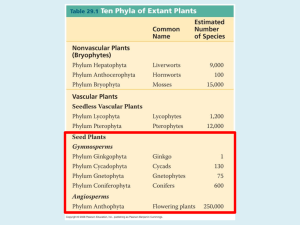
Diversity and evolution of major groups of land plants Robin Allaby (
advertisement

Diversity and evolution of major groups of land plants Robin Allaby (http://www2.warwick.ac.uk/fac/sci/whri/research/archaeobotany/) Notes available from website Notes available from website Overview • • Origin of land plants Evolutionary history of land plants – Major morphological innovations: alternation of generations cuticles and stomata vascular tissues heterospory seeds leaves flowers • • Resultant phylogenetic tree of plants Some evolutionary trends convergence polyploidy genome expansion Invasion of land was really an invasion of the air • • dessication and support are the principal problems adaptation to dessication requires : a cuticle (and consequentially stomata) spores and seeds (ultimately) vascular tissue (when plants are above a certain size) What is a land plant? • Any photosynthetic eukaryote that can survive and sexually reproduce on land What is a land plant? • • Any photosynthetic eukaryote that can survive and sexually reproduce on land All land plants are embryophytes (= embryo bearing plants) Embryophytes • Defined by a true alternation of generations with multicellular diploid and haploid phases, and the two phases remain physically connected. Gametophyte (haploid) stage Sporophyte produces sporangia which make haploid spores Gametophyte produces haploid gametes, which unite in the archegonium of the gametophyte Sporophyte (diploid) stage A brief history of time 545 495 Cambrian 440 Ordovician 417 Silurian 354 Devonian 292 Carboniferous 251 Permian 202 Triassic Jurassic 142 65 Cretaceous 24 Palaeogene 1.8 Neogene Q Evidence of first land plants: 480-360 Mya • • • cuticles & stomata archegonium & sporopollenin walled spores vascular systems (417) (440) 1 cm bar 458 (495) Circa 415 Mya Cooksonia paranensis (Gerrienne et al 2006) Vascular tissues: evolution of xylem and phloem embryophytes better adapted for moist environments than tracheophytes amplification of gametophyte poikilohydric better adapted for dry environments than bryophytes amplification of one stage must reduce the other amplification of sporophyte homoihydric non vascular vascular (bryophytes) (tracheophytes) mosses, liverworts and hornworts ferns, lycopods, horsetails, and seed plants See Proctor 2007 Heterospory unisexual gametophytes, manifest as heterosporous 397 - 391 Mya (Eifelian) phyletic trend for taller sporophytes, bisexual gametophytes, homosporous plants that release spores = pteridophytes (fern like - includes lycopods (L), Horsetails (H) and Ferns (F)) Heterospory homospory heterospory Heterospory • Seems odd - reduces the chances of fertilization by separating egg and sperm. Cannot be good in a harsh environment, this is a cost. • Once separated, makes sense to increase energy investment in the “female” gametophyte which must support the sporophyte, and maximize chances of successful fertilization by making male spore numerous, (and consequently small). • Gives rise to out-crossing. Perhaps this is the advantage (?). • That it is an advantage is proven by the convergence on the habit - possibly as many as 11 times! (Bateman & DiMichele 1994). The logical progression of heterospory e.g. Barinophyton citrulliforme e.g. Chaleuria cirrosa e.g. Cystosporites devonicus e.g. Archaeopteris & Selaginella increasing investment in megaspore causes reduction in megaspore number Seed habit - the next step after heterospory • • • • • • retain megaspore in megasporangium reduce functional megaspores to 1 retain megagametophyte (elimating requirement for external water for fertilization) modification of megasporangia to receive microspores modification of microspores to enable them to deliver sperm cells to eggs (ie pollen tube) integument develops around megasporangia (later) all seed plants = spermatophytes first seed plants = gymnosperms (naked seeds) Oldest seed plant 385 Mya (Mid Devonian) salpinx megasporangium Integument does not fully enclose ovule Runcaria heinzelinii (Gerrienne et al 2004) Basic Gymnosperm architecture integument + megasporangium uncovered Pollen tubes by early Carboniferous (Rothwell 1972) The late arrival of leaves: 360 Mya!! (end of Devonian) Leaf evolution associated with falling CO2 levels (first plants evolved in a CO2 rich atmosphere) (Beerling 2005) A snapshot of the Carboniferous Lycopod trees (Lepidodendron) Equisetoid trees (Calamites) Angiosperms appear 144 Mya (early Cretaceous) Asteropollis sp. Pollen (Laurales) (Friis et al 2005) Evolution of Angiosperms • • • • • • • • Angio = container - megasporangium (and integuments) enclosed in carpel. Flower structure (a determinate shoot built from leaves). 2 integuments not one (as in gymnosperms). Double fertilization (resulting in triploid endosperm). Xylem structure (vessel members and sieve tubes). Other features to such as endopolyploidy ability (weed technology!), vegetative reproductive ability (weed technology!). Very versatile - numerous floral strategies possible - a single mutation can result in sexual isolation and new species formation. Introduction of animal based pollination strategies. Explosion in Angiosperm speciesGnetales is gymnosperm group closest to flowering plants Angiosperms 220 000 species Bryophytes 22 400 species Pteridophytes 9 000 species Gymnosperms 750 species Fabaceae alone have 14000 species Also only land plant group to reinvade the sea (Zostera sp.) Oldest flower fossils circa 125 Mya. Water lily (Nymphaeales) (Friis et al 2001) The diversification of angiosperms: Darwin’s abominable mystery • • • • The rapid appearance of so many species of angiosperm was a problem for Darwin’s theory In his version of events, evolution proceeds gradually, selecting minute changes Saltation was an opposing view point – gives more emphasis to mutation (internally driven) than Natural Selection (externally driven) Darwin discovered the reason, and founded ‘pollination biology’ Floral evolution – protection to attraction Bisexual flowers Nectar reward, bees, predominate birds, moths, bats wind pollination First pollinators: beetles in small inconspicuous flowers Friis et al 2006 Unisexual flowers predominate Angiosperms diversified because of floral morphology and pollinator co-evolution water lillies pepper (spice) magnolia basal complex: primitive dicots bay laurel peace lillies Based on DNA sequences such as rbcL, matK, nadh, atpB, 18S rDNA 30 Mya CO2 levels crashed: C4 metabolism evolved 62 times independently!! 26 in monocots and 36 in eudicots Angiosperm Phylogeny basal monocots asparagus Lillies, daffodils etc yams palms pineapples/air plants reeds and rushes grasses ginger, bird of paradise plants basal buttercups, poppies grapes cactus Docks, rhubarb & sorrels witch hazel, stone crops mistletoe geraniums Oldest tricolpate pollen 120 Mya basal core rosids Euphorbia, willow wood sorrel beans, peas, acacia roses, apples cucumber, melons &pumpkins Oaks, birch, beech evening primrose Cabbages, Arabidopsis cotton, lime trees oranges, lemons dogwood asterids heather, rhododendron, primrose Potato, tomoto, deadly nightshade coffee Mint, basil, rosemary, thyme etc., olives Tricolpate Angiosperms holly carrots, parsley. fennel honeysuckle, elder daisy, asters, thistles, bellflowers Evolutionary Trends: convergence (on tree habit) Evolutionary Trends: convergence (on cactus habit) Evolutionary trends: polyploidy Blanc and Wolfe 2004 Evolutionary trends: genome obesity Leitch et al 2005 Obesity in the Liliales Fritillaria (Liliales) 127 000 Mb Large cells, slow replication - good for bulbs Arabidopsis (Brassicales) 157 Mb Small cells, fast replication good for weed habit Endangered species? Why is there an over-representation of large genomes in the plant Red List? Vinogradov 2003, but read also Cavalier-Smith 2005 Suggested reading Crane PR, Friis EM, Pedersen KR (1995) The origin and early diversification of angiosperms. Nature 374: 27-33. Friis EM, Pederson KR, Crane PR (2005) When the earth started blooming: insights from the fossil record. Current Opinion in Plant Biology 8:5-12. Judd WS, Campbell CS, Kellogg EA, Stevens PF (1999). Plant Systematics: A Phylogenetic Approach. Sinauer Associates, Sunderland, Massachusetts. Kenrick P and Crane PR (1997) The origin and early evolution of plants on land. Nature 389:33-39. Niklas K (1997) The evolutionary biology of plants. University of Chicago Press, Chicago. Other source material Bateman RM and DiMichele WA (1994) Heterospory: The most iterative key innovation in the evolutionary history of the plant kingdom. Biological Reviews of the Cambridge Philosophical Society 69:345-417. Beerling DJ (2005. Leaf Evolution: Gases, Genes and Geochemistry. Annals of Botany 96:345-352. Blanc G, Wolfe KH (2004) Widespread paleopolyploidy in mode plant species inferred from age distributions of duplicate genes. The Plant Cell 16:1667-1678. Cavalier-Smith T (2005) Economy, sped and size matter: evolutionary forces driving nuclear genome miniaturization and expansion. Annals of Botany 95:147-175. Cavalier-Smith T (2004) Only six kingdoms of life. Proceedings of the Royal Society of London Series B. 271:1251-1262. Friis EM, Pederson KR, Crane PR (2001) Fossil evidence of water lilies (Nymphaeales) in the early cretaceous. Nature 410: 357-360. Friis et al 2006 Cretaceous angiosperm flowers: Innovation and evolution in plant reproduction. Palaeogeography, Palaeoclimatology, Palaeoecology 232 (2006) 251–293 Friis et al 2010 Diversity in obscurity: fossil flowers and the early history of angiosperms. Phil. Trans. R. Soc. B 2010 365, 369-382 Gerrienne P, Meyer-Berthaud B, Fairon-Demaret M, Streel M, Steemans P (2004) Runcaria, a Middle Devonian Seed Plant Precursor. Science 306:856-858. Gerrienne P et al. (2006) An exceptional specimen of the early land plant Cooksonia paranensis, and a hypothesis on the life cycle of the earliest eutracheophytes. Review of Palaeontology and Palynology 142:123-130. Leitch IJ, Soltis DE, Soltis PS, Bennett MD (2005) Evolution of DNA amounts across land plants (Embryophyta). Annals of Botany 95:207217. Proctor M (2007) Ferns, evolution, scale and intellectual impedimenta. New Phytologist 176:504-506. Rothwell GW (1972) Evidence of Pollen Tubes in Palaeozoic Pteridosperms. Science 175:772-774. Vinogradov AE (2003) Selfish DNA is maladaptive: evidence from the plant Red List. Trends in Genetics 19:609-614.