20.380 S10 Introduction: the Immune System– the basics, inflammation in health
advertisement

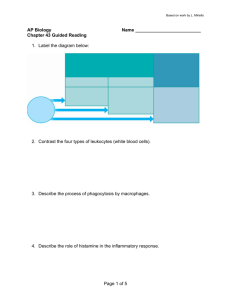
20.380 S10 Introduction: the Immune System– the basics, inflammation in health and disease 20.400 spring 2008 1 Overview of the immune system 2 Two arms of immunity: the innate and adaptive immune systems 20.400 spring 2008 2 KEY EFFECTORS OF ADAPTIVE IMMUNITY Diagram of how B lymphocytes, Helper T lymphocytes, and Cytolytic T lymphocytes recognize particular antigens and effect immunity has been removed due to copyright restrictions. 20.400 spring 2008 (Abbas) 2 THE CLONAL IMMUNE SYSTEM •10^12 total T cells in adult human •25-100 million distinct clones •Only several thousand T cells at most respond to any individual antigen (von Andrian and Mackay 2000) •Precursor frequency of antigen-specific cells: •CD8+ T cells: 1 in 200,000 cells specific for any given antigen (0.0005% antigen-specific cells) Arstila et al. Science 286, 958 (1999) Blattman et al. J. Exp. Med. 195, 657 (2002) 20.400 spring 2008 3 The immune system: evolved to eliminate infectious disease Katakai et al. JEM, 200, 783-792 (2004) PERIPHERAL TISSUE LYMPH NODE LYMPHATICS 5 2 1 B T 3 B cell T cell BLOOD Dendritic cell Source: Katakai, Tomoya, et al. "Lymph Node Fibroblastic Reticular Cells Construct the Stromal Reticulum via Contact with Lymphocytes." Journal of Experimental Medicine 200, no. 6 (2004). 4 B cell activation Diagram of antigen recognition, B cell proliferation, and Ig secretion and isotype switching has been removed due to copyright restrictions. Lecture 20 Spring 2006 (Abbas) 13 Biology of dendritic cells in T cell activation Classical pathways of antigen processing and presentation: CD8+ T cells classical Class I antigen loading pathway CD4+ T cells Class II antigen loading pathway exogenous ANTIGEN Lecture 20 Spring 2006 11 interactions in the lymph node Three electron micrographs of T cells and dendritic cells interacting with reticular fibers have been removed due to copyright restrictions. T-cell activation infected cell or tumor cell (1) antigen recognition target cell T peptide-MHC T cell receptor (TCR) Two electron micrographs removed due to copyright restrictions. T-cell (2) immunological synapse (IS) formation T T 5! Grakoui et al. Science 285, 221 (1999) Monks et al. Nature 395, 82-86 (1998) The immune system: a distributed network 11 lymphocyte trafficking is “addressed” by combinations of adhesion molecules and chemokine signals Diagram of lymphocyte trafficking removed due to copyright restrictions. There are four steps: rolling adhesion, tight binding, diapedesis, and migration. See Figure 2-44 part 3 of 3, Janeway, Charles, et al. Immunobiology. 6th ed. New York: Garland Science, 2005. ISBN: 9780815341017. 12 of TCR, costimulatory, chemokine, and cytokine signals that remain largely unexplored (36, 148). Moreover, the complete Authors’ addresses Summary: Directed cellular migrations underlie i sequencing has and revealed together specify unique microenvironments within cutaneous Daniel J. Campbell1,2, Chang H. Kim3, of the human and mouse ization.genomes Chemokines their receptors (along 1,2 , Eugene C. Butcher many ‘orphan’ chemokine receptors whose ligands, expresmolecules) are central to these migrations, targ and intestinal lymphoid tissues and direct expression of appro1 Laboratory of Immunology and Vascular to tissues and microenvironm poorlyleukocytes understood (149). priate combinations of adhesion and chemoattractant receptors. sion patterns, and function remainmature Biology, Department of Pathology, Stanford differentiation and function. The chemokine CX addition, very the signaling pathways Understanding these complex pathways of University lymphocyte School of In Medicine, Stanford, CA, little USA. is known about CXCR4 play a central role in the migration of he 2 and transcription factors that regulate verychemokine complexreceptors tissue- are transiently ex Center for Molecular Biology and Medicine, differentiation clearly requires further research, willTheundoubtand the several chemokines and chemotaxis: how the immune system gets around Veterans Affairs Palo Alto Health Care System, Palo Alto, CA, USA. 3 Laboratory of Immunology and Hematopoiesis, Department of Veterinary Pathobiology and Purdue Cancer Center, Purdue University, West Lafayette, IN, USA. Correspondence to: Daniel J. Campbell Benaroya Research Institute at Virginia Mason 1201 Ninth Avenue Figure "Known and proposed functions for homeostatic chemokines and Seattle, WA 98101–2795 their receptors in lymphocyte development, trafficking, and function" USA has been removed due to copyright restrictions. Tel.: þ1 206 583 6525 Fax: þ1 206 223 7543 E-mail: campbell@benaroyaresearch.org stages of B- and T-cell development. In the periphe T cells utilize the receptors CCR7, CXCR4, and through specialized microenvironments within th tissues, while effector and memory lymphocyte patterns of adhesion molecules and chemokine them to function within microenvironments and inaccessible to naı̈ve cells. Here, we summarize t and their receptors in the spatial organization of t consider the implications for immune function. The mechanisms of leukocyte homing The mammalian immune system is design tion while maintaining self-tolerance and mediated pathology. In order to accompl Acknowledgements cells and tissues of the immune system are We thank Tracy Staton and Gudrun Debes for comments on the manuscript. E.C.B. is supto ensure the proper development, activati ported by grants from the NIH and the diverse leukocyte populations. Tissue- and Veterans administration. D.J.C. is the recipient selective leukocyte homing is the basis fo of a postdoctoral fellowship from the Arthritis Foundation. C.H.K. is supported by grants and leukocyte responses to the chemoa from The Eli and Edythe L. Broad Foundation (chemokines) function prominently in t and Leukemia and Lymphoma Society. ways (1). Leukocyte homing from blood to tissues specialized post-capillary venules through a Fig. 2. Known and proposed functions for homeostatic chemokines and their receptors in lymphocyte development, trafficking, andthe function. interactions with vascular endothelium Receptors acting predominantly on B cells (red), T cells (green) or both (blue) are color coded. LN, lymph node; PP, Peyer’s patch. sively in 2, 3). Initial interactions mediate Immunological Reviews 2003 sion molecules, either selectins or low-affin Campbell, et al. Vol. 195: 58–71 13 66 ImmunologicalButcher Reviews 195/2003 the leukocytes to roll along the endoth Printed in Denmark. All rights reserved presented on the endothelial surface trigg Copyright ! Blackwell Munksgaard 2003 and avidity activation and mediate le Immunological Reviews the endothelial wall (4–6). Subsequen 0105-2896 unique combinations of chemokine/adhesion molecule “addressins” lead immune cells to different tissue sites, both in “healthy” immune responses and in inflammatory diseases: Reprinted by permission from Macmillan Publishers Ltd: Nature Immunology. Source: Luster, Andrew D., Ronen Alon, and Ulrich H. von Andrian. "Immune cell migration in inflammation: present and future therapeutic targets." Nature Immunology 6 (2005). © 2005. 14 Luster, Alon, and von Andrian Nat. Immunol. 6 1182 (2005) Diagram of chemotaxis directing cell migration has been removed due to copyright restrictions. chemokines also direct cells to their appropriate locations within tissues Source: Okada, Takaharu et al. "Antigen-Engaged B Cells Undergo Chemotaxis toward the T Zone and Form Motile Conjugates with Helper T Cells." PLoS Biology 3, no. 6 (2005). Figure of T cell/B cell removed due to copyright restrictions. Katakai et al. JEM, 200, 783-792 (2004) Steps in the immune response to infection 17 innate immune sentinels HIV illustration removed due to copyright restrictions. 18 http://www.northwestern.edu/newscenter/images/2008/12/hiv-illustration.jpg infection site in peripheral tissue (1) chemoattraction of dendritic cells/DC precursors to sites of infection/ inflammation pathogen resident DCs in skin (3) DCs travel to lymph nodes APC Electron micrograph of resident dendritic cells in skin has been removed due to copyright restrictions. peptide-MHC (2) dendritic cells collect antigen and become activated Dendritic cells and initiation of adaptive immune responses (4) DCs activate lymphocytes by cell-cell contact 5 5 Randolph, Angeli, and Swartz Nat. Rev. Immunol. 5 617 (2005) Reprinted by permission from Macmillan Publishers Ltd: Nature Reviews Immunology. Source: Randolph, Gwendalyn J. et al. "Dendritic-cell trafficking to lymph nodes through lymphatic vessels." Nature Reviews Immunology 5 (2005). © 2005. pattern recognition by the immune system infection site in peripheral tissue dsRNA Diagram of gram-negative bacterium removed due to copyright restrictions. LPS ssDNA, ssRNA flagellin Tri-acyl and diacyl lipopeptides Kawai and Akira, Curr. Opin. Immunol. 17 338-344 (2005) www.invivogen.com Courtesy of Elsevier, Inc., http://www.sciencedirect.com. Used with permission. Source: Kawai, Taro and Shizuo Akira. "Pathogen recognition with Toll-like receptors." Current Opinion in Immunology 17, no. 4 (2005). PAMP recognition of microbes by dendritic cells Immune cells integrate many signals to ‘fingerprint’ pathogens: dsRNA LPS Diagram from Science magazine removed due to copyright restrictions. ssDNA, ssRNA flagellin Tri-acyl and di-acyl lipopeptides Huang et al., Science 294 3870 (2001) Kawai and Akira, Curr. Opin. Immunol. 17 338-344 (2005) Courtesy of Elsevier, Inc., http://www.sciencedirect.com. Used with permission. Source: Kawai, Taro and Shizuo Akira. "Pathogen recognition with Toll-like receptors." Current Opinion in Immunology 17, no. 4 (2005). Lecture 20 Spring 2006 7 TLR signaling is likely one of the earliest steps in the host response to infection 22 what we typically think of as inflammation: recruitment of innate and adaptive immune cells to peripheral tissue sites: The New England Journal of Medicine inflammatory agent applied to epithelium: Figure removed due to copyright restrictions. See Figure 2 from Luster, Andrew D. "Chemokines — Chemotactic Cytokines That Mediate Inflammation." New England Journal of Medicine 338 (2006). Le Borgne, Dubois et al. Immunity 24 191-201 (2006) Courtesy of Elsevier, Inc., http://www.sciencedirect.com. Used with permission. Source: Le Borgne, Marie, et al. " Figure 2. Chemokine Regulation of Leukocyte Movement. Dendritic Cells Rapidly Recruited into Epithelial Tissues via Chemokines are secreted at sites of inflammation and infection by resident tissue cells, resident and recruited leukocytes, and cyCCR6/CCL20 Are Responsible for CD8+ T Cell Crosspriming tokine-activated endothelial cells. Chemokines are locally retained on matrix and cell-surface heparan sulfate proteoglycans, establishing a chemokine concentration gradient surrounding the inflammatory stimulus, as well as on the surface of the overlying In Vivo." Immunity 24 (2006). endothelium. Leukocytes rolling on the endothelium in a selectin-mediated process are brought into contact with chemokines retained on cell-surface heparan sulfate proteoglycans. Chemokine signaling activates leukocyte integrins, leading to firm adherence and extravasation. The recruited leukocytes are activated by local proinflammatory cytokines and may become desensitized to fur- Diagram explaining process of inflammation removed due to copyright restrictions. Robbins and Cotran Pathol. Basis of Disease 7th ed. 24 recruitment of DCs: chemotaxis into inflammation sites chemoattractants bring monocytes and DCs to sites of infection Two electron micrograph images removed due to copyright restrictions. infection site in peripheral tissue (1) chemoattraction of dendritic cells/DC precursors to sites of infection/ inflammation pathogen resident DCs in skin (3) DCs travel to lymph nodes APC peptide-MHC Reprinted by permission from Macmillan Publishers Ltd: Nature Reviews Immunology. Source: Randolph, Gwendalyn J. et al. "Dendritic-cell trafficking to lymph nodes through lymphatic vessels." Nature Reviews Immunology 5 (2005). © 2005. (2) dendritic cells collect antigen and become activated (4) DCs activate lymphocytes by cell-cell contact 5 Randolph, Angeli, and Swartz Nat. Rev. Immunol. 5 617 (2005) Antigen is one of (at least) two signals that must be delivered by a vaccine +DC ACTIVATION +ANTIGEN •T CELLS TOLERIZED •NO T CELL ACTIVATION +ANTIGEN +DC ACTIVATION Signal 1 - antigen Signal 2 - costimulation Signal 3 - cytokines •MAXIMAL T CELL PROLIFERATION •GENERATION OF FULL EFFECTOR FUNCTIONS •GENERATION OF MEMORY T CELLS Lecture 20 Spring 2006 12 (1) antigen carried to lymph nodes: ORCHESTRATION OF THE PRIMARY IMMUNE RESPONSE (3) Antigen-specific T and B cells meet at follicular border: (2a) B cells encounter antigen, likely in follicles: 3.5 min antigen B cells T cells Figure from New England Journal of Medicine removed due to copyright restrictions. See Figure 3 from Ada, Gordon. "Advances in Immunology: Vaccines and Vaccination." New England Journal of Medicine 345 (2001). (Pape et al. Immunity 26 491 (2007)) (2b) T cells encounter antigen in T zones: pMHC B cells Source: Okada, Takaharu et al. "Antigen-Engaged B Cells Undergo Chemotaxis toward the T Zone and Form Motile Conjugates with Helper T Cells." PLoS Biology 3, no. 6 (2005). Courtesy of Elsevier, Inc., http://www.sciencedirect.com . Used with permission. Source: Itano, Andrea A., et al. "Distinct Dendritic Cell Populations Sequentially Present Antigen to CD4 T Cells and Stimulate Different Aspects of Cell-Mediated Immunity." Immunity 19, no. 1 (2003). Katakai et al. JEM, 200, 783-792 (2004) PERIPHERAL TISSUE LYMPH NODE LYMPHATICS 5 1 2 B T 3 B cell T cell BLOOD Dendritic cell Source: Katakai, Tomoya, et al. "Lymph Node Fibroblastic Reticular Cells Construct the Stromal Reticulum via Contact with Lymphocytes." Journal of Experimental Medicine 200, no. 6 (2004). 4 these data suggest that lymphocytes integrate a complex array of TCR, costimulatory, chemokine, and cytokine signals that together specify unique microenvironments within cutaneous and intestinal lymphoid tissues and direct expression of appropriate combinations of adhesion and chemoattractant receptors. Understanding pathways of lymphocyte The New England these Journalcomplex of Medicine differentiation clearly requires further research, will undoubt- growth/survival factors, but these aspects of their biology Eugene C. Butcher1,2, remain largely unexplored (36, 148).1Laboratory Moreover,ofthe complete Immunology and Vascular sequencing of the human and mouse genomes has revealed Biology, Department of Pathology, Stanford many ‘orphan’ chemokine receptorsUniversity whose School ligands, expres- Stanford, CA, USA. of Medicine, 2 The Center for Molecular Biology and Medicine, sion patterns, and function remain poorly understood (149). Veterans Affairs Palo Alto Health In addition, very little is known about the signaling pathways Care System, Palo Alto, CA, USA. and transcription factors that regulate3 the very complex tissue- Adaptive immune cell effectors home back to infection site: Development in primary lymphoid tissues Lymphocyte recirculation and immune surveillance Laboratory of Immunology and Hematopoiesis, Department of Veterinary Pathobiology and Purdue Cancer Center, Purdue University, West Microenvironmental positioning Lafayette, and non-lymphoid tissue homingIN, USA. Correspondence to: Daniel J. Campbell Benaroya Research Institute at Virginia Mason 1201 Ninth Avenue Seattle, WA 98101–2795 USA Tel.: þ1 206 583 6525 Fax: þ1 206 223 7543 E-mail: campbell@benaroyaresearch.org Thymus CXCR4/CXCL12 CCR9/CCL25 CCR4/CCL17/CCL22 CCR7/CCL19/CC21 Figure removed due to copyright restrictions. LN/spleen See Figure 2 from Luster, Andrew D. "Chemokines — Chemotactic CCR7/CCL19/CCL21 Cytokines That Mediate Inflammation." New England Journal of CXCR4/CXCL12 Medicine 338 (2006). Bone marrow CXCR4/CXCL12 CCR9/CCL25 PP CCR7/CCL19/CCL21 CXCR4/CXCL12 CXCR5/CXCL13 Figure removed due to copyright restrictions. Acknowledgements See Campbell, Daniel J., Chang H. Kim, and We thank Tracy Staton and Gudrun Debes for Eugene C. Butcher. "Chemokines in on thethe systemic comments manuscript. E.C.B. is suporganization of immunity." ported Immunological 195,and the by grants Reviews from the NIH no. 1 (2003). Veterans administration. D.J.C. is the recipient of a postdoctoral fellowship from the Arthritis Foundation. C.H.K. is supported by grants from The Eli and Edythe L. Broad Foundation and Leukemia and Lymphoma Society. Fig. 2. Known and proposed functions for homeostatic chemokines and their receptors in lymphocyte development, trafficking, and function. Receptors acting predominantly on B cells (red), T cells (green) or both (blue) are color coded. LN, lymph node; PP, Peyer’s patch. 66 Immunological Reviews 195/2003 Figure 2. Chemokine Regulation of Leukocyte Movement. Chemokines are secreted at sites of inflammation and infection by resident tissue cells, resident and recruited leukocytes, and cytokine-activated endothelial cells. Chemokines are locally retained on matrix and cell-surface heparan sulfate proteoglycans, establishing a chemokine concentration gradient surrounding the inflammatory stimulus, as well as on the surface of the overlying endothelium. Leukocytes rolling on the endothelium in a selectin-mediated process are brought into contact with chemokines retained on cell-surface heparan sulfate proteoglycans. Chemokine signaling activates leukocyte integrins, leading to firm adherence and extravasation. The recruited leukocytes are activated by local proinflammatory cytokines and may become desensitized to further chemokine signaling because of high local concentrations of chemokines. The Duffy antigen receptor for chemokines (DARC), a nonsignaling erythrocyte chemokine receptor, functions as a sink, removing chemokines from the circulation and thus helping to maintain a tissue–bloodstream chemokine gradient. Copyright ! Blackwell Munksgaard 2003 Immunological Reviews 0105-2896 58 molecular warfare involved in clearing infections: Figure of chemical processes within a blood vessel removed due to copyright restrictions. 31 regulatory T-cells play a key role in preventing activation of immune responses in the steady state and are thought to help regulate the close of immune responses in infection: IFN IL-10 IL-4 and IL-10 TH2 cell IL-10 and IFN TH1 cell Conversion DC IDO ion lat du CTLA4, TGF, IL-10, cAMP, adenosine and granzyme B M od ula tio n du lat ion Mo od M n tio a l u Microbial products Modulation Mo WS Turning off the immune response as infection is cleared: Natural TReg cell IL-10 Modulation Induction IL-10 and TR1 cell TGF Induction IDO Figure 1 | Regulatory T cells during infection. Various populations of regulatory T cellshave been shown to have a Reviews | Immunology role during infection. T helper 1 (TH1)-cell and TH2-cell populations can regulate each other viaNature their production of + + + cytokines. Naturally occurring forkhead box P3 (FOXP3)Reprinted CD4 CD25 regulatory cells (natural TReg cells) can limit TH1by permission fromTMacmillan Publishers Ltd: Nature Reviews and TH2-cell responses either indirectly by modulating antigen-presenting cell (APC) function or directly byand cell–cell Immunology. Source: Belkaid, Yasmine. "Regulatory T cells infection: Belkaid Nat. Rev Immunol. 875 (2007)involve the release contact. Indirect regulatory 7 mechanisms of transforming growth factor-B (TGFB, interleukin-10 a dangerous necessity." Nature Reviews Immunology 7 (2007). © 2007. (IL-10) or adenosine, or the induction of the tryptophan-degrading enzyme indoleamine 2,3-dioxygenase (IDO) by APCs recognizing cytotoxic T-lymphocyte antigen 4 (CTLA4) on natural TReg cells. Direct regulation of effector T cells includes 32 anisms that ue hypoxic t for develhat have an roving the issues. We ave evolved anism that gical studies fects on the in different es. unological mediators e factor 1α d adenosine 3 . Turning off the Rimmune E V I E W S response as infection is cleared: REVIEWS role of hypoxia/adenosine receptor signaling Hypoxia [O2]low Inspired air Po2 = 21 kPa (159 mm Hg) A2AR AMP Lungs Po2 = 20 kPa (150 mm Hg) [O2]low NT5 AK Cyclic AMP Adenosine HIF1α A2BR [Adenosine]hi Immune cell Circulation Po2 = 5–13 kPa (40 –100 mm Hg) T cell Inhibited production of interferon-γ Inhibited T-cell response s ny comparttissues14–19. sion of the g a hypoxic med tissues, ecause the rect in vivo poxic enviponses was l-mediated s a marked ygenation. of the anti- TCR Tissues Po2 = 0.5–2.5 kPa (4–20 mm Hg) (hypoxia) Figure 1 | Low oxygen concentration in tissue environments. Oxygen is inspired from the air; the oxygen Sitkovsky Rev Immunol. 712 (2005) in air is 21 kPa (159 mm 5 Hg), which is a tension (Po2)Nat. concentration of 21%. When warmed air arrives in the lungs, it loses some pressure: Po2 = 20 kPa (150 mm Hg). The Reprinted bycontinues permission Macmillan oxygen pressure to from decrease in the Publishers circulation:Ltd: Nature Immunology. 13 kPa (100 mm Hg) Source: in alveoliSitkovsky, and 5 kPaMichail (40 mmand Hg)Dmitriy Po2 =Reviews Lukashev. "Regulation of immune cells by local-tissue in veins that reach the heart. The oxygen pressure in the oxygen tension: HIF1and adenosine receptors." Nature tissues then decreases with increasing distance fromReviews Immunology (2005). 2005. = 0.5©–2.5 kPa (4–20 mm Hg). the blood vessels:5Po 2 Figure 3 | The hypothesis: role of hypoxia in local tissues in the regulation of T cells in inflamed and hypoxic areas. We think that excessive collateral immune damage to the local-tissue microcirculation, and therefore to the oxygen (O2) supply, creates deepening tissue hypoxia, which functions as a signal to stop immune responses. Hypoxia, in turn, inhibits adenosine kinase (AK) and upregulates 5′-nucleotidase (NT5) activity, which results in the accumulation of extracellular adenosine. Adenosine signals through the immunosuppressive adenosine receptors A2AR and/or A2BR at the surface of surrounding activated T cells, and it downregulates T-cellreceptor (TCR)-mediated responses in a delayed negativefeedback manner. The regulatory effects of hypoxia-inducible factor 1α (HIF1α) on T cells remain to be directly established, but it is expected that the increased expression of HIF1 in response to hypoxia will also be inhibitory. HIF1 is a het consists of the con and the oxygenIn the presence o hydroxylation of and 564 of HIF1α, domain, is catalyse containing protein a contribution fro HIF1α is subseque degradation that tumour-suppresso radation of HIF1α low levels under n hypoxic condition PHD3 are, in turn tia homologue 1A degradation77 (FIG mechanism ensu under hypoxic co moxic conditions and activity of the transcription of m ing those encodin poietin, adrenom vascular endothel In addition, th be negatively regu gine residue at po dependent hydrox which inhibits th its co-activators, p Induction of immunological memory (the basis of vaccination) Figure removed due to copyright restrictions. See Kaech, Susan M. and Rafi Ahmed. "CD8 T Cells Remember with a Little Help." Science 300, no. 5617 (2003). MIT OpenCourseWare http://ocw.mit.edu 20.380J / 5.22J Biological Engineering Design Spring 2010 For information about citing these materials or our Terms of Use, visit: http://ocw.mit.edu/terms.