) were used as outgroups. Table 1. Values of Fst, an index of the degree to which pairs of populations are genetically subdivided (on a scale of 0-1), based on mitochondrial (below diagonal) and microsatellite data (above diagonal). Asterisks indicate values significantly greater than 0. NP Collecting samples with biopsy darts, from dung, and from incidental mortality, we compared the genetic makeup of hartebeest in Laikipia with those in Ruma NP, Meru NP, Naivasha, Nairobi NP, Mara-Serengeti NP, Ngorongoro, and elsewhere in Africa, using both mitochondrial DNA sequences and nuclear (microsatellite) markers. CONSERVATION IMPLICATIONS Laikipia Ruma Fig. 2. Distribution of hartebeest in Africa, with different symbols denoting different morphotypes. Symbols are filled grey if previously published mtDNA sequences from that location were used in this analysis, and black where new populations were added by this study. major731 major746 major735 major734 major736 District NP Fig. 1. Variation in hartebeest horn shape among the focal populations. Note the similarity between Nairobi (A. b. cokei) and Meru NP, and the hybrid appearance of Laikipia hartebeest. A conventional approach to conserving hartebeest might aim to preserve or restore populations wherever they occurred naturally, particularly in protected areas. A more contemporary approach would attempt to conserve or even recreate the fragmented and diverse remnants of intricate evolutionary processes that have been operating over vast space and time, such as the one described here involving hybridization between different hartebeest forms. For example, dwindling numbers in Meru NP prompted conservation managers to propose supplementing the Meru hartebeest with individuals imported from elsewhere. Similarities in gross morphology might suggest a suitable source population to be Naivasha or Nairobi NP. A preferable strategy would aim to conserve the evolutionary products of hybridization by breeding up the remaining individuals at Meru in a large, predator-free area. If supplementation from elsewhere becomes mandatory, the goal would be to maintain a population with hybrid characteristics. The Laikipia population is also declining and is nowhere formally protected. A strategic plan should be designed to conserve the remaining hartebeest in Laikipia, particularly those in Solio Ranch in the extreme south, which holds by far the highest densities (if necessary, moving them elsewhere in Laikipia or beyond, but still retaining their identity). Similarly, suitable alternative refuges should be identified to keep the Ruma NP hartebeest intact, should this protected area not survive extreme pressures from surrounding humanity. In Kenya there has been little unnatural mixing of wildlife populations by translocation, so opportunities to define and conserve ongoing evolutionary processes persist, and should be strenuously pursued. As this project has shown, the relevant research can be achieved by Kenyans through collaborations among national and international partners, in this case Mpala Research Centre, Kenyatta University, the Kenya Wildlife Service, the International Livestock Research Institute in Nairobi, and the Laboratory of Genomic Diversity in Maryland.