Comparative genetics of Montana and arctic grayling, Thymallus arcticus
advertisement

Comparative genetics of Montana and arctic grayling, Thymallus arcticus
by Jeremiah Cornelius Lynch
A thesis submitted in partial fulfillment of the requirements for the degree of MASTER OF SCIENCE
in Zoology
Montana State University
© Copyright by Jeremiah Cornelius Lynch (1977)
Abstract:
An investigation was made of the biochemical genetic variation within and among four populations of
the arctic grayling, Thymallus articus. Two populations surveyed were representative of the form found
in the main range of the species, northern Canada and Alaska, and two populations were representative
of the disjunct Montana form of Thymallus articus. Estimates of these parameters were obtained from a
starch gel electrophoretic survey of thirty-five enzyme loci and protein loci. The percent polymorphic
loci (12.5 percent) and average heterozygosity (2.7-3.I percent) are intermediate in the range estimated
for salmonid species and may reflect the limited habitat diversity of grayling compared with other
salmonid species.
No relationship between genetic variability and enzyme function was identified for this species. Both a
rapidly evolving set and a slowly evolving set of proteins appeared to be present.
Comparisons among the four populations were based on allelic protein variation at eight loci. Results
of genetic similarity and genetic distance calculations indicate that genetic divergence has taken place
between the arctic form and Montana form of T. arcticus, which may warrent subspecific status for the
two forms. STATEMENT OF PERMISSION TO COPY
In p r e s e n t i n g t h i s t h e s i s in p a r t i a l f u l f i l l m e n t o f th e
requirem en ts f o r an advanced degree a t Montana S t a t e U n i v e r s i t y ,
I agree t h a t t h e Li b ra r y s h a l l make i t f r e e l y a v a i l a b l e f o r i n s p e c ­
tion.
I f u r t h e r ag re e t h a t pe rm is si on f o r e x t e n s i v e copying o f
t h i s t h e s i s f o r s c h o l a r l y purposes may be g r a n t e d by ny major
p r o f e s s o r , o r , in h i s a bse n c e , by t h e D i r e c t o r o f L i b r a r i e s .
It
i s und erstood t h a t any copying o r p u b l i c a t i o n o f t h i s t h e s i s f o r
f i n a n c i a l gain s h a l l n o t be allowed w i t h o u t my w r i t t e n p e r m i s s i o n .
COMPARATIVE GENETICS OF MONTANA AND ARCTIC GRAYLING,
THYMALLUS ARCTICUS
by
Jeremiah C. Lynch
A t h e s i s s ubm itt e d in p a r t i a l f u l f i l l m e n t
o f the req u ir e m e n ts f o r t h e degree
of
MASTER OF SCIENCE
in
Zoology
Approved:
i
^
=
__________
C h a i rp e r so n , Gj^duate Committee
'ad, Major Department
Graduate Bean
MONTANA,STATE UNIVERSITY
Bozeman, Montana
September, 1977
iii
ACKNOWLEDGMENTS
I would l i k e t o e x p re ss my s i n c e r e g r a t i t u d e t o my major p r o f e s ­
s o r , Dr. E r n e s t R. Vyse, f o r h i s g u id a nce , a s s i s t a n c e and c ontin ue d
s up po rt t h r o u g h o u t t h i s s tu d y .
S pe ci al thanks a re extended t o John D. Varley (Yellowstone Park
F i s h e r i e s Management) f o r h i s i n t e r e s t in t h e s tu d y and a s s i s t a n c e
in o b t a i n i n g samples; Dr. David G. Cameron f o r h i s a s s i s t a n c e through
ou t t h i s s tu d y and c o n s t r u c t i v e review o f t h i s m a n u s c r i p t ; Dr. Fred
W. A l l e n d o r f ( U n i v e r s i t y o f Montana) f o r t h e use o f h i s computer
program f o r a n a l y s i s o f t h e d a t a ; and Dr. Calvin Kaya f o r h i s review
o f t h i s m a nusc ri pt .
F i n a l l y , I would l i k e t o thank my w i f e , T e r i , f o r t h e s p e c i a l
su ppo rt she has given me t h r o u g h o u t t h i s s tu d y .
TABLE OF CONTENTS
Page
VITA
. ...........................................................................................
ACKNOWLEDGMENTS ............................................................................................
LIST OF T A B L E S ...............................................................................................
LIST OF FIGURES...........................................
ABSTRACT...........................................
INTRODUCTION
.............................................................
MATERIALS AND METHODS..........................
Sampling o f P o p ul a tio ns : ..............................................................
Sample P r e p a r a t i o n ..........................
E l e c t r o p h o r e s i s . ......................................................................................
Q u a l i t a t i v e A na ly sis .......................................................................
Nomenclature ........................................................................................
RESULTS..........................
E l e c t r o p h o r e t i c Phenotypes o f Monomorphic P r o t e i n s . .
L a c t a t e dehydrogenase ...............................
Malate dehydrogenase .......................................
Glutamat e - o x a l o a c e t a t e tr a n s a m i n a s e ..............................
Alcohol dehydrogenase ................................................ . . .
Xanthine dehydrogenase .........................................................
S o r b i t o l dehydrogenase .........................................................
I s o c i t r a t e dehydrogenase .....................................................
A lp ha -gl yc erophosph at e dehydrogenase . . . . . . .
E s t e r a s e ...................................
H e x o k i n a s e ....................................................................................
iii
vi
vii
ix
I
I
LO I ''- OO
D i s t r i b u t i o n ........................................................................................
Taxonomy ............................................
Electrophoresis . . . . . . . .
V a r i a t i o n in Natural P o p u la ti o n s
O b j e c t i v e s ............................................................................................
i1
13
19
19
22
22
24
29
31
31
31
35
38
42
42
43
44
45
46
47
V
TABLE OF CONTENTS
(Continued)
Page
E l e c t r o p h o r e t i c Phenotypes o f Polymorphic P r o t e i n s . .
T e tr a z o li u m oxi da se ..................................................................
Phosphoglucomutase ..................................................................
I s o c i t r a t e dehydrogenase ..............................
T r a n s f e r r i n ....................................................................................
Glucose and hexose 6-ph os phat e dehydrogenase . . .
Malic e n z y m e ..........................
Serum p r o t e i n s ..........................................................................
47
47
50
52
56
59
68
71
Q u a n t i t a t i v e A na ly sis o f Gen etic V a r i a b i l i t y
..................
80
DISCUSSION......................................................... .... . ; ...........................
89
Genetic V a r i a b i l i t y o f Thymallus cw ctieus
.......................
Genetic Divergence Between P o p u la ti o n s o f Thymallus
a v o t i o u s ................................................ ' ..............................................
Taxonomic C o n s i d e r a t i o n s . .............................................................
89
97
107
APPENDIX.............................................................................
HO
Bu ffe r Systems .................................................................. .... . . . I l l
S t a i n i n g Procedures ..............................................................................
113
LITERATURE CITED
117
/
vi
LIST OF TABLES
Table
1.
Page
C o l l e c t i o n d a ta f o r t h e f o u r p o p u l a t i o n s o f
.........................................................
21
P r o t e i n s su rveyed, t i s s u e s examined, and b u f f e r
systems employed in e l e c t r o p h o r e t i c a n a l y s i s o f
Thymallus a rctleu s
................................... . . . . . . .
27
Thymallus avotious
2.
.
3.
A l l e l e f r e q u e n c i e s and degree o f h e t e r o z y g o s i t y
in 35 l o c i examined in f o u r p o p u l a t i o n s o f
Thymallus oFctious
4.
Correspondence o f observed genotype f r e q u e n c i e s
t o th o s e expected on t h e b a s i s o f Hardy-Weinberg
e q u i l i b r i u m f o r th e polymorphic l o c i o f Thymallus
a v o tio u s
5.
7.
8.
..............................................................................................................................................................................................................................................................................
86
Es tim a te s o f g e n e t i c v a r i a b i l i t y in Thymallus
avotious
6.
81
......................................................................................
......................................................................................................................................................
88
Amount o f polymorphism and t h e degree o f h e t e r o ­
z y g o s i t y in some f i s h s p e c i e s ........................................ .... .
91
In d ic e s o f s i m i l a r i t y and g e n e t i c d i s t a n c e f o r
. f o u r p o p u l a t i o n s o f Thymallus avotious ...........................
99
Genetic s i m i l a r i t i e s between p o p u l a t i o n s a t
d i f f e r e n t s t a g e s o f e v o l u t i o n a r y d iv e rg e nc e in
s e v e r a l groups o f f i s h ..................................................... .... . 102
vi i
LIST OF FIGURES
Figure
1.
'
Page
Map o f Alaska and w e st e rn Canada showing g r a y l i n g
d i s t r i b u t i o n ............................................................................... .
2
2.
D i s t r i b u t i o n o f indigenous g r a y l i n g in Montana
4
3.
L a c t a t e dehydrogenase (LDH) .....................................................
4.
T i ss u e d i s t r i b u t i o n o f m a la te dehydrogenase (MDH)
from t h e same f i s h .............................................................................37
5.
Glu t a m a t e - o x a l o a c e t a t e t ra n s a m i n a s e (GOT) t i s s u e
d i s t r i b u t i o n .............................................................
40
6.
T e tr a z o li u m oxi da se (TO) polymorphism ...............................
49
7.
Phosphoglucomutase (PGM)
.........................................................
51
8.
Phosphoglucomutase (PGM) polymorphism ...............................
53
9.
I s o c i t r a t e dehydrogenase (IDH)
............................................
55
10.
T r a n s f e r r i n (Tfn) polymorphism
. ..............................................58
11.
Gluc ose -6 -p hos pha te dehydrogenase (G6PD) and hexose6-pho sph ate dehydrogenase (H6PD) e x p r e s s i o n in
e r y t h r o c y t e s and eye t i s s u e ...........................................................63
12.
Gluc os e -6 -p hos pha te dehydrogenase-3 (G6PD-3)
13.
Hexose-6-phosphate dehydrogenase (H6PD) p o l y ­
morphism .................................................................................................. 65
14.
Malic enzyme ( M E ) ...........................................................................
70
15.
Electropherograms o f serum p r o t e i n s
74
16.
Diagrammatic r e p r e s e n t a t i o n o f e l e c t r o p h o r e t i c
p a t t e r n o f serum p r o t e i n s o f Thymallus a v c tio u s . . .
. . .
. . . .
..................................
34
64
76
viii
LIST OF FIGURES
(Continued)
Figure
17.
18.
Page
The twelve observed phen oty pic p a t t e r n s o f
e l ectropherograms o f g r a y l i n g serum p r o t e i n s
in Zone 5 ............................................................................................
Dendrogram f o r f o u r p o p u l a t i o n s o f T. aroti-cus
77
. . . 109
ix
ABSTRACT
An i n v e s t i g a t i o n was made o f t h e biochemical g e n e t i c v a r i a t i o n
w i t h i n and among f o u r p o p u l a t i o n s o f t h e a r c t i c g r a y l i n g , Tkym allus
a vo tio u s. Two p o p u l a t i o n s surveyed were r e p r e s e n t a t i v e o f t h e form
found in t h e main range o f t h e s p e c i e s , n o r t h e r n Canada and A la sk a ,
and two p o p u l a t i o n s were r e p r e s e n t a t i v e o f t h e d i s j u n c t Montana form
o f Tkymallus a v o tia u s. E s ti m a te s o f t h e s e pa ra m et e rs were o b t a i n e d
from a s t a r c h gel e l e c t r o p h o r e t i c surve y o f t h i r t y - f i v e enzyme l o c i
and p r o t e i n l o c i . The p e r c e n t polymorphic l o c i (12. 5 p e r c e n t ) and
average h e t e r o z y g o s i t y ( 2 . 7 - 3 . I p e r c e n t ) a r e i n t e r m e d i a t e in th e
range e s t i m a t e d f o r salmonid s p e c i e s and may r e f l e c t t h e l i m i t e d
h a b i t a t d i v e r s i t y o f g r a y l i n g compared with o t h e r salmonid s p e c i e s .
No r e l a t i o n s h i p between g e n e t i c v a r i a b i l i t y and enzyme f u n c t i o n
was i d e n t i f i e d f o r t h i s s p e c i e s . Both a r a p i d l y e v o l v i n g s e t and a
slowly e v o l v i n g s e t o f p r o t e i n s appeared t o be p r e s e n t .
Comparisons among th e f o u r p o p u l a t i o n s were based on a l l e l i c
p r o t e i n v a r i a t i o n a t e i g h t l o c i . R e s u lt s o f g e n e t i c s i m i l a r i t y
and g e n e t i c d i s t a n c e c a l c u l a t i o n s i n d i c a t e t h a t g e n e t i c div e rg e n c e
has taken p l a c e between t h e a r c t i c form and Montana form o f T.
a p c tio u s, which may w a r r e n t s u b s p e c i f i c s t a t u s f o r t h e two forms.
i
INTRODUCTION
D istribution
Thymallus a v o tie u s , th e a r c t i c g r a y l i n g , i s a f r e s h w a t e r f i s h
which i n h a b i t s cold o r a r c t i c r e g i o n s .
The n a t i v e range o f t h e s p e c i e s
is hoi a r c t i c , o c c u r r i n g in n o r t h e r n d r a i n a g e s o f North America and
E u r a s ia .
In E u r a s i a , i t i s found from t h e Kara and Ob R i v e r s , in
t h e west ern U.S.S.R. t o t h e e a s t e r n S i b e r i a n Coast ( i n c l u d i n g a l l
streams d r a i n i n g i n t o th e Bering Sea, and t h e Penzhina R ive r d r a i n i n g
i n t o t h e sea o f O kho tos k), s outh t o n o r t h e r n Mongolia and t h e Yalu
River (Walters 1955, S c o t t and Crossman 1973).
In Canada and Ala ska , T. a ro tio u s oc cur s from V a n s i t t a r t I s l a n d
o f f t h e M e l v i l l e P e n i n s u l a ; sou th al ong t h e west c o a s t o f Hudson Bay
t o t h e Owl R i v e r , Manitoba; west th ro u g h o u t t h e ^Northwest and Yukon
T e r r i t o r i e s t o th e Bering Sea d r a i n a g e s in A la ska ; so ut h in S a s k a t c h e ­
wan t o Rein dee r Lake bu t a b s e n t in most o f t h e C h u r c h il l R i v e r ; south
t o Central A l b e r t a ; in n o r t h e r n B r i t i s h Columbia from t h e Pease and
S t i k i n e R ive r no rt h (Walters 1955, S la s te n en k o 1950, S c o t t and
Crossman 1973).
Figure I shows th e d i s t r i b u t i o n o f Thymallus a v c tio u s
in Canada and Alaska.
In t h e co nti guous United S t a t e s Thymallus a ro tio u s was indigen ous
in Michigan and Montana.
I s o l a t e d p o p u l a t i o n s were p r e s e n t in
Michigan in t h e upper p a r t o f t h e Lower P e n i n s u l a , and in t h e O t t e r
3
River o f t h e Upper Pe n in s u la (Hubbs and L a g le r 1949).
The Michigan
form, however, has been e x t i n c t s i n c e 1936 ( S c o t t and Crossman 1973,
U.S. Dept, o f I n t e r i o r 1966).
Another p o p u l a t i o n was found in Montana
in th e headwaters o f t h e Missouri River above t h e G reat F a l l s
(Henshall 1906).
This southward e x t e n s i o n and th e su bse que nt e s t a b ­
lis hm e nt o f t h e s e two p o p u l a t i o n s was e v i d e n t l y t h e r e s u l t o f g l a c i a l
action.
In Montana, th e o r i g i n a l range as d e s c r i b e d by Henshall (1906)
has been g r e a t l y reduced.
The d e c l i n e o f t h e s p e c i e s has been r e p o r ­
te d and r e a f f i r m e d by v a r io u s i n v e s t i g a t o r s (Ke lly 1931, Brown 1943,
Nelson 1954, 1956).
The p r e s e n t d i s t r i b u t i o n in n a t i v e r i v e r s and
streams i s d e s c r i b e d by Brown (1971) with t h e s t a t e m e n t t h a t "a few
a re found in t h e Sun, Big Hole, Red Rock, and Madison R i v e r s . ". Gray­
l i n g a r e e n t i r e l y a b s e n t from t h e Missouri R i v e r , t h e G a l l a t i n River
and t h e main stem o f th e J e f f e r s o n Riv er.
Two small remnant popu la ­
t i o n s remain in t h e J e f f e r s o n t r i b u t a r i e s : one in t h e Big Hole River
\
and t h e o t h e r in t h e Red Rock Lakes a r e a (Nelson 1954).
The range o f
indigenous p o p u l a t i o n s o f Thymallus a ^o tio u s in Montana i s shown in
Figure 2.
There has been widespread t r a n s p l a n t i n g o f Canadian s t o c k s i n t o
Montana p o p u l a t i o n s (McPhail and Lindsey 1971) and h a t c h e r y p l a n t i n g
from a s i n g l e so urc e ( t h e Red Rock Lakes) i n t o a l l bu t one o f the
4
G re a t
F a lls
C olum bia Riirer
D ra ina ge
H ele n a
p IF n rk s
Y e l lo w s to n e
N a tio n a l
Park
Idaho
Figure 2,
D is tr ib u tio n of In d ig en o u s grayling
in M o n t a n a .
5
n a t u r a l p o p u l a t i o n s (Ke lly 1931).
At t h i s time re p ro d u c in g g r a y l i n g
p o p u l a t i o n s a re known t o e x i s t in 39 l a k e s and 14 s tre am s in w e ste rn
Montana on both s i d e s o f t h e C o n ti n e n ta l Divide ( Hblten 1971).
The
only indigenous Montana p o p u l a t i o n known not t o be co ntam inated by
p l a n t i n g s i s t h e Red Rock Lakes p o p u la ti o n (Nelson 1954).
In Yellowstone National Park t h e g r a y l i n g o c c u r r e d n a t u r a l l y in
th e Madison River system and t h e G a l l a t i n R iv er.
I t i s no lo n g e r
p r e s e n t in t h e G a l l a t i n River and i s r a r e in t h e Madison R iv e r (Dean
and Varley 1974).
T r a n s p l a n t i n g o f g r a y l i n g t o l a k e s in Yellowstone
National Park has been d e s c r i b e d by Kruse (1959).
Successful s e l f - '
pr op a g a ti n g p o p u l a t i o n s were e s t a b l i s h e d in Grebe Lake, Wolf Lake,
and Ice Lake, above t h e V i r g i n i a Cascades on t h e Gibbon R i v e r , and
Cascade Lake in t h e Yellowstone d r a i n a g e .
Taxonomy
The g r a y l i n g s a r e s o f t rayed t e l e o s t f i s h be lo ngin g t o t h e o r d e r
Is o sp o n d y li , s u b o rd e r S a lm on oid e i.
T h e i r f u r t h e r taxonomic c l a s s i f i c a ­
t i o n has been one f r a u g h t with c o n fu s io n .
The genus Thymallus was
s e p a r a t e d from t h e genus Salmo (C u r v ie r 1829), b u t t h e i r fa m ily
c l a s s i f i c a t i o n was debated f o r some time by ta xonom is ts and remains
unresolved toda y ( S c o t t and Crossman 1973).
Some a u t h o r s have a s s ig n e d
a l l t h r e e ge ographic groups ( A r c t i c , A si an , and Montana-Mic h i gan) t o
t h e Salmonidae (Boulenger 1895, Regean 1914) w hil e o t h e r s (Jordan and
6
Everman 1896, Berg 1940, 1955) have p la ce d t h e g r a y l i n g s in a s e p a r a t e
f a m i l y , t h e Thymmalli d a e .
The most r e c e n t c l a s s i f i c a t i o n , based on
o s t e o l o g i c a l c h a r a c t e r i s t i c s (Norden 1961), p l a c e s t h e g r a y l i n g in
th e s u b - f a m i l y ThymaU l n a e , o f t h e f a m ily Salmonldae.
Four s p e c i e s
a r e re c o g n iz e d : T. b r e v ir o s tv is (Mongolia), T. thymallus (E u r o p e ) ,
T. nigvesoens (Lake Kosogol, Mongolia) and T. arotious ( e a s t e r n
S i b e r i a and North America).
The s t a t u s o f th e v a r i o u s forms o f
f o r some tim e.
araticu s has been de bated
For decades i t was c o n s i d e r e d t h a t t h e North American
g r a y l i n g s c o n s i s t e d o f t h r e e s p e c i e s : t . sig n ife v (Richardson 1823)
found in n o r t h e r n Canada and A la sk a , T. tr ic o lo r (Cope 1865) found
in Michigan, and T. montanus (Milner 1873) found in Montana.
This
c l a s s i f i c a t i o n was p r i n c i p a l l y based on geog raphic i s o l a t i o n , and
s e v e r a l morphological c h a r a c t e r i s t i c s ( s i z e and shape o f d o r s a l f i n ,
m a x i l l a r y l e n g t h , and c o l o r v a r i a t i o n ) .
More r e c e n t l y , T. s ig n ife r
has been c o n s i d e r e d c o n s p e c i f i c with t . arotious ( P a l l a s ) , and th e
o t h e r American forms r e l e g a t e d s u b s p e c i f i c s t a t u s (Walters 1955).
Walters showed t h a t t h e Canadian and Alaskan form was i d e n t i c a l with
two A s i a t i c forms (y. a. p a lla s i and T. a. gruberi n atio m ertensi)
and sug ges te d t h a t they be d e s i g n a t e d as Thymallus arotious s ig n ife r
(Richardson 1823).
Walters (1955) f u r t h e r re c o g n iz e d t h e Montana-
Michigan form as a n o t h e r s u b s p e c i e s tric o lo r..
The v a l i d i t y o f t h e
7
North American s u bs pe c ie s has n o t been a d e q u a t e l y demo nst rat ed ( S c o t t
and Crossman 1973, Norden 1961).
At t h e p r e s e n t time no s u b s p e c ie s
should be rec og ni ze d w i t h i n T. a rc tic u s u n t i l f u r t h e r ev idence
w a rr a n ts such d i s t i n c t i o n (McPhail and Lindsey 1971).
Electrophoresis
P r o t e i n s a r e ampholytes an d, t h e r e f o r e , may c a r r y a n e t n e g a t i v e
or p o s i t i v e ch ar ge.
The n e t charge depends on t h e i o n i z a t i o n o f I )
f r e e carboxyl groups (COOK™) o f glu ta mi c a c i d and a s p a r t i c a c i d
r e s i d u e s and 2) f r e e amino groups (NH^+ ) o f l y s i n e and a r g i n i n e (and
to a le s s e r ex ten t h i s tid in e ) .
The n e t charge o f t h e p r o t e i n depends,
on which group predomin ate s.
The degree o f i o n i z a t i o n depends on th e
pH o f th e p r o t e i n s o l u t i o n .
In a b u f f e r o f high pH t h e a c i d i c groups
a r e p r o g r e s s i v e l y n e u t r a l i z e d by t h e a l k a l i component o f t h e b u f f e r ,
thu s a ll ow in g t h e b a s i c groups t o predominate.
This r e s u l t s in th e
p r o t e i n molecule having a n e t n e g a t i v e ch ar ge.
At a low pH th e
r e v e r s e o c c u r s , and th e p r o t e i n w i l l have a n e t p o s i t i v e c h ar g e.
At
a c e r t a i n pH, t h e i s o e l e c t r i c p o i n t , th e p o s i t i v e and n e g a t i v e charg es
a r e balanced and t h e r e i s no n e t ch arge.
E l e c t r o p h o r e s i s m a n ip ula te s t h e ampholytic b e h a v io r o f p r o t e i n s
by appl yin g an e l e c t r i c a l f i e l d t o a s o l u t i o n o f p r o t e i n s , s e p a r a t i n g
them on t h e b a s i s o f t h e i r n e t c h ar g e.
I f t h e pH i s l e s s than t h e
i s o e l e c t r i c p o i n t o f t h e p r o t e i n , i t w i l l m ig r a te toward t h e cathode
8
and i f t h e pH i s g r e a t e r th a n t h e i s o e l e c t r i c p o i n t o f t h e p r o t e i n ,
i t w i l l m ig r a te toward t h e anode.
The r a t e o f m i g r a t i o n depends on
t h e number o f c h a r g e s , t h e m o le c u la r s i z e and on t h e v o l t a g e a p p l i e d .
T h e r e f o r e , by th e s e l e c t i o n o f th e a p p r o p r i a t e b u f f e r and e l e c t r i c a l
c u r r e n t , p r o t e i n d i f f e r e n c e s can be de ter mi ne d.
E l e c t r o p h o r e t i c te c h n i q u e s vary as t o t h e s u p p o r t i n g media used.
Sta rc h gel was employed in t h e p r e s e n t s t u d y .
Isozyme s e p a r a t i o n
using s t a r c h gel depends not only on n e t i o n i c c h a r g e , but t o a
l e s s e r e x t e n t on d i f f e r e n c e s o f molp cu la r s i z e .
S ta r c h gel a c t s as
a m ole c ula r s i e v e , m e ch a ni c al ly s e p a r a t i n g molecules o f d i f f e r e n t
s i z e s by a f f e c t i n g t h e i r r a t e o f m ig r a ti o n ( S m ith ie s 1955).
V a r i a t i o n in Natural Po p u la ti o n s
The development o f s t a r c h gel e l e c t r o p h o r e s i s (S mi th ie s 1955),
and t h e i n t r o d u c t i o n o f simple s t a i n i n g t e c h n i q u e s f o r t h e d e t e c t i o n
o f s p e c i f i c enzyme a c t i v i t y (Hunter and Markert 1957), has allowed
th e v i s u a l i z a t i o n o f i n d i v i d u a l p r o t e i n s , hence , s i n g l e gene pro d u c ts
The combination o f t h e s e te c h n i q u e s provide d a means by which h e t e r o ­
g e n e i t y o f p r o t e i n s and enzymes could e a s i l y be d e t e c t e d .
This
allowed t h e c h a r a c t e r i z a t i o n a t t h e m o le c u la r l e v e l o f t h e amount
o f g e n e t i c v a r i a b i l i t y in p o p u l a t i o n s and an e s t i m a t e o f t h e e x t e n t
o f g e n e t i c d iv e rg e nce among c l o s e l y r e l a t e d s p e c i e s ( G o t t l i e b 1971).
9
Kimura and Crow (1964) h y poth e si ze d t h a t t h e number o f a l l e l e s
t h a t can be mainta ine d a t a s i n g l e locus in a f i n i t e p o p u l a t i o n i s
large.
Shaw (1965) p o in te d o u t t h a t isozymes which vary w i t h i n popu­
l a t i o n s i s t h e r u l e r a t h e r than t h e e x c e p t i o n .
Approximately TOO
l o c i a re known t o have e l e c t r o p h o r e t i c v a r i a n t s in p o p u l a t i o n s o f
many or ga nis ms , i n c l u d i n g man. Drosophila, a n t s , f i s h , mice , f r o g s
and many p l a n t s p e c i e s ( G o t t l i e b 1971).
The amount o f such genic
v a r i a t i o n , measured by t h e p r o p o r t i o n o f polymorphic l o c i (common
a l l e l e fr eq ue nc y l e s s than o r equal t o 0 .9 9 ) can be determine d
d i r e c t l y from e l e c t r o p h o r e t i c a n a l y s i s .
In t h e o r y , a l a r g e number o f s t r u c t u r a l l y d i f f e r e n t a l l e l e s may
be g e n er at e d by independent m u ta ti o n s w i t h i n t h e c o n f i n e s o f a s i n g l e
gene (H a r r i s 1976).
A c e r t a i n p r o p o r t i o n o f t h e s e m u ta ti o n s can be
expected t o r e s u l t in a s u b s t i t u t i o n a f f e c t i n g t h e n e t charg e o f th e
protein.
Such a mutation would be r e f l e c t e d in t h e m o b i l i t y o f t h e
protein.
The p r o b a b i l i t y t h a t such a mut at ion w i l l occu r i s c a l c u l a ­
t e d t o be 25-30% (Shaw 1965, Nei 1975).
This means t h a t a l a r g e
number o f amino a c i d s u b s t i t u t i o n s go u n d e t e c t e d s i n c e th e y do not
r e s u l t in a charge change.
The e s t i m a t e s o f g e n e t i c v a r i a b i l i t y based
on e l e c t r o p h o r e t i c a l l y d e t e c t a b l e d i f f e r e n c e s may, t h e r e f o r e , be
c o n s e r v a t i v e ( H a r r i s and Hopkinson 1976, Nei 1975).
10
Data from a random sample o f l o c i coding f o r p r o t e i n s can be
e x t r a p o l a t e d t o e s t i m a t e t h e amount o f g e n e t i c v a r i a b i l i t y in th e
e n t i r e genome ( Lewontin and Hubby 1966).
The e s t i m a t e s o f t h e amount
o f polymorphism in th e sample can be used t o c a l c u l a t e t h e p r o p o r t i o n
o f polymorphic l o c i and i n d i v i d u a l h e t e r o z y g o s i t y in a s p e c i e s .
These e s t i m a t e s can pro vid e a b a s i s f o r comparison between s p e c i e s
( U t t e r e t a l. 1973) and may a l s o be used t o compare p o p u l a t i o n s
within species.
The l e v e l o f g e n e t i c polymorphism has been e s t i m a t e d f o r a
v a r i e t y o f s p e c i e s : man - 25 p e r c e n t o f 12 l o c i were polymorphic
( H a rr is 1966) and more r e c e n t l y man - 31 p e r c e n t o f 71 l o c i ( H a r r i s
and Hopkinson 1972); Mus rmsoulus - 30 p e r c e n t polymorphic (S e la n d e r
e t a l. 1969) and 40 p e r c e n t polymorphic (S e l a n d e r and Yang 1969);
Pevomysous polionotus
-
23 p e r c e n t polymorphic (S e la n d e r e t a l. 1971);
q u a i l , Cotuvnix ootuvnix - 54 t o 58 p e r c e n t polymorphic (Baker and
Manwell 1967); p h e a s a n t , Phasianus ooldhious - 43 p e r c e n t polymorphic
(Baker e t a l. 1966); Dvosophila ( v a r i e t y o f s p e c i e s ) - 30 t o 67 p e r ­
c e n t polymorphic ( Lewontin and Hubby 1966, Prakash e t a l, 1969, Ayala
e t a l. 1970, Berger 1970, O'Brien and MacIntyre 1969).
In f i s h
s p e c i f i c a l l y , e s t i m a t e s a r e : Astynax - 29 t o 41 p e r c e n t f o r i n l a n d
p o p u l a t i o n s and 0 t o 20 p e r c e n t f o r cave d w e l l e r s (Avise and S e l a n d e r
1972), h e r r i n g - 45 p e r c e n t polymorphic (Altukhov e t a l. 1972), brook
11
t r o u t , Salvelinus fo n tin a lis - 38 p e r c e n t polymorphic (Wright and
Atherton 1970); chum salmon - 11 t o 18 p e r c e n t polymorphic (Altukhov
e t a l. 1972); r o c k f i s h , Sebastes ( v a r i e t y o f s p e c i e s ) - 4 t o 8 p e r c e n t
polymorphic (Johnson e t a l. 1973), P a c i f i c salmon
-
8 t o 13 p e r c e n t
( U t t e r e t a l. 1 9 7 3 ) rainbow t r o u t - 26 p e r c e n t ( U t t e r e t a l. 1973).
In sur ve yin g t h e l i t e r a t u r e as a whole, a ppro xim a te ly 30 p e r c e n t o f
t h e s t r u c t u r a l gene l o c i a r e polymorphic ( G o t t l i e b 1971; Nei 1975).
H e te r o z y g o s it y v a r i e s c o n s i d e r a b l y with locus (King and Wilson 1975,
Nei and Roychoudhury 1974, S e l a n d e r and Johnson 1973, U t t e r e t a l.
1973).
The e x i s t e n c e o f lo cu s dependent r a t e s o f change i s well
i l l u s t r a t e d by amino a c i d sequencing d a t a in d i v e r s e organisms
(Dickerson 1972), which i n d i c a t e s t h a t p r o t e i n s evo lve a t d i f f e r e n t
rates.
Gene l o c i a r e polymorphic when a l l e l e s u b s t i t u t i o n i s in t r a n s i ­
t i o n , when b a l a n c i n g s e l e c t i o n s t a b i l i z e s f r e q u e n c i e s , o r when a
mutant a l l e l e becomes f r e q u e n t by chance.
Since each loc us may under-
I
go a l l e l e s u b s t i t u t i o n i n d e p e n d e n t l y , a high degree o f i n t e r l o c u s
v a r i a t i o n in h e t e r o z y g o s i t y may r e s u l t .
I n t e r l o c u s v a r i a t i o n may
a l s o be produced i f t h e mu ta tio n r a t e o r t h e type and i n t e n s i t y o f
n a t u r a l s e l e c t i o n v a r i e s among l o c i (Nei 1975).
That l o c i v a r i a t i o n
and, hence, p r o t e i n h e t e r o g e n e i t y i s p r e s e n t in a wide v a r i e t y o f
v e r t e b r a t e s p e c i e s was shown by S e l a n d e r and Johnson (1973).
12
I n t e r l o c u s v a r i a t i o n has a l s o been found in many s p e c i e s o f f i s h
( U t t e r e t a t. 1973).
This i n t e r l o c u s v a r i a t i o n can be s e p a r a t e d i n t o two gro u p s , a
r a p i d l y e v o l v i n g s e t and a slow ly e vo lv in g s e t .
The r a p i d l y e v o lv in g
s e t , i n c l u d i n g plasma p r o t e i n s and e s t e r a s e s , acc umulates e l e c t r o p h o r e t i c a l l y d e t e c t ! ble s u b s t i t u t i o n s a t a r a t e t e n f o l d g r e a t e r than
th e slow er s e t , which i n c l u d e s enzymes in volv e d in m e t a b o l i c pathways
(S ar ic h 1977).
C o r r e l a t i o n s have been proposed between enzyme
f u n c t i o n and h e t e r o z y g o s i t y ( G i l l i s p i e and Kojima 1968, Kojima e t a t.
1970, Johnson 1971, 1974, Powell 1975), and more r e c e n t l y a r e l a t i o n ­
sh ip between h e t e r o z y g o s i t y and q u a t e r n a r y s t r u c t u r e has been proposed
(Ward 1977).
The b a s i s f o r t h e d i f f e r e n c e in h e t e r o z y g o s i t y between
l o c i i s n o t a d e q u a t e l y r e s o l v e d , but e s t i m a t e s o f average h e t e r o ­
z y g o s i t y o r g e n e t i c d i s t a n c e would be o v e r e s t i m a t e d o r u n d e r e s t i m a t e d
i f th e p r o t e i n s chosen did n o t in c l u d e an a deq ua te mi xtu re o f both
sets.
In t h e p r e s e n t s tu dy a l a r g e number o f l o c i have been examined
with no p r e f e r e n c e given t o e i t h e r t h e r a p i d l y e v o lv in g o r t h e slo wer
evo lv in g s e t o f p r o t e i n s .
Nei and Roychoudhury ( 1974) have s u g g es te d t h a t t o e s t i m a t e t h e
average h e t e r o z y g o s i t y p e r l o c u s , a l a r g e number o f l o c i r a t h e r than
a l a r g e number o f i n d i v i d u a l s sho uld be used.
Avise and Ayala (1975)
s t a t e t h a t with r e s p e c t t o g e n e t i c s i m i l a r i t y , t h e v a r i a n c e about
13
i n d i v i d u a l s is. small r e l a t i v e t o t h e v a r i a n c e about l o c i ; t h e r e f o r e ,
th e p r e c i s i o n o f t h e s e e s t i m a t e s i s much more dependent on t h e number
o f l o c i than on t h e numbers o f i n d i v i d u a l s sampled.
I f the in te n t is
to p r e d i c t Hardy-Weinberg e q u i l i b r i u m as well as e s t i m a t e av erage
h e t e r o z y g o s i t y , a r e l a t i v e l y l a r g e number o f i n d i v i d u a l s s hould be
examined f o r each polymorphic lo cu s (Nei 1975).
I f both o f t h e s e
e s t i m a t e d a r e w i t h i n t h e scope o f a s t u d y , a l a r g e number o f i n d i v i d u ­
a l s should be scr een ed a t a l a r g e number o f l o c i with t h e hope o f
minimizing any b i a s .
A g r e a t deal o f d a t a has been c o l l e c t e d as t o th e g e n e t i c v a r i a ­
b i l i t y o f f i s h p r o t e i n s (deLigny 1969, Kirpi ch nik ov 1973).
Protein
h e t e r o g e n e i t y appea rs t o be p r e s e n t in n e a r l y e ver y s p e c i e s s t u d i e d .
A r e l i a b l e s e t o f p r o t e i n s which have been s t u d i e d in o t h e r f i s h
s p e c i e s , and r e p o r t e d in t h e l i t e r a t u r e , were chosen f o r a n a l y s i s so
t h a t r e l i a b l e e s t i m a t e s o f av erage h e t e r o z y g o s i t y and Hardy-Weinberg
f r e q u e n c i e s could be c a l c u l a t e d .
In a d d i t i o n , comparisons can be
made t o t h e p u b li s h e d r e s u l t s f o r o t h e r s p e c i e s .
The a v a i l a b i l i t y o f
s u b s t r a t e f o r s t a i n i n g and t h e c l a r i t y o f r e s o l u t i o n were t h e f i n a l
f a c t o r s in d e te r m in in g what p r o t e i n s were i n c lu d e d .
f
O b je ct iv es
The North American forms o f Thymallus avctieu s have been i s o l a t e d
s i n c e b e f o r e t h e l a s t Wisconsin g l a c i a t i o n (Vincent 1962).
Isolated
14
p o p u l a t i o n s were e s t a b l i s h e d in f a v o r a b l e h a b i t a t s s outh o f t h e main
range o f Thymallus ccrotious.
For p o p u l a t i o n s t o become g e n e t i c a l l y
d i f f e r e n t i a t e d , e v o l u t i o n i s t s b e l i e v e th e y must be com ple tely
i s o l a t e d from one a n o t h e r .
This i s o l a t i o n may oc cu r g e o g r a p h i c a l l y
o r r e p r o d u c t i v e Iy (Dobzhansky 1951 , 1970).
Complete i s o l a t i o n has
oc cu rr ed in t h e case o f Thymallus a ro tio u s, between t h e a l l o p a t r i c
p o p u l a t i o n s , which a t one time s har ed t h e same gene p o o l , b u t have
s i n c e become i s o l a t e d from one a n o t h e r .
T h e r e f o r e , an o p p o r t u n i t y
f o r e v o l u t i o n a r y div e rg e nce o f t h e v a r io u s s t o c k s has been p r e s e n t .
An e s t i m a t i o n o f th e amount o f dive rg e nc e which has ta ken p l a c e could
be o b ta in e d by a survey o f e l e c t r o p h o r e t i c d i f f e r e n c e s ( d i s c u s s e d
previously).
However, t h e comparison o f t h e t h r e e a m er ican forms o f
T. avetious meets with some d i f f i c u l t y .
The Michigan form has been
e x t i n c t s i n c e 1936 ( S c o t t and Crossman 1973, U.S. Dept, o f t h e
I n t e r i o r 1966) and thu s i t s d iv e rg e nc e from th e o t h e r two cann ot be
determined.
The Montana form as d e s c r i b e d by HenshalI (1906) d e a l t
almost e x c l u s i v e l y in r i v e r s and s t r e a m s , hence i t was an a d f l u v i a l
form (s tream d w e l l i n g - s t r e a m spawning) and only s e c o n d a r i l y a l a c u s ­
t r i n e form (l a k e d w e l l i n g - s t r e a m spawning).
From a g e n e t i c and
taxonomic p o i n t o f view, t h e s t u d y o f th e a d f l u v i a l form may be
impos si ble due t o t h e reduced numbers (Brown 1971), t h e c o n ta m in at io n
o f independent p o p u l a t i o n s by t h e i n t r o d u c t i o n o f Canadian s t o c k s
15
(MePha iI and Lindsey 1971), and th e u n i v e r s a l p l a n t i n g o f l a c u s t r i n e
forms from a s i n g l e donor (Red Rock Lakes) i n t o d i s c r e t e a d f l u v i a l
populations.
The Red Rock Lakes p o p u l a t i o n i s a pure deme o f th e
Montana form o f Thymallus Ca1O tio u s i with no t r a n s p l a n t s o f any o t h e r
s to ck s having ta ken pla ce (Nelson 1954).
The Red Rock Lakes po pula ­
t i o n , a lt h o u g h known t o be n a t i v e , i s a r e s t r i c t e d headwaters popula ­
t i o n and may have been g e n e t i c a l l y d i s t i n c t in i t s own r i g h t .
The
use o f i t as a donor s to c k in widespread p l a n t i n g may have d i l u t e d
th e e n z o o t i c p o p u l a t i o n s in r i v e r s and stream s t h r o u g h o u t t h e n a t i v e
range.
The v a l i d i t y o f a g e n e t i c b a s i s f o r t h e a d f l u v i a l and l a c u s ­
t r i n e b e h a v io r a l d i f f e r e n c e s has not been s t u d i e d , however, t h e i n n a t e
g e n e t i c c o n t r o l o f m ig r a ti o n in o t h e r salmonid s p e c i e s has been
su ggested (Ral ei gh 1967, Brannon 1967, N ort hc ote 1969, Raleigh and
Chapman 1971).
I f t h e r e a r e d i s t i n c t g e n e t i c a l l y c o n t r o l l e d behav­
i o r a l d i f f e r e n c e s between t h e two forms, i t may be s u g g e s te d t h a t t h e
\
l a c u s t r i n e form, h i s t o r i c a l l y r a r e in t h e Montana range o f th e s p e c i e s ,
i s now widespread w hile th e h i s t o r i c a l l y common a d f l u v i a l ty pe i s
t h r e a t e n e d with e x t i n c t i o n .
In th e p r e s e n t s t u d y , t h e Grebe and Wolf Lakes p o p u l a t i o n s ,
which were e s t a b l i s h e d by t r a n s p l a n t i n g from a s t o c k d e r i v e d from the
Madison R ive r system (Ke lly 1931, Kruse 1959), were used t o r e p r e s e n t
t h e Montana form.
Grayling were no t n a t i v e t o e i t h e r o f t h e s e l a k e s .
16
but both were stocke d with g r a y l i n g d e r i v e d only from t h e Madison R i v e r
system.
Eggs taken from g r a y l i n g n a t i v e t o Meadow Creek in t h e Madison
River d r a i n a g e were r e a r e d in t h e S t a t e Fish and Game's Anaconda
ha tc he ry and t h e progeny were p l a n t e d in Georgetown Lake (Ke lly 1931).
Subsequent t o t h i s p l a n t i n g , eggs taken from t h e s e Georgetown g r a y l i n g
were t r a n s p l a n t e d t o t h e s e l a k e s in Yellowstone Park (Kruse 1958).
The d a t a o b ta in e d from t h e s e demes. i s , t h e r e f o r e , presumed t o be
o f an a d f l u v i a l form which have s i n c e become adap te d t o a l a c u s t r i n e
existence.
The d a t a o b t a i n e d can be used t o e s t i m a t e t h e amount o f
divergence which has ta ken p la c e between t h e A r c t i c form and th e
Montana forms o f Thymallus arotiou s.
This e s t i m a t e o f t h e amount o f
g e n e t i c div e rg e n c e can be based on th e g e n e t i c c h a r a c t e r s o f th e
p o p u l a t i o n s o b t a i n e d through e l e c t r o p h o r e t i c d a t a .
The use o f th e
Madison R iver p o p u la ti o n i t s e l f i s a p r a c t i c a l i m p o s s i b i l i t y due to
th e dim inished numbers o f i n d i v i d u a l s in t h i s d e c l i n i n g p o p u l a t i o n
(Dean and Varley 1974).
In a d d i t i o n t o th e comparison o f th e A r c t i c form and t h e Montana
form, th e s t u d y can a l s o de ter mi ne i f g e n e t i c d i f f e r e n t i a t i o n has
occu rr ed w i t h i n two Montana p o p u l a t i o n s , t h e Grebe Lake and Wolf
Lake p o p u l a t i o n s .
Salmonids have a tendency t o evolv e g e n e t i c a l l y
d i s c r e t e , e c o l o g i c a l l y s p e c i a l i z e d p o p u l a t i o n s with d i f f e r e n t i a t i o n
based on l i f e h i s t o r y c h a r a c t e r s such as time and p la c e o f spawning
17
( Behnke 1972).
As p r e v i o u s l y mentioned, i n n a t e g e n e t i c c o n t r o l o f
m ig ra tio n h a b i t s in salmonids has been s u g g e s te d .
The s t r o n g homing
beh avi or o f most salmonids i s an im p o r ta n t f a c t o r in t h e g e n e r a t i o n
and maintenance o f t h i s g e n e t i c d i v e r s i t y ( A l l e n d o r f e t a l . 1971).
Such appea rs t o be t h e case wit h t h e s e p o p u l a t i o n s , t h e Wolf Lake
po p u la ti o n sampled was an o u t l e t spawning p o p u l a t i o n w h il e t h e Grebe
Lake p o p u l a t i o n sampled was an i n l e t spawning p o p u l a t i o n .
Thus, t h e
Grebe and Wolf Lakes p o p u l a t i o n s have p a r t i a l Iy e s t a b l i s h e d e t h o l o g i cal r e p r o d u c t i v e i s o l a t i o n , which may be t h e f i r s t s t e p in g e n e t i c
isolation.
The amount o f g e n e t i c dive rg e nc e between t h e s e two popu­
l a t i o n s thu s give s an e s t i m a t e o f t h e amount o f g e n e t i c change, a t
t h e s t r u c t u r a l gene l e v e l , t h a t has accompanied t h i s e v e n t .
I f th e
magnitude o f t h e div e rg e nce between t h e s e p o p u l a t i o n s and t h e Canadian
p o p u l a t i o n s i s l a r g e , t h e e v o l u t i o n a r y p o s i t i o n o f t h e s e forms could
be c l a r i f i e d .
The phe notypic f r e q u e n c i e s r e v e a l e d by p a t t e r n s o f p r o t e i n s on
e l e c t r o p h o r e t i c g e l s , can be i n t e r p r e t e d in terms o f g e n o ty p ic f r e ­
quencies and t h e p o p u la ti o n a l l e l i c f r e q u e n c i e s , which a r e th e
parameters o f e v o l u t i o n a r y g e n e t i c s .
With t h e s e two pa ra m et e rs o f
th e dernes known, th e i n l e t and o u t l e t spawning p o p u l a t i o n s can be
compared with one a n o t h e r and with t h e A r c t i c form with r e g a r d t o
t h e unique p r o p o r t i o n o f t h e genome t h a t d i s t i n g u i s h e s t h e s e
18
p o p u l a t i o n s , and t h e l e v e l o f h e t e r o z y g o s i t y under t h e d i f f e r e n t
environments.
Both o f t h e s e a r e im p o r ta n t p r o p e r t i e s o f d i v e r g i n g
g e n e t i c systems.
E l e c t r o p h o r e t i c te c h n i q u e s provid e c o n s i d e r a b l e i n f o r m a t i o n to
help e l u c i d a t e e v o l u t i o n a r y r e l a t i o n s h i p s among, c l o s e l y r e l a t e d
s p e c i e s (Avise 1974).
Biochemical g e n e t i c v a r i a t i o n among c l o s e l y
r e l a t e d p o p u l a t i o n s can be used t o examine s y s t e m a t i c r e l a t i o n s h i p s
(Nei 1975, S a r i c h 1977).
The use o f such d a t a t o de ter mi ne taxonomic
s t a t u s has been s u c c e s s f u l l y a p p l i e d t o salmonid s p e c i e s (Payne e t d l.
1971, Nyman 1972, U t t e r e t a t. 1973, R e i n i t z 1974).
The d a t a o b t a i n e d
in th e p r e s e n t s tu d y w i l l c o n t r i b u t e t o th e d a t a needed t o help
c l a r i f y t h e confused taxonomic p o s i t i o n t h a t p r e s e n t l y e x i s t s through
t h e s u b s p e c ie s c l a s s i f i c a t i o n o f Thymallus aratiou s.
The r e s u l t s
w i l l h o p e f u l l y provid e some ev ide nc e as t o whether s u b s p e c i a t i o n
has oc cu rr ed between t h e A r c t i c and Montana forms o f Thymallus
OXtC t 1I o U S .
MATERIALS AND METHODS
Sampling o f Po pul a tio ns
D i s c r e t e p o p u la ti o n s o f e i t h e r t h e Canadian form o r t h e Montana
form o f Thymallus avctious were chosen f o r a n a l y s i s .
Populations,
which a c c o rd in g t o Montana Fi sh and Game s t o c k i n g r e c o r d s , were known
no t t o be a mi xtu re o f both forms were sampled as r e p r e s e n t a t i v e o f
th e Montana form.
The Canadian a r c t i c p o p u l a t i o n was assumed t o be
n a t i v e , but d e s i g n a t i o n o f any p o p u la ti o n as a Montana form had t o
be confirmed from t r a n s p l a n t r e c o r d s .
The p o p u l a t i o n s used as r e p r e s e n t a t i v e s o f t h e Canadian form were
taken from t h e Donnelly R i v e r , N.W.T. and Fuse Lake, Montana.
The
Donnelly R iv e r l i e s in th e Mackenzie River d r a i n a g e and g r a y l i n g a re
n a t i v e to i t s w a te rs (McPhail and Lindsey 1970).
Fuse Lake in
G ra ni te County, Montana, i s an a l p i n e la ke which had no n a t i v e f i s h
fauna.
Fuse Lake was s to cke d in 1930 with g r a y l i n g from t h e Saskatche
wan River d r a i n a g e ( S t a t e Fish and Game Records) with no s ubseq ue nt
p l a n t i n g s , th u s i t i s a pure p o p u la ti o n o f th e Canadian form.
The
Saskatchewan R iv e r p o p u la ti o n in t u r n was d e r i v e d from a Canadian
A r c t i c g r a y l i n g p o p u l a t i o n (Lindsey 1956).
The Montana form was sampled from Grebe and Wolf Lakes p o p u l a t i o n s
in Yellowstone National Park.
Grebe and Wolf Lakes a r e connected by
f i v e hundred meters o f th e Gibbon River which flows from Grebe Lake
20
i n t o Wolf Lake.
Both la k e s have i n l e t and o u t l e t spawning ad apted
populations of grayling.
In sampling t h e s e p o p u l a t i o n s , only th e
1
o u t l e t spawning Wolf Lake p o p u la ti o n and t h e i n l e t spawning Grebe
Lake p o p u l a t i o n were sampled in o r d e r t h a t any g e n e t i c d i f f e r e n c e s
a s s o c i a t e d with t h i s spawning be h a v io r could be e s t i m a t e d in th e
e l e c t r o p h o r e t i c survey.
The g r a y l i n g were c o l l e c t e d e i t h e r by a n g l i n g o r by use o f an
e l e c t r i c backpack sh ock er.
The number o f i n d i v i d u a l s c o l l e c t e d , t h e
d a t e , the method, and t h e l o c a t i o n s i t e a r e l i s t e d in Table I .
Upon c a p t u r e t h e weight and l e n g th o f t h e f i s h were re c o rd e d .
Blood samples were taken by making a l o n g i t u d i n a l i n c i s i o n from th e
isthmus t o t h e abdominal re g io n o f the f i s h , opening t h e p e r i c a r d i a l
sac with an i n c i s i o n and removing a ppro xim a te ly I ml o f blood.
The
blood was p la ce d in a p l a s t i c tube and e i t h e r c e n t r i f u g e d a t 5000 g
f o r ap pr ox im a te ly 3 minutes when done in t h e f i e l d , o r t h e tu be of
blood was pla c e d on i c e and t r a n s p o r t e d t o t h e l a b o r a t o r y where
c e n t r i f u g i n g was done.
A f t e r c e n t r i f u g a t i o n t h e serum was s e p a r a t e d
from th e blood c e l l s , s t o r e d in a microfuge tube and both were
immediately f r o z e n .
Ti ssu e samples o f th e l i v e r , muscle, h e a r t and eye were taken
and placed immediately on dry i c e and were t r a n s f e r r e d t o a f r e e z e r
maintained a t -50°C upon r e t u r n t o t h e l a b o r a t o r y .
A f t e r removal
Table I.
C o l l e c t i o n da ta f o r the fo u r popu la ti o n s of Thymallus arotiaus.
Population
Grebe Lakea
Y.N.P.
Wolf Lakeb
Y.N.P.
Donnelly River
Fuse Lake
Mont.
a I n l e t Creek
^O u tl e t Creek
N.W.T.
Date
Number o f
In d i v i d u a l s
A ncestral Stock
6/25/75
30
Madison River System
6/11/76
30
6/17/76
18
6/7 /7 7
22
6/25/75
8
7/22/75
12
6/10/76
10
5/24/77
22
5/31/77
8
9/1/ 76
44
Native
8/15/75
19
Mackenzie River
Madison River System
22
o f t i s s u e samples t h e sex o f t h e i n d i v i d u a l was noted when i t could
be de te r m in e d , oth e rw is e i t was c l a s s i f i e d as immature.
Sample P r e p a r a t i o n
T i ss u e e x t r a c t s were pre p a re d by g r i n d i n g th e t i s s u e in an e q u i v a ­
l e n t volume o f b u f f e r (.01 M I r i s ;
.001 M EDTA; 5 x 10 ^ M NADP; pH
a d j u s t e d t o 6 . 8 ) , in a g l a s s homogenizer.
The homogenate was then
c e n t r i f u g e d a t 15,000 g f o r 20 minutes in a r e f r i g e r a t e d c e n t r i f u g e
ma intained a t -10°C.
The s u p e r n a t a n t was then removed f o r e l e c t r o ­
p h o r e s i s o r s t o r e d a t -50°C f o r l a t e r use.
Electrophoresis
H o ri z ont a l s t a r c h gel e l e c t r o p h o r e s i s was used in t h e e l e c t r o ­
phoretic analysis of pro te in s.
The s t a r c h g e l s were 11.2% hydrolyzed
s t a r c h ( E l e c t r o s t a r c h C. Madison, W isconsin).
The s t a r c h was mixed
thor ou gh ly with th e a p p r o p r i a t e amount o f b u f f e r in a s i d e arm f l a s k
and pla ced on a hot p l a t e w ith a magnetic s t i r r e r , with a d d i t i o n a l
hand s h a k in g , u n t i l th e s o l u t i o n b o i l e d .
The r a t e o f s t i r r i n g was
in c r e a s e d as t h e v i s c o s i t y o f t h e f l u i d i n c r e a s e d t o keep t h e f l u i d
homogeneous.
A vacuum was then a p p l i e d t o t h e f l a s k f o r a ppro xim a te ly
60 seconds t o remove a i r bub b le s.
on g l a s s p l a t e s 25 cm by 18 cm.
The hot s t a r c h s o l u t i o n was poured
P l e x i g l a s s s t r i p s 1.5 cm wide and
I cm t h i c k , he ld in pla ce by l a r g e paper clamps, were used t o form
23
th e edges o f t h e g e l .
A f t e r an i n i t i a l c o o l i n g (I t o 2 hours a t room
te m p e r a t u r e ) t h e ge ls were covered with p l a s t i c wrap w i t h o u t t r a p p i n g
any bubbles and s t o r e d in a r e f r i g e r a t o r .
Gels were used f o r e l e c t r o ­
p h o r e s i s any time up t o 24 hours l a t e r .
P r i o r t o a p p l i c a t i o n o f samples t o t h e g e l , 26 sample s l o t s
.5x1
cm were made a c r o s s t h e gel by a s l o t former.
This allowed
each sample s l o t t o be c omp le tely s e p a r a t e d by .5 cm o f g e l .
A
f i l t e r pap er wick (ap pro xi m a te ly I cm x .5 cm) was soaked in a sample
s u p e r n a t a n t , b l o t t e d on f i l t e r pa per t o remove e x c e s s , and p la ce d in
th e sample s l o t by t h e use o f small f o r c e p s .
P l a s t i c b u f f e r t r a y s (15 cm x 5 cm x 3 cm) were f i l l e d with 250 ml
buffer.
Disposable c l o t h s (Handy Wipes) 25 cm wide, were used as t h e
e l e c t r o d e sponges.
The sponges were pla ced 3 cm from th e ends o f th e
g e l , and t h e e n t i r e top o f th e a p p a r a t u s was covered with p l a s t i c
wrap.
A d i r e c t e l e c t r i c a l c u r r e n t , th e v o l t a g e o f which v a r i e d as
t o b u f f e r system used was a p p l i e d t o t h e g e l .
The gel was cooled by
a pan o f i c e suspended 2 mm o ve r t h e e n t i r e gel s u r f a c e by p l e x i g l a s s
strips.
E l e c t r o p h o r e s i s was co ntin ued u n t i l a marker ( Bromophenol Blue)
reached the anodal sponge.
The b u f f e r systems employed, v o l t a g e us ed ,
and le ngt h o f run a r e l i s t e d in t h e Appendix.
24
A f t e r e l e c t r o p h o r e s i s was completed, t h e g e l s were s l i c e d i n t o
. 2 0 - .2 5 cm t h i c k s l i c e s by a s t a i n l e s s s t e e l b l a d e .
The top s l i c e
was always d i s c a r d e d due t o d i s t o r t i o n and t h e bottom 3 s l i c e s were
s t a i n e d by t h e a p p r o p r i a t e procedure f o r v i s u a l i z a t i o n o f t h e p r o t e i n s
desired.
The s t a i n i n g proc ed ures f o r d i f f e r e n t p r o t e i n systems a r e
l i s t e d in t h e Appendix.
A f t e r t h e a p p r o p r i a t e time f o r adequate
development o f th e zymogram, t h e g e l s were washed t h r e e times in
d i s t i l l e d w a te r and f i x e d in a m e t h a n o l / w a t e r / a c e t i c a c i d ( 5 / 5 / 1 )
solution.
Permanent re c ord s o f t h e zymograms were made by keeping a w r i t t e n
■
record o f t h e r e s u l t s , and photog raphing each gel on 35 mm Kodak High
C o n tr a s t Copy o r Panotomic X f i l m .
produce b e t t e r r e s u l t s .
The l a t t e r type appeared t o
The f i x e d g e l s were covered w it h Saran Wrap
and r e f r i g e r a t e d f o r l a t e r comparison with t h e photo gra phs.
Q u a l i t a t i v e A na ly sis
Isozymes a r e d e fi n e d as m u l t i p l e m o le c u la r forms o f an enzyme
o c c u r r i n g in a s i n g l e i n d i v i d u a l o r in d i f f e r e n t members o f th e same
s p e c i e s (Markert 1968).
Such isozymes may occu r t o g e t h e r in t h e same
c e l l , but t h e r e may a l s o be marked d i f f e r e n c e s in isozyme p a t t e r n s
between t i s s u e s ( H a rr is 1975).
Several a u t h o r s (ManweTl and Baker
1970, H a r r i s 1976) have d i s c u s s e d t h e f a c t t h a t isozyme systems may
be ge ner ate d in a v a r i e t y o f ways.
The v a r i o u s causes o f isozymes may
25
be c l a s s i f i e d i n t o 3 main c a t e g o r i e s (H a r r i s 1969, 1975; Hopkinson and
H a r r i s 1971):
I) o c cu r re nc e o f m u l t i p l e gene l o c i coding f o r s t r u c ­
t u r a l l y d i s t i n c t p o ly p e p ti d e ch ai ns o f th e enzyme; 2) o c cu r re nc e o f
m u l t i p l e a l l e l i s m a t a s i n g l e locus de te r m in in g s t r u c t u r a l l y d i s t i n c t
v e r s io n s o f a p a r t i c u l a r p o l y p e p t i d e c h a i n ; 3) oc cu r re n c e o f s o - c a l l e d
"secondary isozyme for mation due t o p a s t t r a n s l a t i o n a l m o d i f i c a t i o n s
o f th e enzyme s t r u c t u r e .
Furt her mo re, a p p a r e n t v a r i a t i o n observed
on zymograms o f s t a r c h gel e l e c t r o p h o r e s i s may be a r t i f a c t s due t o
s t o r a g e and p r e s e r v a t i o n o f m a t e r i a l
( Eppenberger e t at. 1971), o r i t
may be th e r e s u l t o f d i f f e r e n t i a l a c t i v a t i o n o f genes due t o e n v i r o n ­
mental c o n d i t i o n s (Hochachka and Somero 1971).
S t r i n g e n t c r i t e r i a must be used in a n a l y z i n g biochemical v a r i a ­
t i o n o f zymograms b e fo re one can assume t h a t t h e r e i s a g e n e t i c b a s i s
f o r th e v a r i a t i o n .
The s t r o n g e s t d a ta f o r d e te r m in in g i f t h e v a r i a ­
t i o n has a g e n e t i c b a s i s comes from b re e din g e xper im ent s and th e
subsequent a n a l y s i s o f progeny from p a r e n t s having known biochemical
differences.
In t h e absence o f such d a t a , as i s t h e case in t h e
p r e s e n t s t u d y , o t h e r c r i t e r i a must be imposed as s ug ges te d by v a r i o u s
a u t h o r s ( U t t e r e t a l . 1973, Avise and Smith 1974, .Gall e t a l . 1976).
The c r i t e r i a used in t h e p r e s e n t s tu dy t o v e r i f y t h e g e n e t i c b a s i s o f
observed v a r i a t i o n were:
26
1)
The banding p a t t e r n observed was t y p i c a l f o r t h e presumed
s t r u c t u r e o f t h e enzyme found in c l o s e l y r e l a t e d s p e c i e s .
2)
The p a t t e r n was i n t e r p r e t a b l e on t h e b a s i s o f a simple
g e n e t i c h y p o t h e s i s ; w it h t y p i c a l homozygotes and h e t e r o z y g o t e s being
e x pre ss ed .
3)
The Observed g e not ypic ,class p r o p o r t i o n s were in c l o s e a g r e e ­
ment with th o s e ex pec te d on t h e b a s i s o f Hardy-Weinberg e q u i l i b r i u m ;
any s i g n i f i c a n t d e v i a t i o n s from th e p r e d i c t e d v a lu es must be e x p l a i n e d .
4)
P a t t e r n s must be r e p e a t a b l e upon su bse que nt sampling o f th e
same i n d i v i d u a l , with p a r a l l e l e x p r e s s i o n in o t h e r t i s s u e s t o co nfirm
any polymorphism.
V a r i a t i o n which d id n o t conform t o a l l o f t h e s e c r i t e r i a could
not be c o n c l u s i v e l y v e r i f i e d t o be g e n e t i c .
However, i f t h e ev ide nc e
p r e s e n te d was s t r o n g l y s u g g e s t i v e o f a g e n e t i c b a s i s , i t was in c lu d e d
in t h e a n a l y s i s .
Since isozymes may have d i f f e r e n t i a l t i s s u e e x p r e s s i o n , s e v e r a l
d i f f e r e n t t i s s u e s were a nal yz ed t o deter mine t h e p a t t e r n o f such
expression.
The t i s s u e s examined f o r a given enzyme system a re
summarized in Table 2.
This allowed th e d e t e r m i n a t i o n o f an added
number o f l o c i whose p r o t e i n may be e x p re s s e d in one t i s s u e and not
a n o t h e r (eg. LDH-5 l o c u s ) .
A v a r i e t y o f b u f f e r systems were employed
in e a r l y s c r e e n i n g runs s i n c e v a r i a t i o n may be e x p re s s e d in one b u f f e r
Table 2.
P r o te i n s surveyed, t i s s u e s examined, and b u f f e r systems employed in e l e c t r o ­
p h o r e t i c a n a l y s i s o f Thymallus a r a tic u s .
Tissue
Liver Muscle Heart Eye Serum
Buffer
System
Pr o te i n
E.C. No.
Abbreviation
Alcohol Dehydrogenase
(1.1.1.1)
(ADH)
Alpha-glycerophosphate
Dehydrogenase
( I . I . I . 8)
(AGPD)
Es te ra se
(3.I .I . I )
(EST)
Glucose-6-Phosphate
Dehydrogenase
(1.1.1.49)
(G-6-PD)
+
+
+
+
A,F
Glutamate O xa loacetate
Transaminase
(2.6.I.I)
(GOT)
+
+
+
+
C1H
Hexose-6-Phosphate
Dehydrogenase
(1.1.1.47)
(H-6-PD)
+
+
+
+
A,F
Hexokinase
(2.7.I.I)
(HK)
+
I s o c i t r a t e Dehydrogenase ( 1 . 1 . 1 . 4 2 )
(IDH)
+
+
L a c ta te Dehydrogenase
(1.1.1.27)
(LDH)
+
+
+
+
+
F1G1H
Malate Dehydrogenase
(1.1.1.37)
(MDH)
+
+
+
+
+
A,B,D
Malic Enzyme
(1.1.1.40)
(ME)
+
+
+
C
C,D,F
+
I
G
B1D
A,B
Table 2.
(Continued)
Pro tei n
Phosphoglucomutase
E.C. No.
Abbre­
viation
( 2 . 7 . 5 . I)
(PGM)
Transferrin
(TFN)
Tetraz olium Oxidase
(TO)
Serum P r o te i n s
(SP)
Tissue
Liver Muscle Heart Eye Serum
+
+
A,E
+
+
+
(1.1.1.14)
(SDH)
+
Xanthine Dehydrogenase
( I . 2.3.2)
(XDH)
+
I
A1F
+
S o r b i t o l Dehydrogenase
Buffer
System
I
H
+
A1F
29
but not a n o t h e r ( U t t e r e t a l . 1973).
The d i f f e r e n t b u f f e r systems
employed f o r r e s o l u t i o n o f a given p r o t e i n a r e l i s t e d in Table 2.
Salmonid f i s h a r e b e l i e v e d t o have undergone e x t e n s i v e gene
d u p l i c a t i o n and, t h e r e f o r e , may be t e t r a p l o i d organisms (Ohno 1969).
Grayling a r e salm onids , hence in t h e g e n e t i c i n t e r p r e t a t i o n o f
e l e c t r o p h o r e t i c p a t t e r n s , i t i s im por ta nt t o r e a l i z e t h a t t h e r e may
be two o r more gene l o c i which determine t h e primary s t r u c t u r e o f
proteins.
The presence o f such d u p l i c a t e d l o c i has been found to
e x i s t in many salmonids ( A l l e n d o r f e t a l . 1975, B a i le y e t a l . 1970,
Morrison and Wright 1966, Wolf e t a l . 1970).
Therefore, grayling
would be assumed t o have s i m i l a r l y d u p l i c a t e d l o c i , c o m p li c a ti n g th e
a n a l y s i s o f zymogram p a t t e r n s .
Genetic i n t e r p r e t a t i o n was f u r t h e r
c l a s s i f i e d by d e te r m in in g i f t h e l o c i were d u p l i c a t e d which e x p l a i n e d
observed bra nding p a t t e r n s o f t h e s e l o c i .
This allowed v a r i a t i o n t o
be a t t r i b u t e d t o e i t h e r m u l t i p l e l o c i o r t o m u l t i p l e a l l e l e s a t a
s i n g l e lo c u s .
All o f t h e above p o s s i b i l i t i e s were c o n s i d e r e d and t h e c r i t e r i a
imposed b e f o r e th e g e n e t i c b a s i s o f v a r i a b i l i t y was assumed.
Nomenclature
The system o f nomenclature fo ll ows t h a t o f Richmond (1972) and
P r ak a s h , Lewontin, and Hubby (1969).
a b b r e v i a t i o n o f th e p r o t e i n name.
Each loc us was named us in g an
When p r o t e i n s o f two o r more l o c i
30
with i d e n t i c a l s u b s t r a t e s p e c i f i c i t i e s were o b s e r v e d , th e l o c i were
numbered a c c o rd in g t o t h e m i g r a t i o n r a t e o f t h e p r o t e i n p ro d u c ts from
t h e o r i g i n , i . e . , th e l o c i whose p r o t e i n had th e s l o w e s t m ig r a ti o n
was as s ig n e d numeral one, th e n e x t f a s t e s t t h e numeral two, and so on.
When a l l e l i c v a r i a t i o n was found a t a l o c u s , t h e most common a l l e l e
was as s ig n e d a number 1.00 and a l l o t h e r a l l e l e s were a s s i g n e d numbers
t h a t r e p r e s e n t e d t h e i r p r o t e i n s m ig r a t i o n d i s t a n c e r e l a t i v e t o t h e
most common a l l e l e ' s p r o t e i n , i . e . , i f a p r o t e i n m ig r a te d a d i s t a n c e
h a l f as f a r , i t s a l l e l e would be l a b e l e d 0.50 o r i f one m ig r at e d a
d i s t a n c e one and a h a l f times as f a r , i t s a l l e l e would be d e s i g n a t e d
1.50.
RESULTS
E l e c t r o p h o r e t i c Phenotypes o f Monomprphic P r o t e i n s
L a c ta t e dehydrogenase - (LDH) i s a t e t r a m e r composed o f s u b u n i t s
o f equal s i z e (Appel!a and Markert 1961).
LDH isozymes have been
re s o lv e d e l e c t r o p h o r e t i c a l Iy i n t o two main ty p e s d e s i g n a t e d A and
B (Markert 1962).
In mammals th u s f a r s t u d i e d , f i v e p r i n c i p a l
isozymes a r e u s u a l l y found which r e s u l t from t h e random combination
o f th e s e s u b u n i t s A and B, i n t o a l l p o s s i b l e t e t r a m e r i c s t r u c t u r e s .
Shaw and Barto (1963) provided g e n e t i c c o n f i r m a t i o n o f t h e h y p o th e s i s
t h a t th e s u b u n i t s were under d i s t i n c t g e n e t i c c o n t r o l by showing
t h a t each was c o n t r o l l e d by s e p a r a t e gene l o c i .
An a d d i t i o n a l isozyme, d e s i g n a t e d C, has been observed in mature
t e s t i s e x t r a c t s o f mammals and b i r d s (Blanco and Zinkham 1963,
Goldberg 1963).
I t has s u b s e q u e n tl y been shown t o be coded by a
s e p a r a t e locus (Blanco e t a t . 1964).
Isozyme p a t t e r n s o f t e l e o s t
f i s h pr ovide ev ide nc e o f an a d d i t i o n a l LDH l o c u s , fo r m er ly d e s i g ­
na te d E, in t h e s e animals (Markert and F a ulh ab er 1965, Whitt 1969,
1970, Markert and Holmes 1969, ShakTee e t a t . 1973).
This locus
i s now c o n s id e r e d t o be homologous t o t h e C locus o f b i r d s and
mammals, with t i s s u e e x p r e s s i o n va ryi ng with t h e f i s h s t u d i e d
(Markert e t a t . 1975).
Genetic s t u d i e s o f e l e c t r o p h o r e t i c v a r i a n t s
have e s t a b l i s h e d t h e e x i s t e n c e o f t h i s locus (Whitt e t a t . 1971).
V
32
The isozyme i s e xpr ess ed predomina ntly in nervous t i s s u e , e s p e c i a l l y
in t h e r e t i n a o f t h e eye (Goldberg 1966, Whitt 1969, 1970, Whitt
and Horowitz 1970, 1972).
In t h e ca se o f Sa1Imonid f i s h , e s p e c i a l l y t h e t r o u t , a complex
p a t t e r n o f more than f i f t e e n isozymes i s o b s e r v a b l e , w it h g e n e t i c
s t u d i e s showing t h a t t r o u t LDH i s determined by a t l e a s t f i v e d i s ­
t i n c t l o c i (Morrison and Wright 1966, Morrison 1970, Massaro and
Markert 1968, U t t e r e t a t . 1973).
t h e h y p o th e s i s o f Ohno e t a t.
Relative to t h i s observation is
(196 8) , based on c y t o l o g i c a l e v i d e n c e ,
t h a t salmonids a r e t e t r a p l o i d s .
This i n d i c a t e s t h a t t h e r e has been
d u p l i c a t i o n and su bsequent div e rg e n c e o f A and B l o c i .
The v a l i d i t y
o f th e e x i s t e n c e o f d u p l i c a t e d l o c i i s su pporte d by t h e m o le c u la r
h y b r i d i z a t i o n and immunochemical, s t u d i e s o f Massaro and Markert
(1968).
T h e r e f o r e , th e l o c i in salmonids a r e comprised o f th e
d u p l i c a t e d A (A and A1) and B (B and B1) l o c i and an a d d i t i o n a l C
lo c u s .
The complex zymogram p a t t e r n s o f LDH isozymes a r e e x p l a i n e d
by: f i v e isozymes c o n t a i n i n g A and A1 s u b u n i t s , f i v e isozymes con­
t a i n i n g B and B1 s u b u n i t s and hybrid isozymes c o n t a i n i n g B, B1 and
C subunits.
1
The v a r i o u s l o c i have d i f f e r e n t i a l t i s s u e e x p r e s s i o n .
The B
and B1 a r e e x p re s s e d in most t i s s u e s (see Massaro and Markert 1968),
e x c e p t th e l i v e r where t h e B1 predominates ( U t t e r and Hodgins 1972).
33
The A and A' l o c i a r e e x p re ss e d p r i m a r i l y .in t h e s k e l e t a l muscle
(Massaro and Markert 1968, U t t e r e t a l . 1973).
The C lo cu s i s
ex pressed in t h e eye and o t h e r neu ral t i s s u e (Horowitz and Whitt
1972).
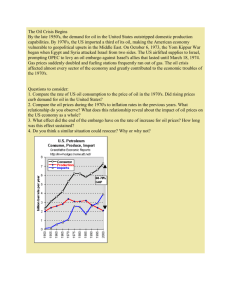
The isozyme p a t t e r n s found in T. avctious were t y p i c a l o f t h a t
found in o t h e r salmonids (F ig. 3 ) .
Five l o c i d e s i g n a t e d LDH-I,
LDH-2, LDH-3, LDH-4, and LDH-5 ( e q u i v a l e n t t o A, A ' , B, B1 and C,
r e s p e c t i v e l y ) in o r d e r o f i n c r e a s i n g anodal m i g r a t i o n , were e x p re ss e d
In muscle t i s s u e , 5 isozymes r e s u l t i n g from t h e combination o f LDH-I
and LDH-2 s u b u n i t s and 5 isozymes r e s u l t i n g from t h e combination o f
LDH-3 and LDH-4 s u b u n i t s were e x p r e s s e d .o n LDH zymograms.
group predominated as e xpect ed f o r salm onids.
The A
In h e a r t , serum and
e y e , f i v e isozymes r e s u l t i n g from t h e combination o f LDH-3 and LDH-4
s u b u n i t s (B group) were e x p r e s s e d .
An a d d i t i o n a l locus LDH-5 was
ex pr e sse d in eye t i s s u e which i s t h e C locus common t o salmonids
(Horowitz and Whitt 1972).
In l i v e r , t h e LDH-3 loc us predomi na te d,
e x h i b i t i n g a s i n g l e band in most b u f f e r syste ms .
However, in b u f f e r
system A f i v e isozymes could be r e s o l v e d , t h u s both t h e LDH-3 and
LDH-4 lo c i a r e e xp re ss ed in t h e l i v e r .
The predominance o f t h e
LDH-3 locus (B) i s in c o n t r a s t t o t h e predominance o f t h e LDH-4
( B ' ) locus in t h e l i v e r o f o t h e r salmonids ( U t t e r and Hodgins 1972,
U t t e r e t a t . 1973).
34
L
E M
M
L
b.
+
— LDH-4
^LDH-3
c.
Figure 3.
L actate Dehydrogenase (LDH). T issue d is tr ib u tio n and the
e f f e c ts o f d if f e r e n t b u ffe rs on re s o lu tio n o f th e LDH
isozym es.
a. L iv er, eye and muscle tis s u e o f th e same f is h . B uffer
system H.
b. Muscle and l i v e r tis s u e from the same f is h . B uffer
system G.
c. L iver tis s u e from th re e in d iv id u a ls . B uffer system A.
35
No a l l e l i c v a r i a t i o n was found a t any o f t h e l o c i in any o f th e
p o p u l a t i o n s examined.
I n d i v i d u a l s were homozygous a t a l l l o c i ,
t h e r e f o r e , el even isozyme ty p e s were e x p r e s s e d .
The fewer number
o f isozymes r e s u l t e d from t h e a p p a r e n t la c k o f h e t e r o t e t r a m e r s being
formed between t h e LDH-5 and LDH-3 o r LDH-4 l o c i .
The number o f LDH
isozymes in T. a p e tic u s i s e vid enc e t h a t gene d u p l i c a t i o n has ta ken
pl a ce a t t h e A and B l o c i o f LDH.
Malate dehydrogenase (MDH) - Two major c l a s s e s o f m al at e dehy­
drogenase isozymes a r e found t o e x i s t in v e r t e b r a t e s , th e m ito c h o n d ri a l
form and t h e s u p e r n a t a n t ( o r c y to p la sm ic ) form ( K i t t o and Kaplan
1966, Sie ga l and England 1961).
Both have a dim e ri c s t r u c t u r e and
a r e coded by d i s t i n c t n u c l e a r genes (Shows e t a l . 1970, Wheat e t at.
1971).
Three ty p e s o f s u p e r n a t a n t MDH isozymes AA, AB, BB, coded by 2
gene lo c i A and B, a re found in many s p e c i e s o f f i s h (Wheat and Whitt
1971, Clayton e t a l. 1971).
The i n v e s t i g a t i o n s o f B a i le y e t a t .
(1970) i n d i c a t e d t h a t t h i s was th e ca se in salm oni ds.
In t h e s a l ­
mon i d s , not only i s t h e r e evidenc e o f g e n e t i c v a r i a t i o n of. th e B form,
o f s u p e r n a t a n t MDH ( S y l n 1ko 1972, U t t e r e t a l . 1973, Clayton e t al.
1975, B a i le y e t o l . 1970, AspinwalI 1974), but t h e r e i s a l s o ev ide nc e
o f d u p l i c a t i o n o f t h e B loc us (B a il e y e t a l . 1970, U t t e r and Hodgins
1972, A l l e n d o r f 1973).
B ai le y e t a l .
(1970) pro vid ed ev id en c e from
36
isozyme dosage s t u d i e s which s ug ges te d t h a t t h e A locus i s a l s o
d u p l i c a t e d in the brown t r o u t {Salmo t r u t t a ) , w hil e Aspinwall (1974)
sug ges te d t h a t t h i s was a l s o t r u e f o r pink salmon (Oncorhynahus,
gorbusoha) .
However, t h i s has not been found in o t h e r s a l mom" d
s p e c i e s ( S y l n 1ko 1976, Clayton e t a l . 1975, A l l e n d o r f 1973).
There i s t i s s u e s p e c i f i c e x p r e s s i o n o f t h e A and B l o c i with
t h e B form pre dominating in s k e l e t a l muscle and t h e A form predom­
i n a t i n g in t h e l i v e r (Clayton e t a l . 1975, A l l e n d o r f e t a l . 1973,
A l l e n d o r f 1973, U t t e r and Hodgins 1972, B a i le y e t a l . 1970).
Massaro (1973) s t u d i e d t h e MDH isozymes o f g r a y l i n g and r e p o r t e d
t h a t th e y p o s se ss t h r e e major isozymes o f s u p e r n a t a n t MDH in s k e l e t a l
muscle and eye t i s s u e , which may correspo nd t o t h e AA, AB, and BB
isozymes p r e s e n t in o t h e r salmonids as r e p o r t e d by B a i le y e t a l .
(1970).
In t h e p r e s e n t s t u d y , t i s s u e samples o f l i v e r , s k e l e t a l
muscle, h e a r t muscle, eye and serum were surveyed for. s u p e r n a t a n t MDH
activity.
The r e s u l t s a r e shown in Fig ur e 4.
A s i n g l e band o f s t r o n g
i n t e n s i t y was found in l i v e r samp le s, whereas 3 bands were e x p re ss e d
in a l l o t h e r t i s s u e s su rveyed.
I t i s p o s t u l a t e d t h a t t h e r e a r e two
l o c i MDHg-I and MDH5 - E , c o rr e sp o n d i n g t o t h e A and B l o c i , r e s p e c ­
t i v e l y , o f o t h e r sal m on id s, coding f o r s u p e r n a t a n t MDH isozymes in T.
arotiaus.
The A form, encoded by t h e MDH5-I l o c u s , i s predominant
in th e l i v e r , while both forms A and B, (B encoded by t h e MDH5-E
37
Figure 4.
T issue d is tr ib u tio n o f M alate Dehydrogenase (MDH) from the
same f is h . Samples are o f l i v e r , m uscle, h e a r t , serum and
eye tis s u e . The MDH isozyme n ear th e o r ig in is common to
a l l tis s u e s b u t s ta in s weakly in m uscle, h e a rt and serum
samples. MDHs -I predom inates in l i v e r tis s u e . Both MDH5-I
and MDHg-2 are more eq u ally expressed in m uscle, h e a rt and
serum tis s u e .
38
lo c u s) a r e e xpr e ss ed in t h e o t h e r t i s s u e s su rv ey ed .
The t h r e e banded
phenotype seen f o r t h i s d im e ri c enzyme i s b e s t e x p l a i n e d by t h e r a n - .
dom combination o f s u b u n i t pro d u c ts from two l o c i with d i f f e r e n t
alleles.
This r e s u l t s in a f i x e d h e t e r o z y g o t e e f f e c t w it h every
i n d i v i d u a l e x h i b i t i n g t h e t h r e e banded phenotype.
No v a r i a t i o n a t e i t h e r locus was found in any o f t h e p o p u l a t i o n s
studied.
Since g r a y l i n g p o s se ss no polymorphism a t t h e l o c i coding
f o r both t h e A ty pe and B ty pe s u b u n i t s , i t was n o t p o s s i b l e t o
determine i f d u p l i c a t i o n has ta ken pla ce a t t h e s e two l o c i .
A f o u r t h band o f a c t i v i t y was a l s o found on zymograms s t a i n e d
f o r MDH a c t i v i t y .
This isozyme had a s h o r t e r m ig r a t i o n d i s t a n c e
and weaker s t a i n i n g i n t e n s i t y than t h e o t h e r fo r m s .
This isozyme
i s b e l i e v e d t o be th e m it o c h o n d ri a l form o f MDH ( d e s i g n a t e d MDHm)
based on c o r r e l a t i o n with zymograms o f o t h e r salmonids ( S y l n 'k o 1976,
Aspinwall 1973, B ai ley e t a t . 1970, B a i le y e t a t. 1969), and i s con­
t r o l l e d by a s e p a r a t e gene locus ( d i s c u s s e d p r e v i o u s l y ) .
No v a r i a t i o n
was observed f o r t h i s isozyme in any o f t h e p o p u l a t i o n s s t u d i e d .
G l u t a m a t e - o x a l o a c e t a t e t r a n s a m i n a s e (GOT) - There a r e two d i s ­
t i n c t forms o f g l u t a m a t e - o x a l o a c e t a t e t r a n s a m i n a s e in v e r t e b r a t e c e l l s
(Moore and Lee 1960), both o f which have a d im e ri c s t r u c t u r e (DeLorenzo
and Ruddle 1970).
One o f t h e forms o f GOT i s found in t h e mitochon­
d r i a l f r a c t i o n w hil e t h e o t h e r one occurs in t h e s u p e r n a t a n t (o r
39
c y to pla sm ic ) f r a c t i o n o f t h e c e l l .
The s u p e r n a t a n t form m ig r a te s
a n o d a ll y a t a n e u t r a l pH, w hi le th e m it o c h o n d r ia l form m ig r a te s
c a t h o d a l Iy ( Schmidtke and Engel 1972).
The s u p e r n a t a n t form o f
GOT has been shown t o be coded f o r by two l o c i in s e v e r a l f i s h
s p e c i e s (Schmidtke and Engel 1972).
In s a l m o n i d s , GOT has been
r e p o r t e d t o be coded by two disomic l o c i in. brown t r o u t (Schmidtke
and Engel 1972), chum salmon ( A l l e n d o r f e t a l . 1975, May e t a t .
1975) and c u t t h r o a t t r o u t ( A l l e n d o r f and U t t e r 1976).
In t h e p r e s e n t s t u d y , g r a y l i n g were surveyed f o r GOT a c t i v i t y
in l i v e r , s k e l e t a l muscle, h e a r t muscle, and eye t i s s u e :
a r e shown in Figure 5.
The r e s u l t s
The b u f f e r system employed in t h e s e p a r a t i o n
o f GOT isozymes had a high pH ( 8 . 6 ) , which r e s u l t s in t h e anodal
m ig ra tio n o f both forms ( m it och ondr ia l and s u p e r n a t a n t ) .
The r e s u l t s i n d i c a t e t h a t t h e r e a r e a t l e a s t two l o c i (GOT5-I
and GOT5 -2 ) coding f o r th e s u p e r n a t a n t form o f GOT and two l o c i
(GOTm-I and GOTm- 2) coding f o r t h e m it o c h o n d r ia l form.
There a r e two bands o f a c t i v i t y f o r t h e m it o c h o n d ri a l form with
no he te ro dim e r formed between t h e two.
This r e s u l t i s b e s t e x p l a i n e d
by th e pres enc e o f 2 l o c i (GOTm-I and G0Tm~2) coding f o r d i s t i n c t
p o ly p e p ti d e s u b u n i t s which do not i n t e r a c t with one a n o t h e r t o form
a he te ro dim e r c o n s i s t i n g o f a s u b u n i t from each l o c u s .
No v a r i a t i o n
was observed a t e i t h e r locus in any o f t h e p o p u l a t i o n s surv ey ed.
40
Figure 5.
Glutamate o x a lo a c e ta te transam inase (GOT) tis s u e d i s t r i ­
b u tio n .
a. Liver samples from th re e f is h w ith ex p ressio n o f two
GOTs and two GOTm l o c i .
b. T issue d i s tr ib u tio n o f GOT isozymes from one f is h ;
eye, m uscle, l i v e r . GOTm isozymes d id not s ta in on
th i s g e l.
41
The s u p e r n a t a n t GOT e x h i b i t e d d i f f e r e n t p a t t e r n s in t h e v a r io u s
t i s s u e s examined.
In t h e l i v e r a t h r e e banded phenotype with
asymmetrical s t a i n i n g i n t e n s i t y was found in a l l i n d i v i d u a l s .
This
p a t t e r n may be e x p l a i n e d by t h e p resen ce o f two l o c i with d i f f e r e n t
a l l e l e s coding f o r s u b u n i t s o f d i f f e r e n t e l e c t r o p h o r e t i c m o b i l i t y .
This f i x e d h e t e r o z y g o t e e f f e c t i s evidenc e f o r t h e p r e s e n c e o f a
d u p l i c a t e d lo cu s ( A l l e n d o r f e t d l . 1975).
In eye t i s s u e a s i m i l a r
t h r e e banded p a t t e r n was t h e phenotype o f a l l i n d i v i d u a l s , however,
the asymmetry o f s t a i n i n g i n t e n s i t y was in t h e o p p o s i t e d i r e c t i o n
(Fig ure 5b).
This p a t t e r n ag ain s u g g e s ts a d u p l i c a t e d l o c u s .
The
asymmetry o f i n t e n s i t y cannot be e x p la i n e d by t h e p o s t u l a t e t h a t one
o f t h e l o c i i s monomorphic f o r t h e common a l l e l e w hil e t h e o t h e r i s
polymorphic s i n c e : I ) a l l i n d i v i d u a l s e x p re s s t h e same p a t t e r n which
would not be ex pec te d i f t h e locus was polymorphic (some would be
sy m m e tr ic a l) , and 2) t h e r e i s o p p o s i t e asymmetry in a d i f f e r e n t t i s s u e
o f t h e same i n d i v i d u a l .
The l a c k o f p a r a l l e l e x p r e s s i o n between eye
and l i v e r s u g g e s t s t h a t t h e p a t t e r n i s t h e r e s u l t o f d i f f e r e n t i a l
a c t i v a t i o n o f t h e two l o c i in t h e s e d i f f e r e n t t i s s u e s .
In muscle, only one band o f s u p e r n a t a n t GOT a c t i v i t y was
e x p re ss e d .
The band had t h e same e l e c t r o p h o r e t i c m o b i l i t y as th e
GOTg-I homozygote in l i v e r .
There i s evidenc e in some salmonids
t h a t t h i s band may r e p r e s e n t a d i s t i n c t locus ( A l l e n d o r f p e rs onal
42
comm., A l l e n d o r f and U t t e r 1976).
However, t h e la c k o f v a r i a t i o n
a t t h i s locus does not al low t h e c o n f i r m a t i o n o f t h i s p o s t u l a t e in
grayling.
T h e r e f o r e , t h e c o n s e r v a t i v e e s t i m a t e o f two s u p e r n a t a n t
GOT lo c i and two mit oc ho nd ria l GOT l o c i was used in t h e p r e s e n t
stu dy.
Alcohol dehydrogenase - (ADH) has been shown t o have a dim e ri c
s t r u c t u r e in v e r t e b r a t e s ( B u t l e r e t d l . 1969).
I t i s b e l i e v e d t o be
under th e c o n t r o l o f a s i n g l e gene loc us in salmonids ( A l l e n d o r f
e t d l. 1975).
The enzyme in t r o u t a ppea rs t o be n e g a t i v e l y c ha r g e d ,
m ig r a ti n g t o t h e cat hode.
In most salmonids s t u d i e d , ADH appea rs
as a s i n g l e band with l i t t l e o r no v a r i a t i o n w i t h i n t h e s p e c i e s
( K r i s t i a n s s o n and McIntyre 1965, Gall e t d l . 1977, U t t e r e t d l . 1973,
A l l e n d o r f e t d l . 1975).
In T. are tic us l i v e r samples were examined f o r ADH a c t i v i t y .
In
a l l i n d i v i d u a l s ADH was e x p r e s s e d as an i d e n t i c a l s i n g l e band o f
a c t i v i t y , migrating cathodally.
On th e b a s i s o f t h e s e r e s u l t s i t
was p o s t u l a t e d t h a t ADH i s encoded by a s i n g l e loc us in g r a y l i n g .
The ADH loc us was monomorphic and i d e n t i c a l in a l l p o p u l a t i o n s
surveyed with no v a r i a t i o n between p o p u l a t i o n s .
Xanthine dehydrogenase - (XDH) i s r e p r e s e n t e d in salmonids by a
s i n g l e band o f a c t i v i t y which m i g r a t e s a n o d a l l y ( K r i s t i a n s s o n e t a t.
1976, A l l e n d o r f e t d l . 1975, A l l e n d o r f 1973).
Since no v a r i a t i o n
43
e x i s t s a t t h i s locus in t h e s p e c i e s s t u d i e d , no c o n c l u s i o n s can be
made as t o whe the r XDH i s encoded by a s i n g l e lo c u s o r d u p l i c a t e d
locus ( A l l e n d o r f e t a t . 1975).
In T. arct-ious a s i n g l e band o f XDH a c t i v i t y was found which
migrated a n o d a l l y .
All p o p u l a t i o n s were monomorphic with no v a r i a ­
t i o n d e t e c t e d between p o p u l a t i o n s .
i s coded f o r by a s i n g l e l o c u s .
I t i s assumed t h a t t h e enzyme
However, t h e p o s s i b i l i t y o f two
l o c i with i d e n t i c a l e l e c t r o p h o r e t i c gene p ro d u c ts cann ot be ex clu ded .
S o r b i t o l dehydrogenase - (SDH) i s presumed t o have a t e t r a m e r i c
s t r u c t u r e in v e r t e b r a t e s (O p tl Hof 1969, O p t1Hof e t a l . 1969, Engel
e t a l. 1970).
Engel e t a l .
(1970) proposed t h a t . t h i s i s a l s o the
s t r u c t u r e o f SDH found in rainbow t r o u t , and has r e p o r t e d th e
pres enc e o f a polymorphism.
I n v a r i a n t m ult i- ban de d phenotypes have
been r e p o r t e d in v a r io u s o t h e r salmonid s p e c i e s (May e t a l . 1975,
Khanna e t a l . 1975, A l l e n d o r f e t a l . 1975, U t t e r e t a l . 1973).
The
p a t t e r n s in t h e s e s p e c i e s i s i n d i c a t i v e o f two disomic l o c i f i x e d
f o r a l l e l e s coding f o r s u b u n i t s o f d i f f e r e n t e l e c t r o p h o r e t i c m o b i l i t y
( A ll e n d o rf e t a l . 1975, U t t e r e t a l . 1973).
c o n t r a s t to t h a t o f Engel e t a l .
This p o s t u l a t e i s in
(1970) which proposed a t e t r a s o m i c
mode o f i n h e r i t a n c e f o r a s i n g l e lo cu s based on t h e ph e noty pic d i s ­
t r i b u t i o n o f isozyme p a t t e r n s with no bre e d in g ex periments performed.
Allendorf e t al.
(1975) on t h e b a s i s o f p r e l i m i n a r y i n h e r i t a n c e
44
d a ta o f a v a r i a n t in c u t t h r o a t t r o u t , prov ide f u r t h e r ev id en c e in
su ppo rt o f t h e former p o s t u l a t e .
In t h e p r e s e n t s t u d y , g r a y l i n g e x h i b i t e d a d i s t i n c t s i n g l e band
phenotype o f SDH a c t i v i t y , which mi grated a n o d a l l y .
l a t i o n a l o r i n t e r p o p u l a t i o n a l v a r i a t i o n was ob ser ved .
No i n t r o p o p u This r e s u l t
allows no c o n c lu s io n s t o be drawn as t o t h e pre s e n c e o f a d u p l i c a t e d
locus in T. o p e tic u s as proposed f o r rainbow t r o u t ( A l l e n d o r f e t a l .
1975).
For t h e purposes o f t h i s s tu dy t h e c o n s e r v a t i v e e s t i m a t e o f
one locus coding f o r SDH was used.
I s o c i t r a t e dehydrogenase (mi to ch on dri al form) - (IDH) e x i s t s
in d i s t i n c t m it oc hond ri a l (he re d e s i g n a t e d IDHm) and s u p e r n a t a n t
(IDHg) forms which a re determined by d i s t i n c t gene l o c i in v e r t e b r a t e s
(Henderson 1968).
Wolf e t a l .
( 1 9 7 0 ) , showed t h a t IDHffl m a n i f e s t s a
dimeric s t r u c t u r e in e l e c t r o p h o r e t i c s t u d i e s o f rainbow t r o u t {salmo
g airdn eri) .
Engel e t a l.
(1971) showed t h a t in some d i p l o i d groups
o f f i s h , a s i n g l e gene lo cu s presumably coded f o r IDHffl, whereas in
some t e t r a p l o i d groups two d i f f e r e n t gene l o c i were r e s p o n s i b l e f o r
t h e f i x e d h e t e r o z y g o t e p a t t e r n observed on zymograms.
Allendorf
e t a l . (1975) r e p o r t e d t h a t in rainbow t r o u t {Salmo galvdnevi) IDHffl
i s a l s o r e p r e s e n t e d by t h r e e n o n v a r i a n t bands i n d i c a t i n g t h e pr e s en c e
o f two monomorphic disomic l o c i with common a l l e l e s coding f o r su b­
units of d if f e r e n t e lec tro p h o retic m o b ilitie s .
IDHffl a c t i v i t y i s
b e s t v i s u a l i z e d on zymograms o f h e a r t muscle e x t r a c t s .
45
In g r a y l i n g , h e a r t muscle e x t r a c t s were surveyed f o r IDHfil
activity.
IDHjfi phenotypes were r e p r e s e n t e d by t h r e e i n v a r i a n t bands
which i n d i c a t e t h e pres enc e o f two monomorphic disomic l o c i , IDHjfi-I
and IDHjfi-Z, w it h common a l l e l e s coding f o r s u b u n i t s o f d i f f e r e n t
electrophoretic m obilities.
The p a t t e r n observed i s ev ide nc e t h a t
t h e IDHjfi loc us in T. arotious i s d u p l i c a t e d as presumed t o . b e t h e
case in rainbow t r o u t ( A l l e n d o r f e t a t . 1975).
All i n d i v i d u a l s
ex pre sse d t h e same phenotype in a l l p o p u l a t i o n s s t u d i e d .
Alpha -g ly c er op ho sp ha te dehydrogenase - (AGPDH) i s a d im e ric
molecule f o r which g e n e t i c v a r i a t i o n has been d e s c r i b e d in v a ri o u s
s p e c i e s o f f i s h (Aspinwall 1972, U t t e r and Hodgins 1972, AlTendorf
e t a l . 1975, Engel e t a t . 1971, Johnson e t a l . 1970, McCabe e t a t.
1970).
Engel e t a l . (1971) proposed t h e e x i s t e n c e o f t h r e e d i f f e r e n t
gene lo c i (A, B, and C) coding f o r AGPDH in t h e brown t r o u t {Salmo
t ru tta ) and t h e rainbow t r o u t {,Salmo gaivdneri) with t h e random
combination o f s u b u n i t s from a l l e l e s a t t h e v a r i o u s l o c i a c c o u n ti n g
f o r t h e complex zymogram p a t t e r n s .
Other a u t h o r s (AspinwalT 1972,
U t t e r and Hodgins 1 9 7 2 ) .have proposed th e e x i s t e n c e o f a s i n g l e
l o c u s , two codominant a l l e l e system in o t h e r salmonids on t h e b a s i s
of electrophoretic re su lts.
Aspinwall (1972) s u g g es te d t h e p resen ce
o f a s i n g l e loc us in salmonids may be due t o a " s i l e n c i n g " o f
d u p l i c a t e d l o c i in salmonids.
However, A l l e n d o r f e t a l .
(1975)
46
provided ev id en c e t h a t t h e b u f f e r system employed in t h e e l e c t r o ­
p h o r e s i s o f AGPDH deter mine s t h e number o f isozymes and hen ce, t h e
number o f l o c i which a r e d e t e c t e d .
A low pH phosphate b u f f e r allows
th e d e t e c t i o n o f an added number o f l o c i which a r e n o t d e t e c t e d with
th e use o f a high pH t r i s - b o r a t e system.
In t h e p r e s e n t s t u d y , t h r e e l o c i (AGPDH-I, AGPDH-2 and AGPDH-3)
with a l l e l e s coding f o r p r o t e i n s o f d i f f e r e n t e l e c t r o p h o r e t i c
m o b i l i t i e s a r e p o s t u l a t e d t o e x i s t in T. arot-ious.
The zymograms
ob ta in e d r e f l e c t t h e pr e s en c e o f t h e s e t h r e e l o c i w ith a c t i v e dimers
formed from t h e s u b u n i t s o f th e d i f f e r e n t l o c i , r e s u l t i n g in s i x
isozymes being formed.
L i v e r and muscle t i s s u e e x t r a c t s e x h i b i t e d
p a r a l l e l e x p r e s s i o n o f t h e i d e n t i c a l number o f isozymes.
Examination
o f t h i s isozyme system with t h e high pH b u f f e r system r e s u l t e d in
th e d e t e c t i o n o f only t h e AGPDH-I lo c u s .
F u r t h e r s t u d i e s usi ng
the low pH b u f f e r system r e s u l t e d in th e d e t e c t i o n o f t h e two
additional lo c i.
These f i n d i n g s a r e s i m i l a r t o th o s e o f Engel e t at.
(1971) and r e f l e c t th e d i f f e r e n c e s which a r e observed depending on
t h e b u f f e r system employed as su g g e s te d by A l l e n d o r f e t a t .
(1975).
No a l l e l i c v a r i a t i o n a t any o f th e AGPDH l o c i was d e t e c t e d in
any o f th e p o p u l a t i o n s surveyed.
E s t e r a s e - The banding p a t t e r n s observed in l i v e r samples o f
Thymallus arotiaus were i n c o n s i s t e n t in number and i n t e n s i t y .
Due
47
t o th e i n c o n s i s t e n c y , t h e l i v e r e s t e r a s e s were n o t d e a l t with in
any d e t a i l .
The e s t e r a s e o f th e serum was r e p r e s e n t e d by a s i n g l e i n v a r i a n t
band in a l l p o p u l a t i o n s .
I t was assumed t h e band was r e p r e s e n t a t i v e
o f a s i n g l e gene lo c u s.
Hexokinase - (HK) has been r e p o r t e d t o be r e p r e s e n t e d by a
s i n g l e band in rainbow t r o u t ( A l l e n d o r f 1973).
L i v e r samples o f
T. arcticus surveyed f o r hexokinase a c t i v e l y e x h i b i t e d an i d e n t i c a l
s i n g l e band in a l l p o p u l a t i o n s examined.
I t i s assumed t h e band
i s r e p r e s e n t a t i v e o f a s i n g l e gene lo c u s .
E l e c t r o p h o r e t i c Phenotypes o f Polymorphic P r o t e i n s
T e tr a z o li u m oxi da se ( Indophenol o x id a s e ) - TO i s th e d e s i g n a t i o n
f o r an enzyme f i r s t d e s c r i b e d by Brewer (1967) which o x i d i z e d reduced
t e t r a z o l i urn dyes r e s u l t i n g in an achr oma ti c re gi on a g a i n s t the
c o lo re d background o f reduced dye on e l e c t r o p h o r e t i c zymograms.
I n t r a s p e c i f i c v a r i a n t s o f TO were found t o e x i s t in f i f t e e n s p e c i e s
o f P a c i f i c r o c k f i s h [Sebastodes) (Johnson e t a t . 1970b).
Inter­
s p e c i e s v a r i a t i o n o f TO has been r e p o r t e d t o e x i s t in th e salmonids
( U t t e r 1971, U t t e r e t a t. T973, May e t a t . 1975).
The e x i s t e n c e o f
i n t r a s p e c i e s TO polymorphisms have a l s o been found in t h e rainbow
t r o u t ( U t t e r 1971, U t t e r e t a t . 1973, Cederbaum and Yoshida 1972,
U t t e r and Hodgins 1972, A l l e n d o r f 1973) and Chinook salmon ( U t t e r
48
1971, K r i s t i a n s s o n and McIntrye 1976).
The p a t t e r n s obser ved ap pea r
t o r e f l e c t one locus w it h two a l l e l e s encoding a d im e r ic p r o t e i n
( U t t e r 1971, Cederbaum and Yoshida 1972).
In T. are tie us i n d i v i d u a l s e x h i b i t e d e i t h e r one zone o r t h r e e
zones o f a c t i v i t y (Fig ure 6) s i m i l a r t o t h e p a t t e r n s found in o t h e r
salmonids ( d i s c u s s e d p r e v i o u s l y ) .
P a r a l l e l t i s s u e e x p r e s s i o n was
found in l i v e r and muscle e x t r a c t s o f t h e same i n d i v i d u a l .
No
i n t e r p o p u l a t i o n a l v a r i a t i o n was found.
The common a l l e l e (TO^" ^ ) , and a v a r i a n t a l l e l e ( t O " ^ ) , were
found in t h e Donnelly, Wolf and Grebe p o p u l a t i o n s .
Homozygotes
( 1 . 0 0 / 1 . 0 0 ) f o r t h e common a l l e l e e x p re ss e d a s i n g l e band o f a c t i v i t y
which mi gr at e d th e f a r t h e s t a n o d a l l y (F ig. 6 ) , h e t e r o z y g o t e s ( 1 . 0 0 /
.50) e x h i b i t e d a 3 banded p a t t e r n w it h a h e t e r o d i meric zone o f
g r e a t e r i n t e n s i t y , and homozygotes ( . 5 0 / . 5 0 ) f o r the v a r i a n t a l l e l e
e x h i b i t e d a s i n g l e band with t h e . s l o w e s t m i g r a t i o n .
I t i s assumed
t h a t th e p a t t e r n s r e f l e c t a s i n g l e l o c u s , two codominant a l l e l e
system s i m i l a r t o t h a t su g g e s te d f o r rainbow t r o u t ( U t t e r 1971,
Cederbaum and Yoshida 1972).
No v a r i a t i o n was found in t h e Fuse Lake p o p u l a t i o n with a l l
i n d i v i d u a l s being homozygous f o r t h e common a l l e l e .
The la ck o f
v a r i a t i o n in t h i s p o p u la ti o n i s presumably a t t r i b u t a b l e t o a founder
effect.
49
1
Figure 6.
2
3
4
5
T etrazolium Oxidase (TO) polymorphism. Column I (0.5 0 /0 .5 0 )
homozygote. Column 2, 3, 4, (1 .0 0 /0 .5 0 ) h e te ro z y g o te s .
Column 5 (1 .0 0 /1 .0 0 ) homozygote.
50
Phosphoglucomutase - (PGM) isozyme v a r i a t i o n was f i r s t s t u d i e d
in human p o p u l a t i o n s by Spencer e t a t . (1964).
Further genetic
s t u d i e s in humans showed t h e PGM a c t i v i t y could be d i v i d e d i n t o
t h r e e z o n e s , with each zone a p p a r e n t l y c o n t r o l l e d by an indepen de nt
locus ( F a r r i n g t o n e t a t . 1968).
Roberts e t a t .
(1969) r e p o r t e d
f i n d i n g t h r e e d i s t i n c t zones o f a c t i v i t y on PGM zymograms o f Satmo
Qaivdnerii and p o s t u l a t e d t h e e x i s t e n c e o f t h r e e l o c i , PGM-I, PGM-2
and PGM-3 ( i n o r d e r o f i n c r e a s i n g anodal m ig r a ti o n o f t h e coded
protein).
Polymorphism o f PGM have been found in rainbow t r o u t
(Roberts e t a t . 1969, U t t e r e t a t . 1972, 1973) and in v a r i o u s s p e c i e s
o f salmon ( K r i s t i a n s s o n and McIntyre 1976, May e t a t . 1975, U t t e r
e t a t . 1973, U t t e r and Hodgins 1970).
In t h e p r e s e n t s t u d y , l i v e r and muscle t i s s u e e x t r a c t s were
surveyed f o r PGM a c t i v i t y .
Three zones o f a c t i v i t y were d e t e c t e d
on PGM zymograms o f l i v e r sa mp le s, a l l o f which m ig r a te a n o d a l Iy
(Figure 7).
Three l o c i (PGM-I , PGM-2 and PGM-3) were p o s t u l a t e d to
be encoding PGM s u b u n i t s in T. avetieus-.
ex pr e sse d in muscle t i s s u e .
Only PGM-I and PGM-2 were
The f a i n t s t a i n i n g o f t h e PGM-3 in
l i v e r t i s s u e and i t s absence in muscle t i s s u e i s c o n s i s t e n t with
th e f i n d i n g s o f o t h e r a u t h o r s (Roberts e t a t . 1969, Hopkinson and
H a r r i s 1968).
PGM-I and PGM-3 were monomorphic in a l l p o p u l a t i o n s s t u d i e d ,
r e p r e s e n t e d by s i n g l e i n v a r i a n t bands.
PGM-2 was polymorphic in t h e
51
+
A
PGM-5
PGM-2
PGM-I
Figure 7.
Phosphoglucomutase (PGM). The products o f th re e PGM
lo c i appear to be p re se n t in liv e r samples. PGM-I ,
PGM-2 and PGM-3. PGM2 is polym orphic, PGM-I and PGM3
are monomorphic.
52
Donnelly R ive r p o p u l a t i o n , but was monomorphic in t h e o t h e r po pula ­
t i o n s su rveyed.
Two a l l e l e s , PGM-2^'^ and PGM-2^
in th e Donnelly p o p u l a t i o n .
, were p r e s e n t
Homozygous i n d i v i d u a l s f o r e i t h e r
a l l e l e were r e p r e s e n t e d by a s i n g l e band w hile he ter ozy gous i n d i ­
v i d u a l s e x h i b i t e d two bands o f a c t i v i t y (F igu re 8) which i s t y p i c a l
o f a monomeric p r o t e i n .
The p a t t e r n s found a r e s i m i l a r t o th o s e
r e p o r t e d in o t h e r salmonids (noted ab ove).
The e x i s t e n c e o f th e
polymorphism a t th e PGM-2 lo cu s c l a r i f i e s t h e i n t e r p r e t a t i o n o f
zymogram p a t t e r n s and s t r e n g t h e n s t h e v a l i d i t y o f t h e t h r e e l o c i
postulate.
I s o c i t r a t e dehydrogenase ( s u p e r n a t a n t form) - The s u p e r n a t a n t
form o f i s o c i t r a t e dehydrogenase (IDHg) behaves e l e c t r o p h o r e t i c a l Iy
as a d im e ric molecule (Henderson 1968, Darnall and Klotz 1972).
A l l e l i c v a r i a t i o n o f IDHg isozymes has been d e s c r i b e d in a v a r i e t y
o f salmonid s p e c i e s ( A l l e n d o r f
a t . 1975, May e t a t . 1975, A l l e n -
d o r f T973, Ropers e t a t . 1973, Engel e t a t . 1975, Wolf e t a t . 1970)
which co nfirm t h e d im e ri c s t r u c t u r e in salmonid f i s h .
The mode o f
i n h e r i t a n c e o f IDHg was i n i t i a l l y proposed t o be t e t r a s o m i c in
rainbow t r o u t (Wolf e t a t . 1970) on t h e b a s i s o f p h e noty pic e x p r e s s i o n
a lon e.
A l l e n d o r f (1973) and A l l e n d o r f and U t t e r (1973) d e s c r i b e d a
system o f IDHg isozymes i d e n t i c a l t o t h a t found by Wolf e t a t . . (1970).
Based on th e number o f bands and r e l a t i v e dos age , a two l o c u s , f o u r
53
+
1.25
1. 00
PGM-I
1
Figure 8.
2
3
4
5
Phosphoglucomutase (PGM). PGM-I, c a th o d a l, is monomoro h ic . PGM2 is polym orphic. Column 2 and 3 (1 .0 0 /1 .0 0 )
homozygotes. “Column I and 4 (1 .0 0 /1 .2 5 ) h etero zy g o tes.
Column 3 (1 .2 5 /1 .2 5 ) homozygote.
54
a l l e l e system was proposed t o be encoding IDHg in t h e s e i n d i v i d u a l s .
Breeding experiments v e r i f i e d t h a t in t h e s t o c k o f rainbow t r o u t
examined by t h e s e a u t h o r s , IDHg followed a disomic mode o f i n h e r i t a n c e
with t h e lo cu s having been d u p l i c a t e d .
Subsequent t o t h e s e f i n d i n g s ,
bre ed in g ex periments were performed with th e s t o c k examined by Wolf
e t a l . (1970) which c o n c l u s i v e l y v e r i f i e d t h a t , i n d e e d , IDHg followe d
a disomic mode o f i n h e r i t a n c e (Engel e t a l . 1975).
L i v e r t i s s u e e x t r a c t s o f T. arcticus were examined f o r IDHg
a c t i v i t y and re v e a le d t h e system d e p i c t e d in Fig ur e 9.
The IDHg
p a t t e r n s were polymorphic in t h r e e o f t h e p o p u l a t i o n s su rv ey ed .
The
e x p r e s s i o n o f only t h r e e phenotypes in any one p o p u l a t i o n i s i n d i c a ­
t i v e o f a s i n g l e disomic lo cu s with a common and a v a r i a n t a l l e l e .
This h y p o t h e s i s o f a s i n g l e disomic lo cu s f o r T. avcticus i s in
c o n t r a s t t o th e two disomic l o c i p o s t u l a t e d f o r rainbow t r o u t
( A l l e n d o r f and U t t e r 1973, Engel e t a l . 1975).
The IDHg locus does
no t a p p ea r t o have undergone gene d u p l i c a t i o n as in rainbow t r o u t
( A l l e n d o r f e t a l . 1975).
A s i n g l e disomic loc us encoding IDHg has
been r e p o r t e d in chum salmon {Oneorhynchus keta) and confirmed
through bre e d in g experiments (May e t a l . 1975).
A l l e l i c v a r i a t i o n was observed a t t h e IDHg locus in t h e Donnelly
R iv e r, Wolf Lake, and Grebe Lake p o p u l a t i o n s .
d e s i g n a t e d IDH
The common a l l e l e
encoded s u b u n i t s which were e l e c t r o p h o r e t i c a l l y
55
Figure 9.
I s o c i t r a t e Dehydrogenase (IDH) is polym orphic. Three
p o p u latio n s have an IDH v a ria n t and one a l l e l e common to
a l l p o p u la tio n s. Column I (1 .2 5 /1 .0 0 ) h etero zy g o te.
Column 2 3, 4, 5, 7 (1 .0 0 /1 .0 0 ) homozygotes. Column 6
(1 .5 0 /1 .0 0 ) h etero zy g o te. Column 8 (1 .2 5 /1 .2 5 ) homozygote.
Column 9 (1 .5 0 /1 .5 0 ) homozygote. Samples I , 2, 3, 8 are
from th e Donnelly r iv e r p o p u latio n . Samples 4, 5, 6, 7,
9 from the Grebe lake p o p u latio n .
f
56
i d e n t i c a l in a l l p o p u l a t i o n s .
I 25
A v a r i a n t a l l e l e , d e s i g n a t e d IDH5 " ,
was unique t o t h e Donnelly R ive r p o p u l a t i o n .
A homozygous i n d i v i d u a l
f o r t h e v a r i a n t a l l e l e ( 1 . 2 5 / 1 : 2 5 ) e x p re ss e d a s i n g l e band o f a c t i v i t y
with a g r e a t e r m ig r a ti o n d i s t a n c e than a homozygous i n d i v i d u a l f o r
th e common a l l e l e ( I . 0 0 / 1 . 0 0 ) .
A hetero zyg ous i n d i v i d u a l
(1.00/1.25)
e x h i b i t e d a t h r e e banded p a t t e r n with symmetrical s t a i n i n g i n t e n s i t y ,
c h a r a c t e r i s t i c of a d im e ri c molecule.
I 50
Another v a r i a n t a l l e l e , IDH5 " , was unique t o t h e two Yellow­
s ton e Park p o p u l a t i o n s .
The isozyme o f an i n d i v i d u a l homozygous f o r
t h i s a l l e l e had a c h a r a c t e r i s t i c m ig r a ti o n d i s t a n c e g r e a t e r than
e i t h e r the 1.00/1.00 in d iv id u als or the 1.25/1.25 in d iv id u a ls .
A
heterozygous i n d i v i d u a l f o r t h i s v a r i a n t ( 1 . 0 0 / 1 . 5 0 ) a gai n e x p re s s e d
a t h r e e banded phenotype.
As noted in Fig ur e 9 a 1 . 0 0 / 1 . 5 0 i n d i v i d u a l
has an asymmetrical s t a i n i n g i n t e n s i t y which may r e f l e c t a d i f f e r ­
e n t i a l a c t i v a t i o n o f t h e two a l l e l e s o r d i f f e r e n t i a l a c t i v i t y o f
these a l l e l e products.
Only 1 . 0 0 / 1 . 0 0 i n d i v i d u a l s were found in t h e Fuse Lake popula ­
t i o n which may ag ain r e f l e c t a fou nde r e f f e c t .
T r a n s f e r r i n - (Tfn) i s a monomeric B -g lo b u li n which e x h i b i t s a
high degree o f polymorphism in most v e r t e b r a t e s p e c i e s (Manwell and
Baker 1970).
The e x i s t e n c e o f an e x t e n s i v e amount o f g e n e t i c v a r i a ­
t i o n has been r e p o r t e d in t e l e o s t s , with more than t h i r t y s p e c i e s
57
being polymorphic (reviewed by Kirpic hnikov 1973, DeLigny 1969).
The
m a j o r i t y o f t h e s e polymorphisms r e f l e c t codominant i n h e r i t a n c e o f
a l l e l e s a s s o c i a t e d with a s i n g l e locus (K ir pic hnik ov 1973).
Inheri­
tance s t u d i e s v e r i f i e d t h e i n t e r p r e t a t i o n o f one polymorphic disomic
locus (Valenta e t a l . 1976, U t t e r e t a l. 1973).
The l a r g e amount o f
v a r i a b i l i t y o f t r a n s f e r r i n i s well documented in salmonid s p e c i e s
(Hershberger 1970, Wright and A therton 1970, Mo lle r 1970, U t t e r
e t a l . 1970, U t t e r and Hodgins 1972, Eckroat 1973, A l l e n d o r f 1973,
U t t e r e t a l . 1973).
In a l l c ase s t h e r e i s a simple a d d i t i v e p a t t e r n
in h e t e r o z y g o t e s f o r a t r a n s f e r r i n polymorphism.
The pre s e n c e o f
t h r e e o r more a l l e l e s has been observed in rainbow t r o u t and v a r i o u s
salmon s p e c i e s ( U t t e r e t a l . 1970, 1973, U t t e r and Hodgins 1972,
Reichenbach-Klinke 1973) and a l s o in brook t r o u t (Eckr oat 1973).
Serum samples o f i n d i v i d u a l g r a y l i n g were an al yz e d f o r t r a n s ­
f e r r i n phenotypes.
The t r a n s f e r r i n bands (F ig ur e 10) were i d e n t i f i e d
as t h e bands having an e l e c t r o p h o r e t i c m o b i l i t y s i m i l a r in p o s i t i o n ,
t o th os e o f rainbow t r o u t and salmon ( U t t e r e t a l . 1970, 1973,
Reichenbach-Klinke 1973).
I t i s assumed t h a t t h e s e bands in T.
a p o tieu s a r e a l s o t r a n s f e r r i n s because o f t h i s e l e c t r o p h o r e t i c
i d e n t i t y and t h e taxonomic r e l a t i o n s h i p o f t h e s e s p e c i e s .
T r a n s f e r r i n was found t o be polymorphic in t h e Donnelly R i v e r ,
Wolf Lake, and Grebe Lake p o p u l a t i o n s .
On t h e b a s i s o f observed
> +
58
Figure 10.
T ra n s fe rrin polymorphism (T fn ). Two a l l e l e s were found
in th e Donnelly r iv e r p o p u latio n and th re e in the
Yellowstone Park p o p u la tio n s. Columns I , 2, 3, 4, 5
7, 8, 9, 11, 12 (1 .0 0 /1 .0 0 ) homozygotes. Column 6
(1 .2 0 /1 .2 0 ) homozygote. Column 10 (1 .1 0 /1 .0 0 ) h e te r o ­
zygote. Column 1 , 3 , 7 Donnelly r iv e r . A ll o th e rs from
Grebe lak e.
59
phenotypes and th e monomeric s t r u c t u r e o f T f n , t h r e e codominant
a l l e l e s ( T f n ^ T f n ^ ' and T f n ^ in o r d e r o f i n c r e a s i n g anodal
m ig r a ti o n ) a t a s i n g l e disomic locus were p o s t u l a t e d t o encode Tfn in
th e two Yellowstone Park p o p u l a t i o n s .
Six phenotypes o f e i t h e r one
o r two o f t h r e e d i f f e r e n t bands a r e ex pec te d t o r e s u l t from t h e
codominant e x p r e s s i o n o f t h r e e a l l e l e s .
The s i x phenotypes a c c o r ­
d i n g ly a r e d e s i g n a t e d 1 . 0 0 / 1 . 0 0 , 1 . 0 0 / 1 . 1 0 , I . 0 0 / 1 . 2 0 , I . 1 0 / 1 . 1 0 ,
1 .1 0 / 1 . 2 0 and 1 .2 0 / 1 . 2 0 .
Figure 10.
Four o f t h e s e phenotypes a r e shown in
The only phenotype no t found in t h e i n d i v i d u a l s sampled
was I . 1 0 / 1 . 2 0 .
The l a c k o f o b s e r v a t i o n o f t h i s phenotype i s most
l i k e l y t h e r e s u l t o f th e small sample s i z e r e s u l t i n g in no d e t e c t i o n
o f I . 1 0 / 1 . 2 0 whose maximum ex pec te d val ue i s .013.
E l e c t r o p h o r e t i c a l l y i d e n t i c a l a l l e l e s t o T f n ^ and Tfn^
I 20
were e x p r e s s e d in th e Donnelly R ive r p o p u l a t i o n , bu t t h e Tfn *
a l l e l e was a b s e n t .
The t h r e e e xpect ed phenotypes ( 1 . 0 0 / 1 . 0 0 , 1 . 0 0 /
1.10 and 1 . 1 0 / 1 . 1 0 ) r e f l e c t i n g two codominant a l l e l e s were obser ve d.
The common a l l e l e T f n ^ ’ ^
was t h e only one e x p re s s e d in t h e Fuse
Lake p o p u l a t i o n , thus only th e common phenotype ( 1 . 0 0 / 1 . 0 0 ) was
observed.
The lack o f v a r i a t i o n a t t h e Tfn locus in t h i s p o p u l a t i o n
i s presumably due t o a fou nde r e f f e c t .
Glucose and hexose 6-p ho sp hat e dehydrogenase - Two major e l e c t r o p h o r e t i c a l l y d i s t i n c t components o f glu co se 6-p hosp ha te dehydrogenase
60
a c t i v i t y a r e found in v e r t e b r a t e organisms.
One i s h i g h l y s p e c i f i c
f o r t h e u t i l i z a t i o n o f g lu c os e 6-ph os pha te with NADP as s u b s t r a t e
and coenzyme, r e s p e c t i v e l y (Noltman and Kuby 1963).
The second form
i s d i s t i n g u i s h e d by i t s a b i l i t y , t o c a t a l y z e t h e o x i d a t i o n o f glu co se
6 -p h o s p h a te , g a l a c t o s e 6 - p h o s p h a te , 2-deoxyglucose 6-p hos phat e and
glucose as s u b s t r a t e s , with e i t h e r NAD o r NADP f u n c t i o n i n g as th e
coenzyme (Shaw 1966, Ohno e t a t . 1966, B e u t l e r and Morrison 1967,
Shaw and Koen 1968).
The isozyme with g lu c ose 6-p hos ph a te s p e c i f i c
dehydrogenase a c t i v i t y i s d e s i g n a t e d glu co se 6-p hos phat e dehydrogen­
ase (G6PD), w hi le t h e enzyme w it h t h e b r o a d e r range o f s u b s t r a t e
s p e c i f i c i t y i s d e s i g n a t e d hexose 6-ph os pha te dehydrogenase (H6PD)
f o r convenience o f d i s t i n c t i o n (Shaw 1966, Ohno e t a l . 1966, Shaw
and Koen 1968).
V e r t e b r a t e G6PD and H6PD isozymes have been shown t o be encoded
by d i s t i n c t gene l o c i .
G6PD i s sex l i n k e d in mammals ( Kirkman and
Hendrickson 1963, Richardson e t a l . 1971) bu t i s under autosomal
gene c o n t r o l in b i r d s and f i s h (Manwell and Baker 1969, Yamauchi
and Goldberg 1973).
G6PD i s l o c a l i z e d in t h e n u c l e a r and c y to pl a sm ic
f r a c t i o n s o f t h e c e l l ( B e u t l e r and Morrison 1967).
In c o n t r a s t ,
H6PD i s a uto so m al ly i n h e r i t e d in mammals (Shaw 1966, Ohno e t a l.
1966, Shaw and Koen 1968) and i s l o c a l i z e d in th e microsomal f r a c t i o n
o f t h e c e l l ( B e u t l e r and Morrison 1967, Metzger e t a l . 1965).
61
, G6PD e x i s t s as two c a t a l y t i c a l l y a c t i v e f o r m s , dimer and t e t r a m e r
(Bonsign or e e t a l . 1970)» being mostly t e t r a m e r i c in some organisms
(Yamauchi and Goldberg 1973) b u t dim e ri c in o t h e r s (Shaw and Koen
1968).
In c o n t r a s t H6PD e x h i b i t s a dim e ri c s t r u c t u r e as ev idenced
in zymograms o f a l l e l i c v a r i a n t s (Stegeman and Goldberg 1971).
The t i s s u e d i s t r i b u t i o n o f G6PD appea rs u b i q u i t o u s in v e r t e b r a t e
c e l l s (B e u t l e r and Morrison 1967).
H6PD i s most a c t i v e in t h e l i v e r
and kidney ( B e u t l e r and Morrison 1967) with a c t i v i t y in o t h e r t i s s u e s
only f i v e t o t e n p e r c e n t o f t h a t in l i v e r e x t r a c t s ( Stegeman and
Goldberg 1971).
The o c c u r re n c e o f both G6PD and H6PD in th e l i v e r s o f salmonids
has been c l e a r l y demo nstrated by v a r i o u s a u t h o r s ( Stegeman and
Goldberg 1971, Shatton e t a l . 1971s Ohno e t a l . 1966).
The e x a c t
number o f isozymes in th e m ulti -b an de d zymograms o f rainbow t r o u t
i s , however, n o t known.
Stegeman and Goldberg (1971, 1972) demon­
s t r a t e d t h a t in t h e genus S a lv e lin u s G6PD a ppea rs t o be t e t r a m e r i c
( e x h i b i t i n g f i v e bands) w it h H6PD e x i s t i n g as a d im e r ic mo le cu le.
The e x i s t e n c e o f two codominant gene l o c i w it h d i f f e r e n t a l l e l e s has
been p o s t u l a t e d t o be de te r m in in g t h e G6PD bands in t r o u t (Ohno .1966,
Yamauchi and Goldberg 1973, Stegeman and Goldberg 1971).
However,
Cederbaum and Yoshida (1975) r e c e n t l y proposed t h a t G6PD in rainbow
t r o u t l i v e r may be encoded by a s i n g l e gene l o c u s , two a l l e l e system.
62
with p o s t t r a n s l a t i o n a l m o d i f i c a t i o n o f t h e enzyme a c c o u n ti n g f o r th e
complex banding p a t t e r n s .
On t h e b a s i s o f a l l e l i c v a r i a t i o n s t u d i e s
by Stegeman and Goldberg (1971, 1972), H6PD i s b e l i e v e d t o be t h e
product o f a s i n g l e autosomal gene lo c u s .
E l e c t r o p h o r e t i c a n a l y s i s o f G6PD a c t i v i t y in l i v e r t i s s u e
e x t r a c t s o f T. a ro tio u s r e v e a l e d t h a t t h i s t i s s u e c o n t a i n e d f o u r
d i s t i n c t zones o f a c t i v i t y when glu co se 6-p hosp ha te was used as sub­
s t r a t e in s t a i n i n g (F ig u re s 11 and 13).
The band a p p e a ri n g second
from t h e c a th oda l end ( l i v e r samples) was i d e n t i f i e d as H6PD,
e x h i b i t i n g broad s u b s t r a t e s p e c i f i c i t y . i n d i c a t e d by s t a i n i n g with
g a l a c t o s e 6-ph os phat e and glu c ose with NAD o r NADP as t h e coenzyme
(Figure 11).
T h e r e f o r e , both forms o f G6PD a r e p r e s e n t in
t.
a ro tio u s as in o t h e r v e r t e b r a t e s .
Three zones o f G6PD a c t i v i t y , each r e p r e s e n t e d by a s i n g l e band,
i s th e common phenotype.
The middle zone o f G6PD a c t i v i t y i s g r e a t e r
in s t a i n i n g i n t e n s i t y than e i t h e r o f th e o t h e r two zones (F ig u re 11,
e r y t h r o c y t e s , and Figure 13).
This phen oty pic p a t t e r n i s s u g g e s t i v e
o f two l o c i with a l l e l e s encoding s u b u n i t s o f d i f f e r e n t e l e c t r o ­
p h o r e t i c m o b i l i t y f o r a dim e ri c molecule.
The two l o c i a r e d e s i g ­
nat ed G6PD-2 and G6PD-3 in o r d e r o f i n c r e a s i n g anodal m i g r a t i o n .
A
v e r i f i c a t i o n o f t h i s model o f i n h e r i t a n c e was o b t a i n e d by f i n d i n g
i n d i v i d u a l s in t h e Grebe and Wolf Lake p o p u l a t i o n s p o s s e s s i n g e l e c t r o ­
p h o r e t i c p a t t e r n s as shown in la ne two and f o u r o f Fi gure 12.
These.
63
+
-G6PD-3
H6PD
-G6PD-2
-G6PD-1
1
Figure 11.
2
3
4
5
6
7
G lucose-6-Phosphate Dehydrogenase (G6PD) and Hexose-6Phosphate Dehydrogenase (H6PD) ex pression in e ry th ro c y te s
and eye tis s u e . L iver tis s u e samples have th e same
banding p a tte r n as e ry th ro c y te s . Column I , 2, 3, 6 and
7 are eye samples from in d iv id u a l f is h . Column 4 and
5 e ry th ro c y te s from in d iv id u a l f is h .
64
1
Figure 12.
2
3
4
Glucose-6 -Phosphate Dehydrogenase-5 (G6PD-3) is p o ly ­
morphic in th e Yellowstone Park p o p u la tio n s. Column
I m ixture o f homogenates a p p lied to column 2 and 4.
Column 2 G6PD-3 (1 .0 0 /1 .0 0 ) homozygote. Column 3
G6PD-3 (1 .0 0 /1 .1 0 ) h etero zy g o te. Column 4 G6PD-3
(1 .1 0 /1 .1 0 ) homozygote. Liver samples.
65
1 2
Figure 13.
5
4
5
6
7
8
9
10 11 12
Hexose-6- Phosphate Dehydrogenase (H6PD) polymorphism.
Donnelly River samples have a s li g h t l y f a s t e r anodal
m ig ratin g form than the Grebe Lake (YNP) samples.
Columns I , 2, 3, 7, 8 and 9 (1 .1 0 /1 .1 0 ) homozygotes
from Donnelly R iver. Columns 4, 5, 6, 10, 11 and 12
(1 .0 0 /1 .0 0 ) homozygotes from Grebe Lake. L iver
homogenates.
66
p a t t e r n s i n d i c a t e t h e pre s e n c e o f a v a r i a n t a l l e l e a t t h e G6PD-3
lo c u s .
A homozygous i n d i v i d u a l f o r t h e v a r i a n t a l l e l e ( d e s ig n a te d
G 6 P D - 3 ^ '^ ) i s r e p r e s e n t e d by a s i n g l e band with a g r e a t e r e l e c t r o ­
p h o r e t i c m o b i l i t y than an i n d i v i d u a l homozygous f o r t h e common
a l l e l e (G6PD- 3^*^).
Heter ozy gote s ( 1 . 0 0 / 1 . 1 0 ) e x h i b i t a t h r e e
banded phenotype c h a r a c t e r i s t i c o f t h e d im e ric s t r u c t u r e w it h a band
i n t e r m e d i a t e between t h e two homozygote bands.
The v a r i o u s isozyme
p a t t e r n s a r e d e p i c t e d in Figure 12.
The pre s en c e o f t h i s polymorphism p r o v i d e s a d d i t i o n a l evidence
t o s u p p o r t t h e two locus p o s t u l a t e .
Since a h e te r o d im e r i s formed
o f s u b u n i t s from t h e two l o c i , a band o f g r e a t e r e l e c t r o p h o r e t i c
m o b i l i t y in t h e h e t e r o d i meric re g io n would be e x pe c te d in a homo­
zygous i n d i v i d u a l f o r t h e v a r i a n t a l l e l e a t th e G6PD-3 l o c u s .
Furt her mo re, an i n d i v i d u a l hetero zyg ous f o r th e v a r i a n t G6PD-3
a l l e l e would have two h e t e r o d i m e r i c ba nds, formed between s u b u n i t s ,
o f t h e G6PD-2 and t h e two G6PD-3 a l l e l e s .
t h i s t o be t h e c a s e .
Figure 12 c l e a r l y shows
These r e s u l t s a r e found whether NADP i s used
in t h e g r i n d i n g s o l u t i o n o r n o t , a l t h o u g h t h e pre s en c e o f NADP does
enhance r e s o l u t i o n .
The d a ta i s in agreement with t h e two codominant
l o c i p o s t u l a t e d by Ohno e t a t. (1966) f o r rainbow t r o u t and Stegeman
and Goldberg (1971) f o r brook t r o u t .
The r e s o l u t i o n o b t a i n e d in
t h e s e zymograms o f T,. a r c tie u s i s much more conv inc in g t h a n th o s e
67
used by Cederbaum and Yoshida (1975) in p o s t u l a t i n g t h e p resen ce o f
a sin g le locus.
A na ly sis o f eye t i s s u e and red blood c e l l s r e s u l t e d in th e
p a r a l l e l e x p r e s s i o n o f t h e two G6PD, l o c i with t h e h e t e r o d i meric
regi on a ga in e v i d e n t (F igu re 11).
The la ck o f H6PD a c t i v i t y in the
eye t i s s u e c l a r i f i e s i n t e r p r e t a t i o n o f l i v e r zymograms s i n c e one
can assume t h a t none o f t h e a c t i v i t y in t h i s re g io n i s due t o G6PD.
An a d d i t i o n a l band c l o s e t o t h e o r i g i n was e x p r e s s e d in eye
t i s s u e and red blood c e l l s , zymograms (F ig ur e 11).
No d i s t i n c t
h e t e r o d i m e r i c r e g i o n was formed between t h i s band and t h e s u b u n i t s
o f G6PD-2 o r G6PD-3.
The band may r e p r e s e n t a d i s s o c i a t i o n produ ct
o f t h e G6PD-2 but i t s c o n s i s t e n t pre s en c e in a l l samples in equal
i n t e n s i t y and absence in l i v e r s u g g e s t s an a d d i t i o n a l l o c u s , d e s i g ­
n a te d G6PD-1, which i s a c t i v e in t h e s e t i s s u e s but n o t in t h e l i v e r .
In summary, t h e G1SPD-I and G6PD-2 l o c i were monomorphic in a l l
populations.
The G6PD-3 loc us was monomorphic in t h e Donnelly and
Fuse p o p u l a t i o n s , but was polymorphic in t h e Grebe and Wolf Lake
populations.
A re g io n o f HSPD a c t i v i t y was e x p r e s s e d in a l l p o p u l a t i o n s
su rveyed.
As noted in Figure 13, HSPD a c t i v i t y i s r e p r e s e n t e d by
an a r e a o f d i f f u s e s t a i n i n g .
I t i s p o s t u l a t e d t h a t a s i n g l e locus
i s r e p r e s e n t e d by t h i s HSPD a c t i v i t y , a lth oug h i t a p p e a rs two bands
68
a r e p r e s e n t a t t im e s .
As shown in F ig ur e 13, t h e H6PD band in
Donnelly River and Fuse Lake , i n d i v i d u a l s has a c h a r a c t e r i s t i c migra­
t i o n d i s t a n c e g r e a t e r than t h e i n d i v i d u a l s from t h e Yellowstone
populations.
A m ix tu r e o f an i n d i v i d u a l sample from each pop u la ti o n
r e s u l t s in an a r e a s t a i n i n g which i s c l e a r l y a d d i t i v e o f t h e a r e a s
s t a i n e d in i n d i v i d u a l samples.
The r e s u l t s provid e ev id en c e t h a t
t h e H6PD loc us i s f i x e d f o r a l l e l e s which code f o r s u b u n i t s o f
d if f e r e n t e le c tro p h o re tic mobility.
The a l l e l e in t h e Grebe and
Wolf Lake p o p u l a t i o n s i s d e s i g n a t e d H6PD1 ‘00 and t h a t in th e Donnelly
I TO
R ive r and Fuse Lake p o p u l a t i o n s i s H6PD * .
No i n t r a p o p u l a t i o n a l
v a r i a t i o n was found t o e x i s t .
Malic enzyme (NADP - MDH) - Malic enzyme o t h e r w i s e known as
NADP dependent ma late dehydrogenase e x i s t s in mammalian t i s s u e in
two forms, s u p e r n a t a n t and m ito c h o n d ri a l (Henderson 1966, Shows e t d l
1970).
Evidence from e l e c t r o p h o r e t i c s t u d i e s o f t h e mouse su g g es t
a t e t r a m e r i c s t r u c t u r e f o r both t h e s u p e r n a t a n t (MEg ) and mitochon­
d r i a l (MEffl) form o f t h i s enzyme (Shows and Ruddle 1968, Baker and
Mintz 1969, Povey e t a l . 1975).
The m it o c h o n d ri a l form m i g r a t e s l e s s
a n o d a l l y than t h e s o l u b l e form in some s p e c i e s (Cohen and Omen 1972)
bu t may move a g r e a t e r d i s t a n c e in o t h e r s (Henderson 1966).
Two autosomal gene l o c i (MEg and MEm) determine t h e s o l u b l e and
m it o c h o n d ri a l f o r m s , r e s p e c t i v e l y (Cohen and Omen 1972, Povey e t a l .
1975).
69
The ME systems have n o t been as well s t u d i e d in t e l e o s t s .
The
la c k o f v a r i a t i o n in s p e c i e s s t u d i e d has n o t allowed d e t e r m i n a t i o n o f
t h e m o le c u la r s t r u c t u r e in t e l e o s t f i s h ( U t t e r e t a l . 1973, A l l e n d o r f
1973, K r i s t i a n s s o n and McIntyre 1973).
Frydenberg and Simonsen
(197 3) , on t h e b a s i s o f e l e c t r o p h o r e t i c p a t t e r n s , s u g g e s te d t h a t a
s i m p l e r molecule e x i s t e d in t h e t e l e o s t , Z paroes , than t h e t e t r a m e r
found in t h e house mouse (Shows and Ruddle 1968, Povey e t a l., 1975).
They c o n s i d e r e d t h e enzyme in Zoareee t o be c o n t r o l l e d by a s i n g l e
locus.
Allendorf e t a l.
(1975) have r e p o r t e d a d i f f e r e n t band o f
a c t i v i t y p r e s e n t in muscle than in l i v e r in rainbow t r o u t , bu t the
e x a c t number o f l o c i was no t det ermi ne d.
L i v e r and muscle t i s s u e e x t r a c t s were surveyed f o r ME a c t i v i t y
in T. a r e tie u s .
An i d e n t i c a l phen oty pic p a t t e r n (F ig ure 14) was
e x p re ss e d in both t i s s u e s .
A group o f f i v e bands which m ig r a te s
f u r t h e r than any MDH isozymes was i d e n t i f i e d as t h e s u p e r n a t a n t form:
o f ME (MEg).. A s lo w er d i f f u s e s t a i n i n g band with a m i g r a t i o n d i s ­
t a n c e i n t e r m e d i a t e between MDH5 and MDHm isozymes was i d e n t i f i e d as
t h e m it o c h o n d ri a l form o f ME (ME ).
m
The pre s en c e o f f i v e bands in e ver y i n d i v i d u a l i n d i c a t e s the
pr e s en c e o f two l o c i with a l l e l e s encoding s u b u n i t s o f d i f f e r e n t
e l e c t r o p h o r e t i c m o b i l i t y f o r a t e t r a m e r i c molecule.
were d e s i g n a t e d ME5-I and ME5 -Z.
The two lo c i
The i n t e n s i t y o f t h e i n d i v i d u a l
70
4-
Figure 14.
M alic enzyme (M.E.) . The su p ern atan t form of M.E.
appears to have a m ultim eric s tr u c tu r e perhaps produced
by lo c i which are d i f f e r e n t i a l l y a c tiv e . L iver homo­
g en ates. The m ito c h o rd ria l form appears to be monom orphic.
71
bands v a r i e d w it h t h e i n d i v i d u a l sample s u g g e s t i n g t h a t one o f th e
l o c i may be polymorphic w hi le t h e o t h e r i s monomorphic.
A rare
v a r i a n t was a l s o i d e n t i f i e d in one i n d i v i d u a l in t h e Grebe Lake
population.
Since v a r i a t i o n i s s u s p e c t e d but t h e number o f a l l e l e s
i s n o t known, t h e s e l o c i were excluded from th e q u a n t i t a t i v e c a l c u ­
lations.
However, t h e p resen ce o f two l o c i coding f o r a t e t r a m e r i c
molecule i s s u g g e s te d .
The s t r u c t u r e i s s i m i l a r t o t h a t r e p o r t e d
in o t h e r v e r t e b r a t e s ( d i s c u s s e d p r e v i o u s l y ) .
The ME5 locus appears
t o be d u p l i c a t e d in T. a ro tio u s .
The MEm band was e l e c t r o p h o r e t i c a l l y i d e n t i c a l in a l l p o p u l a t i o n s .
Since no v a r i a t i o n i s p r e s e n t , n o th in g can be deduced a bout t h e s t r u c ­
t u r e o f t h i s molecule.
One s t r u c t u r a l gene pro bably codes f o r t h i s
enzyme.
Serum p r o t e i n s - T e l e o s t s p e c i e s have been shown t o pos se ss
i n t r a s p e c i e s s p e c i f i c i t y o f plasma p r o t e i n p a t t e r n s (Woods and Engle
1957, Tsuyki and Roberts 1966).
Although f i s h serum p r o t e i n s a r e as
y e t i m p e r f e c t l y s t u d i e d from a g e n e t i c p o i n t o f view ( Kirpi ch nik ov
1973), t h e y have been used s u c c e s s f u l l y in work on i n t r a s p e c i f i c
s y s t e m a t i c s (Lukjanenko and Popov 1969, Wright and H a s l e r 1967,
R e i n i t z 1973, Booke 1964, DeLigny 1969).
In a n a l y z i n g serum p r o t e i n s
f o r g e n e t i c v a r i a t i o n , i t must be n o t e d t h a t serum p r o t e i n p a t t e r n s
r e s o l v e d e l e c t r o p h o r e t i c a l l y vary with p h y s i o l o g i c a l and enviro nme nta l
72
c o n d i t i o n s ( Booke 1964, Thurston 1967).
A p r i n c i p a l v a r i a t i o n noted
i s r e l a t e d t o sex and m a t u r i t y in females (reviewed by DeLigny
1969, Feeney and Brown 1974).
The n o n - g e n e t i c v a r i a t i o n o f serum
p r o t e i n s appea rs t o in v o lv e q u a n t i t a t i v e changes r a t h e r than th e
p resen ce o r absence o f bands ( Booke 1964, Thurston 1967, Feeney and
Brown 1974, DeLigny 1969).
As DeLigny (1969) s u g g e s t s , in any study
o f v a r i a t i o n in n o n - i d e n t i f i e d components o f serum p r o t e i n s , s u f f i ­
c i e n t p o p u l a t i o n d a t a and a n a l y s i s o f t h e composition o f th e samples
wit h r e g a r d t o s e x , m a t u r i t y , and development s t a g e a p p e a rs an
a b s o l u t e re qu ir e m e n t.
Furt her mo re, a good r e s o l u t i o n o f t h e p a t t e r n
which allow s a s har p d i s t i n c t i o n between i n d i v i d u a l f r a c t i o n s i s
needed in o r d e r t o avoid wrong i n t e r p r e t a t i o n o f th e observed
variation.
The plasma p r o t e i n s o f f i s h as y e t do not ap pea r t o have a c c u r a t e
classification.
No c o n c l u s i o n s as t o s i m i l a r i t y t o human plasma
f r a c t i o n s can be made (Feeney and Brown 1974).
Recent a t t e m p t s a t
c l a s s i f i c a t i o n o f t h e v a r i o u s f r a c t i o n s o f plasma p r o t e i n s o f v a ri o u s
salmonid s p e c i e s have been done ( P e r r i e r e t a t . 1973, ReichenbachKlinke 1973).
However, i t was beyond t h e scope o f t h e p r e s e n t study
t o a tt e m p t t o i d e n t i f y and b i o c h e m i c a l l y c h a r a c t e r i z e t h e v a ri ous
f r a c t i o n s o f serum p r o t e i n s observed on s t a r c h g e l s o f Thymallus
a v o tio u s .
73
Serum samples o f t h e f o u r p o p u l a t i o n s o f Thymallus a vo tio u s
under i n v e s t i g a t i o n were s u b j e c t e d t o e l e c t r o p h o r e s i s and s t a i n e d
wit h a g e ner al p r o t e i n s t a i n .
a r e shown in Fi g u r es 14 and 15.
El ectropherograms o f t h e serum p r o t e i n s
Samples were normally f r o z e n be fo re
e l e c t r o p h o r e s i s bu t s u bs e qu e nt s t u d i e s r e v e a l e d t h a t f r e e z i n g and
thawing had no e f f e c t on t h e e l e c t r o p h o r e t i c p a t t e r n s o f t h e serum
proteins.
El ect ropherograms o f serum samples r e v e a l e d 6 zones o f
s t a i n i n g as shown diagra mma tica l!. / in Fig ur e 16.
Zone 2 was i d e n t i ­
f i e d as t r a n s f e r r i n which has been d e a l t with p r e v i o u s l y .
o t h e r zone a d e q u a t e l y r e s o l v e d was Zone 5.
The only
The r e s o l u t i o n of t h i s
zone was e x c e l l e n t and t h e number o f bands could be det ermi ne d p r e ­
cisely.
I n d i v i d u a l s e x h i b i t e d from t h r e e t o f i v e bands in t h i s
region.
A t o t a l o f twelve d i f f e r e n t e l e c t r o p h o r e t i c phenotypes were
observed as d e p i c t e d in Fig ur e 17.
Further analysis revealed th a t
t h e zone could be f u r t h e r d i v i d e d i n t o two groups d e s i g n a t e d A (most
a n o d a l ) , B ( l e a s t a n o d a l ).
Group A was found t o be r e p r e s e n t e d by
two i n t e n s e s t a i n i n g bands in a l l i n d i v i d u a l s in both t h e Grebe Lake
and Wolf Lake p o p u l a t i o n s whose e x p r e s s i o n appeared in de pen de nt of
t h e B group (F ig ure 17).
I t was p o s t u l a t e d t h a t t h e s e bands r e p r e ­
s e n t e d two gene l o c i , SP-2 and SP-3, in o r d e r o f i n c r e a s i n g anodal
m i g r a t i o n , encoding p r o t e i n s o f d i f f e r e n t e l e c t r o p h o r e t i c m o b i l i t y .
I n d i v i d u a l s from t h e Donnelly R ive r p o p u l a t i o n , however,
e x h i b i t e d e i t h e r two o r t h r e e bands in t h i s r e g i o n .
One band o f
74
F igure 15.
Electropherogram s o f serum p r o te n s .
a.
Phenotypes o f the Yellowstone Park p o p u la tio n s. A ll
in d iv id u a ls are homozygous a t the SP-2 ( I .0 0 /1 .OOf
locus and th e SP-3 (1 .0 0 /1 .0 0 ) lo cu s. The in d iv id u a ls
are polymorphic a t th e SP-I locus: Column I - homo­
zygote (1 .1 0 /1 .1 0 ); column 2 and 4 - homozygote
(1 .0 0 /1 .0 0 ); column 3, 5 and 6 - heterozygotes
( 1 . 00 / 1 . 10 ) .
b.
Phenotypes o f Donnelly R iver in d iv id u a ls . A ll
in d iv id u a ls are homozygous (1.00/1.00) a t the SP-%
lo cu s. In d iv id u als are polymorphic a t th e SP-I and
SP-2 l o c i : Columns I and 3 - SP-I (1 .1 0 /1 .1 0 ), SP-2
(1 .1 0 /1 .2 0 ); column 2 - SP-I (1 .1 0 /1 .0 0 ), SP-2
(1 .1 0 /1 .1 0 ); column 5 - SP-I (1 .1 0 /1 .0 0 ), SP-2
(1 .1 0 /1 .2 0 ); column 4 - Yellowstone Park in d iv id u a l SP-I (1 .0 0 /1 .0 0 ), SP-2 (1 .0 0 /1 .0 0 ), SP-3 (1 .0 0 /1 .0 0 ).
75
4
-
I---- SP-3
— I i— SP-2
I - S P - I ( 1 . 10)
— '---- SP-I (1.00)
I
2
3
4
5
6
+
----- SP-2
r— SP-3
^ i— SP-2
F l- S P - 2
1---- SP-I
-----SP-I
1
2 3
4
5
(1.20
(1 .10)
(1.00)
(1.10)
(1.00)
76
Zone I
Zone 2
Zone 3
Zone 4
— Zone 5
: z= z
F igure 16.
Zone 6
C%3
C .Z 3
Group
era
era
I S
I
■■
S P -2 1 *20
era
S P -2 1 ' 10
«9
era
era
S P - I 1 *10
■
b
era
S P - I 1 - 00
n
c .z 3
ii
t t a
mm
U
C Z 3
I
Group B
S P -2 1 ' 00
n
n
U
—
Figure 17. The tw elv e o b serv ed phenotypic p a tte rn s of electropherogram s
of grayling serum p ro tein s in Zone 5. Colum ns 1 -9 , D onnelly
River; Colum ns 1 0 -1 2 , Y ellow stone Park p o p u la tio n s. The
g en o ty p es of th e in d iv id u als are: I - S P -I (1.0 0 /1 .0 0 ) , S P -2
( 1 .1 0 /1 .1 0 ) ; 2 - S P -K l . 0 0 / 1 . 1 0 ),S P -2 (1 .1 0 /1 .1 0 ) ; 3 -S P -l
( 1 .1 0 / 1 .1 0 ) , SP-2 (1.1 0 /1 .1 0 ) ; 4 - S P - I (1 .0 0 /1 .0 0 ), SP-2
(1 .2 0 /1 .2 0 ) ; 5- S P -I ( 1 .0 0 /1 .1 0 ) , SP-2 (1 .2 0 /1 .2 0 ) ; 6 -S P -l
( 1 .1 0 /1 .1 0 ) , S P - 2 ( 1 .2 0 /1 .20) ; 7 - S P -I (1.0 0 /1 .0 0 ) , SP-2
( 1 .1 0 /1 .2 0 ) ; 8 - S P -I (I. 0 0 /1 .1 0 ) , SP-2 (1 .1 0 /1 .2 0 ) ; 9 - S P -I
( 1 .1 0 /1 .1 0 ) , SP-2 (1.1 0 /1 .2 0 ) ; 10- S P -I (1.0 0 /1 .0 0 ) , SP-2
( 1 .0 0 /1 .0 0 ) ; 11- S P -I ( 1 .0 0 /1 .1 0 ) , SP-2 (1.0 0 /1 .0 0 ) ; 12S P -I ( l . 1 0 /1 .1 0 ) , SP-2 (1.0 0 /1 .0 0 ) . All in d iv id u a ls are homo­
zygous for th e common a lle le ( 1 .0 0 /1 .0 0 ) at the SP-3 lo c u s .
78
s t r o n g i n t e n s i t y was c o n s i s t e n t in a l l sam ples, and was e l e c t r o p h o r e t i c a l l y i d e n t i c a l t o t h e SP-3 band o f t h e Yellowstone popula ­
t i o n s (F igu re 17), and was d e s i g n a t e d as such.
Two o t h e r bands were
a l s o observed in t h i s r e g i o n , one which had a s l i g h t l y f a s t e r migra­
t i o n d i s t a n c e than t h e SP-3 band and one which had a s l i g h t l y slower
m i g r a t i o n d i s t a n c e , i n t e r m e d i a t e between t h e SP-3 and t h e SP-2 band
o f t h e Yellowstone i n d i v i d u a l s .
These bands a r e c o n s i d e r e d t o be
t h e e q u i v a l e n t o f t h e bands produced by th e SP-2 locus in th e Yellow­
st o n e p o p u l a t i o n s , s i n c e no band e q u i v a l e n t in m i g r a t i o n d i s t a n c e
t o th e SP-2 band o f t h e Yellowstone p o p u l a t i o n s i s found in the
Donnelly p o p u l a t i o n .
I n d i v i d u a l s e x h i b i t e d e i t h e r an i n t e n s e s t a i n i n g
f a s t band, an i n t e n s e s t a i n i n g slow band, o r had both bands p r e s e n t
in weaker i n t e n s i t y .
These r e s u l t s le a d t o the i n t e r p r e t a t i o n o f a
one l o c u s , two a l l e l e system encoding a monomeric p r o t e i n , with
he terozygous i n d i v i d u a l s e x p r e s s i n g two weaker s t a i n i n g ban ds, the
m o b i l i t i e s o f which a r e i d e n t i c a l t o t h e bands e x p re ss e d in both
ty pe s o f homozygous i n d i v i d u a l s .
The a l l e l e common t o t h e Yellow­
s to n e p o p u l a t i o n s was d e s i g n a t e d S P - 2 ^ ' 0 0 , th o s e in t h e Donnelly
River p o p u l a t i o n SP-2
migration.
I TD
and SP-2
I PD
in o r d e r o f i n c r e a s i n g anodal
On t h e b a s i s o f t h i s h y p o t h e s i s , a l l e l e f r e q u e n c i e s were
determined f o r t h e two a l l e l e s in t h e Donnelly River p o p u l a t i o n .
In t h e Group B r e g i o n , i n d i v i d u a l s i n a l l p o p u l a t i o n s , e xce pt
t h e Fuse Lake p o p u l a t i o n , e x h i b i t e d e i t h e r one o r two bands o f
79
a c t i v i t y (F igu re 17).
A s i n g l e l o c u s , two a l l e l e system encoding a
monomeric p r o t e i n was p o s t u l a t e d t o be r e s p o n s i b l e f o r t h i s p a t t e r n
o f v a r i a t i o n , with homozygous i n d i v i d u a l s e x h i b i t i n g a s i n g l e band
o f s t r o n g i n t e n s i t y and hetero zyg ous i n d i v i d u a l s having two bands
o f weaker i n t e n s i t y .
a l l e l e s termed SP-V
This lo cu s was d e s i g n a t e d SP-I w it h t h e two
and SP-V ’ ^
in o r d e r o f i n c r e a s i n g anodal
migration.
Evidence in s u p p o r t o f t h i s g e n e t i c i n t e r p r e t a t i o n l i e s in the
e x p r e s s i o n o f t h e twelve ex pec te d phen otypes.
Nine phenotypes would
be ex pec te d t o r e s u l t in th e Donnelly R ive r p o p u l a t i o n , and t h r e e in
t h e Wolf and Grebe Lake p o p u l a t i o n s , as i s indeed t h e c a s e .
Fuse Lake i n d i v i d u a l s e x p r e s s e d an i d e n t i c a l SP-3 band as t h a t
found in both t h e Yellowstone Park and Donnelly R ive r p o p u l a t i o n s .
The Fuse Lake p o p u l a t i o n was n o t polymorphic a t t h e SP-2 l o c u s , but
I 10
appeared f i x e d f o r t h e SP-2 ’
a l l e l e which was p r e s e n t in the
Donnelly p o p u l a t i o n s , e x h i b i t i n g a band e l e c t r o p h o r e t i c a l Iy i d e n t i c a l
t o t h a t observed in t h e Donnely p o p u l a t i o n .
At t h e SP-I lo cu s the
Fuse Lake p o p u l a t i o n was f i x e d f o r t h e I . TO a l l e l e w it h no a p p a r e n t
variation.
As p r e v i o u s l y n o t e d , any s t u d y o f v a r i a t i o n o f serum p r o t e i n
p a t t e r n s must have s u f f i c i e n t p o p u l a t i o n d a t a t o det ermi ne i f the
v a r i a t i o n has a b a s i s o t h e r than g e n e t i c .
In th e p r e s e n t c a s e .
80
r e c o r d s were kept as t o t h e s e x , age and r e p r o d u c t i v e c o n d i t i o n o f
t h e i n d i v i d u a l s sampled.
No c o r r e l a t i o n e x i s t e d between t h e s e
f a c t o r s and t h e p a t t e r n e x h i b i t e d by th e i n d i v i d u a l .
Since t h e v a r i a t i o n observed f o r t h e s e serum p r o t e i n s has not
been a d e q u a t e l y s t u d i e d in r e l a t e d s p e c i e s , t h e q u a n t i t a t i v e a n a l y s i s
o f t h e s e systems w i l l be d e a l t with s e p a r a t e l y .
Q u a n t i t a t i v e A na ly si s o f Ge netic V a r i a b i l i t y
A l l e l e f r e q u e n c i e s o f t h i r t y - f i v e enzyme and p r o t e i n l o c i a re
p r e s e n t e d in Table 3, f o r t h e f o u r p o p u l a t i o n s o f Thymallus a vctlo u s
su rveyed.
The t a b l e shows t h e number o f a l l e l e s d e t e c t e d by e l e c t r o ­
p h o r e s i s a t each lo c u s .
calculated.
The h e t e r o z y g o s i t y a t each lo c u s i s a l s o
O
Homozygosity a t a lo cu s ( j ) i s d e f i n e d as E x where
x i s t h e fr eq ue ncy o f t h e i - t h a l l e l e .
H e te r o z y g o s it y f o r the
p
lo cu s (h) i s d e f i n e d as I-E x
(Nei 1975).
The a l l e l e f r e q u e n c i e s
used a r e t h o s e o b t a i n e d d i r e c t l y from t h e observed d a t a .
The enzymes and p r o t e i n s surveyed in t h i s st ud y were div id e d
i n t o t h r e e groups.
cose metabolism.
Group I in c lu d e s t h o s e enzymes in v o lv e d in g l u ­
Group I I in c l u d e s non-g lu co se m e t a b o l i z i n g enzymes
and Group I I I in c l u d e s non-enzymatic p r o t e i n s .
The g e n e t i c b a s i s o f isozyme polymorphisms s c o r e d on t h e ge ls
were no t v e r i f i e d d i r e c t l y by progeny s t u d i e s .
The g e n e t i c i n t e r ­
p r e t a t i o n o f t h e obser ved v a r i a t i o n in simple Medelian terms i s
81
Table 3.
A l l e l e f r e q u e n c i e s and degree o f h e t e r o z y g o s i t y in 35 lo c i
examined in f o u r p o p u l a t i o n s o f Thymallus a r o tic u s . (h =
heterozygosity)
Locus
Alleles
Group I .
Grebe
Wolf
DonnelIy
Fuse
Glucose M et a bol iz in g Enzymes
AGPD-I
(h)
1.00
1.00
0.0
1.00
0 .0
1.00
0 .0
1.00
0 .0
AGPD-2
(h)
1.00
1.00
0.0
1.00
0 .0
1.00
0 .0
1.00
0 .0
AGPD-3
(h)
1.00
1.00
0 .0
1.00
0 .0
1.00
0 .0
1.00
0 .0
Hexokinase-I
(h)
1.00
1.00
0 .0
1.00
0 .0
1 .00
0 .0
1.00
0 .0
H6PD-1
1.00
1.25
1.00
————
0 .0
1.00
————
0 .0
1.00
0 .0
1.00
0 .0
1.00
1.10
1.20
0.98
————
0.02
0.04
0.98
————
0.02
0.04
0.97
0.03
————
0.06
1.00
————
————
0 .0
1.00
1.00
0 .0
1.00
0.0
1.00
0 .0
1.00
0 .0
IDH -2
<h)m
1.00
1.00
0 .0
1.00
0 .0
1.00
0 .0
1.00
0 .0
G6PD-1
(h)
1.00
1.00
0 .0
1.00
0 .0
1.00
0 .0
1.00
0 .0
G6PD-2
1.00
1.10
0.92
0.08
0.14
0.98
0.02
0.04
1.00
————
0 .0
1.00
————
0 .0
(h)
I D H -I
S
(h)
ID H -I
(h)m
(h)
82
Table 3.
(Continued)
Locus
Alleles
Grebe
Wolf
DonnelIy
Fuse
LDH-I
1.00
1.00
0 .0
1.00
0.0
1.00
0 .0
1.00
0 .0
1.00
1.00
0 .0
1.00
0 .0
1.00
0 .0
1.00
0 .0
1.00
1.00
0 .0
1.00
0 .0
1.00
0 .0
1.00
0.0
1.00
1.00
0 .0
1.00
0 .0
1.00
0 .0
1.00
0 .0
1.00
1.00
0 .0
1.00
0 .0
1.00
0 .0
1.00
0 .0
1.00
1.00
0 .0
1.00
0 .0
1.00
0 .0
1.00
0.0
1.00
1.00
0 .0
1.00
0 .0
1.00
0.0
1.00
0 .0
1.00
1.00
0 .0
1.00
0.0
1.00
0 .0
1.00
0.0
1.00
1.00
0 .0
1.00
0.0
1.00
0.0
1.00
0 .0
1.00
1.00
0 .0
1.00
0.0
1.00
0 .0
1.00
0.0
1.00
1.00
m. ^
— — — —
— — — —
0.0
1.00
0.0
(h)
LDH-2
(h)
LDH-3
(h)
LDH-4
(h)
LDH-5
(h)
MDH-I
(h)S
MDH -2
(h)S
MDH -I
(h)m
MEm-I
(hT
PGM-I
(h)
PGM-2
1.00
1.10
(h)
PGM-3
(h)
1.00
0 .0
0 .0
0.89
1.10
0.20
1.00
0.0
1.00
0.0
1.00
0 .0
mm m,
1.00
83
Table 3.
Locus
(Continued)
Alleles
Group I I .
Grebe
Wolf
DonnelIy
Fuse
Non-glucose M et a boliz in g Enzymes
ADH-I
(h)
1.00
1.00
0 .0
1.00
0 .0
1.00
0 .0
1.00
0 .0
XDH-I
(h)
1.00
1.00
0 .0
1.00
0 .0
1.00
0 .0
1.00
0 .0
SDH-I
(h)
1.00
1.00
0 .0
1.00
0.0
1.00
0 .0
1.00
0.0
GOT-I
(h)S
1.00
1 .00
0 .0
1.00
0.0
1.00
0 .0
1.00
0 .0
GOT -2
(h)s
1.00
1.00
0.0
1.00
0.0
1.00
0 .0
1.00
0 .0
GOT-I
(h)m
1.00
1.00
0 .0
1.00
0.0
1.00
0 .0
1.00
0 .0
GOT -2
(h)m
1.00
1.00
0 .0
1.00
0.0
1.00
0 .0
1.00
0 .0
EST-I
(h)
1.00
1.00
0 .0
1.00
0.0
1.00
0 .0
1.00
0 .0
TO-I
1.00
OTBTT
0.65
0.35
0.46
0.56
0.44
0.49
0.65
0.35
0.46
— — — —
(h)
1.00
0 .0
84
Table 3.
Locus
(Continued)
Alleles
Group I I I .
TFN-I
1.00
LW
1.20
(h)
SP-I
1.00
1.10
(h)
SP-2
1.00
1.10
1.20
(h)
SP-3
(h)
1.00
Grebe
Wolf
Donnelly
Fuse
Nonenzymatic P r o t e i n s
0.87
0.11
0.02
0.22
0.75
0.17
0.08
0.41
0.81
0.19
— — — —
— — — —
0.30
0 .0
0.60
0.40
0.48
0.51
0.49
0.50
0.65
0.35
0.45
1.00
0 .0
1.00
1.00
— — — —
— — — —
— — — —
— — — —
0 .0
1.00
0 .0
1.00
— — — —
0 .0
0.30
0.70
0.42
1.00
0 .0
1.00
0 .0
1.00
0 .0
1.00
0 .0
— — — —
85
s u p p o rt e d by agreement between t h e obser ved and ex pec te d p r o p o r t i o n s
o f genotypes in t h e p o p u l a t i o n s , assuming Hardy-Weinberg e q u i l i b r i u m .
Table 4 shows t h e co rre spondence o f observed genotype f r e q u e n c i e s t o
t h o s e ex pe c te d on t h e b a s i s o f Hardy-Weinberg e q u i l i b r i u m .
The c l a s s
s i z e s in s e v e r a l systems were small and r e q u i r e d grouping f o r adequate
a n a l y s i s (Sokal and Rohlf 1969).
The d i f f e r e n c e s observed from Hardy-
Weinberg e q u i l i b r i u m a r e not s i g n i f i c a n t .
The t e t r a z o l i urn oxida se
polymorphism in t h e Donnelly River p o p u l a t i o n e x h i b i t s a s l i g h t but
not s t a t i s t i c a l l y s i g n i f i c a n t deficiency of heterozygotes.
These
r e s u l t s and moreover t h e s i m i l a r i t y o f t h e p r o t e i n p a t t e r n s with
t h o s e e x h i b i t e d in o t h e r f i s h s p e c i e s whose g e n e t i c s have been
th or ou ghl y s t u d i e d , a p p ea r t o make t h e proposed g e n e t i c i n t e r p r e t a ­
t i o n s sound.
The serum p r o t e i n s o f Zone 5 on zymograms o f T. a r o tia u s a r e an
e x c e p ti o n t o t h e above c r i t e r i a as p r e v i o u s l y no te d.
The serum p r o ­
t e i n s o f f i s h a r e as y e t i m p e r f e c t l y s t u d i e d from a g e n e t i c s t a n d ­
p o i n t ( K irp ic hniko v 1975).
However, in t h e p r e s e n t s t u d y t h e r e s o l u ­
t i o n o f a number o f t h e s e p r o t e i n s e x h i b i t e d such c l a r i t y t h a t
p o s t u l a t e s as t o t h e i r g e n e t i c b a s i s a r e r e a s o n a b l e .
Si nce the
e l e c t r o p h o r e t i c p a t t e r n s allowed such i n t e r p r e t a t i o n s , th e y were
in c lu de d in a second s e t o f q u a n t i t a t i v e d a t a .
The a l l e l e f r e q u e n c i e s
a t t h e proposed l o c i a r e , t h e r e f o r e , in c l u d e d in Table 3.
86
Table 4.
Correspondence o f observed genotype f r e q u e n c i e s t o thos e
ex pec te d on th e b a s i s o f Hardy-Weinberg e q u i l i b r i u m f o r
t h e polymorphic l o c i o f Thymallus a r o r tc u s .a
Ch i-Squareb
P opu la tio n
TFN-I
Grebe
Wolfc
Donnelly
.2496
I
P = .62
TO-I
Grebe
Wolf
DonnelIy
.1124
.1115
3.4550
2
2
2
P = .95
P = .95
P = .18
SP-I
Grebe
Wolf
Donnelly
.1817
.0568
.5958
I
2
2
P = .67
P = .97
P = .75
SP-2
Donnelly
.0854
I
P = .77
PGM-2
DonnelIy
.0030
I
P = .96
G-6-PD-2
Grebe
Wolfc
.0003
I
P = .99
.0013
— — — —
----
--
(DF)
P robability of
G r e a t e r Chi-Square
Locus
I
-
-
P = .97
— — — —
—
a IDS-I not in c lu d e d s i n c e c l a s s s i z e s were too small f o r adequate
analysis.
^ C o n ti n u it y c o r r e c t i o n f a c t o r used s i n c e number o f c l a s s e s l e s s than
3.
c Locus n o t in cl uded f o r t h i s p o p u la ti o n s i n c e c l a s s s i z e s too small
f o r adequa te a n a l y s i s .
87
The g e n e t i c v a r i a t i o n o f a p o p u l a t i o n i s u s u a l l y measured by th e
p r o p o r t i o n o f polymorphic lo c i (P) and t h e average h e t e r o z y g o s i t y per.
locus (H).
These two para mete rs were used t o measure t h e i n t r a p o p u -
l a t i o n a l g e n e t i c v a r i a t i o n in t h e p r e s e n t s tu d y .
A locus was here
d e f i n e d as polymorphic in a p o p u l a t i o n i f t h e fr eq ue ncy o f t h e
commonest a l l e l e i s equal t o o r l e s s tha n 0.9 9.
Average h e t e r o ­
z y g o s i t y i s t h e mean o f t h e h e t e r o z y g o s i t y over a l l l o c i examined.
Average h e t e r o z y g o s i t y (H) was e s t i m a t e d usi ng the method o f Nei
(1975):
K " L=I
V
r
where gene f r e q u e n c i e s f o r r l o c i a r e s t u d i e d , and s u b s c r i p t L r e f e r s
t o t h e Lth l o c u s .
each p o p u l a t i o n .
Sampling v a r i a n c e o f H was a l s o c a l c u l a t e d f o r
Several o t h e r measures o f g e n e t i c v a r i a t i o n a re
l e s s s a t i s f a c t o r y f o r most purposes (Nei 1975).
These para mete rs f o r
t h e f o u r p o p u l a t i o n s s t u d i e d a r e summarized in Table 5.
Perc en t
polymorphic l o c i and average h e t e r o z y g o s i t i e s were c a l c u l a t e d separ a t e l y f o r Group I and Group I I enzymes.
In a d d i t i o n t h e s e pa rameters
were c a l c u l a t e d f o r Group I I and Group I I I enzymes and p r o t e i n s
t r e a t e d as a n . e n t i r e s e t .
F i n a l l y , t h e s e para mete rs were c a l c u l a t e d
f o r t h e t o t a l number o f l o c i surveyed in t h e s t u d y , one i n c l u d i n g
t h e g en eral serum p r o t e i n s and one e x c lu d i n g them.
Table 5.
Estimates o f g e n e t ic v a r i a b i l i t y in
Pop ulation
Number of
Individuals
Grebe
Group I
Group II
Group II & I I I a
Total I b
Total3
102
Wolf
Group I
Group II
Group. II & I I I a
Totalb
Total3
58
Donnelly
Group I
Group II
Group. II & I I I d
Totalb
Total3
63
Fuse
Group I
Group. II
Totalb
Total3
20
a In c lu d in g S P - I , SP-2, SP-3.
bExcluding S P - I , SP-2, SP-3.
T h y m a llu s a r c H o u s .
Number of
Loci
% Polymorphic
Loci
22
9
13
32
35
9.1
11.1
23.1
12.5
14.3
.0081
.0507
.0893
.0268
.0382
(±.0065)
(±.0507)
(±.0998)
(±.0161)
(±.0197)
22
9
13
32
35
9.1
11.1
23.1
12.5
14.3
.0039
.0547
.1069
.0303
.0418
(±.0024)
(±.0547)
(±.0566)
(±.0195)
(±.0223)
22
9
13
32
35
9.1
11.1
30.8
12.5
17.1
.0118
.0508
.1259
.0318
.0542
(±.0094)
(±.0508)
(±.0555)
(±.0177)
(±.0230)
22
13
32
35
0.0
0.0
0.0
0.0
.0000
.0000
.0000
.0000
Average
Heterozygosity
DISCUSSION
Genetic V a r i a b i l i t y o f Thymallus arotious
Es tim a te s o f g e n e t i c v a r i a t i o n in p o p u l a t i o n s a r e based on the
amount o f h e t e r o g e n e i t y d e t e c t e d in s t r u c t u r a l gene p r o d u c t s , mainly
p r o t e i n s and enzymes.
Gen etic v a r i a t i o n has been s t u d i e d in many
organisms a lt h o u g h t h e number o f l o c i i s n o t always l a r g e .
In the
p r e s e n t s t u d y , t h i r t y - f i v e enzyme and p r o t e i n l o c i in Thymallus
ccrctiaus were surveyed e l e c t r o p h o r e t i c a l I y .
The number o f l o c i i s
c o n s i d e r a b l y l a r g e in comparison with t h e number o f l o c i surveyed in
comparable s t u d i e s (reviewed by Powell 1975).
This h e t e r o z y g o s i t y
e s t i m a t e i s e x t r a p o l a t e d t o th e e n t i r e genome o f T. avcticu s.
One
must keep in mind t h e l i m i t a t i o n s o f e l e c t r o p h o r e t i c s urv ey s in
s tu d y i n g m o le c u la r v a r i a t i o n in p o p u l a t i o n s .
These l i m i t a t i o n s are
r e p e a t e d l y d i s c u s s e d in c u r r e n t l i t e r a t u r e on t h e s u b j e c t and need
n o t be d i s c u s s e d h e re .
The p e r c e n t polymorphic l o c i (common a l l e l e 0.99 o r l e s s ) and
aver age h e t e r o z y g o s i t y f o r o u t b r e e d i n g organisms a p p e a r s , u s u a l l y ,
t o be 25-50 p e r c e n t and 5-T5 p e r c e n t , r e s p e c t i v e l y (.Selander 1976,
Nei 1975).
studied.
These e s t i m a t e s vary c o n s i d e r a b l y with t h e organism
In g e n e r a l , v e r t e b r a t e s a p p e a r l e s s v a r i a b l e than i n v e r t e ­
b r a t e s , which may r e f l e c t th e s m a l l e r p o p u l a t i o n s i z e s o f v e r t e b r a t e s
(Nei 1975).
In v e r t e b r a t e s p e c i e s t h e p e r c e n t polymorphic l o c i
.
90
ranges from 20-35 p e r c e n t , and av erage h e t e r o z y g o s i t y 3-8 p e r c e n t .
These e s t i m a t e s a r e ta ken from th e aver age s o f t h e s e pa ra m et e rs f o r
v a ri o u s v e r t e b r a t e groups (S e la n d e r 1976, Nei 1975).
Es tim a te s f o r f i s h a r e h ig h l y het er o g en e o u s .
In Table 6 the
v a r i a b i l i t y e s t i m a t e s for. T. apcticus .are compared w it h th o s e o f .
other fis h species.
The v a lu es o b t a i n e d f o r g r a y l i n g a r e w it h in
t h e range o f th o s e found in o t h e r f i s h s p e c i e s (with o r w it h o u t
S P - I , SP-2, and S P -3 ), b u t a t t h e lower end o f t h i s range.
The
s t a n d a r d e r r o r s f o r t h e average h e t e r o z y g o s i t y e s t i m a t e s a r e l a r g e ,
which a p p ea r t o be common in e l e c t r o p h o r e t i c surveys (Nei 1975).
The comparisons o f v a r i a b i l i t y between T. a rc tie u s and o t h e r
salmonid s p e c i e s become more r e l i a b l e when th e general serum
p r o t e i n s a r e excluded s i n c e th e l o c i sampled a r e more homologous.
The importance o f comparing s i m i l a r l o c i i s e v i d e n t in Table 5,
s i n c e t h e h e t e r o g e n e i t y o f l o c i surveyed can g r e a t l y b i a s the
estim ates of v a r i a b i l i t y .
T h e r e f o r e , in any i n t e r s p e c i e s comparisons
t h e groups o f l o c i s t u d i e d should be s i m i l a r t o e l i m i n a t e p o t e n t i a l
bias.
The e s t i m a t e s o f p r o p o r t i o n o f polymorphic l o c i (1 2 .5 p e r c e n t )
and p r o p o r t i o n o f t h e genome he terozygous ( 2 . 7 - 3 . I p e r c e n t ) f o r
Thymallus arotiaus a r e in t h e mid-range o f e s t i m a t e s f o r a l l salmonid
s p e c i e s (Table 6 ) .
The v a r i a b i l i t y e s t i m a t e i s h i g h e r than t h a t
found in salmon s p e c i e s ( U t t e r e t a l. 1973, Altukhov e t a l. 1972),
bu t lower than t h a t found in rainbow t r o u t ( U t t e r e t a l. 1973).
Table 6.
Amount o f polymorphism and the degree o f h ete ro zy g o sity in some f i s h s p e c i e s .
Species
Number of
Loci
% Polymorphic
Loci
Average
H e ter ozy gosi ty
ThymallsUS arotiaus
32
12.5
2.9
Zoaraes viviparus
32
28-31
8.9
s u r f a c e dwelling
cave dw elling
17
17
29-41
0-29
11.2
3.6
Sebastes alutus
Sebastes aaurinus
Sebastes elongatus
25
25
24
8
4
8
3.8
1.8
3.2
Reference
Frydenberg and
Simonsen 1973
Astyanax mexioanus
P a c i f i c salmon
(v ariety of species)
19-23
Salmo gairdneri
19-23
8.7 -1 3
26
Avise and Selander
1972
Johnson e t a l. 1973
Il
Il
0.6-1.8
U t t e r e t a l. 1973
3.7
U t t e r e t a l. 1973
92
Within t h e s al m onida e, t h e h i g h e r g e n e t i c v a r i a b i l i t y found in
rainbow t r o u t may be a r e f l e c t i o n o f t h e h a b i t a t d i v e r s i t y o f t h i s
s p e c i e s when c o n t r a s t e d with t h e more s p e c i a l i z e d h a b i t a t s o f P a c i f i c
salmon ( U t t e r e t a l. 1973).
In l i g h t o f t h i s p o s s i b i l i t y , t h e lower
v a r i a b i l i t y e s t i m a t e s f o r g r a y l i n g may r e f l e c t t h e l e s s d i v e r s e
h a b i t a t o f t h i s s p e c i e s , as compared t o rainbow t r o u t .
This hypothe­
s i s i s su p p o rt e d by t h e r e s t r i c t i o n o f g r a y l i n g t o n o r t h e r n l a t i t u d e s ,
t h e i r f a i l u r e t o s u r v i v e both in modif ied h a b i t a t s and in t r a n s p l a n t s
i n t o new w a te r s (Vincent 1962).
This h y p o th e s i s i s s u g g e s t i v e o f a.
s e l e c t i v e advantage f o r t h e p r o t e i n polymorphisms ob s e r v e d .
However/
t h e r e s u l t s in no way r u l e out t h e p o s s i b i l i t y o f t h e p r o t e i n p o l y ­
morphisms being s e l e c t i v e l y n e u t r a l .
Other p o s s i b l e r e a s o n s f o r
t h e low amount o f g e n e t i c v a r i a b i l i t y in y. arotious may be s i m i l a r
t o th o s e s ugge s te d by Johnson e t a l. (1973) f o r Onoorhynchus ,.th a t
e i t h e r t h e s p e c i e s has evolved in an environment where s u r v i v a l
depends on a unique geno type , o r t h a t th e genome became f i x e d f o r
an optimal genotype and very l i t t l e change i s needed f o r s u r v i v a l
in a slowly changing environment.
The la c k o f v a r i a b i l i t y observed in t h e Fuse Lake p o p u l a t i o n i s
a p p a r e n t l y due t o a foun de r e f f e c t , which reduces v a r i a b i l i t y through
sampling e r r o r (Nei 1975).
The Grebe and.Wolf Lake p o p u l a t i o n s ,
with a s i m i l a r t r a n s p l a n t o r i g i n , may have e x p e r i e n c e d a s i m i l a r
93
" b o t t l e n e c k " in t h e i r e s t a b l i s h m e n t .
The v a r i a b i l i t y found in t h e s e
p o p u l a t i o n s i s n o t s i g n i f i c a n t l y d i f f e r e n t from t h e v a r i a b i l i t y
found in t h e Donnelly p o p u l a t i o n in t h e main range o f t h e s p e c i e s ,
suggesting t h a t t h i s i s not the case.
These p o p u l a t i o n s have
m a in ta in ed a l e v e l o f v a r i a b i l i t y common t o t h e s p e c i e s .
In s urv ey s e s t i m a t i n g average h e t e r o z y g o s i t y , i t becomes a p p a r e n t
t h a t h e t e r o z y g o s i t y v a r i e s c o n s i d e r a b l y among l o c i , w it h enzymes
having d i f f e r e n t l e v e l s o f g e n e t i c v a r i a t i o n (S e la n d e r 1975, Powell
1975, S e l a n d e r and Johnson 1973, Ayala 1974).
This high degree o f
i n t e r l o c u s v a r i a t i o n i s t h e o r e t i c a l l y ex pec te d i f each loc us under­
goes gene s u b s t i t u t i o n in d e p e n d e n tl y a t a low r a t e (Nei 1975).
The
r e l a t i v e deg rees o f v a r i a b i l i t y o f any p a r t i c u l a r enzyme te n d s t o
c u t a c r o s s taxonomic l i n e s , some p r o t e i n s a r e almost u n i v e r s a l l y
v a r i a b l e w hi le o t h e r s a r e r a r e l y polymorphic.
G i l l i s p i e and Kojima
(1968) a tt e m p te d t o acc oun t f o r t h i s degree o f v a r i a t i o n by specu­
l a t i n g t h a t t h e degree o f v a r i a t i o n in enzymes v a r i e s a c c o rd in g t o
function.
These a u t h o r s d i v i d e d enzymes i n t o two gro u p s , th o s e
invol ve d in glu co se metabolism (Group I ) and th o s e enzymes ho t
invol ve d in gluco se metabolism (Group I I ) .
A g r e a t deal o f d a ta
from d i v e r s e organisms a ppea rs t o s u p p o r t t h i s idea as reviewed by
Powell (1975).
Kojima e* a t .
(1970) s ubseq ue nt t o t h e above
h y p o t h e s i s , s u g g es te d t h a t t h e g r e a t e r v a r i a b i l i t y o f Group I I may
'I
94
be due t o t h e f a c t t h a t t h e s e enzymes a c t on a v a r i e t y o f s u b s t r a t e s
o f va ry in g c o n c e n t r a t i o n s , many o f which o r i g i n a t e e x t e r n a l t o the
organism.
The Group I enzymes, however, a c t on a s i n g l e s u b s t r a t e
whose c o n c e n t r a t i o n i s r e l a t i v e l y c o n s t a n t .
The c o n c l u s i o n being
t h a t enzymes in Group I I should be more g e n e t i c a l l y v a r i a b l e .
Johnson (1973) proposed a s i m i l a r h y p o th e s i s t o acco unt f o r v a r i a t i o n
in Drosophila.
The h y p o th e s i s o f G i l l i s p i e and Kojima (1968) i s
s up po rt e d by d a t a on gene d i v e r s i t y in some s p e c i e s ( Kojima e t a l.
1970, Ayala and Powell 1972, Cohen e t a l. 1973), bu t n o t in o t h e r s
(Nair e t a l. 1971, Frydenberg and Simonsen 1973).
In a d d i t i o n , primary s t r u c t u r e ( Zouros 1975) and t h e q u a t e r n a r y
s t r u c t u r e (Ward 1977) o f t h e p r o t e i n have been proposed as im por ta nt
f a c t o r s d e te r m in in g t h e e x t e n t o f polymorphism.
S e l a n d e r (1976)
pro vi de s an e x c e l l e n t review c once rn in g t h e s e proposed hypotheses
and t h e degree o f polymorphism.
The most pra gm at ic way t o examine t h i s problem, as Nei (1975)
s u g g e s t s , i s t o use a wide v a r i e t y o f organisms.
Since t h e p r e s e n t
s tu d y p r e s e n t s d a t a from Thymallus a ro tia u s, which may s e r v e to
he lp e l u c i d a t e t h e q u e s t i o n by adding d a t a t o t h a t a l r e a d y c o l l e c t e d
f o r o t h e r o rg a nis m s, t h e enzymes s t u d i e d were d i v i d e d i n t o two
groups as s u g g es te d by Kojima e t a l. (1970).
The enzymes surveyed
were c l a s s i f i e d as Group I (g lu c o s e m e t a t r o l i z i n g enzyme) and
95
Group I I (non g lu c os e m e t a b o l i z i n g ) as shown in Table 3.
Non-
enzymatic p r o t e i n s o f serum were r e p o r t e d s e p a r a t e l y and c l a s s i f i e d
as Group I I I .
The p e r c e n t polymorphic l o c i and av erage h e t e r o z y g o s i t i e s f o r
t h e Group I and Group IT enzymes o f T. apotious a r e summarized in
Table 5.
The p e r c e n t polymorphic l o c i f o r t h e Group I ( 9 . 1 ) and
Group I I ( 11 .1 ) do n o t ap pe a r t o be s i g n i f i c a n t l y d i f f e r e n t .
The
aver age h e t e r o z y g o s i t i e s f o r t h e Group I and Group I I enzymes appear
d i f f e r e n t , b u t , the. l a r g e s t a n d a r d e r r o r s s u g g e s t t h a t t h i s d i f f e r ­
ence i s n o t s i g n i f i c a n t .
Oh t h e b a s i s o f t h e d a ta o b t a i n e d , i t i s
concluded t h a t t h e r e i s no s i g n i f i c a n t d i f f e r e n c e in h e t e r o z y g o s i t y
between t h e Group I and Group I I enzymes in T. a rc tie u s.
The d a ta
do n o t s u p p o r t t h e s u g g e s t i o n t h a t l e v e l s o f v a r i a b i l i t y between.
Group I and Group I I a r e d i f f e r e n t .
In a few c as e s t h e enzyme c l a s s i f i c a t i o n in th e p r e s e n t stud y
may be c h a l l e n g e d : XDH i s g e n e r a l l y c l a s s i f i e d in Group I I s i n c e i t
has v a r i a b l e s u b s t r a t e s (Glassman 1965), bu t was c l a s s i f i e d in Group
I in Drosophila ( G i l l i s p i e and Langley 1974); TO i s c l a s s i f i e d as
Group I I a lt h o u g h i t has an unknown f u n c t i o n .
When a c c e p t a b l e
r e c l a s s i f i c a t i o n s a r e made, th e c o n c l u s i o n remains t h e same.
There
i s no ev id en c e t h a t t h e glu co se m e t a b o l i z i n g enzymes a r e l e s s v a r i a b l e
th a n o t h e r enzymes in Thymallus arotiau s.
-
96
An im p o r ta n t c o n s i d e r a t i o n in any e l e c t r o p h o r e t i c survey i s the
s e t o f l o c i used in e s t i m a t i n g aver age h e t e r o z y g o s i t y o f a p o p u la ti o n
Since t h e r e i s c o n s i d e r a b l e i n t e r l o c u s v a r i a t i o n , t h e s e t o f p r o t e i n s
examined may b i a s t h e r e s u l t s .
S a r i c h (1977) has shown t h a t t h e r e
a pp ear s t o e x i s t a r a p i d l y e v o l v i n g s e t o f p r o t e i n s (eg. e s t e r a s e s ,
t r a n s f e r r i n , plasma p r o t e i n s and many o f t h e c l a s s I I and I I enzymes)
( G i l l i s p i e and Kojima 1968, Kojima e t a t. 1970, Johnson 1974), and
a s e t o f s low ly e v o l v i n g l o c i (enzymes o f complex m e t a b o l i c pathways)
The d a t a o b ta in e d in t h e p r e s e n t s tu d y a ll ow a comparison o f such
s e t s t o be made.
The enzymes and p r o t e i n s o f Group I I and Group I I I
were c o n s id e r e d as one s e t w hile t h e Group I enzymes were co n si d e re d
as a n o t h e r ( th o s e inv olv ed in complex m e ta b o li c pathways) .
The
r e s u l t s a r e shown in Table 5 f o r t h e p o p u l a t i o n s o f T. arct-Lcus.
The
p e r c e n t polymorphic l o c i and av erage h e t e r o z y g o s i t y f o r t h e former
s e t , 23-31 p e r c e n t and 9-12 p e r c e n t , r e s p e c t i v e l y , a r e s i g n i f i c a n t l y
h i g h e r than f o r t h e Group I p r o t e i n s , 9 p e r c e n t and 0.4 -1 p e r c e n t ,
respectively.
The e x i s t e n c e o f a r a p i d l y e v ol vin g s e t o f p r o t e i n s
and a slow ly e v o l v i n g s e t i s su p p o rt e d by t h e r e s u l t s in T. a reticu s.
These r e s u l t s emphasize t h e importance o f su rv ey in g a wide v a r i e t y o f
l o c i in any e l e c t r o p h o r e t i c sur ve y.
I f one s e t o r t h e o t h e r was
c o n s i s t e n t l y used in e s t i m a t i n g h e t e r o z y g o s i t y , t h e r e s u l t s would
be b i a s e d .
97
Ge netic Divergence Between P o p u la ti o n s o f Thymallus arotiaus
P r o t e i n e l e c t r o p h o r e s i s allo w s q u a n t i f i c a t i o n o f t h e amount o f
g e n e t i c d i f f e r e n c e s between p o p u l a t i o n s based on a sample o f the
genome.
An i n h e r e n t b i a s o f e l e c t r o p h o r e s i s , however, i s t h a t only
s t r u c t u r a l genes which code f o r s o l u b l e p r o t e i n s a r e sampled.
Thus
r e g u l a t o r y genes and o t h e r s t r u c t u r a l genes a r e n o t sampled, which
means t h a t o n l y a minimal e s t i m a t e o f g e n e t i c d i f f e r e n t i a t i o n between
d i f f e r e n t ta x a can be made.
E l e c t r o p h o r e t i c d a t a c o n s i s t s o f a l l e l e and genotype f r e q u e n c i e s
determined from a sample o f a p o p u l a t i o n .
A common in de x which
summarized t h i s in f o r m a t io n i n t o a common meter o f g e n e t i c dive rge nce
between p o p u l a t i o n i s g e n e t i c d i s t a n c e .
Ge netic d i s t a n c e i s th e
g e n e t i c d i f f e r e n c e between p o p u l a t i o n s as e x p re ss e d by a f u n c t i o n
o f gene f r e q u e n c i e s .
The measure o f g e n e t i c d i s t a n c e used in the
p r e s e n t s tu d y was t h a t proposed by Nei (1971, 1972, 1973), by which
t h e av erage number o f codon d i f f e r e n c e s p e r locus can be e s t i m a t e d
from t h e gene fr equ en cy d a t a .
The method can be a p p l i e d t o any p a i r
o f ta x a whether th e y a r e l o c a l p o p u l a t i o n s , s p e c i e s , o r ge ner a (Nei
1975).
The s t a n d a r d g e n e t i c d i s t a n c e as proposed by Nei (1972) was
t h e measure used in t h e p r e s e n t s t u d y , where th e norm al iz ed g e n e t i c
i d e n t i t y o f genes between two p o p u l a t i o n s a t th e j lo c u s i s d e fi n e d as
I
zxy
/ ( s x 2y 2 )
98
where x, and
t i o n s x and y .
r e p r e s e n t t h e f r e q u e n c i e s o f th e i t h a l l e l e in popula­
For a l l l o c i i n a sample t h e g e n e t i c i d e n t i t y i s
defined as:
T
1
Jxy
=
/ (JxJy)
o
where J x , J y , and Jxy a r e t h e a r i t h m e t i c means o ve r a l l l o c i o f x ,
2
y , and x, y , r e s p e c t i v e l y .
D =
The g e n e t i c d i s t a n c e i s d e f i n e d a s :
Ioge I
which e s t i m a t e s t h e accumulated number o f codon d i f f e r e n c e s per
lo cu s s i n c e t h e tim e o f d iv e rg e nc e o f two p o p u l a t i o n s .
The g e n e t i c d i s t a n c e s among t h e v a r i o u s p o p u l a t i o n s o f Thymallus
avatious a r e summarized in Table 7.
Another index commonly employed i s Rogers simi l a r i Iy c o e f f i c i e n t
(S) (Rogers 1972).
Es ti m a te s o f I and S a r e c a l c u l a t e d from the
same d a t a and a r e f a i r l y s i m i l a r a lt h o u g h S give s lower numerical
v a lu es than I .
The v a l u e s o f S among t h e p o p u l a t i o n s o f Thymallus
aratiau s a r e a l s o summarized in Table 7.
The Rogers s i m i l a r i t y
c o e f f i c i e n t s were a l s o c a l c u l a t e d , so t h a t comparisons w ith valu es
o b t a i n e d in o t h e r s p e c i e s by a u t h o r s employing t h i s i n d e x , could
be made.
In a n a l y z i n g t h e s i g n i f i c a n c e o f t h e amount o f g e n e t i c dive rge nce
between p o p u l a t i o n s , t h e q u e s t i o n o f how much g e n e t i c d i f f e r e n t i a t i o n
oc cur s d urin g s p e c i a t i o n , which i s t h e most im p o r ta n t q u e s t i o n in
99
Table 7.
In d ic e s o f s i m i l a r i t y 3 (below d i a g o n a l ) and g e n e t i c
d i s t a n c e 13 (above d i a g o n a l ) f o r f o u r p o p u l a t i o n s o f
Thymallus a rcticu s.
Grebe
Total ( w ith ou t serum)
Wolf
DonnelIy
Grebe
.0000
.0007
.0335
.0368
Wolf
.9922
(.9993)
.0000
.0337
.0401
Donn.
.9597
(.9670)
.9587
(.9668)
.0000
.0054
Fuse
.9511
(.9638)
.9470
(.9606)
.9786
(.9946)
.0000
Grebe
Total (with serum)
Wolf
DonnelIy
Fuse
Fuse
Grebe
.0000
.0007
.0359
.0406
Wolf
.9917
(.9993)
.0000
.0359
.0405
Donn.
.9363
(.9647)
.9343
(.9647)
.0000
.0183
Fuse
.9097
(.9602)
.9071
(.9603)
.9420
(.9818)
.0000
3Rogerl S C o e f f i c i e n t o f Genetic S i m i l a r i t y ( N e i ' s C o e f f i c i e n t
of Sim ilarity)
bN e i l S Measure o f Sta ndard Genetic D is tan ce
100
e v o l u t i o n a r y g e n e t i c s , must be a d d r e s s e d .
The most common mode o f
s p e c i a t i o n i s ge ogr ap hic i s o l a t i o n , which i s t h e usual p r e r e q u i s i t e
t o g e n e t i c d i v e rg e n c e and hence, s p e c i a t i o n .
Two s t a g e s may be
rec og ni ze d in t h e p ro c e ss o f geog raphic s p e c i a t i o n (Ayala e t d l.
1974): I ) p o p u l a t i o n s become i s o l a t e d by geog raphic b a r r i e r s and
accumulate g e n e t i c d i f f e r e n c e s , 2) r e p r o d u c t i v e i s o l a t i n g mechanisms
a r e developed.
The second s t a g e be gin s when g e n e t i c a l l y d i f f e r ­
e n t i a t e d p o p u l a t i o n s r e g a i n geog raphic c o n t a c t .
A s t r a t e g y employed
t o a t t e m p t t o de te rm in e t h e p r o p o r t i o n o f gene l o c i a l t e r e d d u ri n g
t h e s p e c i a t i o n p ro c e s s in v o lv e s a s s a y i n g p o p u l a t i o n s which appear
t o be in v a r i o u s s t a g e s o f t h e s p e c i a t i o n p ro c e ss (Avise 1976).
Ayala e£ d l. (1974) have done t h e most e x t e n s i v e s t u d y o f
g e n e t i c d i f f e r e n t i a t i o n d u ri n g geog raphic s p e c i a t i o n u s in g populat i o n s o f Drosophila w illis to n i.
)
E s ti m a te s were made o f l e v e l s o f
g e n e t i c d i f f e r e n t i a t i o n between p o p u l a t i o n s a t f i v e l e v e l s , of
e v o l u t i o n a r y d i v e r g e n c e : I ) ge ogra phic p o p u l a t i o n s w i t h i n a taxon
(I = 0.970 ± . 0 0 6 ) , 2) s u b s p e c i e s , in t h e f i r s t s t a g e o f geographic
s p e c i a t i o n (I = 0.795 ± 0 . 0 1 3 ) , 3). semi s p e c i e s , in t h e second s t a g e
o f s p e c i a t i o n (I = 0.798 ± 0 . 0 2 6 ) , 4) s i b l i n g s p e c i e s (I = 0.517 +
0 . 0 2 4 ) , 5) n o n - s i b l i n g s p e c i e s (I = 0 . 3 5 2 ) .
Many s t u d i e s e x i s t on g e n e t i c d i f f e r e n t i a t i o n between v e r t e b r a t e
p o p u l a t i o n s in e a r l y s t a g e s o f e v o l u t i o n a r y d iv e rg e nce (reviewed by
101
Ayala 1975).
The r e s u l t s o f such s t u d i e s in s e v e r a l s p e c i e s o f
f i s h a r e summarized in Table 8.
The degre e o f g e n e t i c d i f f e r e n t i a t i o n
between s u b s p e c ie s in f i s h s p e c i e s i s comparable t o t h a t in Dvosoyhila
W illis to n is r e p r e s e n t i n g an e i g h t f o l d i n c r e a s e over t h e aver age d i s ­
ta n c e between l o c a l p o p u l a t i o n s .
However, general c o n c l u s i o n s about
g e n e t i c d i f f e r e n t i a t i o n d u ri n g s p e c i a t i o n a r e n o t w a r r a n t e d due to
t h e tremendous h e t e r o g e n e i t y o f t h e b i o l o g i c a l world (Avise 1976).
Mayr (1963) s t r e s s e d t h e p o i n t t h a t s p e c i e s a r e n o t c h a r a c t e r i z e d by
a given number o f counted gene d i f f e r e n c e s .
I t is highly unlikely
t h a t a l l s p e c i a t i o n e v e n t s w i l l i n v o lv e t h e same amount o f g e n e t i c
change.
Even when new s p e c i e s a r i s e a c c o rd in g t o t h e g en eral model
o f geog raphic s p e c i a t i o n , t h e amount o f g e n e t i c change in volve d may
vary from one c a s e t o a n o t h e r .
A ca se in p o i n t a r e t h e e s t i m a t e s o f
g e n e t i c d iv e r g e n c e in nin e genera o f C a l i f o r n i a minnows (Avise and
Ayala 1976, Avise e t q l. 1975).
L i t t l e genetic d i f f e r e n tia tio n
e x i s t e d between l o c a l p o p u l a t i o n s (I = 0 . 9 9 ) .
The av er ag e g e n e t i c
i d e n t i t y between nine s p e c i e s i s I = 0.592 ± 0.023.
However, the
most g e n e t i c a l l y s i m i l a r s p e c i e s a r e Hesyevoleuous syrm etvicus and
■Latina exilioavda with I = 0.946 and D = 0.055.
The two s p e c i e s
have v i r t u a l l y i d e n t i c a l g e n e t i c c o n s t i t u t i o n s a t 23 o u t o f 24 l o c i ,
but a r e h i g h l y d i f f e r e n t i a t e d a t a s i n g l e locus which i s indeed
s p e c i e s d i a g n o s t i c f o r most p o p u l a t i o n s (Ayala 1975).
U t t e r e t at . -
Table 8.
Genetic s i m i l a r i t i e s between popu la ti o n s a t d i f f e r e n t s t a g e s o f e v o l u t i o n a r y
diverge nce in s e v e r a l groups o f f i s h e s .
Local
Po pul a tio ns
Subspecies
Species
Genera
Lepomis
0.97*
0.85*
0.54 ± .016
- - —-
Avise and
Smith 1974
Cyprinodon
-----
-- - —
0.89 ± .016*
- ———
Turner 1974
Sc ia en id ae
---- -
—- ——
——- —
0.17 ± .027*
C alifornia
minnows
0.99
—-- -
0.59 ± .029
---- -
Avise and
Ayala 1975
Salmonidae
————
-----
0.46 ± .032*
-----
Utter e t al.
1973
Group
^ S i m i l a r i t i e s c a l c u l a t e d usi ng methods o t h e r than N e i 's (1972).
Source
Shaw 1970
103
(1973) have s t u d i e d allozyme d i f f e r e n t i a t i o n in two s p e c i e s o f t r o u t
{Sabno) and s i x s p e c i e s o f salmon [Oncorhynohus).
The average
s i m i l a r i t y (in de x s i m i l a r t o methods o f Nei and Rogers) between the
s i x salmon s p e c i e s i s 0.422 ± 0 . 0 4 ; t h a t between t h e two t r o u t s p e c i e s
i s 0. 9 0 .
The average g e n e t i c s i m i l a r i t y between a l l e i g h t s p e c ie s
i s 0.456 + 0.032.
In g e n e r a l , l i t t l e g e n e t i c d i f f e r e n t i a t i o n e x i s t s between lo c al
populations within a species.
Subspecies, rep resen tin g populations
in t h e f i r s t s t a g e o f s p e c i a t i o n , show moderate bu t s u b s t a n t i a l
degre es o f g e n e t i c d i f f e r e n t i a t i o n .
These o b s e r v a t i o n s a r e p e r t i n e n t
t o t h e p r e s e n t s t u d y s i n c e l o c a l p o p u l a t i o n s and g e o g r a p h i c a l l y
i s o l a t e d p o p u l a t i o n s o f T. ca-ctious were surveyed f o r t h e amount o f
genetic d if f e r e n tia tio n .
I t i s assumed t h a t s i n c e t h e Canadian
p o p u l a t i o n s and Montana p o p u l a t i o n s have been i s o l a t e d f o r more than
7,000 y e a r s (Vincent 1962), t h a t th e y r e p r e s e n t p o p u l a t i o n s in th e
f i r s t stage of sp ec ia tio n .
To deter mine what le vel , o f e v o l u t i o n a r y
d iv e rg e nc e t h e s e two forms a c t u a l l y r e p r e s e n t , t h e b e s t i n d i c a t i o n
o f d i v e rg e n c e would be t h e magnitude o f t h e d i f f e r e n t i a t i o n between
l o c a l p o p u l a t i o n s and t h e g e o g r a p h i c a l l y i s o l a t e d p o p u l a t i o n s .
Two
f a c t o r s which s u p p o r t t h i s l i n e o f r e a s o n i n g a re I ) t h e a p p a r e n t
h e t e r o g e n e i t y o f t h e degree o f g e n e t i c d i f f e r e n t i a t i o n invol ve d in
t h e s p e c i a t i o n p ro c e s s in d i v e r s e organisms ( p r e v i o u s l y d i s c u s s e d )
;v
;•
.
704
and 2) no comparable e s t i m a t e s o f t h e amount o f g e n e t i c d i f f e r e n t i a ­
t i o n between l o c a l p o p u l a t i o n s o r s u b s p e c i e s in salmonid s p e c i e s
are presently a v ailab le .
The Grebe and Wolf Lakes p o p u l a t i o n s r e p r e s e n t l o c a l p o p u l a t i o n s
o f T. apoticus.
The r e s u l t s (Table 7) i n d i c a t e t h a t l i t t l e g e n e t i c
dive rg e nce has o c c u r re d between t h e s e p o p u l a t i o n s (I = .99 and D =
.000 7).
The p o p u l a t i o n s a r e , however, r e p r o d u c t i v e l y i s o l a t e d by
means o f an e t h o l o g i c a l b a r r i e r .
The p o p u l a t i o n s s t u d i e d a t Grebe
Lake i s i n l e t spawning w hi le t h e Wolf Lake p o p u la ti o n
o u t l e t spawning ad ap te d.
studied is
A proposed g e n e t i c b a s i s f o r such be hav io ra l
c h a r a c t e r i s t i c s has been d i s c u s s e d p r e v i o u s l y .
A p p a r e n t ly , r e p r o ­
d u c t i v e i s o l a t i o n through e t h o l o g i c a l mechanisms does n o t r e q u i r e
changes in a s u b s t a n t i a l p r o p o r t i o n o f t h e genome, as s u g g es te d in
th e case o f Drosophila paulistorium (Ayala 1975).
An a l t e r n a t i v e
e x p l a n a t i o n as o f f e r e d by Ayala (1975) may be t h a t t h e completion
o f such r e p r o d u c t i v e i s o l a t i o n may r e q u i r e g e n e t i c c h an ge s , b u t not
in t h e c l a s s o f genes s t u d i e d by e l e c t r o p h o r e t i c t e c h n i q u e s .
The
changes r e q u i r e d may i n v o lv e o t h e r t y p e s o f s t r u c t u r a l o r r e g u l a t o r y
genes.
Wilson e t a t. (1974 a , b ) have s u g g es te d t h a t t h e r e may be two
ty p e s o f m o le c u la r e v o l u t i o n , one i n v o l v i n g s t r u c t u r a l g e n e s , which
goes on a t a more o r l e s s c o n s t a n t r a t e , and a second f o r r e g u l a t o r y
105
ge n e s , which a r e p r i m a r i l y r e s p o n s i b l e f o r r e p r o d u c t i v e i n c o m p a ta b il i
t i e s and morphological e v o l u t i o n .
They f u r t h e r p o i n t o u t t h a t e v o l u ­
t i o n a r y d iv e rg e nce as measured by p r o t e i n d i f f e r e n t i a t i o n on one
s i d e and by morphological div e rg e nce and r e p r o d u c t i v e i n c o m p a t i b i l i t y
on t h e o t h e r do not always go hand in hand.
A g r e a t deal o f morpho­
l o g i c a l d iv e rg e nce and r e p r o d u c t i v e i n c o m p a t i b i l i t y w it h only
moderate p r o t e i n d i f f e r e n t i a t i o n i s observed among mammals, while
t h e r e v e r s e i s observed in some amphibians and b i r d s .
The s u g g e s ti o n o f changes in r e g u l a t o r y genes c o n t r o l l i n g the
r e p r o d u c t i v e b e h a v i o r o f T. arotiaus i s a p l a u s i b l e e x p l a n a t i o n .
This may he lp t o e x p l a i n why t h e r e i s l i t t l e g e n e t i c d i f f e r e n t i a t i o n
a t th e s t r u c t u r a l gene l e v e l d e s p i t e t h e proba ble e t h o l o g i c a l
i s o l a t i n g mechanism p r e s e n t in t h e Grebe and Wolf Lakes p o p u l a t i o n s .
Although a t p r e s e n t t h e r e a r e no good e s t i m a t e s o f t h e p r o p o r­
t i o n o f t h e genome encoding s o l u b l e gene pro duc ts d e t e c t e d by
e l e c t r o p h o r e s i s , p r o t e i n d i f f e r e n c e s w i t h i n p a r t i c u l a r animal, groups
correspo nd c l o s e l y t o l e v e l s o f morphological and o t h e r dive rge nce
d e s c r i b e d by c l a s s i c a l s y s t e m a t i s t s (Avise 1974).
To t h i s e x t e n t
(
s t r u c t u r a l genes prov ide i n fo r m a t io n which i s o f e v o l u t i o n a r y
significance.
The Fuse Lake and Donnelly River p o p u l a t i o n s were a l s o c onsi de re d
l o c a l p o p u l a t i o n s s i n c e t h e i r o r i g i n s were both in t h e same geograph­
i c a l range.
The g e n e t i c div e rg e nc e (0.032) between t h e s e p o p u la ti o n s
106
i s c o n s i d e r a b l y l a r g e r than between t h e lo c a l Montana p o p u l a t i o n s
(. 0 0 0 7 ) .
I t i s b e l i e v e d t h a t t h i s e s t i m a t e o f t h e div e rg e n c e i s no t
e n t i r e l y r e p r e s e n t a t i v e o f t h e d i s t a n c e between l o c a l p o p u l a t i o n s o f
t h e Canadian form due t o t h e " b o t t l e n e c k " e f f e c t on t h e Fuse Lake
population.
IDH
The p o s s i b l e l o s s o f a l l e l e s a t some l o c i (PGM-2^
S P - 2 ^ * ^ ) in t h e Fuse Lake p o p u l a t i o n , which a r e . p r e s e n t
a t r e l a t i v e l y high f r e q u e n c i e s in t h e Donnelly River p o p u l a t i o n ,
r e s u l t s in t h e Fuse Lake p o p u l a t i o n bei ng f i x e d f o r t h e a l l e l e s
common t o both forms.
This r e s u l t s in a g r e a t e r g e n e t i c d i s t a n c e
between t h e Canadian p o p u l a t i o n s than might normally be found.
This
id ea i s s uppo rt e d by t h e o b s e r v a t i o n t h a t t h e Fuse Lake p o p u la ti o n
c o n t a i n s a l l e l e s common t o t h e Donnelly p o p u la ti o n a t t h e H6PD and
SP-2 l o c i .
While t h e r e i s complete s e p a r a t i o n o f t h e Montana and
A rctic populations a t these lo c i.
The g e n e t i c d i s t a n c e between t h e g e o g r a p h i c a l l y i s o l a t e d popula­
t i o n s o f t h e Montana and Canadian forms (Table 7) i s s u b s t a n t i a l in
comparison t o t h e d iv e rg e nce between l o c a l p o p u l a t i o n s .
The g e n e t i c
d i s t a n c e between t h e g e o g r a p h i c a l l y i s o l a t e d p o p u l a t i o n s ( .0 5 6 - .0 7 7 )
and t h e l o c a l p o p u l a t i o n s (. 001) r e p r e s e n t s a magnitude g r e a t e r than
t h a t between l o c a l p o p u l a t i o n s and s u b s p e c i e s in o t h e r organisms
(discussed p rev io u sly ).
A s u b s t a n t i a l degre e o f g e n e t i c d i f f e r e n t i a ­
t i o n a t t h e s t r u c t u r a l gene l e v e l has accumulated between t h e Montana
107
and A r c t i c forms o f T. arctio u s s i n c e t h e i r s e p a r a t i o n from a common
ancestor.
The two forms ap pe a r t o be well advanced i n t o t h e f i r s t
s t a g e o f ge og ra phi c s p e c i a t i o n , r e p r e s e n t a t i v e o f t h e s u b s p e c ie s
level.
The g e n e t i c s i m i l a r i t y between t h e two forms ( 0 . 9 1 - 0 . 9 4 ) i s
n o t much g r e a t e r than t h e s i m i l a r i t y ( 0.9 0) between two salmonids
a t t h e s p e c i e s l e v e l , Sabm gaivdnevi
Sdlmo c la r k ii ( U t t e r e t a t.
1973).
Taxonomic C o n s i d e r a t i o n s
Ph y lo g e n e ti c r e l a t i o n s h i p s can be i n f e r r e d t o some e x t e n t by
s tu d y in g morphological a f f i n i t y , however, t h e morphological a f f i n i t y
o f ta x a does n o t n e c e s s a r i l y r e p r e s e n t t h e r e a l phylogeny (Nei 1975).
Sokal and Sneath (1963) s t r e s s e d t h e s e p a r a t i o n o f t h e p h e n e t i c
( s i m i l a r i t y ) and p h y l e t i c (phylogeny) r e l a t i o n s h i p s .
,
Numerical
taxonomy a p p l i e d t o morphological c h a r a c t e r s give s only t h e p h e n e t i c .
r e l a t i o n o f ta x a (Nei 1975).
Genetic d i s t a n c e may be used t o stu dy
t h e phylogeny o f a group o f ta x a prod uc ing a more r e l i a b l e and
q u a n tita tiv e phylogenetic t r e e .
This method allows d i f f e r e n t i a t i o n
t o be s t u d i e d a t t h e codon l e v e l .
The p r o b a b i l i t y o f back muta tio ns
o r p a r a l l e l m u ta ti o n s a t a codon i s n e g l i g i b l y small u n l e s s e v o l u t i o n
a ry time i s very l a r g e (Nei 1975).
This method, t h e r e f o r e , has a
g r e a t advantage o ve r t h e method o f co mparative morphology, in which
108
d iv e rg e nce and convergence in morphological changes may make th e
r e s u l t u n c e r t a i n (Sokal and Sneath 1963).
A dendrogram o f t h e f o u r populations, o f Thymallus a rc tic u s s u r ­
veyed in t h e p r e s e n t s t u d y i s p r e s e n t e d in Figure 18.
The t r e e i s
c o n s t r u c t e d from t h e g e n e t i c d i s t a n c e s e s t i m a t e d in t h e p r e s e n t
study.
The p o p u l a t i o n s were grouped using t h e unweighted p a i r - g r o u p
method o f c l u s t e r i n g o f Sokal and Sneath (1963).
The g e n e t i c d i s t a n c e s e s t i m a t e d in t h e p r e s e n t s t u d y allows th e
p h y l o g e n e t i c p o s i t i o n o f the''two forms o f Thymallus arotiaus t o be
estimated.
At t h e p r e s e n t time no s u b s p e c i e s a r e re c o g n iz e d w i t h i n
T. a ra ticu s (McPhail and Lindsey 1970).
The g e n e t i c d i s t a n c e found
in t h e p r e s e n t s tu d y s u g g e s t s t h a t such a d i s t i n c t i o n may be w arrante d
The Montana form o f T. arotiaus appea rs g e n e t i c a l l y d i s t i n c t from t h e
A r c t i c form d e s e r v i n g s u b s p e c i f i c . s t a t u s .
These f i n d i n g s appear
cong rue nt with t h o s e p r e v i o u s l y proposed by v a r io u s a u t h o r s on th e
b a s i s o f morphological c h a r a c t e r i s t i c s (s e e i n t r o d u c t i o n ) .
I t is
proposed t h a t t h e Montana form o f r . arotiaus be. re c o g n iz e d as the
s u b s p e c ie s montccnus (nomenclature o f M iln e r 1874).
F u r t h e r , th e
A r c t i c form shou ld be d e s i g n a t e d a s . t h e s u b s p e c ie s s ig n ife r (nomen-,
c l a t u r e o f Richardson 1823).
Grebe
L-
-
Wolf
Donnelly
Fuse
I_ _ _ _ _ _ I_ _ _ _ _ _ I_ _ _ _ _ _ I_ _ _ _ _ _ I_ _ _ _ _ _ I_ _ _ _ _ _ I_ _ _ _ _ _ I_ _ _ _ _ _ I
.040
.035
.030
.025
.020
.015
.010
.005
0
Genetic Distance (D)
Figure 18.
Dendrogram f o r f o u r p o p u la ti o n s o f T. aro ticu s.
r e p r e s e n t e d on th e h o r i z o n t a l a x i s .
Genetic d i s t a n c e i s
APPENDIX
B uff e r Systems
B uff e r System
PH
Power
(V)
Time
(h r )
250
3
S e la n d e r e t a t.
1971
170
3
S e l a n d e r e t d l.
1971
100
4
S e la n d e r e t a l.
1971
150
5
May e t a l. 1975
120
5
S e la n d e r e t a l.
1971
Reference
A. Poulik
Electrode:
Gel:
0 .3 M Borate
8.2
0.076 M T r i s
0.005 M C i t r a t e
8 .7
B. Continuous T r i s C i t r a t e I
Electrode:
Gel:
0.223 M T r i s
0.086 M C i t r a t e
6 .3
0.008 M T r i s
0.003 M C i t r a t e
6 .7
C. Continuous T r i s C i t r a t e II
0.687 M T r i s
0.157 M C i t r a t e
8 .0
G e l : 22.890 M T r i s
5.220 M C i t r a t e
8 .0
Electrode:
D. Sodium Phosphate
Electrode:
Gel:
0.04 M NaH,PO.
0.06 M NaH^POJ
8 .3
D i l u t e Elec 10:1
8.3
E. Potassium Phosphate
Electrode:
Gel:
0.138 M KH9PO.
0.062 M NaOH 4
6.7
D i l u t e Elec 19:1
6 .7
112
B uff e r System
pH
Power
(V)
Time
(hr )
200
4
S e la n d e r e t a l.
1971
250
3
Markert and
Faulh ab er 1965
250
3
Ridgeway e t a l.
1970
350
3
Se la n d e r e t a l .
1971
Reference
F. I r i s - B o r a t e EDTA
Electrode:
Gel:
0 .5 M I r i s
0.65 M Borate
0.02 M EDTA
8 .0
D i l u t e Elec 9:1
8 .0
G. I r i s - B o r a t e EDTA
Stock:
Gel:
Electrode:
0 .9 M T r i s
0 .5 M Borate
0.02 M EDTA
8 .6
D i l u t e 20:1
D i l u t e 4:1
H. Ridgeway Bu ffe r
Electrode:
Gel:
.06 M LiOH
0 .3 M Borate
.03 M T r i s
.005 M C i t r a t e
8 .3
8 .0
I. Lithium Hydroxide
Stock S o l . A: .03 M LiOH
.19 M Borate
8.1
Stock S o l . B: .05 M T r i s
.008 M C i t r a t e
8 .4
Electrode:
Stock S o l u t i o n A
Gel:
1:9 Mixture Stock
S o l u t i o n A and B
113
S t a i n i n g Procedures
( S e la n d e r e t a l . 1971, Shaw and Prasad 1965)
IDH
0 .2 M t r i s - H C l (pH 8 .0 )
0.25 M manganese c h l o r i d e
0.10 M t r i - s o d i u m D L - i s o c i t r i c a c i d
NADP
NBT
PMS
Inc ubate 30-60 mintues (dark)
50
0 .2
3
10
5
7
ml
ml
ml
mg
mg
mg
30
5
20
10
20
5
ml
ml
ml
mg
mg
mg
50
0 .5
200
150
100
ml
mg
mg
mg
mg
50
I
50
20
13
4
ml
ml
mg
mg
mg
mg
MDH
0. 2 M t r i s - H C l (pH 8 . 0 )
2 .0 M mala te
Ho0
N&D
NBT
PMS
Inc ubat e 1-2 hours (dark)
GOT
0. 2 M t r i s - H C l (pH 8 .0 )
Pyridoxial-5'-phosphate
- a s p a r t i c acid
F a s t blue BB
-ketoglutarate
Inc ubate 10-15 minutes
a-GPD
0 .2 M t r i s - H C l (pH 8 . 0 )
0.1 M MgCl2
Disodium Dc -glycerophosphate
NAD
NBT
PMS
Inc ubat e 1-2 hours (dark)
114
PGM
0 .2 M t r i s - H C l (pH 8 . 0 )
H2O
0.05 Mrdis o d i um-D-glucose-1-phosphate
0.0005 M d i p o t a s s i um-D-glucose-1,6d ip hos ph at e
0.1 M MgCl2
10 u n i t s / m i H2O g lu c o s e - 6 - p h o s p h a te
dehydrogenase
NADP
MTT
PMS
Inc ubat e I hour (da rk)
5 ml
25 ml
5 ml
5 ml
5 ml
4
5
5
2
ml
mg
mg
mg
20
30
4 .5
20
4
8
ml
ml
ml
mg
mg
mg
30
5
20
10
20
5
ml
ml
ml
mg
mg
mg
45
5
15
10
2
ml
ml
mg
mg
mg
LDH
0 .2 M t r i s - H C l (pH 8 . 0 )
HO
1.0 M f a c t a t e
NAD
NBT
PMS
Incubate 1-2 hours (dark)
ME
0 .2 M t r i s - H C l (pH 8 .0 )
2 .0 M malate
H2O
NADP
NBT
PMS
In cu ba te 5-6 hours (dark)
G6PD
0 .2 M t r i s - H C l (pH 8 .0 )
0.25 M d i sodium g l uc os e -6 -p hos ph a te
NADP
NBT
PMS
Incubate 6 hours (dark)
115
H6PD
Same as G6PD but s u b s t i t u t e d i sodium g a l a c t o s e 6-pho sp ha te f o r g l uc os e -6 -p hos pha te
TO
0 .2
M t r i s - H C l (pH 8 . 0 )
NBT (MTT)
PMS
In cu ba te 1-2 hours ( l i g h t )
50 ml
I 5 mg
10 mg
Esterase
0.1
M sodium phosphate
HO
Napthyl p r o p r i o n a t e o r
B Napthyl p r o p r i o n a t e
(I gm/100 ml a c e to n e )
F a s t blue RR
4 ml
45 ml
I ml
25 mg
XDH
0 .2
t r i s - H C l (pH 8 . 0 )
Hypoxanthine
NAD
NBT
MTT
PMS
Inc ubate 3 hours (dark)
50 ml
25 mg
10 mg
10 mg
10 mg
5 mg
Serum P r o t e i n s
2% B uff al o Black NBT in f i x i n g s o l u t i o n
S t a i n 20 minutes a t 20°C. Wash gel s e v e r a l times in f i x i n g s o l u t i o n .
ADH
.5 M KPO4 (pH7 .0 )
H2O4
95% e th an ol
NAD
NBT
PMS
Inc ub a te 3-5 hours (d a rk ) (m ig rate s c a t h o d a l ly)
5 ml
40 ml
5 ml
30 mg
20 mg
5 mg
116
SDH
0 .2 M t r i s - H C l (pH 8 . 0 )
Sorbitol
NAD
MTT
PMS
Incubate 3 hours (dark)
50 ml
10 mg
10 mg
5 mg
2 mg
Hexokinase
0 .2 M t r i s - H C l (pH 8 . 0 )
H2O
GTucose
•1 M MgClp
G6PD
NADP
ATP
NBT
PMS
Incubate 3 hours (dark)
10 ml
40 ml
45 mg
10 ml
40 u n i t s
15 mg
15 mg
10 mg
2 mg
LITERATURE CITED
A l l e n d p r f , F. W. (1973) Genetic v a r i a t i o n , i n h e r i t a n c e , and
p r e l i m i n a r y p o p u l a t i o n d i s t r i b u t i o n o f some p r o t e i n s o f Salmo
gaivdnevl. M.S. T h e s i s , Univ. o f Washington.
A l l e n d o r f , F. W. and F. M. U t t e r .
(1973) Gene d u p l i c a t i o n w i t h i n
t h e fa m il y Salmonidae: Disomic i n h e r i t a n c e o f two l o c i r e p o r t e d
t o be t e t r a s o m i c in rainbow t r o u t . Genet ic s 74: 647-654.
A l l e n d o r f , F. W. and F. M. U t t e r .
(1976) Gene d u p l i c a t i o n in th e
f a m ily Salmonidae. I I I . Linkage between two d u p l i c a t e d l o c i
coding f o r a s p a r t a t e a m i n o t r a n s f e r a s e in t h e c u t t h r o a t t r o u t
[Salmo o la v k i) . H e r e d i t a s 82: 19-24.
All e n d o r f , F. W., F. M. U t t e r and B. P. May. (1975) Gene d u p l i ­
c a t i o n w i t h i n t h e f a m ily Salmonidae: I I . D e te c ti o n and d e t e r ­
mi nation o f t h e g e n e t i c c o n t r o l o f d u p l i c a t e l o c i through
i n h e r i t a n c e s t u d i e s and t h e ex amination o f p o p u l a t i o n s . In
M a r k e r t , C. L. ( e d ) , Isozymes IV. G en etic s and Ev o l u ti o n .
Acad. P r e s s , New Yorkl pp. 415-432.
A l l e n d o r f , F. W., N. Ryman, A. Stennek and G. S t a h l .
(1976)
Genetic v a r i a t i o n in Scandanavian brown t r o u t {Salmo tv u tta L . )
Evidence o f d i s t i n c t s ym pat ri c p o p u l a t i o n s . H e r e d i t a s 8 3 :1 ;
73-82.
Altukhov, J . P . , E. A. Salmenkova, V. T. Omelchenko, T. D. Sachko,
V. I . Slynko.
(1972) Number o f mono- and polymorphic l o c i in
t h e p o p u l a t i o n o f t e t r a p l o i d salmon s p e c i e s Onoorhynchus keta.
Gen etica 8 ( 2 ) : 67-75.
Appel l a , E. and C. L. M a r k e r t . (1961) D i s s o c i a t i o n o f l a c t a t e
dehydrogenase i n t o s u b u n i t s with gu an id in e h y d r o c h l o r i d e .
Biochem. Biophys. Res. Commun. 6: 171-176.
A s p i n w a l l , N. (1973) I n h e r i t a n c e o f a lp h a - g l y c e r o p h o s p h a t e
dehydrogenase in t h e pink salmon, Onoorhynohus gorbusoha
(Walbaum). G e net ic s 73: 639-643.
Aspinwall,, N. (1974) Ge netic a n a l y s i s o f d u p l i c a t e m a la te dehy­
drogenase l o c i in t h e pink salm on, Onoorhynohus gorbusoha.
G en etics 76: 65-72.
118
Avis e , J . C. (1974) S y s te m a tic val ue o f e l e c t r o p h o r e t i c d a t a .
System. Z o o l. 23: 465-481.
A vi se, J . C. (1976) Ge netic d i f f e r e n t i a t i o n d urin g s p e c i a t i o n .
In Ayala, F. J . ( e d ) , Molecular E v o l u t i o n . S i n a u e r A s s o c . ,
Su nd e rla nd , Mass.
A vi s e, J . C. and F. J . Ayala.
(1976) Ge net ic d i f f e r e n t i a t i o n in
s p e c i o s e ve rsus de pa upe ra te ph yla ds: Evidence from t h e
C a l i f o r n i a minnows. Evdlu t i on 30: 46-58.
A vi s e, J . C. and R. K. S e l a n d e r .
(1972) E v o l u ti o n a r y g e n e t i c s
o f c a v e - d w e l l i n g f i s h e s o f t h e genus Astyanax . Evo lu tio n
26: 1-19,
A vi se, J . C. and M. H. Smith.
(1974) Biochemical g e n e t i c s o f
s u n f i s h . I . Geographic v a r i a t i o n and s u b s p e c i f i c i n t e r g r a d a ­
t i o n in t h e b l u e g i l l , Lepomls maeroekirus . E v olu tio n 28:
42-56.
Av ise, J . C ., J . J . Smith, and F. J . Ayala.
(1975) Adaptive
d i f f e r e n t i a t i o n with l i t t l e gen ic change between two n a t i v e
C a l i f o r n i a minnows. Evo lution 29: 3; 411-426.
Ayala, F. J .
(1975) Gen etic d i f f e r e n t i a t i o n d uri ng s p e c i a t i o n .
In Dobzhansky, I . , M. K. Hecht and W. C. S t e e r e ( e d s ) ,
E v o l u t i o n a r y B io lo g y , Plenum P r e s s , New York.
Ayala, F. J . and J . R. P ow e ll. (1972) Enzyme v a r i a b i l i t y in the
Drosophila w illis to n i group. VI. Levels o f polymorphism and
t h e p h y s i o l o g i c a l f u n c t i o n o f enzymes. Biochem. G e n et : 7:
334-345.
Ayala, F. J . , M. L. T race y, D. Hedgecock, and R. C. Richmond.
(1974) Gen etic d i f f e r e n t i a t i o n d u r i n g th e s p e c i a t i o n process
\x\ Drosophila. Evol uti on 28 (4 ) : 576-592.
B a i l e y , G. S . , G. T. Cocks, and A. C. Wilson. (1969) Gene
d u p l i c a t i o n in f i s h e s : Malate dehydrogenase o f salmon and
t r o u t . Biochem. Biophys. Res. Comm. 34: 605-612.
119
B a i l e y , G. S . , A. C. Wilson, J . E. H a lv e r , and C. L. Johnson.
(1970) M u l t i p l e forms o f s u p e r n a t a n t m al at e dehydrogenase in
salmonid f i s h e s . J . BioT. Chetn. 245(22) : 5927-5940.
Baker, C.
av ian
qu a il
Comp.
M. A. and C. Manwell. (1967) Molecular g e n e t i c s o f
p r o t e i n s . V I I I . Egg w hit e p r o t e i n s o f t h e m ig r a to r y
Cotwmix ootw m ix - new co ncep ts o f "h ybrid v i g o u r " .
Biochem. P h y s i o l . 23: 21-42.
Baker, C. M. A ., C. Manwel I , R. F. Labi s ky, and J . A. Harper.
(1966) Molecular g e n e t i c s o f a via n p r o t e i n s . V. Egg, blood
and t i s s u e p r o t e i n s o f t h e ri n g - n e c k e d p h e a s a n t , Phasiccnus
oolohious L. Comp. Biochem. P h y s i o l , 17: 467-499.
Baker, W. W. and B. Mintz.
(1969) Su buni t s t r u c t u r e and gene
c o n t r o l o f mouse NADP mala te dehydrogenase. Biochem. Genet.
2: 351.
Behnke, R. J .
(1972) The systematises o f salmonid f i s h e s o f
r e c e n t l y g l a c i a t e d l a k e s . J . F is h . Res. Bd. o f Canada
29: 639-671.
Berg, L. (1940) C l a s s i f i c a t i o n o f f i s h e s , both r e c e n t and f o s s i l .
Dok. Z o o l. I n s t . Akad. Nauk. SSSR. 5 ( 2 ) : 85-517.
Berg, L. (1955) C l a s s i f i c a t i o n o f f i s h e s , both r e c e n t and f o s s i l .
Ed. 2. Dok. Z o o l. I n s t . Akad. Nauk SSSR. 20: 1-286.
B er g e r , E.
(1970)
A comparison o f gene-enzyme v a r i a t i o n between
Genet ic s 66: 667-683.
Drosophila melccnogaster and D. simulccns.
B e u t l e r , E. and M. Morrison.
(1967) L o c a l i z a t i o n and c h a r a c t e r i s ­
t i c s o f he xos e -6 -p ho s pha te dehydrogenase (gluc os e d e h y d r o g e n a s e ) .
J . B i o l . Chem. 242: 5298.
Blanco, A. and W. H. Zinkham. (1963) L a c t a t e dehydrogenases in
human t e s t e s . Scie nce 139: 601-602.
Blanco, A ., W. H. Zinkham, and L. Kupchyk. (1964). G e n et ic c o n tr o l
and ontogeny o f l a c t a t e dehydrogenase in pigeon t e s t e s . J .
Exp. Z ool. 156: 137-152.
Booke, H. E. (1964) A review o f v a r i a t i o n s found in f i s h serum
p r o t e i n s . N.Y. Fish and Game J o u r .
11: 47-57.
.
120
B oni ng so re , A . , R. Canceddas A. N i c o l i n i , G. Damiani and A. DeFlora
(1971) Metabolism o f human e r y t h r o c y t e g l u c o s e -6 - p h o s p h a te
dehydrogenase. VI. I n t e r c o n v e r s i o n o f m u l t i p l e m o le c u la r for ms .
Arch. Biochem. and Biophys. 147: 493-501.
B ou le ng er s G. A. (1895) Remarks on some c r a n i a l c h a r a c t e r s o f
t h e salm onids. Proc. Zoo l. Soc. London f o r 1895. pp. 299-302.
Brannon, E. L. (1967). Genetic c o n t r o l o f m i g r a t i n g b e h a v io r of
newly emerged sockeye salmon f r y .
I n t . P a c i f i c Salmon F i s h .
Comm. P r o g r . R e p t. 16: 31 pp.
Brewer, G. J .
(1967) Achromatic re g i o n s o f t e t r a z o l i u m s t a i n e d
s t a r c h g e l s : i n h e r i t e d e l e c t r o p h o r e t i c v a r i a t i o n . Amer. J .
Hum. Genet. 19: 674-680.
Brown, C. J . D. (1943) Age and growth o f Montana g r a y l i n g .
Wild!. Mgmt. 7 ( 4 ) : 353-364.
Brown, C. J . D. (1971) F is he s o f Montana.
S t a t e U n i v e r s i t y , Bozeman. 207 pp.
CL
Big Sky Books, Montana
B u t l e r , P. J . G ., H. J o r n v a l l and J . I . H a r r i s .
L e t t . 2: 239.
(1969)
FEBS
Cederbaum, S. E. and A. Yoshida.
(1972) Te tr a z o li u m o xid a se
polymorphism in rainbow t r o u t . G e net ic s 72: 363-367.
Cederbaum, S. E. and A. Yoshida.
(1976) Glucose 6-ph os pha te
dehydrogenase in rainbow t r o u t . Biochem. Genet. 14: 245..
C la yt on , J . W., D. N. T r e t i a k and A. H. Kooythan. (1971) Genetics
o f m u l t i p l e m al at e dehydrogenase isozymes in s k e l e t a l muscle
o f w a lle ye {S tizo sted io n vitreum vitreu m ) . J . F i s h . Res. Bd.
Can. 28: 1005-1008.
C la y to n , J . W., D. N. T r e t i a k , B. N. B i l l e c k and. P. I h s s e n . (1975)
G en etics o f m u l t i p l e s u p e r n a t a n t and m it oc hondr ia l ma late
dehydrogenase isozymes in rainbow t r o u t [Salmo g a ird n eri).
In Isozymes IV: G en etic s and E v o l u t i o n , (ed. C. L. M a r k e rt ).
Acad. P r e s s , New York. pp. 415-432.
121
Cohen, P. I . W. and G. S. Omenn. (1972) Ge netic v a r i a t i o n o f the
c y to pl a sm ic and m ito c h o n d ri a l malic enzymes in t h e monkey
Maoaca nemestv-ina. Biochem. G e n et . 7: 289-301.
Cohen, P. I . W., G. S. Omenn, A. G. Motu lsky , S. Chen, and E. R.
G iblett.
(1973) R e s t r i c t e d v a r i a t i o n in g l y c o l y t i c enzymes
o f human b r a i n and e r y t h r o c y t e s . Nat. New. B i o l . 241: 228-233
Cope, E. D. (1865) P a r t i a l c a t a l o g u e o f t h e cold blooded
v e r t e b r a t e o f Michigan, P a r t TI. P r o c . Acad. N a t u r . S c i .
P h i l a d e l p h i a 17: 78-88.
C u r v i e r , G. (1829) Le regne animal d i s t r i b u e d ' a p r e s son
o r g a n i z a t i o n . P a r i s 2: 1-306.
D a rn al I , D. W. and I . M. Klotz.
(1972) P r o t e i n s u b u n i t s : A t a b l e
( r e v i s e d e d i t i o n ) . Arch. Biochem. Biophys. 149: 1-14.
Dean, J . D. and J . D. V a r l e y . (1974) Annual P r o j e c t Report - 1973
F is h e ry Management Program - Yellowstone National Park . U.S.
Fish and W i l d l i f e S e r v i c e Processe d R pt. 170 pp.
DeLigny, W. (1969) S e r o l o g i c a l and biochemical s t u d i e s on f i s h
p o p u l a t i o n s . Oceanographic Mar. B i o l . Ann. Rev. 7: 411-513.
DeLorenzo, R. J . and F. H. Ruddle.
(1970) G l u t a m i c - o x a l o a c e t i c
t r a n s a m i n a s e (GOT) g e n e t i c s in Mus musoulus:- Linkage, p o ly ­
morphism and phenotypes o f th e GOT-2 and GOT-I l o c i . Biochem.
Genet. 4: 259-273.
D ic ke rs on , R. E. (1972)
p r o t e i n . S c i . Amer.
The s t r u c t u r e and h i s t o r y o f an a n c i e n t
226 (4) : 58-72.
Dobzhansky, T. (1951) Genet ic s and t h e O rigin o f t h e S p e c i e s .
3rd ed. Columbia Univ. P r e s s , New York.
Dobzhansky, T. (1970) Genet ic s o f t h e Ev o l u ti o n a r y P r o c e s s .
Columbia Univ. P r e s s , New York.
E c k r o a t , L. R. (1973) A l l e l e fr eq ue ncy a n a l y s i s o f f i v e s o l u b l e
p r o t e i n l o c i in brook t r o u t , Salvelinu s fo n tin a lis ( M i t c h i l l ) .
Tran. Amer. F is h . Soc . 102: 335-340.
122
Engel, W., J . Faust and U. Wolf.
(1971) Isoenzyme polymorphism o f
th e s o r b i t o l and t h e NADP-dependent i s o c i t r a t e dehydrogenases
in th e f i s h fa m il y Cyprin i d a e . Am'm. Blood Grps. Biochem.
Gen et. 2: 127-133.
E n g e l, W. J . , J . O p t 1Hof and U. Wolf.
(1970) Ge nd uplik at io n durch
p o l y p i oid e e v o l u t i o n : d i e isoenzyme d e r s o r b i t dehydrogenase
bei h e r r i n g s und l a c h s a r t i g e n f i s c h e n [ ls o s p o n d y li) . Human^
g e n e t i k 9: 157-163.
E n g e l, W., J . Schmidtke and U. Wolf. (1971) Ge netic v a r i a t i o n of
a - g l y c e r o p h o s p h a t e dehydrogenase isoenzymes in c l u p e o i d and
salmonid f i s h . E x p e r e n ti a 27(12 ): 1489-1491.
E n g e l, W., J . Schmidtke and U. Wolf.
(1975) Diplo i d - t e t r a p l oid
r e l a t i o n s h i p s in t e l e o s t e a n f i s h e s . In M a r k e r t , C. L. ( e d ) ,
Isozymes IV: G e net ic s and E v o l u t i o n . Acad. P r e s s , New York,
pp . 449-462.
~
Eppe nbe rge r, H. M., A. Scholl and H. Ursprung.
L e t t e r s 14: 317-319.
(1971)
FEBS
Feeney, R. E. and W. D. Brown.
(1974) Plasma p r o t e i n s in f i s h .
In M. F l o r k i n and B. I . Scheer ( e d s ) , Chemical Zoology. V I I I .
Acad. P r e s s , New York.
Fr yd en berg, 0. and V. Simonsen. (1973) Ge netics o f Zoanoes popula­
t i o n s . V. Amount o f p r o t e i n polymorphism and degree o f genic
h e t e r o z y g o s i t y . H e r e d i t a s 75: 221-232.
G a l l , G. A. E . , C. A. Busack, R. C. Smith, J . R. Gold, and B. J .
Kornbla t t .
(1976) Biochemical g e n e t i c v a r i a t i o n in p o p u l a t i o n s
o f golden t r o u t , Satmo aquabonita. J . o f Hered. 67: 330-335.
G i l l i s p i e , J . H. and K. Kojima. (1968) The degree o f polymorphisms
in enzymes in vol ve d in energy compared t o t h a t in n o n - s p e c i f i c
enzymes in two Drosophila ananassae p o p u l a t i o n s . P r o c . N a t l .
Acad. S c i . US. 61: 582-585.
G i l l i s p i e , J . H. and C. H. Langley.
(1974) A gener al model to
acc oun t f o r enzyme v a r i a t i o n in n a t u r a l p o p u l a t i o n s , Genetics
76: 837-848.
123
Glassman, E. (1965) Gen etic r e g u l a t i o n o f x a n th in e dehydrogenase
in Drosoph-Lla melanogaster . Fed. P r o c . 24: 1243-1251.
Goldberg, E. (1963) L a c t i c and m a li c dehydrogenases in human
spermatozoa. Scie nce 139: 602-603.
Goldberg, E.
(1966)
L a c t a t e dehydrogenase o f t r o u t : h y b r i d i z a t i o n
Science 151: 1091-1093.
-Ln v-Lvo and -Ln v-Ltro.
G o t t l i e b , L. D. (1971)
s tu dy o f e v o l u t i o n .
Gel e l e c t r o p h o r e s i s : New approach t o th e
B io sc ie nc e 21: 939-944.
H a r r i s , H. (1966) Enzyme polymorphisms in man.
S e r . B. 164: 298-310.
H a r r i s , H.
174: I .
(1969)
Genes and isozymes.
P r o c . Roy. Soc.
P r o c . Roy. Soc. Lond. B.
H a r r i s , H. (1975) The P r i n c i p l e s o f Human Biochemical G e n e t i c s .
2nd ed. North Ho lla nd , Amsterdam.
H a r r i s , H. and D. A. Hopkinson. (1972) Average h e t e r o z y g o s i t y
p e r locus in man: an e s t i m a t e based on in c id e n c e o f enzyme
polymorphism. Ann. Hum. Genet. 36: 9-20.
H a r r i s , H. and D. A. Hopkinson. (1976) Handbook o f Enzyme
E l e c t r o p h o r e s i s in Human G e n e t i c s . North Ho lla nd, Amsterdam.
Henderson, N. S. (1965) Isozymes o f i s o c i t r a t e dehydrogenase:
s u b u n i t s t r u c t u r e and i n t r a c e l l u l a r l o c a t i o n . J . Exp. Z ool.
158: 263-274.
Henderson, N. S. (1966) Isozymes and g e n e t i c c o n t r o l o f NADPmala te dehydrogenase in mice. Arch. ,Biochem. Biop hys. 117: 28.
Henderson, N. S. (1968) I n t r a c e l l u l a r l o c a t i o n and g e n e t i c
c o n t r o l o f isozymes o f NADP-dependent i s o c i t r a t e dehydrogenase
and mala te dehydrogenase. Ann. N.Y. Acad. S c i . 151: 429-440.
H e n s h a l l , J . A. (1906) A l i s t o f t h e f i s h e s o f Montana.
Univ . o f Montana, No. 34(11) : 12 pp.
B ull.
H e r s b e r g e r , W. K. (1970) Some physiochemical p r o p e r t i e s o f t r a n s ­
f e r r i n s in brook t r o u t . Tr ans . Am. F i s h . Soc. 99: 207.
124
H i t z e r o t h , H . , J . Klose, S. Ohno9 and LI. Wolf. (1968). Asynchronous
a c t i v a t i o n o f p a r e n t a l a l l e l e s a t t h e t i s s u e s p e c i f i c gene lo c i
observed on h ybrid t r o u t d urin g e a r l y development. Biochem.
G e n e t. I : 287-300.
Hochachka9 P. W. and G. A. Somero. (1971) Biochemical a d a p t a t i o n
t o t h e environment. In Hoar9 W. S. and D. Randall ( e d s ) , Fish
P h y s i o l o g y , Vol. G9 Acad. P r e s s , New York. pp. 99-156.
Holton, G. D. (1971) Montana g r a y l i n g : th e lady o f t h e s tr e a m s .
Montana Outdoors S e p t/ O c t: 18-23.
Hopkinson9 D. A. and H. H a r r i s .
(1968) A t h i r d phosphoglucomutase
lo cu s in man. Ann. Hum. Genet. Lond. 30: 167.
Hopkinson9 D. A. and H. H a r r i s .
(1971)
in man. Ann. Rev. Gen et. 5: 5.
Recent work on isozymes
Ho row itz 9 J . J . and G. S. Whitt.
(1972) Evolution o f a nervous
system s p e c i f i c l a c t a t e dehydrogenase isozyme in f i s h . J . Exp.
Z o o l. 180: 13-32.
Hubbs9 C. I . and K. :F. L a g le r.
(1949) Fi sh es o f t h e G reat Lakes
r e g i o n . Cranbook I n s t . S c i . B u l l . No. 26. 186 pp.
H u n t e r 9 R. L. and C. L. M a r k e r t. (1957) Histochemical de m onst rat ion
o f enzymes s e p a r a t e d by zone e l e c t r o p h o r e s i s in s t a r c h g e l s .
Science 125: 1294-1295.
Jo hns on 9 A. G ., F. M. U t t e r and H. 0. Hodgins. (1970a) E l e c t r o ­
p h o r e t i c v a r i a n t s o f L - al p h a - g ly c e r o p h o s p h a te dehydrogenase in
P a c i f i c ocean perch {Sebastodes g lu tu s ) . J . F i s h . Res. Bd. Can.
27: 943-945.
Jo hns on9 A. G. , F. M. U t t e r and H. 0. Hodgins.
(1970b) I n t e r ­
s p e c i f i c v a r i a t i o n o f t e t r a z o l ium o xid a se in Sebastodes (rockf i s h ) . Comp. Biochem. P h y s i o l . 37: 281-286.
Jo h n so n 9 A. G., F. M. U t t e r and H. 0. Hodgins. (1973) Estimates
o f g e n e t i c polymorphism and h e t e r o z y g o s i t y in t h r e e s p e c i e s
o f r o c k f i s h (genus S ebastes). Comp. Biochem. P h y s i o l . 44B:
397-406.
(
125
Johnson, G. B. (1971) Metabolic i m p l i c a t i o n s o f polymorphism as
an a d a p t i v e s t r a t e g y . Nature 232: 347-349.
Johnson, G. B. (1974)
184: 28-37.
Enzyme polymorphism and metabolism.
Science
J o r d a n , D. S. and B. W. Evermann. (1923) American Food and Game
F i s h e s . Doubleday, Page C o ., New York. 574 pp.
K e ll y , J . L. (1931)
3 ( 8 ) : 9-11.
Nation watches Montana g r a y l i n g .
Mt. W i l d l .
Khanna, N. D., R. K. J u n e j a , B. L a r s s o n , and B. Gahne. (1975)
E l e c t r o p h o r e t i c s t u d i e s on p r o t e i n s and enzymes in t h e A t l a n t i c
salmon, Salmo sdlca> L. Swed. J . Ag. Res. 5: 185-192.
Kimura, M. and J . F. Crow. (1964) The number o f a l l e l e s t h a t can
be m a in ta in ed in a f i n i t e p o p u l a t i o n . Genetics 49: 725-738.
King, M. and A. C. Wilson.
(1975) Evol uti on a t two l e v e l s :
m o le c u la r s i m i l a r i t i e s and b i o l o g i c a l d i f f e r e n c e s between
humans and chimpanzees. Science
Kirkman, H. and K. M. Hendrickson.
(1963) Sex l i n k e d e l e c t r o ­
p h o r e t i c d i f f e r e n c e in g l uc ose -6 -p hos pha te dehydrogenase.
Am. J . Hum. Gen et. 15: 241.
Kir p i c h n i k o v , V. S. (1973) Biochemical polymorphism and micro­
e v o l u t i o n p r o c e s s e s in f i s h . In Sc hr ode r, J . H. ( e d ) ,
G en etics and Mutagenesis o f F i s h , S p r i n g e r - V e r l a g , New York,
pp. 223-241.
K i t t o , G. B. and N. 0. Kaplan. (1966) P u r i f i c a t i o n and p r o p e r t i e s
o f chicken h e a r t m it o c h o n d ri a l and s u p e r n a t a n t m a li c dehydrogen­
a s e s . Bioc hem is try 5: 3966-3980.
Kojima, K., J . G i l l i s p i e , and Y. N. T o b a r i . (1970) A p r o f i l e o f
Drosophila s p e c i e s ' enzymes ass ay ed by e l e c t r o p h o r e s i s . I . Number
o f a l l e l e s , h e t e r o z y g o s i t i e s , and li n k a g e d i s e q u i l i b r i u m in
g l u c o s e - m e t a b o l i z i n g systems and some o t h e r enzymes. Biochem.
Genet. 4: 627-637.
126
K r i s t i a n s s o n , A. C. and J . D. McIntyre.
(1976) Gen etic v a r i a t i o n
in chinook salmon {Onaorhynahus tshawytsaha) from t h e Columbia
River and t h r e e Oregon c o a s t a l r i v e r s . Trans. Am. F is h . Soc.
105(6): 620-623.
Kruse, I . E. (1959) The g r a y l i n g o f Grebe Lake, Yellowstone
National Park , Wyoming. U.S. Fish and W i l d l i f e S e r v i c e ,
Fish B u l l . 149, v o l . 59: 307-351.
Lewontin , R. C. and J . L. Hubby.
(1966) A m o le c u la r approach to
t h e s tu d y o f ge ni c h e t e r o z y g o s i t y in n a t u r a l p o p u l a t i o n s . I I .
Amount o f v a r i a t i o n and degree o f h e t e r o z y g o s i t y in n a t u r a l
p o p u l a t i o n s o f Drosophila pseudoobsoura. G en etic s 54: 595-609.
Li ndsey, C. C. (1956) D i s t r i b u t i o n and taxonomy o f f i s h e s in the
Mackenzie d r a in a g e o f B r i t i s h Columbia. J . F i s h . Res. Bd. Can.
13: 759-798.
Lukjanenko, V. T. and A. V. Popov.
(1969) P r o t e i n s o f t h e serum
in two a l l o p a t r i c p o p u l a t i o n s o f t h e S i b e r i a n s tu r g e o n Aoipenser
baeri Br. Dokl. Acad. Nauk. 186(1) : 233-235.
Manwell, C. and C. M. A. Baker.
(1969) Hybrid p r o t e i n s , h e t e r o s i s ,
and t h e o r i g i n o f s p e c i e s . I . Unusual v a r i a t i o n o f po ly c h ae te
H y a lin oec ia " not hin g dehydrogenases" and o f q u a il Cotumix
e r y t h r o c y t e enzymes. Comp. Biochem. P h y s i o l . 28: 1007.
Manwell, C. and C. M. A. Baker. (1970) Molecular Biology and
O rig in o f S p e c ie s : H e t e r o s i s , P r o t e i n Polymorphism, and Animal
B r e e d in g . Sidgwick and J a c k s o n , London.
M a r k e r t , C. L. (1962) In M e t c o f f , J . ( e d ) , H e r e d i t a r y , Develop­
mental and Immunological Aspects o f Kidney D is ea s e. North­
west ern Univ. P r e s s , I l l i n o i s ,
pg. 54.
M a r k e r t , C. L.
Acad. S c i .
(1968) Molecular b a s i s f o r isozymes.
151: 14-40.
Ann. N.Y.
M a r k e r t , C. L. and I . F a u l h a b e r . (1965) L a c t a t e dehydrogenase
isozyme p a t t e r n s o f f i s h . J . Exp. Z o o l. 159: 319-332.
M a r k e r t , C. L. and R. S. Holmes. (1969) L a c ta t e dehydrogenase
isozymes o f t h e f l a t f i s h , P l e u r o n e c t i formes: k i n e t i c , m o l e c u l a r , .
and immunochemical a n a l y s i s . J . Exp. Z ool. 171: 85-104.
127
Massaro, E. J .
(1973) T i ss u e d i s t r i b u t i o n and p r o p e r t i e s o f the
l a c t a t e and ma la te dehydrogenase isozymes o f t h e g r a y l i n g
ThymaVlus avct-ious ( P a l l a s ) . J . Exp. Z ool. 186: 151-158.
Massaro, E. J . and C. L. M a r k e r t. (1968) Isozyme p a t t e r n s of
salmonid f i s h e s : e vid enc e f o r m u l t i p l e c i s t r o n s f o r l a c t a t e
dehydrogenase p o l y p e p t i d e s . J . Exp. Z ool. 168: 223-228.
May, B. M., F. M. U t t e r and F. W. A l l e n d o r f . (1975) Biochemical
g e n e t i c v a r i a t i o n in pink and chum salmon: I n h e r i t a n c e o f
i n t r a s p e c i e s v a r i a t i o n and a p p a r e n t abscence o f i n t e r s p e c i e s
i n t r o g r e s s i o n f o ll o w i n g massive h y b r i d i z a t i o n o f h a t c h e r y
s t o c k s . J . Hered. 66: 227-232.
Mayr, E. (1963) Animal Spec ies and E v o l u t i o n .
P r e s s , Cambridge, Mass.
Harvard Univ.
McCabe, M. M., D. M. Dean and C. S. Olson.
(1970) M u lt ip le
forms o f 6-p hosp hogluconate dehydrogenase and a lp h a g l y c e r o ­
phosphate dehydrogenase in t h e s h i p j a c k t u n a , Katsuwonus
pelamis. Comp. Biochem. P h y s i o l . 34: 755-757.
McPhail, J . D. and C. C. Lindsey.
(1970) Fres hw at er f i s h e s of
no rt h w e s t Canada and Alaska. F i s h . Res. Bd. o f Canada
Bull No. 173.
Metzger, R. P . , S. Wilcox and A. Wick.
(1965) S u b c e l l u l a r d i s t r i ­
b u ti o n and p r o p e r t i e s o f h e p a t i c g lu c ose dehydrogenases o f
s e l e c t e d v e r t e b r a t e s . J . B io l. Chem. 240(7): 2767.
M i l n e r , J . W. (1873) Notes on t h e g r a y l i n g s o f North America.
In Fr es hw at er F i s h e r i e s o f t h e United S t a t e s . U.S. Govt.
P r i n t i n g O f f i c e , Washington, D.C. pp. 730-742.
M o ll e r , D.
(1970)
{Salmo s a la r ) .
T r a n s f e r r i n polymorphism in A t l a n t i c salmon
J . F is h . Res. Bd. Can. 27: 1617-1625.
Moore, B. W. and R. H. Lee. (I960) Chromatography o f r a t l i v e r
s o l u b l e p r o t e i n s and l o c a l i z a t i o n o f enzyme a c t i v i t y . J v
B i o l . Chem. 234: 1359.
Mor ris on, W. J .
(1970) Nonrandom s e g r e g a t i o n on two l a c t a t e
dehydrogenase s u b u n i t l o c i in t r o u t . Trans. Amer. F i s h . Soc.
99: 193-206.
128
Mo rrison, W. J . and 0. E. Wright.
(1966) Genetic a n a l y s i s o f .
t h r e e l a c t a t e dehydrogenase systems in t r o u t : Evidence f o r
l in k a g e o f genes coding s u b u n i t s A and B. J . Exp. Z o o l .
163: 259-270.
Nair , P. S . , D. Brncic and K. Kojima. (1971) I I . Isozyme v a r i a t i o n s
and e v o l u t i o n a r y r e l a t i o n s h i p s in t h e mesophragmatica s p e c i e s
group o f Drosophila. S t u d i e s in G en etic s VI. (Univ. Texas
P u b l . No. 7103) pp. 17-28.
N e i , M. (1971) I n t e r s p e c i f i c gene d i f f e r e n c e s and e v o l u t i o n a r y
time e s t i m a t e d from e l e c t r o p h o r e t i c d a ta on p r o t e i n i d e n t i t y .
Amer. N a t- 105: 385-398.
N e i , M. (1972) Ge net ic d i s t a n c e between p o p u l a t i o n s .
106: 283-292.
Amer. N a t.
N e i, M. (1975) Molecular Evol uti on and Po pu la ti o n G e n e t i c s .
North Holland', Amsterdam.
N e i , M. and R. C h a k r a b o r ty . (1973) Ge net ic d i s t a n c e and e l e c t r o ­
p h o r e t i c i d e n t i t y o f p r o t e i n s between t a x a . J . Molec. Evo l.
2: 323-328.
N e i , M. and A. K.. Roychoudhury. (1974)
Genic v a r i a t i o n w i t h i n
and between t h e t h r e e major r a c e s o f man, C au ca soi ds , Negroids,
and Mongoloids. Amer. J . Hum. G e net. 26: 421-443.
N e i , M. and A. K. Roychoudhury. (1974)
h e t e r o z y g o s i t y and g e n e t i c d i s t a n c e .
Sampling v a r i a n c e s o f
Genet ic s 76: 379-390.
N e i , M., I . Maruyama and R. C h a k r a b o r ty . (1975) The b o t t l e n e c k
e f f e c t and g e n e t i c v a r i a b i l i t y in p o p u l a t i o n s . Evo lu tio n
29: 1-10.
Nelson, P. H. (1954) L i f e h i s t o r y and management o f t h e American
g r a y l i n g (Thymallus s ig n ife r tr ic o lo r ) in Montana. J . W ild !.
. Mgmt. 18( 3): 324-342.
Nelson, P. H. (1956) The g r a y l i n g - an endangered s p e c i e s .
Wild!. 6 ( 1 ) : 20-21.
Mt.
Noltman, F. A. and S. A. Kuby. (1963) D-gl ucose-6-pho.sphate and
6-phosp hogluc ona te de h y d ro g e n as es . In Boyer, P . , H. Lard y, and
R. Myrback ( e d s ) , The Enzymes, V ol. 7. Acad. P r e s s , New York.
f
129
Norderii C. R. (1961) Comparative o s t e o l o g y o f r e p r e s e n t a t i v e
salmonid f i s h e s w it h p a r t i c u l a r r e f e r e n c e t o t h e g r a y l i n g
(Thymallus capctious) and i t s phylogeny. J . F is h . Res. Bd.
Can. 18: 679-791.
N o r t h c o t e , I . G. (1969). P a t t e r n s and mechanisms in t h e lakeward
m ig r a to r y b e h a v i o r o f j u v e n i l e t r o u t , in Symposium on Salmon
and Trout in S tr e a m s . N o r t h c o t e , I . G. ( e d j , Univ. B r i t . Co!. ,
Vancouver. pp\ 183-204.
Nyman, L.
(1972)
A new approach t o t h e taxonomy o f t h e Salvelinus
Rep. I n v t . Freshw. Res. Drottingholm
alpinus s p e c i e s complex.
52: 103-131.
O ' B r i e n , S. J . and R. J . MacIntyre.
(1969) An a n a l y s i s o f geneenzyme v a r i a b i l i t y in n a t u r a l p o p u l a t i o n s o f Drosophila
melanogaster and Drosophila simulans. American N a t u r a l i s t
103: 97-113.
Ohno, S . , U. Wolf, and N. B. Atkin.
t o mammals by gene d u p l i c a t i o n .
(1968) Evol uti on from f i s h
H e r e d i t a s 59: 169-187.
Ohno, S . , H. W. Payne, M. Morrison and E. B e u t l e r .
6-p ho sp ha te dehydrogenase found in human l i v e r .
1015.
(1966) HexoseScie nce !53:
Ohnoi S . , J . Muramato, J . Klein and N. B. Atkin.
(1969) D ip ld id t e t r a p l o i d r e l a t i o n s h i p in Clupeoid and Salmonid f i s h .
Chromosomes Today 2: 139-147.
O p t1Hof, J .
(1969) Isoenzymes and p o p u l a t i o n g e n e t i c s o f s o r b i t
dehydrogenase (E.C. : 1 . 1 . 1 . 1 4 ) in swine {Sus sorofa)..
Humangenetik 7: 258-259.
O p t' Hof, J . , U. Wolf and W. Krone. (1969) S t u d i e s on isozymes
o f s o r b i t o l dehydrogenase in some v e r t e b r a t e s p e c i e s . Humang e n e t i k 7: 178-182.
F a r r i n g t o n , J . M., G. Cruic kshank, D. A. Hopkinson, E. B. Robson,
and H. H a r r i s .
(1968) Linkage r e l a t i o n s h i p s between t h e t h r e e
phosphoglucomutase l o c i PGM, PGM0 and PGM0 . Ann. Hum. Genet.
32: 37.
*
*
:
130
Payne, R. H., A. R. C h i l d , A. F o r r e s t .
(1971) Geographical v a r i a ­
t i o n in th e A t l a n t i c salmon. Nature 231: 250-252.
P e r r i e r , H., J . P. D e l c r o i x , C. P e r r i e r , and J . G ro s. (1973) An
a t t e m p t t o c l a s s i f y t h e plasma p r o t e i n s o f t h e rainbow t r o u t
{Salmo gaivdneri Richardson) us in g d i s c e l e c t r o p h o r e s i s , gel
f i l t r a t i o n and s a l t s o l u b i l i t y f r a c t i o n a t i o n . Comp. Biochem.
P h y s i o l . 4 6 ( b ) : 475-482.
Povey, S . , D. E. Wilson and H. H a r r i s .
(1975) Su bunit s t r u c t u r e
o f s o l u b l e and m it o c h o n d ri a l ma lic enzyme: d e m on st ra ti on o f
human m it o c h o n d r i a l enzyme in human-mouse h y b r i d s . Ann. Hum.
Genet. Lond. 39: 203.
Powell, J .
(1975) P r o t e i n v a r i a t i o n in n a t u r a l p o p u l a t i o n o f
a ni m a ls . E v o l u t i o n a r y B io lo g y , VoT. 8. Plenum P r e s s , New
York, pp. 79-119.
P r a k a s h , S . , R. C. Lewontin and J . L. Hubby. (1969) A m ol e c ula r
approach t o t h e s t u d y o f genic h e t e r o z y g o s i t y in n a t u r a l
p o p u l a t i o n s . IV. P a t t e r n s o f ge nic v a r i a t i o n in c e n t r a l ,
marginal and i s o l a t e d p o p u l a t i o n s o f Drosophila pseudoobsouva.
G en etics 61: 84T-858.
R a l e ig h , R. F. (1967) Genetic c o n t r o l in t h e lakeward m ig r a ti o n s
o f sockeye salmon, Onoorhynohus nerka, f r y . J . F i s h . Res.
Bd. Can. 24: 2613-2622.
"
R a l e ig h , R. F. and D. W. Chapman. (1971) Genetic c o n t r o l in
lakeward m i g r a t i o n s o f c u t t h r o a t t r o u t f r y . Tran s. Amer . Fish.
Soc. 100(1) : 33-40.
:
i
:
Reagan, C. I .
(1914) The s y s t e m a t i c arrangement o f t h e f i s h e s o f
t h e f a m ily Salmonidae. Ann. Mag. Nat. H i s t . 13 (8 ) : 405-408.
R e i n i t z , G. L. (1974) I n t r o g r e s s i v e h y b r i d i z a t i o n and v a r i a t i o n
in Salmo o la r k ii and S. gairdneri in Montana. M.S. T h e s i s ,
Univ. o f Montana, Missoula.
R ic ha rd s on , J . R. (1823). Notice o f t h e f i s h e s , in F r a n k l i n , 0.
( e d ) , N a r r a t i v e o f a Journey t o t h e Shores o f t h e P o l a r Sea in
t h e y e a r s 1819, 1820, 1 8 2 1 , 1 8 2 2 . John Murray, London. pp%
705-728.
131
(1972) Enzyme v a r i a b i l i t y in the Drosophila
w illis to n i group. I I I . Amounts o f v a r i a b i l i t y in th e s u p e r ­
s p e c i e s D. paulistorwn. Genet ic s 70: 87-112.
Richmond, R. C.
Ridgeway, G. J . , S. W. Sherburne and R. D. Lewis. (1970)
Polymorphism in t h e e s t e r a s e s o f A t l a n t i c h e r r i n g . Trans.
Amer. Fi s h. Soc. 99: 147-151.
Riechenbach-Klin k e , H. H. (1973) I n v e s t i g a t i o n s on t h e serum
polymorphism o f t r o u t and c ar p. I n Sc h ro d e r, J . H. ( e d ) ,
G en etics and Mutagenesis o f F i s h , S p r i n g e r - V e r l a g , B e r l i n ,
pp . 3Y5-318.
R o b e r t s , F. L . , J . F. Wohnus and S. Ohno. (1969) Phosphoglucomutase polymorphism in t h e rainbow t r o u t , Salmo gaivdnevi.
E x p e re n ti a 25( 10): 1109-1110.
Rogers, J . S. (1972) Measures o f g e n e t i c s i m i l a r i t y and g e n e t i c
d i s t a n c e . S t u d i e s in Genet ic s VII (Univ. Texas P u b l . No.
7213). pp. 145-153.
Ropers, H. H., W. E n g e l , and U. Wolf.
(1973). I n h e r i t a n c e o f the
s -form o f NADP-dependent i s o c i t r a t e dehydrogenase polymorphism
in rainbow t r o u t . In Sc h ro d e r, J . H. ( e d ) , G e net ic s and
Mutagenesis o f F i s h , S p r i n g e r - V e r l a g , New York. pp. 319-327.
S a r i c h , V. M. (1977) R a t e s , sample s i z e s , and t h e n e u t r a l i t y
h y p o th e s i s f o r e l e c t r o p h o r e s i s in e v o l u t i o n a r y s t u d i e s .
Nature 265: 24-28.
Schmid t k e , J . and W. E nge l. (1972) D u p l i c a t i o n o f t h e gene lo c i
coding f o r t h e s u p e r n a t a n t a s p a r t a t e a m i n o t r a n s f e r a s e by
p o l y p i o i d i z a t i o n in t h e f i s h f a m ily Cyprin i d a e . E x p e r i e n t i a
28: 976-978.
~
S c o t t , W. B. and E. J . Crossman. (1973) Fr eshwater f i s h e s o f
Canada. F i s h . Res. Bd. Can. B u l l . 184: 300-305.
S e l a n d e r , R. K. (1976) Genic v a r i a t i o n in n a t u r a l p o p u l a t i o n s .
In Ayala, F. J . ( e d ) , Molecular E v o l u t i o n . S in a e u r A s s o c . ,
Sund erla nd, Mass.
S e l a n d e r , R. K. and W. E. Johnson.
(1973) Genetic v a r i a t i o n among
v e r t e b r a t e s p e c i e s . Ann. Rev. E c o l . S y s t . 4: 75-91.
132
S e l a n d e r , R. K. and S. Yang.
(1969) P r o t e i n polymorphism and genic
h e t e r o z y g o s i t y in a w il d p o p u l a t i o n o f t h e house mouse {Mus
musculus) . Genet ic s 63: 653-667.
S e l a n d e r , R. K., W. G. Hunt and S. Y. Yang. (1969) P r o t e i n p oly ­
morphism and gen ic h e t e r o z y g o s i t y in two European s u b s p e c ie s
o f t h e house mouse. Evol uti on 23: 379-390.
S e l a n d e r , R. K., M. H. Smith, S. Y. Yang, W. E. Johnson and J . B.
Gentry.
(1971) Biochemical polymorphism and s y s t e m a t i c s in
t h e genus Pevomyseus. I . V a r i a t i o n in t h e o ld f i e l d mouse
(Peromyscus p o lio n o tu s) . S t u d i e s o f Genetics VI. (Univ.
o f Texas P u b l . No. 7103) pp. 49-90.
S ha kl e e, J . B ., K. L. Kepes, and G. S. Wh itt . (1973) S p e c i a l i z e d
l a c t a t e dehydrogenase isozymes: t h e m ol e c ula r and g e n e t i c b a s i s
f o r t h e unique eye and l i v e r LDHs o f T e l e o s t f i s h e s . J . Exp.
Z ool. 185: 217-240.
S h a t t o n , J . B ., J . E. Halver and S. Weinhouse. (1971). Glucose
(h ex o s e- 6 - p h o sp h a te ) dehydrogenase in l i v e r o f rainbow t r o u t .
J . B i o l . Chem. 246: 4878.
Shaw, C. R. (1965)
149: 936-943.
E l e c t r o p h o r e t i c v a r i a t i o n in enzymes.
Science
"
Shaw, C. R. (1966) G luc ose -6- phos phat e dehydrogenase homologous
molecules in d e e r mouse and man. Science 153: 1013.
Shaw, C. R. (1970)
275-283.
How many genes evo lve ?
Biochem. Genet.
:
:
4:
Shaw, C. R. and E. Bar to.
(1963) Ge net ic evidence f o r t h e s u b u n it
s t r u c t u r e o f l a c t a t e dehydrogenase. Proc. N a t l . Acad. S c i . U.S.
50: 21 V 214 .
:
~
Shaw, C. R. and L. Koen. (1968) G1ucose-6-phosphate dehydrogenase
and he xo se -6 -p ho s pha te dehydrogenase o f mammalian t i s s u e s .
Ann, N.Y. Acad. S c i . 151: 149.
Shaw, C. R. and R. Pras ad .
(1970) S ta r c h gel e l e c t r o p h o r e s i s o f
enzymes - a c o m p il a ti o n o f r e c i p e s . Biochem. Genet. 4: 297-320.
133
Shows, I . B. and F. H. Ruddle. (1968) Malate dehydrogenase. Evidence
f o r t e t r a m e r i c s t r u c t u r e in Mus musoulus. S c i e n c e , N.Y. 160: 1356.
Shows, I . B ., V. M. Chapman, and F. H. Ruddle.
(1970) Mi tochondrial
mala te dehydrogenase and malic enzyme: Mendelian i n h e r i t e d
e l e c t r o p h o r e t i c v a r i a n t s in th e mouse. Biochem. Genet. 4: 707718.
"
S i e g e l , L. and S. England.
(1961) Beef h e a r t mala te dehydrogenase:
p r o p e r t i e s o f the enzyme p u r i f i e d from e x t r a c t s o f a c e to n e d r i e d
powder. Biochim. Biophys. Acto 54: 67-76.
S l a s t e n e n k o , E. P. (1958).
P r i n t e r s , Toronto.
The F resh w at er Fi sh e s o f Canada.
Kiev
S m i t h i e s , 0. (1955). Zone e l e c t r o p h o r e s i s in s t a r c h g e l s : group
v a r i a t i o n s in t h e serum p r o t e i n s o f normal a d u l t s . Biochem. J .
61: 629-641.
S o k a l , R. and F. J . Rohlf . (1969) Biometry. The P r i n c i p l e s and
P r a c t i c e o f S t a t i s t i c s in B i o l o g i c a l Research’. W.R. Freeman,
San F r a n c i s c o .
S o k a l , R. R. and P. H. A. S n e a t h . (1963). P r i n c i p l e s o f Numerical
Taxonomy. W.H. Freeman, San F r a n c i s c o .
Spe nc er , N ., D. A. Hopkinson and H. H a r r i s .
(1964) Phosphoglucomutase polymorphism in man. Nature 204: 742-745.
Stegeman, J . J . and E. Goldberg.
(1971) D i s t r i b u t i o n and c h a r a c ­
t e r i z a t i o n o f he xo se -6 -p hos pha te dehydrogenase in t r o u t .
Biochem. Genet. 5: 579.
Stegeman, J . J . and E. Goldberg. (1972) I n h e r i t a n c e o f hexose6-p hosp ha te dehydrogenase polymorphism in brook t r o u t . Biochem.
G e net . 7: 279-288.
~~
S y l n ' k o , V. I . (1976) E l e c t r o p h o r e t i c a n a l y s i s o f isoenzymes o f
mala te dehydrogenase from f i s h o f th e Salmonidae f a m il y .
Doklady Akad. Nauk SSSR, 226(2 ): 448-451.
Th u r st o n , R. V. (1967) E l e c t r o p h o r e t i c p a t t e r n s o f blood serum
p r o t e i n s from rainbow t r o u t {Salmo g a ir d n e v i). J . F i s h . Res.
Bd. Can. 24: 2169-2188.
:
134
Ts u y u k i9 H. and E. R oberts.
(T966) I n t e r s p e c i e s r e l a t i o n s h i p s
w i t h i n t h e genus Onaorhynchus based on biochemical s y s t e m a t i c s .
J . F is h. Res. Bd, Can. 23: 101-107.
T u r n e r , . B. J .
(1974) Ge net ic div e rg e n c e o f Death V a ll ey pupf is h
s p e c i e s : Biochemical vs. morphological e v id en c e . Evo lution
28: 281-294.
—
U.S. Department o f I n t e r i o r .
(1966) Rare and endangered f i s h and
w i l d l i f e o f th e United S t a t e s . Bur. S p t . Fi s h. W i l d l . Res.
Pub. 24 Sheet F-7.
U t t e r , F. M. (1971) T e tr a z o li u m ox id a se phenotypes o f rainbow
t r o u t (Salmo g a ird n eri ) and P a c i f i c salmon ( Onoovhynohus s p p . ).
Comp. Biochem. P h y s i o l . 39B: 891-895.
U t t e r , F. M. and H. 0. Hodgins.
morphism in s o c keye salmon.
195-200.
(1970) Phosphoglucomutase p oly ­
Comp. Biochem. P h y s i o l . 86 (1 ) :
U t t e r , F. M. and H. 0. Hodgins. (1972) Biochemical g e n e t i c
v a r i a t i o n a t s i x Toci in f o u r s t o c k s o f rainbow t r o u t . Tran.
Amer. F is h . Soc. 101: 494-502.
U t t e r , F. M., W. E. Ames, and H. 0. Hodgins.
(1970) T r a n s f e r r i n
polymorphism in coho salmon (Onoovhynohus kisu toh ). J . Fish .
Res. Bd. Can. 27: 2371-2373.
:
U t t e r , F. M., F. W. A l l e n d o r f , and H. 0. Hodgins. (1973) Genetic
v a r i a b i l i t y and r e l a t i o n s h i p s in P a c i f i c salmon and r e l a t e d
t r o u t based on p r o t e i n v a r i a t i o n s . S y s t - Z o o l . 22: 257-270.
V in c e n t, R. E. (1962) Bio geo graphical and e c o l o g i c a l f a c t o r s
c o n t r i b u t i n g t o t h e d e c l i n e o f a r c t i c g r a y l i n g , Thymallus
avotious P a l l a s , in Michigan and Montana. Ph.D. T h e s i s , Univ.
o f Mighigan.
W a l t e r s , V. (1955) Fi sh es o f w e st e rn A r c t i c America and e a s t e r n
A r c t i c S i b e r i a : Taxonomy and zoogeography. B u l l . Amer. Mus.
Nat. H i s t . 106(5): 255-368.
Ward, R. D. (1977) R e l a t i o n s h i p between enzyme h e t e r o z y g o s i t y and
q u a t e r n a r y s t r u c t u r e . Biochem. Genet. 15: 123-135.
135
Wheat, I . E. and G. S. Whitt.
(1971) In vi-vo and in v itv o m ole c ul ar
h y b r i d i z a t i o n o f mala te dehydrogenase isozymes. E x p e re n ti a 27:
647-648.
Wheat, T. E . , W. F. C h i l d e r s , E. T. M i l l e r and G . S . Whi tt.
(1971)
Ge netic and in v itr o m o le c u la r h y b r i d i z a t i o n o f m al at e dehydrogen
a se isozymes in i n t e r s p e c i e s bass [Mioropterus) h y b r i d s . Anim.
Blood Groups Biochem. Gen. 2: 3-14.
W h it t, G. S. (1969) Homology o f l a c t a t e dehydrogenase genes: E
gene f u n c t i o n in t h e nervous system. Science 166: 1156^-1158.
W h i t t , G. S. (1970) Developmental g e n e t i c s o f t h e l a c t a t e
dehydrogenase isozymes o f f i s h . J . Exp. Z ool. 175: 1-36.
W hi tt, G. S. and G. M. Booth.
(1970) L o c a l i z a t i o n o f l a c t a t e
dehydrogenase a c t i v i t y in t h e c e l l s o f t h e f i s h {Xiphophorus
h e lle r i) eye. J . Exp. Zo ol. 174: 215-224.
W h it t, G . S . and J . J . Horowitz.
(1970) Evo lution o f r e t i n a l
s p e c i f i c l a c t a t e dehydrogenase isozyme in t e l c o s t s .
E xpe renti a
26: 1302-1304.
Wilson, A. C . , L. R. Maxson, and V. M. S a r i c h .
(1974a) Two types
o f m o le c u la r e v o l u t i o n : Evidence from s t u d i e s o f i n t e r s p e c i f i c
h y b r i d i z a t i o n . P r p c . N a tl . Acad. S c i . 71: 2843-2847.
Wilson, A. C . , V. M. S a r i c h and L. R. Maxson. (1974b) The
importance o f gene re arrangem en t in e v o l u t i o n : Evidence from
s t u d i e s on r a t e s o f chromosomal, p r o t e i n and anatomical
e v o l u t i o n . P r o c . N a t l . Acad. S c i . 71: 3028-3030.
Wolf, U., W. Engel, J . F a u s t.
(1970) Zum mechanisms d e r
d i p l o i d i s i e r u n g in d e r w i r b e l t i e r e v o l u t i o n : Koexistehz von
te tra so m en und disomen gen lo ci d e r i s o c i t r a t d e h y d r o g e n a s e
bei der' r e g e n h o g e n f o r e l l e , SaZmo irrid e u s. Humangenetik
9: 150-156.
Woods, K. and R. Engle.
(1957) Ph ylo ge ne si s o f plasma p r o t e i n s ,
and plasma c e l l s . I . S t a r c h gel zone e l e c t r o p h o r e s i s o f s e r a
from marine i n v e r t e b r a t e s and f i s h e s . B io l. B u l l . 113(2):
362-363.
136
Wright, J . E. and L. M. A th e rt o n .
(1970) Polymorphism f o r LDH
and t r a n s f e r r i n l o c i in brook t r o u t p o p u l a t i o n s . Tr an s . Amer.
F is h . Soc. 9 9 ( 1 ) : 179-192.
Wright, I . D. and A. D. H a s le r .
(1967) An e l e c t r o p h o r e t i c a n a l y s i s
o f t h e e f f e c t s o f i s o l a t i o n and homing b e ha vio r o f t h e serum
p r o t e i n s o f t h e w hi te bass {Rooous ohrysaps) in Wisconsin. Amer.
Natur. 101(921): 401-413.
Yamuchi, I . and E. Goldberg.
(1973) Glu c os e -6 -p hos ph a te dehydrogen­
a se from brook, la ke and s p la k e t r o u t : an isozymic and immuno­
l o g i c a l s tu d y . Biochem. G e net . 10: 121.
Zo ur os, E. (1975). E l e c t r o p h o r e t i c v a r i a t i o n in allozymes r e l a t e d
t o f u n c t i o n o r s t r u c t u r e ? Nature 254: 446-448.
Montana Ctatf ...___
3 1762 10014743
*
*
N3J 8
L 988
cop . 2
Lynch, Jeremiah C
Comparative g en etics
o f Montana and a r c tic
grayling . . .
IS S U E D TO
DATE
JAN
4 I
-