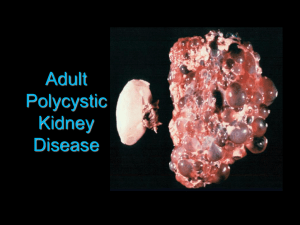
RENAL AUTOREGULATION IN HEALTH AND DISEASE
advertisement

Physiol Rev 95: 405–511, 2015 doi:10.1152/physrev.00042.2012 RENAL AUTOREGULATION IN HEALTH AND DISEASE Mattias Carlström, Christopher S. Wilcox, and William J. Arendshorst Department of Medicine, Division of Nephrology and Hypertension and Hypertension, Kidney and Vascular Research Center, Georgetown University, Washington, District of Columbia; Department of Physiology and Pharmacology, Karolinska Institutet, Stockholm, Sweden; and Department of Cell Biology and Physiology, UNC Kidney Center, and McAllister Heart Institute, University of North Carolina at Chapel Hill, Chapel Hill, North Carolina Carlström M, Wilcox CS, Arendshorst WJ. Renal Autoregulation in Health and Disease. Physiol Rev 95: 405–511, 2015; doi:10.1152/physrev.00042.2012.—Intrarenal autoregulatory mechanisms maintain renal blood flow (RBF) and glomerular filtration rate (GFR) independent of renal perfusion pressure (RPP) over a defined range (80 –180 mmHg). Such autoregulation is mediated largely by the myogenic and the macula densatubuloglomerular feedback (MD-TGF) responses that regulate preglomerular vasomotor tone primarily of the afferent arteriole. Differences in response times allow separation of these mechanisms in the time and frequency domains. Mechanotransduction initiating the myogenic response requires a sensing mechanism activated by stretch of vascular smooth muscle cells (VSMCs) and coupled to intracellular signaling pathways eliciting plasma membrane depolarization and a rise in cytosolic free calcium concentration ([Ca2⫹]i). Proposed mechanosensors include epithelial sodium channels (ENaC), integrins, and/or transient receptor potential (TRP) channels. Increased [Ca2⫹]i occurs predominantly by Ca2⫹ influx through L-type voltage-operated Ca2⫹ channels (VOCC). Increased [Ca2⫹]i activates inositol trisphosphate receptors (IP3R) and ryanodine receptors (RyR) to mobilize Ca2⫹ from sarcoplasmic reticular stores. Myogenic vasoconstriction is sustained by increased Ca2⫹ sensitivity, mediated by protein kinase C and Rho/Rho-kinase that favors a positive balance between myosin light-chain kinase and phosphatase. Increased RPP activates MD-TGF by transducing a signal of epithelial MD salt reabsorption to adjust afferent arteriolar vasoconstriction. A combination of vascular and tubular mechanisms, novel to the kidney, provides for high autoregulatory efficiency that maintains RBF and GFR, stabilizes sodium excretion, and buffers transmission of RPP to sensitive glomerular capillaries, thereby protecting against hypertensive barotrauma. A unique aspect of the myogenic response in the renal vasculature is modulation of its strength and speed by the MD-TGF and by a connecting tubule glomerular feedback (CT-GF) mechanism. Reactive oxygen species and nitric oxide are modulators of myogenic and MD-TGF mechanisms. Attenuated renal autoregulation contributes to renal damage in many, but not all, models of renal, diabetic, and hypertensive diseases. This review provides a summary of our current knowledge regarding underlying mechanisms enabling renal autoregulation in health and disease and methods used for its study. L I. II. III. IV. V. VI. VII. VIII. INTRODUCTION MAJOR MECHANISMS AND METHODS ... MECHANOSENSITIVE MECHANISM... CONDUCTANCE CHANGES... CALCIUM SIGNALING PATHWAYS... MODULATING AGENTS RENAL AUTOREGULATION IN DISEASE... CONCLUSIONS 405 408 431 436 439 444 452 471 I. INTRODUCTION A. Overview and Historical Perspective Arteries from various vascular beds often share functional characteristics. However, prominent differences exist in myogenic responses to changes in transmural pressure. These responses are greater in cerebral and renal than in mesenteric and hindlimb arteries (748, 750, 859, 886). Vascular smooth muscle cells (VSMCs) are derived from diverse embryological and developmental origins, and such lineage may account for heterogeneity of specialized function (1142, 1179). Renal vessels are formed by angiogenesis and vasculogenesis (479). VSMCs express fast and slow contractile gene programs, accounting for phasic and tonic phenotypes (1213). The aorta and efferent arterioles are examples of the tonic phenotype, whereas small resistance arteries and arterioles including the renal cortical radial (interlobular) artery and afferent arteriole are examples of the phasic phenotype. Moreover, there are significant differences in the magnitude of vasoconstrictor responses to KCl, norepinephrine (NE), and serotonin and to endothelium-dependent and -independent vasodilation between mouse aorta and smaller arterial segments (804). These variations likely reflect, in part, functional adaptations to meet the diverse roles of different arterial beds. 0031-9333/15 Copyright © 2015 the American Physiological Society 405 RENAL AUTOREGULATORY MECHANISMS Cerebral artery tone is modulated primarily by local metabolic, paracrine, and mechanical factors such as the partial pressure of carbon dioxide. The cerebral vasculature adjusts blood flow to the local metabolic demand independent of systemic hemodynamics. The cerebral vascular myogenic response mediates rapid and efficient autoregulation of cerebral blood flow that maintains a steady cerebral capillary pressure (356, 739). In contrast, mesenteric arteries are strongly influenced by transmitter release from perivascular sympathetic nerves. They have a major role in the control of peripheral vascular resistance and arterial blood pressure (BP) (410). In the splanchnic circulation, the portal vein and hepatic artery are arranged in parallel and supply blood to the liver for detoxification and metabolism. The specialized pulmonary circulation is characterized by its low vascular pressure and resistance and inherent vasoconstrictor response to hypoxia. These organ-specific regulatory actions interact with myogenic vasoconstriction. Semple and DeWardener (1347) quantified the efficiency of autoregulation by an “autoregulatory index” (AI) defined as the steady-state change in RBF factored by the sustained change in mean RPP (1347). An AI of zero indicates perfect autoregulation, whereas an AI close to 1.0 or above signifies very ineffective RBF autoregulation. Low AI values (⬍0.2), reflecting effective steady-state RBF autoregulation, have been reported in the kidneys of anesthetized dogs (90, 269, 745, 763, 799, 953, 954, 1295, 1344, 1531), mice (583, 751, 756, 1360), rabbits (80, 385, 902, 1141, 1270, 1623), and rats (41, 45, 46, 269, 422, 709, 790, 833, 1105, 1131, 1188, 1270) as well as in conscious rats (270, 423, 668, 1195, 1376) and dogs (69, 97, 421, 510, 511, 753–755, 760, 799, 1172, 1172–1174, 1263, 1618) in both short and prolonged settings (42, 1241). Autoregulation in the kidneys of conscious dogs and anesthetized rats is considerably more complete than in the mesenteric or hindlimb vascular beds (602, 750). The kidney is richly perfused and renal blood flow (RBF) normally accounts for ⬃25% of cardiac output. Autoregulation is a fundamental component of renal function. It integrates intrinsic intrarenal mechanisms that stabilize RBF and glomerular filtration rate (GFR) during changes in renal perfusion pressure (RPP) over a defined range. This requires that the renal vascular resistance (RVR) changes in proportion to the RPP. The cortical radial arteries, and especially the afferent arterioles, are the major preglomerular resistance vessels whose tone mediates most of pressureinduced autoregulation of RBF and GFR. The efferent arterioles usually do not participate in RBF autoregulation, although they may contribute to autoregulation of GFR at low RPP under certain conditions such as low-salt diet and activation of the renin-angiotensin system (RAS) with high angiotensin II (ANG II) levels. Renal autoregulation is achieved primarily by a unique orchestrated action of two major mechanisms: the myogenic response and the macula densa tubuloglomerular feedback (MD-TGF) response. Together they adjust the tone of the preglomerular VSMCs to buffer the challenge of a constantly changing RPP, whether spontaneous or induced. The fast myogenic response relates the tone of the afferent arterioles to their intraluminal pressure by intrinsic adjustments in the tension of VSMCs. The more delayed MD-TGF response is activated by tubular electrolyte delivery and reabsorption primarily by a luminal Na⫹/K⫹/2Cl⫺ type 2 (NKCC2) cotransporter and a Na⫹/H⫹ exchanger on the MD cells at the end of the thick ascending limb (TAL) of Henle’s loop to elaborate positive and negative modulators of vasoconstriction of the adjacent afferent arteriole. The ensuing collective response entails extensive, bidirectional, and time-dependent interactions between these two mechanisms. Renal autoregulation is intrinsic to the kidney and thus largely independent of extrinsic nerves or circulating hormones (428, 998, 1053, 1344, 1345, 1360, 1465, 1578). The myogenic phenomenon was first described for large conduit arteries by Bayliss et al. (76) more than a century ago as part of his investigation of reactive hyperemia in the kidney. Renal autoregulation to acute changes in RPP was characterized originally in rabbits by Forster and Maes in 1947 (428) and in dogs by Selkurt et al. in 1949 (1345). The renal myogenic response was initially described by Gilmore et al. (473) in 1980 in hamster preglomerular arterioles. The unique renal MD-TGF occurs within the juxtaglomerular apparatus (JGA) that is composed of the MD cells at the junction of the TAL of the loop of Henle and the distal tubule, the granular renin-containing cells in the distal afferent arteriole and the extraglomerular mesangium that is interspersed between, or surrounds, these cells. The anatomy of different cell types in the JGA is shown in FIGURE 1. The concept of the JGA coordinating tubular and glomerular functions was introduced originally by Goormaghtigh in 1937 (487), with subsequent hypotheses by Guyton, Harsing, and Thurau in the early 1960s (533, 581, 1465, 1466). The specific contribution of MD-TGF to renal autoregulation of GFR was demonstrated initially by the laboratories of Navar and Schnermann in the 1970s (1022, 1064, 1070, 1190, 1191, 1317). 406 A myogenic response is observed in the renal, cerebral, coronary, pulmonary, mesenteric, and skeletal muscle vasculature (300 –302, 703, 739, 1249). This response limits vascular wall strain by reciprocal adjustments in vessel radius during changes in the transmural pressure gradient. This assumes that the wall thickness remains the same, as described by the LaPlace relationship. Mechanotransduction in VSMCs to increased PP is intrinsic to VSMC and entails stretch or distortion of the plasma membrane coupled to mechanisms that decrease membrane conductance, thereby depolarizing the resting membrane potential (EM) from approximately ⫺40 mV to activate voltage-operated Ca2⫹ channels (VOCCs). The ensuing increase in Ca2⫹ entry el- Physiol Rev • VOL 95 • APRIL 2015 • www.prv.org CARLSTRÖM ET AL. PSF Grease block Efferent arteriole Glomerulus PGC Granular renincontaining cells Afferent arteriole Blood Connecting tubule Extraglomerular mesangial cells Macula densa cells Distal tubule Proximal convoluted tubule Thick ascending limb FIGURE 1. Schematic diagram illustrating the anatomy of the juxtaglomerular apparatus (JGA) showing cell types involved in renal autoregulation, i.e., myogenic response and macula densa (MD)-mediated tubuloglomerular feedback (MD-TGF). MD cells are located at the junction of the thick ascending limb of Henle’s loop and the distal convoluted tubule. Their basolateral membrane contacts some extraglomerular mesangial cells, which in turn are contiguous with vascular smooth muscle cells and renin-containing juxtaglomerular granular cells at the end of the afferent arteriole. Endothelial nitric oxide synthase (eNOS) is expressed primarily in the endothelium of the afferent arteriole and neuronal NOS (nNOS) in the MD cells. MD-TGF can be assessed conveniently by measuring proximal stop-flow pressure (PSF) upstream of a grease block in the nephron as an index of glomerular capillary pressure (PGC) during variations in fluid delivery to MD cells by perfusion from another micropipette (not shown) placed downstream from the block. Recent in vitro and in vivo PSF measurements have demonstrated functional implications of a connecting tubule glomerular feedback (CT-GF) in regulating afferent arteriolar vasomotor tone. evates cytosolic Ca2⫹ concentration ([Ca2⫹]i) that subsequently mobilizes Ca2⫹ stored in the sarcoplasmic reticulum (a process termed Ca2⫹-induced Ca2⫹ release, CICR), combined with signaling pathways that enhance the sensitivity of myosin to a given [Ca2⫹]i (i.e., Ca2⫹ sensitivity). These coordinated actions activate myosin light chain kinase (MLCK) and/or inhibit myosin light chain phosphatase (MLCP) to promote contraction of VSMCs (1328, 1392). This sequence of events is summarized in FIGURE 8. The relative importance of Ca2⫹ influx, release, and sensitization varies considerably among published studies. The MD-TGF is sensitive to renal metabolism. It relates the GFR to the NaCl delivery to and reabsorption by MD cells. MD NaCl delivery, in turn, is related to the filtered load of NaCl and reabsorption in nephron segments upstream of the MD, all of which therefore can affect MD-TGF. Accordingly, MD-TGF integrates tubular and vascular function and is thus a unique renal mechanism of autoregulation. Activated MD cells generate signaling molecule(s) such as ATP and/or its breakdown product adenosine that traverse the interstitial space and extraglomerular mesangial cells, likely involving connexins (Cxs), to stimulate Ca2⫹ signaling and contract the afferent arteriole. The ATP and aden- osine responses are primarily mediated by purinergic P2X receptors and adenosine A1 receptors, respectively, on VSMC and possibly mesangial cells. Mesangial cells may contribute to MD-TGF by providing an extraglomerular conduit pathway, but these cells lack the expression of contractile ␣-smooth muscle actin in vivo that is present in vitro in cell culture, and therefore probably are not contractile in life (341, 378, 796). Because of difficulty in isolating fresh mesangial cells from glomeruli, they are usually studied in culture to obtain a homogeneous population (3, 607, 1615). However, it is recognized that phenotypic changes take place as a function of passage number, for example, in the relative amounts of contractile proteins. VSMC in culture can undergo phenotypic switching from a nondividing contractile phenotype to a proliferative dedifferentiated phenotype characterized by reduced mRNA and protein levels of VSMC markers (1142). Thus results for contraction mechanisms using cultured mesangial cells should be interpreted with caution. Although the general operation of MD-TGF in renal autoregulation is appreciated, the understanding of its fine details is still evolving. It is generally accepted that an increase Physiol Rev • VOL 95 • APRIL 2015 • www.prv.org 407 RENAL AUTOREGULATORY MECHANISMS in RPP initially increases single-nephron glomerular filtration rate (SNGFR) and might reduce proximal tubule reabsorption, thereby increasing NaCl delivery from Henle’s loop to the MD (FIGURE 1). Increased NaCl reabsorption by MD cells generates a signal that mediates contraction of the afferent arteriole. Thus MD-TGF functions as a negativefeedback circuit that increases preglomerular vascular resistance to restrain single-nephron glomerular blood flow (SNGBF) and glomerular capillary hydraulic pressure (PGC), thereby stabilizing GFR and RBF during changes in RPP. The MD-TGF system is sensitive to factors that regulate tubular reabsorption. Therefore, MD-TGF relates renal autoregulation to extracellular fluid volume (ECV) homeostasis. MD-TGF is sensitive to tubular metabolism because it couples epithelial cell salt transport to preglomerular vascular tone utilizing metabolic intermediates such as ATP and adenosine as well as other tubular and vascular metabolites or signaling molecules such as nitric oxide (NO), superoxide anion (O2⫺), prostaglandin endoperoxide (PGH2), thromboxane A2 (TxA2), carbon monoxide (CO), and prostaglandin E2 (PGE2) (FIGURE 6). Ultimately, both myogenic and MD-TGF mechanisms impact regulation of RBF and GFR, primarily by varying the degree of afferent arteriolar vasoconstriction (51, 703, 1436). Their individual contributions, as well as their interactions, determine the overall autoregulatory efficacy. This review focuses on mechanisms responsible for renal autoregulation in health and its role and regulation in several models of hypertension, chronic kidney disease (CKD) and diabetes mellitus (DM). GFR normally is autoregulated with RBF since both respond in parallel to induced changes in preglomerular resistance. However, GFR is determined not only by PGC but also by SNGBF and by the capillary ultrafiltration coefficient (KUF). Selective changes in efferent arteriolar resistance, for example, during inhibition of the RAS at low RPP, can dissociate autoregulation of GFR from RBF (550, 1261). This review concentrates on the changes in preglomerular vascular resistance responsible for regulation of RBF and glomerular perfusion. The reader is referred to recent reviews of renal autoregulation (286, 748, 933, 934) and of mathematical modeling of renal autoregulatory mechanisms (179, 228, 395, 541, 628, 805, 877, 973, 1023, 1350, 1614). B. Functional Significance Renal autoregulation subserves four important functions. First, it sets a basal level of vasomotor tone upon which neurohumoral agents, paracrine/autocrine substances, and metabolic factors adjust RVR (48, 1065, 1071). Second, it buffers the transmission of RPP to the sensitive glomerular capillaries and parenchyma, thereby preventing intrarenal barotrauma during systemic arterial hypertension (108). Impaired renal autoregulation contributes to progressive pressure-sensitive glomerular and interstitial injury in many 408 models of hypertension, CKD, and DM as described in detail in section VII. Third, it isolates the subtle tubular transport processes and sodium (Na⫹) excretion from abrupt changes in GFR that would induce large fluctuations in tubular fluid flow and peritubular physical forces (532). Fourth, a somewhat less complete regulation of medullary blood flow during changes in RPP has been proposed as a mechanism mediating pressure-natriuresis and may be important in maintaining salt and water balance and controlling BP, as discussed in section IIA2. Unfortunately, there are only limited studies of renal autoregulation in normal human subjects or those with hypertension. A study of African-American hypertensive patients reported higher values for GFR and RBF than matched Caucasians (FIGURE 2, A–C) (826). A pressor infusion of NE increased the GFR only in the African-Americans, which led the authors to propose that African-American hypertensives had impaired renal autoregulation, although the RBF was maintained independent of RPP in both groups. In another study of hypertensive patients, the systolic BP was reduced incrementally by infusion of the vasodilator sodium nitroprusside (FIGURE 2, D AND E) that releases NO (20). The GFR and effective renal plasma flow were wellmaintained across the range of systolic BPs of 170 to 130 mmHg in patients with moderate hypertension, but both fell progressively in the range of 200 –135 mmHg in those with severe hypertension. This demonstrated preservation of renal autoregulation in patients with moderate hypertension, but a breakdown in autoregulation in those with severe hypertension. Thus renal autoregulation can provide a remarkably stable GFR and RBF during changes in RPP in human subjects with hypertension. The observation of impaired autoregulation in hypertensive African-Americans and those with severe hypertension is consistent with the hypothesis that renal autoregulation normally functions to protect the kidney from barotrauma. Its failure in these two populations increases risk for the development of nephrosclerosis and hypertensive CKD. II. MAJOR MECHANISMS AND METHODS USED TO STUDY RENAL AUTOREGULATION Autoregulation has been characterized in different preparations of the renal circulation ranging from in vivo measurements of RPP-induced changes in whole kidney RBF, GFR, and single-nephron hemodynamics during induced changes in RPP to direct assessment of pressureinduced contraction of isolated, perfused afferent arterioles, or flow-induced MD-TGF responses in isolated JGAs. Examples of autoregulatory myogenic responses from six commonly used preparations are described below and are highlighted in FIGURE 3, A–F. Physiol Rev • VOL 95 • APRIL 2015 • www.prv.org CARLSTRÖM ET AL. 160 150 * 140 130 White Black ERBF 1,500 ** 1,400 1,300 1,200 1,100 1,000 900 C GFR 200 P<0.001 180 160 140 * 120 100 White Black White Black D GFR 150 130 110 90 70 ** 50 130 140 150 160 170 180 190 200 E CHipurran (ml · min-1 ·1.73 m-2) P<0.001 B CCr (ml · min-1 ·1.73 m-2) SBP (mmHg) 170 P<0.001 GFR (ml · min-1 ·1.73 m-2) BP 180 ERBF (ml · min-1 ·1.73 m-2) A ERPF 450 400 350 300 250 ** 200 150 130 140 150 160 170 180 190 200 FIGURE 2. Studies of renal autoregulation in hypertensive human subjects. Blood pressure (BP) was determined at regular intervals, and the glomerular filtration rate (GFR) was measured from the clearance of inulin or creatinine and the effective renal plasma flow (ERPF) from the clearance of hippuran and used to calculate the effective renal blood flow (ERBF). A–C: data were compared in 33 Caucasian (white) and 29 African-Americans (black) matched patients with essential hypertension in the basal state (open boxes) and during infusion of norepinephrine (0.05 g·kg⫺1·min⫺1 for 30 min) to increase BP (closed boxes). The ERBF was higher in African-American than in Caucasian subjects but did not change during a short-term elevation of BP by norepinephrine in either patient group. However, the GFR also was higher in the African-Americans and increased further with the rise in BP, implying a selective defect in GFR autoregulation. Compared with Caucasians in the basal state: *P ⬍ 0.05; **P ⬍ 0.01. [Data from Kotchen et al. (826).] D and E: data were compared in patients with moderate hypertension (average: 170/108 mmHg; n ⫽ 10, open circles) and severe hypertension (average: 198/127 mmHg; n ⫽ 10; closed circles), in the basal state and during graded infusion of sodium nitroprusside (0.2–12 g·kg⫺1·min⫺1) to provide incremental reductions in systolic BP (SBP). After acute BP normalization, the GFR and the ERPF were reduced significantly in patients with severe hypertension (⫺44.7 ⫾ 6.8 and ⫺41.6 ⫾ 8.3%, respectively), but did not change in those with moderate hypertension, implying that renal autoregulation was preserved in subjects with essential hypertension, but was impaired or absent in those with severe hypertension. **P ⬍ 0.01 vs. recordings in the basal state with high BP. [Data from Almeida et al. (20).] A. In Vivo Studies Using Whole Kidney Preparations 1. Steady-state autoregulation A) STEADY-STATE AUTOREGULATION OF WHOLE KIDNEY BLOOD FLOW. The BP in animal studies can be increased by carotid arterial occlusion to elicit baroreceptor activation of the sympathetic nervous system. Graded reductions of the elevated RPP by constriction of the suprarenal aorta for 1–2 min can provide a range of RPPs above, at, and below the ambient RPP (FIGURE 3A) (41, 46). Ideally, this should be achieved by coupling an aortic or renal artery constrictor to a servo-controlled pressure sensor to provide more stable levels of RPP (46, 376, 549, 610). The kidneys should be denervated to minimize the influence of changes in renal sympathetic nerve activity. Alternatively, the RPP can be increased by constriction of the abdominal aorta below the renal vessels. This can be combined with occlusion of the superior mesenteric and celiac arteries to increase the RPP, with a further rise in PP achieved by infusion of a cocktail of vasoconstrictors such as NE and arginine vasopressin (AVP) (1247). A limitation of this method is that the infused hormones themselves affect renal vascular tone and may modulate autoregulation. Another approach has been to study an isolated, pump-perfused kidney of dogs (1106, 1108, 1110, 1112, 1120) or rats (1285, 1323). The oxygenated perfusate usually contains albumin or a colloid, but has a low hematocrit to minimize erythrocyte damage by the perfusion pump. The reduced red cell mass limits the vessel wall shear stress and NO release, but this is offset by some degree by less oxyhemoglobin-mediated oxidation of NO, whose influence may thereby become more pronounced. Perfusion is usually at a fixed mean RPP rather than a pulsatile pressure. Limitations are that the isolated kidneys become edematous, fail to reabsorb salt normally, and have limited autoregulatory efficiency (870, 871, 1332, 1468). B) STEADY-STATE AUTOREGULATION IN DIFFERENT REGIONS OF THE KIDNEY. Most current studies of renal cortical blood flow use a fiberoptic probe connected to a laser Doppler flowmeter to estimate the erythrocyte (RBC) velocity in a small tissue area (⬃1 mm3). Highly efficient autoregulation was reported in the outer-, mid- and deep-cortical regions of rat kidneys during hydropenia (990, 1195, 1248, 1252) and during acute expansion of ECV (990, 1248). Autoregulation of cortical blood flow generally parallels that of the whole kidney. Measurement of medullary blood flow is considerably more complex (283, 1256, 1423). For example, measurements of average RBC velocity by Doppler methods applied to the medulla do not differentiate between descending and ascending flow in vasa recta capillaries. Volume flux depends on the number of vessels being perfused and the fluid reabsorbed by the tubules. This may explain in part why the efficiency of medullary autoregulation has been more vari- Physiol Rev • VOL 95 • APRIL 2015 • www.prv.org 409 RENAL AUTOREGULATORY MECHANISMS able among studies and species and in some depends on the state of hydration. Both cortical and medullary blood flow in nondiuretic rats are autoregulated efficiently between RPP of 90 –160 mmHg (267, 288, 990, 1248, 1417). An early study that measured the RBC velocity in individual vessels on the surface of the renal papilla reported excellent autoregulation during hydropenia and volume expansion (267). However, other studies using similar methodology 110 RBF (% basal) 100 Rat dynamic autoregulation: time domain RVR (mmHg) B Rat steady state autoregulation 120 RVR (% of perfect AR) A reported weak autoregulation of RBC velocity in descending and ascending vasa recta of euvolemic rats, with AI values of 0.5– 0.8 (391). Many subsequent studies in rats that have used the laser Doppler technique to measure RBC velocity in multiple capillaries in a fixed volume of tissue reported highly efficient autoregulation of blood flow in the outer and inner portions of the renal medulla in hydropenia (990), but this became substantially less efficient during ECV expansion (277, 411, 717, 990, 1243, 1251, 1252, 1254, 1257, 1476). However, another study reported only moderate medullary autoregulation (AI ⫽ 0.4) in hydro- 100 105 AI < 0.1 90 80 70 AI < 0.05 60 50 40 45 55 65 75 85 95 50 TGF: ~35% of total 0 MR: ~50% of total -50 0 105 115 125 135 145 60 80 100 120 Time after release (sec) Rat dynamic autoregulation: frequency domain D Rat juxtamedullary preparation 140 2.0 AI = 0.32 MR - 65% CCB 120 Diameter (% of control) Admittance gain 40 20 Renal PP (mmHg) C 3rd 90 1.0 TGF - 35% 0.5 MR - 65% 100 TGF block TGF - 35% 80 Control 60 0.25 0.01 0.1 1 60 E 100 80 120 140 Renal PP (mmHg) Frequency (Hz) Rat hydronephrotic kidney F Mouse isolated perfused afferent arteriole 0 -5 Active wall tensions (dynes/cm) Afferent arteriolar diameter (% change) 400 -10 -15 -20 300 200 100 -25 0 80 100 120 140 160 Renal PP (mmHg) 410 180 40 60 80 100 Arteriolar PP (mmHg) Physiol Rev • VOL 95 • APRIL 2015 • www.prv.org 120 140 CARLSTRÖM ET AL. penic rats with little change (AI ⫽ 0.5) during ECV expansion (1248). An important study concluded that ECV expansion increases renal medullary blood flow by increasing the number of capillaries perfused with moving RBCs without a change in their velocity in either ascending or descending vasa recta (415). Further evidence of incomplete medullary blood flow autoregulation came from the observation that the hydrostatic pressure in the vasa recta capillaries was autoregulated less completely than in the cortical peritubular capillaries (1248). The renal medulla is perfused from cortical efferent arterioles. Therefore, the finding that medullary perfusion in the rat can be poorly autoregulated, especially during ECV expansion, while deep cortical blood flow is well autoregulated located the postglomerular vasculature as the site of impaired control of medullary blood flow (990). Highly efficient autoregulation of both cortical and medullary blood flow that is independent of the ECV is observed in whole kidney studies in the dog (950, 951, 954, 960, 961) and in the rabbit (381). On the other hand, one study reports that blood flow to the exposed papilla of the dog was poorly autoregulated (AI ⫽ 0.82) compared with whole kidney RBF (AI ⫽ 0.33) (1423), but this may have been a consequence of the impaired function of the exposed papilla. In summary, in contrast to cortical blood flow, the degree of autoregulation of medullary blood flow in the rat is controversial. Although most studies have reported efficient autoregulation of medullary blood flow during hydropenia, this appears to become less efficient during ECV expansion, at least in the rat (277, 990, 1243, 1251, 1252, 1254, 1257). However, most studies in dogs and rabbits report efficient medullary autoregulation during both hydropenia and ECV expansion. Autoregulatory responses of the preglomerular vasculature can be assessed directly in the in vitro rat juxtamedullary nephrovascular (JMN) preparation. As discussed in section IIB3, these studies generally have reported efficient autoregulation, involving both myogenic and MDTGF components. 2. Pressure-natriuresis The reports of highly efficient overall renal autoregulation, and often also of intrarenal distribution of perfusion in the dog, rabbit, and nondiuretic rat, pose a conundrum for the understanding of how the kidney regulates NaCl and fluid excretion to maintain long-term BP homeostasis. Renal Na⫹ excretion changes acutely with RPP, even in studies in which the intrarenal mechanisms maintain a stable blood flow in the renal cortex and medulla. Further uncertainty derives from the recognition that steady-state measurements of proximal reabsorption and peritubular capillary fluid uptake do not permit firm conclusions regarding causality (1490). Thus the rate of reabsorption must correlate with the effective reabsorptive pressure determined by Starling physical forces and the peritubular capillary reabsorptive coefficient. Moreover, the events that initiate the response may be transient yet the GFR and the filtered Na⫹ load remain constant during changes in PP. This indicates that the changes in Na⫹ excretion must be due to variations in tubular Na⫹ reabsorption. The key question is: What intrarenal mechanism(s) can mediate pressure-natriuresis when the pressures in the tubules and vasculature downstream of the afferent arteriole remain stable because of highly efficient autoregulation? A step change in RPP usually yields almost no change in steady-state whole kidney RBF, GFR, efferent arteriolar blood flow (956, 1636), or intrarenal hydrostatic pressures FIGURE 3. Data redrawn from 6 published studies to illustrate different methods for assessing renal autoregulation or the contributions of myogenic and macula densa-tubuloglomerular feedback (MD-TGF) mechanisms. A: measurements of renal blood flow (RBF) by electromagnetic flow meter recordings in anesthetized rats as a function of induced changes in renal perfusion pressure (PP). The autoregulatory index (AI) ⬍0.05 implies almost perfect independence of RBF from renal PP across the range 105–145 mmHg (41, 46). B: study of dynamic renal autoregulation in the time domain in anesthetized rats (749). The renal PP had been reduced previously by a suprarenal aortic clamp to ⬃95 mmHg. Abrupt release of the aortic clamp led to a rapid increase in renal PP (RPP) to ⬃110 mmHg, which was maintained throughout the 2 min. The renal vascular resistance (RVR) fell initially, but this was followed rapidly by a 3-component rise. Over the first 5 s, the increase restored ⬃50% of the final RVR response to the level of perfect autoregulation. This was attributed to the myogenic response (MR) and was followed over 10 –20 s by a further rise in RVR restoring ⬃35% of the RVR. This was attributed to macula densa-tubuloglomerular feedback (MD-TGF) response and was followed by a third, slower component contributing ⬃15% of the final RVR response. C: study of dynamic renal autoregulation in the frequency domain by transfer function analysis in rats of measurements of spontaneous changes of renal PP, measured by a pressure transducer in the aorta, and RBF measured by ultrasonic blood flow meter (297). The tracing shows normalized admittance gain (a relative autoregulatory index) as a function of frequency of spontaneous fluctuations in renal PP. Values at or above unity imply very weak to no active autoregulation. Between 0.3 and 0.1 Hz oscillations in renal PP, the admittance gain fell steeply, corresponding to the myogenic response (MR). There was a further sharp fall in admittance gain between 0.03 and 0.01 Hz, corresponding to the delayed MD-TGF mechanism. D: measurements of the steady-state diameter of the afferent arteriole of the rat juxtamedullary nephron (JMN) preparation during induced changes in renal PP (1020). An increase in renal PP, from 60 to 140 mmHg, reduced arteriolar diameter by ⬃25–30% (control). A progressive autoregulatory reduction in diameter during increased renal PP was attenuated by blockade of MD-TGF (by furosemide) and was totally abolished by the CCB nimodipine that eliminated all active tone. E: directly visualized changes in diameter of the rat afferent arteriole in the hydronephrotic kidney (HNK) preparation lacking MD-TGF (586). An increase in renal PP from 80 to 180 mmHg reduced the arteriolar diameter (i.e., myogenic response). F: development of active wall tension as a function of arteriolar PP in a mouse isolated perfused afferent arteriole (861). The slope of this line defined the myogenic response over the PP range tested, and its intercept on the x-axis defined the threshold. Physiol Rev • VOL 95 • APRIL 2015 • www.prv.org 411 RENAL AUTOREGULATORY MECHANISMS (42, 276, 1167, 1188, 1241). Spontaneously hypertensive rats (SHR) had excellent RBF and GFR autoregulation (41, 43, 465, 672, 706, 1247) and stable mean PGC and peritubular capillary pressures in their superficial cortical nephrons, which were attributed to proportionate adjustments in preglomerular vascular resistance with RPP (42, 58, 394, 1064, 1247, 1564, 1567). Nevertheless, these SHR displayed a pressure-natriuresis, albeit dampened compared with normotensive Wistar-Kyoto (WKY) rats that also autoregulated efficiently (42, 276, 412, 808). Early micropuncture studies reported that steady-state proximal tubular reabsorption and fluid delivery to the distal tubule of superficial cortical nephrons were maintained during an acute change in RPP, implying minimal change in input signal to modulate MD-TGF (42, 43, 342, 535, 901, 1067) and minimal change in fractional Na⫹ delivery to the late distal tubule in the superficial cortex (844). One in vivo microperfusion study in rats reported that ⬃25 mmHg increase in RPP inhibited proximal Na⫹ reabsorption by almost 40%, but a similar decrease in RPP did not affect Na⫹ reabsorption (798). However, previous free-flow micropuncture studies from the same laboratory but in a different strain of rat reported unchanged proximal tubular Na⫹ reabsorption of superficial nephrons, but decreased reabsorption of deep nephrons, in response to increased RPP (535). Nevertheless, other investigators have reported that increased RPP inhibited Na⫹ reabsorption by the proximal tubule of surface nephrons (248, 1245). Marsh, Roman, and colleagues reported that increased RPP reduced proximal tubular reabsorption rapidly and increased fluid delivery to Henle’s loop despite highly efficient autoregulation of RBF and GFR (249, 1248). Reduced absolute and fractional proximal tubular reabsorption after increased RPP has been confirmed in other studies in rats, but autoregulatory efficiency was not verified (814, 995, 996). A more consistent finding has been that increased RPP inhibits Na⫹ reabsorption by proximal tubules of deep nephrons (to a greater extent than superficial nephrons) (535, 1245). This likely reflects pressure-induced reductions in Na⫹ and fluid reabsorption in straight descending portions of the late proximal tubule. In studies in which an increased PP inhibited proximal tubular reabsorption, the anticipated increase in Cl⫺ reabsorption by Henle’s loop in response to the increased tubular fluid delivery was lacking (248, 249, 1245). Studies of fractional Li⫹ excretion as an index of fractional proximal Na⫹ reabsorption concluded that increased RPP reduces fractional Na⫹ reabsorption in the proximal (536, 736) and distal tubules (736). Increased RPP also reduced Na⫹ reabsorption by the nephron segments distal to Henle’s loop, since pharmacological blockade of distal tubular Na⫹ reabsorption markedly attenuated the pressure-natriuresis re- 412 sponse (956). It is uncertain whether the medullary collecting duct (CD) also participates in these changes in reabsorption with PP (1245, 1394). Thus changes in Na⫹ reabsorption by Henle’s loop and the distal nephron can participate in pressure-natriuresis. The preglomerular vasculature of the juxtamedullary cortex maintained an appropriate myogenic and MD-TGF response that stabilized SNGFR in deep nephrons during increases in RPP (535, 1248). Similarly, the afferent arterioles of the JMN preparation responded appropriately to changes in PP (see below). Nevertheless, the hydrostatic pressure in vasa recta capillaries and the cortical renal interstitial hydrostatic pressure (RIHP), which in turn is thought to reduce tubular reabsorption, both increase with PP. An increase in RPP increased RIHP by only a few mmHg in nondiuretic rats, but to a larger extent after volume expansion (459, 486, 524, 790, 792). Indeed, both the cortical and medullary RIHP increased after volume expansion. Decapsulation of the kidney reduced the changes in RIHP with PP and attenuated pressure-natriuresis (459, 537, 791, 792). Changes in RIHP independent of a change in RPP are commonly observed, but may be more associative than causal since changes in hydrostatic pressure are less influential on transepithelial transport than changes in osmotic pressure (493, 539). Since the efferent arterioles from the deep cortical nephrons supply the medullary vasa recta capillaries, any attenuation of autoregulation by the preglomerular vasculature of JMN would likely transmit some of the changed RPP into the vasa recta capillaries and the medullary interstitium. Consistent with this prediction, the hydrostatic pressure in vasa recta capillaries was autoregulated less completely than in renal cortical capillaries (1248). Three factors have been identified that could increase the medullary RIHP during increased PP in diuretic states. First, the descending vasa recta provide a fourfold greater flow resistance than the ascending vasa recta perhaps because of the fewer descending capillaries that results in a markedly lesser vascular cross-sectional area (71, 1149, 1643). Second, the hydrostatic pressure in the medulla is determined primarily by the outflow resistance of the CDs (972). Third, the descending vasa recta are surrounded by contractile pericytes that can generate a myogenic response (908). Large-diameter human outer medullary descending vasa recta contracted to increased luminal pressure, whereas smaller vasa recta did not (1348). The medullary RIHP should determine the cortical RIHP since the kidney is an encapsulated organ (277, 790, 1140). Thus this sequence of events provides a mechanism whereby cortical RIHP and proximal fluid reabsorption could vary with RPP despite no change in PGC. Indeed, Na⫹ Physiol Rev • VOL 95 • APRIL 2015 • www.prv.org CARLSTRÖM ET AL. reabsorption was closely related to even very small changes in cortical RIHP (1607). Paracrine agents including NO, prostaglandins (PGs), 20HETE, and ATP have been proposed to modulate pressurenatriuresis. The slope of the natriuretic response to increased RPP is attenuated by inhibition of NOS (385, 411, 411, 429, 524, 666, 736, 946, 947, 955, 957, 959, 962, 963, 1277, 1476), or COX (190, 486, 536, 538, 797, 1074, 1258), or cytochrome P-450 4A to reduce 20-HETE generation (340, 1612). Moreover, renal interstitial ATP concentration paralleled RPP and regulated both MD-TGF (1090, 1091) and Na⫹ transport via epithelial sodium channels (ENaC) in the CDs (1475). Activation of AT1 receptors can inhibit pressure-natriuresis (317, 321, 513, 515, 547, 569, 946, 995–997, 1074, 1258, 1516, 1549), whereas activation of AT2 receptors enhanced pressure-natriuresis in some (514, 516) but not all studies (911, 918). Inhibition of ACE-dependent ANG II generation enhanced pressure natriuresis (1074), whereas chronic ANG II infusion increased BP and suppressed pressure natriuresis (1549). Moreover, clamping systemic ANG II levels during acute hypertension blunted the magnitude of the pressure-natriuresis response (884). Increased RAS activity, AVP (433), renal sympathetic nerve activity (RSNA) (328, 337, 376, 775, 806, 1130, 1382), and O2⫺ or hydrogen peroxide (H2O2), especially in the renal medulla (273, 734, 1026, 1099), may also inhibit pressure natriuresis. Despite this evidence of the importance of weak autoregulation of the medullary blood flow during volume expansion for initiating pressure-natriuresis, there remain two divergent schools of thought concerning the major cause of pressure-natriuresis in an autoregulating kidney with an unchanged overall filtered load of Na⫹ and increased RIHP. The major difference between them concerns the mediating factor(s) and the relative importance of a primary change in renal cortical NO generation or in medullary blood flow (FIGURE 4, A AND B). Majid and Navar demonstrated strong correlations between intrarenal levels of NO, RIHP, and pressure-natriuresis in dog kidneys exhibiting highly efficient autoregulation of cortical and medullary blood flow (FIGURE 4A) (955, 957–959, 961, 963). During changes in RPP, the Na⫹ excretion, renal cortical and medullary NO production, and RIHP changed in parallel. Inhibition of renal NOS suppressed pressure-natriuresis in autoregulating kidneys without changing the filtered Na⫹ load (537). However, although administration of a NO donor during NOS inhibition increased basal Na⫹ excretion and RIHP, it failed to normalize the slope of the pressure-natriuresis or RIHP relationships. This suggests that a combination of pressuredependent changes in renal NO production and RIHP are required to mediate pressure-natriuresis. A Renal PP Excellent corticol and medullary autoregulation Vessel wall shear stress Arteriolar vasoconstriction Vascular NO production Interstitial hydrostatic pressure No change in corticol or medullary blood flow or GFR Tubular Na+ reabsorption Renal Na+ excretion B Renal PP Incomplete medullary autoregulation Excellent corticol autoregulation Medullary and papillary blood flow No change in corticol blood flow or GFR Medullary interstitial hydrostatic pressure Corticol interstitial hydrostatic pressure Na+ reabsorption in the proximal tubule Renal Na+ excretion FIGURE 4. Flow diagrams summarizing current hypotheses for interactions between renal autoregulation and pressure natriuresis in response to increased renal perfusion pressure (PP) with excellent autoregulation of cortical blood flow with and without efficient autoregulation of medullary blood flow. A: proposed intrarenal events causing pressure-natriuresis when both renal cortical and medullary blood flows are excellently autoregulated. B: proposed intrarenal events causing pressure-natriuresis when renal cortical blood flow is excellently autoregulated but autoregulation of medullary blood flow is less complete. GFR, glomerular filtration rate; RBF, renal blood flow; NO, nitric oxide; ROS, reactive oxygen species. For details, see text (sect. IIA2). Endothelial NO is released in response to increased vessel wall shear stress. The constriction of the renal resistance vessels during increased PP would increase wall stress without changing blood flow (171, 747, 1039, 1184, 1416, 1425) and could thereby provide a signal for NO release in fully autoregulating kidneys. NO inhibits Na⫹ reabsorption in the TAL and the CDs (379, 460, 608, 1185, 1419), whereas the actions in the proximal tubule are more controversial, with reports of NO inhibiting (1419) or stimu- Physiol Rev • VOL 95 • APRIL 2015 • www.prv.org 413 RENAL AUTOREGULATORY MECHANISMS lating (1560) Na⫹ reabsorption. However, studies in mice with deletion of specific NOS isoforms concluded that NO derived from nNOS or eNOS stimulated Na⫹ reabsorption and H⫹ secretion by the proximal tubule (1561). Thus it is unlikely that the proximal tubule is the primary site of NO-mediated inhibition of tubular reabsorption during increased PP. As mentioned previously, blockade of distal nephron Na⫹ transport markedly attenuated pressure-natriuresis (956), demonstrating the importance of active transport inhibition in mediating this response. The TAL and the CD are rich sources of NO and sites at which NO reduced Na⫹ reabsorption (1129, 1148, 1304, 1627). Indeed, medullary NO preferentially inhibited Na⫹ reabsorption in the CDs (1304), which participated in inhibition of fractional NaCl reabsorption during increased PP (1394). Carey and colleagues have proposed a modified view for the participation of NO in pressure natriuresis via effects on the generation of cGMP and activation of PKG. Increased NOcGMP-PKG signaling modulated pressure-natriuresis during elevated PP by increasing RIHP and inhibiting proximal tubular Na⫹ reabsorption independent of changes in GFR or renal cortical or medullary blood flow (736, 737, 900). Renal cortical interstitial cGMP accumulated in proximal tubule cells where it activated PKG, inhibited tubular Na⫹ reabsorption, thereby increasing Na⫹ excretion independent of measurable hemodynamic changes (736, 737, 1153). This was regional specific since infusion of cGMP into the renal cortex produced a natriuresis, whereas infusion into the renal medulla was ineffective (737). Knox and colleagues presented evidence that increased RPP increases prostaglandin E2 (PGE2) production by COX-1. The PGE2 was believed to derive from endothelial cells of the preglomerular vasculature and, during increased RIHP, to inhibit proximal tubular Na⫹ reabsorption (486, 509, 536, 538, 790, 797, 823, 1207). The mechanism may involve PGE2-induced enhancement of paracellular Na⫹ backflux from the interstitium into the tubular lumen during increased RIHP that would decrease net Na⫹ reabsorption (1207). PGE2 also inhibited Na⫹ reabsorption by the TAL and the CDs (37, 150, 151, 1061). An alternative view of pressure natriuresis is based largely on studies in the rat where there is reportedly less efficient autoregulation of medullary and papillary blood flow during states of relatively high Na⫹ excretion (FIGURE 4B) (277, 1248). Cowley, Mattson, Roman, and colleagues have implicated medullary blood flow as a major determinant of Na⫹ excretion and BP (274, 277). They reported that medullary blood flow and NO were reduced, and ROS production was increased, during states of Na⫹ retention and hypertension (275, 989, 991, 1670), whereas increased medullary blood flow was associated with states of a low BP (273, 274, 1014). NO could have a causal role also in the 414 medulla since its blockade attenuated the increases in renal medullary blood flow and Na⫹ excretion during increased RPP (411). An increased RPP in volume-expanded rats did not change RBF, GFR, or SNGFR in either superficial or deep cortical nephrons (1248). Nevertheless, proximal tubular reabsorption was decreased by 10%, and NaCl delivery to the bend of Henle’s loop of JMN was nearly doubled (1245), thereby likely providing a signal to activate MDTGF. As is summarized in FIGURE 4B, these investigators have demonstrated that RPP increases papillary blood flow and vasa recta capillary hydrostatic pressure that is translated into an increased medullary RIHP which, after transmission to the cortex, reduces proximal tubular Na⫹ reabsorption. Increased Na⫹ delivery to the TAL of Henle’s loop may stimulate production of O2⫺ which can be countered by flow-induced NO production by the vasa recta capillaries. The balance of vasoconstrictor O2⫺ and vasodilator NO in the renal medulla is held to regulate the medullary blood flow by determining the degree of contraction of pericytes in the outer medullary vasa recta, which, in turn, can determine the strength of the pressure-natriuresis (1099). Moreover, the oxygen tension (PO2) of the renal parenchyma could also account for a relatively impaired medullary and papillary autoregulation. The kidneys have a high O2 supply that is necessary to match the demand for tubular O2 consumption related to active tubular Na⫹ reabsorption. The PO2 in the proximal and distal tubules, the efferent arterioles, and the interstitium of the outer cortex of rat kidneys were all similar and average 40 – 45 mmHg, indicating diffusional O2 equilibrium (1582). The PO2 in the glomerulus, measured by a different technique, also averaged 45 mmHg (1331). These values were all lower than PO2 measured in the renal vein (i.e., 55– 65 mmHg), indicating a preglomerular diffusional shunt for O2 between the arterial and venous systems, likely via the arcuate vessels (1605). This shunt stabilized renal cortical PO2 during changes in arterial PO2 (883, 1605) but maintained a low renal parenchymal PO2. The PO2 in the deep cortex and the outer medulla of rat (FIGURE 5) (1260, 1582, 1583) and mouse kidneys (860) was reduced to 30 – 40 mmHg. A study in the rat hydronephrotic kidney (HNK) preparation reported substantial reductions in the myogenic response as the perfusate PO2 was reduced from 80 to 20 mmHg (936). Thus a lower parenchymal PO2 in the medulla might contribute to a lower efficiency of the myogenic response (FIGURE 5) and thereby to the relatively impaired medullary autoregulation. However, studies in dogs reported that hypoxia (PO2 between 40 – 45 mmHg) did not affect autoregulation of RBF or GFR (26), but the diffusional O2 shunt rendered renal PO2 largely independent of arterial PO2, at least in the rat (1605). It remains to be demonstrated that medullary PO2 varies with PP as would be required to modulate pressure natriuresis. Physiol Rev • VOL 95 • APRIL 2015 • www.prv.org CARLSTRÖM ET AL. A B Myogenic responses and PO2 PO2 in rat kidney 0 20 -10 30 40 -20 60 -30 PO2 (mmHg) 0 PO2 (mmHg) Change in arteriolar diameter (%) 10 OC IC 10 20 30 40 50 80 -40 80 120 140 160 180 PO2 in mouse kidney OC OM FIGURE 5. Oxygen tension (PO2) in different renal compartments and its influence on the myogenic response of afferent arterioles. A depicts changes in the diameter of afferent arterioles during increases in renal PP (from 80 to 180 mmHg) in the atubular rat HNK preparation (reflecting myogenic responses) during perfusion with fluids of different PO2 ranging from 20 to 80 mmHg. Note the graded impairment of myogenic responses across the range of PO2 from 80 to 20 mmHg (936). B and C present values for PO2 measured with microelectrodes in the kidney. In B, PO2 values are shown for the outer cortex (OC) and inner cortex (IC) in anesthetized rats (1582). In C, PO2 values are shown for the outer cortex and outer medulla (OM) of anesthetized mice (860). Renal PP (mmHg) In summary, the renal pressure-natriuresis response is important for control of ECV and BP. Although a rise in RIHP and reduction in tubular reabsorption following an increase in RPP are crucial events, the underlying mechanisms are still debated. The contrasting two major hypotheses for how the kidney senses a rise in RPP and generates a pressure-natriuresis, despite excellent autoregulation of the cortical nephrons, are summarized in FIGURE 4, A AND B. Functional abnormalities in disease models will be discussed later in the sections on hypertension, CKD, and DM (see sect. VII). 3. Conventional macula densa tubuloglomerular feedback The anatomical arrangement of different cell types in the JGA that are involved in macula densa (MD)-mediated tubuloglomerular feedback (TGF) is shown schematically in FIGURE 1. MD-TGF relates afferent arteriolar tone to MD cell tubular electrolyte delivery and reabsorption. It can be studied in superficial nephrons of anesthetized animals by microperfusion of Henle’s loop with artificial tubular fluid to vary NaCl reabsorption by the MD segment (48, 1066, 1075, 1305, 1316, 1513). Details about methodologies and approaches to investigate MD-TGF by renal micropuncture have been reviewed (923, 1311, 1513). Afferent arteriolar responses have been assessed from changes in the SNGFR, PGC, or proximal tubular stop-flow pressure (PSF) upstream from an oil or wax block in the proximal tubule. MD-TGF in longer deep cortical nephrons has been assessed in the exposed papilla by perfusing fluid through the TAL of Henle’s loop and measuring glomerular responses at an upstream site of the same nephron (1042, 1043). The afferent arteriole is the effector site, and its terminal segment is the most responsive site for MD-TGF activation (200, 695, 703, 1020). Ambient MD-TGF activity has been assessed from the difference in SNGFR measured from the proximal tubule (which prevents activation of the MD) and the distal tubule with MD function intact. This approach characterizes MD-TGF at zero and ambient, but not at supranormal rates of tubular fluid flow. The role of MD-TGF in autoregulation has been evaluated by measuring vascular function during changes in RPP before and during inhibition of MD-TGF by blocking the flow of tubular fluid to the MD of the test nephron or by blocking NKCC2 activation of MD cells with a loop diuretic. Inhibition of MD function abolished the effects of MD-TGF signaling on afferent arteriolar diameter, PGC, single-nephron blood flow, and GFR of superficial and deep cortical nephrons and attenuated autoregulation of SNGFR during changes in RPP (200, 1019, 1022, 1064, 1075, 1249, 1636). A contemporary approach has been to assess afferent arteriolar or glomerular responses using genetic mouse models that lack MDTGF such as the adenosine A1 receptor knockout (155, 205, 686, 1123, 1312, 1318, 1508) and other transgenic models with impaired MD-TGF signaling (152, 496, 806, 942, 1093). Autoregulation was less complete after inhibition of MD-TGF by genetic deletion of adenosine A1 receptors (583, 751) or P2X1 receptors (686) or Cx40 (756, 1395), all of which impair MD-TGF responses. Glomerular autoregulation has been reported to be less efficient in the absence of MD-TGF in several species, including rats (1019, 1022, 1313, 1444), dogs (351, 760, 1064, 1090, 1091, 1622), and mice (583). For example, the MD-TGF in the rat contributed ⬃50% to autoregulation of SNGFR when the RPP was reduced from 115 to 95 mmHg and 30% when it was reduced from 95 to 80 mmHg (1313). Studies made in dogs indicated that a reduction in RPP from 124 to 94 mmHg in kidneys in which RBF and GFR were efficiently autoregulated led to a 30% reduction in proximally measured SNGFR (when flow to the MD is zero), whereas distally determined SNGFR (when delivery to the MD is intact) was unchanged (1613). Steady-state RBF autoregulation was reduced ⬃50% in nonfiltering dog kidneys (1284). In mice, a 15 mmHg reduction in RPP reduced RBF and GFR by 6 and 12%, respectively, but the adjustments were less complete (with 15 and 40% respective reductions) in mice lacking functional MD-TGF (583). Thus MD-TGF clearly contributes to autoregulation of RBF and GFR. Physiol Rev • VOL 95 • APRIL 2015 • www.prv.org 415 RENAL AUTOREGULATORY MECHANISMS delay was ⬃30 s in the dog kidney (86). This response was completed during the following 15–20 s in rodent kidneys (FIGURE 3B) (296, 296, 629, 748, 749) or 20 – 40 s in dog kidneys (748, 753). This delay sets up MD-TGF-based oscillations in proximal tubular pressure (PPT) at ⬃0.035 Hz (626, 893, 1512) which are in phase with fluctuations in early distal tubule fluid flow and [Cl⫺] that are signals for activation of MD-TGF. All of these cycled at 25–35 s (35 mHz) (627). Measurement of these oscillations has been used to assess endogenous MD-TGF activity during changes in RPP. Afferent arteriolar autoregulatory responses in the JMN preparation were reduced 25–50% by elimination of MDTGF by papillectomy or furosemide in the rat (686, 1028, 1364, 1437) or mouse (686, 687). Papillectomy or tubular oil block inhibited afferent arteriolar responses similar to that of furosemide, indicating that the primary effect of furosemide in this preparation was on MD transport and not directly on the afferent arteriole. However, furosemide also attenuated the autoregulation of GFR when measured in proximal tubules in the JMN preparation, which prevents delivery of tubular fluid to the MD and negates the MD-TGF (200, 1022). This might represent nephronnephron interactions whereby inhibition of MD-TGF in neighboring nephrons attenuates the myogenic response in the test nephron. Other studies reported that furosemide abolished pressure-dependent autoregulation of afferent arterioles in the JMN preparation (485). Furosemide also inhibited MD-TGF-mediated constriction of the afferent arteriole in the doubly perfused, isolated JGA (695). These studies reinforce the conclusion that MD-TGF contributes importantly to renal autoregulation. A number of vasoactive factors may link MD transport of electrolytes by the NKCC2 cotransporter, ROMK channel, Na⫹/H⫹ exchanger, and Na⫹-K⫹-ATPase to the release of vasoconstrictors including ATP, adenosine, PGH2, TxA2, and O2⫺ and vasodilators such as NO, PGE2, and carbon monoxide (CO) to modulate afferent arteriolar tone (FIGURE 6). The following sections discuss the role of purinergic signaling molecules (e.g., ATP and adenosine) in regulation of MDTGF and renal autoregulation, and the role of Cxs and extraglomerular mesangial cells. MD-TGF in rodent kidneys had a delay of 10 –15 s before initiation of a PSF response following a rapid step change in RPP and increased tubular fluid flow (87, 296, 1547); the A) ATP RECEPTOR SIGNALING. ATP can be released into the interstitium by activated MD cells (83, 203, 1064, 1510); Renal PP Connecting tubule NaCl delivery Macula densa NaCl delivery ENaC transport NO NKCC2 transport Na+/H+ exchange EETS PGE2 NO O2·- PGH2 TxA2 Adenosine ATP PGE2 Afferent arteriolar tone FIGURE 6. Flow diagram for the modulation of renal afferent arteriolar tone by conventional macula densa tubuloglomerular feedback (MD-TGF) and connecting tubule glomerular feedback (CT-GF). Changes in renal perfusion pressure (PP) are transduced by alterations in tubular fluid NaCl delivery to the MD and the CT. Subsequent changes in tubular transport lead to release of specific mediators and modulators that impact on afferent arteriolar vasomotor tone. Vasoconstrictor agents increase (⫹) arteriolar tone. Vasodilator agents reduce (⫺) arteriolar tone. ENaC, epithelial Na⫹ channel; NKCC2, Na⫹-K⫹-Cl⫺ cotransporter; EETs, epoxyeicosatrienoic acids; PGE2, prostaglandin E2; NO, nitric oxide; O2·⫺, superoxide; PGH2, prostaglandin H2; TxA2, thromboxane A2; ATP, adenosine triphosphate; CO, carbon monoxide. For details, see text (sects. IID and VIE ). 416 Physiol Rev • VOL 95 • APRIL 2015 • www.prv.org CO CARLSTRÖM ET AL. however, uncertainty exists regarding the mechanism(s) of the associated vasoconstriction of the parent afferent arteriole (FIGURE 6). ATP is thought to directly activate P2 receptors on the afferent arteriole or to activate different purinergic receptors on extraglomerular mesangial cells to initiate a Ca2⫹ wave that is transmitted to the afferent arteriole through gap junctional coupling (83, 85, 682, 1071, 1176). Alternatively, ATP and AMP may activate MD-TGF indirectly following metabolism by nucleotidases to adenosine (1510) (discussed in sect. IIA3B). ATP, adenosine, and other endogenous agents can activate vasoconstrictor purinergic homomeric P2X1, heteromeric P2X1/4, or adenosine A1 receptors, respectively, on preglomerular VSMCs (521, 565, 1510, 1540). Activation of P2X receptors by ATP induced ligand-gated cation influx, membrane depolarization, and Ca2⫹ entry via VOCCs that were reinforced by inositol trisphosphate (IP3) receptor-mediated Ca2⫹ mobilization (565, 683, 685, 1197). Studies examining the potential role of P2X receptors in renal autoregulation, in particular MDTGF responses in different nephron populations, have yielded conflicting results. ATP and P2X1 receptors are required for MD-TGF responses in JMN of both rats and mice (683, 686, 690). Knockout of P2X1 receptors attenuated MD-TGF responses and the contribution of MD-TGF to autoregulation of total or juxtamedullary blood flow (686). In one rat study, peritubular capillary infusion of ATP caused transient reduction in PSF and attenuated subsequent maximal MD-TGF response of superficial nephrons by more than 80% (1013). Continuous administration of ATP caused P2X receptor desensitization and attenuated MD-TGF, suggesting that P2 receptor signaling participated in MD-TGF (1013). However, emerging evidence from assessment of MD-TGF in superficial nephrons elicited by elevated loop flows in vivo does not support a role for P2 receptors in superficial nephrons. For example, PSF measurements revealed normal MD-TGF in mice lacking P2X1 receptors (1312), and administration of the P2 receptor inhibitors PPADS or suramin did not alter the PSF response (1306). Stepwise reductions in RPP in dogs elicited parallel reductions in whole kidney RVR and renal cortical interstitial ATP concentration (1090). Moreover, stimulation of MDTGF by increasing distal fluid delivery using acetazolamide increased renal interstitial ATP, whereas blockade of MD reabsorption with furosemide reduced ATP and renal autoregulation (1091). Thus a positive correlation between interstitial ATP and RVR adjustments has been demonstrated, and changes in ATP concentration could provide a renal interstitial signal of changed RPP and an effector agent for vasoconstriction by activation of purinergic signaling. Desensitization, saturation, or blockade of the P2purinoceptors attenuated whole kidney (952, 1131, 1438) or nephron autoregulation via blockade of the MD-TGF (683, 686, 690). Available evidence indicates that P2X1 receptors participate in renal autoregulation primarily by reg- ulation of the MD-TGF mechanism in the JMN (203, 683, 1065, 1312, 1510). In contrast to the renal microcirculation, myogenic autoregulation in the cerebral vasculature depended strongly on P2Y4 and P2Y6 pyrimidine receptors (142). Pharmacological blockade or molecular suppression of either of these P2 receptors using antisense oligodeoxynucleotides reduced myogenic constrictor tone by ⬃45%. Thus the roles of purinergic agonists and receptors in autoregulation vary according to the vascular bed. B) ADENOSINE RECEPTOR SIGNALING. Other studies strongly implicate adenosine in the MD-TGF regulation of RBF, GFR, and PGC (203, 1316, 1459, 1510, 1511). Vasoconstriction occurs primarily via activation of adenosine A1 receptors on the afferent arteriole (155, 778, 1216, 1316, 1320, 1426, 1540), but A1 receptors on mesangial cells also may modulate Ca2⫹ signaling (1122). Activation of high-affinity A1 receptors caused vasoconstriction by stimulation of G␣i proteins that inhibited cAMP production and by activation of phospholipase C (PLC) that liberated IP3 and mobilized intracellular Ca2⫹ (555, 1510). Activation of the lower affinity adenosine A2 receptors elicited vasodilatation by stimulation of G␣s proteins that activated the cAMP/protein kinase A (PKA) pathway and stimulated NOS (188, 555, 557). A2 receptor activation attenuated PP-induced arteriolar vasoconstriction by opening KATP channels to cause hyperpolarization of the plasma membrane (1450). A2 receptor-mediated vasodilation during MD-TGF can oppose A1 receptor-induced vasoconstriction (187, 188). Maximum stimulation of A2 receptors impaired renal autoregulation of afferent arteriolar tone in one study (409), but not of whole kidney RBF in another (1198). Adenosine formed by nucleotidase metabolism of ATP and AMP mediates the MD-TGF response in superficial nephrons of mice (205, 647, 1123, 1459). A1 receptor antagonists inhibit MD-TGF markedly (1320). Mice genetically deficient in A1 receptors lacked MD-TGF in superficial nephrons (155, 1426), whereas MD-TGF was enhanced in mice overexpressing A1 receptors (1125). Steady-state autoregulation of RBF was reduced by 40 –50% in A1⫺/⫺ mice (583, 751). This was ascribed to a weaker contribution of MD-TGF while the myogenic component was well preserved. Selective deletion of A1 receptors in VSMCs reduced MD-TGF responsiveness by 65%, thereby demonstrating that the main effect of A1 receptors is to mediate MD-TGFinduced afferent arteriolar vasoconstriction rather than to activate MD cells or enhance signal transmission through extraglomerular mesangial cells (896, 1122). Thus activation of A1 receptors mediates MD-TGF in superficial nephrons and contributes significantly to autoregulation in rodents, but these receptors are probably not required for the myogenic response. Pharmacological blockade of vascular A1 receptors in the isolated rabbit JGA preparation Physiol Rev • VOL 95 • APRIL 2015 • www.prv.org 417 RENAL AUTOREGULATORY MECHANISMS abolished MD-TGF (1216), similar to inhibition of 5’-nucleotidase conversion of AMP to adenosine (1229). However, some pharmacological studies in dogs, rats, and mice failed to detect a major role of adenosine or A1 receptors in MD-TGF signaling or afferent arteriolar autoregulatory responses (438, 686). Systemic administration of an A1 receptor antagonist did not impair RBF autoregulation in the dog kidney (654) and did not blunt PP-induced changes in afferent arteriolar diameter in the mouse JMN preparation (686). Propagation of a MD-TGF-induced Ca2⫹ wave along the afferent arteriole in rats also was unaffected by A1 receptor blockade (1176). In summary, activated MD cells release ATP that can be metabolized to adenosine. Uncertainties about the roles of adenosine and A1 receptors and ATP and P2X1 receptors in preglomerular autoregulatory responses to MD activation are based on controversies concerning their respective contributions to MD-TGF. There are studies supporting an involvement of both adenosine and ATP in the MD-TGF response, and contrasting conclusions may be explained by different experimental designs or perhaps by regional differences in ATP and adenosine signaling within the kidney. Available evidence demonstrates that P2X1 receptors participate in renal autoregulation primarily by regulation of the MD-TGF mechanism in JMN, whereas A1 receptors are essential for MD-TGF responses in superficial nephrons. Nevertheless, it seems clear that neither of these purinergic agents nor their specific receptors mediates the renal myogenic response. C) CXS AND EXTRAGLOMERULAR MESANGIAL CELLS. Cxs form gap junctions to connect cells. They provide intercellular channels that can facilitate the exchange of ions, second messengers, electrical signals, and metabolites. Cxs coupled multiple cell types in the JGA in the kidney, including endothelial, VSMCs, and juxtaglomerular granular cells (JGC) (553, 1545). The preglomerular vasculature and the JGA expressed primarily Cx37, Cx40, Cx43, and Cx45 (50, 553, 1439, 1546). Cx40 predominated in endothelial cells and Cx45 in VSMCs (553), although the latter has been questioned (1296). The endothelium of the proximal afferent arteriole possessed all of these Cxs, whereas the VSMCs in the terminal segment of the arteriole expressed only Cx40. Moreover, Cx40 was the major isoform in the JGA where it was expressed on endothelial cells, granular reninproducing cells, and extraglomerular mesangial cells. However, MD cells lacked Cxs, and therefore any effects of Cx40 on autoregulation likely involved transmission of MD-TGF signals to the afferent arteriole. Destruction of glomerular mesangial cells in rats by mesangiolysis or pharmacological disruption of gap junctions prevented MD-TGF responses (1218) and impaired GFR autoregulation (706, 1438), but left RBF autoregulation rela- 418 tively intact (706). Administration of 5’-nucleotidase improved GFR autoregulation following mesangiolysis, implicating attenuated adenosine signaling in the impaired GFR autoregulation (1444). Putative inhibitory peptides directed against specific Cxs have implicated them in autoregulation, but have not yet distinguished between myogenic and MD-TGF mechanisms. Blockade of Cx37 and Cx40, but not Cx43, in normal rats increased the BP and reduced RBF autoregulation (1439), whereas blockade of Cx37, Cx40, or Cx43 all impaired the RBF autoregulation in Zucker lean rats (1441). Cx37 and Cx40 in Zucker lean rats were implicated in transducing purinergic signals involved in autoregulatory responses, likely involving MD-TGF (1438). RBF autoregulation following putative Cx inhibition was improved by increasing adenosine production from AMP with 5’-nucleotidase. There is some concern about the specificity of inhibitors of gap junction function. Nonspecific effects of the gap junction inhibitors heptanol and 18-glycyrrhetinic acid have been noted even at concentrations that have no or little effect on gap junctions (979). Cx-mimetic peptides appear to be more selective in inhibiting gap junctions. However, the most definitive results have derived from Cx gene-deleted mice. Genetic deletion of Cx40 and Cx43 increased renin synthesis and secretion and disrupted the regulation of renin release by intravascular pressure and ANG II (545, 1546). Restoration of Cx40 in renin-producing JGC but not endothelial cells reduced hypertension and renin secretion of Cx40 null mice (879). Cx40 knockout mice had impaired whole kidney RBF autoregulation with a markedly reduced MD-TGF component (756). However, they retained an unchanged myogenic and third mechanism and a preserved dampening of the myogenic response by NO. Thus Cx40 contributes to RBF autoregulation via MD-TGF-mediated signal transduction from the MD. This is independent of NO but likely includes traffic across extraglomerular mesangial cells that are heavily endowed with Cx40 (756). These findings and conclusions are supported by the disruption of autoregulatory adjustments of the afferent arteriole in the JMN preparation in Cx40-deficient mice (1395). These mice lack the MD-TGF mechanism, whereas the myogenic response is preserved. Micropuncture studies of superficial nephrons in kidneys lacking Cx40 demonstrated 50% attenuation in MD-TGF (1122). In contrast, Cx37-deficient mice retain a highly efficient RBF autoregulation with normal contributions of myogenic and MD-TGF components (A. Just, personal communication). Thus Cx40 appears to be a key element for transmission of MD-TGF signal(s) that contribute to renal autoregulation. Physiol Rev • VOL 95 • APRIL 2015 • www.prv.org CARLSTRÖM ET AL. Conducted vasomotor responses in preglomerular arterioles are mediated largely by passive electronic spread of the EM transmitted through gap junctions in the arteriolar wall connecting endothelial cells and/or VSMCs, but the specific Cxs involved remain controversial. Electrically induced Ca2⫹ responses conducted along rat isolated cortical radial arteries were preserved after inhibitory peptides directed against Cx37, 40, 43, and 45 (1399), whereas propagation in mice was abolished by genetic deletion of Cx40 (1395). Interestingly, the Ca2⫹ response to NE along this arterial segment was unaffected by deletion of Cx40. In vivo studies demonstrated that vasoconstriction produced by NE, or vasodilation elicited by acetylcholine (ACh), was preserved in the renal vasculature of mice lacking Cx40 (756). Moreover, mechanical stimulation of cultured glomerular endothelial cells triggered a Ca2⫹ wave propagated from cell to cell, which was dependent on Cx40, extracellular ATP, and P2 receptors, but was independent of L-type VOCCs (1473). On the other hand, the propagation of mechanically induced Ca2⫹ waves was slower in cultured VSMCs deficient in Cx45 and in control VSMCs treated with a peptide that inhibited Cx45 gap junctional communication (554). Direct gap junction coupling was suggested since Ca2⫹ wave propagation was unaffected by the ATP P2 receptor blocker suramin. In contrast, VSMC-specific deletion of Cx45 did not affect conduction of vasomotor responses to high KCl, ACh, or adenosine along the VSMC layer of mouse cremaster arterioles tested in vivo (1296). A Ca2⫹ wave originating by activation of MD cells and passing through extraglomerular mesangial cells was dependent on ATP and P2 receptors and was blocked by pharmacological uncoupling of gap junctions (1176). ATP traversed Cx43 hemichannels in astrocytes (766). As discussed in more detail in section VIG, major differences have been noted between positive in vitro and negative in vivo findings in these studies. The Cx-like channel pannexin 1 was expressed in VSMCs of mouse afferent and efferent arterioles and may play a role in purinergic signaling (552). Pannexin 1 also was localized to the apical membrane of several tubular segments where it mediated ATP release into tubular fluid. Gap junctional communication has been implicated in myogenic responses of nonrenal arteries. Nonselective inhibition of gap junctions attenuated myogenic contractions of cerebral arteries (856), whereas inhibition of Cx37 and Cx43 attenuated myogenic vasoconstriction of rat mesenteric resistance arteries (369). VSMCs were hyperpolarized and had diminished changes in [Ca2⫹]i. Although gap junctions mediated pressure-induced VSMC cell depolarization, changes in [Ca2⫹]i and myogenic vasoconstriction in vitro, they were not required for vasoconstriction to PE. Genetic alteration of Cx40 in endothelial cells of mouse mesenteric arteries demonstrated its role in transmission of electrical (but not chemical) signals that increased the sensitivity of the myogenic response (215). In summary, in vivo and in vitro observations suggest a role for gap junctions and Cxs, particularly Cx40, in MD-TGF signaling to the afferent arteriolar cells via mesangial cells. Further studies are required to elucidate how Cxs, either as monomers or multimers, coordinate MD-TGF signal transmission and renal autoregulation. Cxs also are implicated in Ca2⫹ signaling and electrotonic and purinergic signaling along the preglomerular vasculature which may participate in nephron synchronization and renal autoregulation. The initial signaling between MD and JGC is diffusive while transmission from JGC to the AA is mainly electrotonic. The role of Cxs in modulating autoregulation is also discussed in section VIG. 4. Dynamics of autoregulatory responses Steady-state assessments of RBF autoregulation in response to graded changes in RPP have been used widely to study the effectiveness of autoregulation and the range of RPP over which autoregulation takes place (FIGURE 3A). Dynamic analyses allow a more detailed evaluation of the contributions of individual underlying mechanisms to the overall adjustments in vascular resistance or admittance. A) STUDIES IN THE TIME DOMAIN. Measurements of dynamic responses of RVR in the time domain have employed a sudden, single-step change in RPP within the autoregulatory range. Such studies revealed a fingerprint of distinct intrarenal mechanisms as a function of time after the change in RPP based on clearly separable time constants with minimal nonlinear interactions. A rapid 20 mmHg step increase in RPP led to a biphasic or triphasic increase in RVR in dogs (753, 755, 1465, 1578), rats (423, 748 –750, 756, 1313, 1342, 1640), and mice (299, 462, 505, 751, 756). This approach separated the most rapid myogenic component (0 – 8 s in rodents) from the slower MD-TGF component (10 –30 s in rodents) (FIGURE 3B) (748, 749, 753). The early myogenic response was initiated by an immediate change in transmural pressure since a similar rapid response followed a step reduction of extravascular tissue pressure (57, 265). The onset of the MD-TGF-dependent changes in RVR was delayed by 10 –15 s and completed within 20 –30 s in rodent kidneys (FIGURE 3B) (296, 749, 1547) and within 30 – 45 s in dog kidneys (753). Autoregulatory kinetics have been characterized in normal rats using the isolated, blood-perfused JMN preparation. A 50 mmHg increase in RPP elicited a biphasic contractile response with a transition point at ⬃24 s. In the absence of MD-TGF (papillectomy), the response of myogenic origin retained the same rate of contraction but terminated 11 s earlier at 13 versus 24 s with intact MD-TGF (1547), suggesting that MD-TGF enhances the myogenic mechanism Physiol Rev • VOL 95 • APRIL 2015 • www.prv.org 419 RENAL AUTOREGULATORY MECHANISMS by prolonging its response. This important study by Walker and Navar is discussed in more detail in section IIC. Additional autoregulatory mechanisms have been identified recently. A third mechanism has been proposed to increase RVR progressively from 30 to 120 s after the abrupt increase in RPP. It contributed 10 –20% to overall autoregulation in mice and rats (FIGURE 3B) and 30% in dogs (299, 749, 751, 753, 1626). Its participation may increase at low RPP in the rat (749). It was preserved in knockout mice lacking neuronal NOS, inducible NOS, and adenosine A1 receptors (299, 751) and was independent of MD-TGF in some, but not all, studies (749 –751, 753). The mediators of this response are unclear, but may reflect an interaction of known mechanisms, a delayed engagement of MD-TGF of long JMN, a contribution of renal autocrine/paracrine factors, or a component of pressure-natriuresis. ATP and P2X1 receptors may also mediate the third mechanism, or perhaps ANG II acting on AT1 receptors (284, 491, 1397). Inhibition of MD-TGF by drugs or by genetic deletion of adenosine A1 receptors unmasked an apparent fourth mechanism that operated within the 5- to 25-s time window after an abrupt increase in RPP and contributed ⬃25% to overall autoregulation (751). This time-frame overlapped that of the conventional MD-TGF and was unmasked only when MD-TGF was prevented by furosemide. However, some MD-TGF might operate independent of furosemidesensitive NKCC2 cotransport, perhaps modulated by a Na/H exchanger in MD cells (84, 431, 800, 1177, 1178, 1210). The influence of the connecting tubule glomerular feedback (CT-GF) vasodilator system, involving Na⫹ reabsorption via ENaC in the CT, is presently unclear. However, a vasodilatory phase observed 60 –200 s after a step increase in RPP in the rat (1342) may represent the contribution of the CT-GF (see sect. IIB2). Thus a third, and possibly a fourth, mechanism may contribute to renal autoregulation in the dog, rat, and mouse kidneys. Their importance and underlying signaling pathways require further study. Several rat studies have characterized dynamic adjustments of the renal vasculature following a brief period of complete occlusion of the aorta (1342, 1397, 1398, 1626), but this evoked metabolic and vasoactive factors, in addition to basic pressure-dependent autoregulatory mechanisms. Studies in the rat of complete renal ischemia for 30 s followed by a rapid single step increase in RPP reported time-dependent oscillations in the RBF responses at ⬃10, 35, and 115 s, reflecting the operation of three distinct autoregulatory mechanisms (1626), likely the myogenic, MD-TGF, and third mechanisms. A strength of dynamic analysis of vascular adjustments in the time domain is that in addition to providing useful in- 420 sight into individual mechanisms and their relative conributions, it provides reliable quantiative data for total autoregulation that agree with steady-state AI values for RBF autoregulation. B) STUDIES IN THE FREQUENCY DOMAIN. Kidneys normally exhibit oscillations in microvascular tone. Transfer function analyses of the gain of renal vascular admittance (the reciprocal of impedance and similar to steady-state conductance that is the reciprocal of resistance) to spontaneous or imposed changes in RPP can distinguish between autoregulatory components in the frequency domain (19, 230, 297, 629, 631, 722, 971, 1274, 1357, 1358, 1563, 1569, 1570). This has been investigated in conscious dogs (753, 755, 760, 1618), mice (668), rabbits (722), and rats (927, 1182, 1183, 1376). Renal autoregulation in conscious animals is efficient and similar to that recorded using the same methodology under anesthesia. One discordant study reported no blood flow autoregulation in the kidneys of conscious rats during spontaneous fluctuations in BP between 0.03 and 2 Hz (723). An admittance gain of less than 0 dB or a relative normalized admittance gain of 1.0 (analogous to an AI of 1.0) indicates active autoregulatory adjustments within the vasculature to minimize RBF changes during fluctuations in RPP. An admittance gain above 0 dB or 1.0 relative unit implies that passive vascular elasticity contributes to the amplification of the effect of RPP fluctuations on RBF. A lower relative value between 1.0 and zero quantitates the relative strength of overall autoregulation with the fractional variation in RBF being smaller than that of the RPP. As with AI, a normalized admittance gain of zero indicates perfect autoregulation. A representative recording of normalized admittance gain as a function of frequency for normal rats is shown in FIGURE 3C (297). Interpretation of admittance gain commonly is best achieved by reading from right to left, from high to low frequencies. A decline in the gain values indicates the onset of an active contribution to autoregulation. With the use of this methodology, the myogenic mechanism operating in the 0.20 – 0.10 Hz frequency band accounts for ⬃65% of total autoregulation of RBF, whereas the MD-TGF mechanism in the 0.04 – 0.011 Hz frequency band appeared to contribute ⬃35% of total autoregulation. The slower apparent third mechanism operates at frequencies of ⬃0.01 Hz and below which is below the frequency range normally examined with confidence. Most studies of dynamic autoregulation utilizing frequency analysis have reported that the myogenic response in normal rat kidneys reduces the admittance gain from approximately ⫹7.5 to ⫺5 dB (or fraction gain from ⬃2.3 to 0.56) over the frequency range of 0.20 to 0.07 Hz, whereas the MD-TGF response reduces the admittance gain by a lesser degree from ⫺2.5 to ⫺12.5 dB (fractional gain from 0.75 to 0.24) over the frequency range 0.05 to 0.020 Hz (FIGURE 7B). Summarized Physiol Rev • VOL 95 • APRIL 2015 • www.prv.org CARLSTRÖM ET AL. B RVR after a step increases in renal PP 150 100 50 Control L-NAME 0 Frequency domain analysis Admittance gain (dB) RVR (percent of perfect autoregulation) A 15 10 5 0 -5 Control L-NAME -10 -15 -50 0 5 10 15 20 25 30 35 40 45 50 55 60 0.01 Time after renal PP increase (sec) Afferent arteriolar diameter (µm) 24 Afferent arteriolar myogenic response in HNK 20 16 12 Control L-NAME 8 4 80 100 120 140 160 180 Perfusion Pressure (mmHg) D Change in afferent arteriolar diameter (µm) C 0.1 1 Frequency (Hz) Juxtamedullary nephron preparation 0 -10 -20 -30 Control L-SMTC -40 100 130 160 Perfusion Pressure (mmHg) FIGURE 7. Effect of nitric oxide synthase (NOS) inhibition of renal autoregulation mediated by myogenic and macula densa tubuloglomerular feedback (MD-TGF) mechanisms. Illustrations of the effects of inhibition of NOS with L-nitro-arginine methyl ester (L-NAME) on whole kidney autoregulation or renal blood flow (RBF), renal vascular resistance (RVR), and myogenic responses of individual vessel segments. A depicts time-dependent changes in RVR in rat kidneys following an abrupt step increase in renal perfusion pressure (PP) by release of a suprarenal aortic clamp at time zero. After generalized inhibition of NOS with L-NAME, the initial increase in RVR (corresponding to the myogenic response over the first 5 s) was greatly exaggerated and accounted for almost all of the autoregulatory increase in RVR without major contributions of other autoregulatory components, such as the MD-TGF component seen in control kidneys from 5 to 25 s or the more delayed and gradual third component thereafter (750). B shows the effect of general NOS inhibition with L-NAME on the transfer function analysis of the admittance gain as a function of frequency for spontaneous changes in renal PP and RBF in rat kidneys. After inhibition of NOS with L-NAME, the reduction in admittance between 0.07 and 0.2 Hz (corresponding to the myogenic response) was much more abrupt and the regression of admittance on frequency was steeper. Whereas control rat kidneys had a second reduction in admittance between 0.03 and 0.01 Hz (corresponding to the MD-TGF), this was hard to distinguish from the myogenic change in admittance after L-NAME (1358). C depicts changes in the diameter of the afferent arterioles of the rat hydronephrotic kidney (HNK) preparation as a function of renal PP. The decline in diameter with increasing renal PP (myogenic response) was similar in control rats and those given L-NAME, indicating no change in the myogenic response in this preparation that lacks renal tubules and MD cells (1358). D shows the change in afferent arteriolar diameter in the rat JMN preparation to increased renal PP before and after inhibition of nNOS with SMTC (S-methyl-L-thiocitruline) (662). The arteriolar autoregulatory response was increased after inhibition of MD nNOS. data for gain expressed in dB are from References 19, 286, 297, 629, 631, 748, 749, 1358, 1570. Data for fractional gain are from References 109, 499, 1193. The myogenic response in the kidneys of conscious mice reduces the admittance gain from ⫹7 to ⫺4 dB over the frequency range of 0.3 to 0.1 Hz, whereas MD-TGF reduces the gain from ⫺1 to ⫺5 dB over the range of 0.04 to 0.01Hz (668). Thus the myogenic mechanism provides the majority of the buffering of spontaneous fluctuations in RPP in rats and mice. An early comparison of methodologies in conscious resting dogs revealed that stepwise changes in mean RPP produced stronger RBF autoregulation than overall autoregulation analyzed by the transfer function admittance gain during spontaneous fluctuations in RPP (760). Dynamic autoregu- lation measured by the transfer function was only one-third as strong as steady-state adjustments produced by the step changes (⫺6.3 vs. ⫺19.5 dB). Just et al. (760) concluded that MD-TGF was not required for RBF autoregulation under normal physiological conditions, but contributed during more extreme changes in PP. Simultaneous measurements of steady-state RBF autoregulation and frequency domain analysis of whole kidney autoregulation in normal rats confirmed these methodological differences and yielded disparate AIs of 0.2 and 0.4, respectively (109). Moreover, the methodological differences were more pronounced in rats with reduced renal mass (RRM). Whereas steady-state RBF autoregulation in 3/4 nephrectomized (Nx) rats was reduced compared with uni- Physiol Rev • VOL 95 • APRIL 2015 • www.prv.org 421 RENAL AUTOREGULATORY MECHANISMS nephrectomized (UNx) or control rats (AIs: 0.7 vs. 0.2 or 0.1, respectively), frequency analysis revealed no defect in autoregulatory efficiency, with fractional admittance gain of 0.4 – 0.5 at very low frequencies in all three groups (109). Such a methodological difference may represent insensitivity of admittance analysis at low frequencies (⬍0.05 Hz) compounded by low coherence of RBF and PP determinations at these frequencies and the inclusion of time points when RPP is below the lower limit of renal autoregulation. Moreover, caution should be taken when interpreting dynamics at low PP, since this may interfere with natural operating frequencies of the myogenic and MD-TGF components resulting in added complexity due to nonlinear dynamics, interactions, and time-dependent variability, as discussed in section IIC. Another considerations is that the steady-state method assesses predominantly the response of RVR to mean RPP, whereas the dynamic method assesses primarily the admittance response to slow fluctuations in RPP (⬍0.25 Hz), which constitute a substantially smaller component of the total BP power. Bidani, Griffin, and associates (108, 109, 496, 499, 933) concluded the steadystate results were more accurate depictions of the overall impaired autoregulation that leads to hypertensive glomerular damage after RRM. More systematic comparisons of the characteristics and contributions of intrarenal mechanisms and their interactions need to be made using these different approaches. Such results will advance the field by providing a better understanding of the strengths and weaknesses of the static and dynamic methodologies. Most frequency domain studies have found that normalized admittance gain is close to unity (or 0 dB) or greater for frequencies ⬎0.3 Hz, indicating little autoregulation and a passive, compliant vasculature (FIGURE 3C) (286, 297, 629, 748). Nevertheless, one study reported that forcing of RPP between 1 and 6 Hz produced strong, sustained adjustments in afferent arteriolar diameter in the HNK preparation (927). Renal autoregulation clearly acts normally as a high-pass filter in adjusting afferent arteriolar diameter to limit changes in PGC caused by fluctuations in RPP. PGC has a pulse pressure of 7–10 mmHg which is roughly one-third of arterial BP pulsations (47, 148, 347). This demonstrates damping of the very rapid pulsations at heart rate frequencies prior to pressure transmission into the glomerulus, as is expected from the known preglomerular resistance. Nevertheless, high systolic BP may periodically exceed the autoregulatory limit and thereby contribute to glomerular injury and eventual fibrosis (108, 282). Dynamic analysis reported an active 0.01- to 0.02-Hz lowfrequency component of RBF autoregulation in conscious and anesthetized rats, which may represent the third mechanism (753, 755, 1375). An early study with limited ability to quantify fast events (⬎0.08 Hz) reported an apparent fourth mechanism operating in the 0.04 – 0.08 Hz frequency band in the dog (1618). 422 An advantage of the admittance analysis is that it can reveal the contributions of myogenic and MD-TGF mechanisms without necessitating blockade of MD-TGF. However, a limitation is that it does not provide absolute values for the relative strengths, especially of MD-TGF and slower frequency components (109, 289, 297, 631, 1313). Oscillations of arteriolar blood flow and PPT in the renal cortex were synchronous with oscillations of total RBF (1636), indicating that they originate from the vasculature. Recordings of SNBF in the efferent arteriole by laser Doppler flowmetry and of PPT in nephron pairs from the surface of rat kidneys identified two oscillatory modes: a faster (0.1 to 0.2 Hz) myogenic mode and a slower (0.02– 0.05 Hz) MD-TGF mode. A third oscillation at 0.0125– 0.0130 Hz was thought to reflect an interaction between MD-TGF and myogenic systems or possibly the third mechanism. Multiple studies indicate that the myogenic mechanism of normal rats and SHR generates spontaneous oscillations of 0.1– 0.3 Hz, whereas MD-TGF produces oscillations of ⬃0.035 Hz (289, 297, 628, 629, 631). However, the oscillations were more irregular in rats with spontaneous or renovascular hypertension (230, 625). Step changes in RPP elicited vascular oscillations at two frequencies corresponding to the myogenic and MD-TGF responses (631). 5. Relative contributions of the different mechanisms Most in vivo studies reported a ⬃40 –55% contribution of myogenic and 25–35% contribution of MD-TGF responses to dynamic autoregulation in dog, mouse, and rat kidneys to a sudden, step increase in RPP (286, 748 –751, 753, 756). Aside from frequency analysis of admittance gain, the individual components have been studied only after selective blockade of MD-TGF, since prevention of the myogenic response, for example, by blockade of VOCCs, abolished vascular responses to all autoregulatory mechanisms (749, 1064). Blockade of MD reabsorption with loop diuretics partially impairs overall renal autoregulation in kidneys of dogs (749, 760, 1622), rats (749 –751, 1626), and mice (668). Clausen and Aukland, using a previously described box technique (1601), placed a rat kidney in an airtight chamber and acutely reduced the atmospheric pressure of the chamber by 35 mmHg. This reduced RIHP and increased vascular transmural pressure in this case by stretching the vessel from outside. These authors observed a rapid, robust renal vasoconstriction that was unaffected by blockade of MDTGF by furosemide (265). They concluded that the myogenic vasoconstriction is much stronger than that induced by MD-TGF. The MD-TGF component of renal autoregulation can be abolished by ureteral obstruction (297), furosemide (19, 631, 749, 754, 1358), or genetic deletion of adenosine A1 receptors (751). As expected, all TGF components were Physiol Rev • VOL 95 • APRIL 2015 • www.prv.org CARLSTRÖM ET AL. absent from the admittance pattern in the HNK preparation lacking tubules, but the myogenic mechanism at 0.1– 0.3 Hz persisted in all of these conditions and generally became stronger and faster in the absence of MD-TGF (287, 1358), as might be expected in the absence of MD-derived NO (see sect. VIC). Overall, the myogenic mechanism appears to be dominant in that it is essential and sufficient for efficient RBF autoregulation. MD-TGF tends to provide a secondary or back-up system to enhance total efficiency when the myogenic mechanism is attenuated. One study of frequency analysis concluded that an increase in RPP increased the strength of both the myogenic and MD-TGF mechanisms (1636). The efficacy of the myogenic mechanism varies over time after a change in PP (247, 875). There are additional actions of furosemide besides inhibition of NKCC2 transport in MD cells that may contribute to its effects on renal autoregulation. Furosemide has a direct vasorelaxant action in isolated, preconstricted afferent arterioles of mice by blockade of the NKCC1 transporter (1124). Indeed, inhibition of this transporter in the afferent arteriole of the HNK preparation lacking MD-TGF attenuated ⬃70% of myogenic response to changes in RPP (1568). The natriuresis following blockade of tubular NaCl reabsorption with furosemide increased the PPT (170, 749, 1124), which might attenuate the afferent arteriolar myogenic response if the transmural pressure was reduced, although RIHP was reportedly unchanged (789). On the other hand, swollen tubules could compress peritubular capillaries to increase vascular resistance and reduce RBF (1124). Nevertheless, in one study a furosemide diuresis did not change SNGBF, although RIHP was increased (1489). Thus reduced autoregulation after furosemide may not necessarily indicate an exclusive contribution of MD-TGF. In other studies, furosemide reduced RBF or GFR in intact kidneys (1124), but the mechanism was not clear. Thus furosemide may increase or decrease RBF and GFR by mechanisms that have little to do with inhibition of MDTGF. In summary, the myogenic response normally contributes ⬃50%, MD-TGF ⬃35%, and the third mechanism ⬃10 – 20% to dynamic renal autoregulation following a step increase in RPP (FIGURE 3B). Frequency analysis admittance gain suggests less effective overall autoregulation with a rather stronger contribution of the myogenic (⬃65%) than the MD-TGF mechanism (⬃35%) (FIGURE 3C). The myogenic response contributes ⬃65% and MD-TGF ⬃35% at high RPP in the JMN preparation (FIGURE 3D). There are variable interactions between myogenic and MD-TGF mechanisms, as discussed in section IIC. B. In Vitro Studies in Isolated Preparations The myogenic response has been studied from videometric measurements of the diameter of afferent arterioles in re- sponse to changes in RPP in the JMN preparation after inactivation of MD-TGF (FIGURE 3D) or in the HNK preparation lacking tubules and thus devoid of any MD-TGF or CT-GF (FIGURE 3E). Moreover, use of single isolated, perfused afferent arterioles allows calculation of the slope of the regression of active wall tension on PP, which provides a quantitative measure of the myogenic response throughout the PP range studied (FIGURE 3F). The intercept of this regression on the x-axis indicates the threshold for initiating a myogenic response. Myogenic and MD-TGF mechanisms also have been studied using the isolated double-perfused JGA preparation (702, 703). 1. The isolated JGA Bell and associates (801) developed a rabbit nephrovascular unit in which a glomerulus was attached to its MD segment that was perfused via the TAL to characterize epithelial transport properties. Early studies of renin release from the JGC of the microdissected afferent arteriole were performed with the glomerulus attached or absent (698, 1377). Mechanisms regulating renin release for JGC were also investigated in the HNK preparation (162, 163). Subsequent studies demonstrated pressure-dependent renin secretion from a perfused rabbit afferent arteriole with parent glomerulus present (122, 1181). In 1987, Skott and Briggs (1378) used the isolated JGA to show that luminal NaCl concentrations influenced MD signaling that regulated renin secretion from JGC. Shortly thereafter, Ito and Carretero (702, 703) characterized the myogenic and MD-TGF responses in a rabbit JGA preparation that was doubleperfused via the MD and the afferent arteriole. Activation of MD reabsorption by increasing tubular [NaCl] constricted the afferent arteriole adjacent to the glomerulus, whereas a pressure-induced myogenic constriction occurred in more proximal segments of the afferent arteriole. This preparation has been used commonly to characterize MD metabolites that modulate the strength of afferent arteriolar responses to MD-TGF signaling. Examples include potentiation by ATP, adenosine, O2⫺ (913, 1217), and attenuation by NO and CO (702, 1225, 1226). Recent studies in the mouse isolated JGA indicated that MD cells can detect variations in tubular fluid flow, independent of the salt composition, by activation of primary cilia on MD cells, which subsequently increases [Ca2⫹]i in afferent arteriolar VSMCs and causes vasoconstriction (1374). 2. The isolated connecting tubule and glomerular/ afferent arteriolar unit A variation of the isolated JGA is a tubulovascular unit consisting of an individual CT and attached afferent arteriole and glomerulus (822, 1035, 1230). Many CTs, especially in the outer cortex, return to their parent glomerulus and attached afferent arteriole (339, 1538). Three-dimensional tracing of 168 nephrons indicated that the terminal Physiol Rev • VOL 95 • APRIL 2015 • www.prv.org 423 RENAL AUTOREGULATORY MECHANISMS portion of the distal convoluted tubule contacted the afferent arteriole of the parent glomerulus in 90% of shortlooped and 75% of long-looped nephrons of the mouse kidney (1215). Such findings reaffirmed previous observations of 100% contacts in superficial nephrons and 60% of midcortical nephrons of the rat kidney (339). Earlier serial sections of 60 nephrons revealed that the late distal tubule came within 3 m of the afferent arteriole in 90% of superficial glomeruli, 87% from midcortical glomeruli, and 73% of juxtamedullary glomeruli (67). Carretero and associates have conducted a series of in vitro and in vivo studies establishing the functional implications of CT-GF regulation of afferent arteriolar vasomotor tone. Tubular flow-dependent reabsorption of Na⫹ by the rabbit or rat CT was mediated primarily by the amiloride/benzamil-sensitive ENaC (1230). Increased Na⫹ reabsorption promoted epithelial cell production of PGE2 by COX-1 and an EET by a cytochrome P-450 epoxygenase (1219, 1221), both of which dilate the afferent arteriole. PGE2 induced endothelium-independent dilatation via activation of the EP4 receptor (1223). Earlier immunolabeling studies of principal cells that express ENaC suggested colocalization of membrane-associated PGE synthase with COX-1 but not COX-2 (176, 1539). In contrast, COX-2 but not COX-1 was expressed in MD cells in both rats and mice. COX-2 was also expressed in the intra- and extraglomerular mesangium. Tubular ANG II enhanced CT-GF, probably by simulating Na⫹ reabsorption in the CT (1219). The effect of ANG II on CT-GF was mediated via a protein kinase C (PKC)/NOX2/O2⫺ pathway (1227). In contrast to the conventional MDTGF, this suggests a vasodilator component to ANG II action due to CT-GF. Moreover, inhibition of tubular NOS paradoxically increased the vasodilation induced by CTGF, suggesting that NO is not a mediator of the vasodilation but that NO rather inhibits Na⫹ reabsorption in CT, thereby preventing the generation of the signal for the vasodilator response (1230). In vivo micropuncture studies showed that increased tubular NaCl delivery activated the CT-GF circuit to reduce the estimated PGC and offset the MD-TGF-mediated constriction of the preglomerular vasculature (1558). CT-GF contributed also to acute resetting of MD-TGF during changes in ECV and Na⫹ excretion (1554). Such vasodilation and resetting were more pronounced in Dahl SS than in SHR rats consuming a high-salt diet, thereby leading to increased PGC in the Dahl SS rats (1555). In the absence of the MD-TGF circuit, activation of CT-GF in vitro blunts the magnitude of the PP-induced myogenic contraction of the afferent arteriole (Carretero and associates, personal communication). 3. The juxtamedullary nephrovascular unit Casellas and Navar (202) developed an in vitro JMN preparation perfused with low hematocrit blood or an albumin 424 solution to study the microvascular reactivity and the MDTGF of deep cortical nephrons. A rat kidney was sectioned longitudinally and the papilla reflected to expose the overlying pelvic mucosa on the inner surface of the cortex where the glomerular arterioles were visualized directly. Increased RPP reduces the luminal diameters of the cortical radial artery and afferent arteriole, but the diameter of the efferent arteriole is unchanged or even increased (193, 199 –201, 580, 611, 1281, 1395, 1437). This preparation allows separation of the myogenic response from the MD-TGF without the potential complication attendant on the use of loop diuretics by preventing MD perfusion by a tubular oil block or papillectomy (200, 202, 686, 1020, 1281, 1395, 1437). A doubling of RPP reduced afferent arteriolar diameter by 15–30% (1020, 1437), which was sufficient to yield nearperfect autoregulation of blood flow since resistance to flow is a fourth power relation to the radius of the vessel. The reduction in luminal diameter was limited to 7–10% after elimination of MD-TGF by papillectomy, oil blockade to halt tubular flow, or furosemide (FIGURE 3D) (1020, 1437). Abolition of an active response with a Ca2⫹-free solution increased arteriolar diameter by 20% when RPP was doubled (1020, 1437). Thus the modest reduction in arteriolar diameter during a doubling of RPP after blockade of MDTGF still represents a considerable increase in vascular tone. Afferent arteriolar autoregulatory constriction following increased RPP from 60 to 140 mmHg was attenuated by ⬃35% by blockade of MD-TGF, indicating the myogenic mechanism accounts for ⬃65% (FIGURE 3D). Both mechanisms were abolished by blockade of VOCCs with nimodipine. Measurements of hydrostatic pressures along the preglomerular vasculature during increased RPP indicated that 20% of the increase in resistance occured before the end of the cortical radial artery, 65% by the end of the afferent arteriole, and 80% along the composite preglomerular vascular tree (201). The terminal glomerular segment of the afferent arteriole was twice as responsive as the upstream segments (200, 1020, 1437). Afferent arteriolar blood flow remained constant during changes in RPP above 95 mmHg (580, 1437). Increasing RPP from 80 to 120 to 180 mmHg increased PGC only modestly from 45 to 50 to 53 mmHg, respectively (1281). Similar studies reported a 0.10- to 0.19-mmHg change in PGC per mmHg change in RPP within the autoregulatory range (193, 201). Thus this preparation retains excellent autoregulation of nephron flow and PGC within the physiological range. 4. The isolated perfused afferent arteriole Edwards (375) developed a rabbit isolated, perfused renal arteriolar preparation in 1983 to study the myogenic response and reactivity to vasoactive agents. Initial studies reported maintenance of luminal diameter of cortical radial arteries and afferent arterioles or constriction up to 11% to increased PP compared with dilation of efferent arterioles Physiol Rev • VOL 95 • APRIL 2015 • www.prv.org CARLSTRÖM ET AL. (375). Thereafter, the myogenic response was characterized in isolated preglomerular vessels from dogs (562, 777, 1059), rabbits (697, 747), rats (700, 1224, 1637), and more recently from mice (462, 728, 729, 861, 864, 866). A doubling of PP led to an 11% reduction in diameter of mouse afferent arterioles that was complete within 5 s due to a linear increase in active wall tension as a function of PP (FIGURE 3F) (861). While this preparation excludes nonvascular paracrine factors, it requires an artificial perfusate free of erythrocytes. Moreover, it is technically challenging to measure the precise PP at the tip of the perfusion pipette. Myogenic contraction, assessed as active wall tension, is determined as the difference between the tension in physiological versus Ca2⫹-free EGTA-containing solutions. Myogenic contraction was associated with increased VSMC [Ca2⫹]i and increased ROS measured by fluorometric markers (864, 1224). The slope of the calculated regression of active wall tension on PP quantifies the myogenic response with the intercept on the x-axis defining the threshold PP for initiating a response (FIGURE 3F). The threshold pressure was 35– 40 mmHg for the mouse isolated afferent arteriole (861, 864), 60 – 80 mmHg for the preglomerular vasculature of the rat (700, 1637) and rabbit (747), and 80 mmHg for the dog (562, 1059). The estimated or directly measured PGC in the euvolemic rat averaged 55 mmHg (42, 47, 58, 74, 87, 118, 199, 320, 334, 416, 1008, 1488) and was calculated to be similar in the mouse (861). PGC was estimated to be 60 –70 mmHg in the dog kidney (86, 1068, 1070, 1073, 1138, 1139) and 31 mmHg in the anesthetized rabbit (323). Thus these data suggest that myogenic responses generally start at PPs below ambient levels of PGC, except in the rabbit, and may thereby contribute to both dilation of the afferent arteriole during reductions in RPP and constriction during increases in PP. 5. The HNK Steinhausen and associates (1413, 1415) developed the atubular, postischemic HNK preparation to facilitate direct visualization of the renal cortical microcirculation and glomeruli. This preparation allows intravital microscopic measurements of vessel diameter and erythrocyte velocity in an exclusive vascular preparation perfused with blood or artificial solutions. During the development of hydronephrosis, the kidney parenchyma becomes progressively thinner due to tubular atrophy, resulting in a tissue sheet of cortical vessels of ⬃200 m thick. Approximately 6 – 8 wk after unilateral ureteral obstruction, there was complete tubular atrophy (162, 1415). This preparation has been used to investigate mechanisms of renin release and electrophysiological characteristics of JGC of the afferent arteriole (162, 164). In addition to being readily amenable to study of electrophysiology of the renal microcirculation, the HNK provides an opportunity to investigate the myogenic response of various segments of the preglomerular vasculature in the absence of input from either MD-TGF or CT-GF. Videometric measurements were made of the diameter of renal arterioles during perfusion in situ with blood or in vitro with an artificial saline solution (1413, 1415). Loutzenhiser and associates (586) have used the HNK extensively to characterize the myogenic response in the absence of any TGF. Increased RPP contracted cortical radial arteries and afferent arterioles but not efferent arterioles (FIGURE 3E) (586). Furthermore, frequency domain analysis reported a relatively well-maintained myogenic mechanism with a threshold for activation of 60 mmHg (287) and preglomerular vasoconstriction of 7–23% with a doubling of RPP (586) that was complete within 5–10 s (802, 927, 931). The EM of cortical radial arterial and afferent arteriolar VSMCs averaged ⫺40 mV at 80 mmHg, which was just below the threshold required to initiate a myogenic response (928). Overall, RBF autoregulation in this preparation, which lacks MD-TGF, was weak when perfused with a salt solution devoid of colloid or erythrocytes. This was apparent from a steady-state AI of ⬃0.85 over the RPP range of 60 –140 mmHg and a dynamic autoregulatory fractional admittance gain of only ⬃0.8 in the PP range of 100 –140 mmHg (287). These values are considerably higher than the same determinations in efficiently autoregulating intact kidneys during inhibition of MD-TGF (287, 297, 749, 753, 760). The HNK preparation suffers from some other limitations including the low viscosity of a cellfree perfusion solution which influences normal NO production and precludes its uptake into oxyhemoglobin. Nevertheless, the preparation responded well to ACh-induced NO release (1486). C. Interactions Between MD-TGF and the Myogenic Mechanisms Interactions between the myogenic and MD-TGF mechanisms provide renal autoregulation with unique opportunities to regulate preglomerular vascular tone (200, 231, 245, 285, 625). The most distal portion of the afferent arteriole was the major effector site for both the myogenic and the MD-TGF responses (200). Studies have demonstrated positive, negative, or absent interactions between these two mechanisms. The participation of a glomerular feedback includes both the conventional MD-TGF and the more recently described CT-GF. It is firmly established that the MD-TGF influences the myogenic response, which in turn impacts the MD-TGF (244 –246, 630, 1163, 1205, 1341, 1401, 1402, 1547). Interactions between the oscillating frequencies of the myogenic and the MD-TGF mechanisms have been observed in whole kidney blood flow and single-nephron studies. Early studies of myogenic fluctuations in SNGBF in the superficial cortex revealed large amplitude oscillations at 0.1– 0.2 Hz in response to inhibition of MD-TGF with furosemide, whereas the myogenic oscillations were strongly damped when MD-TGF was stimulated maximally by tubular per- Physiol Rev • VOL 95 • APRIL 2015 • www.prv.org 425 RENAL AUTOREGULATORY MECHANISMS fusion (1636). This suggested that MD-TGF tonically inhibits the amplitude of intrinsic vascular tone driven by the myogenic mechanism. Also inhibited were MD-TGF-dependent communications with adjacent afferent arterioles and glomeruli (626). Recent wavelet analysis of blood flow and tubular pressure confirmed that MD-TGF modulated the frequency and amplitude of oscillations initiated by myogenic mechanisms (1400). These dynamic interactions occur within and between nephrons and are nonlinear and time-varying (405, 630, 973, 975). Conversely, delivery of the tubular fluid to the MD and the CT is altered by the myogenic mechanism and thereby modulates TGF responses. Such coupling has been modeled extensively with descriptions of nonlinear modulation of the amplitude and frequency of myogenic oscillations by MD-TGF (244, 245, 395, 875, 973, 973, 975, 976, 1205, 1258, 1400). These interactions appeared to vary between cortical and medullary nephrons (974) and may contribute to fluctuations in the RPP set-point for RBF autoregulation (1172). The interactions were more variable in SHR than in SD rats (247, 626). Based on simulation studies (395), Holstein-Rathlou and associates concluded that the myogenic response needs to be active to have a MD-TGF mechanism participate in RBF autoregulation. Indeed, there are examples when MDTGF is present and yet RBF autoregulation is very weak to absent. However, studies using frequency domain analysis have concluded that autoregulatory responses in one afferent arteriole can generate both positive and negative interactions with others (231, 245, 246, 628). MD-TGF can modulate both the frequency and amplitude of oscillations arising from the myogenic mechanism. Most nephrons in normotensive rats originating from a common renal cortical radial artery had highly regular periodic synchronization of MDTGF and myogenic modes. However, the interactions in SHR were more complex with transient, irregular coupling and synchronization (231, 875, 1205, 1401). Also, the lowfrequency (0.01 Hz) oscillatory component of renal autoregulation, perhaps representing the third component, modulateed both the myogenic response and MD-TGF activity (1163, 1341, 1375), with weaker interactions in SHR than in SD rats (1375). In vivo micropuncture studies by Schnermann and Briggs (1308) reported that the maximum MD-TGF-mediated reduction in the estimated PGC was 13 mmHg when RPP was 120 mmHg, but was reduced to 3 mmHg when RPP was lowered to 80 mmHg. Thus the magnitude MD-TGF increased in parallel with RPP, and effective autoregulation of the estimated PGC was strongly dependent on tubular flow past the MD. These results demonstrate two-way positive interactions between myogenic and MD-TGF mechanisms in vivo. The MD-TGF response is required for a full myogenic response, and conversely, the degree of myogenic contraction regulates the reactivity of MD-TGF. 426 Recent studies have provided further evidence of a positive reinforcement between the MD-TGF and myogenic mechanisms. Dynamic studies using frequency analysis in the rat found that MD-TGF inhibition with furosemide attenuated characteristics associated with myogenic reactivity (reducing the slope of gain reduction and the associated phase peak) (1358), suggesting removal of positive MD-TGF modulation of the myogenic mechanism. In the conscious mouse, furosemide eliminated the frequency domain signature of MD-TGF and reduced the shoulder operating frequency of the myogenic mechanism (from 0.3 to 0.2 Hz), indicative of a slowing of the myogenic response (668). Dynamic studies by Walker and Navar (1547) on the rat JMN preparation in the time domain revealed a positive interaction between MD-TGF and the myogenic response of afferent arterioles. A rapid step increase in RPP produced a prompt but relatively short-lived (⬃13 s) monotonic myogenic vasoconstriction of the afferent arteriole when MDTGF was inactivated by papillectomy, whereas there was a biphasic contractile response with the initial myogenic response lasting ⬃24 s when MD-TGF was intact, indicating positive interaction between these two mechanisms (1547). With MD-TGF operational, a biphasic response included a decline in afferent arteriolar diameter after 24 s, albeit at a slower rate, to reach a 26% reduction at 93 s. The initial myogenic response had a time constant of 6 –11 s (0.16 – 0.09 Hz), whereas that of MD-TGF was 37 s (0.027 Hz). The delay in the initiation of the myogenic response was 6 s, and the rate of myogenic contraction was the same whether MD-TGF was engaged or inoperative. There was no evidence for a very slow third mechanism in this study. Autoregulation of the in vitro JMN preparation was stronger in the initial but not the sustained phase after MD-TGF was activated by increasing NaCl delivery to the MD (662). It is noteworthy that inhibition of MD-TGF with furosemide did not affect the myogenic response in vivo after NOS was inhibited (749, 750), a finding consistent with an action of furosemide to inhibit MD-TGF and NO generation and modulation of myogenic tone. However, several studies suggested weak to no interaction between MD-TGF and the myogenic mechanism. Transfer function analysis revealed that furosemide inhibited the MD-TGF signature at 0.03– 0.05 Hz with a weak effect of the myogenic response between 0.1– 0.2 Hz in the rat, reducing the gain at frequencies below 0.15 without changing the peak of the phase angle (19). Similar inhibition of MDTGF had little effect on the myogenic response in the dog (760) or mouse (299), but furosemide paradoxically augmented the myogenic response in adenosine A1 receptor deficient mice lacking MD-TGF (751). However, inhibition of MD-TGF by ureteral obstruction in the rat abolished the MD-TGF signature of admittance gain without affecting dynamic features of the myogenic response (297). Consistent with a lack of effect of MD-TGF on myogenic respon- Physiol Rev • VOL 95 • APRIL 2015 • www.prv.org CARLSTRÖM ET AL. siveness, activation of MD-TGF by inhibition of proximal NaCl reabsorption by acetazolamide to increase MD delivery of NaCl had little influence on the myogenic mechanism in vitro (662) or in vivo (749). An increased myogenic tone elicited by reducing perirenal pressure in the isolated perfused rat kidney was unaffected by MD-TGF inhibition with furosemide (265). Thus myogenic vasoconstriction can function independently from MD-TGF. RVR responses to a sudden, step increase in RPP gave the impression that the initial myogenic response was enhanced during inhibition of MD-TGF with furosemide, increasing its participation in the first 10 s from 40 to 60% of the total response in the dog (751, 753) and from 40 to 55% of total efficiency with an enhanced response rate in the rat (749, 750). However, the appearance of coupling between MDTGF and myogenic mechanisms depends crucially on the protocol used and the operational status of MD-TGF while RBF is normal and PP is varied within the autoregulatory range. During frequency analysis to generate a transfer function of admittance gain, spontaneous or forced oscillations of RPP continually change tubular flow past MD cells to modulate MD-TGF activity in positive and negative directions. In contrast, analysis based on a sudden step change in RPP usually starts with a reduced PP and diminished tubular flow and MD-TGF activity. The resultant vasodilation tends to offset or counter the stimulus for rapid myogenic vasoconstriction when RPP increases. This may explain the increased strength and speed of the myogenic response observed following a rapid increase in RPP when MD-TGF is inhibited since the opposing preexisting vasodilator input to arterial tone is removed. This spurious result is less evident during inhibition of the more symmetrical activity of MD-TGF during frequency analysis of admittance gain. Inhibition of MD-TGF by acute volume expansion tends to increase the contribution of the myogenic mechanism (from ⬃33 to 50%) of total autoregulatory adjustments in RVR to a sudden step increase in RPP in the wild-type mouse (462, 505). Again, this could be explained by removing the restraining effect of preexisting MD-TGFinduced vasodilation at low tubular flow on the initial myogenic vasoconstriction following the increase in RPP. Total steady-state autoregulatory efficiency was unaffected by such inhibition of MD-TGF. Collectively, these results suggest that the myogenic mechanism is more designed to protect against increases in BP and that the MD-TGF negativefeedback system is more supportive of RBF and GFR during reductions in BP. Furosemide might cause some renal vasodilation by increasing NaCl delivery to the CT and stimulating CT-GF. Afferent arteriolar vasodilation mediated by CT-GF attenuated the strength of vasoconstriction by MD-TGF and myogenic mechanisms (1554, 1555, 1558) (O. A. Carretero, personal communication). However, furosemide had little effect on the contribution of the putative third mechanism in either the dog or rat (749, 753), although it was abolished by furosemide in a mouse study (751). Furosemide may also impact autoregulation by inhibiting Na⫹-K⫹-2Cl⫺ transport in VSMC to affect preglomerular vascular tone (522, 835, 1124, 1568). Effects of loop diuretics on NKCC1 transport by VSMC and their impact on myogenic tone of the afferent arteriole in vitro are discussed in section IVA. Four mechanisms have been proposed to account for observed positive interactions between the MD-TGF and the myogenic responses. First, a MD-TGF-induced constriction of the terminal afferent arteriole should increase the upstream intravascular pressure and thereby enhance the signal for a myogenic contraction in more proximal afferent arteriolar segments (200, 1021). Second, MD-TGF triggered a Ca2⫹ wave that spread from the MD throughout the JGA to the parent afferent arteriole that could enhance myogenic responsiveness (1176). Third, membrane depolarization of the afferent arteriole by an active MD-TGF should enhance Ca2⫹ entry through VOCCs and thereby myogenic contractions. Fourth, MD-TGF-induced depolarization of the afferent arteriolar VSMCs can be transmitted upstream (976, 1176) to constrict neighboring afferent arterioles stemming from the same cortical radial artery (231, 762). Electrical stimulation of the distal afferent arteriole initiated electrical and Ca2⫹ entry responses that were conducted upstream along the afferent arteriole (527) and cortical radial artery (1279), presumably mediated by gap junctions (527). Depolarizing current caused vasoconstriction and hyperpolarizing current caused vasodilation (1414). High KCl-induced depolarization also elicited a constrictor response of upstream arterioles and neighboring afferent arterioles, with a long mechanical length constant of ⬃325 m (1279, 1544). The strength of internephron coupling was greater in SHR than in SD rats (1544). Such coupling could account for synchronized MD-TGF-mediated contractions in pairs or triplets of nephrons (630). MD-TGF initiated stronger interactions with afferent arterioles originating from a common cortical radial artery in SHR than in SD rats (231). The molecular mechanisms by which MD-TGF interacts with the myogenic mechanism are not certain. Positive interactions may reflect vasoconstrictor signaling Ca2⫹ pathways or perhaps involving ATP and P2X1 receptors or adenosine and A1 receptors, as discussed in section IIA3, A and B. Alternatively, as is discussed in section VIB, NO generated by neuronal NOS in MD cells during activation of MDTGF could account for a negative interaction since endogenous NO markedly inhibits both the strength and the speed of the myogenic response. In summary, it is clear that there are communications and interactions among vascular and tubular mechanisms mediating autoregulation in the kidney, but their physiological significance remains elusive. Both reinforcing and opposing Physiol Rev • VOL 95 • APRIL 2015 • www.prv.org 427 RENAL AUTOREGULATORY MECHANISMS interactions between myogenic and MD-TGF mechanisms have been reported. This may relate to coupling based on the balance of positive and negative modulators of vasomotor tone of afferent arterioles, which can be generated by activated MD or CT cells, as summarized in FIGURE 6 and discussed in more detail in section VI. The complexity is increased further by the fact that the degree of information transfer can vary in a nonlinear manner over time and MDTGF may contribute intermittently to autoregulation. The cellular events and consequences of interactions between MD-TGF as well as CT-GF mechanisms and the myogenic response are poorly understood, but modulation of NO and Ca2⫹ signaling appear to be involved. As is discussed in section VIC, NO modulates the renal myogenic response and interactions between tubular and vascular mechanisms, but the primary source is still controversial. D. Kinetics of the Myogenic Response Both the magnitude and the kinetics of the myogenic response vary among different circulatory beds. There is a tendency for the speed of the myogenic response to be greatest in the smallest diameter arterioles. The renal vasculature has the fastest and most complete response, followed by the cerebral and then mesenteric arterioles. The myogenic response of renal afferent arterioles of mice, rats, and dogs (internal ID: 8 –20 m) is often complete within 5–10 s, whereas in other vascular beds it ranges from ⬃10 –15 s for small cremaster muscle arterioles (15–25 m) (613, 613, 615) to 60 –120 s for larger mesenteric and cremaster arterioles (⬃100 m) (303, 740, 1671) and up to 5 min for mesenteric arteries and skeletal muscle arterioles (⬎180 m) (243, 667, 1002). This diversity is probably related to a specific set of VSMC mechanosensation-contraction signaling systems. Whereas the depolarization and Ca2⫹ entry through VOCCs are ubiquitous, distinct upstream signaling systems appear to provide tissue-specific mechanisms of adaptation to local metabolic needs involving the supply of oxygen and fuels that are triggered by changes in BP. The myogenic response in the renal microcirculation is engaged within 1 s and is complete in ⬍5–10 s in intact rodent kidneys (265, 462, 505, 749 –751, 756, 1640), mouse isolated afferent arterioles (861) or rabbit (747), or rat afferent arteriole of the JMN (1547) or the HNK preparation (927). This time frame corresponds closely to the 0.1– 0.3 Hz frequency band for the myogenic admittance transfer function (FIGURE 3C) (109, 289, 297, 629, 749). Slower responses of 40 s were reported in one study of rat pressurized JMN arterioles (1637) and of 60 –120 s in hamster preglomerular arterioles transplanted to the cheek pouch (473). As already discussed in the previous section, Walker and Navar (1547) detected two components in the autoregula- 428 tory contractile response to a single step increase in PP of the JMN preparation. The prolonged delay (⬃6 s) in the initiation of the myogenic response reported in those studies of the JMN preparation may reflect methodological problems, since much more rapid responses have been reported in other studies among isolated afferent arterioles or in the HNK preparation. Loutzenhiser et al. (927) used the rat HNK preparation to characterize the kinetics of the myogenic response in the absence of any tubular influence. The vasoconstrictor response to an 80-mmHg rise in RPP was a monotonic contraction after a delay of ⬃0.3 s, with a time constant of 4 s. The onset of the myogenic response of the HNK, based on changes in arteriolar wall diameter, was very rapid and averaged ⬃0.30 – 0.35 Hz, which is considerably faster than values reported in vivo. A possible contributor to this unusually rapid response is the use of a low viscosity, colloid, and RBC-free saline perfusion solution as well as the lack of tubular modulatory agents. The vasodilator response to an 80 mmHg reduction in RPP was slower and more complex. After a delay of 1 s, the response was biexponential with time constants of 1 and 14 s, requiring almost 25 s for complete relaxation. Based on those results, the authors concluded that rapid pulsations should evoke sustained vasoconstriction due to the faster contraction when RPP rose compared with the more sluggish vasodilatation when PP fell. Consistent with this prediction, the magnitude of afferent arteriolar contraction in the HNK preparation was related to the peak systolic pressure for PP oscillations between 1 and 6 Hz. These investigators proposed that the primary function of the myogenic response at the normal heart rate frequency is to protect glomerular capillaries from the damaging effects of the systolic BP rather than to maintain a constant GFR to regulate body fluid homeostasis (933, 934). In contrast to these results for the HNK preparation, most whole kidney studies of frequency domain analysis of renal vascular admittance have reported a frequency for the myogenic response of 0.20 – 0.25 Hz (FIGURE 3C), as discussed earlier in section IIA5. Moreover, Just and Arendshorst (749, 751) reported that intact kidneys had a similar rate of myogenic contraction and relaxation to a 20-mmHg step change in RPP. Both responses were complete within 8 s (749, 751). The myogenic contraction to an increase in RPP in intact kidneys occurred slightly earlier, with a shorter delay time (0.39 vs. 0.53 s) and contributed a smaller fraction of total autoregulation of ⬍37% compared with the myogenic relaxation to a decreased RPP which was ⬍52% (750). Moreover, the time for complete contraction of 5.1 s was longer than for complete relaxation of 2.6 s. Accordingly, the in vivo renal myogenic response buffered increased and decreased RPP similarly and thus should respond primarily to the mean RPP, rather than preferentially to the systolic BP. Support for this conclusion comes from Physiol Rev • VOL 95 • APRIL 2015 • www.prv.org CARLSTRÖM ET AL. earlier studies that failed to detect an influence of arterial pulse pressure on RBF autoregulation (1238, 1344). The observed dynamic values for autoregulation in the studies of Just and Arendshorst (750) were likely underestimates since the in vivo changes in RPP, although abrupt, were not instantaneous. strictors, e.g., ANG II or AVP, to increase RPP had no major effect on the renal myogenic response (750, 1193, 1563). The rate of change in RPP is another factor that may modulate renal autoregulatory responses. During stepwise increases in RPP in conscious rats, larger changes in RVR were associated with more rapid changes in PP (423). These striking differences reported for the myogenic contraction between the in vitro atubular HNK and in vivo intact kidney preparations may relate to the presence of active tubular cells only in the latter, since MD metabolites impact on VSMC kinetics. Specifically, NO production affects the myogenic mechanism, as discussed in section VIC. Moreover, activation of MD cells can accelerate the rate of relaxation associated with the generation of PGE2 (218, 1036, 1334) and NO (298, 1036). Indeed, Loutzenhiser et al. demonstrated later that adding exogenous NO or cGMP to the HNK preparation to more closely simulate the in vivo paracrine modulation, preferentially reduced the delay in the vasodilation response to a reduced RPP (R. Loutzenhiser, personal communication), thereby reducing the time difference between myogenic relaxation and constriction which more closely approximated to those observed in vivo. The myogenic response appears to be somewhat faster in conscious than in anesthetized animals. Studies by Cupples, Bidani, Griffin and colleagues reported the resonance frequency of the myogenic mechanism to be 0.23– 0.25 Hz in conscious rats (886, 1193), compared with the general finding of 0.18 – 0.21 Hz in rats anesthesized with isofluorane or barbiturates (109, 297, 499, 597, 749, 1274, 1358, 1569, 1570, 1572). There were similar responses during anesthesia with thiobutabarbital (597, 1274) and pentobarbital (297, 749), but halothane slowed the operating frequency of the myogenic mechanism in SD, SHR, and WKY rats to 0.10 – 0.15 Hz (230, 289, 631, 886, 1636). The resonance peak in admittance gain of dynamic autoregulation is an index of the natural frequency of the system (628). The dynamic autoregulation of arterial blood flow in euvolemic Wistar and SHR rats was excellent in the renal and in the superior mesenteric circulation, although the myogenic response was faster (0.22 Hz) in the denervated kidney than in the denervated gut (0.13 Hz) (7). This was confirmed in two other studies (886, 1572). In contrast, blood flow to the hindquarters lacks any dynamic autoregulatory signature. Cupples and associates (1572) reported that the range of RPP tested in the HNK preparation perfused with artificial medium determined the speed of the myogenic response assessed as transfer function admittance gain. Thus an increase in mean RPP from 80 to 140 mmHg increased the myogenic operating frequency from ⬃0.265 to 0.37 Hz. In a similar in vivo study in Wistar rats, inhibition of NOS with L-NAME increased the RPP from 115 to 140 mmHg and increased the myogenic resonance frequency from 0.21 to 0.25 Hz. This change was attributed primarily to PP since it was prevented by maintaining PP by aortic clamping during NOS inhibition. This pressure sensitivity was unique to the kidney as the myogenic resonance frequency in the mesenteric circulation (0.15– 0.16 Hz) was unaffected by LNAME hypertension. In another study by the same laboratory, however, hypertension (115–145 mmHg) induced by L-NAME was reported to increase the strength of the myogenic response (both the slope and the amplitude of admittance gain in the 0.1– 0.2 Hz window) without affecting its speed; the myogenic resonance frequency was unchanged at 0.18 – 0.20 Hz (1569). Other studies employing vasocon- Neurohormonal and paracrine agent also modulate the speed and strength of the myogenic response, as exemplified by the effect of NO, discussed earlier and first reported by P. Persson and associates (1626). ANG II is reported to augment the myogenic resonance peak from 0.23 to 0.27 Hz in rats and increases the fractional gain by 25% and the slope of the reduction in gain, whereas phenylephrine (PE) decreases fractional gain by 30% and the slope of the gain reduction without affecting the resonance peak (0.24 – 0.25 Hz). PE had little effect on MD-TGF (1193). The myogenic resonance peak also was increased by ANG II in conscious dogs (755). As is discussed in section VIC, NOS inhibition in the intact kidney markedly accelerates and magnifies the myogenic mechanism. Expression of different isoforms of smooth muscle myosin heavy chain (MHC) has been proposed to explain the more rapid contraction of the afferent arteriole compared with the efferent arteriole. The B-isoform of MHC inserts into the ATP binding pocket to activate VSMC contraction. The afferent arteriole expressed predominantly the rapidly recycling MHC-B isoform, whereas the efferent arteriole expressed only the slower-cycling, alternatively 5’-spliced MHC-A isoform (1161, 1361). However, the rate of afferent arteriolar contraction was unaltered in mice with a mutated form of MHC-B (1161). Thus variable expression of VSMC MHC isoforms could not explain the differential speed of contraction of afferent and efferent arterioles. In summary, myogenic tone changes rapidly with PP and adjusts the RVR to the systolic or mean BP according to the preparation used. The more rapid constriction than relaxation reported in preparations lacking tubules likely relates to the absence of MD-derived NO and PGE2 that are implicated in accelerating relaxation in vivo. The times of contraction and relaxation in vivo, when measured in response Physiol Rev • VOL 95 • APRIL 2015 • www.prv.org 429 RENAL AUTOREGULATORY MECHANISMS to a step increase or decrease in RPP, are more symmetrical. Moreover, the myogenic response appears to be somewhat faster in conscious animals. E. Vasomotion The myogenic mechanism of most microvessels generates oscillations in vascular tone and organ blood flow termed “vasomotion” (1, 1169) which oscillate in vivo with frequencies of 0.05– 0.2 Hz, likely representing summation of vasomotion in many individual small arteries (2, 854, 1087, 1408). The underlying mechanisms are still not fully understood and may even differ between vascular beds. Although vasomotion can be induced by PP and involve similar Ca2⫹ signaling pathways as autoregulation, it has some unique features. Vasomotion entails the coordinated activation of VSMCs whose oscillations in EM and [Ca2⫹]i evoke rhythmic contractions. Phasic cycles in Ca2⫹ signaling were mediated both by periodic changes in Ca2⫹ influx and in Ca2⫹ mobilization from intracellular stores (1, 979). One proposed sequence of events is that activation of either IP3 or RYRs/ channels on the sarcoplasmic reticulum mobilizes Ca2⫹ that activates the plasma membrane ClCa channels to allow Cl⫺ efflux that depolarizes the membrane. The resultant activation of L-type VOCC allows Ca2⫹ entry that increases [Ca2⫹]i further to promote MLC phosphorylation and cross-bridge formation leading to contraction (1, 123, 529, 531). Ca2⫹ sparks (i.e., highly localized intracellular Ca2⫹ transients generated by RyRs acting on the sarcoplasmic reticulum adjacent to the plasma membrane) also can elicit global Ca2⫹ waves and oscillations (845). An alternative view is that Ca2⫹ mobilization activates BKCa to cause hyperpolarization and reduce Ca2⫹ entry through L-type VOCC and thereby cause vasodilation. Subsequently, Ca2⫹ is taken up into the sarcoplasmic reticulum via Ca2⫹-ATPase to restore the basal tone. A third scenario derives from studies of isolated irideal arterioles whose vasomotion was independent of EM and the endothelium and is mediated by IP3 induced changes in [Ca2⫹]i (544). A periodic depolarization of VSMCs generates electrical signals that propagate along a vessel via cell-to-cell-coupling involving gap junction connections to cause synchronized oscillations in global Ca2⫹ signaling in the vasculature network (979). Oscillations are evoked by NE, KCl, or other agents that depolarize the EM (1333, 1349). They are dependent on gap junctions and Cxs, notably Cx37 (981). Rhythmic Ca2⫹ signaling of VSMCs may oscillate rapidly (⬍10 s) but can be dampened by neurotransmitters, vasoactive hormones, or local autocrine and paracrine factors such as NO. Release of cGMP in some vascular beds may facilitate Ca2⫹ activation of ClCa channels, perhaps via bestrophin 3, to reduce EM, thereby locking the gap junctions between VSMCs into electrical phase (979). 430 The renal afferent arteriole studied in vitro is viewed normally as a damped oscillator that does not exhibit spontaneous vasomotion. However, in vivo oscillations of superficial cortical blood flow are readily detected, as mentioned earlier in section IIA4B. Indeed, measurements of SNGBF on the kidney surface in vivo reveal two oscillations driven by the more rapid myogenic mechanism (0.15– 0.25 Hz) and the slower MD-TGF (0.02– 0.05 Hz) (289, 625, 629, 1340, 1400, 1636). MD-TGF-mediated oscillations in PPT have a period of 25–35 s. This slow process may oscillate because of the relatively long delay in filtered tubular fluid reaching the MD segment at the end of Henle’s loop to elicit a change in arteriolar tone (626, 627, 631, 892, 893). The faster myogenic oscillations observed in vivo with a period of 5–10 s represent a rhythmic oscillation of vascular tone and diameter of preglomerular vessels mediated by a coordinated and periodic fluctuation of intrinsic VSMC activity. Afferent arteriolar oscillations of ⬃0.2 Hz with amplitudes of ⬃1 m have been detected in these renal vessels (289). The cellular mechanisms for vasomotion of the preglomerular vasculature in vivo are thought to resemble those of the myogenic mechanism, but have been studied more extensively in extra-renal vessels, as reviewed below. Vasomotion was diminished in hamster skin by anesthetics (268, 869, 1087), primarily by alternating the amplitude of vasomotion in large arteries (653). The operating frequency of the myogenic mechanism in the renal vasculature of SD rats, based on transfer function gain, was 0.18 – 0.20 Hz under barbiturate or isoflorane anesthesia, but was reduced to ⬃0.14 Hz under halothane anesthesia (289). Stimulation of Ca2⫹ entry in VSMCs of the HNK preparation through L-type VOCC by BAY K 8644 induced large, 10-m oscillations of afferent arteriolar diameter with a period of ⬃20 s which were dependent on RyRmediated Ca2⫹ mobilization from the sarcoplasmic reticulum (1411, 1443). An increased Ca2⫹ release was associated with an increased and more rapid constriction which depended on Rho-kinase (1340). NE can stimulate afferent arteriolar vasomotion and cycling of [Ca2⫹]i in VSMCs (930, 1637). Pacemaker areas enriched in L-type VOCCs occur at branch points along the cortical radial arteries (477). The requirement of the endothelium for vasomotion and the influence of NO varied among vascular beds (1, 817). NO acted as a stimulant in some, whereas in others, endothelium-derived factors dampened or even abolished vasomotion by desynchronizing the Ca2⫹ signals in VSMCs. For example, vasomotion in mesenteric basilar or hamster cheek pouch arteries was stimulated by increased NO or by reduced KATP channel activity (1, 312) and abolished by endothelial removal or NOS inhibition (312, 530). NO and cGMP can modulate vasomotion at gap junctions to synchronize Ca2⫹ oscillations by inhibiting Ca2⫹ mobilization, Physiol Rev • VOL 95 • APRIL 2015 • www.prv.org CARLSTRÖM ET AL. Ca2⫹ entry, or Ca2⫹ sensitivity (1, 530). NOS inhibition abolished vasomotion in hamster cheek pouch arterioles by activating K⫹ channels. cGMP facilitated vasomotion in some (1, 477), but not other studies (312). Myogenic tone and vasomotion were diminished in small mesenteric arteries in mice lacking caveolae with deficient NO generation (1430). In contrast, NO suppressed vasomotion of the rat middle cerebral artery (336, 1645) independent of sGC but mediated by RyRs and VOCCs resulting in less synchronous Ca2⫹ oscillations and propagating Ca2⫹ waves (1645). TRPC1 channels in rat cerebral arteries generate oscillations in EM generated by Na⫹ entry following activation of PLC and Ca2⫹ entry through L-type VOCCs (1620). Endothelial dysfunction in the cerebral vasculature led to lowfrequency (4 –12 cpm) spontaneous oscillations of blood flow (635). Inhibition of oxidative stress caused oscillations in cerebral blood flow mediated by TxA2 and TP receptors (24, 635). The endothelium of the small cerebral basilar arteries modulated vasomotion by direct myoendothelial coupling by Cx37 and Cx40 that synchronized [Ca2⫹]i waves in adjacent VSMCs (542). Endothelial signaling by EDHF or TP receptors, but not by AT1 or adrenergic receptors, also elicited vasomotion in the hamster cheek pouch microcirculation (1532). Pial arteriolar vasomotion was modulated by somatosensory cortical neuronal activation (1536) and was blocked by NE or ACh (1239). NOS inhibition or dibutyryl cGMP failed to resynchronize [Ca2⫹]i waves, but K⫹ channel blockade depolarized constricted vessels and abolished vasomotion (542). Communication between endothelial cells was enhanced by gap junctions consisting of Cx37, Cx40, and Cx43, whereas VSMCs were more weakly coupled by Cx37. The two cell types were linked by myoendothelial gap junctions using Cx37 and Cx40. Electrotonic coupling of EM spread a hyperpolarization wave along endothelial and VSMCs accompanied by a propagated Ca2⫹ wave. The endothelium was essential for vasomotion in some (530, 651, 992, 1117, 1169), but not all (544, 868), studies of mesenteric arteries and may even be promoted by removal of the endothelium or inhibition of NOS (98, 970), which can desynchronize Ca2⫹ signaling in VSMCs (1346), PP regulated vasomotion and autoregulation. An increase in PP of mesenteric or cerebral arteries (528) reduced the amplitude but increased the frequency of rhythmic contractions, independent of the endothelium (529, 1135, 1525). This was dependent on RyR-mediated Ca2⫹ mobilization and Ca2⫹ entry through VOCCs (529, 1525). Moreover, cyclic variations in BP in vivo modulated the arterial diameter and VSMC [Ca2⫹]i (818, 819). Hamster cheek pouch arterioles had spontaneous vasomotion with oscillations of 20 mV in EM and 14 m in diameter, which were most frequent in the smallest arterioles (143, 268, 294). Isolated hamster cheek pouch and rat cerebral and mesenteric arterioles had both rate- and pressure-sensitive vasomotion (303, 1135, 1525). Small-amplitude, positive-pressure steps elicited monophasic constriction, whereas higher pressure steps elicited biphasic constrictions that were only partially dependent on a myogenic mechanism. Measurements of oscillations of blood flow have been used to study myogenic mechanisms in humans. Spectral analysis of microcirculatory peripheral blood flow in the skin of the hand and sternum by laser Doppler flowmetry showed two principal spectral rhythms of 0.04 – 0.15 Hz and ⬃0.17 Hz (907), which reflected myogenic activity (1262). Whereas the skin of the leg had more myogenic (0.05– 0.14 Hz) activity than the forearm at 33°C (622), this was reversed by warming to 42°C. Insulin increased the amplitude of the myogenic vasomotion in human skin, (1262), as in rat skeletal muscle (1085). A large-amplitude ⬃0.1 Hz oscillation in cerebral cortical perfusion was observed in conscious humans in association with wave-like propagation and oscillations in the diameter of pial arterioles (1211). The lowfrequency oscillations (⬃0.1 Hz) of cerebral blood flow were reduced in the aged (1533). In summary, a key component of vasomotion is spontaneous oscillation of EM that is generally mediated by either ClCa or BKCa channels to modulate Ca2⫹ entry through L-type VOCCs. This is followed by a prominent role for RyRs in triggering Ca2⫹ mobilization. The contribution of voltage-dependent channels and the endothelium varies among vessels (543). Cell coupling by gap junctions enhances synchronization of oscillations in Ca2⫹ release among adjacent cells, at least in isolated vessels in vitro. The speed of the myogenic mechanism varies by vascular bed. It is generally most rapid in the renal microcirculation (0.2 Hz), followed by the cerebral and mesenteric vascular beds (⬃0.15 Hz), with slower vasomotion in the cutaneous network (0.05– 0.15 Hz). Smaller arteries and arterioles have higher frequency oscillation than large vessels. Recent studies of oscillation in blood flow have given promise of a new technique to study myogenic mechanisms in the human circulation. III. MECHANOSENSITIVE MECHANISM INITIATING THE MYOGENIC RESPONSE Mechanotransduction implies an integrated interaction of the extracellular environment with the three-dimensional structure of the vessel wall that links changes in membrane stretch to intracellular signaling mechanisms (678, 899). Pressure-induced deformation of extracellular matrix proteins and their cell surface receptors such as integrins are Physiol Rev • VOL 95 • APRIL 2015 • www.prv.org 431 RENAL AUTOREGULATORY MECHANISMS thought to initiate contraction and cytoskeletal remodeling through modulation of ion channels, membrane depolarization, increased [Ca2⫹]i, and actomyosin crossbridge cycling (304, 617). Thus integrin interaction with the extracellular matrix transduced extracellular mechanical forces such as shear and tension into intracellular biochemical signals to affect VSMC contractility (64, 213, 304). Myogenic constriction of VSMCs resulted from activation of VOCCs secondary to reduced EM (301, 562, 810). Activation of mechanosensitive ion channels initiated pressureinduced depolarization and subsequent Ca2⫹ signaling and myogenic constriction. was initiated by localized RyR-dependent Ca2⫹ sparks that amplified global [Ca2⫹]i waves (214). Blockade of RyRs inhibited integrin-mediated myogenic constriction of afferent arterioles (FIGURE 8) (64). RyR-dependent Ca2⫹ signaling was independent of Ca2⫹ entry via L-type VOCCs in the renal (214), but not in nonrenal vessels (525). Ca2⫹-induced Ca2⫹ release in the afferent arteriole may be more critical in the initiation of the myogenic response than in the sustained phase when Ca2⫹ entry through VOCCs is essential. Mechanical force that perturbed the interactions between fibronectin and ␣51 integrins prolonged the oscillatory Ca2⫹ signals and the tractional force developed by renal VSMCs (65). A. Integrins How integrins detect mechanical distortion is poorly understood. The mechanosensitive myogenic response of cerebral arteries involves cytoskeleton remodeling and actin polymerization (262). Changes in the intraluminal pressure of arterioles may cause conformational changes of the extracellular matrix and fibronectin that expose binding sites for integrins, which trigger an intracellular signaling cascade. Alternatively, integrins may sense the distending force directly and signal across the extracellular-integrin-cytoskeleton axis. Disruption of the cytoskeleton can impair the myogenic response. Deletion of the serum response factor in endothelial cells or VSMCs of tail arteries markedly attenuated myogenic constriction and reduced actin polymerization, myosin light chain, and myosin light chain kinase (1234) while maintaining vasoconstriction produced by PE or U-46619. At present, the specific actions of particular integrins in the renal microcirculation are unclear, but they likely constitute an important component of arteriolar mechanosensation. Integrins are a diverse family of transmembrane heterodimeric proteins composed of one ␣- and one -subunit that link the extracellular matrix to the cytoskeleton of VSMCs. Deformation of integrins during changes in cell shape or stress conducts pressure-induced signals across the plasma membrane (304, 525, 614) to activate protein tyrosinephosphorylation cascades and mitogen-activated protein kinases (MAPKs) including extracellular receptor kinases (ERKs) (614). Although inhibition of ERK in rat cerebral arteries inhibited myogenic constriction and KCl-induced tone (857, 1600), the dynamics of PP-induced tyrosine phosphorylation in rat cremaster arterioles was considerably slower than the myogenic response, which argues against a causal relationship (1045, 1046). Both L-type VOCCs and BK channels in isolated VSMCs and cremaster arterioles were regulated by ␣51 integrins that interacted with fibronectin in the extracellular matrix to cause c-Srcmediated phosphorylation (305, 525). Thus both VOCCs that excite the myogenic response and BK channels that generally inhibit the response are under constitutive control by an interaction between ␣51 integrins and fibronectins. The balance of their activities could determine the degree of pressure-induced myogenic tone and perhaps vascular remodeling (525). Rat afferent arteriolar VSMCs expressed the ␣3-, ␣5-, ␣Vand the 1-, 3-integrin subunits (1638). A synthetic peptide with an arginine-glycine-aspartic acid (RGD) motif that interacts with integrin binding sites mobilized Ca2⫹ and contracted VSMCs (1638). In contrast, a different peptide with a RGD sequence reduced [Ca2⫹]i and dilated rat cremaster arterioles (291). Thus there is an extensive diversity and regional specificity among this group of proteins with other residues possibly playing a role. As discussed in the introduction, the embryological origins of VSMCs differ according to their vascular bed and thus may contribute to this diversity. Shear stress in preglomerular VSMCs was transduced by cell surface ␣51 integrins into cytoskeleton-dependent Ca2⫹ release from sarcoplasmic reticular stores (64). This 432 B. ENaCs The ENaC in the CT and CDs is a trimeric protein complex composed of ␣-, -, and ␥-subunits, which facilitates the Na⫹ reabsorption that initiates CT-GF. A vascular -ENaC has been proposed to function as a pressure- or stretchsensitive mechanosensor by forming an ion channel with degenerin (DEG) that is a closely related neuronal acidsensing ion channel (ASIC) that is tethered to the extracellular matrix and cytoskeletal proteins (343, 344, 346, 1431). The non-voltage-gated -ENaC-DEG complex may permit Na⫹ entry in response to increased intraluminal pressure and thereby cause depolarization of VSMC to initiate the myogenic response (FIGURE 8) (728). Stretch activation of -ENaC in mouse freshly isolated renal VSMCs increased Na⫹ entry (258), whereas in the CD, shear stress gated and opened ENaCs (1356). Rat cerebral arterial VSMCs expressed - and ␥-ENaC subunits (344). Most, but not all, of the myogenic constrictor tone was inhibited by blockade of ENaC with amiloride (1 M) or benzamil (30 nM) (344). Likewise, amiloride, ben- Physiol Rev • VOL 95 • APRIL 2015 • www.prv.org CARLSTRÖM ET AL. PERFUSION PRESSURE VOCC ENaC +Deg GPCR PLC Membrane events TRPC Integrins DAG ADP ribosyl cyclase Em p47phox NAPDH oxidase cADPR [Ca2+]i GPCR RyR ROS PLC Cytosolic signaling Ca2+ induced IP3 R Ca2+ release IP3 DAG Rho / rho kinase PKC Actin / myosin crosslinking Myosin light chain kinase Myosin light chain phosphotase Contractile response MYOGENIC CONTRACTION FIGURE 8. An overview of the major pathways proposed to mediate or modulate the renal afferent arteriolar myogenic contractile response to an increase in perfusion pressure (PP). The mechanisms are grouped as membrane events, cytosolic signaling events, and contractile responses. VOCC, voltage-operated calcium channel; ENaC, epithelial Na⫹ channel; DEG, degenerin; GPCR, G protein-coupled receptor; TRPC, transient receptor potential channel; PLC phospholipase C; DAG, diacylglycerol; Em, membrane potential; cADPR, cyclic ADP-ribose; NADPH, nicotinamide adenine dinucleotide phosphate; [Ca2⫹]i, cytosolic concentration of ionized calcium; RyR, ryanodine receptor; ROS, reactive oxygen species; IP3R, inositol trisphosphate receptor; PKC, protein kinase C. For discussion, see text. zamil (1 M) or siRNA knockdown of - or ␥-ENaC subunits attenuated some of the myogenic constriction in rat posterior cerebral arteries (793). Myogenic constriction of middle cerebral arteries of ASIC2 knockout mice was very weak, whereas constrictor responses to high KCl and PE were normal, suggesting specificity to mechanical stimulation (454). DEG-like currents in cerebral arterial VSMCs were inhibited by ROS (257). -ENaC in rat posterior cerebral arteries colocalized with TRPM4 but not TRPC6 (793, 794), although all three channels participated in myogenic constriction. ENaC ␣- and ␥ -subunit expression in mesenteric arteries was reduced by high-salt diet, whereas -ENaC movement to the plasma membrane of VSMCs was increased (730). Benzamil (1 M) inhibited the myogenic response only in arteries from rats fed a high-salt diet (730). There is controversy about the presence of mRNA and protein for specific ENaC subunits in VSMCs of the cortical radial artery and afferent arteriole. Early studies of single VSMCs in the renal microcirculation, either freshly isolated or cultured from relatively large cortical radial arteries of rats or mice, reported expression of - and ␥-, but not ␣-, ENaC protein subunits (729). However, subsequent immunofluorescent studies in VSMC freshly isolated from cortical radial arteries and afferent arterioles revealed expression of ␣- as well as - and ␥-subunits of ENaC (522). The finding of the ␣-subunit poses a potential problem with ENaC serving as a mechanosensitive channel since it renders the ion channel constitutively active and thus not selectively responsive to a change in transmural pressure (522). On the other hand, another study failed to detect mRNAs for any ENaC subunits in VSMCs disassociated from these arterioles (1574), but all three subunits were Physiol Rev • VOL 95 • APRIL 2015 • www.prv.org 433 RENAL AUTOREGULATORY MECHANISMS evident in intact cortical radial arteries. This implicates expression of ENaC on endothelial or adventitial cells rather than on VSMCs, which would not be compatible with a mechanosensing function. Varying results have been reported for pharmacological blockade of ENaC channels depending on the concentrations of antagonists. Apparent blockade of ENaC activity with 5 M amiloride or benzamil or genetic knockdown of either - or ␥-ENaC subunits attenuated myogenic constriction of cortical radial arteries (344, 728, 729). Moreover, ANG II infusion reduced the immunoreactive expression of - and ␥-ENaC in rat interlobar arteries and reduced myogenic contraction (731). However, the majority of the myogenic response and the regulation of RVR in vivo occur in smaller, more distal cortical radial arteries and afferent arterioles rather than in the relatively large interlobar arteries (201, 611, 1071). Nevertheless, recent studies have reported an impaired myogenic constriction of afferent arterioles isolated from mice with genetically reduced -ENaC expression (462). A mouse model of genetically reduced (⬃50%) -ENaC expression had a 50% reduction in the myogenic component of RBF autoregulation in the 0- to 5-s time window, yet a well-maintained overall steady-state autoregulation (505). This likely implies that the MD-TGF contribution was upregulated since inhibition of MD-TGF by acute volume expansion revealed a 75% reduction in the whole kidney myogenic response in mice with genetic reductions in -ENaC expression and almost complete loss of autoregulation over 30 s after a step change in PP (462). Recent studies by Drummond and associates using VSMCs freshly isolated from the mouse afferent arteriole have shown less frequent and smaller stretch-activated Na⫹ currents in the VSMCs from mice with reduced -ENaC expression, but preserved VOCC and BK channel activity (258, 344). Thus -ENaC appears to be required for normal mechanically gated currents in renal VSMCs. Their disruption reduces myogenic constriction of afferent arterioles. Cortical radial arteries and afferent arterioles isolated from -ENaC knockdown mice have a weakened myogenic tone, but constricted normally to PE and KCl (462), which implies that ENaC is upstream from myogenic contractions. Mice with partial gene knockdown of -ENaC became hypertensive and developed renal inflammation and damage that may be related to reduced renal autoregulatory capacity (345). The myogenic response of the rat perfused JMN preparation after papillectomy to eliminate MD-TGF was blunted by blockade of ENaC with 10 M amiloride or benzamil added to the bath or 5 M amiloride added to the luminal perfusate (522). Similarly, relatively high concentrations of inhibitors of ENaC (amiloride and benzamil, 10 M) 434 blocked more than two-thirds of the autoregulatory response of the afferent arteriole, whereas vasoconstriction produced by ATP or 20-HETE was unaffected (1051). The authors concluded that ENaC is an essential component of myogenic responses. Different conclusions were reported in a recent study with the HNK preparation where the myogenic response or the EM was unaltered by relatively low concentrations (ⱕ1–3 M) of amiloride or benzamil which inhibited ⬎80% ENaC activity in renal tubules (926, 1574). However, higher concentrations (ⱖ5 M), similar to those used in the earlier studies, were confirmed to inhibit myogenic constriction, but this was ascribed to nonspecific inhibition of Na⫹ transporters and ion channels other than ENaC (926). Moreover, myogenic constriction in the rat HNK preparation was independent of extracellular [Na⫹] (1574). Benzamil (ⱖ1 M), which also inhibits the NCX exchanger, depolarized and potentiated, rather than inhibited, the myogenic constriction of the afferent arteriole in this study (1574). However, another study of afferent arterioles of the HNK reported that lowering extracellular [Na⫹] led to a slowly developing attenuation of myogenic constriction, with abolition of the myogenic response when [Na⫹] was below 50 mM (1446). Aldosterone regulates ENaC expression and activity in the distal nephron, but less is known about its role in the regulation of ENaC in VSMCs and endothelium. Elevated aldosterone levels are causally linked to vascular fibrosis, inflammation, and remodeling (152, 816). Stretch of rat cultured aortic VSMCs increased nicotinamide adenine dinucleotide phosphate (NADPH) oxidase activity that was dependent on aldosterone and mineralocorticoids (1114). Aged mice lacking mineralocorticoid receptors in their VSMCs were hypotensive and had a 40% reduced myogenic tone in their mesenteric arteries, weaker TP receptorinduced contraction, and an 80% reduction in the expression and activity of L-type VOCC, whereas BKCa function was normal (994). Aldosterone rapidly (⬍5 min) contracted rabbit isolated afferent and efferent arterioles via nongenomic mechanisms mediated by L- and/or T-type VOCC (53, 54). ENaC is also located in the vascular endothelium of mesenteric arteries where it inhibited NO production (851), which might be another pathway whereby ENaC modulates the myogenic mechanism. In summary, genetic studies with knockdown of -ENaC in the preglomerular vasculature have reported an attenuated myogenic response, implicating -ENaC as a critical mechanosensor. However, the pharmacological evidence for a role of ENaC is more controversial because of lack of specificity of the antagonists at the concentrations tested. Integrins could function as force transducers linked to Physiol Rev • VOL 95 • APRIL 2015 • www.prv.org CARLSTRÖM ET AL. ENaC as part of a mechanosensory complex that couples the extracellular matrix to the cytoskeleton. In turn, ENaC may be linked directly or indirectly to TRP channels. Further investigation is required to clarify whether and how ENaC subunits function as mechanosensors in the renal microcirculation. C. Transient Receptor Potential Channels and Stretch-Activated G Protein-Coupled Receptors Transient receptor potential (TRP) channels are a family of voltage-insensitive, homo- or heterotetrameric proteins that function either as nonselective cation channels allowing entry of Ca2⫹ and Na⫹ to depolarize the EM or as highly selective Ca2⫹ channels to directly increase [Ca2⫹]i (FIGURE 8) (330, 366, 368, 681, 1352). Either mechanism could cause vasoconstriction. TRPC1, TRPC3, and TRPC6 are abundant in VSMCs. TRP channels resided in signaling complexes in caveolar lipid raft domains in the plasma membrane (23). Mechano-gating of TRPC1 and TRPC6 can be transduced through scaffolding proteins and actin filaments of the cytoskeleton. The mRNAs and proteins for multiple canonical (C) TRPs have been identified in rat renal resistance vessels and glomeruli. The mRNAs for TRPC1, 3, 4, 5, and 6 were expressed in renal cortical radial arteries and afferent arterioles (387), and the proteins for TRPC1, 3, 5, and 6 have been detected in glomeruli and preglomerular vessels (387, 1278). TRPC1 was expressed both in the afferent and efferent arterioles (1448). Immunoreactive TRPC1, 3, and 6 were expressed on rat afferent arteriolar VSMCs and TRPC3 on cortical radial arteries (1278). Mechanical stretch of the plasma membrane directly activated TRPC6, presumably via a conformational change to open the channel, independent of GPCR ligand binding or PLC activation, in HEK293 and CHO cells overexpressing the ion channel (1403). DAG generation after GPCR activation can engage TRPC3 and TRPC6 channels in VSMC throughout the vasculature (623, 679). In cerebral vessels, TRPC6 and TRP melastatin (M) 4 and vanilloid (V) channels transduce myogenic responses (141, 366, 371, 681, 1352, 1403, 1594). Cerebral arterial VSMCs also express TRPC3 that is activated by GPCR agonists such as UTP but not by pressure, suggesting coupling of TRPC3 and TRPC6 to distinct excitatory stimuli (1212). TRPC1 was present in VSMC of cerebral arteries, but it is not clear if it has a mechanosensitive role in the vasculature (331). These findings led to the concept that membrane stretch activates a TRPC6-dependent constriction of cerebral arteries. A proposed pathway is that membrane stretch activates PLC to produce DAG, which directly activates TRPC6 to mediate depolarizing cation current to trigger a myogenic response (679). Thus pressure-induced depolarization of cerebral artery myocytes was blocked by PLC inhibition (1383), and mechanosensitive channels in VSMCs resembling TRPC channels were inhibited by PLC blockade and activated by a DAG analog (1154). However, mechanosensitive channel activity and myogenic responses of cerebral arteries were not affected by deletion of TRPC1 (331) or TRPC6 (332, 490). 20-HETE potentiated the activity of TRPC6 channels of the mesenteric artery, thereby enhancing myogenic constriction (680). TRPC6 on the glomerular podocyte slit diaphragm functioned as a mechanosensor coupled to Ca2⫹-activated potassium (KCa) channels (348). Stretch activation of cation entry in the podocyte was independent of PLC or PLA2 activity (25). Gain-of-function mutations of TRPC6 caused podocyte dysfunction and focal and segmental glomerulosclerosis (FSGS) (372, 837), whereas deletion of TRPC5 protected mice from albuminuria (1290). IP3 constricted cerebral arteries via TRPC3 channel activation leading to Na⫹ influx and membrane depolarization, independent of sarcoplasmic reticular Ca2⫹ release (1629). A similar mechanism operated in rat mesenteric arteries where IP3 coupling of TRPC3 channels to caveolae resulted in Na⫹ entry (10). TRPM4 in cerebral artery myocytes was a highly selective monovalent cation channel activated by stretch or Ca2⫹ entry through TRPC6 channels and/or Ca2⫹ mobilization mediated by RyRs (370, 1032). Other studies in cerebral arteries reported that transmural pressure activated a signaling cascade involving activation of Src tyrosine kinase and PLC with subsequent IP3 generation that mobilized Ca2⫹ from the sarcoplasmic reticulum (480 – 482). The resultant increase in [Ca2⫹]i in turn activated plasma membrane TRPM4 that increased Na⫹ entry and depolarized the plasma membrane to allow Ca2⫹ entry and contraction. The Ca2⫹ activation of TPRM4 may be enhanced by Ca2⫹ entry through TRPC6 channels. In contrast, TRPV4 in mesenteric arteries acted as a nonselective cation channel activated by stretch or EETs to cause activation of K⫹ channels that hyperpolarized the plasma membrane and relaxed the vessel (367). A provocative observation was that hyposmotic cell swelling and stretch-activated G protein-coupled receptors (GPCRs) that caused a conformational change independent of ligand binding which conferred a mechanosensitive action on TRPC6 in heterologous systems overexpressing GPRC and TRPC6 (1403). GPCR activation in VSMCs linked to Gq/11-dependent DAG generation can engage TRPC6 channels in VSMCs independent of PKC (FIGURE 8) (1143). Subsequent studies demonstrated that stretch activation of GPCRs stimulated cation flux through TRPC3 and TRPC6 via PLC-dependent DAG formation (1000). Diverse GP- Physiol Rev • VOL 95 • APRIL 2015 • www.prv.org 435 RENAL AUTOREGULATORY MECHANISMS CRs may activate this pathway including ANG II type 1 (AT1), endothelin type A (ETA), vasopressin type 1A (V1A), and muscarinic type 5 (M5) receptors (1000, 1673). In the absence of ANG II, certain antagonists of AT1 receptors, such as losartan or candesartan, prevented stretch-induced myogenic tone of cerebral and renal cortical radial arterial VSMCs via reductions in ERK and ROS (999, 1000). The myogenic tone of cerebral arteries was enhanced by activation of GPCRs linked to PLC-sensitive G proteins (1137), whereas TRPC3 in cerebral arteries was activated by IP3, independent of Ca2⫹ release from the sarcoplasmic reticulum (1629). Collectively, these data suggest that membrane stretch causes an agonist-independent conformational change in the GPCR, which activates PLC and downstream G␣q/ll signaling to cause DAG activation of TRPC6, resulting in Na⫹ and/or Ca2⫹ entry, membrane depolarization, and a myogenic response (1000, 1421). A recent study extends these findings to individual mesenteric arteries and isolated kidneys. This demonstrated a requirement for AT1A receptors for myogenic responses in the absence of its native ligand (1294). However, the downstream signaling relies on an ion channel distinct from TRPC6 or a Kv channel distinct from KCNQ3, 4, or 5 to activate VSMCs and elevate vascular resistance. Renal function studies raise some questions about the role of GPCR in the myogenic response. Although ANG II may potentiate the renal myogenic response, as discussed in section VIA, activation of other GPCR by natural agonists such as adrenoceptors by catecholamines (760, 1193, 1570), ETA receptors by endothelin (1357), or V1A receptors by AVP (1563) do not normally impact the renal myogenic response. Frequency analysis in the kidney generally has found that inhibition of ANG II formation (597) or AT1 receptor antagonism (329) has little effect on the myogenic mechanism, although a recent study reported that prolonged ANG II administration potentiated the renal myogenic response (1193). In other studies, blockade of AT1 receptors with losartan or candesartan did not inhibit myogenic tone in rat mesenteric arteries (519, 985) and only blunted myogenic constriction of cerebral arteries weakly (1000). G␣q/ll-coupled receptor activation in cerebral arteries by uridine triphosphate or thromboxane prostanoid (TP) receptor activation by U-46619 did not enhance mechanosensitivity of TRPC-like currents or amplify myogenic responsiveness (36). Increasing the PP of rat isolated mesenteric arteries activated ERK1/2 that was inhibited by an antagonist of AT1 but not AT2 receptors (383, 984). The myogenic response was also blocked during ACE inhibition, suggesting dependence on ANG II formation. At present, limited data have implicated an agonist-independent, stretch-activated GPCR pathway in TRP channel 436 activation of the myogenic contractile response in renal microvessels. Thus myogenic tone of renal arteries was diminished by the AT1 receptor antagonist losartan independent of ANG II (1000). Cortical radial arteries from regulator G protein signaling 2 knockout mice had enhanced myogenic tone in addition to the expected exaggerated vasoconstriction to ANG II, ET-1, and PE (604). Activation of TRPC channels in the afferent arteriole enhanced Ca2⫹ signaling by ANG II (402) and NE where they functioned as store-operated cation channels (SOC) independent of L-type VOCCs (387, 1278). The myogenic response of cortical radial arteries and afferent arterioles of the HNK depended on ill-defined mechanosensitive cation channels (1446, 1447). GPCRs can activate redox signaling in afferent arteriolar VSMCs (399, 400, 1606) where TRP channels themselves are redox sensitive because of oxidative modifications of specific cysteine residues (507, 1005, 1433). VOCCs also were redox regulated (126). NO activated TRPC channels via cysteine S-nitrosylation (1639). The polycystins represent a special class of TRP cation channels. VSMC TRP polycystin-1 and -2 (TRPP1 and -2) cation channel proteins modulated the myogenic response in mesenteric arteries (1057). Deletion of TRPP1 in VSMCs reduced stretch-activated channel activity and myogenic tone (1353), but depletion of TRPP2 rescued both stretchactivated channel opening and the myogenic response, indicating extensive interaction. TRPP2 interacted with the cytoskeleton and an actin crosslinking protein that was central for stretch-activated channel regulation. In summary, cation entry through TRP channels subserves two functions in VSMC: to relate changes in transmural pressure or stretch to depolarization of the plasma membrane and to respond to GPCR stimulation and PLC activation to modulate SOC function. TRP channels are attractive candidates as mechanosensors, but their linkage to GPCRs remains an intriguing, but uncertain, possibility. The specific roles of particular TRP channels acting as mechanosensors initiating membrane depolarization and increases in [Ca2⫹]i to trigger the myogenic response of afferent arterioles requires further clarification. Integrins, signaling through the cytoskeleton and possible TRP channels are attractive candidates, but are incompletely studied in the renal microcirculation. The role of stretch-activated ENaC channels is controversial and still evolving. IV. CONDUCTANCE CHANGES MEDIATING MYOGENIC CONTRACTION A. Chloride Channels The intracellular [Cl⫺] in VSMCs is high due to accumulation by a Cl⫺/HCO3⫺ exchanger and the NKCC1 cotransporter. Because the [Cl⫺] in VSMCs is above its electro- Physiol Rev • VOL 95 • APRIL 2015 • www.prv.org CARLSTRÖM ET AL. chemical equilibrium, activation of plasma membrane Cl⫺ channels permits Cl⫺ efflux, leading to depolarization of the plasma membrane, with subsequent opening of VOCCs to promote Ca2⫹ entry (489, 1324, 1442). However, the resting Cl⫺ permeability of VSMCs (621), arcuate arteries (1200), and renal mesangial cells (1118) is sufficiently low that the EM is unaffected by removal of extracellular Cl⫺. Likewise, the basal tone of rabbit perfused afferent arterioles is unchanged in Cl⫺-free media (726). These results indicate that there is little basal Cl⫺ conductance in afferent arterioles. On the other hand, VSMCs isolated from arcuate or cortical radial arteries display a Ca2⫹-activated Cl⫺ conductance (ClCa) that depolarizes the plasma membrane after GPCR activation and leads to Ca2⫹ mobilization (489, 664). ClCa channels in afferent arterioles contribute to membrane depolarization (559), Ca2⫹ entry (1324), GPCR activation (726, 1442), and increased [Ca2⫹]i produced by ANG II or ET-1 (451, 727, 1324, 1435). GPCR activates Cl⫺ channels in mesangial cells to depolarize the plasma membrane and enhance [Ca2⫹]i signaling (834, 987). Activated Cl⫺ channels also can enhance myogenic tone by providing a Cl⫺ influx pathway as a charge compensation for influx of Ca2⫹ during activation of VOCCs (559, 726). Indeed, the addition of extracellular Cl⫺ enhanced high K⫹-induced depolarization and VOCC-dependent contractions of rabbit afferent arterioles with an EC50 of 82 mM (559), that is close to [Cl⫺] in extracellular fluid, while removal of extracellular Cl⫺ attenuated Ca2⫹ influx via VOCCs in both mesangial cells (834) and rat aortic VSMCs (1650). Although Cl⫺ channels are important in vasoconstriction of afferent arterioles produced by ANG II and NE (559, 726, 1407), neither blockade of Cl⫺ channels nor reducing extracellular [Cl⫺] inhibits myogenic responses of the HNK preparation (1435, 1442). Thus ClCa regulated VOCCs during stimulation with GPCR agonists, but these channels do not appear to be required for the myogenic response of afferent arterioles (726, 1442). The electroneutral NKCC1 is present in VSMCs throughout the vascular system, in contrast to the NKCC2 isoform that is expressed exclusively in the kidney in the TAL of Henle’s loop and MD cells. NKCC1 may regulate myogenic tone of arteries and arterioles. Ion entry via NKCC1 increased intracellular [Cl⫺] that caused membrane depolarization and thereby Ca2⫹ entry through VOCCs and vasoconstriction (1126). Indeed, afferent arteriolar myocytes expressed NKCC1 but not NKCC2 (1568), and inhibition of this vascular NKCC1 in the mouse isolated and perfused afferent arteriole by furosemide or bumetanide caused vasodilation (1124) and inhibited most of the myogenic response of the afferent arteriole in the rat HNK preparation lacking tubules (1568). Inhibition of NKCC1 by bumetanide decreased myogenic tone in mesenteric arteries as well as contraction by high KCl, PE, and UTP (820). However, as discussed in section IVC, such pronounced inhibition of myogenic tone has not been observed during administration of furosemide or bumetanide in vivo. RBF in vivo is usually unchanged or slightly reduced during furosemide-induced natriuresis (749 –751, 753, 760, 1124, 1183, 1489), although increased RBF has been observed in some cases (170, 351, 760). Tubular hydrostatic pressure and interstitial hydrostatic pressure was increased during furosemide-induced natriuresis (170, 1489), which may contribute to a fall in GFR. Molecular candidates for Cl⫺ channels are transmembrane protein 16A (TMEM16A) or anoctamin 1 (ANO1), bestrophins, as well as the classic ClC-3 (1367). Both “classic” and cGMP-dependent ClCa channels are expressed in VSMCs. Bestrophin-3 is reported as a cGMP-dependent Ca2⫹-activated Cl⫺ conductance and ClC-3, a voltagegated channel (980, 982). Membrane stretch in cerebral arteries activated TMEM16A channels in VSMCs and caused membrane depolarization and the myogenic response via cation entry through mechanosensitive nonselective cation channels, with Ca2⫹ activation of TMEM16A facilitating Cl⫺ efflux (165). Disruption of TMEM16A (ANO1) and thereby preventions of Ca2⫹-activated Cl⫺ currents relaxed VSMC of aorta and carotid arteries (599). Blockade of Cl⫺ channels in pressurized rat cerebral arteries caused hyperpolarization (⫺10 to ⫺15 mV) that inhibited myogenic tone (1080). Bestrophin-3 was important for Ca2⫹ oscillations and synchronization of VSMC during vasomotion in the rat mesenteric small arteries in vivo (153). siRNA knockdown of bestrophins in rat mesenteric arteries did not affect arterial contraction but inhibited the rhythmic contractions, vasomotion (295). Downregulation of TMEM16A also reduced expression of bestrophins and suppressed vasomotion and contractions produced by GPCR agonists (NE, AVP) and KCl, suggesting this ClCa channel participates in depolarization of VSMC. Thus Cl⫺ channels are involved in the regulation of the EM of VSMC and vasoconstriction of mesenteric arteries and myogenic tone of cerebral arteries. Cl⫺ channels in the renal vasculature participate in vasoconstriction by GPCR agonists, but apparently not the myogenic response. B. Potassium Channels The resting EM of VSMCs of pressurized afferent arterioles is approximately ⫺40 mV. This is ascribed largely to a K⫹ diffusion potential. The EM was near the threshold for activation of L-type VOCCs (847, 928). Opening K⫹ channels Physiol Rev • VOL 95 • APRIL 2015 • www.prv.org 437 RENAL AUTOREGULATORY MECHANISMS and potentiating K⫹ efflux from VSMCs elicits vasodilation primarily by hyperpolarization that inactivates L-type VOCCs and reduces Ca2⫹ entry. The large-conductance, Ca2⫹-activated K⫹ channel (BKCa) in the cerebral circulation was a major determinant of the VSMC EM (1081). The BKCa channel has been implicated in several steps of the myogenic response, including channel closure causing membrane depolarization, and channel opening causing hyperpolarization to oppose excessive pressure-induced vasoconstriction. A special case in cerebral arteries is spontaneous or pressure-induced Ca2⫹ sparks representing Ca2⫹ release from RyR channels in the sarcoplasmic reticulum, which can activate BKCa to hyperpolarize the plasma membrane, limit Ca2⫹ entry via VOCCs, and diminish myogenic tone. Accordingly, inhibition of BKCa in many nonrenal vascular beds has a variable effect, but often depolarizes the EM and enhances stretch-induced depolarization, Ca2⫹ entry, and contraction (720, 1595). Caveolae in VSMC of cerebral arteries can modulate the myogenic response. VSMC of mice lacking caveolin-1 responded to an increased pressure with a diminished depolarization, an increase in [Ca2⫹]i, and myogenic response (11) via activation of BKCa channels and hyperpolarization. NOS inhibition did not restore myogenic tone in arteries deficient in caveolin-1, arguing against excess NO production as the cause for inhibition of the myogenic response. Genetic ablation of caveolin-1 in murine cerebral artery VSMCs increased Ca2⫹ sparks and the frequency of transient BKCa currents without altering Ca2⫹ entry via VOCC (233). Stretch of VSMCs in coronary arteries/arterioles and mesenteric arteries stimulated an outward K⫹ current through BKCa, whereas blockade of BKCa channels enhanced stretch-induced depolarization (338, 1628). Inhibition of BKCa channels increased myogenic contraction of pressurized mouse mesenteric arteries, presumably by favoring membrane depolarization and Ca2⫹ entry through VOCCs (836). VSMCs exhibit heterogeneity in BKCa activity or their sensitivity that may contribute to tissue-specific differences in the regulation of myogenic vasoconstriction (525, 618, 1632). For example, the BKCa is more sensitive to increased [Ca2⫹]i resulting from Ca2⫹ sparks generated by Ca2⫹ release from sarcoplasmic reticulum in cerebral than skeletal muscle cremaster arteries. The resultant vasodilation attenuated myogenic vasoconstriction in cerebral arteries (1632). The variation in sensitivity might be explained by the number of 1 subunits of the BKCa channel (1632). The extent to which BKCa are open during basal conditions and modulate basal EM in the renal vasculature is not clear. BKCa channels are present on VSMCs of preglomerular ar- 438 teries and arterioles (390, 467, 949, 1200), but usually are closed under basal conditions. Thus their blockade did not modify resting RBF or renal vasoconstriction elicited by ANG II or NE (949). Moreover, BKCa channels in pressurized afferent arterioles of JMN also were quiescent and did not buffer vasoconstriction by ANG II (390, 1200). ACh caused dose-dependent renal vasodilatation in isolated perfused rat kidneys, which was blunted by NOS inhibition but not by inhibition of sGC. The vasodilation was mediated by KCa3.1 channels sensitive to charbdotoxin which is characteristic of BKCa channels (1370). On the other hand, an increase in PP of renal arterioles was reported to increase 20-HETE generation from arachidonate, which closed BKCa channels and thereby caused membrane depolarization and vasoconstriction (672, 943, 1667). Blockade of 20-HETE production inhibited myogenic responses of arcuate arteries and impaired RBF autoregulation (464, 777, 1668, 1669). Voltage-gated and inward-rectifier K⫹ channels (KV and KIR) in afferent arterioles can regulate basal vascular tone (936, 1484). KIR 2.1 in the afferent arteriole contributed to K⫹ currents and EM of VSMCs (238). Enhanced rectification of KIR in the distal cortical radial artery and afferent arteriole hyperpolarized the plasma membrane and attenuated myogenic tone (239, 240). Stimulation of PKC by ANG II or ET-1 potentiated the myogenic response of afferent arterioles of the HNK preparation by inhibition of delayed rectifier K⫹ channels which depolarizes the plasma membrane (802). Activation of ATP-dependent K⫹ channels (KATP) in JMN had no effect on basal afferent arteriolar tone (665), but blunted PE-induced constriction of rabbit afferent arterioles (924). Activation of these channels by calcitonin generelated peptide dilated the afferent arteriole in the HNK preparation (1232) by causing hyperpolarization of the plasma membrane that inhibited Ca2⫹ entry through VOCCs (1231). Moreover, activation of KATP by pharmacological agents or hypoxia hyperpolarized the plasma membrane and inhibited myogenic responses of afferent arterioles (936, 1232). However, KATP channels in the dog did not affect the MD-TGF response (782) or the autoregulation of coronary blood flow (1060). A recent study of the renal vasculature in rats in vivo demonstrated that pharmacological blockade of individual K⫹ channels (e.g., BKCa, Kir, KATP, Kv) had little effect on large whole kidney RVR responses to GPCR agonists such as ANG II or NE and their activation of VOCCs (1396). However, concurrent closure of multiple types of K⫹ channels effectively attenuated agonist-induced renal vasoconstriction, demonstrating considerable redundancy among the K⫹ channels in the renal microcirculation. Although voltage-dependent K⫹ channels were tonically active in coro- Physiol Rev • VOL 95 • APRIL 2015 • www.prv.org CARLSTRÖM ET AL. nary arteries, they were not required in vivo for autoregulation (99). KCNQ (Kv7) K⫹ channels in cerebral arteries were inhibited by membrane stretch and thereby could modulate myogenic vasoconstriction (1663), although genetic deletion of KCNQ3, 4, or 5 did produce constriction of mouse mesenteric arteries or isolated perfused kidneys (1294). The myogenic tone of skeletal muscle arterioles results from an interaction between Ca2⫹ entry through L-type VOCC secondary to pressure-induced membrane depolarization and CICR and Ca2⫹ mobilization mediated by RyR. BKCa modulation of EM played only a small role in limiting myogenic vasoconstriction in this vascular bed (827). However, inhibition of RyRs in mouse pressurized mesenteric arteries increased myogenic constriction, presumably by reducing BKCa activity to favor membrane depolarization and Ca2⫹ entry through VOCCs (836). In summary, most evidence suggests that BKCa channels are normally quiescent in the renal microcirculation and do not respond to changes in RPP or limit the myogenic response, in contrast to their central role in the cerebral circulation. However, 20-HETE is reported to enhance myogenic constriction by inhibiting BKCa activity in some situations. Activation of other K⫹ channels that hyperpolarize the EM also can inhibit myogenic responsiveness under specific experimental conditions such as hypoxia, whereas inhibition of BKCa channels in some settings can depolarize VSMCs and favor vasoconstriction. MD-TGF responses depend on ROMK channels in MD that mediate K⫹ recycling after reabsorption by NKCC-2 (1312). C. Sodium Channels Evidence relating to the proposal that ENaC subunits are expressed on VSMCs of preglomerular vasculature and function as mechanosensors in the myogenic response was discussed in section IIIB. The VSMC plasma membrane Na⫹/Ca2⫹ exchanger (NCX1) normally promotes Ca2⫹ entry and Na⫹ extrusion (1655), in contrast to the heart where NCX1 primarily mediates Ca2⫹ extrusion (327). Normally, three Na⫹ are exchanged for one Ca2⫹ (119). NCX contributes to the myogenic response by favoring Ca2⫹ entry both in nonrenal vessels and also in the renal circulation where it enhances vasoconstrictor responses. Ca2⫹ entry via NCX1 is implicated in pressure-induced myogenic constriction in mouse mesenteric and hindlimb arteries (713, 1655), rat cremaster first-order arterioles (150 m) (1206), and rat posterior cerebral arteries (771). Reducing extracellular Na⫹ around isolated cremaster muscle arterioles promoted NCX1 activity and Ca2⫹ entry to enhance myogenic vasoconstriction (1206), whereas decreasing the activity of the VSMC NCX impaired arteriolar myogenic reactivity (1206). Knockout of NCX1 in VSMCs reduced vasoconstriction elicited by PE, high KCl, PP, or L-type Ca2⫹ channel currents in mouse mesenteric arteries and lowered BP (1655). Myogenic and ANG II responses of mesenteric arteries were blunted in mutant mice lacking the NCX1 in VSMCs (1655, 1660) accompanied by attenuated Ca2⫹ entry and myogenic responses (1010, 1655). A PKC regulated NCX1 in the afferent arteriole usually operates in the “reverse” mode with Ca2⫹ entering VSMCs down its concentration gradient to drive Na⫹ efflux and promotes vasoconstriction (120, 430, 1079). Reducing extracellular [Na⫹] in the isolated perfused kidney promoted Ca2⫹ entry into afferent arteriolar VSMCs via NCX to increase [Ca2⫹]i (1078, 1079) and RVR (1335). Inhibition or genetic deletion of NCX1 reduced the afferent arteriolar [Ca2⫹]i responses and the vasoconstriction to ANG II (402, 1660). The expressions of NCX1, sarcoplasmic reticular Ca2⫹ATPase-2 (SERCA2), and TRPC6 in VSMCs are inversely related to the activity of the ␣2-subunit of Na⫹-K⫹-ATPase (120), whose reductions in mesenteric arterial VSMCs have been linked to increased Ca2⫹ signaling and augmented myogenic- and PE-induced vasoconstriction mediated by IP3 release of Ca2⫹ from the sarcoplasmic reticulum and Ca2⫹ entry through SOC channels (119). Mesenteric arteries expressing 50% of the normal amount of the ␣2-subunit of the Na⫹ pump had augmented myogenic contraction (1654). VSMC NCX1, SERCA2, and TRPC6 were overexpressed in fourth-order mesenteric arteries of Milan genetically hypertensive rats which had exaggerated myogenic responses (905). However, NCX can operate in the “forward” mode in VSMCs, thereby mediating Na⫹ influx in exchange for Ca2⫹ efflux, for example, in the vasculature of the isolatedperfused kidney (1335) or skeletal muscle arteries where blockade of NCX enhanced myogenic tone, presumably by reducing Ca2⫹ extrusion (634). NCX activity in VSMC can be inhibited by oxidative stress that also increased [Ca2⫹]i (1505). Thus Na⫹/Ca2⫹ exchange via NCX1 has variable effects depending on whether it functions in the forward or reverse mode. It typically functions in the reverse mode in renal afferent arteriolar VSMCs to facilitate Ca2⫹ entry down its concentration gradient, thereby expelling Na⫹ and promoting myogenic contractions. Its activity is linked to Na⫹-K⫹ATPase via change in the ␣2 subunit which can account for the enhanced myogenic tone, and perhaps some of the hypertension, in the Milan genetically hypertensive rat. V. CALCIUM SIGNALING PATHWAYS INVOLVED IN AUTOREGULATORY RESPONSES An abrupt increase in RPP in mouse microperfused outer cortical afferent arterioles increases [Ca2⫹]i within seconds, Physiol Rev • VOL 95 • APRIL 2015 • www.prv.org 439 RENAL AUTOREGULATORY MECHANISMS followed by a sharp decline to a half-maximal level (865), whereas the juxtamedullary afferent arteriole has a rather more sustained rise in [Ca2⫹]i (1637). Both preparations demonstrate a maintained myogenic contraction despite declining [Ca2⫹]i, indicating increased sensitivity of the contractile machinery to [Ca2⫹]i. There are comprehensive reviews on VSMC Ca2⫹ signaling during generalized myogenic vasoconstriction (169, 301, 302, 478, 614, 1001, 1249, 1329) and its role in modulating renal vascular microcirculatory responses (49, 1249, 1280). Also recommended are excellent mathematical models of renal autoregulation and the Ca2⫹ dynamics of the myogenic response of the afferent arteriole (228, 374, 395, 628, 805, 1350, 1351, 1614). The models incorporate ionic transport and cell membrane potential with contraction dynamics of the afferent arteriolar VMSC. A. Calcium Entry and Mobilization Vasoconstriction results from a combination of increased [Ca2⫹]i and augmented Ca2⫹ sensitivity (FIGURE 8). Intracellular Ca2⫹ signaling involves both Ca2⫹ entry across the plasma membrane and Ca2⫹ release from intracellular sarcoplasmic reticular stores (1329, 1625, 1630). Ca2⫹ entry into preglomerular VSMCs can be mediated by L-type and perhaps T-type VOCCs, voltage-independent receptor-operated channels (e.g., TRPCs, TRPMs, and GPCR-coupled channels), and SOC. GPCRs normally recruit more than one type of cation channel. Both the sustained myogenic tone and the MD-TGF-mediated vasoconstriction are highly dependent on membrane depolarization and Ca2⫹ entry through L-type VOCC. As discussed in later sections, Ca2⫹ sensitivity of the contractile proteins is primarily regulated by PKC and Rho/Rho kinase. The mechanisms underlying mechanosensation and initiation of the myogenic response vary among vascular beds, but usually involve an initial depolarization of the plasma membrane that activates Ca2⫹ entry through L-type VOCCs that leads to activation of myosin light-chain phosphorylation and vasoconstriction. A decreased intraluminal pressure has the opposite effects, thereby resulting in vasodilation. Removal of extracellular Ca2⫹, or inhibition of L-type VOCC activity, abolishes the myogenic response. Mechanotransduction is a key triggering process, but the details are poorly understood. Mechanosensitive initiating elements include ion channels (e.g., TRPC, ENaC) or extracellular proteins (e.g., integrins). Pressure activation of ENaC or nonselective cation TRPC channels may cause depolarization by favoring Na⫹ entry. There is considerable regional diversity in subsequent Ca2⫹ signaling and Ca2⫹ sensitivity of the actin/myosin contractile apparatus responsible for regulation of local blood flow and capillary hydrostatic pressure. The myogenic response of the afferent arteriole is initiated by depolarization of its VSMCs which, in turn, activate 440 L-type VOCCs to allow Ca2⫹ entry. Although the precise mechanisms responsible for the initial depolarization of the plasma membrane are unclear, they may entail the opening of a cation channel to allow Na⫹ or Ca2⫹ entry, the opening of a Cl⫺ channel to permit Cl⫺ exit, or the closing of K⫹ channels to prevent K⫹ efflux. All of these possibilities are under current investigation. Little is known about the role of Ca2⫹ mobilization resulting from Ca2⫹-induced Ca2⫹ release (CICR) from the sarcoplasmic reticulum following stimulation by IP3 and RyRs and less still about Ca2⫹ release from lysosome-like acidic vesicles (49, 1456). Ca2⫹ entry through VOCCs can amplify CICR from the sarcoplasmic reticulum therby enhancing arteriolar contraction (401, 1457). Depletion of sarcoplasmic reticular Ca2⫹ stores in nonrenal vessels can promote Ca2⫹ entry through TRPC or Orai channels. Orai channels on VSMCs mediate Ca2⫹ entry by Ca2⫹ releaseactivated Ca2⫹ channels after activation a stromal interaction molecule (STIM) protein linking the sarcoplasmic reticulum to the plasma membrane (1390, 1482, 1658). Although renal resistance arterioles have store-operated Ca2⫹ entry channels (397, 398, 925, 1448, 1616), their molecular identity is not yet known and their role in regulating myogenic or MD-TGF mediated autoregulatory tone has not been studied. 1. Calcium entry through VOCCs The renal preglomerular arteries and arterioles expressed mRNA for three types of VOCCs of the L (Cav 1.2), T (Cav 3.1 and 3.2), and P/Q (Cav 2.1) subtypes (35, 558, 560, 595, 633). Depolarization of the plasma membrane led to Ca2⫹ entry through L-type VOCCs in the afferent arteriole, which was blocked by the dihydropyridine Ca2⫹ channel blockers (CCB) (191, 558, 935). The depolarization mechanism adapts over time. Thus 2 days of hypertension enhanced expression of the ␣1c-subunit of the Cav1.2 Ca2⫹ channel in the rat main renal artery (1175), which may contribute to sustaining the vasoconstriction. Increasing the transmural pressure of a dog cortical radial artery from 20 to 120 mmHg reduced the EM from ⫺57 to ⫺38 mV and increased the contractile tone of its VSMCs (562), whereas increasing the luminal pressure from 60 to 100 and then to 145 mmHg reduced the EM from ⫺45 to ⫺38 and to ⫺33 mV, respectively (920). Pressurized afferent arterioles in the HNK preparation had a resting EM of ⫺40 to ⫺45 mV that was close to the activation threshold of ⫺30 mV of L-type VOCCs (928, 1388). Increasing the luminal pressure from 40 to 140 mmHg in cerebral arteries of WKY rats and cats reduced the EM of VSMCs from ⫺57 to ⫺41 mV, followed by an action potential spike and a contraction (561, 564). Harder and associates (466, 563) presented evidence that increased cerebral transmural pressure increased 20-HETE that activated PKC which inhibited BKCa channels. This depolarized the Physiol Rev • VOL 95 • APRIL 2015 • www.prv.org CARLSTRÖM ET AL. VSMCs and activated L-type VOCCs to increase [Ca2⫹]i. Such a chain of events mediated autoregulation of cerebral blood flow (466, 563). Ca2⫹ entry through L-type VOCCs is required for renal myogenic and MD-TGF autoregulatory adjustments of the preglomerular vasculature (FIGURE 3D). Autoregulation is abolished following blockade of VOCCs, or removal of extracellular Ca2⫹ from the isolated perfused kidney of the dog (1106, 1108, 1110, 1112, 1120) or rat (1285, 1323) or the kidney of the dog (760, 1069, 1089, 1120) or the rat in vivo (102, 499, 646, 749, 852). CCBs prevent preglomerular myogenic responses of isolated cortical radial arteries or afferent arterioles of the dog (562) or rat or mouse (861, 890) or rabbit (52). Ca2⫹ entry via VOCCs in the renal vasculature is essential also for autoregulatory adjustments of the JMN preparation (192, 194, 200, 201, 688, 1020, 1281, 1437) and the chronic HNK model (586, 587, 593, 929, 931, 1446). CCBs eliminated the MD-TGF response in the JMN (194, 201) and the isolated JGA preparations (82). Pharmacological studies implicated T-type VOCCs in the myogenic response of afferent arterioles (407). However, recent patch-clamp studies of voltage-activated Ca2⫹ currents indicated that freshly isolated VSMCs of the rat afferent arteriole have a high density of L-type channels, but do not express functionally active T-type VOCCs (1388). Myogenic tone in the perfused hindlimb was attributed exclusively to Ca2⫹ entry through Cav l.2 L-type channels (1024), whereas both L- and T-type VOCCs participate in the myogenic response in rat cerebral arteries (4, 576). The regulation of CaV1.2 L-type VOCC trafficking has been studied in cerebral arteries where insertion of activated CaV1.2 into the cell membrane enhanced basal and myogenic tone (1058). Stretch-induced influx of Ca2⫹ generated IP3 and released Ca2⫹ from the sarcoplasmic reticulum. Increased myogenic tone and elevated [Ca2⫹]i were greater in rat brain parenchymal arterioles than in middle cerebral arteries (264), likely because of a higher density of VOCC and a lower BKCa activity. PI3K/Akt signaling promoted voltage-dependent trafficking of Ca2⫹ channels to the plasma membrane that increased Ca2⫹ influx via L-type VOCC and thereby was involved in the myogenic response in mouse second-order mesenteric arteries (197, 910). Genetic knockdown of Dicer for 5– 6 wk to reduce the global role of microRNAs (miRNAs) lowered BP and myogenic tone in mesenteric arteries by reducing PI3K/Akt signaling and L-type VOCC activity (specifically the ␣2␦1 subunit) (101). Interestingly, myogenic tone was restored by stimulation of PI3K signaling by short-term ANG II exposure and by activation of L-type VOCC channels by BAY K8644. Thus noncoding miRNAs have been identified as regulators of vascular contractility and the myogenic response. Future studies are warranted to inves- tigate the role of this novel mechanism in renal autoregulatory responses. In summary, L-type VOCCs play an essential role in the transmembrane Ca2⫹ flux that is required to initiate a myogenic contraction, but the role of T-type VOCCs remains controversial and likely depends on the experimental conditions. 2. Calcium mobilization mediated by IP3Rs Ca2⫹ is stored in the sarcoplasmic reticulum in VSMCs at high millimolar concentrations from where it can be released by either IP3 or ryanodine acting on specific receptors/channels (FIGURE 8). Type 1 IP3Rs were expressed on VSMCs of intrarenal arteries, afferent arterioles, and glomerular mesangial cells, and type 3 IP3Rs were expressed on VSMCs and mesangial cells (596, 1018). The mRNAs for IP3 type 1 and 2 receptors have been identified in VSMCs of small renal arteries (1197). Increased PP in renal arteries and afferent arterioles activated PLC which enhanced the production of IP3 and subsequent Ca2⫹ mobilization (1059). On the other hand, inhibition of IP3 receptor-mediated [Ca2⫹]i mobilization in the dog perfused kidney did not affect steady-state RBF autoregulation, although it attenuated NE-induced renal vasoconstriction (1111). Autoregulatory adjustments of afferent arterioles of JMN were attenuated by reducing Ca2⫹ mobilization from internal stores (688). Inhibition of Ca2⫹ uptake into the sarcoplasmic reticulum by Ca2⫹-ATPase prevented myogenic contraction of afferent arterioles in the JMN preparation (688), but the relative involvement of IP3Rs and RyRs in release of stored Ca2⫹ was not investigated. Inhibition of PLC and thus of IP3 generation in the cortical radial artery attenuated myogenic contractions by 50%, but this was limited to the proximal arterial segments (1447). Thus there are different mechanisms for mobilization of Ca2⫹ for the myogenic response along the cortical radial artery of JMN and perhaps the preglomerular vasculature in general. Nevertheless, Ca2⫹ mobilization contributes importantly to the myogenic mechanism, and perhaps also to MD-TGF in the renal microvasculature of at least some nephron populations. The myogenic response of VSMCs can be engaged after activation of PLC, generation of IP3, and activation of IP3sensitive receptors on the sarcoplasmic reticulum to release Ca2⫹ from internal stores. Alternatively, DAG produced by PLC in VSMCs can activate ion channels including TRPC6 in the plasma membrane to increase [Ca2⫹]i by transmembrane flux. Stretch- or pressure-induced recruitment of GPCR may increase PLC production of IP3 and DAG during the myogenic response (1137). Inhibition of PLC in nonrenal vessels hyperpolarized the EM from ⫺44 to ⫺53 mV, diminished Ca2⫹ entry, reduced [Ca2⫹]i, and attenuated the myogenic response (301, 724). The PLC/DAG sig- Physiol Rev • VOL 95 • APRIL 2015 • www.prv.org 441 RENAL AUTOREGULATORY MECHANISMS naling pathway participated in myogenic responses of human subcutaneous arteries (266). DAG may activate TRPC6 channels to depolarize the plasma membrane to initiate a rise in [Ca2⫹]i via VOCCs. However, IP3 of skeletal muscle arterioles did not influence Ca2⫹ release during myogenic responses (827). The specific role of TRPC channel activation by DAG in the myogenic response of the renal microcirculation is poorly understood and warrants further investigations. 3. Calcium mobilization mediated by RyRs and cADP-ribose Ca2⫹-induced Ca2⫹ release (CICR) of VSMCs refers primarily to activation or sensitization of RyRs, in particular RyR2 and RyR3, to mobilize intracellular Ca2⫹, thereby releasing more Ca2⫹ from the sarcoplasmic reticulum (49). Activation of GPCRs is linked to ADP-ribosyl cyclase that generates cADP-ribose that sensitizes the RyR to mobilize Ca2⫹ and enhance vasoconstriction (FIGURE 8) (49, 399, 400, 1456, 1457). This is a relatively newly recognized VSMC signaling pathway. Ca2⫹ entry through L-type VOCCs also enhances RyR-mediated Ca2⫹ mobilization and renal vasoconstriction (401, 1457), thereby implicating cADP-ribose and RyR in Ca2⫹ signaling in the myogenic response especially in the maintenance phase. A similar mechanism may explain the coupling of RyRs to stretchactivated cation channels (1032). O2⫺ produced in VSMCs by nicotinamide adenine dinucleotide phosphate (NADPH) oxidase may link activation of GPCRs to stimulation of ADP-ribosyl cyclase production of cADP-ribose that increases Ca2⫹ signaling and renal vasomotor tone (399, 400, 1651). Stretch of afferent arterioles stimulated NOX-2/p47phox-dependent production of O2⫺ that enhanced the strength of the myogenic response (864, 866). Moreover, O2⫺ generated by NOX-2 in stretched cardiac myocytes and pulmonary VSMCs sensitized RyRs to enhance [Ca2⫹]i (349, 1201). The extent to which RyRs mediate Ca2⫹ mobilization in renal afferent arterioles, and their involvement in the myogenic response, requires further investigation. Afferent arteriolar vasomotion following activation of Ltype VOCCs and Ca2⫹ entry, which is an index of intrinsic myogenic tone, was prevented by inhibition of RyRs, but not by inhibition of IP3Rs (1443). In fact, caffeine enhanced the fast oscillations in vasomotor tone, indicating that Ca2⫹ entry via L-type VOCCs was reinforced by Ca2⫹ mobilization by RyRs (1443). An increase in luminal pressure in cerebral arteries induced Ca2⫹ sparks that were generated by RyRs acting on the sarcoplasmic reticulum adjacent to the plasma membrane (720, 919). The local increase in [Ca2⫹]i activated BKCa in the VSMC plasma membrane to hyperpolarize the cell, and thereby inactivate L-type VOCCs. As a result, there was a 442 reduced Ca2⫹ entry that limited [Ca2⫹]i and opposed myogenic tone (16, 720, 810). The same sequence of events was reported in mesenteric arteries of one study (836), whereas myogenic tone of mesenteric arterioles was independent of Ca2⫹ mobilization in another (1577). Blockade of RyRs in VSMCs of cremaster muscle feed arteries by ryanodine or tetracaine inhibited Ca2⫹ sparks and Ca2⫹ waves, BKCa activity, global [Ca2⫹]i, and myogenic tone. On the other hand, blockade of RyRs in smaller cremaster muscle arterioles did not affect Ca2⫹ signaling or vascular tone (1596, 1597). In fact, Ca2⫹ mobilization mediated by IP3R enhanced global [Ca2⫹]i and myogenic tone in both vessel types without affecting Ca2⫹ sparks. Other studies reported that activation of RyRs increased global [Ca2⫹]i, Ca2⫹ entry through L-type VOCC, and myogenic tone in isolated rat skeletal muscle gracilis arterioles but not in mesenteric arterioles (1576, 1577). Depletion of sarcoplasmic reticulum Ca2⫹ stores by ryanodine in isolated skeletal muscle gracilis arterioles slowed the development of myogenic tone (178). Thus there are striking regional differences in the roles of RyRs in modulating Ca2⫹ signaling and myogenic responses. RyRs in skeletal muscle arterioles mobilize Ca2⫹ and induce myogenic constriction, whereas RyRs in cerebral arteries induce Ca2⫹ sparks that activate BKCa and inhibit myogenic tone. As discussed in section IVB, BKCa channels in the renal preglomerular vessels generally are inactive. Moreover, global Ca2⫹ mobilization by RyRs lead to marked increases in [Ca2⫹]i and pronounced vasoconstriction rather than the vasorelaxation anticipated from activation of BKCa (49). However, little is known about the function of small Ca2⫹ sparks in the renal microcirculation except that they likely are triggered by integrin deformation. Fibronectin in the extracellular matrix surrounding VSMC is a natural ligand for ␣51-integrin whose mechanical distortion in the rat afferent arteriole can activate Ca2⫹ release via RyRs on the sarcoplasmic reticulum and trigger local Ca2⫹ sparks, global Ca2⫹ mobilization, and contraction 2⫹ (FIGURE 8) (64, 214). ␣51 Integrins also initiated Ca waves that propagated along preglomerular VSMCs (214). Myogenic constriction was inhibited by integrin-specific binding peptides or by blockade of Ca2⫹ mobilization by RyRs. Integrins in pulmonary VSMCs activated ADP-ribosyl cyclase to produce cADP-ribose that activated RyR and released Ca2⫹ from the sarcoplasmic reticulum (1500). Repetitive cycles of RyR-mediated Ca2⫹ mobilization and Ca2⫹ entry through plasma membrane SOC of vasa recta pericytes may be responsible for oscillations in membrane currents and global [Ca2⫹]i (1443, 1656). In summary, pressure-induced membrane depolarization and Ca2⫹ entry through VOCCs are fundamental to the arteriolar myogenic response. The initial mechanisms me- Physiol Rev • VOL 95 • APRIL 2015 • www.prv.org CARLSTRÖM ET AL. diating mechanotransduction and triggering the depolarization of the plasma membrane of VSMCs are poorly understood. Ca2⫹ sensitivity of actin/myosin contractile complex is central to the maintenance phase of myogenic contractions. RyR-mediated Ca2⫹ sparks and BKCa activation can oppose myogenic vasoconstriction in some vascular beds such as the cerebral circulation, whereas RyR-induced CICR and amplification of [Ca2⫹]i can potentiate myogenic constriction in other circulations, likely including the afferent arteriole. Increased [Ca2⫹]i may initiate membrane depolarization by activating ClCa channels or by offsetting the effects of membrane hyperpolarization by activation of BKCa channels. The precise role of RyR-mediated Ca2⫹ mobilization in the renal myogenic response requires further investigation, as does its participation in MD-TGF activity. Overall, CICR provides an important amplification of the increase in [Ca2⫹]i initiated by Ca2⫹ entry through L-type VOCCs. B. Mechanisms Increasing Calcium Sensitivity Although an elevated [Ca2⫹]i is a key requirement for initiating the myogenic response in arteries and arterioles, [Ca2⫹]i may wane over time despite a sustained myogenic vasoconstriction (290). This dissociation implies a change in the balance between the MLCK and MLCP and the phosphorylation state of the actin/myosin proteins to favor contraction (59, 1328). The Ca2⫹ sensitivity of the contractile apparatus of VSMCs is defined by the amount of active phosphorylated 20-kDa MLC (MLC20) at a given level of [Ca2⫹]i and is regulated by PKC and the Rho/Rho kinase (FIGURE 8) (1392). Increased vascular transmural pressure facilitated Ca2⫹ sensitization and contraction mediated by the RhoA/Rho-associated kinase pathway that inhibited MLCP by phosphorylating the MLCP targeting regulatory subunit (MYPT1) and/or the 17-kDa PKC (743). The 17kDa form of PKC in VSMCs inhibited MLCP that potentiated protein phosphatase 1 inhibitor protein (CPI-17). This increased MLC20 phosphorylation and thereby favored VSMC contraction. However, other mechanisms contribute to Ca2⫹ sensitivity. Thus PKC in VSMCs also phosphorylates MLC20 via MLCK that magnifies Ca2⫹-induced actin-myosin interaction and force development. Actin polymerization enhanced connections between the actin cytoskeleton, the plasma membrane, and the extracellular matrix which promoted force transmission (1548). Both cytochrome P-450 – 4A/20-HETE and ROS enhanced Ca2⫹ sensitivity (1328). Endothelial factors such as ET-1 and TxA2 also increased Ca2⫹ sensitivity and myogenic vasoconstriction in gracilis muscle arterioles (1502). Inhibition of PKC, but not the cytochrome P450 inhibitor 17-ODYA, in isolated arterioles from rat skeletal muscle reduced myogenic tone (63). Moreover, smoothelin-like proteins have been implicated in VSMC contractility and vascular adaptations during hypertension. Smoothelin-like 1 protein at- tenuated the myogenic response in cerebral arteries possibly by inhibiting the expression of MYPT1 (1496). Thus Ca2⫹ sensitivity is a complex process involving PKC and Rho/Rho kinase signaling and other mechanisms that regulate the phosphorylation state of the contractile proteins. These processes play an important role in myogenic constriction. 1. PKC PP and GPCR agonists stimulate phospholipid turnover that generates DAG that in turn activates PKC (FIGURE 8). Translocation of PKC to the plasma membrane triggered a cascade of signaling that enhanced Ca2⫹ sensitivity (1275). Both the Ca2⫹-dependent PKC␣ and the Ca2⫹-independent PKC⑀ enhanced the sensitivity of myofilaments to [Ca2⫹]i, whereas the cytoskeletal PKC␦ and the nuclear PKC were not effective (324). Stretch of rabbit basilar arteries triggered translocation of PKC␣ (but not PKC⑀) and RhoA from the cytosol to the plasma membrane which increased phosphorylation of MLC and contributed to the Ca2⫹ sensitivity and the contractile response (1634). PKC is considered crucial for renal vasoconstriction produced by ANG II and ET-1 (22, 68, 1265, 1322, 1659). The myogenic tone of afferent arterioles of the rat HNK preparation was mediated in part by the basal level of PKC that enhanced Ca2⫹ sensitivity, independent of voltage-dependent activation (802). Inhibition of PKC attenuates myogenic tone in basilar arteries (1634), cerebral arteries (63, 474, 603, 725, 855, 1136, 1136, 1164), coronary arteries (324, 1006), mesenteric arteries (380), cremaster and skeletal muscle arterioles (63, 977), and subcutaneous arteries (266). Inhibition of PKC in skeletal muscle arteries prevented a myogenic response to a PP increase without affecting the increase in [Ca2⫹]i (768). This was specific for PKC␣ in coronary arteries (324). Pressure-induced constriction of cerebral arteries was dependent on increased Ca2⫹ sensitivity due to PKC and Rho kinase (63, 603, 855). In summary, PKC activation is a central component of Ca2⫹ sensitivity and the myogenic response in most arteries studied, including the renal preglomerular vessels. 2. Rho/Rho-kinase As mentioned above, Rho kinase can enhance contractility by inhibiting MLC phosphatase by targeting MYPT1 (FIGURE 8). Mechanical stimulation of VSMCs translocates the GTPase Rho and its associated Rho kinase to caveolin-1 in the plasma membrane. The myogenic response was enhanced by RhoA/Rho-kinase phosphorylation that inactivateed MLCP and augmented Ca2⫹ sensitivity (350). Inhi- Physiol Rev • VOL 95 • APRIL 2015 • www.prv.org 443 RENAL AUTOREGULATORY MECHANISMS bition of this system, or disruption of caveolin-1, reduced myogenic force in mesenteric and ophthalmic arteries (380, 694). Rat cerebral artery myogenic tone depended on the independent effects of Rho-kinase and PKC mediating increased Ca2⫹ sensitivity, but Rho kinase predominated (694, 725, 855). The myogenic response of gracilis and cremaster muscle arteries involved Rho-kinase and PKCevoked Ca2⫹ sensitization leading to inhibition of MLCP and cytoskeletal reorganization of actin polymerization (1027). Membrane depolarization in pulmonary artery VSMCs generated O2⫺ that activated RhoA and led to Ca2⫹ sensitization (1233). RhoA activation and interaction with caveolin-1 were critical for pressure-induced myogenic tone in rat mesenteric resistance arteries (350). Juxtamedullary preglomerular arterioles expressed Rho-kinase ␣ and  (691), whose inhibition completely abolished autoregulation (691), thereby implying inhibition of both myogenic and MD-TGF mechanisms. Indeed, recent work showed that Rho-kinase participates in afferent arteriolar vasoconstriction mediated by both myogenic and MD-TGF mechanisms in response to elevated RPP in the superficial cortex of WKY rats (632). Rho kinase in the rat HNK preparation participated in the myogenic vasoconstriction of cortical radial arteries and afferent arterioles to changes in increased PP, ANG II, and reduced EM (1259, 1358). Inhibition of Rho-kinase blunted pressure-induced myogenic tone and depolarization-induced constriction of isolated afferent arterioles (1054) and also almost abolished the dynamic RPP-RBF transfer functions representative of both the myogenic and MD-TGF responses (1358). In summary, activation of Rho/Rho-kinase is a quantitatively important step in increasing the Ca2⫹ sensitivity that maintains or enhances both the myogenic and MD-TGF autoregulatory mechanisms in the kidney. VI. MODULATING AGENTS The myogenic response of renal (915) and nonrenal (301) vessels was generally intrinsic to their VSMCs and independent of endothelial input. Thus myogenic constriction clearly was triggered within VSMCs, although it could be modulated by various endothelial factors. Despite the constancy of afferent arteriolar blood flow during increased PP by effective autoregulatory adjustments in vascular resistance, the narrowed vascular lumen will increase vessel wall shear stress that could activate endothelial pathways producing vasoactive agents. For example, NO was released by eNOS in response to ANG II (699, 1160), ET-1 (701, 757), or shear stress (696, 747, 1184). Other potential modulators of myogenic tone in the kidney can originate from structures involved in MD-TGF or CT-GF with signaling from MD or CT tubular cells (FIGURE 6). 444 A. ANG II JG granular cells in the terminal afferent arteriole increase renin release and thereby ANG II production (FIGURE 1) in response to reduced luminal pressure at the end of the afferent arteriole, low NaCl delivery to the MD, or increased sympathetic nerve activity activation of -adrenoceptors (204, 846). Proximal tubular and CD cells also contribute to intrarenal renin and ANG II generation (812, 1076). Most studies have concluded that the RAS was not essential for efficient steady-state autoregulation of RBF. ANG II infusion, or inhibition of AT1 receptors or ACE, did not modify highly efficient whole kidney autoregulation in dogs (5, 27, 90, 97, 103, 194, 548, 550, 755, 1003, 1047, 1072, 1171, 1261) or rats (45, 329, 750) even during low-salt diet producing strong activation of the RAS and did not usually affect the lower limit of RBF autoregulation. ANG II also did not affect renal medullary blood flow autoregulation (288). However, ANG II was more important in steadystate autoregulation of the GFR than the RBF (488, 550, 774, 1261), especially at lower RPP, most likely by maintaining efferent arteriolar tone. However, a few studies have reported that the RAS was essential for effective steadystate autoregulation of RBF and GFR (764, 1479). Moreover, autoregulation of isolated perfused rats kidneys was improved by ANG II (523). Micropuncture studies of superficial nephrons demonstrated that MD-TGF responses were enhanced by ANG II (133, 1012, 1308, 1309) and diminished by blockade of ACE (133, 648, 1189, 1422, 1495), or AT1 receptors (781, 1309, 1590), or by deletion of the genes for ACE or AT1A receptors (582, 1310, 1318, 1481). AT1 receptors may modulate MD-TGF responses by actions on the vasculature or on NKCC2 cotransport activity in MD cells (829, 1012, 1556). These effects were countered by NO production by MD nNOS (656). One mechanism of MD-TGF enhancement by ANG II is synergistic potentiation of the actions of adenosine on the afferent arteriole (440, 556, 862, 863, 1159, 1480, 1580). Thus ANG II appears to be an important modulator of MD-TGF activity, but its presence or absence usually has little impact on highly efficient steady-state autoregulation of RBF, at least during the plateau phase. The effects of ANG II on the myogenic mechanism in the kidney are controversial. ANG II contracted the afferent arteriole of the JMN preparation without affecting its autoregulatory response (690). However, low levels of ANG II enhanced the myogenic response in the HNK preparation by potentiating PKC-induced increases in Ca2⫹ sensitivity (802). ANG-(1–7) had no effect on the myogenic tone of preglomerular vessels in the HNK preparation (1524). Physiol Rev • VOL 95 • APRIL 2015 • www.prv.org CARLSTRÖM ET AL. Short-term infusion of ANG II at pressor rates caused renal vasoconstriction without major effects on the myogenic response, the MD-TGF, or the third mechanism analyzed by time-dependent changes in RVR following a rapid, step increase in RPP (750, 755). ACE inhibition has no effect on the myogenic response, and the MD-TGF was slightly attenuated (755). Short-term administration of vasoconstrictor amounts of ANG II to the conscious dog slightly augmented the admittance gain of the transfer functions representing the myogenic and MD-TGF responses, but without affecting their relative contributions (755). Prolonged infusions of ANG II, but not PE, into conscious or anesthetized rats potentiated the admittance gain of the myogenic response but not the MD-TGF (1193). Whereas ANG II slightly increased the slope of the admittance gain reduction, consistent with increased strength of the myogenic mechanism, it did not modify the signature resonance peak of the myogenic mechanism (at ⬃0.23 Hz), which is an indicator of the speed of contraction. The ANG II effect is attributed to activation of AT1 receptors rather than increased RPP. Other frequency analysis studies indicated that increased endogenous ANG II enhanced the myogenic and MD-TGF responses in rats (329, 597), but reduced ANG II activity produced by high salt diet had little effect (1271, 1273). Other studies employing dynamic frequency analysis reported that acute hypertension produced by PE does not affect the myogenic or the MD-TGF mechanism in Wistar (1358) and Brown-Norway (BN) rats (1570). Staircase changes in RPP can produce hysteresis loops where there are different RVR responses to increases versus decreases in RPP in the anesthetized (284) and conscious rat (423). Autoregulation curves for RBF had a lower plateau level when measured during sequential increases compared with subsequent decreases in RPP. This was attributed to activation of the RAS by hypotension. The outcome was that the RAS enhanced RVR at low RPP where it engaged the MD-TGF for RBF autoregulation (284). Early micropuncture studies in the rat reported reductions in RPP below the normal lower limit of autoregulation abolished MD-TGF, but this was slowly restored by ANG II generation after 20 – 60 min of maintained hypoperfusion (1343). The increased RAS activity reduced the lower limit for RBF autoregulation (284, 624, 707, 1397). The putative third mechanism may be modulated by ANG II (1342). ANG II can enhance CT-GF leading to a paradoxical dilation of the afferent arteriole by activating ENaC and stimulating Na⫹ reabsorption in the CT (1227). Sex differences have been reported for expression of AT1 and AT2 receptors and their functional impact on renal autoregulation, MD-TGF, and pressure-natriuresis. Kidneys of female rats express a higher density of vasodilator AT2 receptors that can increase the lower limit of RPP for RBF autoregulation and shift the pressure-natriuresis relation to excrete more Na⫹ at a lower RPP (619). Augmented AT2 receptor signaling in female rats also accounted for a blunted effect of ANG II to enhance MD-TGF (156). In summary, ANG II is a potent vasoconstrictor of the afferent and efferent arterioles. Reductions in RPP release renin and generate ANG II within the kidney which preferentially activates AT1 receptors on the efferent arteriole, thereby maintaining PGC and GFR. This accounts for the particular effect of ANG II in maintaining autoregulation of GFR, relative to RBF, at reduced RPP. Additionally, agonist-independent, stretch activation of AT1 receptors can interact with TRP channels as mechanosensors (see sect. IIIC). ANG II clearly magnifies MD-TGF responses, but most studies suggest a rather weak effect on the myogenic response. Yet, although the individual components of renal autoregulation may be modified by acute ANG II, this generally does not cause major changes in overall steady-state RBF autoregulation. The exception is that ANG II generated by hypotension can modulate the lower limit of PP steady-state autoregulation, perhaps through its ability to enhance MD-TGF activity. As noted in section VIIB5, chronic ANG II infusion causes hypertension, but the effects on the efficiency of RBF autoregulation are inconsistent. B. Eicosanoids Vascular production of vasodilatory PGs such as PGE2 and PGI2 commonly buffer the actions of vasoconstrictor agents (671), but renal hemodynamics also are modulated by vasoconstrictor PGs and by tubular production of eicosanoids. A reduction in RPP reduced the release of PGE2 and PGI2 from the kidney. However, PGs did not affect steady-state RBF autoregulation in salt-replete rats (422) or dogs (1531) or autoregulation of the PGC in superficial nephrons of normal Munich-Wistar rats (1167). Moreover, arachidonate metabolites of COX or cytochrome P-450 were not required for steady-state RBF autoregulation in isolated perfused kidneys of the dog (763, 1107) or rat (1285) or for myogenic contractions of afferent arterioles (588, 747). Nevertheless, other studies have implicated PGs under certain circumstances. Thus the poor autoregulation of papillary blood flow in the rat kidney was improved by COX inhibition (1127), while whole kidney RBF autoregulation was slowly enhanced following COX inhibition (833). Efficient autoregulation of GFR in superficial nephrons depended on COX metabolites contributing to MD-TGF (1313). COX products also participated in conventional MD-TGF-mediated adjustments of afferent arteriolar tone. Micropuncture studies of superficial nephrons demonstrated that COX inhibition reduced MD-TGF sensitivity to changes in tubular flow (1034, 1307, 1319), but this was Physiol Rev • VOL 95 • APRIL 2015 • www.prv.org 445 RENAL AUTOREGULATORY MECHANISMS restored by tubular perfusion with PGE2 or PGI2 but not PGF2␣ (1319). However, other investigators reported that MD-TGF sensitivity was enhanced by tubular perfusion with PGE2 and PGF2␣, whereas perfusion with PGI2 markedly attenuated feedback sensitivity (1170). omega hydroxylase (445, 446). It augmented autoregulation of the afferent arteriole in the JMN preparation with participation of both myogenic and MD-TGF mechanisms (673, 676) and enhanced the myogenic tone of rabbit and mouse isolated afferent arterioles (464). More consistent is the observation that MD-TGF is enhanced by MD generation of prostaglandin endoperoxide (PGH2) and TxA2 (137, 1589, 1593) or by isoprostanes, all of which activate TP receptors that can constrict afferent arterioles and also can enhance Cl⫺ reabsorption from the TAL of Henle’s loop (38, 437, 932, 1319, 1581, 1591). Thus activation of TP receptors may augment MD-TGF by enhancing both the MD signal generated by NaCl reabsorption and the afferent arteriolar responsiveness. TxA2 in gracilis muscle arteries also increased myogenic tone (1502), which was ascribed to enhanced Ca2⫹ sensitivity of the contractile apparatus. Increased transmural pressure in cerebral arteries increased 20-HETE production that activated L-type VOCC currents, myogenic vasoconstriction, and autoregulation of blood flow (468, 1181). 20-HETE also augmented the myogenic responses of cerebral, skeletal muscle, and mesenteric arterioles (243, 468). These vasoconstrictor actions of 20HETE were ascribed to blockade of BKca channels on VSMCs which caused depolarization and activation of Ca2⫹ entry via VOCCs (677, 943, 1666). 20-HETE also has been implicated in RBF autoregulation in vivo in some (1668) but not all studies (1199). It enhanced MD-TGF activity independent of NKCC2-mediated MD reabsorption, presumably by magnifying the vasoconstrictor response to MD-derived adenosine and/or ATP (464, 1668). In contrast, vasodilator EETs generated by arachidonate cytochrome P-450 epoxygenase in endothelial cells in the JMN preparation blunted the afferent arteriolar autoregulatory vasoconstriction (673). It is not yet clear how cytochrome P-450 enzymes are modulated by RPP is not yet clear. Sex differences have been noted in the control of blood perfusion within the kidney by COX metabolites (1152, 1412). Blood flow autoregulation in cortical and outer medullary regions of kidneys of male rats was efficient and independent of PGs, whereas poor medullary autoregulation in female rats was attributed to overproduction of PGs. COX-2 generated vasodilatory metabolites in the JMN preparation during increased activation of MD-TGF, which counteracted MD-TGF-mediated constriction of the afferent arteriole and blunted its constrictor response to increased RPP (657, 658). In contrast, COX-1 knockout mice have a normal MD-TGF response (39). These actions of COX-2 metabolites were attributed to activation of MD (39, 232, 577, 657). Thus the rather inconsistent effects of COX inhibition on renal autoregulation in health may relate to offsetting effects of MD release of vasodilator and vasoconstrictor prostanoids, confounding effects of sex differences between COX-1 and-2, and moderating effects of NO. Products of COX-2 and nNOS produced in MD cells of the rat JMN preparation attenuated autoregulation (657, 658). Inhibition of nNOS no longer enhanced MD-TGF after blockade of COX-2. Therefore, COX-2 products appear to blunt autoregulation secondary to MD nNOS generation of NO. COX inhibition did not affect the myogenic response of the rat HNK lacking MD-TGF (588), consistent with a primary effect of vasodilator PGs on MD signaling. Global inhibition of cytochrome P-450 production of 20HETE and EETs in the JMN preparation (469) or the dog arcuate artery (777) attenuated the myogenic response. In contrast, myogenic responses were not affected by global inhibition of COX-1 and -2 activity. 20-HETE was generated during increased PP in the preglomerular vasculature and the gracilis muscle arteries by a cytochrome P-450 446 In summary, arachidonate metabolites have complex effects on both myogenic and MD-TGF mechanisms. PGs generally are not required for overall renal autoregulation. However, MD-TGF response can be enhanced by vasoconstrictor prostanoids and TxA2 and vasoconstriction can be blunted by vasodilator PGs. In most studies, myogenic responses are increased by 20-HETE and reduced by EETs. C. NO Endogenous NO has complex effects on renal autoregulation. It can blunt both myogenic and MD-TGF mechanisms. Whereas NO contributes importantly to the regulation of RVR in many preparations, it does not affect the overall efficiency of steady-state whole kidney RBF autoregulation over a defined range of RPP. Short-term inhibition of NOS or administration of an NO donor does not perturb steady-state RBF autoregulation in the normotensive rat, the SHR, or the dog (69, 81, 815, 853, 952, 953, 960, 963, 1052, 1109, 1492) nor does it perturb renal cortical or medullary blood flow autoregulation in the dog (960). NOS inhibition improved the poor papillary autoregulation without affecting efficient RBF autoregulation in one study of the hydropenic Munich-Wistar rat (1127). Although not affecting steady-state RBF autoregulation in the plateau phase, NO modulated the range of RPP over which RBF was autoregulated by shifting the inflection point to a lower RPP in some (831, 832, 1203), but not all Physiol Rev • VOL 95 • APRIL 2015 • www.prv.org CARLSTRÖM ET AL. studies (69, 81, 815, 853, 952, 953, 960, 963, 1109, 1492, 1494). On the other hand, increased NO release in the JMN preparation attenuated PP-induced diameter adjustments of the cortical radial artery and afferent arterioles (675, 1184) by reducing myogenic and MD-TGF responses. Inhibition of NOS improved the myogenic response of afferent arterioles of JMN preparation perfused with a cell-free artificial solution (675, 1184). Autoregulation of this preparation was improved with the use of erythrocytes or albumin in the perfusate that scavenge NO (192, 1168, 1184). Thus endogenous generation of NO can blunt both the MD-TGF and myogenic mechanisms and yet has little effect on steady-state autoregulation of RBF. Moreover, global inhibition of NOS, or selective blockade of nNOS, magnifies MD-TGF responses in superficial nephrons whether assessed by micropuncture and microperfusion (38, 116, 133, 188, 661, 761, 1052, 1228, 1463, 1493, 1515, 1553, 1586, 1587, 1602, 1608, 1609) or by wavelet analysis of blood flow oscillations of superficial nephrons (1402). Oscillations in RBF and RVR often become readily apparent after NOS inhibition. Comparison of oscillations in blood flow in the efferent arteriole and the whole kidney led to the conclusion that NO attenuates the strength of arteriolar oscillations mediated by both the myogenic and the MD-TGF mechanisms while improving their synchronization (1402). Dynamic studies have demonstrated that NO slows and/or attenuates renal myogenic vasoconstriction. Inhibition of NOS in the rat augments the strength and speed of the myogenic response to a rapid, step increase in RPP (FIGURE 7A). This increased the contribution of the myogenic mechanism from 35 to 80% (299, 750) or 45 to 100% (1626). As is shown in FIGURE 7A, NOS inhibition also shortens the time to completion of a myogenic response from ⬃8 to 4 s. Indeed, inhibition of NOS augmented the myogenic response sufficiently to induce complete autoregulation thereby providing transmission of little error signal to MD cells to activate MD-TGF. Vascular admittance transfer function analysis in conscious dogs (754) or in some (750, 1357, 1358, 1402, 1573), but not all (1340), conscious or anesthetized rats confirmed that endogenous NO can attenuate the strength of the renal myogenic response. NOS inhibition primarily enhanced the myogenic mechanism by increasing the slope of reduction in admittance gain between 0.05– 0.20 Hz (FIGURE 7B) and the height of the associated phase peak in normotensive (Long-Evans, SD, and Wistar) rats but not in SHR (1341, 1358, 1563, 1569), which could possibly be explained by reduced NO signaling or bioavailability in their kidney due to excessive ROS. The slope of the decline in admittance gain increased from ⬃30 to ⬃55 dB/decade, suggesting the emergence of a second-order component responding to the rate of change in BP in addition to the actual level of BP. NOS inhibition increased the fractional compensation from 40 to 58%, which indicates an enhanced vigor of the myogenic response. However, in this study, the corner frequency where admittance began to decline was largely unaffected by NOS inhibition, which indicates a maintained rate of myogenic contraction. Moreover, the markedly attenuated myogenic response in normotensive BN rat with forced increases in RPP (1563) was normalized by inhibition of NOS (1563, 1569, 1570) (see sect. VIIB8). There is controversy about the origin of the NO that modulates the renal myogenic response. Early studies of rabbit isolated perfused afferent arterioles or afferent arterioles of the rat HNK indicated that the NO that attenuated the myogenic responses was generated by the endothelium (586, 593, 696, 702, 747). Moreover, stimulation of endothelial-derived NO by activation of vascular ETB receptors blunted the myogenic response of the rat isolated renal and mesenteric arteries (1096) and the whole kidney (1357). In contrast, a unique renal source was implicated by the finding that NO modulated the myogenic response in the rat kidney but not in the hindlimb (750). This was ascribed to blunting of myogenic tone by nNOS-derived NO in the MD rather than by NO produced by the vascular endothelium (FIGURE 7). However, NOS inhibition in some other studies of nonrenal vessels, such as skeletal muscle arteries, mesenteric arteries, and splenic arterioles, accelerated or magnified myogenic constriction, thereby implicating eNOS in these extrarenal vascular beds (154, 377, 519, 944, 1339, 1530). Selective inhibition of nNOS in the JMN preparation enhanced the autoregulatory constrictor responses to increased PP of the afferent arteriole (FIGURE 7D) (657, 658, 662). The NO signal arose from MD cells since the effect of nNOS inhibition was abolished by elimination of MD-TGF by papillectomy. However, it is not clear from these JMN studies whether NO affects primarily the myogenic or the MD-TGF mechanism. Support for the view that MD-derived NO attenuates myogenic tone derives from the finding that NOS inhibition did not enhance myogenic constriction in preparations lacking functional MD cells, i.e., mouse isolated perfused afferent arterioles with endothelium present (866) or rat afferent arterioles in the HNK preparation (FIGURE 7C) (1358). Moreover, isolated perfused afferent arterioles from eNOS-deficient mice retained normal myogenic response (866). In addition, NOS inhibition did not affect afferent arteriolar constrictor responses in the artificial fluid-perfused JMN preparation to increased RPP (675). These studies appear to exclude a major role for NO derived from eNOS in afferent arterioles and implicate nNOS-derived NO in the MD as the primary source that attenuates the strength of the myogenic response in intact kidneys. Physiol Rev • VOL 95 • APRIL 2015 • www.prv.org 447 RENAL AUTOREGULATORY MECHANISMS Collectively, there are several lines of evidence to support the notion that NO generated by MD cells attenuates myogenic response of the preglomerular vasculature by buffering the speed and/or the strength of the myogenic mechanism (750, 1358, 1569). Support derives from studies of renal vascular dynamics based on responses to a rapid, step increase in RPP, on frequency analysis of transfer function admittance gain and a dependence on MD-TGF activity. In some studies, the myogenic response after acute global NOS inhibition became so rapid and robust that the autoregulatory response to a step increase in RPP was complete within 10 s without any evidence of participation of a MD-TGF or other mechanisms. The exaggerated myogenic system during NOS inhibition appeared to have become sufficiently potent to prevent an error signal from reaching the MD to activate MD-TGF and other mechanisms. Global NOS inhibition may prevent modulation of myogenic responses by NO generated from any source. However, the enhancing effect of L-NAME on the myogenic response was linked to MD nNOS since it was reduced (1358) or abolished (750) by furosemide. Frequency analysis reported that global inhibition of NOS in Long-Evans, SD, and Wistar rats (FIGURE 7B) (1569) or selective intrarenal inhibition of nNOS, but not iNOS, augmented the myogenic contribution to dynamic autoregulation in SD rats (1358). Selective inhibition of nNOS in vivo increased the slope of admittance gain decline from 24 to 42 dB/decade, a change similar to that observed with global NOS inhibition with L-NAME (1358). These results indicate that MD-derived NO is an important pool to attenuate the myogenic responses of the preglomerular vasculature in the kidney. A recent study questioned this view. Pharmacological inhibition of NOS, but not nNOS or iNOS, increased the speed and strength of the myogenic response to a step increase in RPP without perturbing overall steady-state RBF autoregulation in rats (299). In agreement, global NOS inhibition magnified the myogenic component of RBF autoregulation in wild-type mice and nNOS mutants, but not in mice lacking eNOS (299). This suggests an NO-independent role for eNOS that warrants further investigation. The cellular events by which NO attenuates VSMC myogenic responses are incompletely understood. In one study of the renal microcirculation, exogenous NO signaled via cGMP-dependent and -independent pathways depending on its concentration (1486). Low renal NO concentrations (⬍1 M) that appear to be in the physiological range signal primarily through cGMP (887, 1486). Kurtz and colleagues (1283) demonstrated that NO generated cGMP that caused renal vasodilation primarily by inhibiting phosphodiesterase 3 that degrades cAMP. NO donors, and high levels of 8-bromo-cGMP, completely abolished autoregulation of preglomerular vessels in the JMN preparation (132), indi- 448 cating inhibition of both myogenic and MD-TGF mechanisms. Since stimulation of cGMP by atrial natriuretic peptide (306, 904, 1644) failed to restore RBF or reverse the exaggerated myogenic response detected by transfer function analysis following NOS inhibition (1573), there must be different pools of renal NO that regulate vascular responses. There are multiple sites where NO, cGMP, or its target PKG and cAMP/PKA could dampen Ca2⫹ signaling or sensitivity, and thereby moderate arteriolar tone (434). NO in the renal vasculature reduced agonist- or stretch-induced increases in Ca2⫹ influx by inhibiting VOCCs or TRPC channels or by activating BKCa channels (403, 408, 832, 898, 1011, 1642, 1645). NO in the afferent arteriole is reported to inhibit both L- and T-type VOCCS (408), which could account for its ability to antagonize both MD-TGF and myogenic responses. Moreover, NO in VSMCs suppressed ADP-ribosyl cyclase activity, thereby reducing RyR-mediated Ca2⫹ mobilization (403, 898, 1642). Both NO and ROS in VSMCs regulated RyR-mediated Ca2⫹ mobilization (384, 1645), but by distinct signaling pathways (752, 861, 866, 1224). NO dilated gracilis muscle arteries and afferent arterioles by activating MLCP (129, 1499, 1503), and attenuated myogenic tone of nonrenal vessels by inhibiting Rho kinase in VSMCs (475, 1214, 1458). This may also occur in the renal microcirculation (1358). NO inhibition of 20-HETE formation or action diminished myogenic constriction of coronary arteries (21, 645, 921, 1144). Thus NO has multiple mechanisms that can reduce vascular tone and/or Ca2⫹ sensitivity. NO also could impact the myogenic response indirectly by attenuating MD-TGF signaling. NO from nNOS can inhibit reabsorption of NaCl by activated MD cells, thereby blunting the signal for MD-TGF responses (702, 1460, 1602). NO and GMP signaling in the MD phosphorylates and inhibits NKCC2 and thereby reduces luminal NaCl uptake (828, 1228). Moreover, NO inhibited ecto-5’-nucleotidase that is the final enzyme in the conversion of ATP to adenosine, which can limit MD-TGF activation (1287). NO and O2⫺ interact in MD cells where O2⫺ inactivates NO and enhances MD-TGF responses during oxidative stress (913, 1393). The effect of NOS inhibition to enhance the renal myogenic response can be dissociated from renal or systemic vasoconstriction in most (750, 754, 1358, 1573) but not all studies (1572). The enhancement of myogenic responses by NOS inhibition in vivo can be prevented by furosemide, thereby implicating MD-TGF (755, 1193, 1563). In summary, NO has complex interactive effects on renal autoregulation. It modulates both the dynamics and the steady-state contribution of individual mechanisms. There are mixed results for a role of NO derived from the endothelium of the afferent arteriole in blunting the myogenic Physiol Rev • VOL 95 • APRIL 2015 • www.prv.org CARLSTRÖM ET AL. response. More convincing evidence demonstrates that NO generated by nNOS in the MD not only attenuates MDTGF but also restricts the speed and strength of the myogenic response. During NOS inhibition, the myogenic tone can become sufficiently strong to account for nearly all of the highly efficient renal autoregulation (FIGURE 7, A AND B). The possible role of NO in vasomotion of isolated arteries/arterioles was discussed in section IIE. D. Carbon Monoxide Constitutively active heme oxygenase 2 (HO-2) and inducible HO-1 catalayze heme to CO, iron, and bilverdin. HO-2, but not HO-1, is highly expressed in endothelial and VSMC. CO produced a biphasic response in the renal vasculature, with vasodilation at low concentrations (100 nM) and vasoconstriction at high concentrations (1–10 M) (1445). These vasoactive effects were ascribed to stimulation of NO production by endothelial cells and inhibition of eNOS, respectively. CO attenuated the actions of vasoconstrictors in isolated interlobar arteries by activating VSMC K⫹ channels (1653). Induction of HO-1 in rat kidneys increased basal RBF but did not impair the efficiency of RBF autoregulation (130). However, induction of HO-1 in tubular epithelial cells in the rat JMN preparation increased CO production that blunted the myogenic tone of afferent arterioles, whereas biliverdin was without effect (131). The reason for discrepancy between whole kidney autoregulation in vivo and that recorded for myogenic responses of the afferent arteriole of JMN in vitro requires further investigation. The MD expressed HO-1 and HO-2 that generated CO and inhibited MD-TGF in vivo via activation of sGC and cGMP signaling (1557). Biliverdin appeared to attenuate MDTGF activity by scavenging O2⫺ (1557). Both CO and biliverdin inhibited MD-TGF in the rabbit isolated JGA preparation (FIGURE 6) (1222, 1225, 1226). The HO inhibitor chromium mesoporphyrin decreased renal HO-1 and CO but did not affect RBF or GFR or NO production in the rat (718). In another study, HO inhibition increased RVR in the rat kidney which was more pronounced when NOS was inhibited (1242). On the other hand, HO and CO in the renal medulla may participate in the regulation of medullary blood flow, Na⫹ excretion, and BP (897, 1665). CO blunted rat gracilis arteriolar myogenic responses (830) and attenuated VSMC myogenic tone by activating BKCa channels to increase EM (881, 1653). In summary, CO at low concentrations in vitro in isolated preparations generates NO that activates sGC signaling to produce vasodilation and inhibit both the myogenic and MD-TGF mechanisms. However, CO has little effect on RBF autoregulation in vivo. Renal HO-1 is often induced during pathophysiological conditions (6, 1062), which might be a pathway to attenuate autoregulation and accelerate barotrauma. Thus, despite this potential, the roles or the underlying mechanisms of CO and HO in autoregulation are not clearly established. E. Endothelin-1 Endothelin is produced primarily by vascular endothelial cells and by tubular cells of the TAL of Henle’s loop and the CD. Renal vascular actions of endothelin (ET)-1 are mediated by vasoconstrictor ETA and ETB receptors on VSMCs and vasodilator ETB receptors on vascular endothelial cells linked to NO production. Steady-state RBF autoregulation was unaffected by ETA receptor blockade (97, 140) or by combined blockade of ETA⫹ETB receptors (831). However, blockade of ETB, but not ETA, receptors enhanced the myogenic response in vivo and intrarenal arteries in vitro by inhibiting NO production (453, 1357). For actions of NO on RBF autoregulation and individual components, see section VIIC. A low concentration of ET-1 that does not contract afferent arterioles in the HNK potentiated myogenic reactivity by increasing PKC and its modulation of K⫹ channels (802). Mechanistically, Kirton and Loutzenhiser (802) proposed activation of PKC and modulation of K⫹ channel activity, but as discussed in next section ET-1 may also activate NADPH oxidases. F. Reactive Oxygen Species Reactive oxygen species (ROS), such as O2⫺, H2O2, and hydroxyl anion (OH.⫺), are reactive byproducts of mitochondrial respiration, uncoupled NOS, or specific oxidases, notably NADPH oxidase, xanthine oxidoreductase, and certain arachidonic acid oxygenases. NADPH oxidase is a major source of renal and vascular ROS (494, 895, 1016, 1086, 1151). It is a multiunit enzyme comprising the membrane-bound neutrophil oxidase (NOX) unit, either NOX-1, NOX-2 (gp91phox), NOX-4, or NOX-5. All vascular NOXs generate ROS, but their intracellular distribution, their requirement for interaction with the NOX subunits p22phox, p47phox (or NOXO1), p67phox (or NOXA1), and p40phox and the small G protein Rac-1 or Rac-2, and their modes of regulation and generated ROS (O2⫺ or H2O2) differ (78, 136, 1086). For activation of the enzyme, participation of Rac 2 (or Rac 1) and Rap 1A (128), but also PKC are considered crucial (136). The kidney has a rich, diverse, and cell-type specific distribution of NADPH oxidases (471). ROS were produced in all renal vascular and tubular cells studied (40). NOX-1 was expressed primarily in large vessels, whereas NOX-2 and NOX-4 were expressed abundantly together with their chaperone protein (p22phox) in resistance arterioles. NOX-5 was expressed in human, but not rodent, vessels Physiol Rev • VOL 95 • APRIL 2015 • www.prv.org 449 RENAL AUTOREGULATORY MECHANISMS (78, 182, 873, 1150, 1477). The afferent arteriole and MD cells both expressed NOX-2 and NOX-4 and the subunits capable of generating O2⫺ or H2O2 (471, 1603, 1657). The rat and rabbit afferent arteriole expressed p22phox that was required for activation of NOX-1, NOX-2, and NOX-4 (471). NADPH oxidase-mediated O2⫺ generation in MD cells enhanced MD-TGF in large part by limiting NO bioavailability (912, 913, 1608). Flow-dependent NaCl reabsorption, or stimulation by ANG II, primarily activated MD NOX-2 and enhanced MD-TGF signaling, whereas NOX-4 contributed to basal ROS production (448, 912). Oxidative stress implies an increased production, or a decreased scavenging or metabolism of ROS. Many effects of oxidative stress can be attributed to NO deficiency. Increased NADPH oxidase-derived O2⫺ production and vascular dysfunction have been reported in hypertension, diabetes, atherosclerosis, and CKD (40, 895, 1604). This section primarily focuses on the roles of vascular O2⫺ and H2O2 in renal autoregulation in health, whereas section VII considers the pathophysiological contribution of ROS. Preliminary studies suggested that renal autoregulation in vivo to a step increase in RPP was dependent on NADPH oxidase whose inhibition with apocynin attenuated myogenic response, but had little effect on MD-TGF and the third mechanism. The enhanced myogenic response following inhibition of NOS was largely prevented by inhibition of NADPH oxidase or by tempol (752). Thus the enhancement of myogenic responses by NADPH oxidase-derived O2⫺ was offset by interactions with NO and by metabolism of O2⫺ by superoxide dismutase (SOD). Indeed, many studies in the renal vasculature have concluded that ROS can regulate vasomotor tone by direct effects on VSMC and by quenching and inactivating NO (758, 1301, 1603, 1604). O2⫺ in the JGA enhanced vasoconstriction by scavenging and inactivating NO (1217, 1301, 1588, 1592, 1603, 1608), but also acted directly on the afferent arteriole. The inhibition of MD-TGF by tempol was enhanced by endothelium removal (913), suggesting a primary action of ROS and modulatory role for NO. Activation of GPCRs in VSMCs rapidly increased O2⫺ that potentiated Ca2⫹ signaling in afferent arterioles and renal vasoconstriction (183, 399, 400, 864). Thus renal vasoconstriction with ANG II, ET-1, NE, or U-46619 was blunted by inhibition of NADPH oxidase, genetic deletion of p47phox or NOX-2, or by metabolism of ROS by SOD or tempol (182, 183, 758, 759, 864, 1037, 1301). O2⫺ generated by NADPH oxidase in afferent arteriolar VSMC activated ADP-ribosyl cyclase to increase Ca2⫹ mobilization by RyRs (399, 400, 1456), whereas NO suppressed ADP-ribosyl cyclase activity to reduce [Ca2⫹]i (403, 898, 1642). 450 The enhancement of myogenic responses by ROS may involve activation of GPCR coupled to second messengers, cytosolic proteins, and [Ca2⫹]i (FIGURE 8) (1328) linked to redox signaling by vascular NADPH oxidases (1086). Moreover, stretch activation of AT1 receptors, in the absence of ANG II, enhanced the myogenic tone of cerebral and renal cortical radial arteries via ROS generation (999). Several studies have demonstrated potentiation of renal vasoconstriction by ROS generation. Thus contraction of rabbit isolated afferent arterioles to U-46619 was enhanced by vascular ROS and blunted by NO (1301, 1303). The sensitivity of afferent arterioles to ANG II was enhanced markedly in mice with SOD1 (Cu-Zn-SOD) deficiency (183). This effect was primarily attributed to enhanced O2⫺ inactivation of NO. Conversely, prevention of NADPH oxidase-derived O2.⫺ generation in afferent arterioles from p47phox knockout mice blunted ANG II and myogenic contractions (183, 864, 1097), whereas myogenic contractions were preserved in afferent arterioles from eNOS-deficient mice (864). Thus O2⫺ generated by ANG II enhances afferent arteriolar contractions by inactivation of NO, whereas O2.⫺ generated by increased PP enhances myogenic contractions independently of NO. PP increased the generation of O2⫺ from NADPH oxidase in large conduit vessels (1621) and in afferent arterioles from rats (1224) or mice (702, 861, 866, 1224). Cell-permeable polyethylene glycol (PEG)-SOD and tempol both blunted the myogenic responses of mouse isolated afferent arterioles, whereas PEGcatalase was ineffective, thereby implicating O2⫺ rather than H2O2 (866). Tempol also attenuated myogenic responses in the rat isolated perfused HNK (1146). Moreover, prolonged infusion of ANG II increased ROS production that enhanced myogenic response of isolated afferent arteriole (864). Taken together, many studies have implicated O2⫺ generated by NADPH oxidase/p22phox/p47phox NOX-2 during stretch of afferent arterioles in potentiating the myogenic response. However, there are contrary findings. For example, the myogenic response of isolated afferent arterioles of normotensive WKY rats did not require ROS generation, although O2⫺ enhanced the myogenic response of SHR afferent arterioles (1224). Transforming growth factor-1 (TGF-1) perfused into the JMN preparation antagonized myogenic constriction by generation of ROS from NADPH oxidase (1354). However, this growth factor caused prominent basal constriction of the afferent arteriole, which may have obscured an additional myogenic contractile response. Moreover, TGF-1 activity and its ability to increase BP and myogenic tone of mesenteric arteries were normally suppressed by Emilin-1, a protein of elastic extracellular matrix (196). Intrarenal infusion of hypoxanthine plus xanthine oxidase increased O2⫺ and caused vasoconstriction as well as natriuresis, but steady-state RBF autoregulation was Physiol Rev • VOL 95 • APRIL 2015 • www.prv.org CARLSTRÖM ET AL. well maintained (1204). A high-salt diet that activated NADPH oxidase and increased ROS production in kidneys (471, 579) failed to affect the myogenic transfer function in rats and, when given with ANG II, actually blunted the myogenic response (1273). Another study of rats fed 8% NaCl for 2 wk reported that RBF autoregulation was abolished (AI increased from 0.0 to 1.4) and the autoregulation of afferent arterioles of JMN was blunted by an apparent increase in ROS which was restored in part by prolonged treatment with apocynin (396). The reason for these discrepancies is presently unclear but may relate to different effects of O2⫺ and H2O2. Increased ROS formation enhanced myogenic responses in other vascular beds including mesenteric resistance arteries (1530) and tail arterioles (1097). Elevated PP in gracilis muscle arteries activated sphingosine kinase-1 to stimulate production of ROS from NADPH oxidase that enhanced Ca2⫹ sensitivity and myogenic responses likely by activation of Rac and RhoA/Rho kinase and inhibition of MLCP (783). Activation of NADPH oxidases by increased PP in bovine coronary and rat small cerebral arteries enhanced myogenic contractions and/or autoregulation of blood flow by activation of the ERK/MAPK cascade (1104), or by activating PI3K/Akt signaling that reduced BKCa activity, and increased EM and VSMC [Ca2⫹]i (468, 470, 1454). However, low levels of O2⫺ generated by subdural perfusion of xanthine plus xanthine oxidase decreased cerebral blood flow, but did not affect autoregulation (1646), whereas high levels of ROS increased blood flow and inhibited blood flow autoregulation and myogenic constriction by activating BKCa channels that hyperpolarized the VMSC plasma membrane. This suggests an effect of H2O2, which is known to hyperpolarize VSMCs (1646). ROS-mediated vasodilation counteracted myogenic tone in rat ophthalmic arteries, but this effect was partially dependent on intact endothelium, which is a source of H2O2 as well as NO (1543). Indeed, ROS, probably H2O2, impaired endothelium-dependent vasodilation by inhibiting BKCa channels in renal artery endothelial cells (135). H2O2 can increase endothelial NO production (721) and decrease Ca2⫹ sensitivity in aortic VSMCs (663). In summary, ROS impacts renal autoregulation by modulating both myogenic and MD-TGF mechanisms. Most studies of isolated afferent arterioles concluded that the NADPH oxidase/p47phox/NOX-2 signaling pathway is activated by increased RPP and is the major source of O2⫺ that enhances the myogenic constrictor response, whereas H2O2 may inhibit this response. The opposing effects of these two ROS may account for some inconsistences in the reported effects of ROS on autoregulation, but this requires further study. The mechanisms whereby O2⫺ sensitizes contractions involve reduced NO bioavailability, but the effects on the myogenic response likely involve direct VSMC effects to mobilize [Ca2⫹] or increase Ca2⫹ sensitivity. G. Endothelium-Derived Hyperpolarizing Factor and Cxs Endothelial cells control vascular tone by changes in EM transmitted to VSMC and by the release of factors that relax or constrict VSMCs. Endothelial dilator factors include NO, PGs, and EDHF. ACh and bradykinin can elicit vasodilation and buffer vasoconstriction in part via EDHF that hyperpolarizes VSMCs and relaxes intrarenal arterioles independent of the vasodilators NO, PGE2, and PGI2. EDHF stimulated Na⫹-K⫹-ATPase and enhanced hyperpolarization of the renal interlobar artery (172). EDHF in mice was estimated to contribute ⬍20% of the renal vasodilation produced by ACh or bradykinin (299). Little is known about the influence of EDHF on RBF autoregulation. Although the myogenic response is not mediated by endothelial factors, it is likely that endothelial cells can modulate VSMC tone by electrical coupling and production of paracrine agents such as NO as well as EDHF and EETs. ACh-induced EDHF production transiently blunted myogenic vasoconstriction of afferent arterioles in the isolated perfused rat HNK (592) where it stimulated K⫹ channels to modulate afferent but not efferent arteriolar tone, presumably by affecting EM and VOCC activity (1571). Interestingly, the inhibition was transient, reversing after 5–10 min. EETs mediated EDHF actions in the rabbit isolated, perfused afferent arteriole. They were generated by a cytochrome P-450 epoxygenase and elicited vasodilation by opening BKCa channels and activating Na⫹-K⫹-ATPase on VSMCs (1551). A metabolite of cytochrome P-450 epoxygenase, presumably an EET, blunted myogenic constriction of afferent arterioles in the JMN preparation (673). Vasodilatory EETs attenuated the myogenic response of isolated gracilis muscle arterioles (1424). Metabolism of EETs by soluble epoxide hydrolase regulated myogenic response and BP via effect on Ca2⫹ entry via TRPC1 and TRPC4 channels (1209). Other candidates for an EDHF include H2O2 and hydrogen sulfide (H2S) (914, 1449, 1652). Gap junctions composed of Cxs may contribute to endothelium-dependent vasorelaxation via conducted vascular responses. The extent to which cell-to-cell communication depends on direct signaling through myoendothelial gap junctions is controversial. Isolated arteries and arterioles displayed bidirectional electrical coupling between endothelial cells and VSMCs. Vasodilator waves passing via cellto-cell gap junctions can spread to upstream feed arteries that control blood flow into arteriolar networks (60). Cx may mediate this spreading vasodilation elicited by EDHF by transfer of hyperpolarization from endothelium via gap junction Cx to adjacent VSMCs to conduct along the vessels and thereby close their VOCCs. However, such coupling between endothelium and VSMC cells has not been verified in intact animals where myoendothelial gap junc- Physiol Rev • VOL 95 • APRIL 2015 • www.prv.org 451 RENAL AUTOREGULATORY MECHANISMS tions may be controlled by yet unknown mechanisms that impede such signaling (313, 314). In nonrenal vascular beds including aorta, pulmonary, and coronary arteries, Cx40 and Cx37 interacted to form heterotypic channels, and they were codependent on each other, or mutually dependent, for optimal expression and function in vascular endothelium (1368, 1369, 1599, 1633). ACh acting on cremaster arteries in vitro elicits local vasodilation that was conducted upstream via Cx40, but not on Cx37 (738). However, vasodilation produced by ACh in mouse cremaster arterioles in vivo was largely independent of this mechanism (125, 314, 315, 1362). Moreover, ACh produced potent vasodilation in Cx40-deficient gracilis arteries in vitro or in endothelium-specific Cx40 deficient vessels, but this was detected only under isobaric conditions (125). ACh-induced EDHF-like dilation observed in vivo was independent of the presence of Cx40 (756). Thus ACh responses appear to be conducted along VSMCs, but not between endothelial cells. Cxs at myoendothelial gap junctions of small arteries in vitro modulated the effects of NO on Ca2⫹ signal propagation which prolonged increases in [Ca2⫹]i in endothelial cells and accelerated Ca2⫹ signal transmission in VSMCs (1192), but this was independent of Cx37 (419). In summary, the roles of Cxs in the EDHF response and in myogenic tone are unclear. The role of gap junctions and Cx in the conduction of vasoactive signals along the vascular tree of the microcirculation requires further investigation and resolution concerning differences in results obtained in vitro and in vivo. H. Modulation of Myogenic Responses by Agents Released From Activated MD or CT Cells Both MD, and some CT, cells abut the afferent arteriole supplying the parent glomerulus. As discussed earlier in section IIA3, MD cell signaling is activated by reabsorption via luminal NKCC2 and Na⫹/H⫹ exchange. As summarized in FIGURE 6, MD-derived metabolites can either enhance (ATP, adenosine, PGH2, TxA2, and O2⫺) or inhibit (NO, CO, and PGE2) afferent arteriolar tone. As discussed in section IIB2, CT cell signaling can be activated by reabsorption via luminal ENaC which release EETs and PGE2 that attenuate afferent arteriolar tone (FIGURE 6). The resultant vasodilation from CT-GF activation may contribute to resetting of MD-TGF during salt loading (1554). Such rich paracrine signaling in the JGA provides for complex interactions between the tubular-glomerular systems and myogenic mechanism of vascular control and contributes to the positive and negative interactions that were described in earlier sections. 452 VII. RENAL AUTOREGULATION IN DISEASE: HYPERTENSION, RENAL DISEASE, AND DIABETES A. Introduction Studies in the 1930s demonstrated that hypertension is associated with arteriolosclerosis of the small renal arterioles and renal tubulointerstitial inflammation, fibrosis, and glomerulosclerosis (795, 1033). There were two contrasting theories concerning the principal causes and the secondary compensations (388, 1033, 1541). One held that hypertension is primary and induces secondary renal and vascular abnormalities. In 1937, Moritz and Old (1033) reported that renal afferent arteriopathy was present in 98% of autopsied kidneys from subjects who had hypertension but in only 15% of these who were normotensive. They concluded that the renal arterial lesions are a consequence of the hypertension. The earliest structural changes with hypertension are in the smallest resistance arterioles, a site where a myogenic response plays a fundamental role. Hypertension is associated with structural remodeling of the vascular wall and vascular hyalinization, but the contribution of wall remodeling to functional changes in myogenic responses is uncertain. The second theory held that renal inflammation and afferent arteriopathy are primary and induce hypertension (1033). In 1934, Goldblatt (476) showed convincingly that creating a renal artery stenosis to reduce the RPP leads to hypertension. Renal artery stenosis, whether caused by atherosclerosis or fibromuscular disease, is detected in 5–10% of patients with hypertension (1455). However, recent controlled clinical trials of renal artery angioplasty and stenting to correct a stenosis have concluded that hypertension, although often lessened in severity, is only rarely cured (72). Thus the debate remains incompletely resolved. Structural renal arterial lesions that limit RPP clearly raise BP. However, the renal arteriolar remodeling that accompanies sustained hypertension likely impairs regulation of RBF and RPP and increases RVR and may thereby contribute to sustaining the hypertension. Thus the structural renal arterial changes can be viewed as a consequence of hypertension, but in some settings, likely also contribute to the development or severity of the hypertension. Therefore, they are of considerable importance. Glomerular hyperfiltration refers to an increase in the SNGFR, but this is not necessarily accompanied by glomerular hypertension, i.e., an increase in the PGC (601). Schmieder and co-workers (1297, 1298) suggested that glomerular hyperfiltration occurs early in hypertensive patients with target-organ damage, and Campese et al. (177) concluded that PGC is likely increased in a group of salt-sensitive hypertensive patients that included many African-Americans. An impaired renal autoregulatory response in hypertensive African-Americans (FIGURE 2A) may contribute to an increased PGC (826). Physiol Rev • VOL 95 • APRIL 2015 • www.prv.org CARLSTRÖM ET AL. Many rodent hypertensive models eventually develop CKD (104). However, the development of CKD, unless associated with malignant hypertension, is uncommon in humans except in African-Americans. Recent studies report that the primary problem in African-Americans with CKD is inheritance of an abnormal genotype that confers increased risk of renal damage. Hypertension is usually a secondary consequence (909). Nevertheless, it is clear that once renal damage has been sustained, hypertension is one factor predicting the rate of further decline in renal function, especially in those with heavy proteinuria (803). Autoregulation normally protects the glomerular and tubulointerstitial tissues from hypertensive injury (108, 112, 933). However, although autoregulation usually adapts to higher BPs in hypertensive models, its efficiency often declines over time. The responsible mechanisms are poorly understood. Some, but not all, models of hypertension, CKD, or diabetes mellitus (DM) have impaired renal autoregulation that permits transmission of more of the BP fluctuations to the glomeruli where it increases the PGC. The degree of glomerular injury in hypertensive models generally increases in proportion to the impairment in renal autoregulation (106, 107, 110). Bidani, Kriz, Byrom, and colleagues (103–110, 173, 839, 840, 1162) proposed that hypertension combined with impaired renal autoregulation cause renal barotrauma and enhance the progression of CKD. Kriz et al. (838) demonstrated that an initial podocyte injury from barotrauma in the context of FSGS misdirects filtration into the interstitium, thereby initiating a spreading renal tubulointerstitial inflammation and later fibrosis with degeneration or fibrosis of the glomerulus and its attached tubule. It follows that the ideal therapy for hypertension should lower the BP while preserving, or improving, the effectiveness of the renal autoregulation (105, 106, 110, 500). Ideally, the preglomerular vasculature should protect the kidney from both sustained hypertension and from rapid oscillatory fluctuations in BP (108, 109, 927, 934) since both increased BP and increased pulse pressure predict hypertensive renal damage (108, 927, 933). Examples of impaired autoregulation in disease models include rodent remnant kidney models of CKD with prolonged RRM, some, but not all, rodent insulinopenic or obesity-associated models of DM, or Dahl salt-sensitive or Goldblatt ischemic renovascular hypertensive models. In contrast, SHR retain highly efficient autoregulation throughout life that likely contributes to their resistance to proteinuria and glomerulosclerosis even with established and prolonged hypertension. Renal cross-transplantation between BN rats and SHR proved that the protection from glomerular damage in the SHR was intrinsic to their kidneys (260), which corresponds to the excellent autoregulatory adjustments of the preglomerular vasculature of the SHR but the much weaker adjustments of BN rats. A similar kidney cross-transplantation study demonstrated that the kidneys from stroke-prone SHR (SHR-SP) exhibit more hypertensive damage and malignant nephrosclerosis than those of SHR during a high-salt diet (261, 498). Moreover, poor RBF autoregulation in Fawn-Hooded (FH) rats is improved by replacing chromosome 1 genetic loci from BN rats (922, 1610). Roman, Provoost, and colleagues have provided extensive information concerning the mechanisms, and their genetic components, that underlie autoregulation in many hypertensive models, as described in detail under the individual models below. These studies are important because they have established that the efficiency of renal autoregulation and the associated protection of the kidneys from hypertensive damage are heritable. B. Hypertension 1. SHR SHR are considered a model of human essential hypertension (1115, 1483). The strain was developed from inbreeding of selected rats that developed hypertension spontaneously. Hypertension develops progressively over 4 – 8 wk while fed a normal-salt diet (161, 386, 744, 867, 1009, 1389, 1467). The hypertension is exacerbated by a high salt intake (34, 256, 325, 326, 447, 498, 880, 917, 978, 1406). The degree of elevation in BP was quite variable among studies. Factors likely contributing to this variability are the methodology for BP measurement, the animals initial age (6 –14 wk), duration of treatment (2–28 wk), the amount of salt in the diet (ranging from 1 to 8%), as well as the genetic background of the SHR and control WKY colonies (848, 849, 1428). Cross-transplantation of kidneys between SHR and normotensive controls established that the kidney of SHR is the primary cause of the development of hypertension (506, 575, 779, 1158, 1235, 1236). Renal hemodynamic abnormalities are important in the initiation of hypertension in the SHR. Basal GFR and RBF are reduced during the development of hypertension in young SHR (4 –10 wk old) (335, 386, 567, 574, 1038, 1252, 1507, 1526), but are restored to the levels of normotensive WKY rats during the established phase at 12–20 wk (42, 418, 443, 574, 809, 1246, 1252). A rightward shift in the pressure-natriuresis curve to a higher level of RPP (43, 443, 997, 1093, 1246) led to an early period of relative Na⫹ retention during the development of hypertension (79, 570, 571, 605, 940). A genetic link between abnormal renal hemodynamics and the development of hypertension was established in cross-breeding studies between SHR and WKY. Renal vasoconstriction (reduced RBF and GFR) and elevated BP cosegregated (572, 710) and was evident during the early development phase of hypertension, suggesting causality. This link has been investigated extensively. Cosegregation of the renin allele of SHR with increased BP has been noted Physiol Rev • VOL 95 • APRIL 2015 • www.prv.org 453 RENAL AUTOREGULATORY MECHANISMS (850, 1427, 1641), and expression of the SA region on chromosome 1 is enhanced in hypertensive SHR (693, 711, 712, 906, 1405), in particular in the proximal tubule (1631). However, studies using congenic SHR carrying a smaller segment of BN chromosome 1 but without the SA gene concluded that molecular variation in this gene was not required for the effect of this region of chromosome 1 on blood pressure (1405). The RAS is especially important in the developmental phase of hypertension. ACE inhibition in young SHR for 4 wk between 2 and 10 wk of age normalized the exaggerated renal vasoconstriction and has long lasting effects in preventing full expression of hypertension later in life (250, 354, 417, 472, 567, 568, 573, 574, 939), beneficial effects independent of changes in renal vasculature structure (786). Administration of an AT1 receptor blocker from 3 to 8 wk of age led to a sustained reduction in BP in adult SHR (95, 1103). Although less well studied, blockade of the RAS may be efficacious in reversing established hypertension (355, 393, 939). Renal nerves have an important role in the pathogenesis of hypertension in the SHR. Renal nerve activity was elevated in SHR at the age of 8 –10 wk (746, 941). Denervation of the kidneys in young (4 – 8 wk old) SHR delayed the development of hypertension (606, 807, 1092, 1617) (775), due at least in part to increased Na⫹ excretion (606, 1268). Young (4 – 6 wk old) SHR had excessive preglomerular vasoconstriction that reduced RBF and GFR (43, 276, 1252, 1507). The lower GFR was ascribed to reductions in the glomerular blood flow and the glomerular hydrostatic filtration coefficient (KUF) while the PGC was maintained by increased afferent and efferent arteriolar resistances (335). Hypertensive SHR between 12 and 16 wk of age have increased preglomerular vascular resistance that parallels the increase in BP (42, 58, 358, 639, 704, 709, 1471), mediates efficient autoregulation of RBF and GFR (8, 41, 107, 276, 640, 709, 1246, 1507), and maintains PGC in the normal range (42, 58, 358, 704, 852, 1471) and normal glomerular structure (787). The total number of glomeruli and mean glomerular volume were similar in kidneys of SHR and WKY at 4 wk of age (115). Albuminuria developed only after ⬃20 wk of age. Although the PGC was normal in 12- to 16-wk-old SHR (1471), it tended to increase in some (1471), but not all (394), studies by 36 mo, while the vascular and glomerular morphology remained normal (1471). Thus glomerular hypertension can precede glomerular injury in SHR. Remarkably, in another study, the PGC of 15-mo-old SHR was lower than age-matched WKY (394). The administration of low doses of a CCB or an ACE inhibitor to SHR reduced their BP, increased their RBF and 454 GFR (412– 414, 567, 574, 765, 1056, 1098) and SNGFR (435), and normalized their preglomerular and efferent arteriolar resistances (1056, 1098). Higher doses of a CCB that blocked both L- and T-type VOCC abolished renal autoregulation and rendered PGC highly dependent on BP (365, 500, 852, 1056). These data suggest that the use of CCBs to treat hypertension in patients with CKD is less protective of renal function than ACE inhibitors, as confirmed in the African-American Study of Kidney Disease (AASK) trial (909) and in experimental studies in rats (29, 365, 500, 503). Moreover, administration of an ACE inhibitor or an AT1 receptor blocker to young SHR caused a predominant reduction in efferent arteriolar resistance, thereby reducing PGC and the lower limit of renal autoregulation (852). Young SHR have exaggerated renal vascular reactivity to ANG II (44, 220, 222, 311, 787, 824, 825, 1409, 1542), ATP (1434), catecholamines (787, 813, 1385, 1498), and AVP (404) and attenuated vasodilation to PGE2, PGI2 (219, 223, 224, 716, 717), and agonists of dopamine D1 receptors (225). Thus vasodilation and reduced buffering of vasoconstriction by GPCRs signaling and activation of the cAMP/ PKA pathway are defective in the renal vasculature of SHR and contribute to the exaggerated vasoconstriction. The exaggerated renal vascular reactivity to ANG II was largely due to abnormal blunting by vasodilator PGs (217, 219, 221, 224, 716, 717), since the density of AT1 receptors in the renal vasculature was similar in SHR and WKY (216, 220). However, vasodilation to ACh, sodium nitroprusside, or bradykinin that is mediated by the cGMP/PKG pathway was preserved or even exaggerated in young SHR (174, 225). Indeed, inhibition of NOS led to stronger increases in RVR and reductions in RBF in adult SHR (10 –16 wk old) than in WKY in some studies (96, 432, 853, 1203), implying enhanced regulation of the renal circulation by NO in the SHR with established hypertension, whereas others reported a normal degree of vasoconstriction in adult and aged (70 wk old) SHR (311, 432, 853). Renal autoregulation operates normally in young and mature SHR. Unlike most other hypertensive models, the RBF and GFR of SHR are very efficiently autoregulated with maintained, or even an exaggerated, with strong influence of myogenic and MD-TGF components throughout the established phase of hypertension (707, 1567). Steady-state experiments demonstrated that SHR autoregulate their RBF effectively with a normal plateau phase over a defined range of RPP as they age from 4 – 6 wk (1252) to 10 –16 wk (8, 42, 598, 640, 853, 1203, 1246, 1252, 1567) or even up to 40 (708, 709, 852, 1567) to 70 wk (853). The plateau phase of RBF autoregulation was unaffected by inhibition of NO (853, 1203), the RAS (352, 1565), ET-1 and ETA receptors (140), COX metabolites, ROS (815), or renal nerve activity (708). Whereas autoregulation was not affected by endogenous ET-1 in normotensive animals (97, Physiol Rev • VOL 95 • APRIL 2015 • www.prv.org CARLSTRÖM ET AL. 140, 831), it was improved in SHR after short-term ETAreceptor blockade (140). However, in animals with established hypertension, these systems contribute to switching the autoregulatory range to higher levels of RPP (705–707, 709, 852, 997, 1188, 1203, 1565–1567). The lower limit of RPP for autoregulation was preserved in young hypertensive SHR (41, 1252). Whereas SHR exhibited excellent autoregulation of renal and outer cortical blood flow (704), JMN have increased PGC with less efficient and more sluggish autoregulation. Thus the PGC of JMN of SHR was 5 mmHg higher than cortical nephrons at 10 wk of age and increased to 10 mmHg higher at 70 wk. The higher PGC corresponded with an earlier development of glomerulosclerosis and with larger diameters of glomerular capillaries in JMN (704). Moreover, an abrupt step increase in RPP produced a rapid adaptive increase in vascular resistance in the superficial renal cortex but a slower, less complete response in the juxtamedullary region (1240). Autoregulation of medullary blood flow was well established in young SHR (1252). By 12–16 wk, the medullary blood flow and the RIHP were reduced in SHR, accompanied by a rightward shift in the pressure-natriuresis relation (412). At that age, autoregulation of medullary blood flow was diminished during acute volume expansion, although excellent autoregulation of whole kidney RBF persists (352). Since medullary blood flow contributes ⬍10% to total RBF, small changes may not be detected in measurements of total RBF. These findings are consistent with the hypothesis of Cowley et al. (274) that an attenuation of medullary blood flow autoregulation during volume expansion enhances the strength of the pressure-natriuresis relation and thereby modulates the level of BP, as was discussed in section IIA2 and is illustrated in FIGURE 4B. Medullary blood flow was especially sensitive to ANG II in SHR during the development phase of hypertension (353). ACE inhibition from 4 to 14 wk of age in volume-expanded SHR attenuated the autoregulation of the medullary blood flow despite a maintained autoregulation of RBF (352). Administration of an AT1 receptor blocker to SHR between 4 and 12 wk of age reduced BP, increased medullary blood flow, and reduced the number of pericytes surrounding vasa recta capillaries (70). These treatments also impaired medullary blood flow autoregulation and increased the RIHP which may contribute to the resetting of the pressure-natriuresis relationship to a lower level of RPP, and limiting the development of hypertension (997, 1567). Indeed, most studies employing infusions of CCBs, ACE inhibitors, tempol, lovastatin, or L-arginine concluded that SHR have a reduced NO and a reduced medullary perfusion secondary to increased medullary oxidative stress and that could account for a rightward resetting of the pressure-natriuresis relationship and a higher ambient BP (276, 406, 412, 733, 872). These studies indicate NO is important in maintaining the medullary blood flow in SHR (96), and a reduction in NO contributes to the lower medullary blood flow and reduced pressure-natriuresis relation in SHR (872). Other studies, however, suggested that NOS activity was elevated in the renal medulla of SHR (585) and its inhibition produced a greater reduction in medullary blood flow in SHR than in WKY (96). Collectively, these data suggest an important role for ANG II and the balance between ROS and NO in the renal medulla during the development of hypertension in SHR. Indeed, several studies have reported enhanced interaction between ANG II, NO, and ROS in renal autoregulation of SHR. A strong MD-TGF response may contribute to the resilient renal autoregulation in SHR. Young (6 wk old) SHR had exaggerated MD-TGF response (333) that was attributed to ANG II acting on AT1 receptors and to a vasoconstrictor eicosanoid acting on TP receptors (138, 139). MD-TGF activity reverted towards normal during established hypertension in adult SHR from the NIH/Chapel Hill colony (333) and from some commercial suppliers (231), but remained exaggerated in adult (12–18 wk of age) SHR from some other sources (894, 1587, 1592). However, there is genetic heterogeneity between colonies of SHR, and especially WKY (848). Increased MD-TGF response in adult SHR was attributed to AT1 receptor activation and oxidative stress (894, 1587, 1588, 1592) and weak buffering by NO. Although SHR had increased renal cortical expression of endothelial NOS and of MD nNOS, neither global inhibition of NOS nor relatively selective inhibition of nNOS with 7-nitroindazole increased the MD-TGF in SHR as it did in WKY (1464, 1587, 1588). On the other hand, peritubular capillary perfusion of tempol blunted MD-TGF activity more in SHR than WKY and restored the effect of nNOS inhibition to enhance MD-TGF in SHR (1588). This suggests that the JGA of hypertensive SHR produces excess O2⫺ that bioinactivates MD-derived NO. O2⫺ in the JGA was dependent on tubular fluid reabsorption activating NADPH oxidase NOX-2 in MD cells (448, 1592). Excess O2⫺ diminished dilation of juxtamedullary afferent arterioles by neuronal NOS-derived NO in SHR but not in WKY (655). Thus SHR generally have enhanced MD-TGF responses that are related to oxidative stress in the JGA that inactivate NO, thereby contributing to their efficient renal autoregulation. Vasoconstriction from activation of MD-TGF can be propagated upstream along the afferent arteriole to reduce the perfusion and the PGC in adjacent nephrons supplied by a common cortical radial artery. The magnitude of such a MD-TGF-initiated nephron-nephron interaction was stronger in SHR, implicating greater signal transduction (231). Chon, Holstein-Rathlou, Marsh, and associates demonstrated that MD-TGF signaling to the parent glomerulus caused oscillations in preglomerular vascular tone and PPT Physiol Rev • VOL 95 • APRIL 2015 • www.prv.org 455 RENAL AUTOREGULATORY MECHANISMS that were more time-variant and irregular in SHR, indicative of more complex coupling between MD-TGF and myogenic mechanisms (230, 244, 247, 625, 626, 1205, 1635, 1672). Thus studies of transfer function analysis generally confirmed an enhanced MD-TGF, and perhaps myogenic component, of autoregulation in SHR, which was consistent with increased ROS and reduced NO signaling in the JGA. Myogenic constriction of isolated perfused afferent arterioles was enhanced in both young and adult SHR (700, 1224). Thus exaggerated myogenic tone seems to be an intrinsic functional abnormality evident during the development of hypertension in young SHR and not a consequence of long-standing hypertension in adult SHR. The PP threshold for a myogenic contraction in the afferent arteriole of the HNK was shifted 40 mmHg higher, while the maximum myogenic constriction was preserved (593). NOS inhibition lowered the threshold PP by 20 mmHg in WKY and by 40 mmHg in SHR (15 wk old), whereas the maximum myogenic response was augmented to a similar degree in both strains (593). Thus NO can modulate the myogenic response with different effects in the renal vasculature of SHR and WKY, perhaps due to different actions on pre- and postglomerular arterioles. Perfusion of cortical radial arteries and afferent arterioles of JMN with a physiological salt solution revealed enhanced myogenic contractions in SHR as young as 3– 4 wk of age (465, 672), which were related to enhanced generation of a cytochrome P-450 metabolite (469, 672), presumably the vasoconstrictor 20HETE. Oxidative stress is frequently associated with increased vascular reactivity, hypertension, and renal failure (1330, 1529). Prolonged tempol administration reduced BP in SHR and shifted the pressure-natriuresis relationship to the left (406, 1300, 1302, 1585). Tempol increased basal RBF in hypertensive SHR, reduced exaggerated renal vascular reactivity to ANG II, independent of NO, suggesting a direct effect of O2.⫺ on vascular responsiveness (311). Tempol quenching of O2.⫺ increased medullary blood flow without perturbing RBF autoregulation in adult SHR (311, 406). Thus ROS apparently restrains renal and medullary blood flow in the SHR without major effects on autoregulation. The afferent arteriole of adult SHR has a fourfold increase in ROS production by NOX-2 (1224). Thus, bioinactivation of NO by O2.⫺ in the JGA of SHR is sufficient to prevent endogenous NO from blunting MD-TGF. It also may explain the lack of effect of NOS inhibition to magnify the renal myogenic response of SHR as it did in normotensive Long-Evans, Sprague-Dawley and Wistar rats (1569). The strong myogenic response of young SHR was sufficient 456 to maintain RBF autoregulation after complete inhibition of MD-TGF by ureteral obstruction (297). Nevertheless, whereas NO is considered a strong modulator of the myogenic component of renal autoregulation in normotensive rat strains, the modulation was weak or absent in the SHR (1569). This indicates a separation of the role of NO to blunt vasoconstriction (evident in SHR and normotensive strains) and to blunt the myogenic response (absent in SHR). Nevertheless, efficient RBF autoregulation and the myogenic response in SHR was preserved in the absence of MD-TGF input during ureteral obstruction (297) and persisted after damage to mesangial cells by antithymocyte antibodies in both SHR and WKY, whereas autoregulation of GFR was attenuated (1564). Thus, although mesangial cells can participate in the regulation of GFR, likely via MD-TGF signaling, they do not seem to contribute to RBF autoregulation, which, in their absence, is maintained by enhanced myogenic tone. SHR also have enhanced myogenic tone in nonrenal vessels including cremaster muscle (389, 643, 644, 1359), basilar (18), cerebral (127, 484, 564, 566, 725, 1469), mesenteric (472, 714), and coronary arteries (458). Enhanced myogenic tone in these nonrenal vessels of SHR has been related to increased Ca2⫹ entry due to depolarization of the plasma membrane (564) and elevated VOCC activity (1359), perhaps secondary to reduced EM and reduced K⫹ channel activity (1469). Also contributing were increased Ca2⫹ mobilization and Ca2⫹ sensitivity due to augmented Rho kinase activity (18, 725, 1164). Enhanced myogenic tone was dependent on increased endothelial-derived ET-1 and PGH2 (643, 644) and reduced NO (484), or other endothelial factors (458). The enhanced myogenic responses occurred without vascular remodeling (389) and were not secondary to hypertension since they predated its onset (127, 714, 1359). Mechanisms that enhanced myogenic tone in these nonrenal vessels of SHR may also occur in the renal microcirculation. However, these effects were specific for resistance vessels since normal myogenic responses are reported in larger conduit vessels from SHR (166, 167, 1084). Remodeling (reorganization of the vessel wall components) of small arteries can limit organ blood flow (425, 1044, 1322) and may contribute to exaggerated renal vasoconstriction. An increased cross-sectional area of the preglomerular arterial media was apparent in young SHR before the development of hypertension (1385). Furthermore, some of the second generation offspring of rats crossbred between SHR and normotensive WKY had a narrowed afferent arteriole lumen diameter at 7 wk which predicted the later development of high BP at 21–23 wk (1094, 1380). This suggests that a narrowed renal afferent arteriole, as Physiol Rev • VOL 95 • APRIL 2015 • www.prv.org CARLSTRÖM ET AL. from remodeling, can contribute to the development of sustained hypertension. Most, but not all (1381), studies reported hypertrophic remodeling of the media with luminal narrowing of cortical radial arteries of SHR that may persist (33) or reverse with prolonged ACE inhibition (1379) and may originate from enhanced sympathetic nerve activity (1474). Thus a variable degree of genetically and neurally determined remodeling of the afferent arteriole of the SHR could restrict the transmission of BP into the kidneys, thereby protecting the glomeruli from barotrauma and maintaining the set-point for pressure-natriuresis at ambient level of RPP. Other studies have identified increased wall thickness in the arcuate and cortical radial arteries of 5- or 10-wk-old SHR but not in the afferent arteriole at either age (33, 1385). The ratio of wall thickness to radius of the afferent arterioles in the inner and outer cortex was not different between SHR and WKY at 6 and 12 wk of age (461). Another morphological study reported that afferent arterioles of 12-wk-old SHR had a smaller, not larger, media cross-sectional area than age-matched WKY (1381). Aged SHR have hypertrophy of the afferent arteriolar wall (882). A hallmark of the SHR is excellent autoregulation of whole kidney and outer cortical blood flow and PGC, which may account for the paucity of glomerulosclerosis until up to 70 wk of age, despite increasing hypertension. Excessive NADPH oxidase-mediated O2⫺ production and downregulation of SOD and other antioxidant enzymes in the kidney of SHR (12, 114, 212, 1649) likely contributes to the enhancement of myogenic and MD-TGF mechanisms. Much, but not all, of the effects of O2⫺ in the JGA were secondary to reduced NO activity (1588). However, as SHR age, their cortical autoregulation eventually becomes slightly less efficient and their outer cortical glomeruli develop modest glomerulosclerosis. The earlier impairment of autoregulation in JMN may account for their earlier development of glomerulosclerosis. Nonrenal resistance arterioles of SHR also generally display enhanced myogenic tone, suggesting that it is a fundamental property of VSMCs in this rat strain. Since augmented myogenic responses of renal and systemic microvessels of young SHR precede the development of hypertension, they are likely primary and genetically determined. A primary increase in O2⫺ with bioinactivation of NO in the renal cortex and medulla of the SHR could contribute to the excellent autoregulation and reduction of transmission of BP into the renal parenchyma. This should buffer the effects of increased BP to raise medullary blood flow and RIHP in the SHR, thereby attenuating the pressure natriuresis and sustaining the hypertension on the one hand, yet protecting the renal parenchyma from the effects of hypertensive barotrauma on the other. 2. Stroke-prone SHR Stroke-prone SHR (SHR-SP) have a genetic predisposition to stroke, a more rapid rise in BP, and a more pronounced salt-sensitivity than SHR (1048, 1116). They were normotensive at 4 wk of age and developed hypertension before 9 wk (908, 1049). Kidneys of SHR-SP required a higher BP than WKY to excrete a given amount of salt and water (1049). Marked proteinuria was evident by 12–16 wk of age (1055, 1478). Glomerulosclerosis was present by 24 wk of age (1055). At 15 mo, the kidneys of SHR-SP on a normal-salt diet displayed reduced GFR, arterial wall hypertrophy and sclerosis, glomerulopathy, and tubular interstitial injury, all of which were prevented by normalization of the BP during ACE inhibition since 4 wk of age (908). Remarakbly, this nephroprotection was associated with almost a doubling of the lifespan. When given 1– 4% NaCl to drink, they develop severe hypertension and usually die from stroke within 14 –20 wk. Transplantation of a kidney from an SHR-SP into an SHR accelerated the rise of BP, the reduction of RBF and GFR (1049), and the development of renal damage (498). Thus SHR-SP kidneys have a genetic propensity to cause hypertension and also an enhanced susceptibility to hypertensive injury (261). The basal RBF and GFR were reduced and the RVR increased in hypertensive 9- or 12-wk-old SHR-SP relative to normotensive controls (1049, 1055). The BP and the RBF increased with a high-salt diet (8). The renal vasculature of 4-, 8-, and 16-wk-old SHR-SP was hyperresponsive to ANG II, AVP, NE (94, 1050), and ET-1 but not to AVP (457). Dopamine D1 receptor stimulation produced enhanced vasodilation via PKC/cAMP signaling in intrarenal arteries of SHR-SP (456). Acute CCB reduced BP and increased RBF without affecting GFR (1031). Six-month-old SHR-SP given CCB for 3 mo had reduced BP with attenuation of the development of glomerulosclerosis, reduced vascular wall thickness, less glomerular and interstitial fibrosis, and increased glomerular volume (692). Similar beneficial structural effects were observed when BP was reduced by blockade of adrenoceptors. ETA receptor blockade starting at 8 wk of age attenuated progressive decline in renal function (measured as creatinine clearance), delayed proteinuria, and doubled the life-span of both male and female SHR-SP on a 4% NaCl intake (1478). Such functional changes took place even though antagonism of ETA receptors had little or no effect on BP, as systolic BP (tail-cuff) rose to 240 mmHg in both treated and untreated animals. AT1 receptor blockade starting at 12 wk reduced BP, normalized RBF and GFR in SHR-SP to WKY levels, and reduced the index of glomerulosclerosis (1055). Prolonged TP receptor antagonism from either 7 or 16 wk of age had no effect on BP, development of proteinuria, cerebrovascular lesions, or stroke in SHR SP drinking 1% NaCl (741). Physiol Rev • VOL 95 • APRIL 2015 • www.prv.org 457 RENAL AUTOREGULATORY MECHANISMS The steady-state values and the autoregulation of total and cortical blood flow of SHR-SP were well maintained during normal salt diet (8, 94, 646), but the autoregulation of medullary blood flow was weak (646). As expected, a CCB abolished autoregulation of renal cortical blood flow (646). Another study reported moderate efficiency of RBF autoregulation with an AI of 0.5 in 9-wk-old SHR-SP (1049). Transfer function analysis confirmed a well-preserved renal autoregulation in conscious 12-wk-old SHR-SP consuming a normal-salt diet, but less efficient autoregulation after an 8% NaCl diet for 3–5 days (8). The sharp increase in BP and the loss of renal autoregulation may contribute to the severe salt-dependent renal damage (8). Enhanced renal vasoconstriction and MD-TGF activity in SHR-SP nephrons were normalized by acute renal denervation (1562). Cortical radial arteries of the HNK preparation retained a normal myogenic response in young SHR-SP but an increased PP threshold (586). Hayashi et al. (587) reported that the myogenic reactivity of the cortical radial artery of the HNK increased along the length of the vessel and that SHR-SP exhibited augmented responsiveness of the intermediate but not the distal segment. This was evident by constriction at a lower threshold PP and greater maximum reductions in diameter compared with WKY. Cerebral arteries of 12-wk-old SHR-SP displayed medial hypertrophy and remodeling, together with attenuated myogenic response (715). Moreover, loss of cerebral blood flow autoregulation was observed prior to the event of stroke (1387). Interestingly, RAS inhibition preserved cerebral autoregulation and the myogenic response and delayed the onset of stroke (1133, 1134, 1559), largely independently of BP (1384). Thus a high-salt diet impairs autoregulation of the renal and cerebral blood flow of SHR-SP that likely contributes to the severe hypertensive renal damage and the propensity for stroke. 3. Dahl salt-sensitive rat Three-week-old Dahl salt-sensitive (DS) and Dahl salt-resistant (DR) rats, when nephrogenesis is still ongoing, were normotensive when receiving a low-salt diet (742), but this has not been studied by telemetry in conscious rats. Renal injuries developed before the loss of dynamic renal autoregulation (769). Indeed, prehypertensive DS rats of the JR (J Rapp) strain already had albuminuria, and glomerular damage accelerated once hypertension was established (1418). Moreover, the GFR of DS rats, even when fed a low-salt diet, was reduced by 6 wk (1526). When fed a high-salt diet, the BP of DS rats increased and both RBF and GFR decreased (642) leading to glomerular hypertension, progressive proteinuria, and renal insufficiency (229, 1208, 1418). This was accompanied by intimal thickening and fibrinoid necrosis (deposits of immune complexes and fibrin) with hyperplasia and necrotizing arteritis (i.e., inflammation of the vascular wall) of the large preglomerular 458 vessels (551, 788), all characteristic for human malignant hypertension. Medullary interstitial fibrosis and tubular necrosis were also severe (278). Many DS rats died after 4 – 8 wk treatment with a high-salt diet (229, 1208). Thus the DS rat has severe structural and functional changes in its renal vasculature that are apparent very early in life even when fed a low-salt diet to prevent hypertension. A switch to a high-salt diet leads to malignant hypertension with pronounced renal damage. Kidney cross-transplantation between DS and DR rats demonstrated that the genotype of the donor kidney determined the BP of the recipient rat (259, 292). Single chromosome substitutions from normotensive BN into DS rats demonstrated that the development of hypertension and proteinuria were determined by genes on chromosomes 1, 5, 7, 8, 13, and 18 in males and on chromosomes 1 and 5 in females, although several chromosomes in males and females affected the development of albuminuria without affecting BP (988). Transfer of the region of chromosome 5 that contained the gene for cytochrome P-450 – 4A from Lewis into male DS rats increased 20-HETE in the outer medulla and attenuated hypertension and renal injury (1611). These responses may relate to the natriuretic actions of 20-HETE (1250). Thus multiple sex-specific genes contribute to hypertension in DS rats. Since some of these differ from those determining CKD, there is a genetically determined dissociation of renal damage from hypertension, but the specific genes involved have yet to be identified. This is somewhat analogous to African-Americans who have a much increased risk of developing hypertensive renal damage (nephrosclerosis), yet lowering their BP below normal failed to slow the progressive loss of their GFR (13), perhaps because of inherited alleles from genes such as apolipoprotein L-1 (APOL-1) or the nonmuscle myosin heavy chain 9 (MYH9) that confer genetic risk for renal damage in African-Americans specifically (442, 909, 1156). RBF and GFR were similar initially in prehypertensive and moderately hypertensive DS and DR rats (1255), but became reduced in DS rats with the development of hypertension and proteinuria during salt loading (229, 259, 769, 1244, 1445). The renal vasculature of DS rats was hyperresponsive to vasoconstriction by ANG II, ET-1, and NE during a high-salt diet (517, 1366) but failed to dilate with atrial natriuretic peptide (ANP) or NO (1366). This vascular phenotype was genetically determined since it was corrected in consomic DS rats receiving chromosome 13 from BN rats (278). The development of hypertension in DS rats was preceded by a shift in the pressure-natriuresis relationship toward a higher BP (1244, 1251), implying a renal mechanism and Na⫹ retention. Normotensive DR and DS rats fed a low-salt diet have effective steady-state whole kidney and outer cortical and medullary blood flow autoregulation (1251) which were maintained in DS receiving a high-salt diet in some (1244, 1445) but not all studies Physiol Rev • VOL 95 • APRIL 2015 • www.prv.org CARLSTRÖM ET AL. (1251). The plateau phase of RBF autoregulation was maintained, although the inflection point for cortical blood flow autoregulation was elevated ⬃30 – 40 mmHg (1244, 1251, 1445). However, others studies reported that neither the renal medullary nor the papillary blood flow was autoregulated in either DS or DR rats whether fed low-salt (1244, 1251, 1366) or high-salt diets (1251), which suggests that the salt-sensitive hypertension in DS rats is independent of changes in medullary autoregulation. Nevertheless, another study reported a high-salt diet fed to DS rats impaired RBF autoregulation and related this to AT1 receptor-mediated ROS production (1286). Moreover, frequency analysis of dynamic RBF autoregulation in DS rats revealed normal autoregulatory pattern of transfer gain when normotensive on a low-salt diet, but a progressive decline in the efficacy of autoregulation when hypertension was induced by a highsalt diet, especially when initiated 2 wk after weaning (769). The absence of dynamic RBF autoregulation in the DS rats with early-onset hypertension was evident as both the myogenic and TGF contributions were defective. The observation that vascular injury preceded the loss of whole kidney autoregulation led to the conclusion that the impaired autoregulation was the result, not the cause, of the renal failure (769). This may explain the variable reports of renal autoregulation described above. Oxidative stress contributes to vascular damage and abnormal renal hemodynamics and Na⫹ excretion in DS rats. NADPH oxidase was overexpressed in the renal cortex of DS rats fed a high-salt diet (449). DS rats have enhanced O2⫺ production in the outer medullary TAL of Henle’s loop that reduced the bioavailability of medullary NO (1030, 1453). This could reduce medullary perfusion and attenuate the pressure-natriuresis (424). Indeed, restoration of NO in DS rats normalized the medullary blood flow and prevented hypertension (1014). MD-TGF was normal in DS and DR rats even on a high-salt diet (770). A mathematical model led to the conclusion that hypertensive DS rats have a severely reduced efficiency of the myogenic mechanism, whereas the MD-TGF response was normal (811). However, hypertensive DS rats also have an enhanced CT-GF response that buffers renal vasoconstriction (1555). This may contribute to vasodilation, increased PGC, and glomerular damage in hypertensive DS rats fed with high-salt diet. Afferent arterioles in the HNK preparation of both normotensive and hypertensive DS rats have greatly diminished myogenic responsiveness and a higher threshold PP (589, 1436). The preglomerular resistance vessels of normotensive DS rats fed a low-salt diet have an enhanced sensitivity to inhibition of myogenic responses by a CCB, suggesting reduced activity or density of L-type VOCCs (1436). Both NO- and EDHF-mediated components of ACh-induced afferent arteriolar dilation were attenuated in DS rats by enhanced ROS (1145). Reduced renal vascular production of 20-HETE accounted for the impaired myogenic and MD-TGF responses of the afferent arteriole and the weak autoregulation of RBF and PGC (463, 1220). Cerebral arteries from salt-fed hypertensive DS rats normally have a fast myogenic response. This was maintained in one study (56), but attenuated in another report, thereby leading to hypertensive encephalopathy, seizures, and disruption of the blood-brain barrier (600, 1164, 1386). The myogenic response of skeletal muscle arterioles that is normally slow and modest was reduced, but the gracilis arteries of hypertensive DS rats maintained myogenic tone related to 20-HETE production that inhibited KCa channels and reduced EM (446). Knockdown of TGF-1 expression slowed the progression of proteinuria, glomerulosclerosis, and renal interstitial fibrosis in DS rats fed a high-salt diet independently of changes in BP (226). Prolonged blockade of TGF-1 increased the medullary blood flow and reduced BP modestly (293). This suggests that the development of hypertension, glomerular and tubular injury, and vasoconstriction all may depend on overproduction of this growth factor in the renal medulla of DS rats. In summary, renal cortical blood flow autoregulation is well maintained in normotensive DS and DR rats fed a low-salt diet and in early hypertensive DS rats during increased salt intake. Since the impairment of autoregulation follows the development of renal injury, which indeed is apparent soon after birth, it is likely the result, not the cause, of the renal failure. The eventual loss of renal autoregulation is related to impaired myogenic responses in hypertensive DS rats consuming salt since MD-TGF activity is maintained. These rats tend to have impaired cerebral arteriolar myogenic tone, which probably contributes to their development of hypertensive encephalopathy. The finding of preserved myogenic responses in skeletal muscle arterioles suggests that the severely impaired responses in most studies of renal vessels is not due to a generalized failure of myogenic mechanisms and is consistent with a secondary response to renal vascular and parenchymal damage. 4. Goldblatt renovascular hypertension In 1937, Goldblatt (476) reported that hypertension commonly developed after constriction of one main renal artery to reduce RPP (two-kidney, one-clip or 2K,1C model). This models renovascular hypertension from unilateral renal artery stenosis. The GFR was similar in the two kidneys of rats with moderate, early 2K,1C hypertension with no obvious renal pathology (1410). Hypertension and renal vasoconstriction are related to an activated RAS since AT1A receptor deficient mice develop little hypertension after the clipping of one renal artery (206, 208), and blockade of the RAS or AT1 receptors in rats reduces the BP and produces Physiol Rev • VOL 95 • APRIL 2015 • www.prv.org 459 RENAL AUTOREGULATORY MECHANISMS marked vasodilation of the nonclipped kidney (209, 210, 649, 650, 669, 1186, 1566), similar to CCB blockade (1186). In contrast, the clipped kidney sustains a further reduction in its RBF and GFR, suggesting either that its autoregulation is severely impaired or that the RPP beyond the clip is reduced below the autoregulatory range. An ACE inhibitor given to rats with 2K,1C hypertension reduced the BP, RBF, PGC, and afferent and efferent arteriolar resistances in the clipped kidneys (310), whereas a CCB reduced afferent arteriolar resistance selectively (310). Micropuncture studies of superficial nephrons in moderately hypertensive 2K,1C rats reported that the nonclipped kidney has well maintained or increased SNGBF, SNGFR, and PGC, findings consistent with a predominant increase in preglomerular vascular resistance (308, 452). However, in others, the PGC was elevated and the SNGBF and SNGFR were reduced, suggesting a reduced KUF (452, 1410). The SNGBF, SNGFR, and PGC were reduced in the clipped kidney of 2K,1C rats, implying increased preglomerular vascular resistance from the clip and/or the vasculature (1336 – 1338). Constriction of one main renal artery and removal of the other kidney (one-kidney, one-clip or 1K,1C model) also causes hypertension, but this model is less dependent on the RAS and is distinctly salt sensitive. The 1K,1C is a model of renal transplant stenosis or bilateral renal artery stenosis. These clinical conditions resemble the 1K,1C model since all lack a “normal” contralateral kidney to correct renal salt retention by the clipped kidney. The clipped kidney of 1K,1C rats had reduced SNGBF and SNGFR with unchanged PGC due to a preferential increase in efferent arteriolar resistance (1336). Dogs (1047) and some rats (705, 707, 1365, 1566) with 2K,1C hypertension have maintained RBF and GFR autoregulation in the nonclipped kidney, although other studies in rats have reported weak autoregulation (1188, 1404, 1494) that was improved by NOS inhibition (1494). Blockade of AT1 receptors in 2K,1C hypertensive rats did not affect RBF autoregulation in the nonclipped kidney but abolished GFR autoregulation (1566), presumably because of a reduced efferent arteriolar resistance. Autoregulation of renal cortical perfusion of the nonclipped kidney of 2K,1C hypertensive rats was relatively resistant to blockade by a CCB (646), suggesting upregulation of L-type VOCCs. The clipped kidney has a weak, or even nonexistent, autoregulation of RBF (705). The post-clip kidney and its blood vessels were protected from barotrauma, whereas there were extensive glomerular and vascular injuries in the contralateral nonclipped kidney (1365). yet maintained NO vasodilation (684), indicating a selective impairment of autoregulation. The myogenic response of afferent arterioles in HNK nonclipped kidneys of rats with 2K,1C hypertension was blunted and shifted to higher levels of PP (589). Increased NO production by the nonclipped kidney (318, 1363) impaired its RBF autoregulation since this was normalized by NOS inhibition (1494), whereas reduced EET production contributed to renal vasoconstriction. However, restoring EETs did not restore autoregulation (1404), indicating that they were not the cause of defective renal autoregulation. Similarly, 20-HETE participated in basal vasoconstriction, but did not account for the attenuated autoregulation in the unclipped kidney of 2K,1C rats. Thus increased levels of NO could account for some of the changes in autoregulatory efficiency, whereas altered production of EETs and 20-HETE are not implicated. The MD-TGF response in the nonclipped kidney of 2K,1C rats generally was normal or enhanced (134, 648). However, MD-TGF responses in the nonclipped kidneys of 2K,1C rats early after developing hypertension were attenuated despite an activated RAS (1191, 1314, 1317). The MD-TGF response in the nonclipped kidneys was enhanced greatly after NOS inhibition (1491, 1494). MD-TGF in the clipped kidney was very hard to measure, but was normalized shortly after removing the renal artery clip (1191, 1317). MD-TGF was attenuated severely in the solitary kidney of 1K,1C rats (1187). Dynamic analysis of proximal tubular pressure in the nonclipped kidney of 2K,1C rats during the first week of hypertension revealed normal MD-TGF oscillations. As the hypertension progresses, these oscillations transitioned into deterministic chaos, similar to that found in adult SHR (1635). To our knowledge, renal vascular admittance has not been analyzed across the full-frequency spectrum in animal models of renovascular hypertension. In summary, results on the efficacy of steady-state autoregulation of RBF in the nonclipped kidney of 2K,1C hypertensive rats are quite mixed, with roughly equal numbers reporting that it is well-maintained or impaired. The reason for this disparity is not known. Likewise, reports of MDTGF responses in nonclipped kidneys have varied from normal to markedly impaired. The MD-TGF activity appears to be the outcome of an enhancement due to ATI-receptor activation from renin released by the clipped kidney that is offset by increased NO. Autoregulation is weak or absent in the clipped kidneys, perhaps because the RPP is at, or below, the lower limit of autoregulation. 5. ANG II-induced hypertension Medullary blood flow autoregulation in the nonclipped kidney was impaired (646), and its preglomerular juxtamedullary blood vessels failed to vasodilate during reduced RPP 460 Prolonged infusion of ANG II at an initially subpressor rate gradually increases BP, oxidative stress, RVR, inflamma- Physiol Rev • VOL 95 • APRIL 2015 • www.prv.org CARLSTRÖM ET AL. tion, and renal parenchymal injury in association with renal vasoconstriction and an early phase of Na⫹ retention (492). The degree of hypertension and renal injuries are related to the dose of ANG II used, choice of species (rats vs mice), and the duration of treatment. Cross-transplantation studies in AT1 receptor-deficient mice demonstrated that the major prohypertensive actions of ANG II are on renal AT1 receptors (281). Development of hypertension was prevented by an AT1 receptor blocker (1550). ANG II receptors, in particular AT1, in the vasculature are usually downregulated by high levels of ANG II (15, 77, 91, 526, 1266). Renal vascular AT1 receptors expression was reduced in ANG II-induced hypertension (39). Most studies of renal function have been conducted during the established phase of hypertension after 1 or 2 wk of ANG II infusion. At this time point, the Na⫹ excretion, RBF, and GFR were normal or modestly reduced (207, 652, 1088). RVR is consistently increased. The SNGBF, SNGFR, and KUF were reduced and afferent and efferent arteriolar resistances increased (436). PGC was increased during albuminuria (1008). Pressure natriuresis in ANG II-induced hypertension was reset to a higher RPP despite efficient autoregulation of medullary blood flow (1516, 1549). Blockade of AT1 receptors in ANG II-induced hypertensive rats reduced BP and increased RBF and cortical blood flow, GFR, and Na⫹ excretion (1550). Reactivity of afferent arterioles of JMN to ANG II, but not PE, was increased after 1 or 2 wk of ANG II infusion (670). The exaggerated response depended on the presence of renal nerves (660) and increased inactivation of EETs by soluble epoxide hydrolase (1662). Reactivity of afferent arterioles of JMN to ATP by P2X1 receptors was reduced after 1 or 2 wk of ANG II infusion (1661). ANG II-induced hypertension and renal vasoconstriction are dependent on enhanced ROS production by NADPH oxidase (1015, 1128, 1584), impaired renal cortical NOS activity (241, 242, 1025), and reduced renal medullary NO production (1432). Although COX inhibition has little impact (483), hypertension and renal vasoconstriction were less in TP receptor gene-deleted mice (441, 780), implicating involvement of a vasoconstrictor prostanoid, perhaps TxA2, acting on TP receptors. Prolonged infusion of ANG II increased NOS activity in the renal cortex, with enhanced expression of eNOS and nNOS, but no change in NOS activity in the renal medulla (241). Accordingly, NOS inhibition produced greater renal vasoconstriction with greater reductions in RBF and cortical blood flow (242). The authors concluded that a compensatory increase in NO helps to counteract the vasoconstrictor influence of ANG II and thus maintains RBF and cortical perfusion. The normal renal vasoconstriction after inhibition of nNOS was lost in ANG II-infused rats (207). A CCB normalized BP, RVR, GFR, and Na⫹ excretion in rats with ANG IIinduced hypertension (652). Vascular remodeling may contribute to ANG II-induced hypertension (373). ANG II produced less hypertension in adenosine A1 receptor-deficient mice (455). Enhanced afferent arteriolar responses to ANG II in hypertensive wild-type mice were reduced by tempol. Contractions in arterioles lacking the A1 receptor were weaker and not attenuated by tempol, suggesting that A1 signaling may modulate ANG II-mediated contraction or oxidative stress. RBF autoregulation was impaired, whereas medullary autoregulation was preserved in nondiuretic ANG II-infused rats (1549). One study of ANG II-induced hypertension in UNx rats reported normal autoregulatory responses of the afferent arteriole of JMN whether or not the kidney was denervated (660). Other investigators reported that the autoregulatory response of the cortical radial artery and afferent arteriole in the JMN preparation was attenuated 14 days after ANG II-induced hypertension (198, 670, 689). This was related to vascular injury (198, 1029) and activation of ET-1 receptors. Normalization of BP by prolonged AT1 receptor blockade or by triple therapy (hydralazine, hydrochlorothiazide, and reserpine) restored autoregulation of afferent arterioles (689). RBF autoregulation was impaired during ANG II-induced hypertension (AI increased from 0.04 to 0.66), which was normalized by blockade of P2Y12 receptors (1132). Administration of the anti-inflammatory agent pentosan polysulfate for 14 days restored autoregulation of afferent arterioles of JMN in ANG II hypertensive rats via normalization of P2X1 receptor activation by ATP during sustained hypertension (520). Other studies on superficial nephrons reported augmented P2X1-mediated constriction of afferent and efferent arterioles in rats with ANG II-induced hypertension while P2X1 receptor density was normal, perhaps due to inhibition of NO production (436). In contrast, UNx rats infused with ANG II for 2 wk have normal afferent arteriolar myogenic responses of JMN (660), as do cortical afferent arterioles from ANG II-infused mice studied ex vivo (864). Afferent arterioles from ANG II-infused rabbits have enhanced contractions to ANG II despite a twofold downregulation of mRNA for AT1 receptors (1552). This was offset by a fivefold upregulation of mRNAs for p22phox (1552). In agreement, enhanced arteriolar reactivity to ANG II was absent in mice with NOX-2 deficiency (182, 184). Moreover, isolated cortical radial arteries of ANG II-infused mice lacking regulator of G protein signaling-2 had stronger myogenic tone and constrictor responses to ANG II, probably by sensitizing AT1-receptor signaling (604). Physiol Rev • VOL 95 • APRIL 2015 • www.prv.org 461 RENAL AUTOREGULATORY MECHANISMS Admittance transfer function in anesthetized ANG II-infused rats demonstrated impaired myogenic and MD-TGF components of renal autoregulation independent of ET-1 (1272) but magnified by a high-salt diet that increased ROS (1273). However, a careful study of dynamic autoregulation in conscious ANG II-infused rats reported a clearly enhanced myogenic mechanism and reduced transmission of BP fluctuations to the kidney (1193), whereas the MDTGF signature was unaltered. Likewise, long-term ANG II infusion in conscious dogs potentiated the myogenic resonance peak (755). Thus these studies in conscious animals with prolonged ANG II infusion may explain the relatively modest degree of hypertensive glomerular injury since increased myogenic responses were lost under anesthesia. ANG II is known to stimulate oxidative stress and increase NADPH oxidase expression in the kidney (211, 455, 578). Tempol normalized the vascular O2⫺ production and reduced BP without affecting the RBF of ANG II-infused rats (1088). Oxidative stress is thought to play an important role in the regulation of medullary blood flow and pressure natriuresis (273, 964, 1099, 1670). ANG II can activate T cells that release cytokines to promote oxidative stress, inflammation, renal vasoconstriction, and Na⫹ retention, all of which are prohypertensive (534, 579). As observed with an AT1 receptor blocker, administration of the anti-inflammatory agent mycophenolate mofetil during ANG II infusion reduced BP, renal vasoconstriction, and renal interstitial inflammation (439). Although the MD-TGF was augmented by short-term ANG II infusions (1012, 1077) and usually attenuated by pharmacological or genetic inhibition of the RAS and AT1 receptors (see sect. VIA), prolonged slow-pressor infusion of ANG II did not change MD-TGF at normal tubular fluid flow (1095). During chronic ANG II infusion, there was a complex regulation of MD-TGF by a vasoconstrictor prostanoid that activated TP receptors and by ROS that impaired NO signaling in the JGA and by reduced expression of AT1 receptors (39). COX-1 products enhanced MD-TGF by activation of TP receptors that elevated ROS in the kidney (137, 1082, 1083, 1581, 1593), whereas COX-2 products attenuated MD-TGF in ANG II-infused mice (39). ANG II-induced hypertension depended in part on activation of TP receptors (780). Increased MD-TGF during prolonged ANG II infusion was related to a reduced blunting by MD-derived NO because of enhanced O2⫺ (133, 184, 659, 1608). Small interference RNA knockdown of p22phox reduced ROS and MD-TGF activity by 50% in ANG IIinfused rats, thereby linking ROS to enhanced MD-TGF (1095). P2X1 receptor signaling in the kidney (436) and preglomerular vessels of JMN was impaired by inflammation caused by ANG II infusion (520, 686, 1661). Prevention of inflammation improved autoregulation, apparently by normalizing MD-TGF activity (686, 1661). 462 In summary, steady-state RBF autoregulation is attenuated in most studies of ANG II-induced hypertension, although medullary blood flow and its autoregulation are generally well maintained. Myogenic tone is enhanced in intact kidneys of conscious animals, but not in isolated arterioles, suggesting that a reduced MD-derived NO may enhance responses in intact kidneys. MD-TGF is normal or exaggerated, likely the outcome of enhanced O2⫺ generated in the JGA in response to vasoconstrictor COX-1 metabolites that activate TP receptors and decrease NO activity. However, MD-TGF is attenuated in JMN, perhaps related to inflammation impairing P2X1 receptor signaling. The reasons for the differential effects of ANG II on MD-TGF responses of superficial and deep nephrons and the variable effects of ROS to dampen or enhance myogenic responses are not yet resolved. Thus there are major hemodynamic differences in the response to chronic versus acute activation of the RAS and perhaps the balance of NO and oxidative stress. Longterm ANG II stimulates ROS production that impairs the bioavailability of NO and enhances production of a vasoconstrictor prostanoid that activates TP receptors. These are offset by adaptive reductions of AT receptor expression and structural changes of the vascular wall. 6. DOCA-salt hypertension Prolonged administration of the mineralocorticosteroid deoxycorticosterone acetate (DOCA) and a high-salt diet (e.g., 1% NaCl) have usually been combined with UNx to model volume-expanded, low plasma renin activity (PRA) hypertension similar to human primary hyperaldosteronism. This model is dependent on increased ET-1 production in blood vessels and kidneys (1291). DOCA-salt mice have a rightward shift in the pressure-natriuresis relation (512). UNx rats with DOCA-salt hypertension have increased PGC and develop proteinuria and focal glomerulosclerosis (361, 363). Nonspecific antihypertensive therapy (hydrochlorothiazide, hydralazine, and reserpine) reduced BP in DOCA-salt rats but failed to prevent elevated PGC, proteinuria, or glomerular injury (361). Combined blockade of Tand L-type VOCCs decreased BP and PGC and reduced proteinuria and glomerular lesions without affecting the RAS (75, 767), whereas selective antagonists of L-type VOCCs also decreased the BP but stimulated the RAS and failed to prevent proteinuria or glomerular lesions (75, 365, 767). This finding in hypertensive animals is perplexing in view of the reported absence of T-type channels in glomerular arterioles of normotensive rats (1388). However, another study reported that an L-type CCB was more effective than ACE inhibition in reducing the BP, proteinuria, and glomerular pathology, although both treatments reduced the PGC (364). A sharp fall in BP sufficient to reduce PGC may account for the protective effects of the CCB in this study. Dietary Ca2⫹ supplementation reduced PGC, proteinuria, and renal injury without changing the BP (359). Thus an elevated PGC and the RAS are more important than hyper- Physiol Rev • VOL 95 • APRIL 2015 • www.prv.org CARLSTRÖM ET AL. tension in predicting proteinuria and renal damage in the DOCA-salt model. Basal RBF and GFR are normal or reduced in DOCA-salt rats and dogs (45, 540, 540, 674, 969, 1047, 1047, 1253). There was a maintained plateau phase of RBF autoregulation, with either a normal (45) or a 20 mmHg increase in its lower limit (674). RBF autoregulation in dogs with DOCAsalt hypertension was well preserved and independent of renal sympathetic nerves or PGs (1420), but GFR was poorly autoregulated (550). Neither RBF nor GFR was autoregulated in the isolated perfused kidney of DOCA-salt dogs (764), but autoregulation generally is less efficient in this preparation. The RBF was less efficiently autoregulated in DOCA-salt hypertensive Yucatan miniature swine, but again there was even less complete autoregulation of GFR (1647). RBF and renal cortical blood flow were well autoregulated in DOCA-salt mice, and surprisingly, medullary blood flow was more efficiently autoregulated in DOCAsalt than in control mice (512). Some studies of DOCA-salt rats reported attenuated MDTGF (1040, 1315) and autoregulation of SNGFR (1022), whereas others reported that the autoregulation of GFR and SNGFR were well maintained despite inhibition the MD-TGF (540). The findings in several studies of DOCAsalt hypertensive rats, dogs, or swine that the GFR was less well autoregulated than the RBF suggest a reduced efferent arteriolar resistance due to the strongly suppressed RAS. Vascular O2⫺ production was increased in DOCA-salt hypertensive rats (1391). Antioxidants (100, 968) or inhibition of NADPH oxidase (641, 735) reduced BP, vascular O2⫺, and renal damage. ET-1 production was increased in the vasculature of DOCA-salt hypertensive rats (227, 1292), and selective blockade of ETA receptors reduced BP, vascular O2⫺ generation, and RVR, but did not always improve renal damage (14, 175, 938, 1264). ETB receptors may provide a protective function. Rats lacking ETB receptors have exaggerated vascular and renal pathology (986) accompanied by reduced NO production (620) that may underlie the enhanced vasoconstriction of their afferent arterioles (1470). However, others have ascribed the renal vasoconstriction and hypertension to enhanced activity of the renal sympathetic nerves (719, 776), since renal denervation delayed the development of hypertension and increased Na⫹ excretion (775) or to inflammation since immunosuppression preserved GFR and reduced proteinuria in DOCA-salt hypertension (124). Another study implicated purinergic signaling since P2X7 receptor-deficient DOCA-salt mice were less hypertensive and infiltration of immune cells and renal injuries were blunted (732). In summary, DOCA-salt is a model of low-renin, volumeexpanded hypertension that resembles human primary hyperaldosteronism. Factors contributing to hypertension, re- nal vasoconstriction, and renal damage include oxidative stress, NO deficiency, inflammation, enhanced P2X7 and ETA but reduced ETB receptor signaling, and enhanced renal nerve activity. Despite the renal vasoconstriction and abnormal receptor signaling, RBF autoregulation is well maintained in most studies, albeit GFR autoregulation is less efficient, likely reflecting a diminished efferent arteriolar resistance in this low renin-angiotensin model. The preservation of RBF autoregulation despite a markedly attenuated MD-TGF response suggests an intact or enhanced myogenic mechanism, but this has not been studied specifically. 7. FH hypertensive rat The FH rat is a genetic, renin-dependent, model of systemic and glomerular hypertension that develops progressive albuminuria and spontaneous focal and segmental glomerulosclerosis (FSGS) that shortens its lifespan (841). It resembles progressive loss of function in humans with CKD. Mild hypertension and increased PGC developed by 8 wk (843, 1371), proteinuria and enhanced myogenic contraction of small mesenteric artery by 20 wk (1527), FSGS and proteinuria by 2– 6 mo, and severe hypertension and CKD with malignant nephrosclerosis by 1 yr (294, 840, 842, 843, 1202, 1527, 1579). The FH rats are reported to die of renal failure at sea level, whereas at altitude (e.g., Denver-raised FH rats) they die of pulmonary hypertension, which has been characterized by a deficiency of pulmonary eNOS and abnormal vascular growth (878). Studies in FH rats, before and after appearance of renal injury, demonstrated that attenuated renal myogenic responses in 7-wk-old FH rats preceded glomerulosclerosis and proteinuria (1102). Increased renal cortical and medullary blood flow and PGC were related to a low ratio of afferent to efferent arteriolar resistance. Autoregulation of RBF, cortical perfusion, and PGC were impaired markedly in adult FH rats (1522, 1534). Prolonged ACE inhibition largely prevented the systemic and glomerular hypertension and renal damage (1534, 1535). Isolated, perfused cortical radial arteries from young FH rats constricted weakly or even dilated with increased PP (1102, 1522, 1527). This severe defect, or even absence, of myogenic contraction was considered renal specific since those contracted normally to PE (1534) and the myogenic responses of isolated mesenteric arteries and the aorta were preserved (1102, 1527). This suggested a profound defect in renal arteriolar mechanosensitivity or mechanotransduction. On the other hand, cortical radial arteries and afferent arterioles in the HNK preparation of FH rats had nonselective impairment in reactivity to PP and to ANG II (1523). Micropuncture studies of FH rats with mild FSGS reported normal MD-TGF activity that increased with age and was independent of ANG II (1534). An increased expression of Physiol Rev • VOL 95 • APRIL 2015 • www.prv.org 463 RENAL AUTOREGULATORY MECHANISMS neuronal NOS and COX-2 in the MD and of renin in the afferent arterioles of FH rats preceded the development of glomerulosclerosis (1579). Thus preglomerular vasodilation by NO, perhaps driven by a COX-2 metabolite, may combine with hypertension caused by high ANG II to impair renal autoregulation and increase PGC that culminates in barotrauma manifest as FSGS. Linkage analysis showed that genetic loci on chromosome 1 of FH rats cause the hypertension, impaired autoregulation, and proteinuria (1518 –1521). Transfer of this region from BN into FH rats improved RBF autoregulation, normalized the elevated PGC, and reduced protein excretion and the progression of renal disease (922, 1610), whereas transfer of this chromosomal region from FH rats into normotensive controls attenuated renal autoregulation and led to FSGS (1518). This provides strong evidence that the selective impairment of the renal myogenic response and the hypertension in this model are genetically determined and are causally related to glomerulosclerosis. The particular genes involved have yet to be identified. Thus the selective impairment of renal myogenic responsiveness in FH rats reported in most studies despite preserved MD-TGF, PE-induced vasoconstriction, and myogenic responses in other vascular beds, suggests that the defect in renal autoregulation is the outcome of genetically determined changes in the preglomerular vasculature. The weak myogenic response likely accounts for the attenuated renal autoregulation that, in the context of hypertension, leads to barotrauma and CKD. This conclusion is supported by recent studies of the FH hypertensive, consomic and congenic rat strains, which demonstrate the importance of autoregulation in preserving glomerular structure and function. 8. BN rat The BN rat is of special interest because it develops renal failure despite a low BP, likely related to weak and inconsistent renal autoregulation because of excessive renal NO generation that can prevent full vasoconstrictor responses (260, 916). Although the BN kidney is susceptible to hypertension-induced kidney disease (260), the BN lives a long life, perhaps due to its low BP (916). Cross-transplantation studies demonstrated that the kidney of the normotensive BN rat was inherently more susceptible to hypertensioninduced damage than the SHR (260, 916), thereby locating the deficit within the kidney itself. Surprisingly, the kidneys of BN rats have efficient steady-state RBF autoregulation with a basal AI of 0.19 (922), but this is unusually fragile since they are susceptible to injury with even moderate hypertension or enhanced oscillations in RPP. Dynamic admittance transfer function analysis identified an apparently normal myogenic response with spontaneous changes in RPP, but a weaker myogenic response to larger, forced changes in RPP (1563). Thus incomplete autoregulation 464 could underlie the susceptibility to hypertensive renal injury. Cupples and co-workers (1569, 1570) reported that the attenuated myogenic autoregulation was improved by nonselective NOS inhibition or by inhibition of iNOS but not of nNOS, implicating high NO generation from iNOS as the cause of the blunted myogenic response. In contrast, the renal autoregulation of normal Wistar rats was unaffected by inhibition of iNOS, but the strength of the myogenic response is enhanced by inhibition of nNOS (1358). Since these BN rats also have exaggerated depressor response to ACh, which was normalized by global NOS inhibition, they also may have an enhanced eNOS-derived NO (1570). Wang and Cupples (1570) concluded that the normal buffering of the myogenic response by MD nNOSderived NO was dominated in BN rats by an inappropriate release of NO by iNOS, and perhaps by eNOS, that attenuated the renal myogenic response. The roles of NO in renal autoregulatory mechanisms were discussed in detail earlier in section VIC. BN rats have a preserved myogenic response of cremaster arteries and gracilis muscle resistance arteries that depended on 20-HETE production (445). Impaired myogenic tone has been reported of cerebral arterioles (322) and may contribute to their susceptibility to cerebral hemorrhage and stroke. Recent studies show that a 2.4-Mbp region of chromosome 1 was critical for normal RBF autoregulation since its substitution from BN into FH rats improved their RBF autoregulatory efficiency and afferent arteriolar myogenic responses, reduced BK channel activity (168), and improved the myogenic tone of cerebral arteries (1147). In summary, BN rats have labile renal autoregulation that breaks down during hypertension. The weak link is identified as an impaired dynamic myogenic response related to dysregulated overproduction of NO, but the molecular mechanisms have not yet been defined. The defective myogenic response is considered genetically determined by sites on chromosome 1 that may impair both renal and cerebral vasoconstriction during increased PP. C. CKD and RRM A fundamental problem with human CKD is its propensity to undergo a detrimental switch from beneficial adaptations that restore GFR to pathological proteinuria and declining renal function (637). This sequence is modeled by a 50 – 85% RRM that is characterized in rodents by initial increases in glomerular size, PGC, and SNGFR and hypertrophy in remnant nephrons and followed by proteinuria and progressive tubulointerstitial fibrosis and glomerulosclerosis usually accompanied by hypertension (29, 144, 147). The rate and the magnitude of these changes depends on the species, age, amount of renal mass removed, and the tech- Physiol Rev • VOL 95 • APRIL 2015 • www.prv.org CARLSTRÖM ET AL. nique (surgical ablation or infarction). The infarction method causes ischemic, renin-dependent hypertension and a more rapid decline of GFR, but is a less representative model for human CKD where renal tissue is not clearly ischemic and PRA is generally suppressed. These defects were accelerated by high dietary intakes of protein or salt (147, 803). Bidani and colleagues related the renal damage to hypertension coupled with impaired autoregulation that allowed transmission of enhanced BP fluctuations to the glomeruli to initiate barotrauma (103, 104, 108, 112, 496, 504). Nephron adaptations have been studied extensively after either UNx or 5/6 Nx and generally produce similar results. The SNGFR of Munich-Wistar rats doubles by day two of RRM and triples by 2 wk (319, 636, 637, 1165), restoring whole kidney GFR and RBF almost to normal (497). The early renal vasodilatation is attributed to enhanced PG production (497, 1166, 1167). Proteinuria and mesangial expansion are apparent by 2 wk and are accompanied by an increased glomerular volume and capillary diameter (518, 637, 1119). There is a ⬃10 mmHg increase in PGC as a result of a preferential decrease in afferent arteriolar resistance (357, 876, 1166). Rats with RRM had persistent systemic hypertension, progressive proteinuria, and glomerular mesangial expansion and segmental sclerosis by 8 wk (31). The decline in renal function was considered to be secondary to an initial increase in PGC, SNGFR, and SNGBF (29, 144, 147). Inhibition of the RAS lowered BP and PGC and lessened the proteinuria and glomerulosclerosis in the RRM infarction rat model. This was accompanied by a normalization of SNGBF and SNGFR by a relatively selective reduction in the efferent arteriolar resistance (32, 1004). Impaired autoregulatory adjustments of the afferent arteriole in rats with RRM likely contributed to the reduced afferent arteriolar resistance despite hypertension, the increased PGC, and the glomerulosclerosis (160, 876, 1167). The renal adaptations and impaired autoregulation are dependent on the strain and the age of the rat. Perinatal UNx in normal rats resulted in salt-sensitive hypertension, exaggerated MD-TGF, proteinuria, increased glomerular volume, and renal injury by 3 mo (186, 1611) which were ascribed to increased ROS and NO deficiency (181, 185). UNx in neonatal SD rats led to renal vasodilation and complete loss of steady-state RBF autoregulation at 5– 6 wk attributed to overproduction of PGs (235–237). However, UNx in adult SD rats increased basal RBF but with little impact on whole kidney RBF autoregulation (237). Weak autoregulation in UNx FH rats accelerated the process of progressive renal failure with increased PGC and SNGFR evident by 12 wk, followed by severe proteinuria and a fall in GFR (1371). SHR kidneys have highly efficient steady-state RBF autoregulation with normal PGC, as discussed in section VIIB1 (42, 58, 358, 394, 707, 1471) and minimal glomerular damage (358, 394, 707, 1471). After UNx, the remnant kidney of young SHR has elevated PGC and they develop proteinuria (93, 358, 360). However, adult SHR retained efficient overall steady-state RBF autoregulation after UNx (709). The renal adaptations were more severe in 5/6 Nx rats. Within 2 wk of 5/6 Nx produced by surgical ablation in adult SHR, hypertension worsened and RBF autoregulation was abolished with evident injury to preglomerular arteries, arterioles, and glomeruli (107). Normal rats subjected to 5/6 Nx also developed hypertension and, by 3 wk, had impaired steady-state RBF autoregulation (503). Their juxtamedullary afferent arterioles became hypertrophic and their arteriolar autoregulatory response was impaired (903). Hypertension and elevated PGC are considered key elements in renal damage in UNx SHR and 5/6 Nx Munich-Wistar rats since injury was mitigated by antihypertensive treatment with hydralazine, hydrochlorothiazide, and reserpine (501); salt restriction (357, 876); or with ACE inhibition or even with a CCB in some studies (360, 362). However, impaired renal autoregulation also was a critical element in renal damage since a low-protein diet improved steadystate RBF autoregulation and prevented proteinuria or renal damage without prominent effects on BP in 5/6 Nx rats (112, 502). Moreover, loss of autoregulation by blockade of VOCCs by CCBs in some other studies exacerbated renal damage in rats with RRM despite lowering their BP (499, 500, 502, 503). The best method to evaluate autoregulation and its mechanisms in RRM is not clear. Frequency analysis of dynamic RBF autoregulation suggests preserved autoregulation in rats with RRM. However, this contrasts with defective RBF autoregulation observed in steady-state studies (109). Bidani et al. (109) concluded that the staircase steady-state method provides a better measure of susceptibility to hypertensive glomerular damage, but the reason for the preserved dynamic RBF autoregulation is unresolved. Most studies have concluded that renal damage requires, or is accelerated by, a high BP. Thus rodent strains that do not develop hypertension after RRM, such as WKY rats (110) or C57Bl/6 mice (861), are relatively resistant to renal damage after RRM despite impaired autoregulation and myogenic responses. However, after the induction of hypertension, WKY rats with RRM indeed sustain renal damage (121, 609, 1282, 1451, 1452) as did C57Bl/6 mice infused with ANG II or treated with DOCA and salt (111, 180, 1237). Some studies have dissociated worsening autoregulation from progression of CKD. Thus the myogenic response of renal afferent arterioles isolated from C57Bl/6 mice was attenuated progressively over 3 mo after RRM (861). While Physiol Rev • VOL 95 • APRIL 2015 • www.prv.org 465 RENAL AUTOREGULATORY MECHANISMS a high-salt diet enhanced the defects in myogenic responses, there was no glomerulosclerosis and only mild tubulointerstitial fibrosis (861), perhaps reflecting the absence of hypertension even with a high-salt diet (861). Rho-kinase inhibition in SHR after subtotal Nx attenuated renal autoregulation but paradoxically reduced the renal damage (594). Blockade of PG production in rats with established RRM increased the afferent and efferent arteriolar resistances, but did not improve renal autoregulatory efficiency or prevent renal damage (497, 1063). ACE inhibition after RRM failed to fully restore RBF autoregulation but was protective of kidney function (503). Anti-inflammatory agents given to rats with RRM prevented the rises in SNGFR and PGC and the development of proteinuria and decreased the mesangial proliferation but increased the hypertrophy of afferent arterioles (121, 609, 1451, 1452). Blockade of ROS generation by tempol or by inhibition of xanthine oxidase protected kidneys of rats (1282) or mice (861) with RRM without affecting BP. Nevertheless, the weight of the data generally supports the proposal of Bidani et al. that both hypertension and impaired renal autoregulation are required after RRM for kidneys to develop glomerulosclerosis. However, changes in Rho/Rho kinase, PGs, ANG II, and ROS can impact RRM progression apparently independent of effects on autoregulation or BP. Anti-inflammatory or antioxidant drugs might maintain renal microvascular function after RRM (903, 1112), but this requires further study. Impaired myogenic constriction of mesenteric arteries of hypertensive 5/6 Nx rats was restored by long-term inhibition of ACE or antagonism of AT1 receptors that reduced BP and renal injury (1528, 1537). Tempol plus catalase improved myogenic constrictor response of mesenteric arteries of rats with RRM but not those with prolonged AT1 receptor blockade (1528), suggesting that ANG II-induced ROS can impair autoregulation of nonrenal arteries. This could provide a link between damaged kidneys that release renin and impaired systemic microvascular function via ROS generation. The response to RRM has been studied most extensively in rodents, and similar patterns are not always reported for other species. Cats with RRM developed increased SNGFR (158) and PGC (158, 159), hypertension, and proteinuria (9, 157). Their remnant nephrons exhibited glomerular hypertrophy and impaired renal autoregulation (160). Unlike rodents, the feline renal parenchyma was quite resistant to structural damage (1598), and glomerular hyperfiltration was not exacerbated by increasing the protein intake (159, 1598) even when studied over several years (1110 –1112). Thus the cat dissociates hemodynamics, and even hypertension, from structural adaptations and the development of CKD. Functional adaptations to RRM in dogs are less well characterized. The progression of CKD was accompanied by glomerular hypertension and glomerular hypertrophy com- 466 bined with proteinuria, tubulointerstitial inflammation, and oxidative stress (17, 965, 1326). Initially, 3/4 and 7/8 RRM led to increased SNGBF, SNGFR, and PGC (158), but RBF autoregulation was impaired severely only in dogs with 7/8 Nx (160). ANG II mediated the progression of CKD in dogs with RRM since prolonged ACE inhibition reduced PGC, proteinuria, and glomerular and tubulointerstitial lesions (1326), although it also reduced BP which may have made a critical impact. Protein loading increased GFR in normal dogs (420) and in 7/8 RRM dogs with increased renal growth but failed to worsen the severity of renal lesions (1598) as was established in rats with RRM (87, 389). However, a low-protein diet in dogs with 11/12 RRM reduced the severity of proteinuria, glomerulosclerosis, and renal interstitial lesions (1196). Compared with rodents, a higher degree and longer dosing of RRM are required for dogs to develop renal damage. Thus rats with even modest degrees of RRM have impaired renal autoregulation, with glomerular and systemic hypertension, and develop renal injury quite rapidly. Some strains of mice (notably the C57BL/6), like cats and dogs, are quite resistant to the detrimental effects of RRM. This raises concern about whether the rat is a valid model for progressive CKD in humans. The hypothesis derived from the rodent studies that a very low protein intake would slow the loss of GFR in patients with moderate or advanced CKD was disproved by the results of a controlled clinical trial which also reported that a low BP goal did not slow loss of GFR unless there was ⬎3 g daily of proteinuria (803). This was confirmed in a trial of BP reduction in African-Americans with hypertensive nephrosclerosis where an ACE inhibitor was not clearly superior to a -adrenoceptor blocker in preventing loss of GFR, unlike the reports in rats (147), although a CCB did accelerate GFR loss, as in most animal models (13). Again, these results of human studies challenge the relevance of the rodent models, at least for a complete understanding of human CKD progression. The rapid and substantial increases in SNGFR that occur in rats within 1 day after RRM indicate that MD-TGF is less active, or reset, to accommodate the likely considerable increase in tubular fluid delivery to the MD. Indeed, MDTGF responses were effectively abolished within 7 days of RRM in most, but not all, rat nephrons (1372, 1373). The immediate response of MD-TGF is less clear. One view is that MD-TGF is reset within 2 h of UNx to higher levels of tubular flow (1041) by vasodilator PGs (546). Another thought is that MD-TGF is essentially normal shortly after UNx despite the increase in SNGFR (117, 1372). Efficient GFR autoregulation persisted in volume-expanded UNx rats in the absence of a demonstrable MD-TGF, implicating a strong myogenic response accounting for complete autoregulation (945). One week of administration of an AT1 receptor blocker attenuated the MD-TGF activity in shamoperated rats, but paradoxically normalized the previously nonexistent MD-TGF response in 5/6 Nx rats (1372). Thus Physiol Rev • VOL 95 • APRIL 2015 • www.prv.org CARLSTRÖM ET AL. there is a complex interplay of time, volume status, and ANG II signaling after RRM that can impact MD-TGF in a rather unpredictable manner. Hypertensive rats with 5/6 Nx lost myogenic responses of their mesenteric arteries, but this was preserved after prolonged inhibition of the RAS or after tempol plus catalase (1528, 1537). Thus ANG II and ROS can impair myogenic tone in nonrenal vessels from models of CKD in contrast to the opposite effects in normal renal afferent arterioles (658, 660). However, cremaster and femoral arteries from rats with RRM displayed enhanced myogenic contractions before the development of hypertension that were normalized by ACE inhibition (1288, 1289). Human subcutaneous arteries from patients with end-stage renal disease (ESRD) had an unchanged myogenic tone and lacked vascular remodeling (937). These different patterns of myogenic response in vessels from models of CKD remain unexplained. In summary, rodent models of UNx or RRM generally display glomerular hypertrophy, increased SNGFR, and PGC. A combination of systemic hypertension and attenuated renal autoregulation generally are followed by proteinuria and glomerulosclerosis. Initially, a vasodilator COX metabolite and NO participate in afferent arteriolar vasodilation and impair renal autoregulation while efferent arteriolar tone is maintained by ANG II, thereby contributing to the increased PGC and SNGFR. Oxidative stress and inflammation impair renal autoregulation in chronic RRM models. The myogenic response is generally attenuated, but there are complex, time-dependent effects on MD-TGF. However, very different patterns of response and renal damage have been reported between studies and between different nonrenal vessels. These may depend on the degree, method of RRM, the age of the animal, and especially its species, which cautions against drawing uncompromising conclusions concerning autoregulation in human CKD. D. Diabetes Mellitus Diabetic nephropathy entails a combination of albuminuria, proliferative glomerulonephritis, and glomerular and tubulointerstitial fibrosis and is the leading cause of endstage renal disease (ESRD) (508, 1517). Patients with type 1 diabetes mellitus (DMT1, insulin deficient) and type 2 diabetes mellitus (DMT2, insulin resistant, usually complicating obesity or metabolic syndrome) begin to develop proteinuria and a decline in GFR after an interval of ⬃12–15 yr (280, 942). Insulin lack or resistance and high blood glucose and hypertension are commonly associated with oxidative stress and can interact synergistically in the pathogenesis of diabetic nephropathy. Diabetic renal disease is commonly initiated by an increase in GFR (termed hyperfiltration) and microalbuminuria (i.e., daily albumin excretion of 30 –300 mg), especially in those with DMT1 (149) and hypertension (29, 30). Usually, the increase in GFR in DMT1 exceeds that of RBF. Studies in rat and some humans reported that inhibition of the RAS reduces oxidative stress and slows, but does not halt, the development of albuminuria and glomerular sclerosis (28, 1121, 1619). However, other studies have failed to disclose a renal benefit from inhibiting the RAS or lowering the BP in preventing DM (17, 113, 966, 967, 1326). However, ACE inhibitors or AT1 receptor blockers reduce proteinuria and slow the rate of loss of GFR after the development of overt nephropathy with proteinuria ⬎3 g daily (146, 891, 1157). Dietary Na⫹ restriction corrected increased SNGFR in a rat study (66), but exacerbated abnormalities in humans with early DM (1007). Thus fresh insight is required to develop novel strategies to slow the progression of DM to ESRD. 1. DMT1 Streptozotocin (STZ) damages pancreatic beta cells and thereby provides a model of DMT1 with insulinopenia and hyperglycemia. As demonstrated and discussed in several studies, there is renal vasodilation, increased PGC, and SNGFR and glomerular enlargement (145, 189, 638, 1321, 1648). Control of hyperglycemia with insulin normalized PGC, whereas SNGFR remained elevated (1321). RBF increased by 20 –50% by 2 wk after STZ if hyperglycemia was restrained by insulin treatment, and this renal vasodilation depended on increased NO (88) but was diminished by severe hyperglycemia (638). Impaired function of VOCCs may explain the relatively selective afferent arteriolar vasodilatation and increase in PGC since the efferent arteriole lack functional VOCCs (1487). Afferent arterioles from STZ rats were remodeled independent of BP (1497). Indeed, most models are normotensive. Studies of renal autoregulation in rat models of DMT1 reported quite variable results. The plateau phase of steadystate RBF autoregulation was maintained in one study (993), but the limit of autoregulation was shifted to a lower RPP in many studies (307, 612, 874, 993, 1365, 1472), independent of PGs (1472). However, other studies reported that RBF autoregulation was impaired within hours of induction of hyperglycemia by STZ and persisted into the established phase of DM with attenuated myogenic (588) and MD-TGF responses (1509). Hyperglycemia itself is considered important since it can attenuate steady-state autoregulation of RBF and GFR in normal dogs (1624). Rat and rabbit models of DMT1 were accompanied by severe endothelial dysfunction of isolated renal afferent arterioles and impaired NO-mediated vasodilation despite an increased basal RBF (821, 1303, 1575). The reduced NO availability may result from increased production of ROS (821, 1121) by renal NADPH oxidase (55). However, these studies on isolated vessels are at variance with reports of increased GFR, persistent vasodilation (1472), and increased glomerular blood flow and PGC in intact kidneys, all of which are attributed to excess NO (309). Thus the NO in Physiol Rev • VOL 95 • APRIL 2015 • www.prv.org 467 RENAL AUTOREGULATORY MECHANISMS question may derive elsewhere from the afferent arteriole, likely the MD cells. Increased proximal tubular Na⫹ reabsorption in rats with early DMT1 via Na⫹-glucose cotransport reduces Na⫹ delivery to the MD and thus reduces the signal for MD-TGF. An inactivated MD-TGF may contribute to an increased SNGFR in STZ rats (1509), perhaps by exaggerated MD generation of NO (1461, 1462, 1509). Thus hyperglycemia in normal dogs increases their RBF and GFR when MDTGF is active but fails to do so in a nonfiltering kidney (1624). An increased MD-TGF can occur despite increased proximal Na⫹ reabsorption and reduced Na⫹ delivery to the MD (1194). However, other factors must participate in the increased SNGFR since adenosine A1 receptor-deficient mice that lack functional MD-TGF (155, 896, 1426) retained a modest (1514), full (1276), or exaggerated increase in GFR (392) in models with DMT1. Some studies of frequency analysis of the RBF dynamics revealed strong contributions to autoregulation by both the myogenic and the MD-TGF mechanisms that were not impaired by diabetes (89, 874). However, another study reported an attenuated RBF autoregulation with weak contributions of myogenic and MD-TGF responses largely due to excess NO (88). Diminished afferent arteriolar vasodilatation in diabetic models can occur despite maintained NO production (1023) probably because of quenching of NO by enhanced O2⫺ generation (1113, 1303, 1325). The afferent arteriolar autoregulatory response in the JMN preparation was normal in rats with STZ-induced DMT1 (993), but attenuated in the HNK preparation where it was restored by blockade of COX or control of hyperglycemia by insulin (588). However, insulin has no effect on myogenic constriction of normal afferent arterioles despite increasing NO and reducing vasoconstrictor responses to ANG II and NE (590). Moreover, acute hyperglycemia in normal rats dilated posterior cerebral arteries and diminished myogenic tone by an endothelium-dependent mechanism involving NO and a COX-derived PG (263), suggesting different regulation of renal and systemic microvessels in DMT1. Cremaster arterioles from STZ-induced diabetic rats initially autoregulated poorly, but eventually reverted to a normal response (616). Thus the variability of autoregulatory responses may be a function of the degree of hyperglycemia and its effects to increase both NO and ROS, the experimental preparation and the duration of the disease process. Defective L-type VOCC function could contribute to the weakened myogenic and MD-TGF mechanisms reported in STZ rats. Afferent arterioles of JMN had a reduced contraction to high KCl-induced membrane depolarization or to the L-type VOCC agonist BAY K 8644 that was normalized by reducing the glucose concentration (195). Impaired 468 VOCC function appeared to result from hyperpolarization of the VSMC plasma membrane due to increased activity of KATP and KIR channels during oxidative stress and hyperglycemia (665, 1484, 1485). Galactose feeding to inhibit aldose reductase and polyol metabolism models DMT1 with a normal insulin level and a normal afferent arteriolar myogenic response. Thus hyperglycemia can inhibit myogenic tone independent from insulin lack (426, 427). Some studies have demonstrated enhanced myogenic responses in nonrenal vessels from diabetic animals. Whereas myogenic regulation of cerebral cortical blood flow in STZinduced diabetic rats was normal in one study (1267), another reported enhanced cerebral myogenic constrictor tone with reduced NO and activation of KATP channels (1664). Femoral and skeletal muscle arterioles of rats with STZ-induced diabetes had enhanced PKC activity that phosphorylated p47phox and augmented NADPH oxidase production of O2⫺ (1501, 1504), which increased speed and strength by upregulation of VOCC activity (1504). However, one should be cautious in extrapolating these findings to the kidney since there are clearly differences in the myogenic response and its mechanisms between vascular beds. Moreover, careful assessment of BP shows it to fall with the onset of DMT1 or DMT2 and to remain reduced for some time in most studies. In summary, the kidneys of rats with early DMT1 generally are vasodilated with increased RBF and GFR attributed to excess NO, which is impaired by diminishing hyperglycemia with insulin. In contrast, later stages of DMT1 are associated with vasoconstriction, exaggerated ROS, and reduced NO. The available evidence does not allow a definitive conclusion as to whether RBF autoregulation and the myogenic and MD-TGF mechanisms are normal or attenuated. Variability in results likely reflects differences in the disease models or its progression. Attenuated myogenic or MD-TGF components could contribute to afferent arteriolar dilation, glomerular hypertension, and increased SNGFR which some studies have ascribed to attenuated activity of VOCCs. Nevertheless, several studies of nonrenal vessels have demonstrated an enhanced myogenic response. Hyperglycemia and lack of insulin might exert differential effects on the renal and systemic arterioles. Thus impaired autoregulation and vasodilation could result from enhanced proximal tubule fluid reabsorption secondary to activated Na⫹-glucose cotransport leading to reduced NaCl delivery to the MD and attenuated MD-TGF. However, increased SNGFR can occur in models lacking a MD-TGF response. Regardless of the mechanism, a combination of a prolonged increase in SNGFR and PGC is ominous since it predisposes to barotrauma and nephropathy (948). Some of the variability in reports between STZ studies likely relates to insulin replacement and prevention of severe hyperglyce- Physiol Rev • VOL 95 • APRIL 2015 • www.prv.org CARLSTRÖM ET AL. mia. Most studies before 2000 did not use insulin such that animals lost weight and often suffered from prolonged glucose osmotic diuresis. 2. DMT2 Albuminuria is a marker for risk of CKD in patients with DMT2 (316). The RAS may be a major mediator of glomerular adaptation, renal inflammation, and renal injury during established diabetic nephropathy with heavy proteinuria since inhibition of the RAS at this stage reduces albuminuria and affords some renal protection (316). There are several rodent models of DMT2, as discussed below. The obese Zucker diabetic fat rat (ZDF) fed a modestly enriched fat diet develops insulin resistance and hyperglycemia followed by proteinuria and glomerular injury (272, 772, 773) related to increased SNGFR either without (1101) or with increased PGC (1299). Renal ANG II was elevated in ZDF rats (1440), and inhibition of RAS reduced systemic and glomerular hypertension, albuminuria, and renal damage (73, 73, 1100, 1299, 1299). ZDF rats have increased GFR but, impaired GFR and RBF autoregulation despite reduced NO production in the renal cortex and medulla attributed to increased O2⫺ (450, 1440). They are modestly hypertensive with a resetting of the pressure-natriuresis relation to favor Na⫹ reabsorption at a lower RPP. Hyperglycemia and oxidative stress can activate PKC and MAPK signaling (271, 279, 1429), which could phosphorylate Cxs. Indeed, kidneys from ZDF rats displayed increased Cx43 phosphorylation and reduced staining of Cx37 in JGC (1441). Inhibitory peptides against Cx37 and Cx40/Cx43, but not Cx43, attenuated RBF autoregulation in control Zucker rats, but did not further decrement the weak autoregulation in kidneys of ZDF rats (1441). Similar changes in Cxs in VSMCs may contribute to abnormal myogenic and MD-TGF responses during diabetes. Myogenic constrictor and ACh-induced vasodilation of afferent arterioles of the HNK of ZDF rats were both blunted but were improved by sensitization to insulin release with troglitazone (591). Insulin buffered ANG II and NE-induced vasoconstriction in the HNK of normal rats that was attributed to increased NO, but these effects were attenuated in ZDF rats (591), perhaps reflecting oxidative stress. A hybrid cross of ZDF with spontaneously hypertensive heart failure rats developed hypertension, proteinuria, and glomerulosclerosis (495) despite efficient steady-state RBF autoregulation (AI ⬍0.1) and dynamic transfer function with normal myogenic and MD-TGF responses (495). Thus factors other than impaired renal autoregulation are responsible for kidney damage in this model of DMT2. The myogenic tone of skeletal muscle arterioles from ZDF rats was normal initially but became enhanced in older hypertensive animals by a ROS-dependent mechanism that diminished NO signaling (444, 885). Cerebral arteries of ZDF rats also had reduced vasodilation to ACh or hypoxia and enhanced ROS-dependent myogenic constriction (1180). Although generation of ROS in the ZDF rat may underlie enhanced myogenic responses in nonrenal vascular beds, it is unclear whether this occurs in the renal microcirculation. Otsuka Long-Evans Tokushima rats develop late-onset, non-insulin-dependent hyperglycemia, moderate obesity, mild hypertension, and increased GFR and RBF (1506). An impaired steady-state autoregulation of total and deep cortical blood flow and MD-TGF preceded the diabetic phase and persisted into the established phase (584). The PGC in glomeruli of deep versus superficial nephrons was increased by 20 mmHg (584). This interesting model with insulin resistance and later development of DMT2 has a weakened MD-TGF mechanism that may underlie increased RBF and PGC, impaired autoregulation, and subsequent renal injury. Goto-Kakizaki rats are a lean Wistar substrain that develops early-onset DMT2 with hyperglycemia despite normal insulin levels followed by renal and glomerular hypertrophy, glomerular basement membrane thickening, mesangial expansion, and albuminuria (1327) despite normal BP and reduced GFR (784, 1327). Hypertension induced by DOCA-salt accelerated the development of proteinuria and nephropathy (234). Thus this is a model of a lean, hyperglycemic, euinsulinemic, normotensive DMT2 with prominent renal functional and structural changes characteristic of human diabetic nephropathy. The myogenic responses were maintained or augmented in cerebral and mesenteric arteries of diabetic Goto-Kakizaki rats (784, 785, 1269), with normalization by glycemic control (784, 1269). Renal hemodynamics and autoregulatory mechanisms have not been characterized in this intriguing model. The leptin receptor-deficient db⫺/db⫺ mouse model of DMT2 develops dyslipidemia, obesity, hyperglycemia, glomerular hypertrophy, and albuminuria (1355) with increased GFR while BP is normal (888, 889). MD-TGF activity was normal (888, 889), but after correction for the depleted ECV by the osmatic diuresis from glucosuria the SNGFR increased further and the MD-TGF activity was abolished (889). The increase in GFR in volume-replete db⫺/db⫺ mice was ascribed to overproduction of NO by nNOS in the MD (889). Afferent arterioles in the JMN preparation of db⫺/db⫺mice were dilated, but had normal autoregulation to increased RPP and normal contractions to ANG II (1155), suggesting a normal myogenic response. MD-TGF was normal in JMN but impaired in superficial nephrons, which might reflect differences in A1 receptor signaling. Myogenic tone of the preglomerular vasculature in the absence of MD-TGF has not been investigated to date. Mixed results have been reported for nonrenal vessels from db⫺/db⫺ mice. Myogenic tone was increased in mesenteric Physiol Rev • VOL 95 • APRIL 2015 • www.prv.org 469 RENAL AUTOREGULATORY MECHANISMS and coronary arteries but was normalized by inhibition of epidermal growth factor receptor tyrosine kinase (92, 983) and was related to activation of a vasoconstrictor prostanoid activating TP receptors and to reduce NO (858). Likewise, myogenic tone of gracilis skeletal muscle arterioles was enhanced by a constrictor prostanoid produced by COX-2 that activated TP receptors secondary to increased H2O2 production (61, 382). However, other studies of coronary arteries reported preserved myogenic tone and normal TP responses (1017) despite reduced NO and increased ROS production (62). of MD-TGF, following enhanced proximal reabsorption, may contribute to afferent arteriolar dilation and increased SNGFR. Glomerular damage can be dissociated from hypertension and impaired renal autoregulation in some models, thereby highlighting the importance of other, as yet undiscovered, mechanism of renal damage in DMT2. 3. Clinical studies of renal autoregulation in diabetes and nephropathy Christensen and co-workers (251–255) studied GFR from measurements of inulin clearance in human diabetics before and after an acute 15–20 mmHg reduction in mean BP produced by oral clonidine (FIGURE 9). The dotted line in FIGURE 9 indicates the GFR response without any autoregulation (AI ⫽ 1.0). Normal subjects (FIGURE 9F) had excellent autoregulation with only a 1–2% reduction in GFR when BP was reduced by 18% (253). GFR autoregulation In summary, studies in rodent models of hyperglycemic, insulin-resistant DMT2 have generally demonstrated renal vasodilation, increased PGC, and GFR. Renal autoregulation and the myogenic and MD-TGF mechanisms are often normal or modestly attenuated. Autoregulation is generally improved by glycemic control or by an insulin-sensitizing agent. Resetting A DMT1 (10 yrs); no nephropathy ΔMAP (%) -15 -20 -10 -5 0 B -20 0 C DMT1 (27 yrs); no nephropathy ΔMAP (%) -15 -10 -5 0 -20 0 -15 -10 -5 -5 Normal blood glucose -10 -5 Control ΔGFR (%) -15 -10 Spironolactone -20 DMT2 (14 yrs); no nephropathy ΔMAP (%) -20 -15 -10 -5 0 E 0 -5 ΔGFR (%) -15 -10 -5 -10 -15 -15 -20 -20 DMT2 (15 yrs); with or without nephropathy ΔMAP (%) -20 0 Control F Non-diabetic nephropathy ΔMAP (%) -20 0 0 ΔGFR (%) -15 -10 -5 0 0 No nephropathy Irsadipine -5 Control -5 -5 Nephropathy -10 ΔGFR (%) -10 ΔGFR (%) Nephropathy -10 -15 -15 -15 -20 -20 -20 FIGURE 9. Data have been redrawn from studies of autoregulation of glomerular filtration rate (GFR) during short-term reductions in mean arterial pressure (MAP) produced by oral clonidine administration to patients with diabetes mellitus type 1 (DMT1) or type 2 (DMT2), and/or nephropathy (broken line and open circle) and appropriate control subjects (solid line and closed circle). Perfect autoregulation (AI ⫽ 0) is equated with no change in GFR with MAP. An absence of autoregulation (AI ⫽ 1.0) with equal changes in GFR and MAP is depicted by the dotted lines in each panel. A: patients with DMT1 but without nephropathy at an average of 10 yr after diagnosis. Those with high blood glucose had a similar autoregulation as those with normal blood glucose (control) (255). B: patients with DMT1 but without nephropathy at an average of 27 yr after diagnosis. Autoregulation was impaired in subjects with DMT1 (control), but was not altered by mineralocorticosteroid blockade with spironolactone (1293). C: patients with DMT2 but without nephropathy at an average of 13 yr after diagnosis. Autoregulation was mildly impaired (control), but was not affected by blockade of AT1 receptors with cansdesartan (254). D: patients with DMT2 without nephropathy at ⬃14 yr after diagnosis. Autoregulation was quite well maintained (control), but was impaired severely by blockade of L-type VOCCs with isradipine (251). E: patients with DMT2 at 15 yr after diagnosis. Autoregulation was quite well maintained in the group without nephropathies (control) but was impaired in those with nephropathy (252). F: autoregulation in normal subjects was excellent (control), but was impaired in those with nondiabetic nephropathy (253). 470 0 Control Blood glucose D DMT2 (13 yrs); no nephropathy ΔMAP (%) Physiol Rev • VOL 95 • APRIL 2015 • www.prv.org ΔGFR (%) CARLSTRÖM ET AL. also was well maintained after ⬃10 yr of DMT1, even with poor glycemic control (FIGURE 9A) (255), but became impaired after ⬃27 yr and was not improved by reduction in BP by a mineralocorticoid receptor blocker (FIGURE 9B) (1293). GFR autoregulation was excellent after ⬃13 yr of DMT2 and was not affected by AT1 receptor blockade (FIGURE 9C) (254), but was almost abolished by VOCC blockade with isradipine (FIGURE 9D) (251). GFR autoregulation was impaired severely with DMT2 and nephropathy (FIGURE 9E) (252) and in other patients with nondiabetic nephropathy (FIGURE 9F) (253). These data demonstrate that GFR autoregulation is well preserved in subjects with early DMT1 or 2, but becomes quite severely impaired later in the disease, with the development of nephropathy or after treatment with a CCB. VIII. CONCLUSIONS Autoregulation relates preglomerular vascular resistance to changes in RPP. It thereby stabilizes the RBF, PGC, and GFR and protects glomerular capillaries from barotrauma during hypertension. Autoregulation in the normal kidney is highly efficient. It involves a unique combination of tubular and vascular control mechanisms (FIGURE 6). The major mechanisms entail a rapid myogenic response and a more delayed MD-TGF response within the JGA. In addition, there appear to be contributions from a poorly characterized third mechanism and modulation by a CT-GF system. Although the myogenic phenomenon was described more than a century ago, the precise mechanisms for sensing mechanical distension and promoting the response in VSMCs are still not fully understood. Increased [Ca2⫹]i in arteriolar VSMCs following mechanical distension is obligate for the initiation of the response, but prolonged, maintained vasoconstriction also entails enhanced Ca2⫹ sensitivity involving PKC, RhoA/Rho-kinase, and/or ROS signaling (FIGURE 8). Initiation can be triggered by stretch-sensitive cation channels, integrin-cytoskeleton interactions, and/or GPCR signaling. VSMC membrane depolarization and Ca2⫹ entry via VOCCs are central to Ca2⫹ signaling for the myogenic and MD-TGF-induced vasoconstrictor components of autoregulation. Impaired renal autoregulation is observed in many, but not all, models of hypertension, diabetes, or intrinsic renal disease. Further investigations are warranted to advance our understanding of the underlying cellular are molecular events in these important basic regulatory mechanisms, since they likely contribute to progression of renal damage. ACKNOWLEDGMENTS Address for reprint requests and other correspondence: M. Carlström, Dept. of Physiology & Pharmacology, Karolinska Institutet, Nanna Svartz Väg 2, S-171 77, Stockholm, Sweden (e-mail: mattias.carlstrom@ki.se). GRANTS The authors’ work was supported by National Heart, Lung, and Blood Institute Grants HL68686 (to C. S. Wilcox) and HL02334 (to W. J. Arendshorst); National Institute of Diabetes and Digestive and Kidney Diseases Grants DK036079 and DK-049870 (to C. S. Wilcox); Swedish Research Council Grant K2012-99X-21971-01-3 (to M. Carlström); Swedish Heart and Lung Foundation Grants 20110589 and 20140448 (to M. Carlström); the Swedish Society for Medical Research (to M. Carlström); Jeanssons Foundation Grants JS20130064 and JS2011-0212 (to M. Carlström); the Wenner-Gren Foundation (to M. Carlström); the Swedish Society of Medicine (to M. Carlström); Karolinska Institutet Research Foundations Grant 2014fobi41264 (to M. Carlström); the George E. Schreiner Chair of Nephrology (to C. S. Wilcox); the Hypertension, Kidney, and Vascular Research Center of Georgetown University (to C. S. Wilcox); the Smith Family Trust Fund (to C. S. Wilcox); as well as the Joseph Gildenhorn/Spiesman Family Foundation Incorporated and by Ms. Alma Gilde (to C. S. Wilcox). DISCLOSURES No conflicts of interest, financial or otherwise, are declared by the authors. REFERENCES 1. Aalkjaer C, Boedtkjer D, Matchkov V. Vasomotion: what is currently thought? Acta Physiol 202: 253–269, 2011. 2. Aalkjaer C, Nilsson H. Vasomotion: cellular background for the oscillator and for the synchronization of smooth muscle cells. Br J Pharmacol 144: 605– 616, 2005. 3. Abboud HE. Mesangial cell biology. Exp Cell Res 318: 979 –985, 2012. 4. Abd El-Rahman RR, Harraz OF, Brett SE, Anfinogenova Y, Mufti RE, Goldman D, Welsh DG. Identification of L- and T-type Ca2⫹ channels in rat cerebral arteries: role in myogenic tone development. Am J Physiol Heart Circ Physiol 304: H58 –H71, 2013. 5. Abe Y, Kishimoto T, Yamamoto K. Effect of angiotensin II antagonist infusion on autoregulation of renal blood flow. Am J Physiol 231: 1267–1271, 1976. 6. Abraham NG, Cao J, Sacerdoti D, Li X, Drummond G. Heme oxygenase: the key to renal function regulation. Am J Physiol Renal Physiol 297: F1137–F1152, 2009. 7. Abu-Amarah I, Ajikobi DO, Bachelard H, Cupples WA, Salevsky FC. Responses of mesenteric and renal blood flow dynamics to acute denervation in anesthetized rats. Am J Physiol Regul Integr Comp Physiol 275: R1543–R1552, 1998. 8. Abu-Amarah I, Bidani AK, Hacioglu R, Williamson GA, Griffin KA. Differential effects of salt on renal hemodynamics and potential pressure transmission in stroke-prone and stroke-resistant spontaneously hypertensive rats. Am J Physiol Renal Physiol 289: F305–F313, 2005. 9. Adams LG, Polzin DJ, Osborne CA, O’Brien TD, Hostetter TH. Influence of dietary protein/calorie intake on renal morphology and function in cats with 5/6 nephrectomy. Lab Invest 70: 347–357, 1994. 10. Adebiyi A, Thomas-Gatewood CM, Leo MD, Kidd MW, Neeb ZP, Jaggar JH. An elevation in physical coupling of type 1 inositol 1,4,5-trisphosphate (IP3) receptors to transient receptor potential 3 (TRPC3) channels constricts mesenteric arteries in genetic hypertension. Hypertension 60: 1213–1219, 2012. Physiol Rev • VOL 95 • APRIL 2015 • www.prv.org 471 RENAL AUTOREGULATORY MECHANISMS 11. Adebiyi A, Zhao G, Cheranov SY, Ahmed A, Jaggar JH. Caveolin-1 abolishment attenuates the myogenic response in murine cerebral arteries. Am J Physiol Heart Circ Physiol 292: H1584 –H1592, 2007. 31. Anderson S, Meyer TW, Rennke HG, Brenner BM. Control of glomerular hypertension limits glomerular injury in rats with reduced renal mass. J Clin Invest 76: 612– 619, 1985. 12. Adler S, Huang H. Oxidant stress in kidneys of spontaneously hypertensive rats involves both oxidase overexpression and loss of extracellular superoxide dismutase. Am J Physiol Renal Physiol 287: F907–F913, 2004. 32. Anderson S, Rennke HG, Brenner BM. Antihypertensive therapy must control glomerular hypertension to limit glomerular injury. J Hypertens Suppl 4: S242–S244, 1986. 13. Agodoa LY, Appel L, Bakris GL, Beck G, Bourgoignie J, Briggs JP, Charleston J, Cheek D, Cleveland W, Douglas JG, Dowie D, Faulkner M, Gabriel A, Gassman J, Greene T, Hall Y, Hebert L, Hiremath L, Jamerson K, Johnson CJ, Kopple J, Kusek J, Lash J, Lea J, Lewis JB, Lipkowitz M, Massry S, Middleton J, Miller ER, Norris K, O’Connor D, Ojo A, Phillips RA, Pogue V, Rahman M, Randall OS, Rostand S, Schulman G, Smith W, Thornley-Brown D, Tisher CC, Toto RD, Wright JT, Xu S. Effect of ramipril vs amlodipine on renal outcomes in hypertensive nephrosclerosis: a randomized controlled trial. JAMA 285: 2719 –2728, 2001. 33. Anderson WP, Kett MM, Alcorn D, Bertram JF. Angiotensin II antagonism and preglomerular arterial wall dimensions in the kidney of the spontaneously hypertensive rat. Clin Exp Hypertens 19: 965–979, 1997. 14. Agrawal S, Temsamani J, Tang JY. Pharmacokinetics, biodistribution, and stability of oligodeoxynucleotide phosphorothioates in mice. Proc Natl Acad Sci USA 88: 7595– 7599, 1991. 15. Aguilera G, Catt KJ. Regulation of vascular angiotensin II receptors in the rat during altered sodium intake. Circ Res 49: 751–758, 1981. 16. Ahmed A, Waters CM, Leffler CW, Jaggar JH. Ionic mechanisms mediating the myogenic response in newborn porcine cerebral arteries. Am J Physiol Heart Circ Physiol 287: H2061–H2069, 2004. 17. Ahmed AK, Kamath NS, El KM, El Nahas AM. The impact of stopping inhibitors of the renin-angiotensin system in patients with advanced chronic kidney disease. Nephrol Dial Transplant 25: 3977–3982, 2010. 18. Ahn DS, Choi SK, Kim YH, Cho YE, Shin HM, Morgan KG, Lee YH. Enhanced stretch-induced myogenic tone in the basilar artery of spontaneously hypertensive rats. J Vasc Res 44: 182–191, 2007. 19. Ajikobi DO, Novak P, Salevsky FC, Cupples WA. Pharmacological modulation of spontaneous renal blood flow dynamics. Can J Physiol Pharmacol 74: 964 –972, 1996. 20. Almeida JB, Saragoca MA, Tavares A, Cezareti ML, Draibe SA, Ramos OL. Severe hypertension induces disturbances of renal autoregulation. Hypertension 19: II279 – II283, 1992. 21. Alonso-Galicia M, Drummond HA, Reddy KK, Falck JR, Roman RJ. Inhibition of 20-HETE production contributes to the vascular responses to nitric oxide. Hypertension 29: 320 –325, 1997. 22. Amann K, Wiest G, Zimmer G, Gretz N, Ritz E, Mall G. Reduced capillary density in the myocardium of uremic rats–a stereological study. Kidney Int 42: 1079 –1085, 1992. 23. Ambudkar IS, Brazer SC, Liu X, Lockwich T, Singh B. Plasma membrane localization of TRPC channels: role of caveolar lipid rafts. Novartis Found Symp 258: 63–70, 2004. 24. Ances BM, Greenberg JH, Detre JA. Interaction between nitric oxide synthase inhibitor induced oscillations and the activation flow coupling response. Brain Res 1309: 19 –28, 2010. 25. Anderson M, Kim EY, Hagmann H, Benzing T, Dryer SE. Opposing effects of podocin on the gating of podocyte TRPC6 channels evoked by membrane stretch or diacylglycerol. Am J Physiol Cell Physiol 305: C276 –C289, 2013. 26. Anderson RJ, Pluss RG, Pluss WT, Bell J, Zerbe GG. Effect of hypoxia and hypercapnic acidosis on renal autoregulation in the dog: role of renal nerves. Clin Sci 65: 533–538, 1983. 27. Anderson RJ, Taher MS, Cronin RE, McDonald KM, Schrier RW. Effect of -adrenergic blockade and inhibitors of angiotensin II and prostaglandins on renal autoregulation. Am J Physiol 229: 731–736, 1975. 28. Anderson S. Antihypertensive therapy in experimental diabetes. J Am Soc Nephrol 3: S86 –S90, 1992. 29. Anderson S, Brenner BM. Intraglomerular hypertension: implications and drug treatment. Annu Rev Med 39: 243–253, 1988. 30. Anderson S, Brenner BM. Pathogenesis of diabetic glomerulopathy: hemodynamic considerations. Diabetes Metab Rev 4: 163–177, 1988. 472 34. Ando K, Ono A, Sato Y, Fujita T. Involvement of prostaglandins and renal haemodynamics in salt-sensitivity of young spontaneously hypertensive rats. J Hypertens 11: 373–377, 1993. 35. Andreasen D, Friis UG, Uhrenholt TR, Jensen BL, Skott O, Hansen PB. Coexpression of voltage-dependent calcium channels Cav1.2, 21a, and 21b in vascular myocytes. Hypertension 47: 735–741, 2006. 36. Anfinogenova Y, Brett SE, Walsh MP, Harraz OF, Welsh DG. Do TRPC-like currents and G protein-coupled receptors interact to facilitate myogenic tone development? Am J Physiol Heart Circ Physiol 301: H1378 –H1388, 2011. 37. Ankorina-Stark I, Haxelmans S, Schlatter E. Receptors for bradykinin and prostaglandin E2 coupled to Ca2⫹ signalling in rat cortical collecting duct. Cell Calcium 22: 269 –275, 1997. 38. Araujo M, Welch WJ. Cyclooxygenase 2 inhibition suppresses tubuloglomerular feedback: roles of thromboxane receptors and nitric oxide. Am J Physiol Renal Physiol 296: F790 –F794, 2009. 39. Araujo M, Welch WJ. Tubuloglomerular feedback is decreased in COX-1 knockout mice after chronic angiotensin II infusion. Am J Physiol Renal Physiol 298: F1059 – F1063, 2010. 40. Araujo M, Wilcox CS. Oxidative stress in hypertension: role of the kidney. Antioxid Redox Signal 20: 74 –101, 2013. 41. Arendshorst WJ. Autoregulation of renal blood flow in spontaneously hypertensive rats. Circ Res 44: 344 –349, 1979. 42. Arendshorst WJ, Beierwaltes WH. Renal and nephron hemodynamics in spontaneously hypertensive rats. Am J Physiol Renal Fluid Electrolyte Physiol 236: F246 –F251, 1979. 43. Arendshorst WJ, Beierwaltes WH. Renal tubular reabsorption in spontaneously hypertensive rats. Am J Physiol Renal Fluid Electrolyte Physiol 237: F38 –F47, 1979. 44. Arendshorst WJ, Chatziantoniou C, Daniels FH. Role of angiotensin in the renal vasoconstriction observed during the development of genetic hypertension. Kidney Int Suppl 30: S92–S96, 1990. 45. Arendshorst WJ, Finn WF. Renal hemodynamics in the rat before and during inhibition of angiotensin II. Am J Physiol Renal Fluid Electrolyte Physiol 233: F290 –F297, 1977. 46. Arendshorst WJ, Finn WF, Gottschalk CW. Autoregulation of blood flow in the rat kidney. Am J Physiol 228: 127–133, 1975. 47. Arendshorst WJ, Gottschalk CW. Glomerular ultrafiltration dynamics: euvolemic and plasma volume-expanded rats. Am J Physiol Renal Fluid Electrolyte Physiol 239: F171–F186, 1980. 48. Arendshorst WJ, Navar LG. Renal Circulation and Glomerular Hemodynamics. In: Diseases of the Kidney & Urinary Tract, edited by Schrier RW, Nielsen EH, Molitoris BA, Coffman , Falk RJ. Philadelphia, PA: Lippincott Williams & Wilkins, 2012. 49. Arendshorst WJ, Thai TL. Regulation of the renal microcirculation by ryanodine receptors and calcium-induced calcium release. Curr Opin Nephrol Hypertens 18: 40 – 49, 2009. 50. Arensbak B, Mikkelsen HB, Gustafsson F, Christensen T, Holstein-Rathlou NH Holstein-Rathlou NH. Expression of connexin 37, 40, and 43 mRNA and protein in renal preglomerular arterioles. Histochem Cell Biol 115: 479 – 487, 2001. 51. Arima S, Ito S. Isolated juxtaglomerular apparatus as a tool for exploring glomerular hemodynamics: application of microperfusion techniques. Exp Nephrol 8: 304 –311, 2000. Physiol Rev • VOL 95 • APRIL 2015 • www.prv.org CARLSTRÖM ET AL. 52. Arima S, Ito S, Omata K, Tsunoda K, Yaoita H, Abe K. Diverse effects of calcium antagonists on glomerular hemodynamics. Kidney Int Suppl 55: S132–S134, 1996. 53. Arima S, Kohagura K, Xu HL, Sugawara A, Abe T, Satoh F, Takeuchi K, Ito S. Nongenomic vascular action of aldosterone in the glomerular microcirculation. J Am Soc Nephrol 14: 2255–2263, 2003. 54. Arima S, Kohagura K, Xu HL, Sugawara A, Uruno A, Satoh F, Takeuchi K, Ito S. Endothelium-derived nitric oxide modulates vascular action of aldosterone in renal arteriole. Hypertension 43: 352–357, 2004. 55. Asaba K, Tojo A, Onozato ML, Goto A, Quinn MT, Fujita T, Wilcox CS. Effects of NADPH oxidase inhibitor in diabetic nephropathy. Kidney Int 67: 1890 –1898, 2005. 56. Aukes AM, Vitullo L, Zeeman GG, Cipolla MJ. Pregnancy prevents hypertensive remodeling and decreases myogenic reactivity in posterior cerebral arteries from Dahl salt-sensitive rats: a role in eclampsia? Am J Physiol Heart Circ Physiol 292: H1071–H1076, 2007. 57. Aukland K. Myogenic mechanisms in the kidney. J Hypertens Suppl 7: S71–S76, 1989. 58. Azar S, Johnson MA, Scheinman J, Bruno L, Tobian L. Regulation of glomerular capillary pressure and filtration rate in young Kyoto hypertensive rats. Clin Sci 56: 203–209, 1979. 59. Baek EB, Kim SJ. Mechanisms of myogenic response: Ca2⫹-dependent and -independent signaling. J Smooth Muscle Res 47: 55– 65, 2011. 60. Bagher P, Segal SS. Regulation of blood flow in the microcirculation: role of conducted vasodilation. Acta Physiol 202: 271–284, 2011. 61. Bagi Z, Erdei N, Toth A, Li W, Hintze TH, Koller A, Kaley G. Type 2 diabetic mice have increased arteriolar tone and blood pressure: enhanced release of COX-2derived constrictor prostaglandins. Arterioscler Thromb Vasc Biol 25: 1610 –1616, 2005. 62. Bagi Z, Koller A, Kaley G. Superoxide-NO interaction decreases flow- and agonistinduced dilations of coronary arterioles in Type 2 diabetes mellitus. Am J Physiol Heart Circ Physiol 285: H1404 –H1410, 2003. 63. Bakker EN, Kerkhof CJ, Sipkema P. Signal transduction in spontaneous myogenic tone in isolated arterioles from rat skeletal muscle. Cardiovasc Res 41: 229 –236, 1999. 64. Balasubramanian L, Ahmed A, Lo CM, Sham JS, Yip KP. Integrin-mediated mechanotransduction in renal vascular smooth muscle cells: activation of calcium sparks. Am J Physiol Regul Integr Comp Physiol 293: R1586 –R1594, 2007. 65. Balasubramanian L, Lo CM, Sham JS, Yip KP. Remanent cell traction force in renal vascular smooth muscle cells induced by integrin-mediated mechanotransduction. Am J Physiol Cell Physiol 304: C382–C391, 2013. 66. Bank N, Lahorra G, Aynedjian HS, Wilkes BM. Sodium restriction corrects hyperfiltration of diabetes. Am J Physiol Renal Fluid Electrolyte Physiol 254: F668 –F676, 1988. 67. Barajas L, Powers K, Carretero OA, Scicli AG, Inagami T. Immunocytochemical localization of renin and kallikrein in the rat renal cortex. Kidney Int 29: 965–970, 1986. 73. Baylis C, Atzpodien EA, Freshour G, Engels K. Peroxisome proliferator-activated receptor [gamma] agonist provides superior renal protection versus angiotensinconverting enzyme inhibition in a rat model of type 2 diabetes with obesity. J Pharmacol Exp Ther 307: 854 – 860, 2003. 74. Baylis C, Ichikawa I, Willis WT, Wilson CB, Brenner BM. Dynamics of glomerular ultrafiltration. IX. Effects of plasma protein concentration. Am J Physiol Renal Fluid Electrolyte Physiol 232: F58 –F71, 1977. 75. Baylis C, Qiu C, Engels K. Comparison of L-type and mixed L- and T-type calcium channel blockers on kidney injury caused by deoxycorticosterone-salt hypertension in rats. Am J Kidney Dis 38: 1292–1297, 2001. 76. Bayliss WM. On the local reactions of the arterial wall to changes of internal pressure. J Physiol 28: 220 –231, 1902. 77. Beaufils M, Sraer J, Lepreux C, Ardaillou R. Angiotensin II binding to renal glomeruli from sodium-loaded and sodium-depleted rats. Am J Physiol 231: 1187–1193, 1976. 78. Bedard K, Krause KH. The NOX family of ROS-generating NADPH oxidases: physiology and pathophysiology. Physiol Rev 87: 245–313, 2007. 79. Beierwaltes WH, Arendshorst WJ, Klemmer PJ. Electrolyte and water balance in young spontaneously hypertensive rats. Hypertension 4: 908 –915, 1982. 80. Beierwaltes WH, Schryver S, Britton SL, Romero JC. Renal pressure-flow relationships in severely hypertensive rabbits. Proc Soc Exp Biol Med 164: 495– 499, 1980. 81. Beierwaltes WH, Sigmon DH, Carretero OA. Endothelium modulates renal blood flow but not autoregulation. Am J Physiol Renal Fluid Electrolyte Physiol 262: F943– F949, 1992. 82. Bell PD. Calcium antagonists and intrarenal regulation of glomerular filtration rate. Am J Nephrol 7 Suppl 1: 24 –31, 1987. 83. Bell PD, Komlosi P, Zhang ZR. ATP as a mediator of macula densa cell signalling. Purinergic Signal 5: 461– 471, 2009. 84. Bell PD, LaPointe JY, Peti-Peterdi J. Macula densa cell signaling. Annu Rev Physiol 65: 481–500, 2003. 85. Bell PD, LaPointe JY, Sabirov R, Hayashi S, Peti-Peterdi J, Manabe K, Kovacs G, Okada Y. Macula densa cell signaling involves ATP release through a maxi anion channel. Proc Natl Acad Sci USA 100: 4322– 4327, 2003. 86. Bell PD, Navar LG. Stop-flow pressure feedback responses during reduced renal vascular resistance in the dog. Am J Physiol Renal Fluid Electrolyte Physiol 237: F204 – F209, 1979. 87. Bell PD, Reddington M, Ploth D, Navar LG. Tubuloglomerular feedback-mediated decreases in glomerular pressure in Munich-Wistar rats. Am J Physiol Renal Fluid Electrolyte Physiol 247: F877–F880, 1984. 88. Bell TD, DiBona GF, Biemiller R, Brands MW. Continuously measured renal blood flow does not increase in diabetes if nitric oxide synthesis is blocked. Am J Physiol Renal Physiol 295: F1449 –F1456, 2008. 68. Bauer J, Parekh N. Variations in cell signaling pathways for different vasoconstrictor agonists in renal circulation of the rat. Kidney Int 63: 2178 –2186, 2003. 89. Bell TD, DiBona GF, Wang Y, Brands MW. Mechanisms for renal blood flow control early in diabetes as revealed by chronic flow measurement and transfer function analysis. J Am Soc Nephrol 17: 2184 –2192, 2006. 69. Baumann JE, Persson PB, Ehmke H, Nafz B, Kirchheim HR. Role of endotheliumderived relaxing factor in renal autoregulation in conscious dogs. Am J Physiol Renal Fluid Electrolyte Physiol 263: F208 –F213, 1992. 90. Belleau LJ, Earley LE. Autoregulation of renal blood flow in the presence of angiotensin infusion. Am J Physiol 213: 1590 –1595, 1967. 70. Baumann M, Janssen BJ, Rob Hermans JJ, Bartholome R, Smits JF, Struijker-Boudier HA. Renal medullary effects of transient prehypertensive treatment in young spontaneously hypertensive rats. Acta Physiol 196: 231–237, 2009. 71. Baumgartl H, Zimelka W, Lubbers DW. Effects of puncturing on the measurement of local oxygen pressure using polarographic microelectrodes. Oxygen Transport Tissue 18: 527–549, 1997. 72. Bax L, Woittiez AJ, Kouwenberg HJ, Mali WP, Buskens E, Beek FJ, Braam B, Huysmans FT, Schultze Kool LJ, Rutten MJ, Doorenbos CJ, Aarts JC, Rabelink TJ, Plouin PF, Raynaud A, van Montfrans GA, Reekers JA, van den Meiracker AH, Pattynama PM, van de Ven PJ, Vroegindeweij D, Kroon AA, de Haan MW, Postma CT, Beutler JJ. Stent placement in patients with atherosclerotic renal artery stenosis and impaired renal function: a randomized trial. Ann Intern Med 150: 840 – 841, 2009. 91. Bellucci A, Wilkes BM. Mechanism of sodium modulation of glomerular angiotensin receptors in the rat. J Clin Invest 74: 1593–1600, 1984. 92. Belmadani S, Palen DI, Gonzalez-Villalobos RA, Boulares HA, Matrougui K. Elevated epidermal growth factor receptor phosphorylation induces resistance artery dysfunction in diabetic db/db mice. Diabetes 57: 1629 –1637, 2008. 93. Benstein JA, Feiner HD, Parker M, Dworkin LD. Superiority of salt restriction over diuretics in reducing renal hypertrophy and injury in uninephrectomized SHR. Am J Physiol Renal Fluid Electrolyte Physiol 258: F1675–F1681, 1990. 94. Berecek KH, Schwertschlag U, Gross F. Alterations in renal vascular resistance and reactivity in spontaneous hypertension of rats. Am J Physiol Heart Circ Physiol 238: H287–H293, 1980. Physiol Rev • VOL 95 • APRIL 2015 • www.prv.org 473 RENAL AUTOREGULATORY MECHANISMS 95. Bergstrom G, Johansson I, Wickman A, Gan L, Thorup C. Brief losartan treatment in young spontaneously hypertensive rats abates long-term blood pressure elevation by effects on renal vascular structure. J Hypertens 20: 1413–1421, 2002. 116. Blantz RC, Deng A, Lortie M, Munger K, Vallon V, Gabbai FB, Thomson SC. The complex role of nitric oxide in the regulation of glomerular ultrafiltration. Kidney Int 61: 782–785, 2002. 96. Bergstrom G, Rudenstam J, Taghipour K, Gothberg G, Karlstrom G. Effect of nitric oxide and renal nerves on renomedullary haemodynamics in SHR and Wistar rats, studied with laser Doppler technique. Acta Physiol Scand 156: 27–35, 1995. 117. Blantz RC, Gabbai FB, Peterson OW, Thomson SC. Tubuloglomerular feedback activity after acute reductions in renal mass. Kidney Int Suppl 32: S102–S105, 1991. 97. Berthold H, Munter K, Just A, Kirchheim HR, Ehmke H. Contribution of endothelin to renal vascular tone and autoregulation in the conscious dog. Am J Physiol Renal Physiol 276: F417–F424, 1999. 98. Bertuglia S, Colantuoni A, Intaglietta M. Capillary reperfusion after L-arginine, LNMMA, and L-NNA treatment in cheek pouch microvasculature. Microvasc Res 50: 162–174, 1995. 99. Berwick ZC, Moberly SP, Kohr MC, Morrical EB, Kurian MM, Dick GM, Tune JD. Contribution of voltage-dependent K⫹ and Ca2⫹ channels to coronary pressureflow autoregulation. Basic Res Cardiol 107: 264 – 0264, 2012. 100. Beswick RA, Zhang H, Marable D, Catravas JD, Hill WD, Webb RC. Long-term antioxidant administration attenuates mineralocorticoid hypertension and renal inflammatory response. Hypertension 37: 781–786, 2001. 101. Bhattachariya A, Dahan D, Turczynska KM, Sward K, Hellstrand P, Albinsson S. Expression of microRNAs is essential for arterial myogenic tone and pressure-induced activation of the PI3-kinase/Akt pathway. Cardiovasc Res 101: 288 –296, 2014. 102. Bidani AK, Griffin KA. Calcium channel blockers and renal protection: is there an optimal dose? J Lab Clin Med 125: 553–555, 1995. 103. Bidani AK, Griffin KA. Long-term renal consequences of hypertension for normal and diseased kidneys. Curr Opin Nephrol Hypertens 11: 73– 80, 2002. 104. Bidani AK, Griffin KA. Pathophysiology of hypertensive renal damage: implications for therapy. Hypertension 44: 595– 601, 2004. 105. Bidani AK, Griffin KA, Bakris G, Picken MM. Lack of evidence of blood pressureindependent protection by renin-angiotensin system blockade after renal ablation. Kidney Int 57: 1651–1661, 2000. 106. Bidani AK, Griffin KA, Picken M, Lansky DM. Continuous telemetric blood pressure monitoring and glomerular injury in the rat remnant kidney model. Am J Physiol Renal Fluid Electrolyte Physiol 265: F391–F398, 1993. 107. Bidani AK, Griffin KA, Plott W, Schwartz MM. Renal ablation acutely transforms “benign” hypertension to “malignant” nephrosclerosis in hypertensive rats. Hypertension 24: 309 –316, 1994. 108. Bidani AK, Griffin KA, Williamson G, Wang X, Loutzenhiser R. Protective importance of the myogenic response in the renal circulation. Hypertension 54: 393–398, 2009. 109. Bidani AK, Hacioglu R, Abu-Amarah I, Williamson GA, Loutzenhiser R, Griffin KA. “Step” vs “dynamic” autoregulation: implications for susceptibility to hypertensive injury. Am J Physiol Renal Physiol 285: F113–F120, 2003. 110. Bidani AK, Mitchell KD, Schwartz MM, Navar LG, Lewis EJ. Absence of glomerular injury or nephron loss in a normotensive rat remnant kidney model. Kidney Int 38: 28 –38, 1990. 111. Bidani AK, Polichnowski AJ, Loutzenhiser R, Griffin KA. Renal microvascular dysfunction, hypertension and CKD progression. Curr Opin Nephrol Hypertens 22: 1–9, 2013. 112. Bidani AK, Schwartz MM, Lewis EJ. Renal autoregulation and vulnerability to hypertensive injury in remnant kidney. Am J Physiol Renal Fluid Electrolyte Physiol 252: F1003–F1010, 1987. 113. Bilous R, Chaturvedi N, Sjolie AK, Fuller J, Klein R, Orchard T, Porta M, Parving HH. Effect of candesartan on microalbuminuria and albumin excretion rate in diabetes: three randomized trials. Ann Intern Med 151: 11–14, 2009. 114. Biswas SK, de Faria JB. Which comes first: renal inflammation or oxidative stress in spontaneously hypertensive rats? Free Radic Res 41: 216 –224, 2007. 115. Black MJ, Wang Y, Bertram JF. Nephron endowment and renal filtration surface area in young spontaneously hypertensive rats. Kidney Blood Press Res 25: 20 –26, 2002. 474 118. Blantz RC, Konnen KS, Tucker BJ. Glomerular filtration response to elevated ureteral pressure in both the hydropenic and the plasma-expanded rat. Circ Res 37: 819 – 829, 1975. 119. Blaustein MP, Hamlyn JM. Signaling mechanisms that link salt retention to hypertension: endogenous ouabain, the Na⫹ pump, the Na⫹/Ca2⫹ exchanger and TRPC proteins. Biochim Biophys Acta 1802: 1219 –1229, 2010. 120. Blaustein MP, Lederer WJ. Sodium/calcium exchange: its physiological implications. Physiol Rev 79: 763– 854, 1999. 121. Bobadilla NA, Tack I, Tapia E, Sanchez-Lozada LG, Santamaria J, Jimenez F, Striker LJ, Striker GE, Herrera-Acosta J. Pentosan polysulfate prevents glomerular hypertension and structural injury despite persisting hypertension in 5/6 nephrectomy rats. J Am Soc Nephrol 12: 2080 –2087, 2001. 122. Bock HA, Hermle M, Fiallo A, Osgood RW, Fried TA. Measurement of renin secretion in single perfused rabbit glomeruli. Am J Physiol Renal Fluid Electrolyte Physiol 258: F1460 –F1465, 1990. 123. Boedtkjer DM, Matchkov VV, Boedtkjer E, Nilsson H, Aalkjaer C. Vasomotion has chloride-dependency in rat mesenteric small arteries. Pflügers Arch 457: 389 – 404, 2008. 124. Boesen EI, Williams DL, Pollock JS, Pollock DM. Immunosuppression with mycophenolate mofetil attenuates the development of hypertension and albuminuria in deoxycorticosterone acetate-salt hypertensive rats. Clin Exp Pharmacol Physiol 37: 1016 –1022, 2010. 125. Boettcher M, de Wit C. Distinct endothelium-derived hyperpolarizing factors emerge in vitro and in vivo and are mediated in part via connexin 40-dependent myoendothelial coupling. Hypertension 57: 802– 808, 2011. 126. Bogeski I, Kappl R, Kummerow C, Gulaboski R, Hoth M, Niemeyer BA. Redox regulation of calcium ion channels: chemical and physiological aspects. Cell Calcium 50: 407– 423, 2011. 127. Bohlen HG. Enhanced cerebral vascular regulation occurs by age 4 to 5 weeks in spontaneously hypertensive rats. Hypertension 9: 325–331, 1987. 128. Bokoch GM, Zhao T. Regulation of the phagocyte NADPH oxidase by Rac GTPase. Antioxid Redox Signal 8: 1533–1548, 2006. 129. Bolz SS, Vogel L, Sollinger D, Derwand R, de Wit C, Loirand G, Pohl U. Nitric oxide-induced decrease in calcium sensitivity of resistance arteries is attributable to activation of the myosin light chain phosphatase and antagonized by the RhoA/Rho kinase pathway. Circulation 107: 3081–3087, 2003. 130. Botros FT, Dobrowolski L, Navar LG. Renal heme oxygenase-1 induction with hemin augments renal hemodynamics, renal autoregulation, and excretory function. Int J Hypertens 2012: 1– 8, 2012. 131. Botros FT, Prieto-Carrasquero MC, Martin VL, Navar LG. Heme oxygenase induction attenuates afferent arteriolar autoregulatory responses. Am J Physiol Renal Physiol 295: F904 –F911, 2008. 132. Bouriquet N, Casellas D. Interaction between cGMP-dependent dilators and autoregulation in rat preglomerular vasculature. Am J Physiol Renal Fluid Electrolyte Physiol 268: F338 –F346, 1995. 133. Braam B, Koomans HA. Nitric oxide antagonizes the actions of angiotensin II to enhance tubuloglomerular feedback responsiveness. Kidney Int 48: 1406 –1411, 1995. 134. Braam B, Navar LG, Mitchell KD. Modulation of tubuloglomerular feedback by angiotensin II type 1 receptors during the development of Goldblatt hypertension. Hypertension 25: 1232–1237, 1995. 135. Brakemeier S, Eichler I, Knorr A, Fassheber T, Kohler R, Hoyer J. Modulation of Ca2⫹-activated K⫹ channel in renal artery endothelium in situ by nitric oxide and reactive oxygen species. Kidney Int 64: 199 –207, 2003. Physiol Rev • VOL 95 • APRIL 2015 • www.prv.org CARLSTRÖM ET AL. 136. Brandes RP, Kreuzer J. Vascular NADPH oxidases: molecular mechanisms of activation. Cardiovasc Res 65: 16 –27, 2005. 137. Brannstrom K, Arendshorst WJ. Thromboxane A2 contributes to the enhanced tubuloglomerular feedback activity in young SHR. Am J Physiol Renal Physiol 276: F758 –F766, 1999. 138. Brannstrom K, Morsing P, Arendshorst WJ. Exaggerated tubuloglomerular feedback activity in genetic hypertension is mediated by ANG II and AT1 receptors. Am J Physiol Renal Fluid Electrolyte Physiol 270: F749 –F755, 1996. 139. Brannstrom K, Morsing P, Arendshorst WJ. Candesartan normalizes exaggerated tubuloglomerular feedback activity in young spontaneously hypertensive rats. J Am Soc Nephrol 10: S213–S219, 1999. 140. Braun C, Lang C, Hocher B, van der Woude FJ, Rohmeiss P. Influence of the renal endothelin A system on the autoregulation of renal hemodynamics in SHRs and WKY rats. J Cardiovasc Pharmacol 31: 643– 648, 1998. 141. Brayden JE, Earley S, Nelson MT, Reading S. Transient receptor potential (TRP) channels, vascular tone and autoregulation of cerebral blood flow. Clin Exp Pharmacol Physiol 35: 1116 –1120, 2008. 142. Brayden JE, Li Y, Tavares MJ. Purinergic receptors regulate myogenic tone in cerebral parenchymal arterioles. J Cereb Blood Flow Metab 33: 293–299, 2013. 143. Brekke JF, Jackson WF, Segal SS. Arteriolar smooth muscle Ca2⫹ dynamics during blood flow control in hamster cheek pouch. J Appl Physiol 101: 307–315, 2006. 144. Brenner BM. Nephron adaptation to renal injury or ablation. Am J Physiol Renal Fluid Electrolyte Physiol 249: F324 –F337, 1985. 145. Brenner BM, Anderson S. Glomerular function in diabetes mellitus. Adv Nephrol Necker Hosp 19: 135–144, 1990. 158. Brown SA, Finco DR, Crowell WA, Choat DC, Navar LG. Single-nephron adaptations to partial renal ablation in the dog. Am J Physiol Renal Fluid Electrolyte Physiol 258: F495–F503, 1990. 159. Brown SA, Finco DR, Crowell WA, Navar LG. Dietary protein intake and the glomerular adaptations to partial nephrectomy in dogs. J Nutr 121: S125–S127, 1991. 160. Brown SA, Finco DR, Navar LG. Impaired renal autoregulatory ability in dogs with reduced renal mass. J Am Soc Nephrol 5: 1768 –1774, 1995. 161. Bruno L, Azar S, Weller D. Absence of a pre-hypertensive stage in post-natal Kyoto hypertensive rats. Jpn Heart J 90 –92, 1997. 162. Buhrle CP, Hackenthal E, Helmchen U, Lackner K, Nobiling R, Steinhausen M, Taugner R. The hydronephrotic kidney of the mouse as a tool for intravital microscopy and in vitro electrophysiological studies of renin-containing cells. Lab Invest 54: 462– 472, 1986. 163. Buhrle CP, Hackenthal E, Nobiling R, Skott O, Baumbach L, Taugner R. Tachyphylaxis of juxtaglomerular epithelioid cells to angiotensin II. Differences between the electrical membrane response and renin secretion. Pflügers Arch 410: 55– 62, 1987. 164. Buhrle CP, Nobiling R, Mannek E, Schneider D, Hackenthal E, Taugner R. The afferent glomerular arteriole: immunocytochemical and electrophysiological investigations. J Cardiovasc Pharmacol 6 Suppl 2: S383–S393, 1984. 165. Bulley S, Neeb ZP, Burris SK, Bannister JP, Thomas-Gatewood CM, Jangsangthong W, Jaggar JH. TMEM16A channels contribute to the myogenic response in cerebral arteries. Circ Res 111: 1027–1036, 2012. 166. Bund SJ. Contractility of resistance arteries of spontaneously hypertensive rats related to their media: lumen ratio. J Hypertens 18: 1223–1231, 2000. 167. Bund SJ. Spontaneously hypertensive rat resistance artery structure related to myogenic and mechanical properties. Clin Sci 101: 385–393, 2001. 146. Brenner BM, Cooper ME, de Zeeuw D, Keane WF, Mitch WE, Parving HH, Remuzzi G, Snapinn SM, Zhang Z, Shahinfar S. Effects of losartan on renal and cardiovascular outcomes in patients with type 2 diabetes and nephropathy. N Engl J Med 345: 861– 869, 2001. 168. Burke M, Pabbidi M, Fan F, Ge Y, Liu R, Williams JM, Sarkis A, Lazar J, Jacob HJ, Roman RJ. Genetic basis of the impaired renal myogenic response in FHH rats. Am J Physiol Renal Physiol 304: F565–F577, 2013. 147. Brenner BM, Lawler EV, Mackenzie HS. The hyperfiltration theory: a paradigm shift in nephrology. Kidney Int 49: 1774 –1777, 1996. 169. Burke M, Pabbidi MR, Farley J, Roman RJ. Molecular mechanisms of renal blood flow autoregulation. Curr Vasc Pharmacol 12: 845– 858, 2014. 148. Brenner BM, Troy JL, Daugharty TM. The dynamics of glomerular ultrafiltration in the rat. J Clin Invest 50: 1776 –1780, 1971. 170. Burke TJ, Duchin KL. Glomerular filtration during furosemide diuresis in the dog. Kidney Int 16: 672– 680, 1979. 149. Breyer MD, Bottinger E, Brosius FC, Coffman TM, Fogo A, Harris RC, Heilig CW, Sharma K. Diabetic nephropathy: of mice and men. Adv Chronic Kidney Dis 12: 128 –145, 2005. 171. Busse R, Fleming I. Vascular endothelium and blood flow. Handb Exp Pharmacol 176: 43–78, 2006. 150. Breyer MD, Breyer RM. Prostaglandin E receptors and the kidney. Am J Physiol Renal Physiol 279: F12–F23, 2000. 151. Breyer MD, Zhang Y, Guan YF, Hao CM, Hebert RL, Breyer RM. Regulation of renal function by prostaglandin E receptors. Kidney Int Suppl 67: S88 –S94, 1998. 172. Bussemaker E, Wallner C, Fisslthaler B, Fleming I. The Na-K-ATPase is a target for an EDHF displaying characteristics similar to potassium ions in the porcine renal interlobar artery. Br J Pharmacol 137: 647– 654, 2002. 173. Byrom FB. Angiotensin and renal vascular damage. Br J Exp Pathol 45: 7–12, 1964. 152. Briet M, Schiffrin EL. Aldosterone: effects on the kidney and cardiovascular system. Nat Rev Nephrol 6: 261–273, 2010. 174. Cachofeiro V, Nasjletti A. Increased vascular responsiveness to bradykinin in kidneys of spontaneously hypertensive rats. Effect of N-nitro-L-arginine. Hypertension 18: 683– 688, 1991. 153. Broegger T, Jacobsen JC, Secher D, V, Boedtkjer DM, Kold-Petersen H, Pedersen FS, Aalkjaer C, Matchkov VV. Bestrophin is important for the rhythmic but not the tonic contraction in rat mesenteric small arteries. Cardiovasc Res 91: 685– 693, 2011. 175. Callera GE, Touyz RM, Teixeira SA, Muscara MN, Carvalho MH, Fortes ZB, Nigro D, Schiffrin EL, Tostes RC. ETA receptor blockade decreases vascular superoxide generation in DOCA-salt hypertension. Hypertension 42: 811– 817, 2003. 154. Brookes ZL, Kaufman S. Myogenic responses and compliance of mesenteric and splenic vasculature in the rat. Am J Physiol Regul Integr Comp Physiol 284: R1604 – R1610, 2003. 176. Campean V, Theilig F, Paliege A, Breyer M, Bachmann S. Key enzymes for renal prostaglandin synthesis: site-specific expression in rodent kidney (rat, mouse). Am J Physiol Renal Physiol 285: F19 –F32, 2003. 155. Brown R, Ollerstam A, Johansson B, Skott O, Gebre-Medhin S, Fredholm B, Persson AE. Abolished tubuloglomerular feedback and increased plasma renin in adenosine A1 receptor-deficient mice. Am J Physiol Regul Integr Comp Physiol 281: R1362– R1367, 2001. 177. Campese VM, Parise M, Karubian F, Bigazzi R. Abnormal renal hemodynamics in black salt-sensitive patients with hypertension. Hypertension 18: 805– 812, 1991. 156. Brown RD, Hilliard LM, Head GA, Jones ES, Widdop RE, Denton KM. Sex differences in the pressor and tubuloglomerular feedback response to angiotensin II. Hypertension 59: 129 –135, 2012. 157. Brown SA, Brown CA. Single-nephron adaptations to partial renal ablation in cats. Am J Physiol Regul Integr Comp Physiol 269: R1102–R1108, 1995. 178. Cancela JM, Gerasimenko OV, Gerasimenko JV, Tepikin AV, Petersen OH. Two different but converging messenger pathways to intracellular Ca2⫹ release: the roles of nicotinic acid adenine dinucleotide phosphate, cyclic ADP-ribose and inositol trisphosphate. EMBO J 19: 2549 –2557, 2000. 179. Carlson BE, Secomb TW. A theoretical model for the myogenic response based on the length-tension characteristics of vascular smooth muscle. Microcirculation 12: 327–338, 2005. Physiol Rev • VOL 95 • APRIL 2015 • www.prv.org 475 RENAL AUTOREGULATORY MECHANISMS 180. Carlstrom M, Brown RD, Sallstrom J, Larsson E, Zilmer M, Zabihi S, Eriksson UJ, Persson AE. SOD1-deficiency causes salt-sensitivity and aggravates hypertension in hydronephrosis. Am J Physiol Regul Integr Comp Physiol 297: R82–R92, 2009. 200. Casellas D, Moore LC. Autoregulation and tubuloglomerular feedback in juxtamedullary glomerular arterioles. Am J Physiol Renal Fluid Electrolyte Physiol 258: F660 – F669, 1990. 181. Carlstrom M, Brown RD, Yang T, Hezel M, Larsson E, Scheffer PG, Teerlink T, Lundberg JO, Persson AE. L-Arginine or tempol supplementation improves renal and cardiovascular function in rats with reduced renal mass and chronic high salt intake. Acta Physiol 207: 732–741, 2013. 201. Casellas D, Moore LC. Autoregulation of intravascular pressure in preglomerular juxtamedullary vessels. Am J Physiol Renal Fluid Electrolyte Physiol 264: F315–F321, 1993. 182. Carlstrom M, Lai EY, Ma Z, Patzak A, Brown RD, Persson AE. Role of NOX2 in the regulation of afferent arteriole responsiveness. Am J Physiol Regul Integr Comp Physiol 296: R72–R79, 2009. 183. Carlstrom M, Lai EY, Ma Z, Steege A, Patzak A, Eriksson UJ, Lundberg JO, Wilcox CS, Persson AE. Superoxide dismutase 1 limits renal microvascular remodeling and attenuates arteriole and blood pressure responses to angiotensin II via modulation of nitric oxide bioavailability. Hypertension 56: 907–913, 2010. 184. Carlstrom M, Persson AE. Important role of NAD(P)H oxidase 2 in the regulation of the tubuloglomerular feedback. Hypertension 53: 456 – 457, 2009. 185. Carlstrom M, Persson AE, Larsson E, Hezel M, Scheffer PG, Teerlink T, Weitzberg E, Lundberg JO. Dietary nitrate attenuates oxidative stress, prevents cardiac and renal injuries, and reduces blood pressure in salt-induced hypertension. Cardiovasc Res 89: 574 –585, 2011. 186. Carlstrom M, Sallstrom J, Skott O, Larsson E, Persson AE. Uninephrectomy in young age or chronic salt loading causes salt-sensitive hypertension in adult rats. Hypertension 49: 1342–1350, 2007. 187. Carlstrom M, Wilcox CS, Welch WJ. Adenosine A2 receptors modulate the tubuloglomerular feedback. Am J Physiol Renal Physiol 299: F412–F417, 2010. 188. Carlstrom M, Wilcox CS, Welch WJ. Adenosine A2A receptor activation attenuates tubuloglomerular feedback responses by stimulation of endothelial nitric oxide synthase. Am J Physiol Renal Physiol 300: F457–F464, 2011. 189. Carmines PK. The renal vascular response to diabetes. Curr Opin Nephrol Hypertens 19: 85–90, 2010. 190. Carmines PK, Bell PD, Roman RJ, Work J, Navar LG. Prostaglandins in the sodium excretory response to altered renal arterial pressure in dogs. Am J Physiol Renal Fluid Electrolyte Physiol 248: F8 –F14, 1985. 191. Carmines PK, Fowler BC, Bell PD. Segmentally distinct effects of depolarization on intracellular [Ca2⫹] in renal arterioles. Am J Physiol Renal Fluid Electrolyte Physiol 265: F677–F685, 1993. 192. Carmines PK, Inscho EW. Perfusate composition modulates in vitro renal microvascular pressure responsiveness in a segment-specific manner. Microvasc Res 43: 347– 351, 1992. 193. Carmines PK, Inscho EW, Gensure RC. Arterial pressure effects on preglomerular microvasculature of juxtamedullary nephrons. Am J Physiol Renal Fluid Electrolyte Physiol 258: F94 –F102, 1990. 194. Carmines PK, Mitchell KD, Navar LG. Effects of calcium antagonists on renal hemodynamics and glomerular function. Kidney Int 41: 43– 48, 1992. 202. Casellas D, Navar LG. In vitro perfusion of juxtamedullary nephrons in rats. Am J Physiol Renal Fluid Electrolyte Physiol 246: F349 –F358, 1984. 203. Castrop H. Mediators of tubuloglomerular feedback regulation of glomerular filtration: ATP and adenosine. Acta Physiol 189: 3–14, 2007. 204. Castrop H, Hocherl K, Kurtz A, Schweda F, Todorov V, Wagner C. Physiology of kidney renin. Physiol Rev 90: 607– 673, 2010. 205. Castrop H, Huang Y, Hashimoto S, Mizel D, Hansen P, Theilig F, Bachmann S, Deng C, Briggs J, Schnermann J. Impairment of tubuloglomerular feedback regulation of GFR in ecto-5’-nucleotidase/CD73-deficient mice. J Clin Invest 114: 634 – 642, 2004. 206. Cervenka L, Horácek V, Vanecková I, Hubácek JA, Oliverio MI, Coffman TM, Navar LG. Essential role of AT1A receptor in the development of 2K1C hypertension. Hypertension 40: 735–741, 2002. 207. Cervenka L, Kramer HJ, Malý J, Heller J. Role of nNOS in regulation of renal function in angiotensin II-induced hypertension. Hypertension 38: 280 –285, 2001. 208. Cervenka L, Vaneckova I, Huskova Z, Vanourkova Z, Erbanova M, Thumova M, Skaroupkova P, Opocensky M, Maly J, Chabova VC, Tesar V, Burgelova M, Viklicky O, Teplan V, Zelizko M, Kramer HJ, Navar LG. Pivotal role of angiotensin II receptor subtype 1A in the development of two-kidney, one-clip hypertension: study in angiotensin II receptor subtype 1A knockout mice. J Hypertens 26: 1379 –1389, 2008. 209. Cervenka L, Wang CT, Mitchell KD, Navar LG. Proximal tubular angiotensin II levels and renal functional responses to AT1 receptor blockade in nonclipped kidneys of Goldblatt hypertensive rats. Hypertension 33: 102–107, 1999. 210. Cervenka L, Wang CT, Navar LG. Effects of acute AT1 receptor blockade by candesartan on arterial pressure and renal function in rats. Am J Physiol Renal Physiol 274: F940 –F945, 1998. 211. Chabrashvili T, Kitiyakara C, Blau J, Karber A, Aslam S, Welch WJ, Wilcox CS. Effects of ANG II type 1 and 2 receptors on oxidative stress, renal NADPH oxidase and SOD expression. Am J Physiol Regul Integr Comp Physiol 285: R117–R124, 2003. 212. Chabrashvili T, Tojo A, Onozato M, Kitiyakara C, Quinn MT, Fujita T, Welch WJ, Wilcox CS. Expression and cellular localization of classic NADPH oxidase subunits in the spontaneously hypertensive rat kidney. Hypertension 39: 269 –274, 2002. 213. Champion D, Simatos D, Kalogianni EP, Cayot P, Le MM. Ascorbic acid oxidation in sucrose aqueous model systems at subzero temperatures. J Agric Food Chem 52: 3399 –3404, 2004. 214. Chan WL, Holstein-Rathlou NH, Yip KP. Integrin mobilizes intracellular Ca2⫹ in renal vascular smooth muscle cells. Am J Physiol Cell Physiol 280: C593–C603, 2001. 195. Carmines PK, Ohishi K, Ikenaga H. Functional impairment of renal afferent arteriolar voltage-gated calcium channels in rats with diabetes mellitus. J Clin Invest 98: 2564 – 2571, 1996. 215. Chaston DJ, Baillie BK, Grayson TH, Courjaret RJ, Heisler JM, Lau KA, Machaca K, Nicholson BJ, Ashton A, Matthaei KI, Hill CE. Polymorphism in endothelial connexin40 enhances sensitivity to intraluminal pressure and increases arterial stiffness. Arterioscler Thromb Vasc Biol 33: 962–970, 2013. 196. Carnevale D, Lembo G. PI3Kgamma in hypertension: a novel therapeutic target controlling vascular myogenic tone and target organ damage. Cardiovasc Res 95: 403– 408, 2012. 216. Chatziantoniou C, Arendshorst WJ. Angiotensin and thromboxane in genetically hypertensive rats: renal blood flow and receptor studies. Am J Physiol Renal Fluid Electrolyte Physiol 261: F238 –F247, 1991. 197. Carnevale D, Vecchione C, Mascio G, Esposito G, Cifelli G, Martinello K, Landolfi A, Selvetella G, Grieco P, Damato A, Franco E, Haase H, Maffei A, Ciraolo E, Fucile S, Frati G, Mazzoni O, Hirsch E, Lembo G. PI3Kgamma inhibition reduces blood pressure by a vasorelaxant Akt/L-type calcium channel mechanism. Cardiovasc Res 93: 200 –209, 2012. 217. Chatziantoniou C, Arendshorst WJ. Impaired ability of prostaglandins to buffer renal vasoconstriction in genetically hypertensive rats. Am J Physiol Renal Fluid Electrolyte Physiol 263: F573–F580, 1992. 198. Casellas D, Bouriquet N, Herizi A. Bosentan prevents preglomerular alterations during angiotensin II hypertension. Hypertension 30: 1613–1620, 1997. 199. Casellas D, Carmines PK, Navar LG. Microvascular reactivity of in vitro blood perfused juxtamedullary nephrons from rats. Kidney Int 28: 752–759, 1985. 476 218. Chatziantoniou C, Arendshorst WJ. Prostaglandin interactions with angiotensin, norepinephrine, and thromboxane in rat renal vasculature. Am J Physiol Renal Fluid Electrolyte Physiol 262: F68 –F76, 1992. 219. Chatziantoniou C, Arendshorst WJ. Renal vascular reactivity to vasodilator prostaglandins in genetically hypertensive rats. Am J Physiol Renal Fluid Electrolyte Physiol 262: F124 –F130, 1992. Physiol Rev • VOL 95 • APRIL 2015 • www.prv.org CARLSTRÖM ET AL. 220. Chatziantoniou C, Arendshorst WJ. Angiotensin receptor sites in renal vasculature of rats developing genetic hypertension. Am J Physiol Renal Fluid Electrolyte Physiol 265: F853–F862, 1993. 240. Chilton L, Smirnov SV, Loutzenhiser K, Wang X, Loutzenhiser RD. Segment-specific differences in the inward rectifier K⫹ current along the renal interlobular artery. Cardiovasc Res 92: 169 –177, 2011. 221. Chatziantoniou C, Arendshorst WJ. Vascular interactions of prostaglandins with thromboxane in kidneys of rats developing hypertension. Am J Physiol Renal Fluid Electrolyte Physiol 265: F250 –F256, 1993. 241. Chin SY, Pandey KN, Shi SJ, Kobori H, Moreno C, Navar LG. Increased activity and expression of Ca2⫹-dependent NOS in renal cortex of ANG II-infused hypertensive rats. Am J Physiol Renal Physiol 277: F797–F804, 1999. 222. Chatziantoniou C, Daniels FH, Arendshorst WJ. Exaggerated renal vascular reactivity to angiotensin and thromboxane in young genetically hypertensive rats. Am J Physiol Renal Fluid Electrolyte Physiol 259: F372–F382, 1990. 242. Chin SY, Wang CT, Majid DSA, Navar LG. Renoprotective effects of nitric oxide in angiotensin II-induced hypertension in the rat. Am J Physiol Renal Physiol 274: F876 – F882, 1998. 223. Chatziantoniou C, Ruan X, Arendshorst WJ. Impaired ability of prostaglandins to buffer renal vasoconstriction in genetically hypertensive rats. Am J Physiol Renal Fluid Electrolyte Physiol 263: F573–F580, 1992. 243. Chlopicki S, Nilsson H, Mulvany MJ. Initial and sustained phases of myogenic response of rat mesenteric small arteries. Am J Physiol Heart Circ Physiol 281: H2176 – H2183, 2001. 224. Chatziantoniou C, Ruan X, Arendshorst WJ. Interactions of cAMP-mediated vasodilators with angiotensin II in rat kidney during hypertension. Am J Physiol Renal Fluid Electrolyte Physiol 265: F845–F852, 1993. 244. Chon KH, Chen YM, Holstein-Rathlou NH, Marmarelis VZ. Nonlinear system analysis of renal autoregulation in normotensive and hypertensive rats. IEEE Trans Biomed Eng 45: 342–353, 1998. 225. Chatziantoniou C, Ruan X, Arendshorst WJ. Defective G protein activation of the cAMP pathway in rat kidney during genetic hypertension. Proc Natl Acad Sci USA 92: 2924 –2928, 1995. 245. Chon KH, Chen YM, Marmarelis VZ, Marsh DJ, Holstein-Rathlou NH. Detection of interactions between myogenic and TGF mechanisms using nonlinear analysis. Am J Physiol Renal Fluid Electrolyte Physiol 267: F160 –F173, 1994. 226. Chen CC, Geurts AM, Jacob HJ, Fan F, Roman RJ. Heterozygous knockout of transforming growth factor-beta1 protects Dahl S rats against high salt-induced renal injury. Physiol Genomics 45: 110 –118, 2013. 246. Chon KH, Raghavan R, Chen YM, Marsh DJ, Yip KP. Interactions of TGF-dependent and myogenic oscillations in tubular pressure. Am J Physiol Renal Physiol 288: F298 – F307, 2005. 227. Chen DD, Dong YG, Yuan H, Chen AF. Endothelin 1 activation of endothelin A receptor/NADPH oxidase pathway and diminished antioxidants critically contribute to endothelial progenitor cell reduction and dysfunction in salt-sensitive hypertension. Hypertension 59: 1037–1043, 2012. 247. Chon KH, Zhong Y, Moore LC, Holstein-Rathlou NH, Cupples WA. Analysis of nonstationarity in renal autoregulation mechanisms using time-varying transfer and coherence functions. Am J Physiol Regul Integr Comp Physiol 295: R821–R828, 2008. 228. Chen J, Sgouralis I, Moore LC, Layton HE, Layton AT. A mathematical model of the myogenic response to systolic pressure in the afferent arteriole. Am J Physiol Renal Physiol 300: F669 –F681, 2011. 229. Chen PY, St. John PL, Kirk KA, Abrahamson DR, Sanders PW. Hypertensive nephrosclerosis in the Dahl/Rapp rat Initial sites of injury and effect of dietary L-arginine supplementation. Lab Invest 68: 174 –184, 1993. 230. Chen YM, Holstein-Rathlou NH. Differences in dynamic autoregulation of renal blood flow between SHR and WKY rats. Am J Physiol Renal Fluid Electrolyte Physiol 264: F166 –F174, 1993. 231. Chen YM, Yip KP, Marsh DJ, Holstein-Rathlou NH. Magnitude of TGF-initiated nephron-nephron interactions is increased in SHR. Am J Physiol Renal Fluid Electrolyte Physiol 269: F198 –F204, 1995. 232. Cheng HF, Zhang MZ, Harris RC. Nitric oxide stimulates cyclooxygenase-2 in cultured cTAL cells through a p38-dependent pathway. Am J Physiol Renal Physiol 290: F1391–F1397, 2006. 2⫹ 233. Cheng X, Jaggar JH. Genetic ablation of caveolin-1 modifies Ca spark coupling in murine arterial smooth muscle cells. Am J Physiol Heart Circ Physiol 290: H2309 – H2319, 2006. 234. Cheng ZJ, Vaskonen T, Tikkanen I, Nurminen K, Ruskoaho H, Vapaatalo H, Muller D, Park JK, Luft FC, Mervaala EM. Endothelial dysfunction and salt-sensitive hypertension in spontaneously diabetic Goto-Kakizaki rats. Hypertension 37: 433– 439, 2001. 235. Chevalier RL, Carey RM, Kaiser DL. Endogenous prostaglandins modulate autoregulation of renal blood flow in young rats. Am J Physiol Renal Fluid Electrolyte Physiol 253: F66 –F75, 1987. 236. Chevalier RL, Kaiser DL. Autoregulation of renal blood flow in the rat: effects of growth and uninephrectomy. Am J Physiol Renal Fluid Electrolyte Physiol 244: F483– F487, 1983. 237. Chevalier RL, Kaiser DL. Effects of acute uninephrectomy and age on renal blood flow autoregulation in the rat. Am J Physiol Renal Fluid Electrolyte Physiol 249: F672– F679, 1985. 248. Chou CL, Marsh DJ. Role of proximal convoluted tubule in pressure diuresis in the rat. Am J Physiol Renal Fluid Electrolyte Physiol 251: F283–F289, 1986. 249. Chou CL, Marsh DJ. Time course of proximal tubule response to acute arterial hypertension in the rat. Am J Physiol Renal Fluid Electrolyte Physiol 254: F601–F607, 1988. 250. Christensen HR, Nielsen H, Christensen KL, Baandrup U, Jespersen LT, Mulvany MJ. Long-term hypotensive effects of an angiotensin converting enzyme inhibitor in spontaneously hypertensive rats: is there a role for vascular structure? J Hypertens 6: S27–S31, 1988. 251. Christensen PK, Akram K, Konig KB, Parving HH. Autoregulation of glomerular filtration rate in patients with type 2 diabetes during isradipine therapy. Diabetes Care 26: 156 –162, 2003. 252. Christensen PK, Hansen HP, Parving HH. Impaired autoregulation of GFR in hypertensive non-insulin dependent diabetic patients. Kidney Int 52: 1369 –1374, 1997. 253. Christensen PK, Hommel EE, Clausen P, Feldt-Rasmussen B, Parving HH. Impaired autoregulation of the glomerular filtration rate in patients with nondiabetic nephropathies. Kidney Int 56: 1517–1523, 1999. 254. Christensen PK, Lund S, Parving HH. Autoregulated glomerular filtration rate during candesartan treatment in hypertensive type 2 diabetic patients. Kidney Int 60: 1435– 1442, 2001. 255. Christensen PK, Lund S, Parving HH. The impact of glycaemic control on autoregulation of glomerular filtration rate in patients with non-insulin dependent diabetes. Scand J Clin Lab Invest 61: 43–50, 2001. 256. Chrysant SG, Walsh GM, Frohlich ED, Townsend SM. Hemodynamic changes induced by prolonged NaCl and DOCA administration in spontaneously hypertensive rats. Angiology 29: 303–309, 1978. 257. Chung WS, Farley JM, Drummond HA. ASIC-like currents in freshly isolated cerebral artery smooth muscle cells are inhibited by endogenous oxidase activity. Cell Physiol Biochem 27: 129 –138, 2011. 258. Chung WS, Weissman JL, Farley J, Drummond HA. betaENaC is required for whole cell mechanically gated currents in renal vascular smooth muscle cells. Am J Physiol Renal Physiol 304: F1428 –F1437, 2013. 238. Chilton L, Loutzenhiser K, Morales E, Breaks J, Kargacin GJ, Loutzenhiser RD. Inward rectifier K⫹ currents Kir2.1 expression in renal afferent and efferent arterioles. J Am Soc Nephrol 19: 69 –76, 2008. 259. Churchill PC, Churchill MC, Bidani AK. Kidney cross transplants in Dahl salt-sensitive and salt-resistant rats. Am J Physiol Heart Circ Physiol 262: H1809 –H1817, 1992. 239. Chilton L, Loutzenhiser RD. Functional evidence for an inward rectifier potassium current in rat renal afferent arterioles. Circ Res 88: 152–158, 2001. 260. Churchill PC, Churchill MC, Bidani AK, Griffin KA, Picken M, Pravenec M, Kren V, St Lezin E, Wang JM, Wang N, Kurtz TW. Genetic susceptibility to hypertension- Physiol Rev • VOL 95 • APRIL 2015 • www.prv.org 477 RENAL AUTOREGULATORY MECHANISMS induced renal damage in the rat. Evidence based on kidney-specific genome transfer. J Clin Invest 100: 1373–1382, 1997. 283. Cupples WA. Renal medullary blood flow: Its measurement and physiology. Can J Physiol Pharmacol 64: 873– 870, 1985. 261. Churchill PC, Churchill MC, Griffin KA, Picken M, Webb RC, Kurtz TW, Bidani AK. Increased genetic susceptibility to renal damage in the stroke-prone spontaneously hypertensive rat. Kidney Int 61: 1794 –1800, 2002. 284. Cupples WA. Angiotensin II conditions the slow component of autoregulation of renal blood flow. Am J Physiol Renal Fluid Electrolyte Physiol 264: F515–F522, 1993. 262. Cipolla MJ, Gokina NI, Osol G. Pressure-induced actin polymerization in vascular smooth muscle as a mechanism underlying myogenic behavior. FASEB J 16: 72–76, 2002. 263. Cipolla MJ, Porter JM, Osol G. High glucose concentrations dilate cerebral arteries and diminish myogenic tone through an endothelial mechanism. Stroke 28: 405– 410, 1997. 264. Cipolla MJ, Sweet JG, Chan SL, Tavares MJ, Gokina NI, Brayden JE. Increased pressure-induced tone in rat parenchymal arterioles vs. middle cerebral arteries: role of ion channels and calcium sensitivity. J Appl Physiol 117: 53–59, 2014. 265. Clausen G, Oien AH, Aukland K. Myogenic vasoconstriction in the rat kidney elicited by reducing perirenal pressure. Acta Physiol Scand 144: 277–290, 1992. 266. Coats P, Johnston F, MacDonald J, McMurray JJ, Hillier C. Signalling mechanisms underlying the myogenic response in human subcutaneous resistance arteries. Cardiovasc Res 49: 828 – 837, 2001. 267. Cohen HJ, Marsh DJ, Kayser B. Autoregulation in vasa recta of the rat kidney. Am J Physiol Renal Fluid Electrolyte Physiol 245: F32–F40, 1983. 268. Colantuoni A, Bertuglia S, Intaglietta M. Quantitation of rhythmic diameter changes in arterial microcirculation. Am J Physiol Heart Circ Physiol 246: H508 –H517, 1984. 269. Conger JD, Burke TJ. Effects of anesthetic agents on autoregulation of renal hemodynamics in the rat and dog. Am J Physiol 230: 652– 657, 1976. 270. Conrad KP, Brinck-Johnsen T, Gellai M, Valtin H. Renal autoregulation in chronically catheterized conscious rats. Am J Physiol Renal Fluid Electrolyte Physiol 247: F229 – F233, 1984. 271. Cooper ME. Interaction of metabolic and haemodynamic factors in mediating experimental diabetic nephropathy. Diabetologia 44: 1957–1972, 2001. 272. Corsetti JP, Sparks JD, Peterson RG, Smith RL, Sparks CE. Effect of dietary fat on the development of non-insulin dependent diabetes mellitus in obese Zucker diabetic fatty male and female rats. Atherosclerosis 148: 231–241, 2000. 273. Cowley AW Jr. Renal medullary oxidative stress, pressure-natriuresis, and hypertension. Hypertension 52: 777–786, 2008. 274. Cowley AW Jr, Mattson DL, Lu S, Roman RJ. The renal medulla and hypertension. Hypertension 25: 663– 673, 1995. 275. Cowley AW Jr, Mori T, Mattson D, Zou AP. Role of renal NO production in the regulation of medullary blood flow. Am J Physiol Regul Integr Comp Physiol 284: R1355–R1369, 2003. 276. Cowley AW Jr, Roman RJ. Abnormal pressure-diuresis-natriuresis response in spontaneously hypertensive rats. Am J Physiol Renal Fluid Electrolyte Physiol 248: F199 – F205, 1985. 277. Cowley AW Jr, Roman RJ, Fenoy FJ, Mattson DL. Effect of renal medullary circulation on arterial pressure. J Hypertens 10: S187–S193, 1992. 278. Cowley AW Jr, Roman RJ, Kaldunski ML, Dumas P, Dickhout JG, Greene AS, Jacob HJ. Brown Norway chromosome 13 confers protection from high salt to consomic Dahl S rat. Hypertension 37: 456 – 461, 2001. 279. Craven PA, Studer RK, Negrete H, DeRubertis FR. Protein kinase C in diabetic nephropathy. J Diabetes Complications 9: 241–245, 1995. 280. Creager MA, Luscher TF, Cosentino F, Beckman JA. Diabetes and vascular disease: pathophysiology, clinical consequences, and medical therapy: Part I. Circulation 108: 1527–1532, 2003. 281. Crowley SD, Coffman TM. In hypertension, the kidney rules. Curr Hypertens Rep 9: 148 –153, 2007. 282. Crowley SD, Zhang J, Herrera M, Griffiths RC, Ruiz P, Coffman TM. Role of AT1 receptor-mediated salt retention in angiotensin II-dependent hypertension. Am J Physiol Renal Physiol 301: F1124 –F1130, 2011. 478 285. Cupples WA. Interactions contributing to kidney blood flow autoregulation. Curr Opin Nephrol Hypertens 16: 39 – 45, 2007. 286. Cupples WA, Braam B. Assessment of renal autoregulation. Am J Physiol Renal Physiol 292: F1105–F1123, 2007. 287. Cupples WA, Loutzenhiser RD. Dynamic autoregulation in the in vitro perfused hydronephrotic rat kidney. Am J Physiol Renal Physiol 275: F126 –F130, 1998. 288. Cupples WA, Marsh DJ. Autoregulation of blood flow in renal medulla of the rat: no role for angiotensin II. Can J Physiol Pharmacol 66: 833– 836, 1988. 289. Cupples WA, Novak P, Novak V, Salevsky FC. Spontaneous blood pressure fluctuations and renal blood flow dynamics. Am J Physiol Renal Fluid Electrolyte Physiol 270: F82–F89, 1996. 290. D’Angelo G, Davis MJ, Meininger GA. Calcium and mechanotransduction of the myogenic response. Am J Physiol Heart Circ Physiol 273: H175–H182, 1997. 291. D’Angelo G, Mogford JE, Davis GE, Davis MJ, Meininger GA. Integrin-mediated reduction in vascular smooth muscle [Ca2⫹]i induced by RGD-containing peptide. Am J Physiol Heart Circ Physiol 272: H2065–H2070, 1997. 292. Dahl LK, Heine M, Thompson K. Genetic influence of the kidneys on blood pressure. Evidence from chronic renal homografts in rats with opposite predispositions to hypertension. Circ Res 34: 94 –101, 1974. 293. Dahly AJ, Hoagland KM, Flasch AK, Jha S, Ledbetter SR, Roman RJ. Antihypertensive effects of chronic anti-TGF-beta antibody therapy in Dahl S rats. Am J Physiol Regul Integr Comp Physiol 283: R757–R767, 2002. 294. Dalle-Donne I, Aldini G, Carini M, Colombo R, Rossi R, Milzani A. Protein carbonylation, cellular dysfunction, and disease progression. J Cell Mol Med 10: 389 – 406, 2006. 295. Dam VS, Boedtkjer DM, Nyvad J, Aalkjaer C, Matchkov V. TMEM16A knockdown abrogates two different Ca-activated Cl currents and contractility of smooth muscle in rat mesenteric small arteries. Pflügers Arch 466: 1391–1409, 2013. 296. Daniels FH, Arendshorst WJ. Tubuloglomerular feedback kinetics in spontaneously hypertensive and Wistar-Kyoto rats. Am J Physiol Renal Fluid Electrolyte Physiol 259: F529 –F534, 1990. 297. Daniels FH, Arendshorst WJ, Roberds RG. Tubuloglomerular feedback and autoregulation in spontaneously hypertensive rats. Am J Physiol Renal Fluid Electrolyte Physiol 258: F1479 –F1489, 1990. 298. Dautzenberg M, Just A. Temporal characteristics of nitric oxide-, prostaglandin-, and EDHF-mediated components of endothelium-dependent vasodilation in the kidney. Am J Physiol Regul Integr Comp Physiol 305: R987–R998, 2013. 299. Dautzenberg M, Keilhoff G, Just A. Modulation of the myogenic response in renal blood flow autoregulation by NO depends on endothelial nitric oxide synthase (eNOS), but not neuronal or inducible NOS. J Physiol 589: 4731– 4744, 2011. 300. Davis MJ. Perspective: physiological role(s) of the vascular myogenic response. Microcirculation 19: 99 –114, 2012. 301. Davis MJ, Hill MA. Signaling mechanisms underlying the vascular myogenic response. Physiol Rev 79: 387– 423, 1999. 302. Davis MJ, Hill MA, Kuo L. Local regulation of microvascular perfusion. In: Handbook of Physiology: Microcirculation. Boston: Academic, 2008. 303. Davis MJ, Sikes PJ. Myogenic responses of isolated arterioles: test for a rate-sensitive mechanism. Am J Physiol Heart Circ Physiol 259: H1890 –H1900, 1990. 304. Davis MJ, Wu X, Nurkiewicz TR, Kawasaki J, Davis GE, Hill MA, Meininger GA. Integrins and mechanotransduction of the vascular myogenic response. Am J Physiol Heart Circ Physiol 280: H1427–H1433, 2001. 305. Davis MJ, Wu X, Nurkiewicz TR, Kawasaki J, Gui P, Hill MA, Wilson E. Regulation of ion channels by integrins. Cell Biochem Biophys 36: 41– 66, 2002. Physiol Rev • VOL 95 • APRIL 2015 • www.prv.org CARLSTRÖM ET AL. 306. De Leon H, Gauquelin G, Thibault G, Garcia R. Characterization of receptors for the atrial natriuretic factor in rat renal microvessels. J Hypertens 11: 499 –508, 1993. 329. DiBona GF, Sawin LL. Effect of endogenous angiotensin II on the frequency response of the renal vasculature. Am J Physiol Renal Physiol 287: F1171–F1178, 2004. 307. De Micheli AG, Forster H, Duncan RC, Epstein M. A quantitative assessment of renal blood flow autoregulation in experimental diabetes. Nephron 68: 245–251, 1994. 330. Dietrich A, Kalwa H, Gudermann T. TRPC channels in vascular cell function. Thromb Haemost 103: 262–270, 2010. 308. De Nicola L, Blantz RC, Gabbai FB. Renal functional reserve in treated and untreated hypertensive rats. Kidney Int 40: 406 – 412, 1991. 331. Dietrich A, Kalwa H, Storch U, Mederos y Schnitzler M, Salanova B, Pinkenburg O, Dubrovska G, Essin K, Gollasch M, Birnbaumer L, Gudermann T Mederos y Schnitzler M, Salanova B, Pinkenburg O, Dubrovska G, Essin K, Gollasch M, Birnbaumer L, Gudermann T. Pressure-induced and store-operated cation influx in vascular smooth muscle cells is independent of TRPC1. Pflügers Arch 455: 465– 477, 2007. 309. De Nicola L, Blantz RC, Gabbai FB. Renal functional reserve in the early stage of experimental diabetes. Diabetes 41: 267–273, 1992. 310. De Nicola L, Keiser JA, Blantz RC, Gabbai FB. Angiotensin II and renal functional reserve in rats with Goldblatt hypertension. Hypertension 19: 790 –794, 1992. 311. De Richelieu LT, Sorensen CM, Holstein-Rathlou NH, Salomonsson M. NO-independent mechanism mediates tempol-induced renal vasodilation in SHR. Am J Physiol Renal Physiol 289: F1227–F1234, 2005. 312. De Souza Md, Bouskela E. Arteriolar diameter and spontaneous vasomotion: importance of potassium channels and nitric oxide. Microvasc Res 90: 121–127, 2013. 313. De Wit C, Boettcher M, Schmidt VJ. Signaling across myoendothelial gap junctions– fact or fiction? Cell Commun Adhes 15: 231–245, 2008. 314. De Wit C, Griffith TM. Connexins and gap junctions in the EDHF phenomenon and conducted vasomotor responses. Pflügers Arch 459: 897–914, 2010. 315. De Wit C, Wolfle SE. EDHF and gap junctions: important regulators of vascular tone within the microcirculation. Curr Pharm Biotechnol 8: 11–25, 2007. 316. De Zeeuw D, Remuzzi G, Parving HH, Keane W, Zhang Z, Shahinfar S, Snapinn S, Cooper ME, Mitch WE, Brenner BM. Proteinuria, a target for renoprotection in patients with type 2 diaetic nephropathy:lessons from RENAAL. Kidney Int 65: 2309 –2320, 2004. 317. DeClue JW, Guyton AC, Cowley AW Jr, Coleman TG, Norman RA Jr, McCaa RE. Subpressor angiotensin infusion, renal sodium handling, and salt-induced hypertension in the dog. Circ Res 43: 503–512, 1978. 318. Dedeoglu IO, Springate JE. Effect of nitric oxide inhibition on kidney function in experimental renovascular hypertension. Clin Exp Hypertens 23: 267–275, 2001. 319. Deen WM, Maddox DA, Robertson CR, Brenner BM. Dynamics of glomerular ultrafiltration in the rat. VII. Response to reduced renal mass. Am J Physiol 227: 556 – 562, 1974. 320. Deen WM, Troy JL, Robertson CR, Brenner BM. Dynamics of glomerular ultrafiltration in the rat. IV. Determination of the ultrafiltration coefficient. J Clin Invest 52: 1500 –1508, 1973. 321. Dehmel B, Mervaala E, Lippoldt A, Gross V, Bohlender J, Ganten D, Luft FC. Pressure-natriuresis and -diuresis in transgenic rats harboring both human renin and human angiotensinogen genes. J Am Soc Nephrol 9: 2212–2222, 1998. 322. Delaney PJ, Burnham MP, Heagerty AM, Izzard AS. Impaired myogenic properties of cerebral arteries from the Brown Norway rat. J Hypertens 30: 926 –931, 2012. 323. Denton KM, Anderson WP. Glomerular ultrafiltration in rabbits with superficial glomeruli. Pflügers Arch 419: 235–242, 1991. 324. Dessy C, Matsuda N, Hulvershorn J, Sougnez CL, Sellke FW, Morgan KG. Evidence for involvement of the PKC-alpha isoform in myogenic contractions of the coronary microcirculation. Am J Physiol Heart Circ Physiol 279: H916 –H923, 2000. 325. Di Nicolantonio R, Silvapulle MJ. Blood pressure, salt appetite and mortality of genetically hypertensive and normotensive rats maintained on high and low salt diets from weaning. Clin Exp Pharmacol Physiol 15: 741–751, 1988. 326. Di Nicolantonio R, Spargo S, Morgan TO. Prenatal high salt diet increases blood pressure and salt retention in the spontaneously hypertensive rat. Clin Exp Pharmacol Physiol 14: 233–235, 1987. 327. Dibb KM, Graham HK, Venetucci LA, Eisner DA, Trafford AW. Analysis of cellular calcium fluxes in cardiac muscle to understand calcium homeostasis in the heart. Cell Calcium 42: 503–512, 2007. 328. DiBona GF, Kopp UC. Neural control of renal function. Physiol Rev 77: 75–197, 1997. 332. Dietrich A, Mederos YS, Gollasch M, Gross V, Storch U, Dubrovska G, Obst M, Yildirim E, Salanova B, Kalwa H, Essin K, Pinkenburg O, Luft FC, Gudermann T, Birnbaumer L. Increased vascular smooth muscle contractility in TRPC6⫺/⫺ mice. Mol Cell Biol 25: 6980 – 6989, 2005. 333. Dilley JR, Arendshorst WJ. Enhanced tubuloglomerular feedback activity in rats developing spontaneous hypertension. Am J Physiol Renal Fluid Electrolyte Physiol 247: F672–F679, 1984. 334. Dilley JR, Corradi A, Arendshorst WJ. Glomerular ultrafiltration dynamics during increased renal venous pressure. Am J Physiol Renal Fluid Electrolyte Physiol 244: F650 –F658, 1983. 335. Dilley JR, Stier CT Jr, Arendshorst WJ. Abnormalities in glomerular function in rats developing spontaneous hypertension. Am J Physiol Renal Fluid Electrolyte Physiol 246: F12–F20, 1984. 336. Dirnagl U, Lindauer U, Villringer A. Nitric oxide synthase blockade enhances vasomotion in the cerebral microcirculation of anesthetized rats. Microvasc Res 45: 318 – 323, 1993. 337. Diz DI, Nasjletti A, Baer PG. Renal denervation at weaning retards development of hypertension in New Zealand genetically hypertensive rats. Hypertension 4: 361– 368, 1982. 338. Dopico AM, Kirber MT, Singer JJ, Walsh JV Jr. Membrane stretch directly activates large conductance Ca2⫹-activated K⫹ channels in mesenteric artery smooth muscle cells. Am J Hypertens 7: 82– 89, 1994. 339. Dorup J, Morsing P, Rasch R. Tubule-tubule and tubule-arteriole contacts in rat kidney distal nephrons. A morphologic study based on computer-assisted threedimensional reconstructions. Lab Invest 67: 761–769, 1992. 340. Dos Santos EA, Dahly-Vernon AJ, Hoagland KM, Roman RJ. Inhibition of the formation of EETs and 20-HETE with 1-aminobenzotriazole attenuates pressure natriuresis. Am J Physiol Regul Integr Comp Physiol 287: R58 –R68, 2004. 341. Drenckhahn D, Schnittler H, Nobiling R, Kriz W. Ultrastructural organization of contractile proteins in rat glomerular mesangial cells. Am J Pathol 137: 1343–1351, 1990. 342. Dresser TP, Lynch RE, Schneider EG, Knox FG. Effect of increases in blood pressure on pressure and reabsorption in the proximal tubule. Am J Physiol 220: 444 – 447, 1971. 343. Drummond HA. B-ENaC is a molecular component of a VSMC mechanotransducer that contributes to renal blood flow regulation, protection from renal injury, and hypertension. Front Physiol 3: 1–11, 2012. 344. Drummond HA, Gebremedhin D, Harder DR. Degenerin/epithelial Na⫹ channel proteins: components of a vascular mechanosensor. Hypertension 44: 643– 648, 2004. 345. Drummond HA, Grifoni SC, Abu-Zaid A, Gousset M, Chiposi R, Barnard JM, Murphey B, Stec DE. Renal inflammation and elevated blood pressure in a mouse model of reduced -ENaC. Am J Physiol Renal Physiol 301: F443–F449, 2011. 346. Drummond HA, Grifoni SC, Jernigan NL. A new trick for an old dogma: ENaC proteins as mechanotransducers in vascular smooth muscle. Physiology 23: 23–31, 2008. 347. Drumond MC, Deen WM. Analysis of pulsatile pressures and flows in glomerular filtration. Am J Physiol Renal Fluid Electrolyte Physiol 261: F409 –F419, 1991. 348. Dryer SE, Reiser J. TRPC6 channels and their binding partners in podocytes: role in glomerular filtration and pathophysiology. Am J Physiol Renal Physiol 299: F689 –F701, 2010. Physiol Rev • VOL 95 • APRIL 2015 • www.prv.org 479 RENAL AUTOREGULATORY MECHANISMS 349. Du W, Stiber JA, Rosenberg PB, Meissner G, Eu JP. Ryanodine receptors in muscarinic receptor-mediated bronchoconstriction. J Biol Chem 280: 26287–26294, 2005. 369. Earley S, Resta TC, Walker BR. Disruption of smooth muscle gap junctions attenuates myogenic vasoconstriction of mesenteric resistance arteries. Am J Physiol Heart Circ Physiol 287: H2677–H2686, 2004. 350. Dubroca C, Loyer X, Retailleau K, Loirand G, Pacaud P, Feron O, Balligand JL, Levy BI, Heymes C, Henrion D. RhoA activation and interaction with Caveolin-1 are critical for pressure-induced myogenic tone in rat mesenteric resistance arteries. Cardiovasc Res 73: 190 –197, 2007. 370. Earley S, Straub SV, Brayden JE. Protein kinase C regulates vascular myogenic tone through activation of TRPM4. Am J Physiol Heart Circ Physiol 292: H2613–H2622, 2007. 351. Duchin KL, Peterson LN, Burke TJ. Effect of furosemide on renal autoregulation. Kidney Int 12: 379 –386, 1977. 371. Earley S, Waldron BJ, Brayden JE. Critical role for transient receptor potential channel TRPM4 in myogenic constriction of cerebral arteries. Circ Res 95: 922–929, 2004. 352. Dukacz SA, Adams MA, Kline RL. Short- and long-term enalapril affect renal medullary hemodynamics in the spontaneously hypertensive rat. Am J Physiol Regul Integr Comp Physiol 276: R10 –R16, 1999. 372. Eckel J, Lavin PJ, Finch EA, Mukerji N, Burch J, Gbadegesin R, Wu G, Bowling B, Byrd A, Hall G, Sparks M, Zhang ZS, Homstad A, Barisoni L, Birbaumer L, Rosenberg P, Winn MP. TRPC6 enhances angiotensin II-induced albuminuria. J Am Soc Nephrol 22: 526 –535, 2011. 353. Dukacz SA, Feng MG, Yang LF, Lee RM, Kline RL. Abnormal renal medullary response to angiotensin II in SHR is corrected by long-term enalapril treatment. Am J Physiol Regul Integr Comp Physiol 280: R1076 –R1084, 2001. 354. Dukacz SA, Kline RL. Differing effects of enalapril and losartan on renal medullary blood flow and renal interstitial hydrostatic pressure in spontaneously hypertensive rats. J Hypertens 17: 1345–1352, 1999. 355. Duke LM, Paull JR, Widdop RE. Cardiovascular status following combined angiotensin-converting enzyme and AT1 receptor inhibition in conscious spontaneously hypertensive rats. Clin Exp Pharmacol Physiol 30: 317–323, 2003. 356. Durao V, Martins Prata M, Pires Goncalves LM. Modification of antihypertensive effect of b-adrenoceptor-blocking agents by inhibition of endogenous prostaglandin synthesis. Lancet 12: 1005–1007, 1977. 357. Dworkin LD, Benstein JA, Tolbert E, Feiner HD. Salt restriction inhibits renal growth and stabilizes injury in rats with established renal disease. J Am Soc Nephrol 7: 437– 442, 1996. 358. Dworkin LD, Feiner HD. Glomerular injury in uninephrectomized spontaneously hypertensive rats. A consequence of glomerular capillary hypertension. J Clin Invest 77: 797– 809, 1986. 359. Dworkin LD, Feiner HD, Parker M, Randazzo J. Calcium carbonate exacerbates glomerular capillary hypertension and injury in rats with desoxycorticosterone-salt hypertension. Am J Hypertens 3: 444 – 450, 1990. 360. Dworkin LD, Feiner HD, Parker M, Tolbert E. Effects of nifedipine and enalapril on glomerular structure and function in uninephrectomized SHR. Kidney Int 39: 1112– 1117, 1991. 361. Dworkin LD, Feiner HD, Randazzo J. Glomerular hypertension and injury in desoxycorticosterone-salt rats on antihypertensive therapy. Kidney Int 31: 718 –724, 1987. 362. Dworkin LD, Grosser M, Feiner HD, Ullian M, Parker M. Renal vascular effects of antihypertensive therapy in uninephrectomized SHR. Kidney Int 35: 790 –798, 1989. 363. Dworkin LD, Hostetter TH, Rennke HG, Brenner BM. Hemodynamic basis for glomerular injury in rats with desoxycorticosterone-salt hypertension. J Clin Invest 73: 1448 –1461, 1984. 364. Dworkin LD, Levin RI, Benstein JA, Parker M, Ullian ME, Kim Y, Feiner HD. Effects of nifedipine and enalapril on glomerular injury in rats with deoxycorticosterone-salt hypertension. Am J Physiol Renal Fluid Electrolyte Physiol 259: F598 –F604, 1990. 365. Dworkin LD, Tolbert E, Recht PA, Hersch JC, Feiner H, Levin RI. Effects of amlodipine on glomerular filtration, growth, and injury in experimental hypertension. Hypertension 27: 245–250, 1996. 366. Earley S, Brayden JE. Transient receptor potential channels and vascular function. Clin Sci 119: 19 –36, 2010. 367. Earley S, Pauyo T, Drapp R, Tavares MJ, Liedtke W, Brayden JE. TRPV4-dependent dilation of peripheral resistance arteries influences arterial pressure. Am J Physiol Heart Circ Physiol 297: H1096 –H1102, 2009. 368. Earley S, Reading S, Brayden JE. Functional significance of transient receptor potential channels in vascular function. In: TRP Ion Channel Function in Sensory Transduction and Cellular Signaling Cascades., edited by Liedtke WB, Heller S. Boca Raton, FL: CRC, 2007. 480 373. Edgley AJ, Kett MM, Anderson WP. Evidence for renal vascular remodeling in angiotensin II-induced hypertension. J Hypertens 21: 1401–1406, 2003. 374. Edwards A, Layton AT. Calcium dynamics underlying the afferent arteriole myogenic response. Am J Physiol Renal Physiol 306: F34 –F48, 2014. 375. Edwards RM. Segmental effects of norepinephrine and angiotensin II on isolated renal microvessels. Am J Physiol Renal Fluid Electrolyte Physiol 244: F526 –F534, 1983. 376. Ehmke H, Persson PB, Seyfarth M, Kirchheim HR. Neurogenic control of pressure natriuresis in conscious dogs. Am J Physiol Renal Fluid Electrolyte Physiol 259: F466 – F473, 1990. 377. Ekelund U, Bjornberg J, Grande PO, Albert U, Mellander S. Myogenic vascular regulation in skeletal muscle in vivo is not dependent of endothelium-derived nitric oxide. Acta Physiol Scand 144: 199 –207, 1992. 378. Elger M, Drenckhahn D, Nobiling R, Mundel P, Kriz W. Cultured rat mesangial cells contain smooth muscle alpha-actin not found in vivo. Am J Pathol 142: 497–509, 1993. 379. Eliahou HE, Cohen D, Hellberg B, Ben-David A, Herzog D, Shechter P, Kapuler S, Kogan N. Effect of the calcium channel blocker nisoldipine on the progression of chronic renal failure in man. Am J Nephrol 8: 285–290, 1988. 380. Enouri S, Monteith G, Johnson R. Characteristics of myogenic reactivity in isolated rat mesenteric veins. Am J Physiol Regul Integr Comp Physiol 300: R470 –R478, 2011. 381. Eppel GA, Bergstrom G, Anderson WP, Evans RG. Autoregulation of renal medullary blood flow in rabbits. Am J Physiol Regul Integr Comp Physiol 284: R233–R244, 2003. 382. Erdei N, Bagi Z, Edes I, Kaley G, Koller A. H2O2 increases production of constrictor prostaglandins in smooth muscle leading to enhanced arteriolar tone in Type 2 diabetic mice. Am J Physiol Heart Circ Physiol 292: H649 –H656, 2007. 383. Eskildsen-Helmond YE, Mulvany MJ. Pressure-induced activation of extracellular signal-regulated kinase 1/2 in small arteries. Hypertension 41: 891– 897, 2003. 384. Eu JP, Xu L, Stamler JS, Meissner G. Regulation of ryanodine receptors by reactive nitrogen species. Biochem Pharmacol 57: 1079 –1084, 1999. 385. Evans RG, Szenasi G, Anderson WP. Effects of NG-nitro-L-arginine on pressure natriuresis in anaesthetized rabbits. Clin Exp Pharmacol Physiol 22: 94 –101, 1995. 386. Evenwel RT, Kasbergen CM, Struyker-Boudier HA. Central and regional hemodynamics and plasma volume distribution during the development of spontaneous hypertension in rats. Clin Exp Hypertens A 5: 1511–1536, 1983. 387. Facemire CS, Mohler PJ, Arendshorst WJ. Expression and relative abundance of short transient receptor potential channels in the rat renal microcirculation. Am J Physiol Renal Physiol 286: F546 –F551, 2004. 388. Fahr T. Handbuch der speziellen pathologischen Anatomie und Histologie. Berlin: Springer, 1934. 389. Falcone JC, Granger HJ, Meininger GA. Enhanced myogenic activation in skeletal muscle arterioles from spontaneously hypertensive rats. Am J Physiol Heart Circ Physiol 265: H1847–H1855, 1993. 390. Fallet RW, Bast JP, Fujiwara K, Ishii N, Sansom SC, Carmines PK. Influence of Ca2⫹-activated K⫹ channels on rat renal arteriolar responses to depolarizing agonists. Am J Physiol Renal Physiol 280: F583–F591, 2001. Physiol Rev • VOL 95 • APRIL 2015 • www.prv.org CARLSTRÖM ET AL. 391. Farrugia E, Lockhart JC, Larson TS. Relation between vasa recta blood flow and renal interstitial hydrostatic pressure during pressure natriuresis. Circ Res 71: 1153–1158, 1992. 413. Fenoy FJ, Milicic I, Mistry M, Mecca TE, Roman RJ. Effect of clentiazem on arterial pressure and renal function in normotensive and hypertensive rats. J Pharmacol Exp Ther 261: 470 – 475, 1992. 392. Faulhaber-Walter R, Chen L, Oppermann M, Kim SM, Huang Y, Hiramatsu N, Mizel D, Kajiyama H, Zerfas P, Briggs JP, Kopp JB, Schnermann J. Lack of A1 adenosine receptors augments diabetic hyperfiltration and glomerular injury. J Am Soc Nephrol 19: 722–730, 2008. 414. Fenoy FJ, Milicic I, Smith RD, Wong PC, Timmermans PB, Roman RJ. Effects of DuP 753 on renal function of normotensive and spontaneously hypertensive rats. Am J Hypertens 4: 321S–326S, 1991. 393. Feld LG, Cachero S, Van Liew JB, Zamlauski-Tucker M, Noble B. Enalapril and renal injury in spontaneously hypertensive rats. Hypertension 16: 544 –554, 1990. 394. Feld LG, Zamlauski-Tucker MJ, Springate J, Van Liew JB. Single nephron hemodynamics in spontaneously hypertensive rats. Proc Soc Exp Biol Med 209: 185–189, 1995. 395. Feldberg R, Colding-Jorgensen M, Holstein-Rathlou NH. Analysis of interaction between TGF and the myogenic response in renal blood flow autoregulation. Am J Physiol Renal Fluid Electrolyte Physiol 269: F581–F593, 1995. 396. Fellner RC, Cook AK, O’Connor PM, Zhang S, Pollock DM, Inscho EW. High salt diet blunts renal autoregulation by a reactive oxygen species-dependent mechanism. Am J Physiol Renal Physiol 307: F33–F40, 2014. 397. Fellner SK, Arendshorst WJ. Capacitative calcium entry in smooth muscle cells from preglomerular vessels. Am J Physiol Renal Physiol 277: F533–F542, 1999. 398. Fellner SK, Arendshorst WJ. Ryanodine receptor and capacitative Ca2⫹ entry in fresh preglomerular vascular smooth muscle cells. Kidney Int 58: 1686 –1694, 2000. 399. Fellner SK, Arendshorst WJ. Angiotensin II, reactive oxygen species, and Ca2⫹ signaling in afferent arterioles. Am J Physiol Renal Physiol 289: F1012–F1019, 2005. 400. Fellner SK, Arendshorst WJ. Endothelin-A and -B receptors, superoxide, and Ca2⫹ signaling in afferent arterioles. Am J Physiol Renal Physiol 292: F175–F184, 2007. 401. Fellner SK, Arendshorst WJ. Voltage-gated Ca2⫹ entry and ryanodine receptor Ca2⫹-induced Ca2⫹ release in preglomerular arterioles. Am J Physiol Renal Physiol 292: F1568 –F1572, 2007. 402. Fellner SK, Arendshorst WJ. Angiotensin II-stimulated Ca2⫹ entry mechanisms in afferent arterioles: role of transient receptor potential canonical channels and reverse Na⫹/Ca2⫹ exchange. Am J Physiol Renal Physiol 294: F212–F219, 2008. 403. Fellner SK, Arendshorst WJ. Complex interactions of NO/cGMP/PKG systems on Ca2⫹ signaling in afferent arteriolar vascular smooth muscle. Am J Physiol Heart Circ Physiol 298: H144 –H151, 2010. 404. Feng JJ, Arendshorst WJ. Enhanced renal vasoconstriction induced by vasopressin in SHR is mediated by V1 receptors. Am J Physiol Renal Fluid Electrolyte Physiol 271: F304 –F313, 1996. 405. Feng L, Siu K, Moore LC, Marsh DJ, Chon KH. A robust method for detection of linear and nonlinear interactions: application to renal blood flow dynamics. Ann Biomed Eng 34: 339 –353, 2006. 406. Feng MG, Dukacz SAW, Kline RL. Selective effect of tempol on renal medullary hemodynamics in spontaneously hypertensive rats. Am J Physiol Regul Integr Comp Physiol 281: R1420 –R1425, 2001. 407. Feng MG, Li M, Navar LG. T-type calcium channels in the regulation of afferent and efferent arterioles in rats. Am J Physiol Renal Physiol 286: F331–F337, 2004. 2⫹ 408. Feng MG, Navar LG. Nitric oxide synthase inhibition activates L- and T-type Ca channels in afferent and efferent arterioles. Am J Physiol Renal Physiol 290: F873– F879, 2006. 409. Feng MG, Navar LG. Adenosine A2 receptor activation attenuates afferent arteriolar autoregulation during adenosine receptor saturation in rats. Hypertension 50: 744 – 749, 2007. 410. Fenger-Gron J, Mulvany MJ, Christensen KL. Mesenteric blood pressure profile of conscious, freely moving rats. J Physiol 488: 753–760, 1995. 411. Fenoy FJ, Ferrer P, Carbonell L, Garcia-Salom M. Role of nitric oxide on papillary blood flow and pressure natriuresis. Hypertension 25: 408 – 414, 1995. 412. Fenoy FJ, Kauker ML, Milicic I, Roman RJ. Normalization of pressure-natriuresis by nisoldipine in spontaneously hypertensive rats. Hypertension 19: 49 –55, 1992. 415. Fenoy FJ, Roman RJ. Effect of volume expansion on papillary blood flow and sodium excretion. Am J Physiol Renal Fluid Electrolyte Physiol 260: F813–F822, 1991. 416. Fenoy FJ, St Lezin E, Kurtz TW, Roman RJ. Genetic heterogeneity and differences in glomerular hemodynamics between inbred colonies of Munich-Wistar rats. J Am Soc Nephrol 3: 66 –72, 1992. 417. Ferrone RA, Antonaccio MJ. Prevention of the development of spontaneous hypertension in rats by captopril (SQ 14,225). Eur J Pharmacol 60: 131–137, 1979. 418. Ferrone RA, Heran CL, Antonaccio MJ. Comparison of the acute and chronic hemodynamic effects of captopril and guanethidine in spontaneously hypertensive rats. Clin Exp Hypertens 2: 247–272, 1980. 419. Figueroa XF, Duling BR. Dissection of two Cx37-independent conducted vasodilator mechanisms by deletion of Cx40: electrotonic versus regenerative conduction. Am J Physiol Heart Circ Physiol 295: H2001–H2007, 2008. 420. Finco DR, Cooper TL. Soy protein increases glomerular filtration rate in dogs with normal or reduced renal function. J Nutr 130: 745–748, 2000. 421. Finke R, Gross R, Hackenthal E, Huber J, Kirchheim HR. Threshold pressure for the pressure-dependent renin release in the autoregulating kidney of conscious dogs. Pflügers Arch 399: 102–110, 1983. 422. Finn WF, Arendshorst WJ. Effect of prostaglandin synthetase inhibitors on renal blood flow in the rat. Am J Physiol 231: 1541–1545, 1976. 423. Flemming B, Arenz N, Seeliger E, Wronski T, Steer K, Persson PB. Time-dependent autoregulation of renal blood flow in conscious rats. J Am Soc Nephrol 12: 2253–2262, 2001. 424. Fliser D, Schroter M, Neubeck M, Ritz E. Coadministration of thiazides increases the efficacy of loop diuretics even in patients with advanced renal failure. Kidney Int 46: 482– 488, 1994. 425. Folkow B, Grimby G, Thulesius O. Adaptive structural changes of the vascular walls in hypertension and their relation to the control of the peripheral resistance. Acta Physiol Scand 44: 255–272, 1958. 426. Forster HG, Ter Wee PM, Hohman TC, Epstein M. Impairment of afferent arteriolar myogenic responsiveness in the galactose-fed rat is prevented by tolrestat. Diabetologia 39: 907–914, 1996. 427. Forster HG, Ter Wee PM, Takenaka T, Hohman TC, Epstein M. Impairment of afferent arteriolar myogenic responsiveness in the galactose-fed rat. Proc Soc Exp Biol Med 206: 365–374, 1994. 428. Forster RP, Maes JP. Effect of experimental neurogenic hypertension on renal blood flow and glomerular filtration rates in intact denervated kidneys of unanesthetized rabbits with adrenal glands demedullated. Am J Physiol 150: 534 –540, 1947. 429. Fortepiani LA, Rodrigo E, Ortiz MC, Cachofeiro V, Atucha NM, Ruilope LM, Lahera V, Garcia-Estan J. Pressure natriuresis in nitric oxide-deficient hypertensive rats: effect of antihypertensive treatments. J Am Soc Nephrol 10: 21–27, 1999. 430. Fowler BC, Carmines PK, Nelson LD, Bell PD. Characterization of sodium-calcium exchange in rabbit renal arterioles. Kidney Int 50: 1856 –1862, 1996. 431. Fowler BC, Chang YS, Laamarti A, Higdon M, LaPointe JY, Bell PD. Evidence for apical sodium proton exchange in macula densa cells. Kidney Int 47: 746 –751, 1995. 432. Fozard JR, Part ML. Haemodynamic responses to NG-monomethyl-L-arginine in spontaneously hypertensive and normotensive Wistar-Kyoto rats. Br J Pharmacol 102: 823– 826, 1991. 433. Franchini KG, Mattson DL, Cowley AW Jr. Vasopressin modulation of medullary blood flow and pressure-natriuresis-diuresis in the decerebrated rat. Am J Physiol Regul Integr Comp Physiol 272: R1472–R1479, 1997. Physiol Rev • VOL 95 • APRIL 2015 • www.prv.org 481 RENAL AUTOREGULATORY MECHANISMS 434. Francis SH, Busch JL, Corbin JD, Sibley D. cGMP-dependent protein kinases and cGMP phosphodiesterases in nitric oxide and cGMP action. Pharmacol Rev 62: 525– 563, 2010. 453. Gandley RE, Conrad KP, McLaughlin MK. Endothelin and nitric oxide mediate reduced myogenic reactivity of small renal arteries from pregnant rats. Am J Physiol Regul Integr Comp Physiol 280: R1–R7, 2001. 435. Francischetti A, Ono H, Frohlich ED. Renoprotective effects of felodipine and/or enalapril in spontaneously hypertensive rats with and without L-NAME. Hypertension 31: 795– 801, 1998. 454. Gannon KP, VanLandingham LG, Jernigan NL, Grifoni SC, Hamilton G, Drummond HA. Impaired pressure-induced constriction in mouse middle cerebral arteries of ASIC2 knockout mice. Am J Physiol Heart Circ Physiol 294: H1793–H1803, 2008. 436. Franco M, Bautista R, Tapia E, Soto V, Santamaria J, Osorio H, Pacheco U, SanchezLozada LG, Kobori H, Navar LG. Contribution of renal purinergic receptors to renal vasoconstriction in angiotensin II-induced hypertensive rats. Am J Physiol Renal Physiol 300: F1301–F1309, 2011. 455. Gao X, Patzak A, Sendeski M, Scheffer PG, Teerlink T, Sallstrom J, Fredholm BB, Persson AE, Carlstrom M. Adenosine A1 receptor deficiency diminishes afferent arteriolar and blood pressure responses during nitric oxide inhibition and angiotensin II treatment. Am J Physiol Regul Integr Comp Physiol 301: R16669 –R16681, 2011. 437. Franco M, Bell PD, Navar LG. Evaluation of prostaglandins as mediators of tubuloglomerular feedback. Am J Physiol Renal Fluid Electrolyte Physiol 254: F642–F649, 1988. 456. Gao YJ, Nishimura Y, Suzuki A. Dopamine-induced relaxation in isolated intrarenal arteries from adult stroke-prone spontaneously hypertensive rats. Clin Exp Pharmacol Physiol 22 Suppl 1: S96 –S98, 1995. 438. Franco M, Bell PD, Navar LG. Effect of adenosine A1 analogue on tubuloglomerular feedback mechanism. Am J Physiol Renal Fluid Electrolyte Physiol 257: F231–F236, 1989. 457. Gao YJ, Nishimura Y, Suzuki A, Higashino H. Reactivity of intrarenal arteries to vasoconstrictor and vasorelaxant polypeptides in adult stroke-prone spontaneously hypertensive rats. J Smooth Muscle Res 35: 125–134, 1999. 439. Franco M, Martinez F, Rodriguez-Iturbe B, Johnson RJ, Santamaria J, Montoya A, Nepomuceno T, Bautista R, Tapia E, Herrera-Acosta J. Angiotensin II, interstitial inflammation, and the pathogenesis of salt-sensitive hypertension. Am J Physiol Renal Physiol 291: F1281–F1287, 2006. 458. Garcia SR, Izzard AS, Heagerty AM, Bund SJ. Myogenic tone in coronary arteries from spontaneously hypertensive rats. J Vasc Res 34: 109 –116, 1997. 440. Franco M, Perez-Mendez O, Martinez F. Interaction of intrarenal adenosine and angiotensin II in kidney vascular resistance. Curr Opin Nephrol Hypertens 18: 63– 67, 2009. 441. Francois H, Athirakul K, Mao L, Rockman H, Coffman TM. Role for thromboxane receptors in angiotensin-II-induced hypertension. Hypertension 43: 364 –369, 2004. 442. Freedman BI, Hicks PJ, Bostrom MA, Cunningham ME, Liu Y, Divers J, Kopp JB, Winkler CA, Nelson GW, Langefeld CD, Bowden DW. Polymorphisms in the nonmuscle myosin heavy chain 9 gene (MYH9) are strongly associated with end-stage renal disease historically attributed to hypertension in African Americans. Kidney Int 75: 736 –745, 2009. 443. Frey BA, Grisk O, Bandelow N, Wussow S, Bie P, Rettig R. Sodium homeostasis in transplanted rats with a spontaneously hypertensive rat kidney. Am J Physiol Regul Integr Comp Physiol 279: R1099 –R1104, 2000. 444. Frisbee JC, Maier KG, Stepp DW. Oxidant stress-induced increase in myogenic activation of skeletal muscle resistance arteries in obese Zucker rats. Am J Physiol Heart Circ Physiol 283: H2160 –H2168, 2002. 445. Frisbee JC, Roman RJ, Falck JR, Krishna UM, Lombard JH. 20-HETE contributes to myogenic activation of skeletal muscle resistance arteries in Brown Norway and Sprague-Dawley rats. Microcirculation 8: 45–55, 2001. 446. Frisbee JC, Roman RJ, Krishna UM, Falck JR, Lombard JH. 20-HETE modulates myogenic response of skeletal muscle resistance arteries from hypertensive Dahl-SS rats. Am J Physiol Heart Circ Physiol 280: H1066 –H1074, 2001. 447. Frohlich ED, Chien Y, Sesoko S, Pegram BL. Relationship between dietary sodium intake, hemodynamics, and cardiac mass in SHR and WKY rats. Am J Physiol Regul Integr Comp Physiol 264: R30 –R34, 1993. 448. Fu Y, Zhang R, Lu D, Liu H, Chandrashekar K, Juncos LA, Liu R. NOX2 is the primary source of angiotensin II-induced superoxide in the macula densa. Am J Physiol Regul Integr Comp Physiol 298: R707–R712, 2010. 449. Fujii S, Zhang L, Igarashi J, Kosaka H. L-Arginine reverses p47phox and gp91phox expression induced by high salt in Dahl rats. Hypertension 42: 1014 –1020, 2003. 450. Fujiwara K, Hayashi K, Matsuda H, Kubota E, Honda M, Ozawa Y, Saruta T. Altered pressure-natriuresis in obese Zucker rats. Hypertension 33: 1470 –1475, 1999. 451. Fuller AJ, Hauschild BC, Gonzalez-Villalobos R, Awayda MS, Imig JD, Inscho EW, Navar LG. Calcium and chloride channel activation by angiotensin II-AT1 receptors in preglomerular vascular smooth muscle cells. Am J Physiol Renal Physiol 289: F760 – F767, 2005. 452. Gabbai FB, Gushwa LC, Peterson OW, Wilson CB, Blantz RC. Analysis of renal function in the two-kidney Goldblatt model. Am J Physiol Renal Fluid Electrolyte Physiol 252: F131–F137, 1987. 482 459. Garcia-Estan J, Roman RJ. Role of renal interstitial hydrostatic pressure in the pressure diuresis response. Am J Physiol Renal Fluid Electrolyte Physiol 256: F63–F70, 1989. 460. Garvin JL, Herrera M, Ortiz PA. Regulation of renal NaCl transport by nitric oxide, endothelin, and ATP: clinical implications. Annu Rev Physiol 73: 359 –376, 2011. 461. Gattone VH, Evan AP, Willis LR, Luft FC. Renal afferent arteriole in the spontaneously hypertensive rat. Hypertension 5: 8 –16, 1983. 462. Ge Y, Gannon KP, Gousset M, Liu R, Murphey B, Drummond HA. Impaired myogenic constriction of the renal afferent arteriole in a mouse model of reduced betaENaC expression. Am J Physiol Renal Physiol 302: F1486 –F1493, 2012. 463. Ge Y, Murphy SR, Fan F, Williams JM, Falck JR, Liu R, Roman RJ. Role of 20-HETE in the impaired myogenic and TGF responses of the Af-Art of Dahl Salt-sensitive rats. Am J Physiol Renal Physiol 307: F509 –F515, 2014. 464. Ge Y, Murphy SR, Lu Y, Falck J, Liu R, Roman RJ. Endogenously produced 20-HETE modulates myogenic and TGF response in microperfused afferent arterioles. Prostaglandins Other Lipid Mediat 102–103: 42– 48, 2013. 465. Gebremedhin D, Fenoy FJ, Harder DR, Roman RJ. Enhanced vascular tone in the renal vasculature of spontaneously hypertensive rats. Hypertension 16: 648 – 654, 1990. 466. Gebremedhin D, Gopalakrishnan S, Harder DR. Endogenous events modulating myogenic regulation of cerebrovascular function. Curr Vasc Pharmacol 12: 810 – 817, 2014. 467. Gebremedhin D, Kaldunski ML, Jacobs ER, Harder DR, Roman RJ. Coexistence of two types of Ca2⫹-activated K⫹ channels in rat renal arterioles. Am J Physiol Renal Fluid Electrolyte Physiol 270: F69 –F81, 1996. 468. Gebremedhin D, Lange AR, Lowry TF, Taheri MR, Birks EK, Hudetz AG, Narayanan J, Falck JR, Okamoto H, Roman RJ, Nithipatikom K, Campbell WB, Harder DR. Production of 20-HETE and its role in autoregulation of cerebral blood flow. Circ Res 87: 60 – 65, 2000. 469. Gebremedhin D, Ma YH, Imig JD, Harder DR, Roman RJ. Role of cytochrome P-450 in elevating renal vascular tone in spontaneously hypertensive rats. J Vasc Res 30: 53– 60, 1993. 470. Gebremedhin D, Terashvili M, Wickramasekera N, Zhang DX, Rau N, Miura H, Harder DR. Redox signaling via oxidative inactivation of PTEN modulates pressuredependent myogenic tone in rat middle cerebral arteries. PLoS One 8: e68498, 2013. 471. Gill PS, Wilcox CS. NADPH oxidases in the kidney. Antioxid Redox Signal 8: 1597– 1607, 2006. 472. Gillies LK, Lu M, Wang H, Lee RMKW. AT1 receptor antagonist treatment caused persistent arterial functional changes in young spontaneously hypertensive rats. Hypertension 30: 1471–1478, 1997. Physiol Rev • VOL 95 • APRIL 2015 • www.prv.org CARLSTRÖM ET AL. 473. Gilmore JP, Cornish KG, Rogers SD, Joyner WL. Direct evidence for myogenic autoregulation of the renal microcirculation in the hamster. Circ Res 47: 226 –230, 1980. 494. Griendling KK, Minieri CA, Ollerenshaw JD, Alexander RW. Angiotensin II stimulates NADH and NADPH oxidase activity in cultured vascular smooth muscle cells. Circ Res 74: 1141–1148, 1994. 474. Gokina NI, Knot HJ, Nelson MT, Osol G. Increased Ca2⫹ sensitivity as a key mechanism of PKC-induced constriction in pressurized cerebral arteries. Am J Physiol Heart Circ Physiol 277: H1178 –H1188, 1999. 495. Griffin KA, Abu-Naser M, Abu-Amarah I, Picken M, Williamson GA, Bidani AK. Dynamic blood pressure load and nephropathy in the ZSF1 (fa/facp) model of type 2 diabetes. Am J Physiol Renal Physiol 293: F1605–F1613, 2007. 475. Gokina NI, Park KM, Elroy-Yaggy K, Osol G. Effects of Rho kinase inhibition on cerebral artery myogenic tone and reactivity. J Appl Physiol 98: 1940 –1948, 2005. 496. Griffin KA, Bidani AK. Hypertensive renal damage: insights from animal models and clinical relevance. Curr Hypertens Rep 6: 145–153, 2004. 476. Goldblatt H, Lynch J, Hanzal RF, Summerville WW. Studies on experimental hypertension. I. The production of persistent elevation of systolic blood pressure by means of renal ischemia. J Exp Med 59: 347–379, 1934. 497. Griffin KA, Bidani AK, Picken M, Ellis VR, Churchill PC. Prostaglandins do not mediate impaired autoregulation or increased renin secretion in remnant rat kidneys. Am J Physiol Renal Fluid Electrolyte Physiol 263: F1057–F1062, 1992. 477. Goligorsky MS, Colflesh D, Gordienko DV, Moore LC. Branching points of renal resistance arteries are enriched in L-type calcium channels and initiate vasoconstriction. Am J Physiol Renal Fluid Electrolyte Physiol 268: F251–F257, 1995. 478. Gollasch M, Lohn M, Furstenau M, Nelson MT, Luft FC, Haller H. Ca2⫹ channels, “quantized” Ca2⫹ release, and differentiation of myocytes in the cardiovascular system. J Hypertens 18: 989 –998, 2000. 479. Gomez RA, Norwood VF, Tufro-McReddie A. Development of the kidney vasculature. Microscopy Res Tech 39: 254 –260, 1997. 480. Gonzales AL, Amberg GC, Earley S. Ca2⫹ release from the sarcoplasmic reticulum is required for sustained TRPM4 activity in cerebral artery smooth muscle cells. Am J Physiol Cell Physiol 299: C279 –C288, 2010. 481. Gonzales AL, Earley S. Regulation of cerebral artery smooth muscle membrane potential by Ca2⫹-activated cation channels. Microcirculation 20: 337–347, 2012. 482. Gonzales AL, Yang Y, Sullivan MN, Sanders L, Dabertrand F, Hill-Eubanks DC, Nelson MT, Earley S. A PLCgamma1-dependent, force-sensitive signaling network in the myogenic constriction of cerebral arteries. Sci Signal 7: ra49, 2014. 483. Gonzalez JD, Llinas MT, Moreno C, Rodriguez F, Salazar FJ. Renal effects of prolonged cyclooxygenase inhibition when angiotensin II levels are elevated. J Cardiovasc Pharmacol 36: 236 –241, 2000. 484. Gonzalez JM, Somoza B, Conde MV, Fernandez-Alfonso MS, Gonzalez MC, Arribas SM. Hypertension increases middle cerebral artery resting tone in spontaneously hypertensive rats: role of tonic vasoactive factor availability. Clin Sci 114: 651– 659, 2008. 485. Gonzalez R, Fernandez-Alfonso MS, Rodriguez-Martinez MA, Fuertes E, Angulo J, Sanchez-Ferrer CF, Marin J. Pressure-induced contraction of the juxtamedullary afferent arterioles in spontaneously hypertensive rats. Gen Pharmacol 25: 333–339, 1994. 486. Gonzalez-Campoy JM, Long C, Roberts D, Berndt TJ, Romero JC, Knox FG. Renal interstitial hydrostatic pressure and PGE2 in pressure natriuresis. Am J Physiol Renal Fluid Electrolyte Physiol 260: F643–F649, 1991. 487. Goormaghtigh N. L’appareil neuro-myo-arteriel juxta-glmerulaire du rein: ses reactions en pathologie et ses rapports avec le tube urinifere. CR Seances Soc Biol Fil 124: 293–296, 1937. 488. Goransson A, Isaksson B, Sjoquist M. Whole kidney response to reduced arterial pressure during converting enzyme inhibition in the rat. Renal Physiol 9: 287–301, 1986. 498. Griffin KA, Churchill PC, Picken M, Webb RC, Kurtz TW, Bidani AK. Differential salt-sensitivity in the pathogenesis of renal damage in SHR and stroke prone SHR. Am J Hypertens 14: 311–320, 2001. 499. Griffin KA, Hacioglu R, Abu-Amarah I, Loutzenhiser R, Williamson GA, Bidani AK. Effects of calcium channel blockers on “dynamic” and “steady-state step” renal autoregulation. Am J Physiol Renal Physiol 286: F1136 –F1143, 2004. 500. Griffin KA, Picken M, Bakris GL, Bidani AK. Comparative effects of selective T- and L-type calcium channel blockers in the remnant kidney model. Hypertension 37: 1268 –1272, 2001. 501. Griffin KA, Picken M, Bidani AK. Radiotelemetric BP monitoring, antihypertensives and glomeruloprotection in remnant kidney model. Kidney Int 46: 1010 –1018, 1994. 502. Griffin KA, Picken M, Giobbie-Hurder A, Bidani AK. Low protein diet mediated renoprotection in remnant kidneys: renal autoregulatory versus hypertrophic mechanisms. Kidney Int 63: 607– 616, 2003. 503. Griffin KA, Picken MM, Bidani AK. Deleterious effects of calcium channel blockade on pressure transmission and glomerular injury in rat remnant kidneys. J Clin Invest 96: 793– 800, 1995. 504. Griffin KA, Picken MM, Bidani AK. Blood pressure lability and glomerulosclerosis after normotensive 5/6 renal mass reduction in the rat. Kidney Int 65: 209 –218, 2004. 505. Grifoni SC, Chiposi R, McKey SE, Ryan MJ, Drummond HA. Altered whole kidney blood flow autoregulation in a mouse model of reduced beta-ENaC. Am J Physiol Renal Physiol 298: F285–F292, 2010. 506. Grisk O, Rettig R. Renal transplantation studies in genetic hypertension. News Physiol Sci 16: 262–265, 2001. 507. Groschner K, Rosker C, Lukas M. Role of TRP channels in oxidative stress. Novartis Found Symp 258: 222–230, 2004. 508. Gross JL, de Azevedo MJ, Silveiro SP, Canani LH, Caramori ML, Zelmanovitz T. Diabetic nephropathy: diagnosis, prevention, treatment. Diabetes Care 28: 164 –176, 2005. 509. Gross JM, Dwyer JE, Knox FG. Natriuretic response to increased pressure is preserved with COX-2 inhibitors. Hypertension 34: 1163–1167, 1999. 510. Gross R, Kirchheim H, Brandstetter K. Basal vascular tone in the kidney. Evaluation from the static pressure-flow relationship under normal autoregulation and at maximal dilation in the dog. Circ Res 38: 525–531, 1976. 489. Gordienko DV, Clausen C, Goligorsky MS. Ionic currents and endothelin signaling in smooth muscle cells form rat renal resistance arterioles. Am J Physiol Renal Fluid Electrolyte Physiol 266: F325–F341, 1994. 511. Gross R, Kirchheim H, Ruffmann K. Effect of carotid occlusion and of perfusion pressure on renal function in conscious dogs. Circ Res 48: 777–784, 1981. 490. Gottlieb P, Folgering J, Maroto R, Raso A, Wood TG, Kurosky A, Bowman C, Bichet D, Patel A, Sachs F, Martinac B, Hamill OP, Honore E. Revisiting TRPC1 and TRPC6 mechanosensitivity. Pflügers Arch 455: 1097–1103, 2008. 512. Gross V, Lippoldt A, Bohlender J, Bader M, Hansson A, Luft FC. Cortical and medullary hemodynamics in deoxycorticosterone acetate-salt hypertensive mice. J Am Soc Nephrol 9: 346 –354, 1998. 491. Gouedard O, Blanc J, Gaudet E, Ponchon P, Elghozi JL. Contribution of the reninangiotensin system to short-term blood pressure variability during blockade of nitric oxide synthesis in the rat. Br J Pharmacol 119: 1085–1092, 1996. 513. Gross V, Lippoldt A, Schneider W, Luft FC. Effect of captopril and angiotensin II receptor blockade on pressure natriuresis in transgenic TGR(mRen-2)27 rats. Hypertension 26: 471– 479, 1995. 492. Granger JP, Schnackenberg CG. Renal mechanisms of angiotensin II-induced hypertension. Semin Nephrol 20: 417– 425, 2000. 514. Gross V, Obst M, Luft FC. Insights into angiotensin II receptor function through AT2 receptor knockout mice. Acta Physiol Scand 181: 487– 494, 2004. 493. Green R, Giebisch G. Luminal hypotonicity: a driving force for fluid absorption from the proximal tubule. Am J Physiol Renal Fluid Electrolyte Physiol 246: F167–F174, 1984. 515. Gross V, Roman RJ, Cowley AW Jr. Abnormal pressure-natriuresis in transgenic renin gene rats. J Hypertension 12: 1029 –1034, 1994. Physiol Rev • VOL 95 • APRIL 2015 • www.prv.org 483 RENAL AUTOREGULATORY MECHANISMS 516. Gross V, Schunck WH, Honeck H, Milia AF, Kargel E, Walther T, Bader M, Inagami T, Schneider W, Luft FC. Inhibition of pressure natriuresis in mice lacking the AT2 receptor. Kidney Int 57: 191–202, 2000. 537. Haas JA, Khraibi AA, Perrella MA, Knox FG. Role of renal interstitial hydrostatic pressure in natriuresis of systemic nitric oxide inhibition. Am J Physiol Renal Fluid Electrolyte Physiol 264: F411–F414, 1993. 517. Grossman E, Hoffman A, Keiser HR. Sodium intake modulates renal vascular reactivity to endothelin-1 in Dahl rats. Clin Exp Pharmacol Physiol 17: 121–128, 1990. 538. Haas JA, Knox FG. Effect of meclofenamate or ketoconazole on the natriuretic response to increased pressure. J Lab Clin Med 128: 202–207, 1996. 518. Gschwend S, Buikema H, Navis G, Henning RH, de Zeeuw D, Van Dokkum RP. Endothelial dilatory function predicts individual susceptibility to renal damage in the 5/6 nephrectomized rat. J Am Soc Nephrol 13: 2909 –2915, 2002. 539. Haas JA, Lockhart JC, Larson TS, Henrikson T, Knox FG. Natriuretic response to renal interstitial hydrostatic pressure during angiotensin II blockade. Am J Physiol Renal Fluid Electrolyte Physiol 266: F117–F119, 1994. 519. Gschwend S, Henning RH, Pinto YM, de Zeeuw D, van Gilst WH, Buikema H. Myogenic constriction is increased in mesenteric resistance arteries from rats with chronic heart failure: instantaneous counteraction by acute AT1 receptor blockade. Br J Pharmacol 139: 1317–1325, 2003. 540. Haberle DA, Konigbauer B, Davis JM, Kawata T, Mast C, Metz C, Dahlheim H. Autoregulation of the glomerular filtration rate and the single-nephron glomerular filtration rate despite inhibition of tubuloglomerular feedback in rats chronically volume-expanded by deoxycorticosterone acetate. Pflügers Arch 416: 548 –553, 1990. 520. Guan Z, Fuller BS, Yamamoto T, Cook AK, Pollock JS, Inscho EW. Pentosan polysulfate treatment preserves renal autoregulation in Ang II-infused hypertensive tats via normalization of P2X1 receptor activation. Am J Physiol Renal Physiol 298: F1276 – F1284, 2010. 541. Hacioglu R, Williamson GA, Abu-Amarah I, Griffin KA, Bidani AK. Characterization of dynamics in renal autoregulation using volterra models. IEEE Trans Biomed Eng 53: 2166 –2176, 2006. 521. Guan Z, Osmond DA, Inscho EW. P2X receptors as regulators of the renal microvasculature. Trends Pharmacol Sci 28: 646 – 652, 2007. 522. Guan Z, Pollock JS, Cook AK, Hobbs JL, Inscho EW. Effect of epithelial sodium channel blockade on the myogenic response of rat juxtamedullary afferent arterioles. Hypertension 54: 1062–1069, 2009. 523. Guan Z, Willgoss DA, Matthias A, Manley SW, Crozier S, Gobe G, Endre ZH. Facilitation of renal autoregulation by angiotensin II is mediated through modulation of nitric oxide. Acta Physiol Scand 179: 189 –201, 2003. 524. Guarasci GR, Kline RL. Pressure natriuresis following acute and chronic inhibition of nitric oxide synthase in rats. Am J Physiol Regul Integr Comp Physiol 270: R469 –R478, 1996. 525. Gui P, Chao JT, Wu X, Yang Y, Davis GE, Davis MJ. Coordinated regulation of vascular Ca2⫹ and K⫹ channels by integrin signaling. Adv Exp Med Biol 674: 69 –79, 2010. 526. Gunther S, Gimbrone MA Jr, Alexander RW. Regulation by angiotensin II of its receptors in resistance blood vessels. Nature 287: 230 –232, 1980. 527. Gustafsson F, Holstein-Rathlou NH. Conducted vasomotor responses in arterioles: characteristics, mechanisms and physiological significance. Acta Physiol Scand 167: 11–21, 1999. 528. Gustafsson H. Vasomotion and underlying mechanisms in small arteries. An in vitro study of rat blood vessels. Acta Physiol Scand Suppl 614: 1– 44, 1993. 529. Gustafsson H, Bulow A, Nilsson H. Rhythmic contractions of isolated, pressurized small arteries from rat. Acta Physiol Scand 152: 145–152, 1994. 530. Gustafsson H, Mulvany MJ, Nilsson H. Rhythmic contractions of isolated small arteries from rat: influence of the endothelium. Acta Physiol Scand 148: 153–163, 1993. 531. Gustafsson H, Nilsson H. Rhythmic contractions of isolated small arteries from rat: role of calcium. Acta Physiol Scand 149: 283–291, 1993. 532. Guyton AC, Hall JE, Coleman TG, Manning RD Jr, Norman RA. The dominant role of the kidneys in long-term arterial pressure regulation in normal and hypertensive states. In: Hypertension: Pathophysiology, Diagnosis and Management, edited by Laragh JH and Brenner BM. New York: Raven, 1995. 533. Guyton AC, Langston JB, Navar LG. Theory for renal autoregulation by feedback at the juxtaglomerular apparatus. Circ Res 15: 187–197, 1964. 542. Haddock RE, Grayson TH, Brackenbury TD, Meaney KR, Neylon CB, Sandow SL, Hill CE. Endothelial coordination of cerebral vasomotion via myoendothelial gap junctions containing connexins 37 and 40. Am J Physiol Heart Circ Physiol 291: H2047– H2056, 2006. 543. Haddock RE, Hill CE. Rhythmicity in arterial smooth muscle. J Physiol 566: 645– 656, 2005. 544. Haddock RE, Hirst GD, Hill CE. Voltage independence of vasomotion in isolated irideal arterioles of the rat. J Physiol 540: 219 –229, 2002. 545. Haefliger JA, Krattinger N, Martin D, Pedrazzini T, Capponi A, Doring B, Plum A, Charollais A, Willecke K, Meda P. Connexin43-dependent mechanism modulates renin secretion and hypertension. J Clin Invest 116: 405– 413, 2006. 546. Hahne B, Selen G, Persson AEG. Indomethacin inhibits renal functional adaptation to nephron loss. Ren Physiol 7: 13–21, 1984. 547. Hall JE. Control of sodium excretion by angiotensin II: intrarenal mechanisms and blood pressure regulation. Am J Physiol Regul Integr Comp Physiol 250: R960 –R972, 1986. 548. Hall JE, Coleman TG, Guyton AC, Balfe JW, Salgado HC. Intrarenal role of angiotensin II and [des-Asp1]angiotensin II. Am J Physiol Renal Fluid Electrolyte Physiol 236: F252–F259, 1979. 549. Hall JE, Granger JP, Hester RL, Coleman TG, Smith MJ Jr, Cross RB. Mechanisms of escape from sodium retention during angiotensin II hypertension. Am J Physiol Renal Fluid Electrolyte Physiol 246: F627–F634, 1984. 550. Hall JE, Guyton AC, Cowley AW Jr. Dissociation of renal blood flow and filtration rate autoregulation by renin depletion. Am J Physiol Renal Fluid Electrolyte Physiol 232: F215–F221, 1977. 551. Hampton JA, Bernardo DA, Khan NA, Lacher DA, Rapp JP, Gohara AF, Goldblatt PJ. Morphometric evaluation of the renal arterial system of Dahl salt-sensitive and salt-resistant rats on a high salt diet. II. Interlobular arteries and intralobular arterioles. Lab Invest 60: 839 – 846, 1989. 552. Hanner F, Lam L, Nguyen MT, Yu A, Peti-Peterdi J. Intrarenal localization of the plasma membrane ATP channel pannexin1. Am J Physiol Renal Physiol 303: F1454 – F1459, 2012. 553. Hanner F, Sorensen CM, Holstein-Rathlou NH, Peti-Peterdi J. Connexins and the kidney. Am J Physiol Regul Integr Comp Physiol 298: R1143–R1155, 2010. 534. Guzik TJ, Hoch NE, Brown KA, McCann LA, Rahman A, Dikalov S, Goronzy J, Weyand C, Harrison DG. Role of the T cell in the genesis of angiotensin II induced hypertension and vascular dysfunction. J Exp Med 204: 2449 –2460, 2007. 554. Hanner F, von Maltzahn J, Maxeiner S, Toma I, Sipos A, Kruger O, Willecke K, Peti-Peterdi J. Connexin45 is expressed in the juxtaglomerular apparatus and is involved in the regulation of renin secretion and blood pressure. Am J Physiol Regul Integr Comp Physiol 295: R371–R380, 2008. 535. Haas JA, Granger JP, Knox FG. Effect of renal perfusion pressure on sodium reabsorption from proximal tubules of superficial and deep nephrons. Am J Physiol Renal Fluid Electrolyte Physiol 250: F425–F429, 1986. 555. Hansen PB, Castrop H, Briggs J, Schnermann J. Adenosine induces vasoconstriction through Gi-dependent activation of phospholipase C in isolated perfused afferent arterioles of mice. J Am Soc Nephrol 14: 2457–2465, 2003. 536. Haas JA, Granger JP, Knox FG. Effect of meclofenamate on lithium excretion in response to changes in renal perfusion pressure. J Lab Clin Med 111: 543–547, 1988. 556. Hansen PB, Hashimoto S, Briggs J, Schnermann J. Attenuated renovascular constrictor responses to angiotensin II in adenosine 1 receptor knockout mice. Am J Physiol Regul Integr Comp Physiol 285: R44 –R49, 2003. 484 Physiol Rev • VOL 95 • APRIL 2015 • www.prv.org CARLSTRÖM ET AL. 557. Hansen PB, Hashimoto S, Oppermann M, Huang Y, Briggs JP, Schnermann J. Vasoconstrictor and vasodilator effects of adenosine in the mouse kidney due to preferential activation of A1 or A2 adenosine receptors. J Pharmacol Exp Ther 315: 1150 – 1157, 2005. 579. Harrison DG, Gongora MC, Guzik TJ, Widder J. Oxidative stress and hypertension. J Am Soc Hypertens 1: 30 – 44, 2007. 580. Harrison-Bernard LM, Navar LG, Cook AK. Renal cortical and medullary microvascular blood flow autoregulation in rat. Kidney Int 50: S23–S29, 1996. 558. Hansen PB, Jensen BL, Andreasen D, Skott O. Differential expression of T- and L-type voltage-dependent calcium channels in renal resistance vessels. Circ Res 89: 630 – 638, 2001. 581. Harsing L, Fonyodi S, Laszlo K, Takacs G. Effect of hypertonic infusions on renal haemodynamics. Acta Physiol Hung 12: 351–361, 1957. 559. Hansen PB, Jensen BL, Skott O. Chloride regulates afferent arteriolar contraction in response to depolarization. Hypertension 32: 1066 –1070, 1998. 582. Hashimoto S, Adams JW, Bernstein KE, Schnermann J. Micropuncture determination of nephron function in mice without tissue angiotensin-converting enzyme. Am J Physiol Renal Physiol 288: F445–F452, 2005. 560. Hansen PB, Poulsen CB, Walter S, Marcussen N, Cribbs LL, Skott O, Jensen BL. Functional importance of L- and P/Q-type voltage-gated calcium channels in human renal vasculature. Hypertension 58: 464 – 470, 2011. 561. Harder DR. Pressure-dependent membrane depolarization in cat middle cerebral artery. Circ Res 55: 197–202, 1984. 562. Harder DR, Gilbert R, Lombard JH. Vascular muscle cell depolarization and activation in renal arteries on elevation of transmural pressure. Am J Physiol Renal Fluid Electrolyte Physiol 253: F778 –F781, 1987. 563. Harder DR, Narayanan J, Gebremedhin D. Pressure-induced myogenic tone and role of 20-HETE in mediating autoregulation of cerebral blood flow. Am J Physiol Heart Circ Physiol 300: H1557–H1565, 2011. 564. Harder DR, Smeda J, Lombard J. Enhanced myogenic depolarization in hypertensive cerebral arterial muscle. Circ Res 57: 319 –322, 1985. 565. Harhun MI, Povstyan OV, Gordienko DV. Purinoreceptor-mediated current in myocytes from renal resistance arteries. Br J Pharmacol 160: 987–997, 2010. 566. Harper SL, Bohlen HG. Microvascular adaptation in the cerebral cortex of adult spontaneously hypertensive rats. Hypertension 6: 408 – 419, 1984. 567. Harrap SB. Angiotensin converting enzyme inhibitors, regional vascular hemodynamics, and the development and prevention of experimental genetic hypertension. Am J Hypertens 4: 212S–216S, 1991. 568. Harrap SB. Cardiovascular disease and genetics of the renin-angiotensin system. Heart 76: 13–17, 1996. 569. Harrap SB, Clark SA, Fraser R, Towrie A, Brown AJ, Lever AF. Effects of sodium intake and aldosterone on the renal pressure-natriuresis. Am J Physiol Renal Fluid Electrolyte Physiol 254: F697–F703, 1988. 570. Harrap SB, Doyle AE. Total body sodium in immature spontaneously hypertensive and Wistar Kyoto rats. Clin Exp Pharmacol Physiol 12: 315–318, 1985. 571. Harrap SB, Doyle AE. Renal haemodynamics and total body sodium in immature spontaneously hypertensive and Wistar-Kyoto rats. J Hypertens Suppl 4: S249 –S252, 1986. 572. Harrap SB, Doyle AE. Genetic co-segregation of renal haemodynamics and blood pressure in the spontaneously hypertensive rat. Clin Sci 74: 63– 69, 1988. 573. Harrap SB, Mirakian C, Datodi SR, Lever AF. Blood pressure and lifespan following brief ACE inhibitor treatment in young spontaneously hypertensive rats. Clin Exp Pharmacol Physiol 21: 125–127, 1994. 574. Harrap SB, Nicolaci JA, Doyle AE. Persistent effects on blood pressure and renal haemodynamics following chronic angiotensin converting enzyme inhibition with perindopril. Clin Exp Pharmacol Physiol 13: 753–765, 1986. 575. Harrap SB, Wang BZ, MacLellan DG. Transplantation studies of the role of the kidney in long-term blood pressure reduction following brief ACE inhibitor treatment in young spontaneously hypertensive rats. Clin Exp Pharmacol Physiol 21: 129 – 131, 1994. 576. Harraz OF, Brett SE, Welsh DG. Nitric oxide suppresses vascular voltage-gated T-type Ca2⫹ channels through cGMP/PKG signaling. Am J Physiol Heart Circ Physiol 306: H279 –H285, 2014. 583. Hashimoto S, Huang Y, Briggs J, Schnermann J. Reduced autoregulatory effectiveness in adenosine 1 receptor-deficient mice. Am J Physiol Renal Physiol 290: F888 – F891, 2006. 584. Hashimoto S, Yamada K, Kawata T, Mochizuki T, Schnermann J, Koike T. Abnormal autoregulation and tubuloglomerular feedback in prediabetic and diabetic OLETF rats. Am J Physiol Renal Physiol 296: F598 –F604, 2009. 585. Hayakawa H, Raij L. Nitric oxide synthase activity and renal injury in genetic hypertension. Hypertension 31: 266 –270, 1998. 586. Hayashi K, Epstein M, Loutzenhiser R. Pressure-induced vasoconstriction of renal microvessels in normotensive and hypertensive rats. Studies in the isolated perfused hydronephrotic kidney. Circ Res 65: 1475–1484, 1989. 587. Hayashi K, Epstein M, Loutzenhiser R. Enhanced myogenic responsiveness of renal interlobular arteries in spontaneously hypertensive rats. Hypertension 19: 153–160, 1992. 588. Hayashi K, Epstein M, Loutzenhiser R, Forster H. Impaired myogenic responsiveness of the afferent arteriole in streptozotocin-induced diabetic rats: role of eicosanoid derangements. J Am Soc Nephrol 2: 1578 –1586, 1992. 589. Hayashi K, Epstein M, Saruta T. Altered myogenic responsiveness of the renal microvasculature in experimental hypertension. J Hypertens 14: 1387–1401, 1996. 590. Hayashi K, Fujiwara K, Oka K, Nagahama T, Matsuda H, Saruta T. Effects of insulin on rat renal microvessels: studies in the isolated perfused hydronephrotic kidney. Kidney Int 51: 1507–1513, 1997. 591. Hayashi K, Kanda T, Homma K, Tokuyama H, Okubo K, Takamatsu I, Tatematsu S, Kumagai H, Saruta T. Altered renal microvascular response in Zucker obese rats. Metabolism 51: 1553–1561, 2002. 592. Hayashi K, Loutzenhiser RD, Epstein M, Suzuki H, Saruta T. Multiple factors contribute to acetylcholine-induced renal afferent arteriolar vasodilation during myogenic and norepinephrine- and KCl-induced vasoconstriction. Studies in the isolated perfused hydronephrotic kidney. Circ Res 75: 821– 828, 1994. 593. Hayashi K, Suzuki H, Saruta T. Nitric oxide modulates but does not impair myogenic vasoconstriction of the afferent arteriole in spontaneously hypertensive rats. Hypertension 25: 1212–1219, 1995. 594. Hayashi K, Wakino S, Kanda T, Homma K, Sugano N, Saruta T. Molecular mechanisms and therapeutic strategies of chronic renal injury: role of rho-kinase in the development of renal injury. J Pharmacol Sci 100: 29 –33, 2006. 595. Hayashi K, Wakino S, Sugano N, Ozawa Y, Homma K, Saruta T. Ca2⫹ channel subtypes and pharmacology in the kidney. Circ Res 100: 342–353, 2007. 596. Hayashi M, Monkawa T, Saruta T. Cell-specific expression of the IP3 receptor gene family in the kidney. Exp Nephrol 8: 215–218, 2000. 597. He J, Marsh DJ. Effect of captopril on fluctuations of blood pressure and renal blood flow in rats. Am J Physiol Renal Fluid Electrolyte Physiol 264: F37–F44, 1993. 598. Heeg JE, de Jong PE, de Zeeuw D. Additive antiproteinuric effect of angiotensinconverting enzyme inhibition and non-steroidal anti-inflammatory drug therapy: a clue to the mechanism of action. Clin Sci 81: 367–372, 1991. 577. Harris RC. An update on cyclooxygenase-2 expression and metabolites in the kidney. Curr Opin Nephrol Hypertens 17: 64 – 69, 2008. 599. Heinze C, Seniuk A, Sokolov MV, Huebner AK, Klementowicz AE, Szijarto IA, Schleifenbaum J, Vitzthum H, Gollasch M, Ehmke H, Schroeder BC, Hubner CA. Disruption of vascular Ca2⫹-activated chloride currents lowers blood pressure. J Clin Invest 124: 675– 686, 2014. 578. Harrison DG, Gongora MC. Oxidative stress and hypertension. Med Clin North Am 93: 621– 635, 2009. 600. Heistad DD, Baumbach GL. Cerebral vascular changes during chronic hypertension: good guys and bad guys. J Hypertens Suppl 10: S71–S75, 1992. Physiol Rev • VOL 95 • APRIL 2015 • www.prv.org 485 RENAL AUTOREGULATORY MECHANISMS 601. Helal I, Fick-Brosnahan GM, Reed-Gitomer B, Schrier RW. Glomerular hyperfiltration: definitions, mechanisms and clinical implications. Nat Rev Nephrol 8: 293–300, 2012. 622. Hodges GJ, Del Pozzi AT. Noninvasive examination of endothelial, sympathetic, and myogenic contributions to regional differences in the human cutaneous microcirculation. Microvasc Res 93: 87–91, 2014. 602. Hellebrekers LJ, Liard JF, Laborde AL, Greene AS, Cowley AW Jr. Regional autoregulatory responses during infusion of vasoconstrictor agents in conscious dogs. Am J Physiol Heart Circ Physiol 259: H1270 –H1277, 1990. 623. Hofmann T, Obukhov AG, Schaefer M, Harteneck C, Gudermann T, Schultz G. Direct activation of human TRPC6 and TRPC3 channels by diacylglycerol. Nature 397: 259 –263, 1999. 603. Henrion D, Laher I. Effects of staurosporine and calphostin C, two structurally unrelated inhibitors of protein kinase C, on vascular tone. Can J Physiol Pharmacol 71: 521–524, 1993. 624. Holm L, Morsing P, Casellas D, Persson AE. Resetting of the pressure range for blood flow autoregulation in the rat kidney. Acta Physiol Scand 138: 395– 401, 1990. 604. Hercule HC, Tank J, Plehm R, Wellner M, da Costa Goncalves AC, Gollasch M, Diedrich A, Jordan J, Luft FC, Gross V. Regulator of G protein signalling 2 ameliorates angiotensin II-induced hypertension in mice. Exp Physiol 92: 1014 –1022, 2007. 605. Herlitz H, Lundin S, Ricksten SE, Gothberg G, Aurell M, Hallback M, Berglund G. Sodium balance and structural vascular changes in the kidney during development of hypertension in spontaneously hypertensive rats. Acta Med Scand Suppl 625: 111– 115, 1979. 625. Holstein-Rathlou NH. Oscillations and chaos in renal blood flow control. J Am Soc Nephrol 4: 1275–1287, 1993. 626. Holstein-Rathlou NH, Leyssac PP. TGF-mediated oscillations in the proximal intratubular pressure: differences between spontaneously hypertensive rats and WistarKyoto rats. Acta Physiol Scand 126: 333–339, 1986. 627. Holstein-Rathlou NH, Marsh DJ. Oscillations of tubular pressure, flow, and distal chloride concentration in rats. Am J Physiol Renal Fluid Electrolyte Physiol 256: F1007– F1014, 1989. 606. Herlitz H, Ricksten SE, Lundin S, Thoren P, Aurell M, Berglund G. Renal denervation and sodium balance in young spontaneously hypertensive rats. Renal Physiol 6: 145– 150, 1983. 628. Holstein-Rathlou NH, Marsh DJ. A dynamic model of renal blood flow autoregulation. Bull Math Biol 56: 411– 429, 1994. 607. Herrera GA, Turbat-Herrera EA, Teng J. Mesangial homeostasis and pathobiology: their role in health and disease. Contrib Nephrol 169: 6 –22, 2011. 629. Holstein-Rathlou NH, Marsh DJ. Renal blood flow regulation and arterial pressure fluctuations: a case study in nonlinear dynamics. Physiol Rev 74: 637– 681, 1994. 608. Herrera M, Ortiz PA, Garvin JL. Regulation of thick ascending limb transport: role of nitric oxide. Am J Physiol Renal Physiol 290: F1279 –F1284, 2006. 630. Holstein-Rathlou NH, Sosnovtseva OV, Pavlov AN, Cupples WA, Sorensen CM, Marsh DJ. Nephron blood flow dynamics measured by laser speckle contrast imaging. Am J Physiol Renal Physiol 300: F319 –F329, 2011. 609. Herrera-Acosta J, Tapia E, Sanchez-Lozada LG, Franco M, Striker LJ, Striker GE, Rodriguez IB. Restoration of glomerular haemodynamics and renal injury independent of arterial hypertension in rats with subtotal renal ablation. J Hypertens Suppl 20: S29 –S35, 2002. 631. Holstein-Rathlou NH, Wagner AJ, Marsh DJ. Tubuloglomerular feedback dynamics and renal blood flow autoregulation in rats. Am J Physiol Renal Fluid Electrolyte Physiol 260: F53–F68, 1991. 610. Hester RL, Granger JP, Williams J, Hall JE. Acute and chronic servo-control of renal perfusion pressure. Am J Physiol Renal Fluid Electrolyte Physiol 244: F455–F460, 1983. 611. Heyeraas KJ, Aukland K. Interlobular arterial resistance: influence of renal arterial pressure and angiotensin II. Kidney Int 31: 1291–1298, 1987. 612. Hill JV, Findon G, Appelhoff RJ, Endre ZH. Renal autoregulation and passive pressure-flow relationships in diabetes and hypertension. Am J Physiol Renal Physiol 299: F837–F844, 2010. 613. Hill MA, Davis MJ, Meininger GA. Cyclooxygenase inhibition potentiates myogenic activity in skeletal muscle arterioles. Am J Physiol Heart Circ Physiol 258: H127–H133, 1990. 614. Hill MA, Davis MJ, Meininger GA, Potocnik SJ, Murphy TV. Arteriolar myogenic signalling mechanisms: Implications for local vascular function. Clin Hemorheol Microcirc 34: 67–79, 2006. 615. Hill MA, Falcone JC, Meininger GA. Evidence for protein kinase C involvement in arteriolar myogenic reactivity. Am J Physiol Heart Circ Physiol 259: H1586 –H1594, 1990. 616. Hill MA, Meininger GA. Impaired arteriolar myogenic reactivity in early experimental diabetes. Diabetes 42: 1226 –1232, 1993. 617. Hill MA, Meininger GA. Arteriolar vascular smooth muscle cells: mechanotransducers in a complex environment. Int J Biochem Cell Biol 44: 1505–1510, 2012. 2⫹ ⫹ 618. Hill MA, Yang Y, Ella SR, Davis MJ, Braun AP. Large conductance, Ca -activated K channels (BKCa) and arteriolar myogenic signaling. FEBS Lett 584: 2033–2042, 2010. 619. Hilliard LM, Nematbakhsh M, Kett MM, Teichman E, Sampson AK, Widdop RE, Evans RG, Denton KM. Gender differences in pressure-natriuresis and renal autoregulation: role of the angiotensin type 2 receptor. Hypertension 57: 275–282, 2011. 632. Homma K, Hayashi K, Wakino S, Tokuyama H, Kanda T, Tatematsu S, Hasegawa K, Fujishima S, Hori S, Saruta T, Itoh H. Rho-kinase contributes to pressure-induced constriction of renal microvessels. Keio J Med 63: 1–12, 2014. 633. Homma K, Hayashi K, Yamaguchi S, Fujishima S, Hori S, Itoh H. Renal microcirculation and calcium channel subtypes. Curr Hypertens Rev 9: 182–186, 2013. 634. Horiguchi S, Watanabe J, Kato H, Baba S, Shinozaki T, Miura M, Fukuchi M, Kagaya Y, Shirato K. Contribution of Na⫹/Ca2⫹ exchanger to the regulation of myogenic tone in isolated rat small arteries. Acta Physiol Scand 173: 167–173, 2001. 635. Horvath B, Lenzser G, Benyo B, Nemeth T, Benko R, Iring A, Herman P, Komjati K, Lacza Z, Sandor P, Benyo Z. Hypersensitivity to thromboxane receptor mediated cerebral vasomotion and CBF oscillations during acute NO-deficiency in rats. PLoS One 5: e14477, 2010. 636. Hostetter TH, Meyer TW, Rennke HG, Brenner BM. Chronic effects of dietary protein in the rat with intact and reduced renal mass. Kidney Int 30: 509 –517, 1986. 637. Hostetter TH, Olson JL, Rennke HG, Venkatachalam MA, Brenner BM. Hyperfiltration in remnant nephrons: a potentially adverse response to renal ablation. Am J Physiol Renal Fluid Electrolyte Physiol 241: F85–F93, 1981. 638. Hostetter TH, Troy JL, Brenner BM. Glomerular hemodynamics in experimental diabetes mellitus. Kidney Int 19: 410 – 415, 1981. 639. Hsu CH, Slavicek JH, Kurtz TW. Segmental renal vascular resistance in the spontaneously hypertensive rat. Am J Physiol Heart Circ Physiol 242: H961–H966, 1982. 640. Hsu CH, Slavicek JM. The effect of renal perfusion pressure on renal vascular resistance in the spontaneously hypertensive rat. Pflügers Arch 393: 340 –343, 1982. 641. Hu L, Zhang Y, Lim PS, Miao Y, Tan C, McKenzie KU, Schyvens CG, Whitworth JA. Apocynin but not L-arginine prevents and reverses dexamethasone-induced hypertension in the rat. Am J Hypertens 19: 413– 418, 2006. 620. Hirata Y, Hayakawa H, Suzuki E, Kimura K, Kikuchi K, Nagano T, Hirobe M, Omata M. Direct measurements of endothelium-derived nitric oxide release by stimulation of endothelin receptors in rat kidney and its alteration in salt-induced hypertension. Circulation 91: 1229 –1235, 1995. 642. Hua JL, Kaskel FJ, Juno CJ, Moore LC, McCaughran JA Jr. Salt intake and renal hemodynamics in immature and mature Dahl salt-sensitive (DS/JR) and salt-resistant (DR/JR) rats. Am J Hypertens 3: 268 –273, 1990. 621. Hirst GD, van Helden DF. Ionic basis of the resting potential of submucosal arterioles in the ileum of the guinea-pig. J Physiol 333: 53– 67, 1982. 643. Huang A, Koller A. Endothelin and prostaglandin H2 enhance arteriolar myogenic tone in hypertension. Hypertension 30: 1210 –1215, 1997. 486 Physiol Rev • VOL 95 • APRIL 2015 • www.prv.org CARLSTRÖM ET AL. 644. Huang A, Sun D, Koller A. Endothelial dysfunction augments myogenic arteriolar constriction in hypertension. Hypertension 22: 913–921, 1993. 665. Ikenaga H, Bast JP, Fallet RW, Carmines PK. Exaggerated impact of ATP-sensitive K⫹ channels on afferent arteriolar diameter in diabetes mellitus. J Am Soc Nephrol 11: 1199 –1207, 2000. 645. Huang A, Sun D, Yan C, Falck JR, Kaley G. Contribution of 20-HETE to augmented myogenic constriction in coronary arteries of endothelial NO synthase knockout mice. Hypertension 46: 607– 613, 2005. 666. Ikenaga H, Suzuki H, Ishii N, Itoh H, Saruta T. Role of NO on pressure-natriuresis in Wistar-Kyoto and spontaneously hypertensive rats. Kidney Int 43: 205–211, 1993. 646. Huang C, Davis G, Johns EJ. Effect of nitrendipine on autoregulation of perfusion in the cortex and papilla of kidneys from Wistar and stroke prone spontaneously hypertensive rats. Br J Pharmacol 111: 111–116, 1994. 667. Ikeoka K, Nishigaki K, Ohyanagi M, Faber JE. In vitro analysis of alpha-adrenoceptor interactions with the myogenic response in resistance vessels. J Vasc Res 29: 313– 321, 1992. 647. Huang DY, Vallon V, Zimmermann H, Koszalka P, Schrader J, Osswald H. Ecto-5’nucleotidase (cd73)-dependent and -independent generation of adenosine participates in the mediation of tubuloglomerular feedback in vivo. Am J Physiol Renal Physiol 291: F282–F288, 2006. 668. Iliescu R, Cazan R, McLemore GR Jr, Venegas-Pont M, Ryan MJ. Renal blood flow and dynamic autoregulation in conscious mice. Am J Physiol Renal Physiol 295: F734 – F740, 2008. 648. Huang WC, Bell PD, Harvey D, Mitchell KD, Navar LG. Angiotensin influences on tubuloglomerular feedback mechanism in hypertensive rats. Kidney Int 34: 631– 637, 1988. 649. Huang WC, Ploth DW, Bell PD, Work J, Navar LG. Bilateral renal function responses to converting enzyme inhibitor (SQ 20,881) in two-kidney, one clip Goldblatt hypertensive rats. Hypertension 3: 285–293, 1981. 650. Huang WC, Ploth DW, Navar LG. Effects of saralasin infusion on bilateral renal function in two-kidney, one-clip Goldblatt hypertensive rats. Clin Sci 62: 573–579, 1982. 651. Huang Y, Cheung KK. Endothelium-dependent rhythmic contractions induced by cyclopiazonic acid in rat mesenteric artery. Eur J Pharmacol 332: 167–172, 1997. 652. Huelsemann JL, Sterzel RB, McKenzie DE, Wilcox CS. Effects of a calcium entry blocker on blood pressure and renal function during angiotensin-induced hypertension. Hypertension 7: 374 –379, 1985. 653. Hundley WG, Renaldo GJ, Levasseur JE, Kontos HA. Vasomotion in cerebral microcirculation of awake rabbits. Am J Physiol Heart Circ Physiol 254: H67–H71, 1988. 654. Ibarrola AM, Inscho EW, Vari RC, Navar LG. Influence of adenosine receptor blockade on renal function and renal autoregulation. J Am Soc Nephrol 2: 991–999, 1991. 655. Ichihara A, Hayashi M, Hirota N, Saruta T. Superoxide inhibits neuronal nitric oxide synthase influences on afferent arterioles in spontaneously hypertensive rats. Hypertension 37: 630 – 634, 2001. 656. Ichihara A, Hayashi M, Koura Y, Tada Y, Sugaya T, Hirota N, Saruta T. Blunted tubuloglomerular feedback by absence of angiotensin type 1A receptor involves neuronal NOS. Hypertension 40: 934 –939, 2002. 657. Ichihara A, Imig JD, Inscho EW, Navar LG. Cyclooxygenase-2 participates in tubular flow-dependent afferent arteriolar tone: interaction with neuronal NOS. Am J Physiol Renal Physiol 275: F605–F612, 1998. 658. Ichihara A, Imig JD, Navar LG. Cyclooxygenase-2 modulates afferent arteriolar responses to increases in pressure. Hypertension 34: 843– 847, 1999. 659. Ichihara A, Imig JD, Navar LG. Neuronal nitric oxide synthase-dependent afferent arteriolar function in angiotensin II-induced hypertension. Hypertension 33: 462– 466, 1999. 660. Ichihara A, Inscho EW, Imig JD, Michel RE, Navar LG. Role of renal nerves in afferent arteriolar reactivity in angiotensin-induced hypertension. Hypertension 29: 442– 449, 1997. 661. Ichihara A, Inscho EW, Imig JD, Navar LG. Neuronal nitric oxide synthase modulates rat renal microvascular function. Am J Physiol Renal Physiol 274: F516 – F524, 1998. 662. Ichihara A, Navar LG. Neuronal NOS contributes to biphasic autoregulatory response during enhanced TGF activity. Am J Physiol Renal Physiol 277: F113–F120, 1999. 663. Iesaki T, Okada T, Shimada I, Yamaguchi H, Ochi R. Decrease in Ca2⫹ sensitivity as a mechanism of hydrogen peroxide-induced relaxation of rabbit aorta. Cardiovasc Res 31: 820 – 825, 1996. 664. Iijima K, Lin L, Nasjletti A, Goligorsky MS. Intracellular signaling pathway of endothelin-1. J Cardiovasc Pharmacol 17 Suppl 7: S146 –S149, 1991. 669. Imamura A, Mackenzie HS, Lacy ER, Hutchison FN, Fitzgibbon WR, Ploth DW. Effects of chronic treatment with angiotensin converting enzyme inhibitor or an angiotensin receptor antagonist in two-kidney, one-clip hypertensive rats. Kidney Int 47: 1394 –1402, 1995. 670. Imig JD. Afferent arteriolar reactivity to angiotensin II is enhanced during the early phase of angiotensin II hypertension. Am J Hypertens 13: 810 – 818, 2000. 671. Imig JD. Eicosanoid regulation of the renal vasculature. Am J Physiol Renal Physiol 279: F965–F981, 2000. 672. Imig JD, Falck JR, Gebremedhin D, Harder DR, Roman RJ. Elevated renovascular tone in young spontaneously hypertensive rats. Role of cytochrome P-450. Hypertension 22: 357–364, 1993. 673. Imig JD, Falck JR, Inscho EW. Contribution of cytochrome P450 epoxygenase and hydroxylase pathways to afferent arteirolar autorregulatory responsiveness. Br J Pharmacol 127: 1399 –1405, 1999. 674. Imig JD, Passmore JC, Anderson GL, Jimenez AE. Chloride alters renal blood flow autoregulation in deoxycorticosterone-treated rats. J Lab Clin Med 121: 608 – 613, 1993. 675. Imig JD, Roman RJ. Nitric oxide modulates vascular tone in preglomerular arterioles. Hypertension 19: 770 –774, 1992. 676. Imig JD, Zou AP, Ortiz de Montellano PR, Sui Z, Roman, RJ. Cytochrome P-450 inhibitors alter afferent arteriolar responses to elevations in pressure. Am J Physiol Heart Circ Physiol 266: H1879 –H1885, 1994. 677. Imig JD, Zou AP, Stec DE, Harder DR, Falck JR, Roman, RJ. Formation and actions of 20-hydroxyeicosatetraenoic acid in rat renal arterioles. Am J Physiol Regul Integr Comp Physiol 270: R217–R227, 1996. 678. Ingber DE. Cellular mechanotransduction: putting all the pieces together again. FASEB J 20: 811– 827, 2006. 679. Inoue R, Hai L, Honda A. Pathophysiological implications of transient receptor potential channels in vascular function. Curr Opin Nephrol Hypertens 17: 193–198, 2008. 680. Inoue R, Jensen LJ, Jian Z, Shi J, Hai L, Lurie AI, Henriksen FH, Salomonsson M, Morita H, Kawarabayashi Y, Mori M, Mori Y, Ito Y. Synergistic activation of vascular TRPC6 channel by receptor and mechanical stimulation via phospholipase C/diacylglycerol and phospholipase A2/omega-hydroxylase/20-HETE pathways. Circ Res 104: 1399 –1409, 2009. 681. Inoue R, Jensen LJ, Shi J, Morita H, Nishida M, Honda A, Ito Y. Transient receptor potential channels in cardiovascular function and disease. Circ Res 99: 119 –131, 2006. 682. Inscho EW. P2 receptors in regulation of renal microvascular function. Am J Physiol Renal Physiol 280: F927–F944, 2001. 683. Inscho EW. ATP, P2 receptors and the renal microcirculation. Purinergic Signal 5: 447– 460, 2009. 684. Inscho EW, Carmines PK, Cook AK, Navar LG. Afferent arteriolar responsiveness to altered perfusion pressure in renal hypertension. Hypertension 15: 748 –752, 1990. 685. Inscho EW, Cook AK. P2 receptor-mediated afferent arteriolar vasoconstriction during calcium blockade. Am J Physiol Renal Physiol 282: F245–F255, 2002. 686. Inscho EW, Cook AK, Imig JD, Vial C, Evans RJ. Physiological role for P2X1 receptors in renal microvascular autoregulatory behavior. J Clin Invest 112: 1895–1905, 2003. Physiol Rev • VOL 95 • APRIL 2015 • www.prv.org 487 RENAL AUTOREGULATORY MECHANISMS 687. Inscho EW, Cook AK, Imig JD, Vial C, Evans RJ. Renal autoregulation in P2X1 knockout mice. Acta Physiol Scand 181: 445– 453, 2004. 688. Inscho EW, Cook AK, Mui V, Imig JD. Calcium mobilization contributes to pressuremediated afferent arteriolar vasoconstriction. Hypertension 31: 421– 428, 1998. 689. Inscho EW, Cook AK, Murzynowski JB, Imig JD. Elevated arterial pressure impairs autoregulation independently of AT(1) receptor activation. J Hypertens 22: 811– 818, 2004. 709. Iversen BM, Sekse I, Ofstad J. Resetting of renal blood flow autoregulation in spontaneously hypertensive rats. Am J Physiol Renal Fluid Electrolyte Physiol 252: F480 – F486, 1987. 710. Iwai N, Kinoshita M, Shimoike H. Chromosomal mapping of quantitative trait loci that influence renal hemodynamic functions. Circulation 100: 1923–1929, 1999. 711. Iwai N, Kurtz TW, Inagami T. Further evidence of the SA gene as a candidate gene contributing to the hypertension in spontaneously hypertensive rat. Biochem Biophys Res Commun 188: 64 – 69, 1992. 690. Inscho EW, Cook AK, Navar LG. Pressure-mediated vasoconstriction of juxtamedullary afferent arterioles involves P2-purinoceptor activation. Am J Physiol Renal Fluid Electrolyte Physiol 271: F1077–F1085, 1996. 712. Iwai N, Tsujita Y, Kinoshita M. Isolation of a chromosome 1 region that contributes to high blood pressure and salt sensitivity. Hypertension 32: 636 – 638, 1998. 691. Inscho EW, Cook AK, Webb RC, Jin LM. Rho-kinase inhibition reduces pressuremediated autoregulatory adjustments in afferent arteriolar diameter. Am J Physiol Renal Physiol 296: F590 –F597, 2009. 713. Iwamoto T, Kita S, Zhang J, Blaustein MP, Arai Y, Yoshida S, Wakimoto K, Komuro I, Katsuragi T. Salt-sensitive hypertension is triggered by Ca2⫹ entry via Na⫹/Ca2⫹ exchanger type-1 in vascular smooth muscle. Nat Med 10: 1193–1199, 2004. 692. Irzyniec T, Mall G, Greber D, Ritz E. Beneficial effect of nifedipine and moxonidine on glomerulosclerosis in spontaneously hypertensive rats. A micromorphometric study. Am J Hypertens 5: 437– 443, 1992. 714. Izzard AS, Bund SJ, Heagerty AM. Myogenic tone in mesenteric arteries from spontaneously hypertensive rats. Am J Physiol Heart Circ Physiol 270: H1–H6, 1996. 693. Ishinaga Y, Nabika T, Shimada T, Hiraoka J, Nara Y, Yamori Y. Re-evaluation of the SA gene in spontaneously hypertensive and Wistar-Kyoto rats. Clin Exp Pharmacol Physiol 24: 18 –22, 1997. 694. Ito I, Jarajapu YP, Grant MB, Knot HJ. Characteristics of myogenic tone in the rat ophthalmic artery. Am J Physiol Heart Circ Physiol 292: H360 –H368, 2007. 695. Ito S. Characteristics of isolated perfused juxtaglomerular apparatus. Kidney Int Suppl 67: S46 –S48, 1998. 715. Izzard AS, Graham D, Burnham MP, Heerkens EH, Dominiczak AF, Heagerty AM. Myogenic and structural properties of cerebral arteries from the stroke-prone spontaneously hypertensive rat. Am J Physiol Heart Circ Physiol 285: H1489 –H1494, 2003. 716. Jackson EK, Herzer WA. Angiotensin II/prostaglandin I2 interactions in spontaneously hypertensive rats. Hypertension 22: 688 – 698, 1993. 717. Jackson EK, Herzer WA. Defective modulation of angiotensin II-induced renal vasoconstriction in hypertensive rats. Hypertension 23: 329 –336, 1994. 696. Ito S, Carretero OA, Abe K. Role of nitric oxide in the control of glomerular microcirculation. Clin Exp Pharmacol Physiol 24: 578 –581, 1997. 718. Jackson KE, Jackson DW, Quadri S, Reitzell MJ, Navar LG. Inhibition of heme oxygenase augments tubular sodium reabsorption. Am J Physiol Renal Physiol 300: F941– F946, 2011. 697. Ito S, Carretero OA, Abe K, Beierwaltes WH, Yoshinaga K. Effect of prostanoids on renin release from rabbit afferent arterioles with and without macula densa. Kidney Int 35: 1138 –1144, 1989. 719. Jacob F, Clark LA, Guzman PA, Osborn JW. Role of renal nerves in development of hypertension in DOCA-salt model in rats: a telemetric approach. Am J Physiol Heart Circ Physiol 289: H1519 –H1529, 2005. 698. Ito S, Carretero OA, Murray RD. Renin release from isolated afferent arterioles. Kidney Int 27: 762–767, 1985. 720. Jaggar JH, Wellman GC, Heppner TJ, Porter VA, Perez GJ, Gollasch M, Kleppisch T, Rubart M, Stevenson AS, Lederer WJ, Knot HJ, Bonev AD, Nelson MT. Ca2⫹ channels, ryanodine receptors and Ca2⫹-activated K⫹ channels: a functional unit for regulating arterial tone. Acta Physiol Scand 164: 577–587, 1998. 699. Ito S, Johnson CS, Carretero OA. Modulation of angiotensin II-induced vasoconstriction by endothelium-derived relaxing factor in the isolated microperfused rabbit afferent arteriole. J Clin Invest 87: 1656 –1663, 1991. 700. Ito S, Juncos LA, Carretero OA. Pressure-induced constriction of the afferent arteriole of spontaneously hypertensive rats. Hypertension 19: II164 –II167, 1992. 701. Ito S, Juncos LA, Nushiro N, Johnson CS, Carretero OA. Endothelium-derived relaxing factor modulates endothelin action in afferent arterioles. Hypertension 17: 1052–1056, 1991. 702. Ito S, Ren Y. Evidence for the role of nitric oxide in macula densa control of glomerular hemodynamics. J Clin Invest 92: 1093–1098, 1993. 703. Itoh S, Carretero OA. An in vitro approach to the study of macula densa-mediated glomerular hemodynamics. Kidney Int 38: 1206 –1210, 1990. 704. Iversen BM, Amann K, Kvam FI, Wang X, Ofstad J. Increased glomerular capillary pressure and size mediate glomerulosclerosis in SHR juxtamedullary cortex. Am J Physiol Renal Physiol 274: F365–F373, 1998. 705. Iversen BM, Heyeraas KJ, Sekse I, Andersen KJ, Ofstad J. Autoregulation of renal blood flow in two-kidney, one-clip hypertensive rats. Am J Physiol Renal Fluid Electrolyte Physiol 251: F245–F250, 1986. 706. Iversen BM, Kvam FI, Matre K, Morkrid L, Horvei G, Bagchus W, Grond J, Ofstad J. Effect of mesangiolysis on autoregulation of renal blood flow and glomerular filtration rate in rats. Am J Physiol Renal Fluid Electrolyte Physiol 262: F361–F366, 1992. 721. Jaimes EA, Sweeney C, Raij L. Effects of the reactive oxygen species hydrogen peroxide and hypochlorite on endothelial nitric oxide production. Hypertension 38: 877– 883, 2001. 722. Janssen BJ, Malpas SC, Burke SL, Head GA. Frequency-dependent modulation of renal blood flow by renal nerve activity in conscious rabbits. Am J Physiol Regul Integr Comp Physiol 273: R597–R608, 1997. 723. Janssen BJ, Oosting J, Slaaf DW, Persson PB, Struijker-Boudier HA. Hemodynamic basis of oscillations in systemic arterial pressure in conscious rats. Am J Physiol Heart Circ Physiol 269: H62–H71, 1995. 724. Jarajapu YP, Knot HJ. Role of phospholipase C in development of myogenic tone in rat posterior cerebral arteries. Am J Physiol Heart Circ Physiol 283: H2234 –H2238, 2002. 725. Jarajapu YP, Knot HJ. Relative contribution of Rho kinase and protein kinase C to myogenic tone in rat cerebral arteries in hypertension. Am J Physiol Heart Circ Physiol 289: H1917–H1922, 2005. 726. Jensen BL, Ellekvist P, Skott O. Chloride is essential for contraction of afferent arterioles after agonists and potassium. Am J Physiol Renal Physiol 272: F389 –F396, 1997. 727. Jensen BL, Skott O. Blockade of chloride channels by DIDS stimulates renin release and inhibits contraction of afferent arterioles. Am J Physiol Renal Fluid Electrolyte Physiol 270: F718 –F727, 1996. 707. Iversen BM, Kvam FI, Matre K, Ofstad J. Resetting of renal blood autoregulation during acute blood pressure reduction in hypertensive rats. Am J Physiol Regul Integr Comp Physiol 275: R343–R349, 1998. 728. Jernigan NL, Drummond HA. Vascular ENaC proteins are required for renal myogenic constriction. Am J Physiol Renal Physiol 289: F891–F901, 2005. 708. Iversen BM, Kvam FI, Morkrid L, Sekse I, Ofstad J. Effect of cyclooxygenase inhibition on renal blood flow autoregulation in SHR. Am J Physiol Renal Fluid Electrolyte Physiol 263: F534 –F539, 1992. 729. Jernigan NL, Drummond HA. Myogenic vasoconstriction in mouse renal interlobar arteries: role of endogenous beta and gammaENaC. Am J Physiol Renal Physiol 291: F1184 –F1191, 2006. 488 Physiol Rev • VOL 95 • APRIL 2015 • www.prv.org CARLSTRÖM ET AL. 730. Jernigan NL, Lamarca B, Speed J, Galmiche L, Granger JP, Drummond HA. Dietary salt enhances benzamil-sensitive component of myogenic constriction in mesenteric arteries. Am J Physiol Heart Circ Physiol 294: H409 –H420, 2008. 751. Just A, Arendshorst WJ. A novel mechanism of renal blood flow autoregulation and the autoregulatory role of A1 adenosine receptors in mice. Am J Physiol Renal Physiol 293: F1489 –F1500, 2007. 731. Jernigan NL, Speed J, Lamarca B, Granger JP, Drummond HA. Angiotensin II regulation of renal vascular ENaC proteins. Am J Hypertens 22: 593–597, 2009. 752. Just A, Arendshorst WJ. The role of reactive oxygen species in renal blood flow autoregulation (Abstract). FASEB J 22: 761.F1489 –19, 2008. 732. Ji X, Naito Y, Weng H, Endo K, Ma X, Iwai N. P2X7 deficiency attenuates hypertension and renal injury in deoxycorticosterone acetate-salt hypertension. Am J Physiol Renal Physiol 303: F1207–F1215, 2012. 753. Just A, Ehmke H, Toktomambetova L, Kirchheim HR. Dynamic characteristics and underlying mechanisms of renal blood flow autoregulation in the conscious dog. Am J Physiol Renal Physiol 280: F1062–F1071, 2001. 733. Jiang J, Roman RJ. Lovastatin prevents development of hypertension in spontaneously hypertensive rats. Hypertension 30: 968 –974, 1997. 754. Just A, Ehmke H, Wittmann U, Kirchheim HR. Tonic and phasic influences of nitric oxide on renal blood flow autoregulation in conscious dogs. Am J Physiol Renal Physiol 276: F442–F449, 1999. 734. Jin C, Hu C, Polichnowski A, Mori T, Skelton M, Ito S, Cowley AW Jr. Effects of renal perfusion pressure on renal medullary hydrogen peroxide and nitric oxide production. Hypertension 53: 1048 –1053, 2009. 755. Just A, Ehmke H, Wittmann U, Kirchheim HR. Role of angiotensin II in dynamic renal blood flow autoregulation of the conscious dog. J Physiol 538: 167–177, 2002. 735. Jin L, Beswick RA, Yamamoto T, Palmer T, Taylor TA, Pollock JS, Pollock DM, Brands MW, Webb RC. Increased reactive oxygen species contributes to kidney injury in mineralocorticoid hypertensive rats. J Physiol Pharmacol 57: 343–357, 2006. 736. Jin XH, McGrath HE, Gildea JJ, Siragy HM, Felder RA, Carey RM. Renal interstitial guanosine cyclic 3’,5’-monophosphate mediates pressure-natriuresis via protein kinase G. Hypertension 43: 1133–1139, 2004. 756. Just A, Kurtz L, de Wit C, Wagner C, Kurtz A, Arendshorst WJ. Connexin 40 mediates the tubuloglomerular feedback contribution to renal blood flow autoregulation. J Am Soc Nephrol 20: 1577–1585, 2009. 757. Just A, Olson AJ, Falck JR, Arendshorst WJ. NO and NO-independent mechanisms mediate ETB receptor buffering of ET-1-induced renal vasoconstriction in the rat. Am J Physiol Regul Integr Comp Physiol 288: R1168 –R1177, 2005. 737. Jin XH, Siragy HM, Carey RM. Renal interstitial cGMP mediates natriuresis by direct tubule mechanism. Hypertension 38: 309 –316, 2001. 758. Just A, Olson AJ, Whitten CL, Arendshorst WJ. Superoxide mediates acute renal vasoconstriction produced by angiotensin II and catecholamines by a mechanism independent of nitric oxide. Am J Physiol Heart Circ Physiol 292: H83–H92, 2007. 738. Jobs A, Schmidt K, Schmidt VJ, Lubkemeier I, van Veen TA, Kurtz A, Willecke K, de Wit C. Defective Cx40 maintains Cx37 expression but intact Cx40 is crucial for conducted dilations irrespective of hypertension. Hypertension 60: 1422–1429, 2012. 759. Just A, Whitten CL, Arendshorst WJ. Reactive oxygen species participate in acute renal vasoconstrictor responses induced by ETA and ETB receptors. Am J Physiol Renal Physiol 294: F719 –F728, 2008. 739. Johnson PC. The myogenic response. In: Handbook of Physiology: Microcirculation, edited by Tuma R, Duran WN, and Ley N. Boston: Academic, 2008. 760. Just A, Wittmann U, Ehmke H, Kirchheim HR. Autoregulation of renal blood flow in the conscious dog and the contribution of the tubuloglomerular feedback. J Physiol 506: 275–290, 1998. 740. Johnson PC, Intaglietta M. Contributions of pressure and flow sensitivity to autoregulation in mesenteric arterioles. Am J Physiol 231: 1686 –1698, 1976. 741. Johnson RA, Belmonte A, Fan NY, Lavesa M, Nasjletti A, Stier CT Jr. Effect of ifetroban, a thromboxane A2 receptor antagonist, in stroke-prone spontaneously hypertensive rats. Clin Exp Hypertens 18: 171–188, 1996. 742. Johnson RJ, Gordon KL, Giachelli C, Kurth T, Skelton MM, Cowley AW Jr. Tubulointerstitial injury and loss of nitric oxide synthases parallel the development of hypertension in the Dahl-SS rat. J Hypertens 18: 1497–1505, 2000. 743. Johnson RP, El-Yazbi AF, Takeya K, Walsh EJ, Walsh MP, Cole WC. Ca2⫹ sensitization via phosphorylation of myosin phosphatase targeting subunit at threonine-855 by Rho kinase contributes to the arterial myogenic response. J Physiol 587: 2537– 2553, 2009. 744. Jones DR, Dowd DA. Development of elevated blood pressure in young genetically hypertensive rats. Life Sci 9: 247–250, 1970. 745. Jose PA, Slotkoff LM, Montgomery S, Calcagno PL, Eisner G. Autoregulation of renal blood flow in the puppy. Am J Physiol 229: 983–988, 1975. 746. Judy WV, Watanabe AM, Murphy WR, Aprison BS, Yu PL. Sympathetic nerve activity and blood pressure in normotensive backcross rats genetically related to the spontaneously hypertensive rat. Hypertension 1: 598 – 604, 1979. 747. Juncos LA, Garvin J, Carretero OA, Ito S. Flow modulates myogenic responses in isolated microperfused rabbit afferent arterioles via endothelium-derived nitric oxide. J Clin Invest 95: 2741–2748, 1995. 748. Just A. Mechanisms of renal blood flow autoregulation: dynamics and contributions. Am J Physiol Regul Integr Comp Physiol 292: R1–R17, 2007. 749. Just A, Arendshorst WJ. Dynamics and contribution of mechanisms mediating renal blood flow autoregulation. Am J Physiol Regul Integr Comp Physiol 285: R619 –R631, 2003. 750. Just A, Arendshorst WJ. Nitric oxide blunts myogenic autoregulation in rat renal but not skeletal muscle circulation via tubuloglomerular feedback. J Physiol 569: 959 – 974, 2005. 761. Kaiser G, Ackermann R, Sioufi A. Pharmacokinetics of a new angiotensin-converting enzyme inhibitor, benazepril hydrochloride, in special populations. Am Heart J 117: 746 –751, 1989. 762. Kallskog O, Marsh DJ. TGF-initiated vascular interactions between adjacent nephrons in the rat kidney. Am J Physiol Renal Fluid Electrolyte Physiol 259: F60 –F64, 1990. 763. Kaloyanides GJ, Ahrens RE, Shepherd JA, DiBona GF. Inhibition of prostaglandin E2 secretion. Failure to abolish autoregulation in the isolated dog kidney. Circ Res 38: 67–73, 1976. 764. Kaloyanides GJ, Bastron RD, DiBona GF. Impaired autoregulation of blood flow and glomerular filtration rate in the isolated dog kidney depleted of renin. Circ Res 35: 400 – 412, 1974. 765. Kanagawa R, Wada T, Sanada T, Ojima M, Inada Y. Regional hemodynamic effects of candesartan cilexetil (TCV-116), an angiotensin II AT1-receptor antagonist, in conscious spontaneously hypertensive rats. Jpn J Pharmacol 73: 185–190, 1997. 766. Kang J, Kang N, Lovatt D, Torres A, Zhao Z, Lin J, Nedergaard M. Connexin 43 hemichannels are permeable to ATP. J Neurosci 28: 4702– 4711, 2008. 767. Karam H, Clozel JP, Bruneval P, Gonzalez MF, Menard J. Contrasting effects of selective T- and L-type calcium channel blockade on glomerular damage in DOCA hypertensive rats. Hypertension 34: 673– 678, 1999. 768. Karibe A, Watanabe J, Horiguchi S, Takeuchi M, Suzuki S, Funakoshi M, Katoh H, Keitoku M, Satoh S, Shirato K. Role of cytosolic Ca2⫹ and protein kinase C in developing myogenic contraction in isolated rat small arteries. Am J Physiol Heart Circ Physiol 272: H1165–H1172, 1997. 769. Karlsen FM, Andersen CB, Leyssac PP, Holstein-Rathlou NH. Dynamic autoregulation and renal injury in Dahl rats. Hypertension 30: 975–983, 1997. 770. Karlsen FM, Leyssac PP, Holstein-Rathlou NH. Tubuloglomerular feedback in Dahl rats. Am J Physiol Regul Integr Comp Physiol 274: R1561–R1569, 1998. 771. Kashihara T, Nakayama K, Matsuda T, Baba A, Ishikawa T. Role of Na⫹/Ca2⫹ exchanger-mediated Ca2⫹ entry in pressure-induced myogenic constriction in rat posterior cerebral arteries. J Pharmacol Sci 110: 218 –222, 2009. Physiol Rev • VOL 95 • APRIL 2015 • www.prv.org 489 RENAL AUTOREGULATORY MECHANISMS 772. Kasiske BL, Cleary MP, O’Donnell MP, Keane WF. Effects of genetic obesity on renal structure and function in the Zucker rat. J Lab Clin Med 106: 598 – 604, 1985. 792. Khraibi AA, Knox FG. Effect of renal decapsulation on renal interstitial hydrostatic pressure and natriuresis. Am J Physiol Regul Integr Comp Physiol 257: R44 –R48, 1989. 773. Kasiske BL, O’Donnell MP, Keane WF. The Zucker rat model of obesity, insulin resistance, hyperlipidemia, and renal injury. Hypertension 19: I110 –I115, 1992. 793. Kim EC, Ahn DS, Yeon SI, Lim M, Lee YH. Epithelial Na⫹ channel proteins are mechanotransducers of myogenic constriction in rat posterior cerebral arteries. Exp Physiol 97: 544 –555, 2012. 774. Kastner PR, Hall JE, Guyton AC. Control of glomerular filtration rate: role of intrarenally formed angiotensin II. Am J Physiol Renal Fluid Electrolyte Physiol 246: F897– F906, 1984. 775. Katholi RE. Renal nerves in the pathogenesis of hypertension in experimental animals and humans. Am J Physiol Renal Fluid Electrolyte Physiol 245: F1–F14, 1983. 776. Katholi RE, Naftilan AJ, Bishop SP, Oparil S. Role of the renal nerves in the maintenance of DOCA-salt hypertension in the rat: influence on the renal vasculature and sodium excretion. Hypertension 5: 427– 435, 1983. 777. Kauser K, Clark JE, Masters BS, Ortiz de Montellano PR, Ma YH, Harder DR, Roman RJ. Inhibitors of cytochrome P-450 attenuate the myogenic response of dog renal arcuate arteries. Circ Res 68: 1154 –1163, 1991. 778. Kawabata M, Ogawa T, Takabatake T. Control of rat glomerular microcirculation by juxtaglomerular adenosine A1 receptors. Kidney Int Suppl 67: S228 –S230, 1998. 779. Kawabe K, Watanabe TX, Shiono K, Sokabe H. Influence on blood pressure of renal isografts between spontaneously hypertensive and normotensive rats, utilizing the F1 hybrids. Jpn Heart J 19: 886 – 894, 1978. 780. Kawada N, Dennehy K, Solis G, Modlinger P, Hamel R, Kawada JT, Aslam S, MOriyama T, Imai E, Welch WJ, Wilcox CS. TP receptors regulate renal hemodynamics during angiotensin II slow pressor response. Am J Physiol Renal Physiol 287: F753–F759, 2004. 781. Kawata T, Hashimoto S, Koike T. The angiotensin receptor antagonist 2-ethoxy-1 [[2’-(1H-tetrazol-5-yl) biphenyl-4-yl]methyl]-1H-benzimidazole-7-carboxylic acid (CV11974) attenuates the tubuloglomerular feedback response during NO synthase blockade in rats. J Pharmacol Exp Ther 277: 572–577, 1996. 782. Kawata T, Mimuro T, Onuki T, Tsuchiya K, Nihei H, Koike T. The K(ATP) channel opener nicorandil: effect on renal hemodynamics in spontaneously hypertensive and Wistar Kyoto rats. Kidney Int Suppl 67: S231–S233, 1998. 783. Keller M, Lidington D, Vogel L, Peter BF, Sohn HY, Pagano PJ, Pitson S, Spiegel S, Pohl U, Bolz SS. Sphingosine kinase functionally links elevated transmural pressure and increased reactive oxygen species formation in resistance arteries. FASEB J 20: 702–704, 2006. 784. Kelly-Cobbs AI, Elgebaly MM, Li W, Ergul A. Pressure-independent cerebrovascular remodelling and changes in myogenic reactivity in diabetic Goto-Kakizaki rat in response to glycaemic control. Acta Physiol 203: 245–251, 2011. 785. Kelly-Cobbs AI, Prakash R, Coucha M, Knight RA, Li W, Ogbi SN, Johnson M, Ergul A. Cerebral myogenic reactivity and blood flow in type 2 diabetic rats: role of peroxynitrite in hypoxia-mediated loss of myogenic tone. J Pharmacol Exp Ther 342: 407– 415, 2012. 794. Kim EC, Choi SK, Lim M, Yeon SI, Lee YH. Role of endogenous ENaC and TRP channels in the myogenic response of rat posterior cerebral arteries. PLoS One 8: e84194, 2013. 795. Kimmelstiel P, Wilson C. Inflammatory lesions in the glomeruli in pyelonephritis in relation to hypertension and renal insufficiency. Am J Pathol 12: 99 –106, 1936. 796. Kimura K, Nagai R, Sakai T, Aikawa M, Kuro-o M, Kobayashi N, Shirato I, Inagami T, Oshi M, Suzuki N. Diversity and variability of smooth muscle phenotypes of renal arterioles as revealed by myosin isoform expression. Kidney Int 48: 372–382, 1995. 797. Kinoshita Y, Knox FG. Role of prostaglandins in proximal tubule sodium reabsorption: response to elevated renal interstitial hydrostatic pressure. Circ Res 64: 1013– 1018, 1989. 798. Kinoshita Y, Knox FG. Response of superficial proximal convoluted tubule to decreased and increased renal perfusion pressure. In vivo microperfusion study in rats. Circ Res 66: 1184 –1189, 1990. 799. Kirchheim HR, Ehmke H, Hackenthal E, Lowe W, Persson P. Autoregulation of renal blood flow, glomerular filtration rate and renin release in conscious dogs. Pflügers Arch 410: 441– 449, 1987. 800. Kirchner KA. Hypertension in hemodialysis patients: more questions than answers. Am J Kidney Dis 30: 577–578, 1997. 801. Kirk KL, Bell PD, Barfuss DW, Ribadeneira M. Direct visualization of the isolated and perfused macula densa. Am J Physiol Renal Fluid Electrolyte Physiol 248: F890 –F894, 1985. 802. Kirton CA, Loutzenhiser R. Alterations in basal protein kinase C activity modulate renal afferent arteriolar myogenic reactivity. Am J Physiol Heart Circ Physiol 275: H467–H475, 1998. 803. Klahr S, Levey AS, Beck GJ, Caggiula AW, Hunsicker L, Kusek JW, Striker G. The effects of dietary protein restriction and blood-pressure control on the progression of chronic renal disease. Modification of Diet in Renal Disease Study Group. N Engl J Med 330: 877– 884, 1994. 804. Kleinbongard P, Schleiger A, Heusch G. Characterization of vasomotor responses in different vascular territories of C57BL/6J mice. Exp Biol Med 238: 1180 –1191, 2013. 805. Kleinstreuer N, David T, Plank MJ, Endre Z. Dynamic myogenic autoregulation in the rat kidney: a whole-organ model. Am J Physiol Renal Physiol 294: F1453– F1464, 2008. 806. Kline RL. Renal nerves and experimental hypertension: evidence and controversy. Can J Physiol Pharmacol 65: 1540 –1547, 1987. 786. Kett MM, Alcorn D, Bertram JF, Anderson WP. Enalapril does not prevent renal arterial hypertrophy in spontaneously hypertensive rats. Hypertension 25: 335–342, 1995. 807. Kline RL, Kelton PM, Mercer PF. Effect of renal denervation on the development of hypertension in spontaneously hypertensive rats. Can J Physiol Pharmacol 56: 818 – 822, 1978. 787. Kett MM, Bergstrom G, Alcorn D, Bertram JF, Anderson WP. Renal vascular resistance properties and glomerular protection in early established SHR hypertension. J Hypertens 19: 1505–1512, 2001. 808. Kline RL, Liu F. Modification of pressure natriuresis by long-term losartan in spontaneously hypertensive rats. Hypertension 24: 467– 473, 1994. 788. Khan NJ, Hampton JA, Lacher DA, Rapp JP, Gohara AF, Goldblatt PJ. Morphometric evaluation of the renal arterial system of Dahl salt-sensitive and salt-resistant rats on a high salt diet. I. Interlobar and arcuate arteries. Lab Invest 57: 714 –723, 1987. 789. Khraibi AA, Granger JP, Haas JA, Burnett JC Jr, Knox FG. Intrarenal pressures during direct inhibition of sodium transport. Am J Physiol Regul Integr Comp Physiol 263: R1182–R1186, 1992. 790. Khraibi AA, Haas JA, Knox FG. Effect of renal perfusion pressure on renal interstitial hydrostatic pressure in rats. Am J Physiol Renal Fluid Electrolyte Physiol 256: F165– F170, 1989. 791. Khraibi AA, Knox FG. Effect of acute renal decapsulation on pressure natriuresis in SHR and WKY rats. Am J Physiol Renal Fluid Electrolyte Physiol 257: F785–F789, 1989. 490 809. Kline RL, McLennan GP. Chronic hydralazine treatment alters the acute pressurenatriuresis curve in young spontaneously hypertensive rats. Can J Physiol Pharmacol 69: 164 –169, 1991. 810. Knot HJ, Standen NB, Nelson MT. Ryanodine receptors regulate arterial diameter and wall [Ca2⫹] in cerebral arteries of rat via Ca2⫹-dependent K⫹ channels. J Physiol 508: 211–221, 1998. 811. Knudsen T, Elmer H, Knudsen MH, Holstein-Rathlou NH, Stoustrup J. Dynamic modeling of renal blood flow in Dahl hypertensive and normotensive rats. IEEE Trans Biomed Eng 51: 689 – 697, 2004. 812. Kobori H, Nangaku M, Navar LG, Nishiyama A. The intrarenal renin-angiotensin system: from physiology to the pathobiology of hypertension and kidney disease. Pharmacol Rev 59: 251–287, 2007. Physiol Rev • VOL 95 • APRIL 2015 • www.prv.org CARLSTRÖM ET AL. 813. Kobrin I, Pegram BL, Frohlich ED. Acute pressure increase and intrarenal hemodynamics in conscious WKY and SHR rats. Am J Physiol Heart Circ Physiol 249: H1114 – H1118, 1985. 834. Kremer SG, Zeng W, Hurst R, Ning T, Whiteside C, Skorecki KL. Chloride is required for receptor-mediated divalent cation entry in mesangial cells. J Cell Physiol 162: 15–25, 1995. 814. Koch KM, Aynedjian HS, Bank N. Effect of acute hypertension on sodium reabsorption by the proximal tubule. J Clin Invest 47: 1696 –1709, 1968. 835. Kreye VA, Bauer PK, Villhauer I. Evidence for furosemide-sensitive active chloride transport in vascular smooth muscle. Eur J Pharmacol 73: 91–95, 1981. 815. Koeners MP, Racasan S, Koomans HA, Joles JA, Braam B. Nitric oxide, superoxide and renal blood flow autoregulation in SHR after perinatal L-arginine and antioxidants. Acta Physiol 190: 329 –338, 2007. 836. Krishnamoorthy G, Sonkusare SK, Heppner TJ, Nelson MT. Opposing roles of smooth muscle BK channels and ryanodine receptors in the regulation of nerveevoked constriction of mesenteric resistance arteries. Am J Physiol Heart Circ Physiol 306: H981–H988, 2014. 816. Koenig JB, Jaffe IZ. Direct role for smooth muscle cell mineralocorticoid receptors in vascular remodeling: novel mechanisms and clinical implications. Curr Hypertens Rep 16: 427– 0427, 2014. 837. Kriz W. TRPC6: a new podocyte gene involved in focal segmental glomerulosclerosis. Trends Mol Med 11: 527–530, 2005. 817. Koenigsberger M, Sauser R, Beny JL, Meister JJ. Role of the endothelium on arterial vasomotion. Biophys J 88: 3845–3854, 2005. 818. Koenigsberger M, Sauser R, Beny JL, Meister JJ. Effects of arterial wall stress on vasomotion. Biophys J 91: 1663–1674, 2006. 819. Koenigsberger M, Sauser R, Seppey D, Beny JL, Meister JJ. Calcium dynamics and vasomotion in arteries subject to isometric, isobaric, and isotonic conditions. Biophys J 95: 2728 –2738, 2008. 820. Koltsova SV, Kotelevtsev SV, Tremblay J, Hamet P, Orlov SN. Excitation-contraction coupling in resistance mesenteric arteries: evidence for NKCC1-mediated pathway. Biochem Biophys Res Commun 379: 1080 –1083, 2009. 821. Komers R, Anderson S. Paradoxes of nitric oxide in the diabetic kidney. Am J Physiol Renal Physiol 284: F1121–F1137, 2003. 822. Komlosi P, Bell PD, Zhang ZR. Tubuloglomerular feedback mechanisms in nephron segments beyond the macula densa. Curr Opin Nephrol Hypertens 18: 57– 62, 2009. 823. Kompanowska-Jezierska E, Berndt TJ, Knox FG. Prostaglandin E2 concentrations in rat renal cortical and medullary interstitium: effect of volume expansion and renal perfusion pressure. Acta Physiol Scand 172: 287–289, 2001. 824. Kost CK Jr, Herzer WA, Li P, Jackson EK. Vascular reactivity to angiotensin II is selectively enhanced in the kidneys of spontaneously hypertensive rats. J Pharmacol Exp Ther 269: 82– 88, 1994. 825. Kost CK, Li P, Williams DS, Jackson EK. Renal vascular responses to angiotensin II in conscious spontaneously hypertensive and normotensive rats. J Cardiovasc Pharmacol 31: 854 – 861, 1998. 826. Kotchen TA, Piering AW, Cowley AW, Grim CE, Gaudet D, Hamet P, Kaldunski ML, Kotchen JM, Roman RJ. Glomerular hyperfiltration in hypertensive African Americans. Hypertension 35: 822– 826, 2000. 827. Kotecha N, Hill MA. Myogenic contraction in rat skeletal muscle arterioles: smooth muscle membrane potential and Ca2⫹ signaling. Am J Physiol Heart Circ Physiol 289: H1326 –H1334, 2005. 828. Kovacs G, Komlosi P, Fuson A, Peti-Peterdi J, Rosivall L, Bell PD. Neuronal nitric oxide synthase: its role and regulation in macula densa cells. J Am Soc Nephrol 14: 2475–2483, 2003. 838. Kriz W, Hartmann I, Hosser H, Hahnel B, Kranzlin B, Provoost A, Gretz N. Tracer studies in the rat demonstrate misdirected filtration and peritubular filtrate spreading in nephrons with segmental glomerulosclerosis. J Am Soc Nephrol 12: 496 –506, 2001. 839. Kriz W, Hosser H, Hahnel B, Gretz N, Provoost AP. From segmental glomerulosclerosis to total nephron degeneration and interstitial fibrosis: a histopathological study in rat models and human glomerulopathies. Nephrol Dial Transplant 13: 2781– 2798, 1998. 840. Kriz W, Hosser H, Hahnel B, Simons JL, Provoost AP. Development of vascular pole-associated glomerulosclerosis in the Fawn-hooded rat. J Am Soc Nephrol 9: 381–396, 1998. 841. Kuijpers MH, de Jong W. Spontaneous hypertension in the fawn-hooded rat: a cardiovascular disease model. J Hypertens Suppl 4: S41–S44, 1986. 842. Kuijpers MH, Gruys E. Spontaneous hypertension and hypertensive renal disease in the fawn-hooded rat. Br J Exp Pathol 65: 181–190, 1984. 843. Kuijpers MH, Provoost AP, de Jong W. Development of hypertension and proteinuria with age in fawn-hooded rats. Clin Exp Pharmacol Physiol 13: 201–209, 1986. 844. Kunau RT Jr, Lameire NH. The effect of an acute increase in renal perfusion pressure on sodium transport in the rat kidney. Circ Res 39: 689 – 695, 1976. 845. Kur J, Bankhead P, Scholfield CN, Curtis TM, McGeown JG. Ca2⫹ sparks promote myogenic tone in retinal arterioles. Br J Pharmacol 168: 1675–1686, 2013. 846. Kurtz A. Renin release: sites, mechanisms, control. Annu Rev Physiol 73: 377–399, 2011. 847. Kurtz A, Penner R. Angiotensin II induces oscillations of intracellular calcium and blocks anomalous inward rectifying potassium current in mouse renal juxtaglomerular cells. Proc Natl Acad Sci USA 86: 3423–3427, 1989. 848. Kurtz TW, Montano M, Chan L, Kabra P. Molecular evidence of genetic heterogeneity in Wistar-Kyoto rats: implications for research with the spontaneously hypertensive rat. Hypertension 13: 188 –192, 1989. 849. Kurtz TW, Morris RC Jr. Biological variability in Wistar-Kyoto rats. Implications for research with the spontaneously hypertensive rat. Hypertension 10: 127–131, 1987. 850. Kurtz TW, Simonet L, Kabra PM, Wolfe S, Chan L, Hjelle BL. Cosegregation of the renin allele of the spontaneously hypertensive rat with an increase in blood pressure. J Clin Invest 85: 1328 –1332, 1990. 829. Kovacs G, Peti-Peterdi J, Rosivall L, Bell PD. Angiotensin II directly stimulates macula densa Na-2Cl-K cotransport via apical AT(1) receptors. Am J Physiol Renal Physiol 282: F301–F306, 2002. 851. Kusche-Vihrog K, Tarjus A, Fels J, Jaisser F. The epithelial Na⫹ channel: a new player in the vasculature. Curr Opin Nephrol Hypertens 23: 143–148, 2014. 830. Kozma F, Johnson RA, Zhang F, Yu C, Tong X, Nasjletti A. Contribution of endogenous carbon monoxide to regulation of diameter in resistance vessels. Am J Physiol Regul Integr Comp Physiol 276: R1–R8, 1999. 852. Kvam FI, Ofstad J, Iversen BM. Effects of antihypertensive drugs on autoregulation of RBF and glomerular capillary pressure in SHR. Am J Physiol Renal Physiol 275: F576 – F584, 1998. 831. Kramp RA, Fourmanoir P, Caron N. Endothelin resets renal blood flow autoregulatory efficiency during acute blockade of NO in the rat. Am J Physiol Renal Physiol 281: F1132–F1140, 2001. 853. Kvam FI, Ofstad J, Iversen BM. Role of nitric oxide in the autoregulation of renal blood flow and glomerular filtration rate in aging spontaneously hypertensive rats. Kidney Blood Press Res 23: 376 –384, 2000. 832. Kramp RA, Fourmanoir P, Ladriere L, Joly E, Gerbaux C, El HA, Caron N. Effects of Ca2⫹ channel activity on renal hemodynamics during acute attenuation of NO synthesis in the rat. Am J Physiol Renal Physiol 278: F561–F569, 2000. 854. Kvandal P, Landsverk SA, Bernjak A, Stefanovska A, Kvernmo HD, Kirkeboen KA. Low-frequency oscillations of the laser Doppler perfusion signal in human skin. Microvasc Res 72: 120 –127, 2006. 833. Kramp RA, Genard J, Fourmanoir P, Caron N, Laekeman G, Herman A. Renal hemodynamics and blood flow autoregulation during acute cyclooxygenase inhibition in male rats. Am J Physiol Renal Fluid Electrolyte Physiol 268: F468 –F479, 1995. 855. Lagaud GJ, Gaudreault N, Moore ED, Van Breemen C, Laher I. Pressure-dependent myogenic constriction of cerebral arteries occurs independently of voltage-dependent activation. Am J Physiol Heart Circ Physiol 283: H2187–H2195, 2002. Physiol Rev • VOL 95 • APRIL 2015 • www.prv.org 491 RENAL AUTOREGULATORY MECHANISMS 856. Lagaud GJ, Karicheti V, Knot HJ, Christ GJ, Laher I. Inhibitors of gap junctions attenuate myogenic tone in cerebral arteries. Am J Physiol Heart Circ Physiol 283: H2177–H2186, 2002. 877. Layton AT, Bowen M, Wen A, Layton HE. Feedback-mediated dynamics in a model of coupled nephrons with compliant thick ascending limbs. Math Biosci 230: 115– 127, 2011. 857. Lagaud GJ, Lam E, Lui A, Van Breemen C, Laher I. Nonspecific inhibition of myogenic tone by PD98059, a MEK1 inhibitor, in rat middle cerebral arteries. Biochem Biophys Res Commun 257: 523–527, 1999. 878. Le Cras TD, Kim DH, Markham NE, Abman AS. Early abnormalities of pulmonary vascular development in the Fawn-Hooded rat raised at Denver’s altitude. Am J Physiol Lung Cell Mol Physiol 279: L283–L291, 2000. 858. Lagaud GJ, Masih-Khan E, Kai S, Van Breemen C, Dube GP. Influence of type II diabetes on arterial tone and endothelial function in murine mesenteric resistance arteries. J Vasc Res 38: 578 –589, 2001. 879. Le Gal L, Alonso F, Wagner C, Germain S, Haefliger DN, Meda P, Haefliger JA. Restoration of connexin 40 (Cx40) in renin-producing cells reduces the hypertension of Cx40 null mice. Hypertension 63: 1198 –1204, 2014. 859. Lagaud GJ, Skarsgard PL, Laher I, Van Breemen C. Heterogeneity of endotheliumdependent vasodilation in pressurized cerebral and small mesenteric resistance arteries of the rat. J Pharmacol Exp Ther 290: 832– 839, 1999. 880. Ledingham JM, Simpson FO, Hamada M. Salt appetite, body sodium, handling of a NaCl load, renin, and aldosterone in genetically and spontaneously hypertensive rats. J Cardiovasc Pharmacol 16 Suppl 7: S6 –S8, 1990. 860. Lai EY, Luo Z, Onozato ML, Rudolph EH, Solis G, Jose PA, Wellstein A, Aslam S, Quinn MT, Griendling K, Le T, Li P, Palm F, Welch WJ, Wilcox CS. Effects of the antioxidant drug tempol on renal oxygenation in mice with reduced renal mass. Am J Physiol Renal Physiol 303: F64 –F74, 2012. 881. Leffler CW, Parfenova H, Jaggar JH, Wang R. Carbon monoxide and hydrogen sulfide: gaseous messengers in cerebrovascular circulation. J Appl Physiol 100: 1065– 1076, 2006. 861. Lai EY, Onozato ML, Solis G, Aslam S, Welch WJ, Wilcox CS. Myogenic responses of mouse isolated perfused renal afferent arterioles: effects of salt intake and reduced renal mass. Hypertension 55: 983–989, 2010. 882. Leh S, Hultstrom M, Rosenberger C, Iversen BM. Afferent arteriolopathy and glomerular collapse but not segmental sclerosis induce tubular atrophy in old spontaneously hypertensive rats. Virchows Arch 459: 99 –108, 2011. 862. Lai EY, Patzak A, Persson AE, Carlstrom M. Angiotensin II enhances the afferent arteriolar response to adenosine through increases in cytosolic calcium. Acta Physiol 196: 435– 445, 2009. 883. Leong CL, Anderson WP, O’Connor PM, Evans RG. Evidence that renal arterialvenous oxygen shunting contributes to dynamic regulation of renal oxygenation. Am J Physiol Renal Physiol 292: F1726 –F1733, 2007. 863. Lai EY, Patzak A, Steege A, Mrowka R, Brown R, Spielmann N, Persson PB, Fredholm BB, Persson AE. Contribution of adenosine receptors in the control of arteriolar tone and adenosine-angiotensin II interaction. Kidney Int 70: 690 – 698, 2006. 884. Leong PKK, Zhang Y, Yang LE, Holstein-Rathlou NH, McDonough AA. Diuretic response to acute hypertension is blunted during angiotensin II clamp. Am J Physiol Regul Integr Comp Physiol 283: R837–R842, 2002. 864. Lai EY, Solis G, Holland S, Luo Z, Carlstrom M, Sandberg K, Wellstein A, Welch WJ, Wilcox CS. P47phox is required for renal and afferent arteriolar contractile responses to angiotensin II and perfusion pressure in mice. Hypertension 59: 415– 420, 2012. 885. Lesniewski LA, Donato AJ, Behnke BJ, Woodman CR, Laughlin MH, Ray CA, Delp MD. Decreased NO signaling leads to enhanced vasoconstrictor responsiveness in skeletal muscle arterioles of the ZDF rat prior to overt diabetes and hypertension. Am J Physiol Heart Circ Physiol 294: H1840 –H1850, 2008. 865. Lai EY, Wang Y, Persson AE, Manning D, Liu R. Pressure induces intracellular calcium changes in juxtaglomerular cells in perfused afferent arterioles. Hypertension Res 34: 942–948, 2011. 866. Lai EY, Wellstein A, Welch WJ, Wilcox CS. Superoxide modulates myogenic contractions of mouse afferent arterioles. Hypertension 58: 650 – 656, 2011. 867. Lais LT, Rios LL, Boutelle S, DiBona GF, Brody MJ. Arterial pressure development in neonatal and young spontaneously hypertensive rats. Blood Vessels 14: 277–284, 1977. 868. Lamboley M, Schuster A, Beny JL, Meister JJ. Recruitment of smooth muscle cells and arterial vasomotion. Am J Physiol Heart Circ Physiol 285: H562–H569, 2003. 869. Landsverk SA, Kvandal P, Bernjak A, Stefanovska A, Kirkeboen KA. The effects of general anesthesia on human skin microcirculation evaluated by wavelet transform. Anesth Analg 105: 1012–1019, 2007. 870. Langston JB, Guyton AC, Gillespie WJ Jr. Acute effect of changes in renal arterial pressure and sympathetic blockade on kidney function. Am J Physiol 197: 595– 600, 1959. 871. Langston JB, Guyton AC, Gillespie WJ Jr. Autoregulation absent in normal kidney but present after renal damage. Am J Physiol 199: 495– 498, 1960. 872. Larson TS, Lockhart JC. Restoration of vasa recta hemodynamics and pressure natriuresis in SHR by L-arginine. Am J Physiol Renal Fluid Electrolyte Physiol 268: F907–F912, 1995. 873. Lassegue B, San MA, Griendling KK. Biochemistry, physiology, and pathophysiology of NADPH oxidases in the cardiovascular system. Circ Res 110: 1364 –1390, 2012. 886. Lessard A, Salevsky FC, Bachelard H, Cupples WA. Incommensurate frequencies of major vascular regulatory mechanisms. Can J Physiol Pharmacol 77: 293–299, 1999. 887. Levine DZ, Iacovitti M. Real-time measurement of kidney tubule fluid nitric oxide concentrations in early diabetes: disparate changes in different rodent models. Nitric Oxide 15: 87–92, 2006. 888. Levine DZ, Iacovitti M, Robertson SJ. Modulation of single-nephron GFR in the db/db mouse model of type 2 diabetes mellitus. II. Effects of renal mass reduction. Am J Physiol Regul Integr Comp Physiol 294: R1840 –R1846, 2008. 889. Levine DZ, Iacovitti M, Robertson SJ, Mokhtar GA. Modulation of single-nephron GFR in the db/db mouse model of type 2 diabetes mellitus. Am J Physiol Regul Integr Comp Physiol 290: R975–R981, 2006. 890. Levy D, Anderson KM, Christiansen JC, Campanile G, Stokes J. Antihypertensive drug therapy and arrhythmia risk. Am J Cardiol 62: 147–149, 1988. 891. Lewis EJ, Hunsicker LG, Clark WR, Berl T, Pohl MA, Lewis JB, Ritz E, Atkins RC, Rohde R, Raz I. Renoprotective effect of the angiotensin-receptor antagonist irbesartan in patients with nephropathy due to type 2 diabetes. N Engl J Med 345: 851– 860, 2001. 892. Leyssac PP. Further studies on oscillating tubulo-glomerular feedback responses in the rat kidney. Acta Physiol Scand 126: 271–277, 1986. 893. Leyssac PP, Baumbach L. An oscillating intratubular pressure response to alterations in Henle loop flow in the rat kidney. Acta Physiol Scand 117: 415– 419, 1983. 874. Lau C, Sudbury I, Thomson M, Howard PL, Magil AB, Cupples WA. Salt-resistant blood pressure and salt-sensitive renal autoregulation in chronic streptozotocin diabetes. Am J Physiol Regul Integr Comp Physiol 296: R1761–R1770, 2009. 894. Leyssac PP, Holstein-Rathlou NH. Tubulo-glomerular feedback response: enhancement in adult spontaneously hypertensive rats and effects of anaesthetics. Pflügers Arch 413: 267–272, 1989. 875. Laugesen JL, Sosnovtseva OV, Mosekilde E, Holstein-Rathlou NH, Marsh DJ. Coupling-induced complexity in nephron models of renal blood flow regulation. Am J Physiol Regul Integr Comp Physiol 298: R997–R1006, 2010. 895. Li JM, Shah AM. Endothelial cell superoxide generation: regulation and relevance for cardiovascular pathophysiology. Am J Physiol Regul Integr Comp Physiol 287: R1014 – R1030, 2004. 876. Lax DS, Benstein JA, Tolbert E, Dworkin LD. Effects of salt restriction on renal growth and glomerular injury in rats with remnant kidneys. Kidney Int 41: 1527– 1534, 1992. 896. Li L, Lai EY, Huang Y, Eisner C, Mizel D, Wilcox CS, Schnermann J. Renal afferent arteriolar and tubuloglomerular feedback reactivity in mice with conditional deletions of adenosine 1 receptors. Am J Physiol Renal Physiol 303: F1166 –F1175, 2012. 492 Physiol Rev • VOL 95 • APRIL 2015 • www.prv.org CARLSTRÖM ET AL. 897. Li N, Yi F, Dos Santos EA, Donley DK, Li PL. Role of renal medullary heme oxygenase in the regulation of pressure natriuresis and arterial blood pressure. Hypertension 49: 148 –154, 2007. 916. Llamas B, Lau C, Cupples WA, Rainville ML, Souzeau E, Deschepper CF. Genetic determinants of systolic and pulse pressure in an intercross between normotensive inbred rats. Hypertension 48: 921–926, 2006. 898. Li N, Zou AP, Ge ZD, Campbell WB, Li PL. Effect of nitric oxide on calcium-induced calcium release in coronary arterial smooth muscle. Gen Pharmacol 35: 37– 45, 2000. 917. Lo M, Liu KL, Clemitson JR, Sassard J, Samani NJ. Chromosome 1 blood pressure QTL region influences renal function curve and salt sensitivity in SHR. Physiol Genomics 8: 15–21, 2002. 899. Li S, Moon JJ, Miao H, Jin G, Chen BP, Yuan S, Hu Y, Usami S, Chien S. Signal transduction in matrix contraction and the migration of vascular smooth muscle cells in three-dimensional matrix. J Vasc Res 40: 378 –388, 2003. 918. Lo M, Liu KL, Lantelme P, Sassard J. Subtype 2 of angiotensin II receptors controls pressure-natriuresis in rats. J Clin Invest 95: 1394 –1397, 1995. 900. Lieb DC, Kemp BA, Howell NL, Gildea JJ, Carey RM. Reinforcing feedback loop of renal cyclic guanosine 3’,5’-monophosphate and interstitial hydrostatic pressure in pressure-natriuresis. Hypertension 54: 1278 –1283, 2009. 919. Lohn M, Jessner W, Furstenau M, Wellner M, Sorrentino V, Haller H, Luft FC, Gollasch M. Regulation of calcium sparks and spontaneous transient outward currents by RyR3 in arterial vascular smooth muscle cells. Circ Res 89: 1051–1057, 2001. 901. Liebau G, Levine DZ, Thurau K. Micropuncture studies on the dog kidney. I. The response of the proximal tubule to changes in systemic blood pressure within and below the autoregulatory range. Pflügers Arch 304: 57– 68, 1968. 920. Lombard JH, Eskinder H, Kauser K, Osborn JL, Harder DR. Enhanced norepinephrine sensitivity in renal arteries at elevated transmural pressure. Am J Physiol Heart Circ Physiol 259: H29 –H33, 1990. 902. Lin HB, Young DB. Verapamil alters the relationship between renal perfusion pressure and glomerular filtration rate and renin release: the mechanism of the antihypertensive effect. J Cardiovasc Pharmacol 6: S57–S59, 1988. 921. Lopez B, Moreno C, Salom MG, Roman RJ, Fenoy FJ. Role of guanylyl cyclase and cytochrome P-450 on renal response to nitric oxide. Am J Physiol Renal Physiol 281: F420 –F427, 2001. 903. Lin JJ, Tonshoff B, Bouriquet N, Casellas D, Kaskel FJ, Moore LC. Insulin-like growth factor-I restores microvascular autoregulation in experimental chronic renal failure. Kidney Int Suppl 67: S195–S198, 1998. 922. Lopez B, Ryan RP, Moreno C, Sarkis A, Lazar J, Provoost AP, Jacob HJ, Roman RJ. Identification of a QTL on chromosome 1 for impaired autoregulation of RBF in fawn-hooded hypertensive rats. Am J Physiol Renal Physiol 290: F1213–F1221, 2006. 904. Lincoln TM, Cornwell TL. Intracellular cyclic GMP receptor proteins. FASEB J 7: 328 –338, 1993. 923. Lorenz JN. Micropuncture of the kidney: a primer on techniques. Comp Physiol 2: 621– 637, 2012. 905. Linde CI, Karashima E, Raina H, Zulian A, Wier WG, Hamlyn JM, Ferrari P, Blaustein MP, Golovina VA. Increased arterial smooth muscle Ca2⫹ signaling, vasoconstriction, and myogenic reactivity in Milan hypertensive rats. Am J Physiol Heart Circ Physiol 302: H611–H620, 2012. 924. Lorenz JN, Schnermann J, Brosius FC, Briggs JP, Furspan PB. Intracellular ATP can regulate afferent arteriolar tone via ATP-sensitive K⫹ channels in the rabbit. J Clin Invest 90: 733–740, 1992. 906. Lindpaintner K, Hilbert P, Ganten D, Nadal-Ginard B, Inagami T, Iwai N. Molecular genetics of the SA-gene: cosegregation with hypertension and mapping to rat chromosome 1. J Hypertens 11: 19 –23, 1993. 925. Loutzenhiser K, Loutzenhiser R. Angiotensin II-induced Ca2⫹ influx in renal afferent and efferent arterioles: differing roles of voltage-gated and store-operated Ca2⫹ entry. Circ Res 87: 551–557, 2000. 907. Lino S, Calcagnini G, Censi F, Congi M, De PF. Cardiovascular neuroregulation and rhythms of the autonomic nervous system: frequency domain analysis (Article in Italian)Cardiologia 44: 281–287, 1999. 926. Loutzenhiser R, Aaronson PI. Role of epithelial sodium channels in the renal myogenic response? Hypertension 55: -e6 – e9, 2010. 908. Linz W, Becker RH, Scholkens BA, Wiemer G, Keil M, Langer KH. Nephroprotection by long-term ACE inhibition with ramipril in spontaneously hypertensive stroke prone rats. Kindey Int 54: 2037–2044, 1998. 909. Lipkowitz MS, Freedman BI, Langefeld CD, Comeau ME, Bowden DW, Kao WH, Astor BC, Bottinger EP, Iyengar SK, Klotman PE, Freedman RG, Zhang W, Parekh RS, Choi MJ, Nelson GW, Winkler CA, Kopp JB. Apolipoprotein L1 gene variants associate with hypertension-attributed nephropathy and the rate of kidney function decline in African Americans. Kidney Int 83: 114 –120, 2013. 910. Litteri G, Carnevale D, D’Urso A, Cifelli G, Braghetta P, Damato A, Bizzotto D, Landolfi A, Ros FD, Sabatelli P, Facchinello N, Maffei A, Volpin D, Colombatti A, Bressan GM, Lembo G. Vascular smooth muscle Emilin-1 is a regulator of arteriolar myogenic response and blood pressure. Arterioscler Thromb Vasc Biol 32: 2178 – 2184, 2012. 927. Loutzenhiser R, Bidani AK, Chilton L. Renal myogenic response: kinetic attributes and physiological role. Circ Res 90: 1316 –1324, 2002. 928. Loutzenhiser R, Chilton L, Trottier G. Membrane potential measurements in renal afferent and efferent arterioles: actions of angiotensin II. Am J Physiol Renal Physiol 273: F307–F314, 1997. 929. Loutzenhiser R, Epstein M. Effects of calcium antagonists on renal hemodynamics. Am J Physiol Renal Fluid Electrolyte Physiol 249: F619 –F629, 1985. 930. Loutzenhiser R, Epstein M, Hayashi K, Horton C. Direct visualization of effects of endothelin on the renal microvasculature. Am J Physiol Renal Fluid Electrolyte Physiol 258: F61–F68, 1990. 931. Loutzenhiser R, Epstein M, Horton C. Inhibition by diltiazem of pressure-induced afferent vasoconstriction in the isolated perfused rat kidney. Am J Cardiol 59: -72A– 75A, 1987. 911. Liu KL, Lo M, Grouzmann E, Mutter M, Sassard J. The subtype 2 of angiotensin II receptors and pressure-natriuresis in adult rat kidneys. Br J Pharmacol 126: 826 – 832, 1999. 932. Loutzenhiser R, Epstein M, Horton C, Sonke P. Reversal of renal and smooth muscle actions of the thromboxane mimetic U-44069 by diltiazem. Am J Physiol Renal Fluid Electrolyte Physiol 250: F619 –F626, 1986. 912. Liu R, Garvin JL, Ren Y, Pagano PJ, Carretero OA. Depolarization of the macula densa induces superoxide production via NAD(P)H oxidase. Am J Physiol Renal Physiol 292: F1867–F1872, 2007. 933. Loutzenhiser R, Griffin K, Williamson G, Bidani A. Renal autoregulation: new perspectives regarding the protective and regulatory roles of the underlying mechanisms. Am J Physiol Regul Integr Comp Physiol 290: R1153–R1167, 2006. 913. Liu R, Ren Y, Garvin JL, Carretero OA. Superoxide enhances tubuloglomerular feedback by constricting the afferent arteriole. Kidney Int 66: 268 –274, 2004. 934. Loutzenhiser R, Griffin KA, Bidani AK. Systolic blood pressure as the trigger for the renal myogenic response: protective or autoregulatory? Curr Opin Nephrol Hypertens 15: 41– 49, 2006. 914. Liu Y, Bubolz AH, Mendoza S, Zhang DX, Gutterman DD. H2O2 is the transferrable factor mediating flow-induced dilation in human coronary arterioles. Circ Res 108: 566 –573, 2011. 915. Liu Y, Harder D, Lombard J. Myogenic activation of canine small renal arteries after nonchemical removal of the endothelium. Am J Physiol Heart Circ Physiol 267: H302– H307, 1994. 935. Loutzenhiser R, Hayashi K, Epstein M. Divergent effects of KCl induced depolarization on afferent and efferent arterioles. Am J Physiol Renal Fluid Electrolyte Physiol 257: F561–F564, 1989. 936. Loutzenhiser R, Parker MJ. Hypoxia inhibits myogenic reactivity of renal afferent arterioles by activating ATP-sensitive K⫹ channels. Circ Res 74: 861– 869, 1994. Physiol Rev • VOL 95 • APRIL 2015 • www.prv.org 493 RENAL AUTOREGULATORY MECHANISMS 937. Luksha N, Luksha L, Carrero JJ, Hammarqvist F, Stenvinkel P, Kublickiene K. Impaired resistance artery function in patients with end-stage renal disease. Clin Sci 120: 525–536, 2011. 938. Lund-Johansen P. Hemodynamic changes in long-term diuretic therapy of essential hypertension. Acta Med Scand 187: 509 –518, 1970. 939. Lundie MJ, Friberg P, Kline RL, Adams MA. Long-term inhibition of the renin-angiotensin system in genetic hypertension: analysis of the impact on blood pressure and cardiovascular structural changes. J Hypertens 15: 339 –348, 1997. 940. Lundin S, Herlitz H, Hallback-Nordlander M, Ricksten SE, Gothberg G, Berglund G. Sodium balance during development of hypertension in the spontaneously hypertensive rat (SHR). Acta Physiol Scand 115: 317–323, 1982. 941. Lundin S, Rickesten SE, Thoren P. Renal sympathetic activity in spontaneously hypertensive rats with normotensive controls as studied by three different methods. Acta Physiol Scand 120: 265–272, 1984. 959. Majid DSA, Omoro SA, Chin SY, Navar LG. Intrarenal nitric oxide activity and pressure natriuresis in anesthetized dogs. Hypertension 32: 262–272, 1998. 960. Majid DSA, Said KE, Omoro SA. Responses to acute changes in arterial pressure on renal medullary nitric oxide activity in dogs. Hypertension 34: 832– 836, 1999. 961. Majid DSA, Said KE, Omoro SA, Navar LG. Nitric oxide dependency of arterial pressure-induced changes in renal interstitial hydrostatic pressure in dogs. Circ Res 88: 347–351, 2001. 962. Majid DSA, Williams A, Kadowitz PJ, Navar LG. Renal responses to intra-arterial administration of nitric oxide donor in dogs. Hypertension 22: 535–541, 1993. 963. Majid DSA, Williams A, Navar LG. Inhibition of nitric oxide synthesis attenuates pressure-induced natriuretic responses in anesthetized dogs. Am J Physiol Renal Fluid Electrolyte Physiol 264: F79 –F87, 1993. 964. Makino A, Skelton MM, Zou AP, Roman RJ, Cowley AW Jr. Increased renal medullary oxidative stress produces hypertension. Hypertension 39: 667– 672, 2002. 942. Luscher TF, Creager MA, Beckman JA, Cosentino F. Diabetes and vascular disease: pathophysiology, clinical consequences, and medical therapy: Part II. Circulation 108: 1655–1661, 2003. 965. Mann JF, Green D, Jamerson K, Ruilope LM, Kuranoff SJ, Littke T, Viberti G. Avosentan for overt diabetic nephropathy. J Am Soc Nephrol 21: 527–535, 2010. 943. Ma YH, Gebremedhin D, Schwartzman ML, Falck JR, Clark JE, Masters BS, Harder DR, Roman RJ. 20-Hydroxyeicosatetraenoic acid is an endogenous vasoconstrictor of canine renal arcuate arteries. Circ Res 72: 126 –136, 1993. 966. Mann JF, Schmieder RE, Dyal L, McQueen MJ, Schumacher H, Pogue J, Wang X, Probstfield JL, Avezum A, Cardona-Munoz E, Dagenais GR, Diaz R, Fodor G, Maillon JM, Ryden L, Yu CM, Teo KK, Yusuf S. Effect of telmisartan on renal outcomes: a randomized trial. Ann Intern Med 151: 1–10, 2009. 944. Macedo MP, Lautt WW. Autoregulatory capacity in the superior mesenteric artery is attenuated by nitric oxide. Am J Physiol Gastrointest Liver Physiol 271: G400 –G404, 1996. 945. Maddox DA, Troy JL, Brenner BM. Autoregulation of filtration rate in the absence of macula densa-glomerulus feedback. Am J Physiol 227: 123–131, 1974. 946. Madrid MI, Garcia-Salom M, Tornel J, de Gasparo M, Fenoy FJ. Effect of interactions between nitric oxide and angiotensin II on pressure diuresis and natriuresis. Am J Physiol Regul Integr Comp Physiol 273: R1676 –R1682, 1997. 947. Madrid MI, Salom MG, Tornel J, Lopez E, Fenoy FJ. Interactions between nitric oxide and renal nerves on pressure-diuresis and natriuresis. J Am Soc Nephrol 9: 1588 – 1595, 1998. 948. Magee GM, Bilous RW, Cardwell CR, Hunter SJ, Kee F, Fogarty DG. Is hyperfiltration associated with the future risk of developing diabetic nephropathy? A metaanalysis. Diabetologia 52: 691– 697, 2009. 949. Magnusson L, Sorensen CM, Braunstein TH, Holstein-Rathlou NH, Salomonsson M. Renovascular BK(Ca) channels are not activated in vivo under resting conditions and during agonist stimulation. Am J Physiol Regul Integr Comp Physiol 292: R345–R353, 2007. 950. Majid DS, Godfrey M, Navar LG. Pressure natriuresis and renal medullary blood flow in dogs. Hypertension 29: 1051–1057, 1997. 951. Majid DS, Godfrey M, Omoro SA. Pressure natriuresis and autoregulation of inner medullary blood flow in canine kidney. Hypertension 29: 210 –215, 1997. 952. Majid DS, Inscho EW, Navar LG. P2 purinoceptor saturation by adenosine triphosphate impairs renal autoregulation in dogs. J Am Soc Nephrol 10: 492– 498, 1999. 953. Majid DS, Navar LG. Suppression of blood flow autoregulation plateau during nitric oxide blockade of canine kidney. Am J Physiol Renal Fluid Electrolyte Physiol 262: F40 –F46, 1992. 954. Majid DS, Navar LG. Medullary blood flow responses to changes in arterial pressure in canine kidney. Am J Physiol Renal Fluid Electrolyte Physiol 270: F833–F838, 1996. 955. Majid DS, Navar LG. Nitric oxide in the control of renal hemodynamics and excretory function. Am J Hypertens 14: 74S– 82S, 2001. 967. Mann JF, Schmieder RE, McQueen M, Dyal L, Schumacher H, Pogue J, Wang X, Maggioni A, Budaj A, Chaithiraphan S, Dickstein K, Keltai M, Metsarinne K, Oto A, Parkhomenko A, Piegas LS, Svendsen TL, Teo KK, Yusuf S. Renal outcomes with telmisartan, ramipril, or both, in people at high vascular risk (the ONTARGET study): a multicentre, randomised, double-blind, controlled trial. Lancet 372: 547– 553, 2008. 968. Manning RD Jr, Tian N, Meng S. Oxidative stress and antioxidant treatment in hypertension and the associated renal damage. Am J Nephrol 25: 311–317, 2005. 969. Marchand GR, Burke TJ, Haas JA, Romero JC, Knox FG. Regulation of filtration rate in sodium-depleted and -expanded dogs. Am J Physiol Renal Fluid Electrolyte Physiol 232: F325–F328, 1977. 970. Marchenko SM, Sage SO. Smooth muscle cells affect endothelial membrane potential in rat aorta. Am J Physiol Heart Circ Physiol 267: H804 –H811, 1994. 971. Marsh DJ. Frequency response of autoregulation. Kidney Int 22: S165–S172, 1982. 972. Marsh DJ, Martin CM. Effects of diuretic states on collecting duct fluid flow resistance in the hamster kidney. Am J Physiol 229: 13–17, 1975. 973. Marsh DJ, Sosnovtseva OV, Chon KH, Holstein-Rathlou NH. Nonlinear interactions in renal blood flow regulation. Am J Physiol Regul Integr Comp Physiol 288: R1143– R1159, 2005. 974. Marsh DJ, Sosnovtseva OV, Mosekilde E, Holstein-Rathlou NH. Vascular coupling induces synchronization, quasiperiodicity, and chaos in a nephron tree. Chaos 17: 015114, 2007. 975. Marsh DJ, Sosnovtseva OV, Pavlov AN, Yip KP, Holstein-Rathlou NH. Frequency encoding in renal blood flow regulation. Am J Physiol Regul Integr Comp Physiol 288: R1160 –R1167, 2005. 976. Marsh DJ, Toma I, Sosnovtseva OV, Peti-Peterdi J, Holstein-Rathlou NH. Electrotonic vascular signal conduction and nephron synchronization. Am J Physiol Renal Physiol 296: F751–F761, 2009. 977. Massett MP, Ungvari Z, Csiszar A, Kaley G, Koller A. Different roles of PKC and MAP kinases in arteriolar constrictions to pressure and agonists. Am J Physiol Heart Circ Physiol 283: H2282–H2287, 2002. 956. Majid DSA, Navar LG. Blockade of distal nephron sodium transport attenuates pressure natriuresis in dogs. Hypertension 23: 1040 –1045, 1994. 978. Matavelli LC, Zhou X, Varagic J, Susic D, Frohlich ED. Salt loading produces severe renal hemodynamic dysfunction independent of arterial pressure in spontaneously hypertensive rats. Am J Physiol Heart Circ Physiol 292: H814 –H819, 2007. 957. Majid DSA, Navar LG. Nitric oxide in the mediation of pressure natriuresis. Clin Exp Pharmacol Physiol 24: 595–599, 1997. 979. Matchkov VV. Mechanisms of cellular synchronization in the vascular wall. Mechanisms of vasomotion. Dan Med Bull 57: 1–50, 2010. 958. Majid DSA, Navar LG. Nitric oxide in the control of renal hemodynamics and excretory function. Am J Hypertens 14: 74S– 82S, 2001. 980. Matchkov VV, Larsen P, Bouzinova EV, Rojek A, Boedtkjer DM, Golubinskaya V, Pedersen FS, Aalkjaer C, Nilsson H. Bestrophin-3 (vitelliform macular dystrophy 494 Physiol Rev • VOL 95 • APRIL 2015 • www.prv.org CARLSTRÖM ET AL. 2-like 3 protein) is essential for the cGMP-dependent calcium-activated chloride conductance in vascular smooth muscle cells. Circ Res 103: 864 – 872, 2008. 1002. Meininger GA, Faber JE. Adrenergic tone selectively augments myogenic constriction in skeletal muscle microcirculation. Am J Physiol Heart Circ Physiol 260: H1424 – H1432, 1991. 981. Matchkov VV, Rahman A, Bakker LM, Griffith TM, Nilsson H, Aalkjaer C. Analysis of effects of connexin-mimetic peptides in rat mesenteric small arteries. Am J Physiol Heart Circ Physiol 291: H357–H367, 2006. 1003. Messick J Jr, Spielman WS, Knox FG. Failure of high doses of Sar1-lle8-angiotensin II to abolish autoregulation of renal blood flow. Mayo Clin Proc 55: 619 – 622, 1980. 982. Matchkov VV, Secher D, V, Bodtkjer DM, Aalkjaer C. Transport and function of chloride in vascular smooth muscles. J Vasc Res 50: 69 – 87, 2012. 1004. Meyer TW, Anderson S, Rennke HG, Brenner BM. Reversing glomerular hypertension stabilizes established glomerular injury. Kidney Int 31: 752–759, 1987. 983. Matrougui K. Diabetes and microvascular pathophysiology: role of epidermal growth factor receptor tyrosine kinase. Diabetes Metab Res Rev 26: 13–16, 2010. 1005. Miller BA, Zhang W. TRP channels as mediators of oxidative stress. Adv Exp Med Biol 704: 531–544, 2011. 984. Matrougui K, Eskildsen-Helmond YE, Fiebeler A, Henrion D, Levy BI, Tedgui A, Mulvany MJ. Angiotensin II stimulates extracellular signal-regulated kinase activity in intact pressurized rat mesenteric resistance arteries. Hypertension 36: 617– 621, 2000. 1006. Miller FJ Jr, Dellsperger KC, Gutterman DD. Myogenic constriction of human coronary arterioles. Am J Physiol Heart Circ Physiol 273: H257–H264, 1997. 1007. Miller JA. Renal responses to sodium restriction in patients with early diabetes mellitus. J Am Soc Nephrol 8: 749 –755, 1997. 985. Matrougui K, Loufrani L, Heymes C, Levy BI, Henrion D. Activation of AT2 receptors by endogenous angiotensin II is involved in flow-reduced dilation in rat resistance arteries. Hypertension 34: 659 – 665, 1999. 1008. Miller PL, Rennke HG, Meyer TW. Glomerular hypertrophy accelerates hypertensive glomerular injury in rats. Am J Physiol Renal Fluid Electrolyte Physiol 261: F459 – F465, 1991. 986. Matsumura Y, Kuro T, Kobayashi Y, Konishi F, Takaoka M, Wessale JL, Opgenorth TJ, Gariepy CE, Yanagisawa M. Exaggerated vascular and renal pathology in endothelin-B receptor-deficient rats with deoxycorticosterone acetate-salt hypertension. Circulation 102: 2765–2773, 2000. 1009. Minami N, Imai Y, Munakata M, Sasaki S, Sekino H, Abe K, Yoshinaga K. Age-related changes in blood pressure, heart rate and baroreflex sensitivity in SHR. Clin Exp Pharmacol Physiol 15 Suppl: 85– 87, 1989. 987. Matsunaga H, Yamashita N, Okuda T, Kurokawa K. Mesangial cell ion transport and tubuloglomerular feedback. Curr Opin Nephrol Hypertens 3: 518 –522, 1994. 1010. Miriel VA, Mauban JR, Blaustein MP, Wier WG. Local and cellular Ca2⫹ transients in smooth muscle of pressurized rat resistance arteries during myogenic and agonist stimulation. J Physiol 518: 815– 824, 1999. 988. Mattson DL, Dwinell MR, Greene AS, Kwitek AE, Roman RJ, Jacob HJ, Cowley AW Jr. Chromosome substitution reveals the genetic basis of Dahl salt-sensitive hypertension and renal disease. Am J Physiol Renal Physiol 295: F837–F842, 2008. 1011. Mistry DK, Garland CJ. Nitric oxide (NO)-induced activation of large conductance Ca2⫹-dependent K⫹ channels [BK(Ca)] in smooth muscle cells isolated from the rat mesenteric artery. Br J Pharmacol 124: 1131–1140, 1998. 989. Mattson DL, Lu S, Cowley AW Jr. Role of nitric oxide in the control of the renal medullary circulation. Clin Exp Pharmacol Physiol 24: 587–590, 1997. 1012. Mitchell KD, Navar LG. Enhanced tubuloglomerular feedback during peritubular infusions of angiotensins I and II. Am J Physiol Renal Fluid Electrolyte Physiol 255: F383–F390, 1988. 990. Mattson DL, Lu S, Roman RJ, Cowley AW Jr. Relationship between renal perfusion pressure and blood flow in different regions of the kidney. Am J Physiol Regul Integr Comp Physiol 264: R578 –R583, 1993. 991. Mattson DL, Roman RJ, Cowley AW Jr. Role of nitric oxide in renal papillary blood flow and sodium excretion. Hypertension 19: 766 –769, 1992. 992. Mauban JR, Wier WG. Essential role of EDHF in the initiation and maintenance of adrenergic vasomotion in rat mesenteric arteries. Am J Physiol Heart Circ Physiol 287: H608 –H616, 2004. 993. Mauer SM, Brown DM, Steffes MW, Azar S. Studies of renal autoregulation in pancreatectomized and streptozotocin diabetic rats. Kidney Int 37: 909 –917, 1990. 994. McCurley A, Pires PW, Bender SB, Aronovitz M, Zhao MJ, Metzger D, Chambon P, Hill MA, Dorrance AM, Mendelsohn ME, Jaffe IZ. Direct regulation of blood pressure by smooth muscle cell mineralocorticoid receptors. Nat Med 18: 1429 –1433, 2012. 995. McDonough AA. Mechanisms of proximal tubule sodium transport regulation that link extracellular fluid volume and blood pressure. Am J Physiol Regul Integr Comp Physiol 298: R851–R861, 2010. 1013. Mitchell KD, Navar LG. Modulation of tubuloglomerular feedback responsiveness by extracellular ATP. Am J Physiol Renal Fluid Electrolyte Physiol 264: F458 –F466, 1993. 1014. Miyata N, Zou AP, Mattson DL, Cowley AW Jr. Renal medullary interstitial infusion of L-arginine prevents hypertension in Dahl salt-sensitive rats. Am J Physiol Regul Integr Comp Physiol 275: R1667–R1673, 1998. 1015. Modlinger P, Chabrashvili T, Gill PS, Mendonca M, Harrison DG, Griendling KK, Li M, Raggio J, Wellstein A, Chen Y, Welch WJ, Wilcox CS. RNA silencing in vivo reveals role of p22phox in rat angiotensin slow pressor response. Hypertension 47: 238 – 244, 2006. 1016. Mohazzab KM, Kaminski PM, Wolin MS. NADH oxidoreductase is a major source of superoxide anion in bovine coronary artery endothelium. Am J Physiol Heart Circ Physiol 266: H2568 –H2572, 1994. 1017. Moien-Afshari F, Ghosh S, Elmi S, Khazaei M, Rahman MM, Sallam N, Laher I. Exercise restores coronary vascular function independent of myogenic tone or hyperglycemic status in db/db mice. Am J Physiol Heart Circ Physiol 295: H1470 –H1480, 2008. 996. McDonough AA, Leong PK, Yang LE. Mechanisms of pressure natriuresis: how blood pressure regulates renal sodium transport. Ann NY Acad Sci 986: 669 – 677, 2003. 1018. Monkawa T, Hayashi M, Miyawaki A, Sugiyama T, Yamamoto-Hino M, Hasegawa M, Furuichi T, Mikoshiba K, Saruta T. Localization of inositol 1,4,5-trisphosphate receptors in the rat kidney. Kidney Int 53: 296 –301, 1998. 997. McLennan GP, Kline RL, Mercer PF. Effect of enalapril treatment on the pressure-natriuresis curve in spontaneously hypertensive rats. Hypertension 17: 54 – 62, 1991. 1019. Moore LC. Tubuloglomerular feedback and SNGFR autoregulation in the rat. Am J Physiol Renal Fluid Electrolyte Physiol 247: F267–F276, 1984. 998. McNay JL, Kishimoto T. Association between autoregulation and pressure dependency of renal vascular responsiveness in dogs. Circ Res 24: 599 – 605, 1969. 1020. Moore LC, Casellas D. Tubuloglomerular feedback dependence of autoregulation in rat juxtamedullary afferent arterioles. Kidney Int 37: 1402–1408, 1990. 999. Mederos y Schnitzler M, Storch U, Gudermann T. AT1 receptors as mechanosensors. Curr Opin Pharmacol 11: 112–116, 2011. 1021. Moore LC, Rich A, Casellas D. Ascending myogenic autoregulation: interactions between tubuloglomerular feedback and myogenic mechanisms. Bull Math Biol 56: 391– 410, 1994. 1000. Medros y Schnitzler M, Storch U, Meibers S, Nurwakagari P, Breit A, Essin K, Gollasch M, Gudermann T. Gq-coupled receptors as mechanosensors mediating myogenic vasoconstriction. EMBO J 27: 3092–3103, 2008. 1022. Moore LC, Schnermann J, Yarimizu S. Feedback mediation of SNGFR autoregulation in hydropenic and DOCA- and salt-loaded rats. Am J Physiol 6: F63–F74, 1979. 1001. Meininger GA, Davis MJ. Cellular mechanisms involved in the vascular myogenic response. Am J Physiol Heart Circ Physiol 263: H647–H659, 1992. 1023. Moore LC, Thorup C, Ellinger A, Paccione J, Casellas D, Kaskel FJ. Advanced glycosylation end-products and NO-dependent vasodilation in renal afferent arterioles from diabetic rats. Acta Physiol Scand 168: 101–106, 2000. Physiol Rev • VOL 95 • APRIL 2015 • www.prv.org 495 RENAL AUTOREGULATORY MECHANISMS 1024. Moosmang S, Haider N, Bruderl B, Welling A, Hofmann F. Antihypertensive effects of the putative T-type calcium channel antagonist mibefradil are mediated by the L-type calcium channel Cav1.2. Circ Res 98: 105–110, 2006. 1046. Murphy TV, Spurrell BE, Hill MA. Cellular signalling in arteriolar myogenic constriction: involvement of tyrosine phosphorylation pathways. Clin Exp Pharmacol Physiol 29: 612– 619, 2002. 1025. Moreno C, Lopez A, Llinas MT, Rodriguez F, Lopez-Farre A, Nava E, Salazar FJ. Changes in NOS activity and protein expression during acute and prolonged ANG II administration. Am J Physiol Regul Integr Comp Physiol 282: R31–R37, 2002. 1047. Murray RD, Malvin RL. Intrarenal renin and autoregulation of renal plasma flow and glomerular filtration rate. Am J Physiol Renal Fluid Electrolyte Physiol 236: F559 –F566, 1979. 1026. Moreno JM, Rodriguez G, I, Wangensteen R, varez-Guerra M, Luna JD, Garcia-Estan J, Vargas F. Tempol improves renal hemodynamics and pressure natriuresis in hyperthyroid rats. Am J Physiol Regul Integr Comp Physiol 294: R867–R873, 2008. 1048. Nagaoka A, Iwatsuka H, Suzuoki Z, Okamoto K. Genetic predisposition to stroke in spontaneously hypertensive rats. Am J Physiol 230: 1354 –1359, 1976. 1027. Moreno-Dominguez A, Colinas O, El-Yazbi A, Walsh EJ, Hill MA, Walsh MP, Cole WC. Ca2⫹ sensitization due to myosin light chain phosphatase inhibition and cytoskeletal reorganization in the myogenic response of skeletal muscle resistance arteries. J Physiol 591: 1235–1250, 2013. 1028. Morgan T. Renin, angiotensin, sodium and organ damage. Hypertension Res 26: 349 –354, 2003. 1029. Mori T, Cowley AW Jr. Role of pressure in angiotensin II-induced renal injury: chronic servo-control of renal perfusion pressure in rats. Hypertension 43: 752–759, 2004. 1049. Nagaoka A, Kakihana M, Suno M, Hamajo K. Renal hemodynamics and sodium excretion in stroke-prone spontaneously hypertensive rats. Am J Physiol Renal Fluid Electrolyte Physiol 241: F244 –F249, 1981. 1050. Nagaoka A, Toyoda S, Iwatsuka H. Increased renal vascular reactivity to norepinephrine in stroke-prone spontaneously hypertensive rat. Life Sci 23: 1159 –1166, 1978. 1051. Nagasawa T, Imig JD. Afferent arteriolar responses to beta,gamma-methylene ATP and 20-HETE are not blocked by ENaC inhibition. Physiol Rep 1: e00082, 2013. 1052. Naguib RE, Contant C, Cupples WA. Atrial natriuretic factor, angiotensin II, and the slow component of renal autoregulation. Can J Physiol Pharmacol 72: 1132–1137, 1994. 1030. Mori T, O’Connor PM, Abe M, Cowley AW Jr. Enhanced superoxide production in renal outer medulla of Dahl salt-sensitive rats reduces nitric oxide tubular-vascular cross-talk. Hypertension 49: 1336 –1341, 2007. 1053. Nahmod VE, Lanari A. Abolution of autoregulation of renal blood flow by acetylcholine. Am J Physiol 207: 123–127, 1964. 1031. Morimoto S, Ohyama T, Hisaki K, Matsumura Y. Effects of CV-4093, a new dihydropyridine calcium channel blocker, on renal hemodynamics and function in strokeprone spontaneously hypertensive rats (SHRSP). Jpn J Pharmacol 51: 257–265, 1989. 1054. Nakamura A, Hayashi K, Ozawa Y, Fujiwara K, Okubo K, Kanda T, Wakino S, Saruta T. Vessel- and vasoconstrictor-dependent role of rho/rho-kinase in renal microvascular tone. J Vasc Res 40: 244 –251, 2003. 1032. Morita H, Honda A, Inoue R, Ito Y, Abe K, Nelson MT, Brayden JE. Membrane stretch-induced activation of a TRPM4-like nonselective cation channel in cerebral artery myocytes. J Pharmacol Sci 103: 417– 426, 2007. 1055. Nakamura T, Obata J, Kuroyanagi R, Kimura H, Ikeda Y, Takano H, Naito A, Sato T, Yoshida Y. Involvement of angiotensin II in glomerulosclerosis of stroke-prone spontaneously hypertensive rats. Kidney Int 55 Suppl: S109 –S112, 1996. 1033. Moritz AR, Oldt MR. Arteriolar sclerosis in hypertensive and non-hypertensive individuals. Am J Pathol 13: 679 –728, 1937. 1056. Nakamura Y, Ono H, Frohlich ED. Differential effects of T- and L-type calcium antagonists on glomerular dynamics in spontaneously hypertensive rats. Hypertension 34: 273–278, 1999. 1034. Morsing P, Persson AE. Effect of prostaglandin synthesis inhibition on the tubuloglomerular feedback control in the rat kidney. Ren Physiol Biochem 15: 66 –72, 1992. 1035. Morsing P, Velazquez H, Ellison D, Wright FS. Resetting of tubuloglomerular feedback by interrupting early distal flow. Acta Physiol Scand 148: 63– 68, 1993. 1036. Moss NG, Kopple TE, Arendshorst WJ. Renal vasoconstriction by vasopressin V1a receptors is modulated by nitric oxide, prostanoids, and superoxide but not the ADP ribosyl cyclase CD38. Am J Physiol Renal Physiol 306: F1143–F1154, 2014. 1037. Moss NG, Vogel PA, Kopple TE, Arendshorst WJ. Thromboxane-induced renal vasoconstriction is mediated by the ADP-ribosyl cyclase CD38 and superoxide anion. Am J Physiol Renal Physiol 305: F830 –F838, 2013. 1038. Moulana M, Hosick K, Stanford J, Zhang H, Roman RJ, Reckelhoff JF. Sex differences in blood pressure control in SHR: lack of a role for EETs. Physiol Rep 2: e12022, 2014. 1039. Muller JM, Chilian WM, Davis MJ. Integrin signaling transduces shear stress-dependent vasodilation of coronary arterioles. Circ Res 80: 320 –326, 1997. 1040. Muller-Suur R, Gutsche HU, Samwer KF, Oelkers W, Hierholzer K. Tubuloglomerular feedback in rat kidneys of different renin contents. Pflügers Arch 359: 33–56, 1975. 1057. Narayanan D, Bulley S, Leo MD, Burris SK, Gabrick KS, Boop FA, Jaggar JH. Smooth muscle cell transient receptor potential polycystin-2 (TRPP2) channels contribute to the myogenic response in cerebral arteries. J Physiol 591: 5031–5046, 2013. 1058. Narayanan D, Xi Q, Pfeffer LM, Jaggar JH. Mitochondria control functional CaV1.2 expression in smooth muscle cells of cerebral arteries. Circ Res 107: 631– 641, 2010. 1059. Narayanan J, Imig JD, Roman RJ, Harder DR. Pressurization of isolated renal arteries increases inositol trisphosphate and diacylglycerol. Am J Physiol Heart Circ Physiol 266: H1840 –H1845, 1994. 1060. Narishige T, Egashira K, Akatsuka Y, Katsuda Y, Numaguchi K, Sakata M, Takeshita A. Glibenclamide, a putative ATP-sensitive K⫹ channel blocker, inhibits coronary autoregulation in anesthetized dogs. Circ Res 73: 771–776, 1993. 1061. Narumiya S, Sugimoto Y, Ushikubi F. Prostanoid receptors: structures, properties, functions. Physiol Rev 79: 1193–1226, 1999. 1062. Nath KA. Heme oxygenase-1: a provenance for cytoprotective pathways in the kidney and other tissues. Kidney Int 70: 432– 443, 2006. 1041. Muller-Suur R, Norlen BJ, Persson AE. Resetting of tubuloglomerular feedback in rat kidneys after unilateral nephrectomy. Kidney Int 18: 48 –57, 1980. 1063. Nath KA, Chmielewski DH, Hostetter TH. Regulatory role of prostanoids in glomerular microcirculation of remnant nephrons. Am J Physiol Renal Fluid Electrolyte Physiol 252: F829 –F837, 1987. 1042. Müller-Suur R, Persson AE, Ulfendahl HR. Tubuloglomerular feedback in juxtamedullary nephrons. Kidney Int 22: S-104 –S-108, 1982. 1064. Navar LG. Renal autoregulation: perspectives from whole kidney and single nephron studies. Am J Physiol Renal Fluid Electrolyte Physiol 234: F357–F370, 1978. 1043. Muller-Suur R, Ulfendahl HR, Persson AE. Evidence for tubuloglomerular feedback in juxtamedullary nephrons of young rats. Am J Physiol Renal Fluid Electrolyte Physiol 244: F425–F431, 1983. 1065. Navar LG, Arendshorst WJ, Pallone TL, Inscho EW, Imig JD, Bell PD. The renal microcirculation. In: Handbook of Physiology: Microcirculation, edited by Tuma R, Duran WN, and Ley N. Boston: Academic, 2008. 1044. Mulvany MJ. Small artery remodelling in hypertension. Basic Clin Pharmacol Toxicol 110: 49 –55, 2012. 1066. Navar LG, Bell PD. Romancing the macula densa at UAB. Kidney Int Suppl: S34 –S40, 2004. 1045. Murphy TV, Spurrell BE, Hill MA. Tyrosine phosphorylation following alterations in arteriolar intraluminal pressure and wall tension. Am J Physiol Heart Circ Physiol 281: H1047–H1056, 2001. 1067. Navar LG, Bell PD, Burke TJ. Autoregulatory responses of superficial nephrons and their association with sodium excretion during arterial pressure alterations in the dog. Circ Res 41: 487– 496, 1977. 496 Physiol Rev • VOL 95 • APRIL 2015 • www.prv.org CARLSTRÖM ET AL. 1068. Navar LG, Bell PD, White RW, Watts RL, Williams RH. Evaluation of the single nephron glomerular filtration coefficient in the dog. Kidney Int 12: 137–149, 1977. 1069. Navar LG, Champion WJ, Thomas CE. Effects of calcium channel blockade on renal vascular resistance responses to changes in perfusion pressure and angiotensinconverting enzyme inhibition in dogs. Circ Res 58: 874 – 881, 1986. 1070. Navar LG, Chomdej B, Bell PD. Absence of estimated glomerular pressure autoregulation during interrupted distal delivery. Am J Physiol 229: 1596 –1603, 1975. 1071. Navar LG, Inscho EW, Majid DSA, Imig JD, Harrison-Bernard LM, Mitchell KD. Paracrine regulation of the renal microcirculation. Physiol Rev 76: 425–536, 1996. 1072. Navar LG, Jirakulsomchok D, Bell PD, Thomas CE, Huang WC. Influence of converting enzyme inhibition on renal hemodynamics and glomerular dynamics in sodium-restricted dogs. Hypertension 4: 58 – 68, 1982. 1073. Navar LG, LaGrange RA, Bell PD, Thomas CE, Ploth DW. Glomerular and renal hemodynamics during converting enzyme inhibition (SQ20,881) in the dog. Hypertension 1: 371–377, 1979. 1074. Navar LG, Paul RV, Carmines PK, Chou CL, Marsh DJ. Intrarenal mechanisms mediating pressure natriuresis: role of angiotensin and prostaglandins. Federation Proc 45: 2885–2891, 1986. 1075. Navar LG, Ploth DW, Bell PD. Distal tubular feedback control of renal hemodynamics and autoregulation. Annu Rev Physiol 42: 557–571, 1980. 1091. Nishiyama A, Majid DS, Walker M, III, Miyatake A, Navar LG. Renal interstitial ATP responses to changes in arterial pressure during alterations in tubuloglomerular feedback activity. Hypertension 37: 753–759, 2001. 1092. Norman RA Jr, Dzielak DJ. Role of the renal nerves in onset and maintenance of spontaneous hypertension. Am J Physiol Heart Circ Physiol 243: H284 –H288, 1982. 1093. Norman RA Jr, Enobakhare JA, DeClue JW, Douglas BH, Guyton AC. Arterial pressure-urinary output relationship in hypertensive rats. Am J Physiol Regul Integr Comp Physiol 234: R98 –R103, 1978. 1094. Norrelund H, Christensen KL, Samani NJ, Kimber P, Mulvany MJ, Korsgaard N. Early narrowed afferent arteriole is a contributor to the development of hypertension. Hypertension 24: 301–308, 1994. 1095. Nouri P, Gill P, Li M, Wilcox CS, Welch WJ. p22phox in the macula densa regulates single nephron GFR during angiotensin II infusion in rats. Am J Physiol Heart Circ Physiol 292: H1685–H1689, 2007. 1096. Novak J, Ramirez RJ, Gandley RE, Sherwood OD, Conrad KP. Myogenic reactivity is reduced in small renal arteries isolated from relaxin-treated rats. Am J Physiol Regul Integr Comp Physiol 283: R349 –R355, 2002. 1097. Nowicki PT, Flavahan S, Hassanain H, Mitra S, Holland S, Goldschmidt-Clermont PJ, Flavahan NA. Redox signaling of the arteriolar myogenic response. Circ Res 89: 114 –116, 2001. 1076. Navar LG, Prieto MC, Satou R, Kobori H. Intrarenal angiotensin II and its contribution to the genesis of chronic hypertension. Curr Opin Pharmacol 11: 180 –186, 2011. 1098. Numabe A, Komatsu K, Frohlich ED. Effects of ANG-converting enzyme and alpha 1-adrenoceptor inhibition on intrarenal hemodynamics in SHR. Am J Physiol Regul Integr Comp Physiol 266: R1437–R1442, 1994. 1077. Navar LG, Saccomani G, Mitchell KD. Synergistic intrarenal actions of angiotensin on tubular reabsorption and renal hemodynamics. Am J Hypertens 4: 90 –96, 1991. 1099. O’Connor PM, Cowley AW Jr. Modulation of pressure-natriuresis by renal medullary reactive oxygen species and nitric oxide. Curr Hypertens Rep 12: 86 –92, 2010. 1078. Nelson LD, Mashburn NA, Bell PD. Altered sodium-calcium exchange in afferent arterioles of the spontaneously hypertensive rat. Kidney Int 50: 1889 –1896, 1996. 1100. O’Donnell MP, Crary GS, Oda H, Kasiske BL, Powell JR, Keane WF. Irbesartan lowers blood pressure and ameliorates renal injury in experimental non-insulindependent diabetes mellitus. Kidney Int Suppl 63: S218 –S220, 1997. 1079. Nelson LD, Unlap MT, Lewis JL, Bell PD. Renal arteriolar Na⫹/Ca2⫹ exchange in salt-sensitive hypertension. Am J Physiol Renal Physiol 276: F567–F573, 1999. 1080. Nelson MT, Conway MA, Knot HJ, Brayden JE. Chloride channel blockers inhibit myogenic tone in rat cerebral arteries. J Physiol 502: 259 –264, 1997. 1101. O’Donnell MP, Kasiske BL, Cleary MP, Keane WF. Effects of genetic obesity on renal structure and function in the Zucker rat. II. Micropuncture studies. J Lab Clin Med 106: 605– 610, 1985. 1081. Nelson MT, Quayle JM. Physiological roles and properties of potassium channels in arterial smooth muscle. Am J Physiol Cell Physiol 268: C799 –C822, 1995. 1102. Ochodnicky P, Henning RH, Buikema HJ, de Zeeuw D, Provoost AP, Van Dokkum RP. Renal vascular dysfunction precedes the development of renal damage in the hypertensive Fawn-Hooded rat. Am J Physiol Renal Physiol 298: F625–F633, 2010. 1082. Nelson RG, Meyer TW, Myers BD, Bennett PH. Clinical and pathological course of renal disease in non-insulin-dependent diabetes mellitus: the Pima Indian experience. Semin Nephrol 17: 124 –131, 1997. 1103. Oddie CJ, Dilley RJ, Bobik A. Long-term angiotensin II antagonism in spontaneously hypertensive rats: effects on blood pressure and cardiovascular amplifiers. Clin Exp Pharmacol Physiol 19: 392–395, 1992. 1083. Nelson RG, Meyer TW, Myers BD, Bennett PH. Course of renal disease in Pima Indians with non-insulin-dependent diabetes mellitus. Kidney Int Suppl 63: S45–S48, 1997. 1104. Oeckler RA, Kaminski PM, Wolin MS. Stretch enhances contraction of bovine coronary arteries via an NAD (P) oxidase-medicated activation of the extracellular signal- regulated kinase mitogen-activated protein kinase cascade. Circ Res 92: 24 – 31, 2003. 1084. New DI, Chesser AM, Thuraisingham RC, Yaqoob MM. Cerebral artery responses to pressure and flow in uremic hypertensive and spontaneously hypertensive rats. Am J Physiol Heart Circ Physiol 284: H1212–H1216, 2003. 1085. Newman JM, Dwyer RM, St-Pierre P, Richards SM, Clark MG, Rattigan S. Decreased microvascular vasomotion and myogenic response in rat skeletal muscle in association with acute insulin resistance. J Physiol 587: 2579 –2588, 2009. 1086. Nguyen Dinh CA, Montezano AC, Burger D, Touyz RM. Angiotensin II, NADPH oxidase, and redox signaling in the vasculature. Antioxid Redox Signal 19: 1110 –1120, 2013. 1087. Nilsson H, Aalkjaer C. Vasomotion: mechanisms and physiological importance. Mol Interv 3: 5179 –5189, 2003. 1105. Ofstad J, Iversen BM, Morkrid L, Sekse I. Autoregulation of renal blood flow (RBF) with and without participation of afferent arterioles. Acta Physiol Scand 130: 25–32, 1987. 1106. Ogawa N. Effect of nicardipine on the relationship of renal blood flow and of renal vascular resistance to perfusion pressure in dog kidney. J Pharm Pharmacol 42: 138 – 140, 1990. 1107. Ogawa N, Ono H. No role for prostaglandins and bradykinin in the autoregulation of renal blood flow. Jpn J Pharmacol 39: 349 –355, 1985. 1108. Ogawa N, Ono H. Different effects of noradrenaline, angiotensin II and BAY K 8644 on the abolition of autoregulation of renal blood flow by verapamil. Naunyn-Schmiedebergs Arch Pharmacol 333: 445– 449, 1986. 1088. Nishiyama A, Fukui T, Fujisawa Y, Rahman M, Tian RX, Kimura S, Youichi A. Systemic and regional hemodynamic responses to tempol in angiotensin II-infused hypertensive rats. Hypertension 37: 77– 83, 2001. 1109. Ogawa N, Ono H. Different effects of various vasodilators on autoregulation of renal blood flow in anesthetized dogs. Jpn J Pharmacol 41: 299 –306, 1986. 1089. Nishiyama A, Jackson KE, Majid DS, Rahman M, Navar LG. Renal interstitial fluid ATP responses to arterial pressure and tubuloglomerular feedback activation during calcium channel blockade. Am J Physiol Heart Circ Physiol 290: H772–H777, 2006. 1110. Ogawa N, Ono H. Role of Ca channel in the renal autoregulatory vascular response analysed by the use of BAY K 8644. Naunyn-Schmiedebergs Arch Pharmacol 335: 189 –193, 1987. 1090. Nishiyama A, Majid DS, Taher KA, Miyatake A, Navar LG. Relation between renal interstitial ATP concentrations and autoregulation-mediated changes in renal vascular resistance. Circ Res 86: 656 – 662, 2000. 1111. Ogawa N, Ono H. Effect of 8-(N,N-diethylamino)octyl-3,4,5-trimethoxybenzoate (TMB-8), an inhibitor of intracellular Ca2⫹ release, on autoregulation of renal blood flow in the dog. Naunyn-Schmiedebergs Arch Pharmacol 338: 293–296, 1988. Physiol Rev • VOL 95 • APRIL 2015 • www.prv.org 497 RENAL AUTOREGULATORY MECHANISMS 1112. Ogawa N, Yokota S, Ono H. Different interaction of bepridil and diltiazem with BAY K 8644 in the abolition of autoregulation of renal blood flow. J Cardiovasc Pharmacol 11: 147–150, 1988. 1113. Ohishi K, Carmines PK. Superoxide dismutase restores the influence of nitric oxide on renal arterioles in diabetes mellitus. J Am Soc Nephrol 5: 1559 –1566, 1995. 1114. Ohmine T, Miwa Y, Takahashi-Yanaga F, Morimoto S, Maehara Y, Sasaguri T. The involvement of aldosterone in cyclic stretch-mediated activation of NADPH oxidase in vascular smooth muscle cells. Hypertension Res 32: 690 – 699, 2009. 1115. Okamoto K, Aoki K. Development of a strain of spontaneously hypertensive rats. Jpn Circ J 27: 282–293, 1963. 1116. Okamoto K, Hazama F, Yamori Y, Haebara H, Nagaoka A. Pathogenesis and prevention of stroke in spontaneously hypertensive rats. Clin Sci Mol Med Suppl 2: 161s–163s, 1975. 1117. Okazaki K, Seki S, Kanaya N, Hattori J, Tohse N, Namiki A. Role of endotheliumderived hyperpolarizing factor in phenylephrine-induced oscillatory vasomotion in rat small mesenteric artery. Anesthesiology 98: 1164 –1171, 2003. ⫺ 1118. Okuda T, Kojima I, Ogata E, Kurokawa K. Ambient Cl ions modify rat mesangial cell contraction by modulating cell inositol trisphosphate and Ca2⫹ via enhanced prostaglandin E2. J Clin Invest 84: 1866 –1872, 1989. 1119. Olson JL, Hostetter TH, Rennke HG, Brenner BM, Venkatachalam MA. Altered glomerular permselectivity and progressive sclerosis following extreme ablation of renal mass. Kidney Int 22: 112–126, 1982. 1120. Ono H, Kokubun H, Hashimoto K. Abolition by calcium antagonists of the autoregulation of renal blood flow. Arch Pharmacol 285: 201–207, 1974. 1121. Onozato ML, Tojo A, Goto A, Fujita T, Wilcox CS. Oxidative stress and nitric oxide synthase in rat diabetic nephropathy: effects of ACEI or ARB. Kidney Int 61: 186 –194, 2002. 1122. Oppermann M, Carota I, Schiessl I, Eisner C, Castrop H, Schnermann J. Direct assessment of tubulogomerular feedback responsiveness in connexin 40-deficient mice. Am J Phyisol Renal Physiol 304: F1181–F1186, 2013. 1123. Oppermann M, Friedman DJ, Faulhaber-Walter R, Mizel D, Castrop H, Enjyoji K, Robson SC, Schnermann J. Tubuloglomerular feedback and renin secretion in NTPDase1/CD39-deficient mice. Am J Physiol Renal Physiol 294: F965–F970, 2008. 1124. Oppermann M, Hansen PB, Castrop H, Schnermann J. Vasodilatation of afferent arterioles and paradoxical increase of renal vascular resistance by furosemide in mice. Am J Physiol Renal Physiol 293: F279 –F287, 2007. 1125. Oppermann M, Qin Y, Lai EY, Eisner C, Li L, Huang Y, Mizel D, Fryc J, Wilcox CS, Briggs J, Schnermann J, Castrop H. Enhanced tubuloglomerular feedback in mice with vascular overexpression of A1 adenosine receptors. Am J Physiol Renal Physiol 297: F1256 –F1264, 2009. 1126. Orlov SN, Koltsova SV, Tremblay J, Baskakov MB, Hamet P. NKCC1 and hypertension: role in the regulation of vascular smooth muscle contractions and myogenic tone. Ann Med 44 Suppl 1: S111–S118, 2012. 1127. Ortiz MC, Atucha NM, Lahera V, Vargas F, Quesada T, Garcia-Estan J. Importance of nitric oxide and prostaglandins in the control of rat renal papillary blood flow. Hypertension 27: 377–381, 1996. 1128. Ortiz MC, Manriquez MC, Romero JC, Juncos LA. Antioxidants block angiotensin II-induced increases in blood pressure and endothelin. Hypertension 38: 655– 659, 2001. 1133. Osol G, Halpern W. Myogenic properties of cerebral blood vessels from normotensive and hypertensive rats. Am J Physiol Heart Circ Physiol 249: H914 –H921, 1985. 1134. Osol G, Halpern W. Effect of antihypertensive treatment on myogenic properties of brain arteries from the stroke-prone rat. J Hypertens Suppl 4: S517–S518, 1986. 1135. Osol G, Halpern W. Spontaneous vasomotion in pressurized cerebral arteries from genetically hypertensive rats. Am J Physiol Heart Circ Physiol 254: H28 –H33, 1988. 1136. Osol G, Laher I, Cipolla M. Protein kinase C modulates basal myogenic tone in resistance arteries from the cerebral circulation. Circ Res 68: 359 –367, 1991. 1137. Osol G, Laher I, Kelley M. Myogenic tone is coupled to phospholipase C and G protein activation in small cerebral arteries. Am J Physiol Heart Circ Physiol 265: H415–H420, 1993. 1138. Osswald H, Haas JA, Marchand GR, Knox FG. Glomerular dynamics in dogs at reduced renal artery pressure. Am J Physiol Renal Fluid Electrolyte Physiol 236: F25– F29, 1979. 1139. Ott CE, Marchand GR, Diaz-Buxo JA, Knox FG. Determinants of glomerular filtration rate in the dog. Am J Physiol 231: 235–239, 1976. 1140. Ott CE, Navar LG, Guyton AC. Pressures in static and dynamic states from capsules implanted in the kidney. Am J Physiol 221: 394 – 400, 1971. 1141. Ott CE, Vari RC. Renal autoregulation of blood flow and filtration rate in the rabbit. Am J Physiol Renal Fluid Electrolyte Physiol 237: F479 –F482, 1979. 1142. Owens GK, Kumar MS, Wamhoff BR. Molecular regulation of vascular smooth muscle cell differentiation in development and disease. Physiol Rev 84: 767– 801, 2004. 1143. Owsianik G, Talavera K, Voets T, Nilius B. Permeation and selectivity of TRP channels. Annu Rev Physiol 68: 685–717, 2006. 1144. Oyekan AO, McGiff JC. Functional response of the rat kidney to inhibition of nitric oxide synthesis: role of cytochrome P450-derived arachidonic metabolites. Br J Pharm 125: 1065–1073, 1998. 1145. Ozawa Y, Hayashi K, Kanda T, Homma K, Takamatsu I, Tatematsu S, Yoshioka K, Kumagai H, Wakino S, Saruta T. Impaired nitric oxide- and endothelium-derived hyperpolarizing factor-dependent dilation of renal afferent arteriole in Dahl saltsensitive rats. Nephrology 9: 272–277, 2004. 1146. Ozawa Y, Hayashi K, Wakino S, Kanda T, Homma K, Takamatsu I, Tatematsu S, Yoshioka K, Saruta T. Free radical activity depends on underlying vasoconstrictors in renal microcirculation. Clin Exp Hypertens 26: 219 –229, 2004. 1147. Pabbidi MR, Juncos J, Juncos L, Renic M, Tullos HJ, Lazar J, Jacob HJ, Harder DR, Roman RJ. Identification of a region of rat chromosome 1 that impairs the myogenic response and autoregulation of cerebral blood flow in fawn-hooded hypertensive rats. Am J Physiol Heart Circ Physiol 304: H311–H317, 2013. 1148. Pallone TL, Mattson DL. Role of nitric oxide in regulation of the renal medulla in normal and hypertensive kidneys. Curr Opin Nephrol Hypertens 11: 93–98, 2002. 1149. Pallone TL, Robertson CR, Jamison RL. Renal medullary microcirculation. Physiol Rev 70: 885–920, 1990. 1150. Pandey D, Patel A, Patel V, Chen F, Qian J, Wang Y, Barman SA, Venema RC, Stepp DW, Rudic RD, Fulton DJ. Expression and functional significance of NADPH oxidase 5 (Nox5) and its splice variants in human blood vessels. Am J Physiol Heart Circ Physiol 302: H1919 –H1928, 2012. 1129. Ortiz PA, Garvin JL. Role of nitric oxide in the regulation of nephron transport. Am J Physiol Renal Physiol 282: F777–F784, 2002. 1151. Paravicini TM, Touyz RM. NADPH oxidases, reactive oxygen species, and hypertension: clinical implications and therapeutic possibilities. Diabetes Care 31 Suppl 2: S170 –S180, 2008. 1130. Osborn JL, Francisco LL, DiBona GF. Effect of renal nerve stimulation on renal blood flow autoregulation and antinatriuresis during reductions in renal perfusion pressure. Proc Soc Exper Biol Med 168: 77– 81, 1981. 1152. Parekh N, Zou AP, Jungling I, Endlich K, Sadowski J, Steinhausen M. Sex differences in control of renal outer medullary circulation in rats: role of prostaglandins. Am J Physiol Renal Fluid Electrolyte Physiol 264: F629 –F636, 1993. 1131. Osmond DA, Inscho EW. P2X(1) receptor blockade inhibits whole kidney autoregulation of renal blood flow in vivo. Am J Physiol Renal Physiol 298: F1360 –F1368, 2010. 1153. Park J, Kemp BA, Howell NL, Gildea JJ, Keller SR, Carey RM. Intact microtubules are required for natriuretic responses to nitric oxide and increased renal perfusion pressure. Hypertension 51: 494 – 499, 2008. 1132. Osmond DA, Zhang S, Pollock JS, Yamamoto T, De Miguel C, Inscho EW. Clopidogrel preserves whole kidney autoregulatory behavior in ANG II-induced hypertension. Am J Physiol Renal Physiol 306: F619 –F628, 2014. 1154. Park KS, Kim Y, Lee YH, Earm YE, Ho WK. Mechanosensitive cation channels in arterial smooth muscle cells are activated by diacylglycerol and inhibited by phospholipase C inhibitor. Circ Res 93: 557–564, 2003. 498 Physiol Rev • VOL 95 • APRIL 2015 • www.prv.org CARLSTRÖM ET AL. 1155. Park S, Bivona BJ, Feng Y, Lazartigues E, Harrison-Bernard LM. Intact renal afferent arteriolar autoregulatory responsiveness in db/db mice. Am J Physiol Renal Physiol 295: F1504 –F1511, 2008. 1175. Pesic A, Madden JA, Pesic M, Rusch NJ. High blood pressure upregulates arterial L-type Ca2⫹ channels: is membrane depolarization the signal? Circ Res 94: e97–104, 2004. 1156. Parsa A, Kao WH, Xie D, Astor BC, Li M, Hsu CY, Feldman HI, Parekh RS, Kusek JW, Greene TH, Fink JC, Anderson AH, Choi MJ, Wright JT Jr, Lash JP, Freedman BI, Ojo A, Winkler CA, Raj DS, Kopp JB, He J, Jensvold NG, Tao K, Lipkowitz MS, Appel LJ. APOL1 risk variants, race, and progression of chronic kidney disease. N Engl J Med 369: 2183–2196, 2013. 1176. Peti-Peterdi J. Calcium wave of tubuloglomerular feedback. Am J Physiol Renal Physiol 291: F473–F480, 2006. 1157. Parving HH, Lehnert H, Brochner-Mortensen J, Gomis R, Andersen S, Arner P. The effect of irbesartan on the development of diabetic nephropathy in patients with type 2 diabetes. N Engl J Med 345: 870 – 878, 2001. 1178. Peti-Peterdi J, Chambrey R, Bebok Z, Biemesderfer D, St John PL, Abrahamson DR, Warnock DG, Bell PD. Macula densa Na⫹/H⫹ exchange activities mediated by apical NHE2 and basolateral NHE4 isoforms. Am J Physiol Renal Physiol 278: F452–F463, 2000. 1158. Patschan O, Kuttler B, Heemann U, Uber A, Rettig R. Kidneys from normotensive donors lower blood pressure in young transplanted spontaneously hypertensive rats. Am J Physiol Regul Integr Comp Physiol 273: R175–R180, 1997. 1159. Patzak A, Lai EY, Fahling M, Sendeski M, Martinka P, Persson PB, Persson AE. Adenosine enhances long term the contractile response to angiotensin II in afferent arterioles. Am J Physiol Regul Integr Comp Physiol 293: R2232–R2242, 2007. 1160. Patzak A, Mrowka R, Storch E, Hocher B, Persson PB. Interaction of angiotensin II and nitric oxide in isolated perfused afferent arterioles of mice. J Am Soc Nephrol 12: 1122–1127, 2001. 1161. Patzak A, Petzhold D, Wronski T, Martinka P, Babu GJ, Periasamy M, Haase H, Morano I. Constriction velocities of renal afferent and efferent arterioles of mice are not related to SMB expression. Kidney Int 68: 2726 –2734, 2005. 1162. Pavenstadt H, Kriz W, Kretzler M. Cell biology of the glomerular podocyte. Physiol Rev 83: 253–307, 2003. 1163. Pavlov AN, Sosnovtseva OV, Pavlova ON, Mosekilde E, Holstein-Rathlou NH. Characterizing multimode interaction in renal autoregulation. Physiol Meas 29: 945–958, 2008. 1164. Payne GW, Smeda JS. Cerebrovascular alterations in pressure and protein kinase C-mediated constriction in Dahl salt-sensitive rats. J Hypertens 20: 1355–1363, 2002. 1165. Pelayo JC, Quan AH, Shanley PF. Angiotensin II control of the renal microcirculation in rats with reduced renal mass. Am J Physiol Renal Fluid Electrolyte Physiol 258: F414 –F422, 1990. 1166. Pelayo JC, Shanley PF. Glomerular and tubular adaptive responses to acute nephron loss in the rat. Effect of prostaglandin synthesis inhibition. J Clin Invest 85: 1761–1769, 1990. 1167. Pelayo JC, Westcott JY. Impaired autoregulation of glomerular capillary hydrostatic pressure in the rat remnant nephron. J Clin Invest 88: 101–105, 1991. 1168. Peled N, Bendayan D, Shitrit D, Fox B, Yehoshua L, Kramer MR. Peripheral endothelial dysfunction in patients with pulmonary arterial hypertension. Respir Med 102: 1791–1796, 2008. 1177. Peti-Peterdi J, Bell PD. Regulation of macula densa Na:H exchange by angiotensin II. Kidney Int 54: 2021–2028, 1998. 1179. Pfaltzgraff ER, Shelton EL, Galindo CL, Nelms BL, Hooper CW, Poole SD, Labosky PA, Bader DM, Reese J. Embryonic domains of the aorta derived from diverse origins exhibit distinct properties that converge into a common phenotype in the adult. J Mol Cell Cardiol 69: 88 –96, 2014. 1180. Phillips SA, Sylvester FA, Frisbee JC. Oxidant stress and constrictor reactivity impair cerebral artery dilation in obese Zucker rats. Am J Physiol Regul Integr Comp Physiol 288: R522–R530, 2005. 1181. Pickering TG, Sos TA, Saddekni S, Rozenblit G, James GD, Orenstein A, Helseth G, Laragh JH. Renal angioplasty in patients with azotaemia and renovascular hypertension. J Hypertens 4 Suppl 6: 667– 669, 1986. 1182. Pires SL, Barres C, Sassard J, Julien C. Renal blood flow dynamics and arterial pressure lability in the conscious rat. Hypertension 38: 147–152, 2001. 1183. Pires SL, Julien C, Chapuis B, Sassard J, Barres C. Spontaneous renal blood flow autoregulation curves in conscious sinoaortic baroreceptor-denervated rats. Am J Physiol Renal Physiol 282: F51–F58, 2002. 1184. Pittner J, Wolgast M, Casellas D, Persson AE. Increased shear stress-released NO and decreased endothelial calcium in rat isolated perfused juxtamedullary nephrons. Kidney Int 67: 227–236, 2005. 1185. Plato CF, Stoos BA, Wang D, Garvin JL. Endogenous nitric oxide inhibits chloride transport in the thick ascending limb. Am J Physiol Renal Physiol 276: F159 –F163, 1999. 1186. Ploth DW, Kleeman K, Morrill L, Rademacher R, Jackson CA. Effects of verapamil and converting enzyme inhibition on bilateral renal function of two-kidney, one-clip hypertensive rats. Clin Sci 72: 657– 667, 1987. 1187. Ploth DW, Mackenzie HS. Tubuloglomerular feedback in one-kidney, one-clip hypertensive rats. Kidney Int Suppl 32: S115–S118, 1991. 1188. Ploth DW, Roy RN, Huang WC, Navar LG. Impaired renal blood flow and cortical pressure autoregulation in contralateral kidneys of Goldblatt hypertensive rats. Hypertension 3: 67–74, 1981. 1169. Peng H, Matchkov V, Ivarsen A, Aalkjaer C, Nilsson H. Hypothesis for the initiation of vasomotion. Circ Res 88: 810 – 815, 2001. 1189. Ploth DW, Rudulph J, LaGrange R, Navar LG. Tubuloglomerular feedback and single nephron function after converting enzyme inhibition in the rat. J Clin Invest 64: 1325–1335, 1979. 1170. Persson AE, Hahne B, Selen G. The effect of tubular perfusion with PGE2, PGF2 alpha, and PGI2 on the tubuloglomerular feedback control in the rat. Can J Physiol Pharmacol 61: 1317–1323, 1983. 1190. Ploth DW, Rudulph J, Thomas C, Navar LG. Renal and tubuloglomerular feedback responses to plasma expansion in the rat. Am J Physiol Renal Fluid Electrolyte Physiol 235: F156 –F162, 1978. 1171. Persson PB, Ehmke H, Kirchheim H. Influence of the renin-angiotensin system on the autoregulation of renal blood flow and glomerular filtration rate in conscious dogs. Acta Physiol Scand 134: 1–7, 1988. 1191. Ploth DW, Schnermann J, Dahlheim H, Hermle M, Schmidmeier E. Autoregulation and tubuloglomerular feedback in normotensive and hypertensive rats. Kidney Int 12: 253–267, 1977. 1172. Persson PB, Ehmke H, Kirchheim HR, Janssen B, Baumann JE, Just A, Nafz B. Autoregulation and non-homeostatic behaviour of renal blood flow in conscious dogs. J Physiol 462: 261–273, 1993. 1192. Pogoda K, Fuller M, Pohl U, Kameritsch P. NO, via its target Cx37, modulates calcium signal propagation selectively at myoendothelial gap junctions. Cell Commun Signal 12: 33–12, 2014. 1173. Persson PB, Ehmke H, Nafz B, Kirchheim HR. Resetting of renal autoregulation in conscious dogs: angiotensin II and alpha1-adrenoceptors. Pflügers Arch 417: 42– 47, 1990. 1193. Polichnowski AJ, Griffin KA, Long J, Williamson GA, Bidani AK. Blood pressure-renal blood flow relationships in conscious angiotensin II- and phenylephrine-infused rats. Am J Physiol Renal Physiol 305: F1074 –F1084, 2013. 1174. Persson PB, Ehmke H, Nafz B, Kirchheim HR. Sympathetic modulation of renal autoregulation by carotid occlusion in conscious dogs. Am J Physiol Renal Fluid Electrolyte Physiol 258: F364 –F370, 1990. 1194. Pollock CA, Lawrence JR, Field MJ. Tubular sodium handling and tubuloglomerular feedback in experimental diabetes mellitus. Am J Physiol Renal Fluid Electrolyte Physiol 260: F946 –F952, 1991. Physiol Rev • VOL 95 • APRIL 2015 • www.prv.org 499 RENAL AUTOREGULATORY MECHANISMS 1195. Pollock DM, Arendshorst WJ. Tubuloglomerular feedback and blood flow autoregulation during DA1-induced renal vasodilation. Am J Physiol Renal Fluid Electrolyte Physiol 258: F627–F635, 1990. 1196. Polzin DJ, Leininger JR, Osborne CA, Jeraj K. Development of renal lesions in dogs after 11/12 reduction of renal mass. Influences of dietary protein intake. Lab Invest 58: 172–183, 1988. 1216. Ren Y, Arima S, Carretero OA, Itoh S. Possible role of adenosine in macula densa control of glomerular hemodynamics. Kidney Int 61: 169 –176, 2002. 1217. Ren Y, Carretero OA, Garvin JL. Mechanism by which superoxide potentiates tubuloglomerular feedback. Hypertension 39: 624 – 628, 2002. 1218. Ren Y, Carretero OA, Garvin JL. Role of mesangial cells and gap junctions in tubuloglomerular feedback. Kidney Int 62: 525–531, 2002. 1197. Povstyan OV, Harhun MI, Gordienko DV. Ca2⫹ entry following P2X receptor activation induces IP3 receptor-mediated Ca2⫹ release in myocytes from small renal arteries. Br J Pharmacol 162: 1618 –1638, 2011. 1219. Ren Y, D’Ambrosio MA, Garvin JL, Carretero OA. Angiotensin II enhances connecting tubule glomerular feedback. Hypertension 56: 636 – 642, 2010. 1198. Premen AJ, Hall JE, Mizelle HL, Cornell JE. Maintenance of renal autoregulation during infusion of aminophylline or adenosine. Am J Physiol Renal Fluid Electrolyte Physiol 248: F366 –F373, 1985. 1220. Ren Y, D’Ambrosio MA, Garvin JL, Peterson EL, Carretero OA. Mechanism of impaired afferent arteriole myogenic response in Dahl salt sensitive rats: role of 20-HETE. Am J Physiol Renal Physiol 307: F533–F538, 2014. 1199. Pretus HA, Williams A, Navar LG. Impaired autoregulation of glomerular capillary hydrostatic pressure in the rat remnant nephron (Abstract). J Am Soc Nephrol 4: 586, 1994. 1200. Prior HM, Yates MS, Beech DJ. Functions of large conductance Ca2⫹-activated (BKCa), delayed rectifier (KV) and background K⫹ channels in the control of membrane potential in rabbit renal arcuate artery. J Physiol 511: 159 –169, 1998. 1201. Prosser BL, Ward CW, Lederer WJ. X-ROS signaling: rapid mechano-chemo transduction in heart. Science 333: 1440 –1445, 2011. 1202. Provoost AP. Spontaneous glomerulosclerosis: insights from the fawn-hooded rat. Kidney Int Suppl 45: S2–S5, 1994. 1203. Racasan S, Joles JA, Boer P, Koomans HA, Braam B. NO dependency of RBF and autoregulation in the spontaneously hypertensive rat. Am J Physiol Renal Physiol 285: F105–F112, 2003. 1204. Racasan S, Turkstra E, Joles JA, Koomans HA, Braam B. Hypoxanthine plus xanthine oxidase causes profound natriuresis without affecting renal blood flow autoregulation. Kidney Int 64: 226 –231, 2003. 1205. Raghavan R, Chen X, Yip KP, Marsh DJ, Chon KH. Interactions between TGFdependent and myogenic oscillations in tubular pressure and whole kidney blood flow in both SDR and SHR. Am J Physiol Renal Physiol 290: F720 –F732, 2006. ⫹ 1206. Raina H, Ella S, Hill MA. Decreased activity of the smooth muscle Na /Ca changer impairs arteriolar reactivity. J Physiol 586: 1669 –1681, 2008. 2⫹ ex- 1207. Ramsey CR, Berndt TJ, Knox FG. Indomethacin blocks enhanced paracellular backflux in proximal tubules. J Am Soc Nephrol 13: 1449 –1454, 2002. 1208. Rapp JP, Dene H. Development and characteristics of inbred strains of Dahl saltsensitive and salt-resistant rats. Hypertension 7: 340 –349, 1985. 1209. Rath G, Dessy C, Feron O. Caveolae, caveolin and control of vascular tone: nitric oxide (NO) and endothelium derived hyperpolarizing factor (EDHF) regulation. J Physiol Pharmacol 60 Suppl 4: 105–109, 2009. 1210. Raychowdhury MK, Yukawa M, Collins LJ, McGrail SH, Kent KC, Ware JA. Alternative splicing produces a divergent cytoplasmic tail in the human endothelial thromboxane A2 receptor. J Biol Chem 269: 19256 –19261, 1994. 1211. Rayshubskiy A, Wojtasiewicz TJ, Mikell CB, Bouchard MB, Timerman D, Youngerman BE, McGovern RA, Otten ML, Canoll P, McKhann GM, Hillman EM. Direct, intraoperative observation of ⬃0.1 Hz hemodynamic oscillations in awake human cortex: implications for fMRI. Neuroimage 87: 323–331, 2014. 1212. Reading SA, Earley S, Waldron BJ, Welsh DG, Brayden JE. TRPC3 mediates pyrimidine receptor-induced depolarization of cerebral arteries. Am J Physiol Heart Circ Physiol 288: H2055–H2061, 2005. 1213. Reho JJ, Zheng X, Fisher SA. Smooth muscle contractile diversity in the control of regional circulations. Am J Physiol Heart Circ Physiol 306: H163–H172, 2014. 1221. Ren Y, D’Ambrosio MA, Garvin JL, Wang H, Carretero OA. Possible mediators of connecting tubule glomerular feedback. Hypertension 53: 319 –323, 2009. 1222. Ren Y, D’Ambrosio MA, Garvin JL, Wang H, Carretero OA. Mechanism of inhibition of tubuloglomerular feedback by CO and cGMP. Hypertension 62: 99 –104, 2013. 1223. Ren Y, D’Ambrosio MA, Garvin JL, Wang H, Carretero OA. Prostaglandin E2 mediated connecting tubule glomerular feedback. Hypertension 62: 1123–1128, 2013. 1224. Ren Y, D’Ambrosio MA, Liu R, Pagano PJ, Garvin JL, Carretero OA. Enhanced myogenic response in the afferent arteriole of spontaneously hypertensive rats. Am J Physiol Heart Circ Physiol 298: H1769 –H1775, 2010. 1225. Ren Y, D’Ambrosio MA, Wang H, Falck JR, Peterson EL, Garvin JL, Carretero OA. Mechanisms of carbon monoxide attenuation of tubuloglomerular feedback. Hypertension 59: 1139 –1144, 2012. 1226. Ren Y, D’Ambrosio MA, Wang H, Liu R, Garvin JL, Carretero OA. Heme oxygenase metabolites inhibit tubuloglomerular feedback (TGF). Am J Physiol Renal Physiol 295: F1207–F1212, 2008. 1227. Ren Y, D’Ambrosio MA, Wang H, Peterson EL, Garvin JL, Carretero OA. Mechanisms of angiotensin II-enhanced connecting tubule glomerular feedback. Am J Physiol Renal Physiol 303: F259 –F265, 2012. 1228. Ren Y, Garvin JL, Carretero OA. Role of macula densa nitric oxide and cGMP in the regulation of tubuloglomerular feedback. Kidney Int 58: 2053–2060, 2000. 1229. Ren Y, Garvin JL, Liu R, Carretero OA. Role of macula densa adenosine triphosphate (ATP) in tubuloglomerular feedback. Kidney Int 66: 1479 –1485, 2004. 1230. Ren Y, Garvin JL, Liu R, Carretero OA. Crosstalk between the connecting tubule and the afferent arteriole regulates renal microcirculation. Kidney Int 71: 1116 –1121, 2007. 1231. Reslerova M, Loutzenhiser R. Divergent mechanisms of ATP-sensitive K⫹ channelinduced vasodilation in renal afferent and efferent arterioles. Evidence of L-type Ca2⫹ channel-dependent and -independent actions of pinacidil. Circ Res 77: 1114 – 1120, 1995. 1232. Reslerova M, Loutzenhiser R. Renal microvascular actions of calcitonin gene-related peptide. Am J Physiol Renal Physiol 274: F1078 –F1085, 1998. 1233. Resta TC, Broughton BR, Jernigan NL. Reactive oxygen species and RhoA signaling in vascular smooth muscle: role in chronic hypoxia-induced pulmonary hypertension. Adv Exp Med Biol 661: 355–373, 2010. 1234. Retailleau K, Toutain B, Galmiche G, Fassot C, Sharif-Naeini R, Kauffenstein G, Mericskay M, Duprat F, Grimaud L, Merot J, Lardeux A, Pizard A, Baudrie V, Jeunemaitre X, Feil R, Gothert JR, Lacolley P, Henrion D, Li Z, Loufrani L. Selective involvement of serum response factor in pressure-induced myogenic tone in resistance arteries. Arterioscler Thromb Vasc Biol 33: 339 –346, 2013. 1235. Rettig R, Bandelow N, Patschan O, Kuttler B, Frey B, Uber A. The importance of the kidney in primary hypertension: insights from cross-transplantation. J Hum Hypertens 10: 641– 644, 1996. 1214. Reimann K, Krishnamoorthy G, Wangemann P. NOS inhibition enhances myogenic tone by increasing rho-kinase mediated Ca2⫹ sensitivity in the male but not the female gerbil spiral modiolar artery. PLoS One 8: e53655, 2013. 1236. Rettig R, Grisk O. The kidney as a determinant of genetic hypertension: evidence from renal transplantation studies. Hypertension 46: 463– 468, 2005. 1215. Ren H, Liu NY, Andreasen A, Thomsen JS, Cao L, Christensen EI, Zhai XY. Direct physical contact between intercalated cells in the distal convoluted tubule and the afferent arteriole in mouse kidneys. PLoS One 8: e70898, 2013. 1237. Rhaleb NE, Pokharel S, Sharma U, Carretero OA. Renal protective effects of Nacetyl-Ser-Asp-Lys-Pro in deoxycorticosterone acetate-salt hypertensive mice. J Hypertens 29: 330 –338, 2011. 500 Physiol Rev • VOL 95 • APRIL 2015 • www.prv.org CARLSTRÖM ET AL. 1238. Ritter ER. Pressure/flow relations in the kidney: Alleged effects of pulse pressure. Am J Physiol 168: 480 – 489, 1952. 1239. Rivadulla C, de LC, Grieve KL, Cudeiro J. Vasomotion and neurovascular coupling in the visual thalamus in vivo. PLoS One 6: e28746, 2011. 1240. Roald AB, Ofstad J, Iversen BM. Attenuated buffering of renal perfusion pressure variation in juxtamedullary cortex in SHR. Am J Physiol Renal Physiol 282: F506 –F511, 2002. 1241. Robertson CR, Deen WM, Troy JL, Brenner BM. Dynamics of glomerular ultrafiltration in the rat. 3. Hemodynamics and autoregulation. Am J Physiol 223: 1191–1200, 1972. 1242. Rodriguez F, Zhang F, Dinocca S, Nasjletti A. Nitric oxide synthesis influences the renal vascular response to heme oxygenase inhibition. Am J Physiol Renal Physiol 284: F1255–F1262, 2003. 1243. Roman RJ. Alterations in renal medullary hemodynamics and the pressure-natriuretic response in genetic hypertension. Am J Hypertens 3: 893–900, 1990. 1244. Roman RJ. Abnormal renal hemodynamics and pressure-natriuresis relationship in Dahl salt-sensitive rats. Am J Physiol Renal Fluid Electrolyte Physiol 251: F57–F65, 1986. 1245. Roman RJ. Pressure-diuresis in volume-expanded rats. Tubular reabsorption in superficial and deep nephrons. Hypertension 12: 177–183, 1988. 1246. Roman RJ, Cowley AW Jr. Abnormal pressure-diuresis-natriuresis response in spontaneously hypertensive rats. Am J Physiol Renal Fluid Electrolyte Physiol 248: F199 – F205, 1985. 1261. Rosivall L, Youngblood P, Navar LG. Renal autoregulatory efficiency during angiotensin-converting enzyme inhibition in dogs on a low sodium diet. Renal Physiol 9: 18 –28, 1986. 1262. Rossi M, Bertuglia S, Varanini M, Giusti A, Santoro G, Carpi A. Generalised wavelet analysis of cutaneous flowmotion during post-occlusive reactive hyperaemia in patients with peripheral arterial obstructive disease. Biomed Pharmacother 59: 233– 239, 2005. 1263. Rothe CF, Nash FD, Thompson DE. Patterns in autoregulation of renal blood flow in the dog. Am J Physiol 220: 1621–1626, 1971. 1264. Rothermund L, Traupe T, Dieterich M, Kossmehl P, Yagil C, Yagil Y, Kreutz R. Nephroprotective effects of the endothelin ET(A) receptor antagonist darusentan in salt-sensitive genetic hypertension. Eur J Pharmacol 468: 209 –216, 2003. 1265. Ruan X, Arendshorst WJ. Role of protein kinase C in angiotensin II-induced renal vasoconstriction in genetically hypertensive rats. Am J Physiol Renal Fluid Electrolyte Physiol 270: F945–F952, 1996. 1266. Ruan X, Wagner C, Chatziantoniou C, Kurtz A, Arendshorst WJ. Regulation of angiotensin II receptor AT1 subtypes in renal afferent arterioles during chronic changes in sodium diet. J Clin Invest 99: 1072–1081, 1997. 1267. Rubin MJ, Bohlen HG. Cerebral vascular autoregulation of blood flow and tissue PO2 in diabetic rats. Am J Physiol Heart Circ Physiol 249: H540 –H546, 1985. 1268. Rudd MA, Grippo RS, Arendshorst WJ. Acute renal denervation produces a diuresis and natriuresis in young SHR but not WKY rats. Am J Physiol Renal Fluid Electrolyte Physiol 251: F655–F661, 1986. 1247. Roman RJ, Cowley AW Jr. Characterization of a new model for the study of pressure-natriuresis in the rat. Am J Physiol Renal Fluid Electrolyte Physiol 248: F190 –F198, 1985. 1269. Sachidanandam K, Hutchinson JR, Elgebaly MM, Mezzetti EM, Dorrance AM, Motamed K, Ergul A. Glycemic control prevents microvascular remodeling and increased tone in type 2 diabetes: link to endothelin-1. Am J Physiol Regul Integr Comp Physiol 296: R952–R959, 2009. 1248. Roman RJ, Cowley AW Jr, Garcia-Estan J, Lombard JH. Pressure-diuresis in volumeexpanded rats Cortical and medullary hemodynamics. Hypertension 12: 168 –176, 1988. 1270. Sadowski J, Kulczykowska E, Kulczykowski M, Badzynska B. Renal vein outflow recording in rats and rabbits: alternative method of RBF measurement. Acta Physiol Hung 72: 335–342, 1988. 1249. Roman RJ, Harder DR. Cellular and ionic signal transduction mechanisms for the mechanical activation of renal arterial vascular smooth muscle. J Am Soc Nephrol 4: 986 –996, 1993. 1271. Saeed A, DiBona GF, Grimberg E, Nguy L, Mikkelsen ML, Marcussen N, Guron G. High-NaCl diet impairs dynamic renal blood flow autoregulation in rats with adenine-induced chronic renal failure. Am J Physiol Regul Integr Comp Physiol 306: R411– R419, 2014. 1250. Roman RJ, Hoagland KM, Lopez B, Kwitek AE, Garrett MR, Rapp JP, Lazar J, Jacob HJ, Sarkis A. Characterization of blood pressure and renal function in chromosome 5 congenic strains of Dahl S rats. Am J Physiol Renal Physiol 290: F1463–F1471, 2006. 1251. Roman RJ, Kaldunski M. Pressure natriuresis and cortical and papillary blood flow in inbred Dahl rats. Am J Physiol Regul Integr Comp Physiol 261: R595–R602, 1991. 1272. Saeed A, DiBona GF, Guron G. Effects of endothelin receptor antagonists on renal hemodynamics in angiotensin II-infused rats on high NaCl intake. Kidney Blood Press Res 36: 258 –267, 2012. 1252. Roman RJ, Kaldunski ML. Renal cortical and papillary blood flow in spontaneously hypertensive rats. Hypertension 11: 657– 663, 1988. 1273. Saeed A, DiBona GF, Marcussen N, Guron G. High-NaCl intake impairs dynamic autoregulation of renal blood flow in ANG II-infused rats. Am J Physiol Regul Integr Comp Physiol 299: R1142–R1149, 2010. 1253. Roman RJ, Kaldunski ML, Mattson DL, Mistry M, Nasjletti A. Influence of eicosanoids on renal function of DOCA-salt hypertensive rats. Hypertension 12: 287–294, 1988. 1274. Sakai T, Hallman E, Marsh DJ. Frequency domain analysis of renal autoregulation in the rat. Am J Physiol Renal Fluid Electrolyte Physiol 250: F364 –F373, 1986. 1254. Roman RJ, Lianos E. Influence of prostaglandins on papillary blood flow and pressurenatriuretic response. Hypertension 15: 29 –35, 1990. 1275. Salamanca DA, Khalil RA. Protein kinase C isoforms as specific targets for modulation of vascular smooth muscle function in hypertension. Biochem Pharmacol 70: 1537– 1547, 2005. 1255. Roman RJ, Osborn JL. Renal function and sodium balance in conscious Dahl S and R rats. Am J Physiol Regul Integr Comp Physiol 252: R833–R841, 1987. 1256. Roman RJ, Smits C. Laser-Doppler determination of papillary blood flow in young and adult rats. Am J Physiol Renal Fluid Electrolyte Physiol 251: F115–F124, 1986. 1276. Sallstrom J, Carlsson PO, Fredholm BB, Larsson E, Persson AE, Palm F. Diabetesinduced hyperfiltration in adenosine A(1)-receptor deficient mice lacking the tubuloglomerular feedback mechanism. Acta Physiol 190: 253–259, 2007. 1257. Roman RJ, Zou AP. Influence of the renal medullary circulation on the control of sodium excretion. Am J Physiol Regul Integr Comp Physiol 265: R963–R973, 1993. 1277. Salom MG, Lahera V, Miranda-Guardiola F, Romero JC. Blockade of pressure natriuresis induced by inhibition of renal synthesis of nitric oxide in dogs. Am J Physiol Renal Fluid Electrolyte Physiol 262: F718 –F722, 1992. 1258. Romero JC, Knox FG. Mechanisms underlying pressure-related natriuresis: the role of the renin-angiotensin and prostaglandin systems. Hypertension 11: 724 –738, 1988. 1278. Salomonsson M, Braunstein TH, Holstein-Rathlou NH, Jensen LJ. Na⫹-independent, nifedipine-resistant rat afferent arteriolar Ca2⫹ responses to noradrenaline: possible role of TRPC channels. Acta Physiol 200: 265–278, 2010. 1259. Roos MH, van Rodijnen WF, Van Lambalgen AA, Ter Wee PM, Tangelder GJ. Renal microvascular constriction to membrane depolarization and other stimuli: pivotal role for rho-kinase. Pflügers Arch 452: 471– 477, 2006. 1279. Salomonsson M, Gustafsson F, Andreasen D, Jensen BL, Holstein-Rathlou NH. Local electric stimulation causes conducted calcium response in rat interlobular arteries. Am J Physiol Renal Physiol 283: F473–F480, 2002. 1260. Rosen S, Epstein FH, Brezis M. Determinants of intrarenal oxygenation: factors in acute renal failure. Renal Failure 14: 321–325, 1992. 1280. Salomonsson M, Sorensen CM, Arendshorst WJ, Steendahl J, Holstein-Rathlou NH. Calcium handling in afferent arterioles. Acta Physiol Scand 181: 421– 429, 2004. Physiol Rev • VOL 95 • APRIL 2015 • www.prv.org 501 RENAL AUTOREGULATORY MECHANISMS 1281. Sanchez-Ferrer CF, Roman RJ, Harder DR. Pressure-dependent contraction of rat juxtamedullary afferent arterioles. Circ Res 64: 790 –798, 1989. 1282. Sanchez-Lozada LG, Tapia E, Soto V, Avila-Casado C, Franco M, Wessale JL, Zhao L, Johnson RJ. Effect of febuxostat on the progression of renal disease in 5/6 nephrectomy rats with and without hyperuricemia. Nephron Physiol 108: 69 –78, 2008. 1283. Sandner P, Kornfeld M, Ruan X, Arendshorst WJ, Kurtz A. Nitric oxide/cAMP interactions in the control of rat renal vascular resistance. Circ Res 84: 186 –192, 1999. 1284. Sanowski J, Wocial B. Renin release and autoregulation of blood flow in a new model of non-filtering non-transporting kidney. J Physiol 266: 219 –233, 1977. 1285. Sarubbi D, Quilley J. Evidence against a role of arachidonic acid metabolites in autoregulatory responses of the isolated perfused rat kidney. Eur J Pharmacol 197: 27–31, 1991. 1286. Satoh M, Haruna Y, Fujimoto S, Sasaki T, Kashihara N. Telmisartan improves endothelial dysfunction and renal autoregulation in Dahl salt-sensitive rats. Hypertension Res 33: 135–142, 2010. 1287. Satriano J, Wead L, Cardus A, Deng A, Boss GR, Thomson SC, Blantz RC. Regulation of ecto-5’-nucleotidase by NaCl and nitric oxide: potential roles in tubuloglomerular feedback and adaptation. Am J Physiol Renal Physiol 291: F1078 –F1082, 2006. 1288. Savage T, McMahon AC, Mullen A, Tribe RM, Yaqoob MM. Ramipril prevents basal arterial constriction and enhanced myogenic tone in the femoral artery in mildly uraemic normotensive rats. Clin Sci 97: 233–237, 1999. 1289. Savage T, McMahon AC, Mullen AM, Nott CA, Dodd SM, Tribe RM, Yaqoob MM. Increased myogenic tone precedes structural changes in mild experimental uraemia in the absence of hypertension in rats. Clin Sci 95: 681– 686, 1998. 1290. Schaldecker T, Kim S, Tarabanis C, Tian D, Hakroush S, Castonguay P, Ahn W, Wallentin H, Heid H, Hopkins CR, Lindsley CW, Riccio A, Buvall L, Weins A, Greka A. Inhibition of the TRPC5 ion channel protects the kidney filter. J Clin Invest 123: 5298 –5309, 2013. 1291. Schiffrin EL. Endothelin:role in experimental hypertension. J Cardiovasc Pharmacol 35: S33–S35, 2000. 1292. Schiffrin EL, Lariviere R, Li JS, Sventek P, Touyz RM. Deoxycorticosterone acetate plus salt induces overexpression of vascular endothelin-1 and severe vascular hypertrophy in spontaneously hypertensive rats. Hypertension 25: 769 –773, 1995. 1293. Schjoedt KJ, Christensen PK, Jorsal A, Boomsma F, Rossing P, Parving HH. Autoregulation of glomerular filtration rate during spironolactone treatment in hypertensive patients with type 1 diabetes: a randomized crossover trial. Nephrol Dial Transplant 24: 3343–3349, 2009. 1294. Schleifenbaum J, Kassmann M, Szijarto IA, Hercule HC, Tano JY, Weinert S, Heidenreich M, Pathan AR, Anistan YM, Alenina N, Rusch NJ, Bader M, Jentsch TJ, Gollasch M. Stretch-activation of angiotensin II type 1a receptors contributes to the myogenic response of mouse mesenteric and renal arteries. Circ Res 115: 263–272, 2014. 1301. Schnackenberg CG, Welch WJ, Wilcox CS. TP receptor-mediated vasoconstriction in microperfused afferent arterioles: role O2⫺ and NO. Am J Physiol Renal Physiol 279: F302–F308, 2000. 1302. Schnackenberg CG, Wilcox CS. Two-week administration of tempol attenuates both hypertension and renal excretion of 8-Iso prostaglandin F2alpha. Hypertension 33: 424 – 428, 1999. 1303. Schnackenberg CG, Wilcox CS. The SOD mimetic tempol restores vasodilation in afferent arterioles of experimental diabetes. Kidney Int 59: 1859 –1864, 2001. 1304. Schneider MP, Ge Y, Pollock DM, Pollock JS, Kohan DE. Collecting duct-derived endothelin regulates arterial pressure and Na excretion via nitric oxide. Hypertension 51: 1605–1610, 2008. 1305. Schnermann J. Micropuncture analysis of tubuloglomerular feedback regulation in transgenic mice. J Am Soc Nephrol 10: 2614 –2619, 1999. 1306. Schnermann J. Maintained tubuloglomerular feedback responses during acute inhibition of P2 purinergic receptors in mice. Am J Physiol Renal Physiol 300: F339 –F344, 2011. 1307. Schnermann J, Briggs JP. Participation of renal cortical prostaglandins in the regulation of glomerular filtration rate. Kidney Int 19: 802– 815, 1981. 1308. Schnermann J, Briggs JP. Interaction between loop of Henle flow and arterial pressure as determinants of glomerular pressure. Am J Physiol Renal Fluid Electrolyte Physiol 256: F421–F429, 1989. 1309. Schnermann J, Briggs JP. Restoration of tubuloglomerular feedback in volume-expanded rats by angiotensin II. Am J Physiol Renal Fluid Electrolyte Physiol 259: F565– F572, 1990. 1310. Schnermann J, Briggs JP. Role of NO in the function of the juxtaglomerular apparatus. In: Nitric Oxide and the Kidney: Physiology and Pathophysiology, edited by Goligorsky MS and Gross SS. New York: Chapman & Hall, 1997. 1311. Schnermann J, Briggs JP. Function of the juxtaglomerular apparatus: control of glomerular hemodynamics and renin secretion. In: Seldin and Giebisch’s The Kidney: Physiology and Pathophysiology, edited by Alpern RJ and Hebert SC. New York: Academic, 2007. 1312. Schnermann J, Briggs JP. Tubuloglomerular feedback: mechanistic insights from gene-manipulated mice. Kidney Int 74: 418 – 426, 2008. 1313. Schnermann J, Briggs JP, Weber PC. Tubuloglomerular feedback, prostaglandins, and angiotensin in the autoregulation of glomerular filtration rate. Kidney Int 25: 53– 64, 1984. 1314. Schnermann J, Gokel M, Weber PC, Schubert G, Briggs JP. Tubuloglomerular feedback and glomerular morphology in Goldblatt hypertensive rats on varying protein diets. Kidney Int 29: 520 –529, 1986. 1315. Schnermann J, Hermle M, Schmidmeier E, Dahlheim H. Impaired potency for feedback regulation of gloemrular filtration rate in DOCA escaped rats. Pflügers Arch 358: 325–338, 1975. 1295. Schmid HE Jr, Spencer MP. Characteristics of pressure-flow regulation by the kidney. J Appl Physiol 17: 201–204, 1962. 1316. Schnermann J, Levine DZ. Paracrine factors in tubuloglomerular feedback: adenosine, ATP, and nitric oxide. Annu Rev Physiol 65: 501–529, 2003. 1296. Schmidt VJ, Jobs A, von MJ, Worsdorfer P, Willecke K, de Wit C. Connexin45 is expressed in vascular smooth muscle but its function remains elusive. PLoS One 7: e42287, 2012. 1317. Schnermann J, Ploth DW, Dahlheim H. Inter-relationship between autoregulation of glomerular filtration rate, tubuloglomerular feedback and juxtaglomerular renin activity in normotensive and hypertensive rats. Clin Sci Mol Med 51: 105s–107s, 1976. 1297. Schmieder RE, Messerli FH, Garavaglia G, Nunez B. Glomerular hyperfiltration indicates early target organ damage in essential hypertension. JAMA 264: 2775–2780, 1990. 1318. Schnermann J, Traynor TR, Yang T, Huang YG, Oliverio MI, Coffman T, Briggs JP. Absence of tubuloglomerular feedback responses in AT1A receptor-deficient mice. Am J Physiol Renal Physiol 273: F315–F320, 1997. 1298. Schmieder RE, Veelken R, Gatzka CD, Ruddel H, Schachinger H. Predictors for hypertensive nephropathy: results of a 6-year follow-up study in essential hypertension. J Hypertens 13: 357–365, 1995. 1299. Schmitz PG, O’Donnell MP, Kasiske BL, Katz SA, Keane WF. Renal injury in obese Zucker rats: glomerular hemodynamic alterations and effects of enalapril. Am J Physiol Renal Fluid Electrolyte Physiol 263: F496 –F502, 1992. 1300. Schnackenberg CG, Welch WJ, Wilcox CS. Normalization of blood pressure and renal vascular resistance in SHR with a membrane-permeable superoxide dismutase mimetic: role of nitric oxide. Hypertension 32: 59 – 64, 1998. 502 1319. Schnermann J, Weber PC. Reversal of indomethacin-induced inhibition of tubuloglomerular feedback by protaglandin infusion. Prostaglandins 24: 351–361, 1982. 1320. Schnermann J, Weihprecht H, Briggs JP. Inhibition of tubuloglomerular feedback during adenosine1 receptor blockade. Am J Physiol Renal Fluid Electrolyte Physiol 258: F553–F561, 1990. 1321. Scholey JW, Meyer TW. Control of glomerular hypertension by insulin administration in diabetic rats. J Clin Invest 83: 1384 –1389, 1989. 1322. Scholz H, Kurtz A. Role of protein kinase C in renal vasoconstriction caused by angiotensin II. Am J Physiol Cell Physiol 259: C421–C426, 1990. Physiol Rev • VOL 95 • APRIL 2015 • www.prv.org CARLSTRÖM ET AL. 1323. Scholz H, Kurtz A. Disparate effects of calcium channel blockers on pressure dependence of renin secretion and flow in the isolated perfused rat kidney. Pflügers Arch 421: 155–162, 1992. 1345. Selkurt EE, Hall PW, Spencer MP. Influence of graded arterial pressure decrement on renal clearance of creatinine, p-aminohippurate and sodium. Am J Physiol 159: 369 –378, 1949. 1324. Scholz H, Kurtz A. Differential regulation of cytosolic calcium between afferent arteriol ar smooth muscle cells from mouse kidney. Pflügers Arch 431: 46 –51, 1995. 1346. Sell M, Boldt W, Markwardt F. Desynchronising effect of the endothelium on intracellular Ca2⫹ concentration dynamics in vascular smooth muscle cells of rat mesenteric arteries. Cell Calcium 32: 105–120, 2002. 1325. Schoonmaker GC, Fallet RW, Carmines PK. Superoxide anion curbs nitric oxide modulation of afferent arteriolar Ang II responsiveness in diabetes mellitus. Am J Physiol Renal Physiol 278: F302–F309, 2000. 1347. Semple SJ, de Wardener HE. Effect of increased renal venous pressure on circulatory autoregulation of isolated dog kidneys. Circ Res 7: 643– 648, 1959. 1326. Schrier RW, Estacio RO, Esler A, Mehler P. Effects of aggressive blood pressure control in normotensive type 2 diabetic patients on albuminuria, retinopathy and strokes. Kidney Int 61: 1086 –1097, 2002. 1348. Sendeski MM, Liu ZZ, Perlewitz A, Busch JF, Ikromov O, Weikert S, Persson PB, Patzak A. Functional characterization of isolated, perfused outermedullary descending human vasa recta. Acta Physiol 208: 50 –56, 2013. 1327. Schrijvers BF, De Vriese AS, Van de Voorde J, Rasch R, Lameire NH, Flyvbjerg A. Long-term renal changes in the Goto-Kakizaki rat, a model of lean type 2 diabetes. Nephrol Dial Transplant 19: 1092–1097, 2004. 1349. Seppey D, Sauser R, Koenigsberger M, Beny JL, Meister JJ. Intercellular calcium waves are associated with the propagation of vasomotion along arterial strips. Am J Physiol Heart Circ Physiol 298: H488 –H496, 2010. 1328. Schubert R, Lidington D, Bolz SS. The emerging role of Ca2⫹ sensitivity regulation in promoting myogenic vasoconstriction. Cardiovasc Res 77: 8 –18, 2008. 1350. Sgouralis I, Layton AT. Autoregulation and conduction of vasomotor responses in a mathematical model of the rat afferent arteriole. Am J Physiol Renal Physiol 303: F229 –F239, 2012. 1329. Schubert R, Mulvany MJ. The myogenic response: established facts and attractive hypotheses. Clin Sci 96: 313–326, 1999. 1330. Schulman IH, Zhou MS, Raij L. Nitric oxide, angiotensin II, and reactive oxygen species in hypertension and atherogenesis. Curr Hypertens Rep 7: 61– 67, 2005. 1331. Schurek HJ, Jost U, Baumgartl H, Bertram H, Heckmann U. Evidence for a preglomerular oxygen diffusion shunt in rat renal cortex. Am J Physiol Renal Fluid Electrolyte Physiol 259: F910 –F915, 1990. 1332. Schurek HJ, Kriz W. Morphologic and functional evidence for oxygen deficiency in the isolated perfused rat kidney. Lab Invest 53: 145–155, 1985. 1333. Schuster A, Lamboley M, Grange C, Oishi H, Beny JL, Stergiopulos N, Meister JJ. Calcium dynamics and vasomotion in rat mesenteric arteries. J Cardiovasc Pharmacol 43: 539 –548, 2004. 1334. Schweda F, Klar J, Narumiya S, Nusing RM, Kurtz A. Stimulation of renin release by prostaglandin E2 is mediated by EP2 and EP4 receptors in mouse kidneys. Am J Physiol Renal Physiol 287: F427–F433, 2004. 1335. Schweda F, Seebauer H, Kramer BK, Kurtz A. Functional role of sodium-calcium exchange in the regulation of renal vascular resistance. Am J Physiol Renal Physiol 280: F155–F161, 2001. 1336. Schweitzer G, Gertz KH. Single-nephron hemodynamics in one-kidney one-clip hypertension in the rat. Renal Physiol 7: 46 –53, 1984. 1337. Schwietzer G, Gertz KH. Changes of hemodynamics and glomerular ultrafiltration in renal hypertension of rats. Kidney Int 15: 134 –143, 1979. 1338. Schwietzer G, Gertz KH. Goldblatt hypertension in rats: a model for unilateral renal artery stenosis in man. Contrib Nephrol 19: 134 –138, 1980. 1339. Scotland RS, Chauhan S, Vallance PJ, Ahluwalia A. An endothelium-derived hyperpolarizing factor-like factor moderates myogenic constriction of mesenteric resistance arteries in the absence of endothelial nitric oxide synthase-derived nitric oxide. Hypertension 38: 833– 839, 2001. 1340. Scully CG, Mitrou N, Braam B, Cupples WA, Chon KH. Detecting physiological systems with laser speckle perfusion imaging of the renal cortex. Am J Physiol Regul Integr Comp Physiol 304: R929 –R939, 2013. 1341. Scully CG, Siu KL, Cupples WA, Braam B, Chon KH. Time-frequency approaches for the detection of interactions and temporal properties in renal autoregulation. Ann Biomed Eng 41: 172–184, 2013. 1342. Seeliger E, Wronski T, Ladwig M, Dobrowolski L, Vogel T, Godes M, Persson PB, Flemming B. The renin-angiotensin system and the third mechanism of renal blood flow autoregulation. Am J Physiol Renal Physiol 296: F1334 –F1345, 2009. 1343. Selen G, Persson AE. Effects of reduced renal artery pressure on feedback control of glomerular filtration. Am J Physiol Renal Fluid Electrolyte Physiol 244: F342–F348, 1983. 1344. Selkurt EE. Effect of pulse pressure and mean arterial pressure modification on renal hemodynamics and electrolyte and water excretion. Circ 4: 541–551, 1951. 1351. Sgouralis I, Layton AT. Theoretical assessment of renal autoregulatory mechanisms. Am J Physiol Renal Physiol 306: F1357–F1371, 2014. 1352. Sharif-Naeini R, Dedman A, Folgering JH, Duprat F, Patel A, Nilius B, Honore E. TRP channels and mechanosensory transduction: insights into the arterial myogenic response. Pflügers Arch 456: 529 –540, 2008. 1353. Sharif-Naeini R, Folgering JH, Bichet D, Duprat F, Lauritzen I, Arhatte M, Jodar M, Dedman A, Chatelain FC, Schulte U, Retailleau K, Loufrani L, Patel A, Sachs F, Delmas P, Peters DJ, Honore E. Polycystin-1 and -2 dosage regulates pressure sensing. Cell 139: 587–596, 2009. 1354. Sharma K, Cook A, Smith M, Valancius C, Inscho EW. TGF-beta impairs renal autoregulation via generation of ROS. Am J Physiol Renal Physiol 288: F1069 –F1077, 2005. 1355. Sharma K, McCue P, Dunn SR. Diabetic kidney disease in the db/db mouse. Am J Physiol Renal Physiol 284: F1138 –F1144, 2003. 1356. Shi S, Carattino MD, Hughey RP, Kleyman TR. ENaC regulation by proteases and shear stress. Curr Mol Pharmacol 6: 28 –34, 2013. 1357. Shi Y, Lau C, Cupples WA. Interactive modulation of renal myogenic autoregulation by nitric oxide and endothelin acting through ET-B receptors. Am J Physiol Regul Integr Comp Physiol 292: R354 –R361, 2007. 1358. Shi Y, Wang X, Chon KH, Cupples WA. Tubuloglomerular feedback-dependent modulation of renal myogenic autoregulation by nitric oxide. Am J Physiol Regul Integr Comp Physiol 290: R982–R991, 2006. 1359. Shibuya J, Ohyanagi M, Iwasaki T. Enhanced myogenic response in resistance small arteries from spontaneously hypertensive rats: relationship to the voltage-dependent calcium channel. Am J Hypertens 11: 767–773, 1998. 1360. Shipley RE, Study RS. Changes in renal blood flow, extraction of inulin, glomerular filtation rate, tissue pressure and urine flow with acute alterations of renla artery blood pressure. Am J Physiol 167: 676 – 688, 1951. 1361. Shiraishi M, Wang X, Walsh MP, Kargacin G, Loutzenhiser K, Loutzenhiser R. Myosin heavy chain expression in renal afferent and efferent arterioles: relationship to contractile kinetics and function. FASEB J 17: 2284 –2286, 2003. 1362. Siegl D, Koeppen M, Wolfle SE, Pohl U, de Wit C. Myoendothelial coupling is not prominent in arterioles within the mouse cremaster microcirculation in vivo. Circ Res 97: 781–788, 2005. 1363. Sigmon DH, Beierwaltes WH. Influence of nitric oxide in the chronic phase of two-kidney, one clip renovascular hypertension. Hypertension 31: 649 – 656, 1998. 1364. Silbernagl S. The renal handling of amino acids and oligopeptides. Physiol Rev 68: 911–1007, 1988. 1365. Sima CA, Koeners MP, Joles JA, Braam B, Magil AB, Cupples WA. Increased susceptibility to hypertensive renal disease in streptozotocin-treated diabetic rats is not modulated by salt intake. Diabetologia 55: 2246 –2255, 2012. Physiol Rev • VOL 95 • APRIL 2015 • www.prv.org 503 RENAL AUTOREGULATORY MECHANISMS 1366. Simchon S, Manger WM, Shi GS, Brensilver J. Impaired renal vascular reactivity in prehypertensive Dahl salt-sensitive rats. Hypertension 20: 524 –532, 1992. 1367. Simon Bulley SB, Jaggar JH. Cl⫺ channels in smooth muscle cells. Pflügers Arch 466: 861– 872, 2013. 1388. Smirnov SV, Loutzenhiser K, Loutzenhiser RD. Voltage-activated Ca2⫹ channels in rat renal afferent and efferent myocytes: no evidence for the T-type Ca2⫹ current. Cardiovasc Res 97: 293–301, 2013. 1389. Smith TL, Hutchins PM. Central hemodynamics in the developmental stage of spontaneous hypertension in the unanesthetized rat. Hypertension 1: 508 –517, 1979. 1368. Simon AM, McWhorter AR. Decreased intercellular dye-transfer and downregulation of non-ablated connexins in aortic endothelium deficient in connexin37 or connexin40. J Cell Sci 116: 2223–2236, 2003. 1390. Soboloff J, Rothberg BS, Madesh M, Gill DL. STIM proteins: dynamic calcium signal transducers. Nat Rev Mol Cell Biol 13: 549 –565, 2012. 1369. Simon AM, McWhorter AR. Role of connexin37 and connexin40 in vascular development. Cell Commun Adhes 10: 379 –385, 2003. 1391. Somers MJ, Mavromatis K, Galis ZS, Harrison DG. Vascular superoxide production and vasomotor function in hypertension induced by deoxycorticosterone acetatesalt. Circulation 101: 1722–1728, 2000. 1370. Simonet S, Isabelle M, Bousquenaud M, Clavreul N, Feletou M, Vayssettes-Courchay C, Verbeuren TJ. KCa 3.1 channels maintain endothelium-dependent vasodilatation in isolated perfused kidneys of spontaneously hypertensive rats after chronic inhibition of NOS. Br J Pharmacol 167: 854 – 867, 2012. 1392. Somlyo AP, Somlyo AV. Ca2⫹ sensitivity of smooth muscle and nonmuscle myosin II: modulated by G proteins, kinases, and myosin phosphatase. Physiol Rev 83: 1325– 1358, 2003. 1371. Simons JL, Provoost AP, Anderson S, Troy JL, Rennke HG, Sandstrom DJ, Brenner BM. Pathogenesis of glomerular injury in the fawn-hooded rat: early glomerular capillary hypertension predicts glomerular sclerosis. J Am Soc Nephrol 3: 1775–1782, 1993. 1372. Singh P, Blantz RC, Rosenberger C, Gabbai FB, Schoeb TR, Thomson SC. Aberrant tubuloglomerular feedback and HIF-1 alpha confer resistance to ischemia after subtotal nephrectomy. J Am Soc Nephrol 23: 483– 493, 2012. 1373. Singh P, Deng A, Blantz RC, Thomson SC. Unexpected effect of angiotensin AT1 receptor blockade on tubuloglomerular feedback in early subtotal nephrectomy. Am J Physiol Renal Physiol 296: F1158 –F1165, 2009. 1374. Sipos A, Vargas S, Peti-Peterdi J. Direct demonstration of tubular fluid flow sensing by macula densa cells. Am J Physiol Renal Physiol 299: F1087–F1093, 2010. 1375. Siu KL, Sung B, Cupples WA, Moore LC, Chon KH. Detection of low-frequency oscillations in renal blood flow. Am J Physiol Renal Physiol 297: F155–F162, 2009. 1376. Skarlatos S, Metting PJ, Britton SL. Spontaneous pressure-flow patterns in the kidney of conscious rats. Am J Physiol Heart Circ Physiol 265: H2151–H2159, 1993. 1377. Skott O, Baumbach L. Effects of adenosine on renin release from isolated rat glomeruli and kidney slices. Pflügers Arch 404: 232–237, 1985. 1378. Skott O, Briggs JP. Direct demonstration of macula densa-mediated renin secretion. Science 237: 1618 –1620, 1987. 1379. Skov K, Fenger-Gron J, Mulvany MJ. Effects of an angiotensin-converting enzyme inhibitor, a calcium antagonist, and an endothelin receptor antagonist on renal afferent arteriolar structure. Hypertension 28: 464 – 471, 1996. 1380. Skov K, Mulvany MJ. Structure of renal afferent arterioles in the pathogenesis of hypertension. Acta Physiol Scand 181: 397– 405, 2004. 1381. Skov K, Mulvany MJ, Korsgaard N. Morphology of renal afferent arterioles in spontaneously hypertensive rats. Hypertension 20: 821– 827, 1992. 1382. Skov K, Nyengaard JR, Korsgaard N, Mulvany MJ. Number and size of renal glomeruli in spontaneously hypertensive rats. J Hypertens 12: 1373–1376, 1994. 1393. Song J, Lu Y, Lai EY, Wei J, Wang L, Chandrashekar K, Wang S, Shen C, Juncos LA, Liu R. Oxidative status in the macula densa modulates tubuloglomerular feedback responsiveness in Ang II-induced hypertension. Acta Physiol Oxf. In press. 1394. Sonnenberg H, Honrath U, Wilson DR. Effects of increased perfusion pressure on medullary collecting duct function. Can J Physiol Pharmacol 68: 402– 407, 1990. 1395. Sorensen CM, Giese I, Braunstein TH, Brasen JC, Salomonsson M, Holstein-Rathlou NH. Role of connexin 40 in the autoregulatory response of the afferent arteriole. Am J Phyisol Renal Physiol 303: F855–F863, 2012. 1396. Sorensen CM, Giese I, Braunstein TH, Holstein-Rathlou NH, Salomonsson M. Closure of multiple types of K⫹ channels is necessary to induce changes in renal vascular resistance in vivo in rats. Pflügers Arch 462: 655– 667, 2011. 1397. Sorensen CM, Leyssac PP, Skott O, Holstein-Rathlou NH. Role of the renin-angiotensin system in regulation and autoregulation of renal blood flow. Am J Physiol Regul Integr Comp Physiol 279: R1017–R1024, 2000. 1398. Sorensen CM, Leyssac PP, Skott O, Holstein-Rathlou NH. NO mediates downregulation of RBF after a prolonged reduction of renal perfusion pressure in SHR. Am J Physiol Regul Integr Comp Physiol 285: R329 –R338, 2003. 1399. Sorensen CM, Salomonsson M, Braunstein TH, Nielsen MS, Holstein-Rathlou NH. Connexin mimetic peptides fail to inhibit vascular conducted calcium responses in renal arterioles. Am J Physiol Regul Integr Comp Physiol 295: R840 –R847, 2008. 1400. Sosnovtseva OV, Pavlov AN, Mosekilde E, Holstein-Rathlou NH, Marsh DJ. Doublewavelet approach to study frequency and amplitude modulation in renal autoregulation. Phys Rev E Stat Nonlin Soft Matter Phys 70: 031915, 2004. 1401. Sosnovtseva OV, Pavlov AN, Mosekilde E, Yip KP, Holstein-Rathlou NH, Marsh DJ. Synchronization among mechanisms of renal autoregulation is reduced in hypertensive rats. Am J Physiol Renal Physiol 293: F1545–F1555, 2007. 1402. Sosnovtseva OV, Pavlov AN, Pavlova ON, Mosekilde E, Holstein-Rathlou NH. The effect of L-NAME on intra- and inter-nephron synchronization. Eur J Pharm Sci 36: 39 –50, 2009. 1403. Spassova MA, Hewavitharana T, Xu W, Soboloff J, Gill DL. A common mechanism underlies stretch activation and receptor activation of TRPC6 channels. Proc Natl Acad Sci USA 103: 16586 –16591, 2006. 1383. Slish DF, Welsh DG, Brayden JE. Diacylglycerol and protein kinase C activate cation channels involved in myogenic tone. Am J Physiol Heart Circ Physiol 283: H2196 – H2201, 2002. 1404. Sporkova A, Kopkan L, Varcabova S, Huskova Z, Hwang SH, Hammock BD, Imig JD, Kramer HJ, Cervenka L. Role of cytochrome P-450 metabolites in the regulation of renal function and blood pressure in 2-kidney 1-clip hypertensive rats. Am J Physiol Regul Integr Comp Physiol 300: R1468 –R1475, 2011. 1384. Smeda JS, Daneshtalab N. The effects of poststroke captopril and losartan treatment on cerebral blood flow autoregulation in SHRsp with hemorrhagic stroke. J Cereb Blood Flow Metab 31: 476 – 485, 2011. 1405. St Lezin E, Liu W, Wang JM, Yang Y, Qi N, Kren V, Zidek V, Kurtz TW, Pravenec M. Genetic analysis of rat chromosome 1 and the Sa gene in spontaneous hypertension. Hypertension 35: 225–230, 2000. 1385. Smeda JS, Lee RM, Forrest JB. Structural and reactivity alterations of the renal vasculature of spontaneously hypertensive rats prior to and during established hypertension. Circ Res 63: 518 –533, 1988. 1406. Stec DE, Trolliet MR, Krieger JE, Jacob HJ, Roman RJ. Renal cytochrome P4504A activity and salt sensitivity in spontaneously hypertensive rats. Hypertension 27: 1329 –1336, 1996. 1386. Smeda JS, Payne GW. Alterations in autoregulatory and myogenic function in the cerebrovasculature of Dahl salt-sensitive rats. Stroke 34: 1484 –1490, 2003. 1407. Steendahl J, Holstein-Rathlou NH, Sorensen CM, Salomonsson M. Effects of chloride channel blockers on rat renal vascular responses to angiotensin II and norepinephrine. Am J Physiol Renal Physiol 286: F323–F330, 2004. 1387. Smeda JS, van Vliet BN, King SR. Stroke-prone spontaneously hypertensive rats lose their ability to auto-regulate cerebral blood flow prior to stroke. J Hypertens 17: 1697–1705, 1999. 504 1408. Stefanovska A. Coupled oscillators. Complex but not complicated cardiovascular and brain interactions. IEEE Eng Med Biol Mag 26: 25–29, 2007. Physiol Rev • VOL 95 • APRIL 2015 • www.prv.org CARLSTRÖM ET AL. 1409. Stegbauer J, Vonend O, Oberhauser V, Sellin L, Rump LC. Angiotensin II receptor modulation of renal vascular resistance and neurotransmission in young and adult spontaneously hypertensive rats. Kidney Blood Press Res 28: 20 –26, 2005. 1410. Steiner RW, Tucker BJ, Leslie MS, Gushwa LC, Gifford J, Wilson CB, Blantz RC. Glomerular hemodynamics in moderate Goldblatt hypertension in the rat. Hypertension 4: 51–57, 1982. 1411. Steinhausen M, Baehr M, Dussel R. Vasomotion and vasoconstriction induced by a Ca-agonist in the split hydronephrotic kidney. Prog Appl Microvasc 14: 25–39, 1989. 1412. Steinhausen M, Ballantyne D, Fretschner M, Parekh N. Sex differences in autoregulation of juxtamedullary glomerular blood flow in hydronephrotic rats. Am J Physiol Renal Fluid Electrolyte Physiol 258: F863–F869, 1990. 1413. Steinhausen M, Blum M, Fleming JT, Holz FG, Parekh N, Wiegman DL. Visualization of renal autoregulation in the split hydronephrotic kidney of rats. Kidney Int 35: 1151–1160, 1989. 1414. Steinhausen M, Endlich K, Nobiling R, Parekh N, Schutt F. Electrically induced vasomotor responses and their propagation in rat renal vessels in vivo. J Physiol 505: 493–501, 1997. 1415. Steinhausen M, Snoei H, Parekh N, Baker R, Johnson PC. Hydronephrosis: a new method to visualize vas afferens, efferens, and glomerular network. Kidney Int 23: 794 – 806, 1983. 1416. Stepp DW, Merkus D, Nishikawa Y, Chilian WM. Nitric oxide limits coronary vasoconstriction by a shear stress-dependent mechanism. Am J Physiol Heart Circ Physiol 281: H796 –H803, 2001. 1430. Sward K, Albinsson S, Rippe C. Arterial dysfunction but maintained systemic blood pressure in cavin-1-deficient mice. PLoS One 9: e92428, 2014. 1431. Syntichaki P, Tavernarakis N. Genetic models of mechanotransduction: the nematode Caenorhabditis elegans. Physiol Rev 84: 1097–1153, 2004. 1432. Szentivanyi M Jr, Maeda CY, Cowley AW Jr. Local renal medullary L-NAME infusion enhances the effect of long-term angiotensin II treatment. Hypertension 33: 440 – 445, 1999. 1433. Takahashi N, Mori Y. TRP channels as sensors and signal integrators of redox status changes. Front Pharmacol 2: 1–11, 2011. 1434. Takata Y, Koga T, Kobayashi K, Takishita S, Fujishima M. Decrease in endothelium dependent hypotension in spontaneously hypertensive rats. Jpn Circ J 54: 183–191, 1990. 1435. Takenaka T, Epstein M, Forster H, Landry DW, Iijima K, Goligorsky MS. Attenuation of endothelin effects by a chloride channel inhibitor, indanyloxyacetic acid. Am J Physiol Renal Fluid Electrolyte Physiol 262: F799 –F806, 1992. 1436. Takenaka T, Forster H, De Micheli AG, Epstein M. Impaired myogenic responsiveness of renal microvessels in Dahl salt-sensitive rats. Circ Res 71: 471– 480, 1992. 1437. Takenaka T, Harrison-Bernard LM, Inscho EW, Carmines PK, Navar LG. Autoregulation of afferent arteriolar blood flow in juxtamedullary nephrons. Am J Physiol Renal Fluid Electrolyte Physiol 267: F879 –F887, 1994. 1438. Takenaka T, Inoue T, Kanno Y, Okada H, Hill CE, Suzuki H. Connexins 37 and 40 transduce purinergic signals mediating renal autoregulation. Am J Physiol Regul Integr Comp Physiol 294: R1–R11, 2008. 1417. Stern MD, Bowen PD, Parma R, Osgood RW, Bowman RL, Stein JH. Measurement of renal cortical and medullary blood flow by laser-Doppler spectroscopy in the rat. Am J Physiol Renal Fluid Electrolyte Physiol 236: F80 –F87, 1979. 1439. Takenaka T, Inoue T, Kanno Y, Okada H, Meaney KR, Hill CE, Suzuki H. Expression and role of connexins in the rat renal vasculature. Kidney Int 73: 415– 422, 2008. 1418. Sterzel RB, Luft FC, Gao Y, Schnermann J, Briggs JP, Ganten D, Waldherr R, Schnabel E, Kriz W. Renal disease and the development of hypertension in salt-sensitive Dahl rats. Kidney Int 33: 1119 –1129, 1988. 1440. Takenaka T, Inoue T, Ohno Y, Miyazaki T, Nishiyama A, Ishii N, Suzuki H. Elucidating mechanisms underlying altered renal autoregulation in diabetes. Am J Physiol Regul Integr Comp Physiol 303: R495–R504, 2012. 1419. Stoos BA, Garvin JL. Actions of nitric oxide on renal epithelial transport. Clin Exp Pharmacol Physiol 24: 591–594, 1997. 1441. Takenaka T, Inoue T, Okada H, Ohno Y, Miyazaki T, Chaston DJ, Hill CE, Suzuki H. Altered gap junctional communication and renal haemodynamics in Zucker fatty rat model of type 2 diabetes. Diabetologia 54: 2192–2201, 2011. 1420. Stoos BA, Metting PJ, Britton SL. Autoregulatory capacity in renal and mesenteric vasculatures of mineralocorticoid hypertensive dogs. Am J Physiol Heart Circ Physiol 261: H1205–H1213, 1991. 1421. Storch U, Mederos y Schnitzler M, Gudermann T. G protein-mediated stretch reception. Am J Physiol Heart Circ Physiol 302: H1241–H1249, 2012. 1422. Stowe N, Schnermann J, Hermle M. Feedback regulation of nephron filtration rate during pharmacologic interference with the renin-angiotensin and adrenergic systems in rats. Kidney Int 15: 473– 486, 1979. 1423. Strick DM, Fiksen-Olsen MJ, Lockhart JC, Roman RJ, Romero JC. Direct measurement of renal medullary blood flow in the dog. Am J Physiol Regul Integr Comp Physiol 267: R253–R259, 1994. 1424. Sun D, Cuevas AJ, Goltlinger K, Hwang SH, Hammock BD, Laniado SM, Huang A. Soluble epoxide hydrolase-dependent regulation of myogenic response and blood pressure. Am J Physiol Heart Circ Physiol 306: H1146 –H1153, 2014. 1425. Sun D, Huang A, Yan EH, Wu Z, Yan C, Kaminski PM, Oury TD, Wolin MS, Kaley G. Reduced release of nitric oxide to shear stress in mesenteric arteries of aged rats. Am J Physiol Heart Circ Physiol 286: H2249 –H2256, 2004. 1426. Sun D, Samuelson LC, Yang T, Huang Y, Paliege A, Saunders T, Briggs J, Schnermann J. Mediation of tubuloglomerular feedback by adenosine: evidence from mice lacking adenosin 1 receptors. Proc Natl Acad Sci USA 98: 9983–9988, 2001. 1427. Sun L, McArdle S, Chun M, Wolff DW, Pettinger WA. Cosegregation of the renin gene with an increase in mean arterial blood pressure in the F2 rats of SHR-WKY cross. Clin Exp Hypertens 15: 797– 805, 1993. 1428. Susic D, Frohlich ED, Kobori H, Shao W, Seth D, Navar LG. Salt-induced renal injury in SHRs is mediated by AT1 receptor activation. J Hypertens 29: 716 –723, 2011. 1429. Suzaki Y, Yoshizumi M, Kagami S, Nishiyama A, Ozawa Y, Kyaw M, Izawa Y, Kanematsu Y, Tsuchiya K, Tamaki T. BMK1 is activated in glomeruli of diabetic rats and in mesangial cells by high glucose conditions. Kidney Int 65: 1749 –1760, 2004. 1442. Takenaka T, Kanno Y, Kitamura Y, Hayashi K, Suzuki H, Saruta T. Role of chloride channels in afferent arteriolar constriction. Kidney Int 50: 864 – 872, 1996. 1443. Takenaka T, Ohno Y, Hayashi K, Saruta T, Suzuki H. Governance of arteriolar oscillation by ryanodine receptors. Am J Physiol Regul Integr Comp Physiol 285: R125– R131, 2003. 1444. Takenaka T, Okada H, Kanno Y, Inoue T, Ryuzaki M, Nakamoto H, Kawachi H, Shimizu F, Suzuki H. Exogenous 5’-nucleotidase improves glomerular autoregulation in Thy-1 nephritic rats. Am J Physiol Renal Physiol 290: F844 –F853, 2006. 1445. Takenaka T, Suzuki H, Ikenaga H, Itaya Y, Yamakawa H, Sakamaki Y, Saruta T. Effects of a calcium channel blocker, nicardipine, on pressure-natriuresis in Dahl salt-sensitive rats. Clin Exp Hypertens 16: 77– 88, 1994. 1446. Takenaka T, Suzuki H, Okada H, Hayashi K, Kanno Y, Saruta T. Mechanosensitive cation channels mediate afferent arteriolar myogenic constriction in the isolated rat kidney. J Physiol 511: 245–253, 1998. 1447. Takenaka T, Suzuki H, Okada H, Hayashi K, Ozawa Y, Saruta T. Biophysical signals underlying myogenic responses in rat interlobular artery. Hypertension 32: 1060 – 1065, 1998. 1448. Takenaka T, Suzuki H, Okada H, Inoue T, Kanno Y, Ozawa Y, Hayashi K, Saruta T. Transient receptor potential channels in rat renal microcirculation: actions of angiotensin II. Kidney Int 62: 558 –565, 2002. 1449. Tang G, Wu L, Wang R. Interaction of hydrogen sulfide with ion channels. Clin Exp Pharmacol Physiol 37: 753–763, 2009. 1450. Tang L, Parker M, Fei Q, Loutzenhiser R. Afferent arteriolar adenosine A2a receptors are coupled to KATP in in vitro perfused hydronephrotic rat kidney. Am J Physiol Renal Physiol 277: F926 –F933, 1999. 1451. Tapia E, Franco M, Sanchez-Lozada LG, Soto V, Avila-Casado C, Santamaria J, Quiroz Y, Rodriguez-Iturbe B, Herrera-Acosta J. Mycophenolate mofetil prevents Physiol Rev • VOL 95 • APRIL 2015 • www.prv.org 505 RENAL AUTOREGULATORY MECHANISMS arteriolopathy and renal injury in subtotal ablation despite persistent hypertension. Kidney Int 63: 994 –1002, 2003. 1452. Tapia E, Gabbai FB, Calleja C, Franco M, Cermeno JL, Bobadilla NA, Perez JM, Alvarado JA, Herrera-Acosta J. Determinants of renal damage in rats with systemic hypertension and partial renal ablation. Kidney Int 38: 642– 648, 1990. 1453. Taylor NE, Glocka P, Liang M, Cowley AW Jr. NADPH oxidase in the renal medulla causes oxidative stress and contributes to salt-sensitive hypertension in Dahl S rats. Hypertension 47: 692– 698, 2006. 1454. Terashvili M, Pratt PF, Gebremedhin D, Narayanan J, Harder DR. Reactive oxygen species cerebral autoregulation in health and disease. Pediatr Clin North Am 53: 1029 –1037, 2006. 1455. Textor SC, Wilcox CS. Renal artery stenosis: a common, treatable cause of renal failure? Annu Rev Med 52: 421– 442, 2001. 1456. Thai TL, Churchill GC, Arendshorst WJ. NAADP receptors mediate calcium signaling stimulated by endothelin-1 and norepinephrine in renal afferent arterioles. Am J Physiol Renal Physiol 297: F510 –F516, 2009. 1457. Thai TL, Fellner SK, Arendshorst WJ. ADP-ribosyl cyclase and ryanodine receptor activity contribute to basal renal vasomotor tone and agonist-induced renal vasoconstriction in vivo. Am J Physiol Renal Physiol 293: F1107–F1114, 2007. 1471. Tolbert EM, Weisstuch J, Feiner HD, Dworkin LD. Onset of glomerular hypertension with aging precedes injury in the spontaneously hypertensive rat. Am J Physiol Renal Physiol 278: F839 –F846, 2000. 1472. Tolins JP, Shultz PJ, Raij L, Brown DM, Mauer SM. Abnormal renal hemodynamic response to reduced renal perfusion pressure in diabetic rats: role of NO. Am J Physiol Renal Fluid Electrolyte Physiol 265: F886 –F895, 1993. 1473. Toma I, Bansal E, Meer EJ, Kang JJ, Vargas SL, Peti-Peterdi J. Connexin 40 and ATP-dependent intercellular calcium wave in renal glomerular endothelial cells. Am J Physiol Regul Integr Comp Physiol 294: R1769 –R1776, 2008. 1474. Tomoda F, Bergstrom G, Evans RG, Anderson WP. Evidence for decreased structurally determined preglomerular resistance in the young spontaneously hypertensive rat after 4 weeks of renal denervation. J Hypertens 15: 1187–1195, 1997. 1475. Toney GM, Vallon V, Stockand JD. Intrinsic control of sodium excretion in the distal nephron by inhibitory purinergic regulation of the epithelial Na⫹ channel. Curr Opin Nephrol Hypertens 21: 52– 60, 2012. 1476. Tornel J, Madrid MI, Garcia-Salom M, Wirth KJ, Fenoy FJ. Role of kinins in the control of renal papillary blood flow, pressure natriuresis, and arterial pressure. Circ Res 86: 589 –595, 2000. 1477. Touyz RM, Schiffrin EL. Reactive oxygen species in vascular biology: implications in hypertension. Histochem Cell Biol 122: 339 –352, 2004. 1458. Thomsen AB, Kim S, Aalbaek F, Aalkjaer C, Boedtkjer E. Intracellular acidification alters myogenic responsiveness and vasomotion of mouse middle cerebral arteries. J Cereb Blood Flow Metab 34: 161–168, 2014. 1478. Touyz RM, Turgeon A, Schiffrin EL. Endothelin-A-receptor blockade improves renal function and doubles the lifespan of stroke-prone spontaneously hypertensive rats. J Cardiovasc Pharmacol 36: S300 –S304, 2000. 1459. Thomson S, Bao D, Deng A, Vallon V. Adenosine formed by 5’-nucleotidase mediates tubuloglomerular feedback. J Clin Invest 106: 289 –298, 2000. 1479. Traynor TR, Schnermann J. Renin-angiotensin system dependence of renal hemodynamics in mice. J Am Soc Nephrol 10 Suppl 11: S184 –S188, 1999. 1460. Thomson SC, Deng A. Cyclic GMP mediates influence of macula densa nitric oxide over tubuloglomerular feedback. Kidney Blood Press Res 26: 10 –18, 2003. 1480. Traynor TR, Yang T, Huang YG, Arend L, Oliverio MI, Coffman T, Briggs JP, Schnermann J. Inhibition of adenosine-1 receptor-mediated preglomerular vasoconstriction in AT1A receptor-deficient mice. Am J Physiol Renal Physiol 275: F922–F927, 1998. 1461. Thomson SC, Deng A, Komine N, Hammes JS, Blantz RC, Gabbai FB. Early diabetes as a model for testing the regulation of juxtaglomerular NOS I. Am J Physiol Renal Physiol 287: F732–F738, 2004. 1462. Thorup C, Ollerstam A, Persson AE, Torffvit O. Increased tubuloglomerular feedback reactivity is associated with increased NO production in the streptozotocindiabetic rat. J Diabetes Complications 14: 46 –52, 2000. 1481. Traynor TR, Yang T, Huang YG, Krege JH, Briggs JP, Smithies O, Schnermann J. Tubuloglomerular feedback in ACE-deficient mice. Am J Physiol Renal Physiol 276: F751–F757, 1999. 1482. Trebak M. STIM/Orai signalling complexes in vascular smooth muscle. J Physiol 590: 4201– 4208, 2012. 1463. Thorup C, Persson AE. Inhibition of locally produced nitric oxide resets tubuloglomerular feedback mechanism. Am J Physiol Renal Fluid Electrolyte Physiol 267: F606 – F611, 1994. 1483. Trippodo NC, Frohlich ED. Similarities of genetic (spontaneous) hypertension. Man and rat. Circ Res 48: 309 –319, 1981. 1464. Thorup C, Persson AE. Impaired effect of nitric oxide synthesis inhibition on tubuloglomerular feedback in hypertensive rats. Am J Physiol Renal Fluid Electrolyte Physiol 271: F246 –F252, 1996. 1484. Troncoso Brindeiro CM, Fallet RW, Lane PH, Carmines PK. Potassium channel contributions to afferent arteriolar tone in normal and diabetic rat kidney. Am J Physiol Renal Physiol 295: F171–F178, 2008. 1465. Thurau K. Autoregulation of renal blood flow and glomerular filtration rate, including data on tubular and peritubular capillary pressures and vessel wall tension. Circ Res 15 Suppl: 132–141, 1964. 1485. Troncoso Brindeiro CM, Lane PH, Carmines PK. Tempol prevents altered K⫹ channel regulation of afferent arteriolar tone in diabetic rat kidney. Hypertension 59: 657– 664, 2012. 1466. Thurau K, Schnermann J. Die Natriumkonzentration an den Macula densa Zellen als regulierender Faktor fuer das Glomerulumfiltrat (Mikropunktionsversuche). Klin Wochenschr 43: 410 – 413, 1965. 1486. Trottier G, Triggle CR, O’Neill SK, Loutzenhiser R. Cyclic GMP-dependent and cyclic GMP-independent actions of nitric oxide on the renal afferent arteriole. Br J Pharmacol 125: 563–569, 1998. 1467. Tobia AJ, Walsh GM, Tadepalli S, Lee JY. Unaltered distribution of cardiac output in the conscious young spontaneously hypertensive rat: evidence for uniform elevation of regional vascular resistances. Blood Vessels 11: 287–294, 1974. 1487. Tucker BJ, Anderson CM, Thies RS, Collins RC, Blantz RC. Glomerular hemodynamic alterations during acute hyperinsulinemia in normal and diabetic rats. Kidney Int 42: 1160 –1168, 1992. 1468. Tobian L, Johnson MA, Lange J, Magraw S. Effect of varying perfusion pressures on the output of sodium and renin and the vascular resistance in kidneys of rats with “post-salt” hypertension and Kyoto spontaneous hypertension. Circ Res 36: 162– 170, 1975. 1469. Tobin AA, Joseph BK, Al-Kindi HN, Albarwani S, Madden JA, Nemetz LT, Rusch NJ, Rhee SW. Loss of cerebrovascular Shaker-type K⫹ channels: a shared vasodilator defect of genetic and renal hypertensive rats. Am J Physiol Heart Circ Physiol 297: H293–H303, 2009. 1470. Tojo A, Onozato ML, Fukuda S, Asaba K, Kimura K, Fujita T. Nitric oxide generated by nNOS in the macula densa regulates the afferent arteriolar diameter in rat kidney. Med Electron Microsc 37: 236 –241, 2004. 506 1488. Tucker BJ, Blantz RC. An analysis of the determinants of nephron filtration rate. Am J Physiol Renal Fluid Electrolyte Physiol 232: F477–F483, 1977. 1489. Tucker BJ, Blantz RC. Effect of furosemide administration on glomerular and tubular dynamics in the rat. Kidney Int 26: 112–121, 1984. 1490. Tucker BJ, Mundy CA, Blantz RC. Can causality be determined from proximal tubular reabsorption and peritubular physical factors? Am J Physiol Renal Fluid Electrolyte Physiol 250: F169 –F175, 1986. 1491. Turkstra E, Boer P, Braam B, Koomans HA. Increased availability of nitric oxide leads to enhanced nitric oxide dependency of tubuloglomerular feedback in the contralateral kidney of rats with 2-kidney, 1-clip Goldblatt hypertension. Hypertension 34: 679 – 684, 1999. Physiol Rev • VOL 95 • APRIL 2015 • www.prv.org CARLSTRÖM ET AL. 1492. Turkstra E, Braam B, Koomans HA. Losartan attenuates modest but not strong renal vasoconstriction induced by nitric oxide inhibition. J Cardiovasc Pharmacol 32: 593– 600, 1998. 1512. Vallon V, Richter K, Huang DY, Rieg T, Schnermann J. Functional consequences at the single-nephron level of the lack of adenosine A1 receptors and tubuloglomerular feedback in mice. Pflügers Arch 448: 214 –221, 2004. 1493. Turkstra E, Braam B, Koomans HA. Nitric oxide release as an essential mitigating step in tubuloglomerular feedback: observations during intrarenal nitric oxide clamp. J Am Soc Nephrol 9: 1596 –1603, 1998. 1513. Vallon V, Schnermann J. Tubuloglomerular feedback. Methods Mol Med 86: 429 – 441, 2003. 1494. Turkstra E, Braam B, Koomans HA. Impaired renal blood flow autoregulation in two-kidney, one-clip hypertensive rats is caused by enhanced activity of nitric oxide. J Am Soc Nephrol 11: 847– 855, 2000. 1495. Turkstra E, Braam B, Koomans HA. Normal TGF responsiveness during chronic treatment with angiotensin-converting enzyme inhibition: role of AT1 receptors. Hypertension 36: 818 – 823, 2000. 1496. Turner SR, Macdonald JA. Novel contributions of the smoothelin-like 1 protein in vascular smooth muscle contraction and its potential involvement in myogenic tone. Microcirculation 21: 249 –258, 2014. 1497. Turoni CM, Reynoso HA, Maranon RO, Coviello A, Peral De BM. Structural changes in the renal vasculature in streptozotocin-induced diabetic rats without hypertension. Nephron Physiol 99: 50 –57, 2005. 1498. Uchino K, Frohlich ED, Nishikimi T, Isshiki T, Kardon MB. Spontaneously hypertensive rats demonstrate increased renal vascular alpha 1-adrenergic receptor responsiveness. Am J Physiol Regul Integr Comp Physiol 260: R889 –R893, 1991. 1499. Uhrenholt TR, Schjerning J, Vanhoutte PM, Jensen BL, Skott O. Intercellular calcium signaling and nitric oxide feedback during constriction of rabbit renal afferent arterioles. Am J Physiol Renal Physiol 292: F1124 –F1131, 2007. 1500. Umesh A, Thompson MA, Chini EN, Yip KP, Sham JS. Integrin ligands mobilize Ca2⫹ from ryanodine receptor-gated stores and lysosome-related acidic organelles in pulmonary arterial smooth muscle cells. J Biol Chem 281: 34312–34323, 2006. 1501. Ungvari Z, Csiszar A, Huang A, Kaminski PM, Wolin MS, Koller A. High pressure induces superoxide production in isolated arteries via protein kinase C-dependent activation of NAD(P)H oxidase. Circulation 108: 1253–1258, 2003. 1502. Ungvari Z, Koller A. Endothelin and prostaglandin H2/thromboxane A2 enhance myogenic constriction in hypertension by increasing Ca2⫹ sensitivity of arteriolar smooth muscle. Hypertension 36: 856 – 861, 2000. 1503. Ungvari Z, Koller A. Selected contribution: NO released to flow reduces myogenic tone of skeletal muscle arterioles by decreasing smooth muscle Ca2⫹ sensitivity. J Appl Physiol 91: 522–527, 2001. 1504. Ungvari Z, Pacher P, Kecskemeti V, Papp G, Szollar L, Koller A. Increased myogenic tone in skeletal muscle arterioles of diabetic rats. Possible role of increased activity of smooth muscle Ca2⫹ channels and protein kinase C. Cardiovasc Res 43: 1018 –1028, 1999. 1505. Unlap MT, Bates E, Williams C, Komlosi P, Williams I, Kovacs G, Siroky B, Bell PD. Na⫹/Ca2⫹ exchanger: target for oxidative stress in salt-sensitive hypertension. Hypertension 42: 363–368, 2003. 1506. Uriu K, Kaizu K, Qie YL, Kai K, Ito A, Ikeda M, Hashimoto O, Sun XF, Morita E, Eto S. Renal hemodynamics in Otsuka Long-Evans Tokushima fatty rat, a model rat of spontaneous non-insulin-dependent diabetes mellitus with obesity. J Lab Clin Med 134: 483– 491, 1999. 1507. Uyehara CF, Gellai M. Impairment of renal function precedes establishment of hypertension in spontaneously hypertensive rats. Am J Physiol Regul Integr Comp Physiol 265: R943–R950, 1993. 1508. Vallon V. Tubuloglomerular feedback in the kidney: insights from gene-targeted mice. Pflügers Arch 445: 470 – 476, 2003. 1509. Vallon V, Blantz RC, Thomson S. Homeostatic efficiency of tubuloglomerular feedback is reduced in established diabetes mellitus in rats. Am J Physiol Renal Fluid Electrolyte Physiol 269: F876 –F883, 1995. 1510. Vallon V, Muhlbauer B, Osswald H. Adenosine and kidney function. Physiol Rev 86: 901–940, 2006. 1511. Vallon V, Osswald H. Adenosine receptors and the kidney. Handb Exp Pharmacol 443– 470, 2009. 1514. Vallon V, Schroth J, Satriano J, Blantz RC, Thomson SC, Rieg T. Adenosine A(1) receptors determine glomerular hyperfiltration and the salt paradox in early streptozotocin diabetes mellitus. Nephron Physiol 111: 30 –38, 2009. 1515. Vallon V, Traynor TR, Barajas L, Huang YG, Briggs JP, Schnermann J. Feedback control of glomerular vascular tone in neuronal nitric oxide synthase knockout mice. Am J Soc Nephrol 12: 1599 –1606, 2001. 1516. Van der Mark J, Kline RL. Altered pressure natriuresis in chronic angiotensin II hypertension in rats. Am J Physiol Regul Integr Comp Physiol 266: R739 –R748, 1994. 1517. Van Dijk PC, Jager KJ, Stengel B, Gronhagen-Riska C, Feest TG, Briggs JD. Renal replacement therapy for diabetic end-stage renal disease: data from 10 registries in Europe (1991–2000). Kidney Int 67: 1489 –1499, 2005. 1518. Van Dijk SJ, Specht PA, Lazar J, Jacob HJ, Provoost AP. Renal damage susceptibility and autoregulation in RF-1 and RF-5 congenic rats. Nephron Exp Nephrol 101: e59 – e66, 2005. 1519. Van Dijk SJ, Specht PA, Lazar J, Jacob HJ, Provoost AP. Absence of an interaction between the Rf-1 and Rf-5 QTLs influencing susceptibility to renal damage in rats. Nephron Exp Nephrol 104: e96 – e102, 2006. 1520. Van Dijk SJ, Specht PA, Lazar J, Jacob HJ, Provoost AP. Synergistic QTL interactions between Rf-1 and Rf-3 increase renal damage susceptibility in double congenic rats. Kidney Int 69: 1369 –1376, 2006. 1521. Van Dijk SJ, Specht PA, Lutz MM, Lazar J, Jacob HJ, Provoost AP. Interaction between Rf-1 and Rf-4 quantitative trait loci increases susceptibility to renal damage in double congenic rats. Kidney Int 68: 2462–2472, 2005. 1522. Van Dokkum RP, Sun CW, Provoost AP, Jacob HJ, Roman RJ. Altered renal hemodynamics and impaired myogenic responses in the fawn-hooded rat. Am J Physiol Regul Integr Comp Physiol 276: R855–R863, 1999. 1523. Van Rodijnen WF, van Lambalgen TA, Tangelder G, Van Dokkum RP, Provoost AP, Ter Wee PM. Reduced reactivity of renal microvessels to pressure and angiotensin II in Fawn-Hooded rats. Hypertension 39: 111–115, 2002. 1524. Van Rodijnen WF, van Lambalgen TA, van Wijhe MH, Tangelder GJ, Ter Wee PM. Renal microvascular actions of angiotensin II fragments. Am J Physiol Renal Physiol 283: F86 –F92, 2002. 1525. VanBavel E, Giezeman MJ, Mooij T, Spaan JA. Influence of pressure alterations on tone and vasomotion of isolated mesenteric small arteries of the rat. J Physiol 436: 371–383, 1991. 1526. Vaneckova I. Function of the isolated perfused kidney in young and adult spontaneously hypertensive and dahl salt-sensitive rats. Kidney Blood Press Res 25: 315–321, 2002. 1527. Vavrinec P, Henning RH, Goris M, Landheer SW, Buikema H, Van Dokkum RP. Renal myogenic constriction protects the kidney from age-related hypertensive renal damage in the Fawn-Hooded rat. J Hypertens 31: 1637–1645, 2013. 1528. Vavrinec P, Van Dokkum RP, Goris M, Buikema H, Henning RH. Losartan protects mesenteric arteries from ROS-associated decrease in myogenic constriction following 5/6 nephrectomy. J Renin Angiotensin Aldosterone Syst 12: 184 –199, 2011. 1529. Vaziri ND. Roles of oxidative stress and antioxidant therapy in chronic kidney disease and hypertension. Curr Opin Nephrol Hypertens 13: 93–99, 2004. 1530. Veerareddy S, Cooke CL, Baker PN, Davidge ST. Gender differences in myogenic tone in superoxide dismutase knockout mouse: animal model of oxidative stress. Am J Physiol Heart Circ Physiol 287: H40 –H45, 2004. 1531. Venuto RC, O’Dorisio T, Ferris TF, Stein JH. Prostaglandins and renal function. II. The effect of prostaglandin inhibition on autoregulation of blood flow in the intact kidney of the dog. Prostaglandins 9: 817– 828, 1975. 1532. Verbeuren TJ, Vallez MO, Lavielle G, Bouskela E. Activation of thromboxane receptors and the induction of vasomotion in the hamster cheek pouch microcirculation. Br J Pharmacol 122: 859 – 866, 1997. Physiol Rev • VOL 95 • APRIL 2015 • www.prv.org 507 RENAL AUTOREGULATORY MECHANISMS 1533. Vermeij A, Meel-van den Abeelen AS, Kessels RP, van Beek AH, Claassen JA. Verylow-frequency oscillations of cerebral hemodynamics and blood pressure are affected by aging and cognitive load. Neuroimage 85: 608 – 615, 2014. 1534. Verseput GH, Braam B, Provoost AP, Koomans HA. Tubuloglomerular feedback and prolonged ACE-inhibitor treatment in the hypertensive fawn-hooded rat. Nephrol Dial Transplant 13: 893– 899, 1998. 1535. Verseput GH, Provoost AP, Braam BB, Weening JJ, Koomans HA. Angiotensinconverting enzyme inhibition in the prevention and treatment of chronic renal damage in the hypertensive fawn-hooded rat. J Am Soc Nephrol 8: 249 –259, 1997. 1555. Wang H, D’Ambrosio MA, Garvin JL, Ren Y, Carretero OA. Connecting tubule glomerular feedback in hypertension. Hypertension 62: 738 –745, 2013. 1556. Wang H, Garvin JL, Carretero OA. Angiotensin II enhances tubuloglomerular feedback via luminal AT(1) receptors on the macula densa. Kidney Int 60: 1851–1857, 2001. 1557. Wang H, Garvin JL, D’Ambrosio MA, Falck JR, Leung P, Liu R, Ren Y, Carretero OA. Heme oxygenase metabolites inhibit tubuloglomerular feedback in vivo. Am J Physiol Heart Circ Physiol 300: H1320 –H1326, 2011. 1536. Vetri F, Menicucci D, Lapi D, Gemignani A, Colantuoni A. Pial arteriolar vasomotion changes during cortical activation in rats. Neuroimage 38: 25–33, 2007. 1558. Wang H, Garvin JL, D’Ambrosio MA, Ren Y, Carretero OA. Connecting tubule glomerular feedback antagonizes tubuloglomerular feedback in vivo. Am J Physiol Renal Physiol 299: F1374 –F1378, 2010. 1537. Vettoretti S, Ochodnicky P, Buikema H, Henning RH, Kluppel CA, de Zeeuw D, Van Dokkum RP. Altered myogenic constriction and endothelium-derived hyperpolarizing factor-mediated relaxation in small mesenteric arteries of hypertensive subtotally nephrectomized rats. J Hypertens 24: 2215–2223, 2006. 1559. Wang H, Smeda JS, Lee RM. Prevention of stroke and preservation of the functions of cerebral arteries by treatment with perindopril in stroke-prone spontaneously hypertensive rats. Can J Physiol Pharmacol 76: 26 –34, 1998. 1538. Vio CP, Figueroa CD, Caorsi I. Anatomical relationship between kallikrein-containing tubules and the juxtaglomerular apparatus in the human kidney. Am J Hypertens 1: 269 –271, 1988. 1539. Vitzhum H, Abt I, Einhellig S, Kurtz A. Gene expression of prostanoid forming enzymes along the rat nephron. Kidney Int 62: 1570 –1581, 2002. 1560. Wang T. Role of iNOS and eNOS in modulating proximal tubule transport and acid-base balance. Am J Physiol Renal Physiol 283: F658 –F662, 2002. 1561. Wang T, Inglis FM, Kalb RG. Defective fluid and HCO3⫺ absorption in proximal tubule of neuronal nitric oxide synthase-knockout mice. Am J Physiol Renal Physiol 279: F518 –F524, 2000. 1540. Vitzthum H, Weiss B, Bachleitner W, Kramer BK, Kurtz A. Gene expression of adenosine receptors along the nephron. Kidney Int 65: 1180 –1190, 2004. 1562. Wang T, Kobayashi Y, Nabika T, Takabatake T. Enhanced sympathetic control of renal function in rats congenic for the hypertension-related region on chromosome 1. Clin Exp Pharmacol Physiol 32: 1055–1060, 2005. 1541. Volhard F. Clinical aspects of Bright s disease. In: The Kidney in Health and Disease, edited by Berglund H, Medes G, and Huber GL. Philadelphia, PA: Lea & Febiger, 1935, p. 665– 688. 1563. Wang X, Ajikobi DO, Salevsky FC, Cupples WA. Impaired myogenic autoregulation in kidneys of Brown Norway rats. Am J Physiol Renal Physiol 278: F962–F969, 2000. 1542. Vyas SJ, Jackson EK. Angiotensin II: enhanced renal responsiveness in young genetically hypertensive rats. J Pharmacol Exp Ther 273: 768 –777, 1995. 1564. Wang X, Aukland K, Bostad L, Iversen BM. Autoregulation of total and zonal glomerular filtration rate in spontaneously hypertensive rats with mesangiolysis. Kidney Blood Press Res 20: 11–17, 1997. 1543. Wagenfeld L, von Domarus F, Weiss S, Klemm M, Richard G, Zeitz O. The effect of reactive oxygen species on the myogenic tone of rat ophthalmic arteries with and without endothelium. Graefes Arch Clin Exp Ophthalmol 251: 2339 –2344, 2013. 1544. Wagner AJ, Holstein-Rathlou NH, Marsh DJ. Internephron coupling by conducted vasomotor responses in normotensive and spontaneously hypertensive rats. Am J Physiol Renal Physiol 272: F372–F379, 1997. 1565. Wang X, Aukland K, Iversen BM. Autoregulation of total and zonal glomerular filtration rate in spontaneously hypertensive rats during antihypertensive therapy. J Cardiovasc Pharmacol 28: 833– 841, 1996. 1566. Wang X, Aukland K, Iversen BM. Acute effects of angiotensin II receptor antagonist on autoregulation of zonal glomerular filtration rate in renovascular hypertensive rats. Kidney Blood Press Res 20: 225–232, 1997. 1545. Wagner C. Function of connexins in the renal circulation. Kidney Int 73: 547–555, 2008. 1546. Wagner C, de Wit C, Kurtz L, Grunberger C, Kurtz A, Schweda F. Connexin 40 is essential for the pressure control of renin synthesis and secretion. Circ Res 100: 556 –563, 2007. 1547. Walker M, Harrison-Bernard LM, Cook AK, Navar LG. Dynamic interaction between myogenic and TGF mechanisms in afferent arteriolar blood flow autoregulation. Am J Physiol Renal Physiol 279: F858 –F865, 2000. 1567. Wang X, Aukland K, Ofstad J, Iversen BM. Autoregulation of zonal glomerular filtration rate and renal blood flow in spontaneously hypertensive rats. Am J Physiol Renal Fluid Electrolyte Physiol 269: F515–F521, 1995. 1568. Wang X, Breaks J, Loutzenhiser K, Loutzenhiser R. Effects of inhibition of the Na⫹/ K⫹/2Cl⫺ cotransporter on myogenic and angiotensin II responses of the rat afferent arteriole. Am J Physiol Renal Physiol 292: F999 –F1006, 2007. 1548. Walsh MP, Cole WC. The role of actin filament dynamics in the myogenic response of cerebral resistance arteries. J Cereb Blood Flow Metab 33: 1–12, 2013. 1569. Wang X, Cupples WA. Interaction between nitric oxide and renal myogenic autoregulation in normotensive and hypertensive rats. Can J Physiol Pharmacol 79: 238 – 245, 2001. 1549. Wang CT, Chin SY, Navar LG. Impairment of pressure-natriuresis and renal autoregulation in ANG II-infused hypertensive rats. Am J Physiol Renal Physiol 279: F319 – F325, 2000. 1570. Wang X, Cupples WA. Brown Norway rats show impaired nNOS-mediated information transfer in renal autoregulation. Can J Physiol Pharmacol 87: 29 –36, 2009. 1550. Wang CT, Zou LX, Navar LG. Renal responses to AT1 blockade in angiotensin II-induced hypertensive rats. J Am Soc Nephrol 8: 535–542, 1997. 1571. Wang X, Loutzenhiser R. Determinants of renal microvascular repsonse to ACh: afferent and efferent arteriolar actions of EDHF. Am J Physiol Renal Physiol 282: F124 –F132, 2002. 1551. Wang D, Borrego-Conde L, Falck JR, Sharma KK, Wilcox CS, Umans JG. Contributions of NO, EDHF and EETs to endothelium-dependent relaxation in rabbit renal afferent arterioles. Kidney Int 63: 2187–2193, 2003. 1552. Wang D, Chen Y, Chabrashvili T, Aslam S, Borrego L, Umans J, Wilcox CS. Role of oxidative stress in endothelial dysfunction and enhanced responses to Ang II of afferent arterioles from rabbits infused with Ang II. J Am Soc Nephrol 14: 2783–2789, 2003. 1572. Wang X, Loutzenhiser RD, Cupples WA. Frequency modulation of renal myogenic autoregulation by perfusion pressure. Am J Physiol Regul Integr Comp Physiol 293: R1199 –R1204, 2007. 1573. Wang X, Salevsky FC, Cupples WA. Nitric oxide, atrial natriuretic factor, and dynamic renal autoregulation. Can J Physiol Pharmacol 77: 777–786, 1999. 1553. Wang H, Carretero OA, Garvin JL. Nitric oxide produced by THAL nitrc oxide synthase inhibitis TGF. Hypertension 39: 662– 666, 2002. 1574. Wang X, Takeya K, Aaronson PI, Loutzenhiser K, Loutzenhiser R. Effects of amiloride, benzamil, and alterations in extracellular Na⫹ on the rat afferent arteriole and its myogenic response. Am J Physiol Renal Physiol 295: F272–F282, 2008. 1554. Wang H, D’Ambrosio MA, Garvin JL, Ren Y, Carretero OA. Connecting tubule glomerular feedback mediates acute tubuloglomerular feedback resetting. Am J Physiol Renal Physiol 302: F1300 –F1304, 2012. 1575. Wang YX, Brooks DP, Edwards RM. Attenuated glomerular cGMP production and renal vasodilation in streptozotocin-induced diabetic rats. Am J Physiol Regul Integr Comp Physiol 264: R952–R956, 1993. 508 Physiol Rev • VOL 95 • APRIL 2015 • www.prv.org CARLSTRÖM ET AL. 1576. Watanabe J, Horiguchi S, Karibe A, Keitoku M, Takeuchi M, Satoh S, Takishima T, Shirato K. Effects of ryanodine on development of myogenic response in rat small skeletal muscle arteries. Cardiovasc Res 28: 480 – 484, 1994. 1598. White JV, Finco DR, Crowell WA, Brown SA, Hirakawa DA. Effect of dietary protein on functional, morphologic, and histologic changes of the kidney during compensatory renal growth in dogs. Am J Vet Res 52: 1357–1365, 1991. 1577. Watanabe J, Karibe A, Horiguchi S, Keitoku M, Satoh S, Takishima T, Shirato K. Modification of myogenic intrinsic tone and [Ca2⫹]i of rat isolated arterioles by ryanodine and cyclopiazonic acid. Circ Res 73: 465– 472, 1993. 1599. White TW, Paul DL, Goodenough DA, Bruzzone R. Functional analysis of selective interactions among rodent connexins. Mol Biol Cell 6: 459 – 470, 1995. 1578. Waugh WH. Circulatory autoregulation in the fully isolated kidney and in the humorally supported, isolated kidney. Circ Res 15 Suppl: 156 –169, 1964. 1600. Wickramasekera NT, Gebremedhin D, Carver KA, Vakeel P, Ramchandran R, Schuett A, Harder DR. Role of dual-specificity protein phosphatase-5 in modulating the myogenic response in rat cerebral arteries. J Appl Physiol 114: 252–261, 2013. 1579. Weichert W, Paliege A, Provoost AP, Bachmann S. Upregulation of juxtaglomerular NOS1 and COX-2 precedes glomerulosclerosis in fawn-hooded hypertensive rats. Am J Physiol Renal Physiol 280: F706 –F714, 2001. 1601. Wiederhielm CA, Bouskela E, Heald R, Black L. A method for varying arterial and venous pressures in intact, unanesthetized mammals. Microvasc Res 18: 124 –128, 1979. 1580. Weihprecht H, Lorenz JN, Briggs JP, Schnermann J. Synergistic effects of angiotensin and adenosine in the renal microvasculature. Am J Physiol Renal Fluid Electrolyte Physiol 266: F227–F239, 1994. 1602. Wilcox CS. Role of macula densa NOS in tubuloglomerular feedback. Nephrol Hypertens 7: 443– 449, 1998. 1581. Welch WJ. Effects of isoprostane on tubuloglomerular feedback: roles of TP receptors, NOS, and salt intake. Am J Physiol Renal Physiol 288: F757–F762, 2005. 1582. Welch WJ, Baumgärtl H, Lübbers D, Wilcox CS. Nephron pO2 and renal oxygen usage in the hypertensive rat kidney. Kidney Int 59: 230 –237, 2001. 1583. Welch WJ, Baumgärtl H, Lübbers D, Wilcox CS. Renal oxygenation defects in the spontaneously hypertensive rat: role of AT1 receptors. Kidney Int 63: 202–208, 2003. 1584. Welch WJ, Chabrashvili T, Solis G, Chen Y, Gill P, Aslam S, Wang X, Ji H, Sandberg K, Jose P, Wilcox CS. Role of extracellular superoxide dismutase in the mouse angiotensin slow pressor response. Hypertension 48: 934 –941, 2006. 1585. Welch WJ, Mendonca M, Blau J, Karber A, Dennehy K, Lao Y, José P, Wilcox CS. Antihypertensive response to prolonged tempol in the spontaneously hypertensive rat. Kidney Int 68: 179 –187, 2005. 1586. Welch WJ, Schmidt HHHW, Murad F, Gross SS, Taylor G, Levi R, Wilcox CS. Nitric oxide synthase in macula densa: localization and functional studies. In: The Biology of Nitric Oxide. 1. Physiologic and Clinical Aspects, edited by Moncada S, Marletta MA, Hibbs JB, and Higgs EA. London: Portland, 1992. 1603. Wilcox CS. Redox regulation of the afferent arteriole and tubuloglomerular feedback. Acta Physiol Scand 179: 217–223, 2003. 1604. Wilcox CS. Oxidative stress and nitric oxide deficiency in the kidney: a critical link to hypertension? Am J Physiol Regul Integr Comp Physiol 289: R913–R935, 2005. 1605. Wilcox CS, Palm F, Welch WJ. Renal oxygenation and function of the rat kidney: effects of inspired oxygen and preglomerular oxygen shunting. Adv Exp Med Biol 765: 329 –334, 2013. 1606. Wilcox CS, Pearlman A. Chemistry and antihypertensive effects of tempol and other nitroxides. Pharm Rev 60: 418 – 469, 2008. 1607. Wilcox CS, Sterzel RB, Dunckel PT, Mohrmann M, Perfetto M. Renal interstitial pressure and sodium excretion during hilar lymphatic ligation. Am J Physiol Renal Fluid Electrolyte Physiol 247: F344 –F351, 1984. 1608. Wilcox CS, Welch WJ. Interaction between nitric oxide and oxygen radicals in regulation of tubuloglomerular feedback. Acta Physiol Scand 168: 119 –124, 2000. 1609. Wilcox CS, Welch WJ, Murad F, Gross SS, Taylor G, Levi R, Schmidt HH. Nitric oxide synthase in macula densa regulates glomerular capillary pressure. Proc Natl Acad Sci USA 89: 11993–11997, 1992. 1587. Welch WJ, Tojo A, Lee JU, Kang DG, Schnackenberg CG, Wilcox CS. Nitric oxide synthase in the JGA of the SHR: expression and role in tubuloglomerular feedback. Am J Physiol Renal Physiol 277: F130 –F138, 1999. 1610. Williams JM, Burke M, Lazar J, Jacob HJ, Roman RJ. Temporal characterization of the development of renal injury in FHH rats and FHH.1BN congenic strains. Am J Physiol Renal Physiol 300: F330 –F338, 2011. 1588. Welch WJ, Tojo A, Wilcox CS. Roles of NO and oxygen radicals in tubuloglomerular feedback in SHR. Am J Physiol Renal Physiol 278: F769 –F776, 2000. 1611. Williams JM, Sarkis A, Hoagland KM, Fredrich K, Ryan RP, Moreno C, Lopez B, Lazar J, Fenoy FJ, Sharma M, Garrett MR, Jacob HJ, Roman RJ. Transfer of the CYP4A region of chromosome 5 from Lewis to Dahl S rats attenuates renal injury. Am J Physiol Renal Physiol 295: F1764 –F1777, 2008. 1589. Welch WJ, Wilcox CS. Modulating role for thromboxane in the tubuloglomerular feedback response in the rat. J Clin Invest 81: 1843–1849, 1988. 1590. Welch WJ, Wilcox CS. Feedback responses during sequential inhibition of angiotensin and thromboxane. Am J Physiol Renal Fluid Electrolyte Physiol 258: F457–F466, 1990. 1591. Welch WJ, Wilcox CS. Potentiation of tubuloglomerular feedback in the rat by thromboxane mimetic. Role of macula densa. J Clin Invest 89: 1857–1865, 1992. 1592. Welch WJ, Wilcox CS. AT1 receptor antagonist combats oxidative stress and restores nitric oxide signaling in the SHR. Kidney Int 59: 1257–1263, 2001. 1593. Welch WJ, Wilcox CS, Folger WH. Mechanism of renal vasoconstriction with thromboxane mimetic: engagement of tubuloglomerular feedback. Adv Prostaglandin Thromboxane Leukotriene Res 21B: 693– 696, 1991. 1594. Welsh DG, Morielli AD, Nelson MT, Brayden JE. Transient receptor potential channels regulate myogenic tone of resistance arteries. Circ Res 90: 248 –250, 2002. 1595. Wesselman JP, Schubert R, VanBavel ED, Nilsson H, Mulvany MJ. KCa-channel blockade prevents sustained pressure-induced depolarization in rat mesenteric small arteries. Am J Physiol Heart Circ Physiol 272: H2241–H2249, 1997. 1596. Westcott EB, Goodwin EL, Segal SS, Jackson WF. Function and expression of ryanodine receptors and inositol 1,4,5-trisphosphate receptors in smooth muscle cells of murine feed arteries and arterioles. J Physiol 590: 1849 –1869, 2012. 1597. Westcott EB, Jackson WF. Heterogeneous function of ryanodine receptors, but not IP3 receptors, in hamster cremaster muscle feed arteries and arterioles. Am J Physiol Heart Circ Physiol 300: H1616 –H1630, 2011. 1612. Williams JM, Sarkis A, Lopez B, Ryan RP, Flasch AK, Roman RJ. Elevations in renal interstitial hydrostatic pressure and 20-hydroxyeicosatetraenoic acid contribute to pressure natriuresis. Hypertension 49: 687– 694, 2007. 1613. Williams RH, Thomas C, Bell PD, Navar LG. Autoregulation of nephron filtration rate in the dog assessed by indicator-dilution technique. Am J Physiol Renal Fluid Electrolyte Physiol 233: F282–F289, 1977. 1614. Williamson GA, Loutzenhiser RD, Wang X, Griffin KA, Bidani AK. Systolic and mean blood pressures and afferent arteriolar myogenic response dynamics: a modeling approach. Am J Physiol Regul Integr Comp Physiol 295: R1502–R1511, 2008. 1615. Wilson HM, Stewart KN. Glomerular epithelial and mesangial cell culture and characterization. Methods Mol Biol 806: 187–201, 2012. 1616. Wilson SM, Mason HS, Smith GD, Nicholson N, Johnston L, Janiak R, Hume JR. Comparative capacitative calcium entry mechanisms in canine pulmonary and renal arterial smooth muscle cells. J Physiol 543: 917–931, 2002. 1617. Winternitz SR, Katholi RE, Oparil S. Role of the renal sympathetic nerves in the development and maintenance of hypertension in the spontaneously hypertensive rat. J Clin Invest 66: 971–978, 1980. 1618. Wittmann U, Nafz B, Ehmke H, Kirchheim HR, Persson PB. Frequency domain of renal autoregulation in the conscious dog. Am J Physiol Renal Fluid Electrolyte Physiol 269: F317–F322, 1995. 1619. Wolf G, Ritz E. Diabetic nephropathy in type 2 diabetes prevention and patient management. J Am Soc Nephrol 14: 1396 –1405, 2003. Physiol Rev • VOL 95 • APRIL 2015 • www.prv.org 509 RENAL AUTOREGULATORY MECHANISMS 1620. Wolfle SE, Navarro-Gonzalez MF, Grayson TH, Stricker C, Hill CE. Involvement of nonselective cation channels in the depolarisation initiating vasomotion. Clin Exp Pharmacol Physiol 37: 536 –543, 2010. 1641. Yu H, Harrap SB, Di NR. Cosegregation of spontaneously hypertensive rat renin gene with elevated blood pressure in an F2 generation. J Hypertens 16: 1141–1147, 1998. 1621. Wolin MS, Gupte SA, Oeckler RA. Superoxide in the vascular system. J Vasc Res 39: 191–207, 2002. 1642. Yu JZ, Zhang DX, Zou AP, Campbell WB, Li PL. Nitric oxide inhibits Ca2⫹ mobilization through cADP-ribose signaling in coronary arterial smooth muscle cells. Am J Physiol Heart Circ Physiol 279: H873–H881, 2000. 1622. Woods LL, DeYoung DR, Smith BE. Regulation of renal hemodynamics after protein feeding: effects of loop diuretics. Am J Physiol Renal Fluid Electrolyte Physiol 261: F815–F823, 1991. 1643. Yuan J, Pannabecker TL. Architecture of inner medullary descending and ascending vasa recta: pathways for countercurrent exchange. Am J Physiol Renal Physiol 299: F265–F272, 2010. 1623. Woods LL, Mizelle HL, Hall JE. Autoregulation of renal blood flow and glomerular filtration rate in the pregnant rabbit. Am J Physiol Regul Integr Comp Physiol 252: R69 –R72, 1987. 1644. Yuen PST, Garbers DL. Guanylyl cyclase-linked receptors. Annu Rev Neurosci 15: 193–225, 1992. 1624. Woods LL, Mizelle HL, Hall JE. Control of renal hemodynamics in hyperglycemia: possible role of tubuloglomerular feedback. Am J Physiol Renal Fluid Electrolyte Physiol 252: F65–F73, 1987. 1645. Yuill KH, McNeish AJ, Kansui Y, Garland CJ, Dora KA. Nitric oxide suppresses cerebral vasomotion by sGC-independent effects on ryanodine receptors and voltage-gated calcium channels. J Vasc Res 47: 93–107, 2010. 1625. Wray S, Burdyga T. Sarcoplasmic reticulum function in smooth muscle. Physiol Rev 90: 113–178, 2010. 1646. Zagorac D, Yamaura K, Zhang C, Roman RJ, Harder DR. The effect of superoxide anion on autoregulation of cerebral blood flow. Stroke 36: 2589 –2594, 2005. 1626. Wronski T, Seeliger E, Persson PB, Forner C, Fichtner C, Scheller J, Flemming B. The step response: a method to characterize mechanisms of renal blood flow autoregulation. Am J Physiol Renal Physiol 285: F758 –F764, 2003. 1647. Zambraski EJ, Ciccone CD. Effects of aortic constriction and renal denervation in DOCA-hypertensive swine. Am J Physiol Renal Fluid Electrolyte Physiol 253: F1223– F1231, 1987. 1627. Wu F, Park F, Cowley AW Jr, Mattson DL. Quantification of nitric oxide synthase activity in microdissected segments of the rat kidney. Am J Physiol Renal Physiol 276: F874 –F881, 1999. 1648. Zatz R, Meyer TW, Rennke HG, Brenner BM. Predominance of hemodynamic rather than metabolic factors in the pathogenesis of diabetic glomerulopathy. Proc Natl Acad Sci USA 82: 5963–5967, 1985. 1628. Wu X, Davis MJ. Characterization of stretch-activated cation current in coronary smooth muscle cells. Am J Physiol Heart Circ Physiol 280: H1751–H1761, 2001. 1649. Zhan CD, Sindhu RK, Vaziri ND. Up-regulation of kidney NAD(P)H oxidase and calcineurin in SHR: reversal by lifelong antioxidant supplementation. Kidney Int 65: 219 –227, 2004. 1629. Xi Q, Adebiyi A, Zhao G, Chapman KE, Waters CM, Hassid A, Jaggar JH. IP3 constricts cerebral arteries via IP3 receptor-mediated TRPC3 channel activation and independently of sarcoplasmic reticulum Ca2⫹ release. Circ Res 102: 1118 –1126, 2008. 1630. Xiong Z, Sperelakis N. Regulation of L-type calcium channels of vascular smooth muscle cells. J Mol Cell Cardiol 27: 75–91, 1995. 1631. Yang T, Hassan SA, Singh I, Smart A, Brosius FC, Holzman LB, Schnermann J, Briggs JP. SA gene expression in the proximal tubule of normotensive and hypertensive rats. Hypertension 27: 541–551, 1996. 1632. Yang Y, Sohma Y, Nourian Z, Ella SR, Li M, Stupica A, Korthuis RJ, Davis MJ, Braun AP, Hill MA. Mechanisms underlying regional differences in the Ca2⫹ sensitivity of BKCa current in arteriolar smooth muscle. J Physiol 591: 1277–1293, 2013. 1633. Yeh HI, Rothery S, Dupont E, Coppen SR, Severs NJ. Individual gap junction plaques contain multiple connexins in arterial endothelium. Circ Res 83: 1248 –1263, 1998. 1634. Yeon DS, Kim JS, Ahn DS, Kwon SC, Kang BS, Morgan KG, Lee YH. Role of protein kinase C- or RhoA-induced Ca2⫹ sensitization in stretch-induced myogenic tone. Cardiovasc Res 53: 431– 438, 2002. 1635. Yip KP, Holstein-Rathlou NH, Marsh DJ. Chaos in blood flow control in genetic and renovascular hypertensive rats. Am J Physiol Renal Fluid Electrolyte Physiol 261: F400 – F408, 1991. 1636. Yip KP, Holstein-Rathlou NH, Marsh DJ. Mechanisms of temporal variation in singlenephron blood flow in rats. Am J Physiol Renal Fluid Electrolyte Physiol 264: F427– F434, 1993. 1637. Yip KP, Marsh DJ. [Ca2⫹]i in rat afferent arteriole during constriction measured with confocal fluorescence microscopy. Am J Physiol Renal Fluid Electrolyte Physiol 271: F1004 –F1011, 1996. 1650. Zhang A, Carella A, Altura BT, Altura BM. Interactions of magnesium and chloride ions on tone and contractility of vascular muscle. Eur J Pharmacol 203: 223–235, 1991. 1651. Zhang AY, Yi F, Teggatz EG, Zou AP, Li PL. Enhanced production and action of cyclic ADP-ribose during oxidative stress in small bovine coronary arterial smooth muscle. Microvasc Res 67: 159 –167, 2004. 1652. Zhang DX, Borbouse L, Gebremedhin D, Mendoza SA, Zinkevich NS, Li R, Gutterman DD. H2O2-induced dilation in human coronary arterioles: role of protein kinase G dimerization and large-conductance Ca2⫹-activated K⫹ channel activation. Circ Res 110: 471– 480, 2012. 1653. Zhang F, Kaide J, Wei Y, Jiang H, Yu C, Balazy M, Abraham NG, Wang W, Nasjletti A. Carbon monoxide produced by isolated arterioles attenuates pressure-induced vasoconstriction. Am J Physiol Heart Circ Physiol 281: H350 –H358, 2001. 1654. Zhang J, Lee MY, Cavalli M, Chen L, Berra-Romani R, Balke CW, Bianchi G, Ferrari P, Hamlyn JM, Iwamoto T, Lingrel JB, Matteson DR, Wier WG, Blaustein MP. Sodium pump alpha2 subunits control myogenic tone and blood pressure in mice. J Physiol 569: 243–256, 2005. 1655. Zhang J, Ren C, Chen L, Navedo MF, Antos LK, Kinsey SP, Iwamoto T, Philipson KD, Kotlikoff MI, Santana LF, Wier WG, Matteson DR, Blaustein MP. Knockout of Na⫹/ Ca2⫹ exchanger in smooth muscle attenuates vasoconstriction and L-type Ca2⫹ channel current and lowers blood pressure. Am J Physiol Heart Circ Physiol 298: H1472–H1483, 2010. 1656. Zhang Q, Cao C, Zhang Z, Wier WG, Edwards A, Pallone TL. Membrane current oscillations in descending vasa recta pericytes. Am J Physiol Renal Physiol 294: F656 – F666, 2008. 1638. Yip KP, Marsh DJ. An Arg-Gly-Asp peptide stimulates constriction in rat afferent arteriole. Am J Physiol Renal Physiol 273: F768 –F776, 1997. 1657. Zhang R, Harding P, Garvin JL, Juncos R, Peterson E, Juncos LA, Liu R. Isoforms and functions of NAD(P)H oxidase at the macula densa. Hypertension 53: 556 –563, 2009. 1639. Yoshida T, Inoue R, Morii T, Takahashi N, Yamamoto S, Hara Y, Tominaga M, Shimizu S, Sato Y, Mori Y. Nitric oxide activates TRP channels by cysteine S-nitrosylation. Nat Chem Biol 2: 596 – 607, 2006. 1658. Zhang SL, Yu Y, Roos J, Kozak JA, Deerinck TJ, Ellisman MH, Stauderman KA, Cahalan MD. STIM1 is a Ca2⫹ sensor that activates CRAC channels and migrates from the Ca2⫹ store to the plasma membrane. Nature 437: 902–905, 2005. 1640. Young DK, Marsh DJ. Pulse wave propagation in rat renal tubules: implications for GFR autoregulation. Am J Physiol Renal Fluid Electrolyte Physiol 240: F446 –F458, 1981. 1659. Zhang Z, Rhinehart K, Kwon W, Weinman E, Pallone TL. ANG II signaling in vasa recta pericytes by PKC and reactive oxygen species. Am J Physiol Heart Circ Physiol 287: H773–H781, 2004. 510 Physiol Rev • VOL 95 • APRIL 2015 • www.prv.org CARLSTRÖM ET AL. 1660. Zhao D, Zhang J, Blaustein MP, Navar LG. Attenuated renal vascular responses to acute angiotensin II infusion in smooth muscle-specific Na⫹/Ca2⫹ exchanger knockout mice. Am J Physiol Renal Physiol 301: F574 –F579, 2011. 1661. Zhao X, Cook AK, Field M, Edwards B, Zhang S, Zhang Z, Pollock JS, Imig JD, Inscho EW. Impaired Ca2⫹ signaling attenuates P2X receptor-mediated vasoconstriction of afferent arterioles in angiotensin II hypertension. Hypertension 46: 562–568, 2005. 1662. Zhao X, Yamamoto T, Newman JW, Kim IH, Watanabe T, Hammock BD, Stewart J, Pollock JS, Pollock DM, Imig JD. Soluble epoxide hydrolase inhibition protects the kidney from hypertension-induced damage. J Am Soc Nephrol 15: 1244 –1253, 2004. 1663. Zhong XZ, Harhun MI, Olesen SP, Ohya S, Moffatt JD, Cole WC, Greenwood IA. Participation of KCNQ (Kv7) potassium channels in myogenic control of cerebral arterial diameter. J Physiol 588: 3277–3293, 2010. 1664. Zimmermann PA, Knot HJ, Stevenson AS, Nelson MT. Increased myogenic tone and diminished responsiveness to ATP-sensitive K⫹ channel openers in cerebral arteries from diabetic rats. Circ Res 81: 996 –1004, 1997. 1665. Zou AP, Billington H, Su N, Cowley AW Jr. Expression and actions of heme oxygenase in the renal medulla of rats. Hypertension 35: 342–347, 2000. 1666. Zou AP, Drummond HA, Roman RJ. Role of 20-HETE in elevating loop chloride reabsorption in Dahl SS/Jr rats. Hypertension 27: 631– 635, 1996. 1667. Zou AP, Fleming JT, Falck JR, Jacobs ER, Gebremedhin D, Harder DR, Roman RJ. Stereospecific effects of epoxyeicosatrienoic acids on renal vascular tone and K⫹-channel activity. Am J Physiol Renal Fluid Electrolyte Physiol 270: F822–F832, 1996. 1668. Zou AP, Imig JD, Kaldunski ML, Ortiz de Montellano PR, Sui Z, Roman RJ. Inhibition of renal vascular 20-HETE production impairs autoregulation of renal blood flow. Am J Physiol Renal Fluid Electrolyte Physiol 266: F275–F282, 1994. 1669. Zou AP, Imig JD, Ortiz de Montellano PR, Sui Z, Falck JR, Roman RJ. Effect of P-450 omega-hydroxylase metabolites of arachidonic acid on tubuloglomerular feedback. Am J Phyisol 266: F934 –F941, 1994. 1670. Zou AP, Li N, Cowley AW Jr. Production and actions of superoxide in the renal medulla. Hypertension 37: 547–553, 2001. 1671. Zou H, Ratz PH, Hill MA. Role of myosin phosphorylation and [Ca2⫹]i in myogenic reactivity and arteriolar tone. Am J Physiol Heart Circ Physiol 269: H1590 –H1596, 1995. 1672. Zou R, Cupples WA, Yip KP, Holstein-Rathlou NH, Chon KH. Time-varying properties of renal autoregulatory mechanisms. IEEE Trans Biomed Eng 49: 1112–1120, 2002. 1673. Zou Y, Akazawa H, Qin Y, Sano M, Takano H, Minamino T, Makita N, Iwanaga K, Zhu W, Kudoh S, Toko H, Tamura K, Kihara M, Nagai T, Fukamizu A, Umemura S, Iiri T, Fujita T, Komuro I. Mechanical stress activates angiotensin II type 1 receptor without the involvement of angiotensin II. Nat Cell Biol 6: 499 –506, 2004. Physiol Rev • VOL 95 • APRIL 2015 • www.prv.org 511