Tailoring Enzymes Acting on Carrier Protein-Tethered Substrates in Natural Product Biosynthesis
advertisement

CHAPTER SIXTEEN
Tailoring Enzymes Acting on
Carrier Protein-Tethered
Substrates in Natural Product
Biosynthesis
Shuangjun Lin*, Tingting Huang{, Ben Shen{,{,},1
*The State Key Laboratory of Microbial Metabolism, School of Life Sciences and Biotechnology, Shanghai Jiao
Tong University, Shanghai, PR China
{
Department of Chemistry, The Scripps Research Institute, Jupiter, Florida, USA
{
Department of Molecular Therapeutics, The Scripps Research Institute, Jupiter, Florida, USA
}
Natural Products Library Initiative at TSRI, The Scripps Research Institute, Jupiter, Florida, USA
1
Corresponding author: e-mail address: shenb@scripps.edu
Contents
1. Introduction
2. Methods
2.1 In vitro characterization of SgcC3-catalyzed chlorination of
(S)-b-tyrosyl-SgcC2
2.2 In vitro characterization of SgcC-catalyzed hydroxylation
of (S)-b-3-chloro-tyrosinyl-SgcC2
2.3 Exploitation of SgcC2-tethered (S)-b-tyrosine analogues for structural
diversification
3. Conclusion
Acknowledgment
References
322
331
331
336
338
339
340
340
Abstract
Carrier proteins (CPs) are integral components of fatty acid synthases, polyketide
synthases, and nonribosomal peptide synthetases and play critical roles in the biosynthesis of fatty acids, polyketides, and nonribosomal peptides. An emerging role CPs play
in natural product biosynthesis involves tailoring enzymes that act on CP-tethered substrates. These enzymes provide a new opportunity to engineer natural product diversity
by exploiting CPs to increase substrate promiscuity for the tailoring steps. This chapter
describes protocols for in vitro biochemical characterization of SgcC3 and SgcC that catalyze chlorination and hydroxylation of SgcC2-tethered (S)-b-tyrosine and analogues in
the biosynthesis of the enediyne chromophore of the chromoprotein C-1027. These
protocols are applicable to mechanistic characterization and engineered exploitation
of other tailoring enzymes that act on CP-tethered substrates in natural product
Methods in Enzymology, Volume 516
ISSN 0076-6879
http://dx.doi.org/10.1016/B978-0-12-394291-3.00008-3
#
2012 Elsevier Inc.
All rights reserved.
321
322
Shuangjun Lin et al.
biosynthesis and structural diversification. The ultimate goal is to use the in vitro findings
to guide in vivo engineering of designer natural products.
1. INTRODUCTION
Acyl carrier proteins (ACPs) and peptidyl carrier proteins (PCPs) are
small (10 kDa) proteins, existing as either a discrete protein in a type II multienzyme complex or a distinct domain interspersed among the catalytic domains of a type I multifunctional megasynthase (Marahiel & Essen, 2009;
Mercer & Burkart, 2007; Shen, 2000; Staunton & Weissman, 2001;
Weissman, 2009). While the overall amino acid sequence identity among
the carrier proteins (CPs) is modest, they are characterized by a highly
conserved signature motif of GxxSL/I. The serine residue in this motif is
the site for 40 -phosphopantetheinylation, a posttranslational modification
catalyzed by 40 -phosphopantetheinyl transferases (PPTases) (Lambalot et al.,
1996; Sanchez, Du, Edwards, Toney, & Shen, 2001). PPTases convert the
apo-CPs into the functional holo-CPs by installing the 20 Å-long 40 phosphopantetheine prosthetic group with a free terminal thiol
(Fig. 16.1A). At this thiol, both substrates and the growing intermediates
are tethered as thioesters. While the 40 -phosphopantetheinyl arm facilitates
the delivery of substrates into each of the active sites and channels the
growing intermediates between each of the elongation cycles, the CPs
provide necessary protein–protein recognition among the various
enzymatic partners.
CPs that carry short carboxylic acids or other acyl intermediates
are known as ACPs, which were first characterized from fatty acid synthases (FASs) (Chan & Vogel, 2010; Gago, Diacovich, Arabolaza, Tsai, &
Gramajo, 2011; Mercer & Burkart, 2007). Type I FASs are multifunctional
proteins consisting of domains for individual activities, while type II
FASs are multienzyme complexes consisting of discrete, monofunctional
proteins. ACPs, either as a domain in type I FASs or a discrete protein in
type II FASs, play a pivotal role in fatty acid biosynthesis by tethering the
starter and extender units for condensation and by channeling the growing
acyl intermediates for complete b-ketoreduction (i.e., b-ketoreduction,
dehydration, and enoylreduction) during each cycle of chain elongation
to afford the fully reduced fatty acid as the final product (Fig. 16.1B).
323
Tailoring Enzymes Acting on Carry Protein-Tethered Substrates
4⬘-Phosphopantetheine
A
PPTase
O
OH
CoA
apo-ACP
apo-PCP
ADP
O
O
O
P
O
SH
N
H
SH
holo-ACP
holo-PCP
b-Keto
reduction
(complete)
B
ACP
O
N
OH H
AT
Elongation
ACP
SH
ACP
O
S
O
ACP
R
ACP
S
S
S
O
O
S-Enz
O
O
n
O
O
R
R
R
Fatty acids
b-Keto
reduction
(selective)
C
ACP
AT
Elongation
ACP
SH
ACP
O
S
O
A
S
O
O
S-Enz
O
PCP
Elongation
R
R
PCP
PCP
R1
SH
S
R1
S
PCP
O
NH2
R1
O
R2
S
O
S
NH2
O
N
H
NH
O
OH
Rn+1
H
N
Rn
O
Polyketides
n
NH2
O
Peptides
O
H 2N
R
O
OH
O
PCP
ACP
S
S
R
O
D
ACP
R2
E
A/P-CP
SH
AT/A
A/P-CP
S
O
Tailoring enzymes
A/P-CP
Cyclization
Halogenation
Methylation
Oxidation
Reduction
(see Table 16.1 for examples)
Rs:
Natural products
(see Fig. 16.2 for examples)
S
O
Rs
Groups introduced
by tailoring enzymes
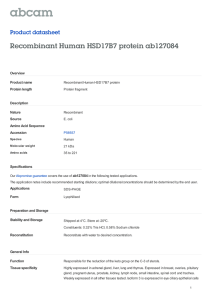
Figure 16.1 Carrier proteins and their roles in fatty acid, polyketide, and nonribosomal
peptide biosynthesis: (A) posttranslational modification of an apo-ACP or apo-PCP into a
holo-ACP or holo-PCP by a PPTase; (B) ACP-mediated substrate activation and intermediate channeling in fatty acid biosynthesis; (C) ACP-mediated substrate activation and
intermediate channeling in polyketide biosynthesis; (D) PCP-mediated substrate activation and intermediate channeling in nonribosomal peptide biosynthesis; and (E) tailoring enzymes acting on ACP- or PCP-tethered substrates in natural product biosynthesis.
See Table 16.1 for specific tailoring enzymes, ACP- or PCP-tethered substrates and their
corresponding products, and the types of modification and Fig. 16.2 for structures of
natural products with moieties modified by tailoring enzymes highlighted in gray.
A, adenylation enzyme; ACP, acyl carrier protein; AT, acyltransferase; PCP, peptidyl
carrier protein; PPTase, 40 -phosphopantetheinyl transferase.
324
Shuangjun Lin et al.
ACPs were subsequently characterized from polyketide synthases (PKSs),
which catalyze the biosynthesis of polyketides, a large family of natural products with profound biological activities (Mercer & Burkart, 2007; Shen, 2000;
Staunton & Weissman, 2001; Weissman, 2009). Following the convention of
FASs, PKSs have also been classified into types I and II according to their
enzyme architectures (Shen, 2003). Thus, similar to FASs, ACPs in type I
PKSs are domains, ACPs in type II PKSs are discrete proteins, and
regardless of their architectural difference, both ACP domains and proteins
tether the acyl CoA substrates for condensation and channel the growing
acyl intermediates during each cycle of chain elongation. However, in
contrast to FASs, the b-ketone groups of the ACP-tethered growing acyl
intermediates in PKSs can undergo no, partial, or full reduction,
depending on the given cycle of elongation, thereby providing a
mechanistic basis to account for the vast structural diversity of polyketide
natural products (Fig. 16.1C).
CPs from nonribosomal peptide synthetases (NRPSs) are known as
PCPs, carrying amino acids or peptidyl intermediates. NRPSs catalyze
the biosynthesis of nonribosomal peptides, another major family of natural
products including many clinically important drugs (Marahiel & Essen,
2009; Mercer & Burkart, 2007). Although PKSs and NRPSs catalyze the
biosynthesis of two distinct classes of natural products from two different
pools of substrates, they apparently use a very similar molecular logic for
substrate activation and intermediate channeling. While the type I and II
nomenclature for FASs and PKSs has not been widely accepted to classify
NRPSs, both multifunctional NRPSs with distinct domains and discrete
NRPSs with largely monofunctions are known. In a mechanism
analogous to FASs and PKSs, NRPSs use PCPs to tether the amino acid
substrates for condensation and channel the growing peptidyl
intermediates during each cycle of chain elongation (Fig. 16.1D). These
striking structural and mechanistic similarities between PKSs and NRPSs
have inspired the discovery and characterization of natural NRPS–PKS
megasynthases for the biosynthesis of hybrid peptide–polyketide natural
products and the construction of engineered hybrid NRPS–PKS systems
to further expand the size and diversity of natural product libraries
(Du et al., 2001; Fischbach & Walsh, 2006, 2010).
CP-dependent PKSs and NRPSs catalyze the assembly of a myriad of
polyketide, peptide, and hybrid polyketide–peptide backbones from a vast
array of short carboxylic acids and amino acids. The nascent scaffolds are
often heavily modified by the coordinated action of specialized enzymes,
Tailoring Enzymes Acting on Carry Protein-Tethered Substrates
325
known as tailoring enzymes, to further imbue structural and functional
diversity. While tailoring enzymes that act during chain elongation, that
is, with the growing intermediates still tethered to specific ACPs or PCPs,
are known, most tailoring enzymes act on the peptide, polyketide, or hybrid
peptide–polyketide intermediates after they are released from the PKS or
NRPS megasynthases as free substrates (Fischbach & Walsh, 2010; Walsh
et al., 2001).
A subset of tailoring enzymes is emerging that specifically act on CPtethered substrates; the corresponding free substrates are not recognized.
This strategy is most commonly associated with biosynthesis of unusual
building blocks incorporated into many polyketide and nonribosomal peptide natural products. Modifications catalyzed by tailoring enzymes acting
on both ACP- and PCP-tethered substrates are known, including cyclization, halogenation, methylation, oxidation (dehydrogenation, epoxidation,
and hydroxylation), and reduction (Fig. 16.1E). Table 16.1 summarizes the
tailoring enzymes known to date that have been biochemically characterized
and act on CP-tethered substrates in natural product biosynthesis (Fig. 16.2).
Tailoring enzymes that act on CP-tethered substrates therefore represent a
new molecular logic for natural product biosynthesis. The tethering of precursors to CPs ensures that the resultant building blocks will be sequestered
from endogenous metabolite pools and efficiently incorporated into the final
natural products.
The enediyne chromophore of the C-1027 chromoprotein, one of the
most potent antitumor antibiotics known to date, features a highly modified
b-amino acid moiety (Fig. 16.3; Van Lanen & Shen, 2008). The gene cluster
for C-1027 biosynthesis was cloned and sequenced from Streptomyces
globisporus (Liu, Christenson, Standage, & Shen, 2002). Bioinformatics analysis of the genes within the C-1027 biosynthetic gene cluster predicted, and
biochemical characterizations subsequently confirmed, that the biosynthesis
of the b-amino acid moiety from the a-tyrosine precursor involved tailoring
enzymes that act on PCP-tethered substrates (Van Lanen et al., 2005). Thus,
a-tyrosine is first converted by the SgcC4 aminomutase to (S)-b-tyrosine
(Christenson, Liu, Toney, & Shen, 2003; Christenson, Wu, Spies, Shen, &
Toney, 2003), which is then tethered by the SgcC1 adenylation enzyme to
the SgcC2 PCP (Van Lanen, Lin, Dorrestein, Kelleher, & Shen, 2006).
Sequential chlorination and hydroxylation of the SgcC2-tethered (S)-btyrosine by the SgcC3 halogenase (Lin et al., 2007) and SgcC
monooxygenase (Lin et al., 2008), respectively, affords the fully modified
b-tyrosine building block, which, still tethered to the SgcC2 PCP, is
Table 16.1 Tailoring enzymes acting on carrier protein-tethered substrates that have been biochemically characterized from natural product
biosynthetic pathways.
Tailoring Carrier
Products
Reference
Natural productsa enzyme protein Type of reaction Substrates
L-aminobutanoic
acid
g,g-Dichloro-Laminobutanoic acid
Ueki et al. (2006)
PCPBarA Chlorination
L-leucine
d-trichloro-L-leucine
Galonić,
Vaillancourt, and
Walsh (2006)
Flatt et al. (2006)
SgcC3
SgcC2
Chlorination
(S)-b-tyrosine
(S)-3-chloro-b-tyrosine
Lin, Van Lanen, and
Shen (2007)
C-1027
SgcC
SgcC2
Hydroxylation
(S)-3-chloro-btyrosine
(S)-3-chloro-5-hydroxy-btyrosine
Lin, Van Lanen, and
Shen (2008)
CDA
HxcO
ACP
Dehydrogenation hexanoic acid
hex-2-enoic acid
Kopp, Linne,
Oberthür, and
Marahiel (2008)
CDA
HcmO
ACP
Oxidation
2,3-Epoxyhexanoic acid
Kopp et al. (2008)
Chloramphenicol
CmlA
PCPCmlP Hydroxylation
Chlorobiocin
CloN3
CloN5
Armentomycin
CytC3
CytC2
Barbamide
BarB1
BarB2
C-1027
Chlorination
hex-2-enoic acid
L-pb-Hydroxy-L-paminophenylalanine aminophenylalanine
Dehydrogenation L-proline
Pyrrole-2-carboxylic acid
Makris, Chakrabarti,
Münck, and
Lipscomb (2010)
Garneau-Tsodikova,
Dorrestein, Kelleher,
and Walsh (2005)
Coronatine
CmaB
CmaD
Chlorination
L-allo-isoleucine
g-Chloro-L-allo-isoleucine
Coronatine
CmaC
CmaD
Cyclization
g-Chloro-L-alloisoleucine
Vaillancourt, et al.
(1S,2S)-1-amino-2ethylcyclopropanecarboxylic (2005)
acid
Dapdiamide
DdaC
PCPDpaD Epoxidation
Nb-fumaramoyl-L- Nb-epoxysuccinamoyl-L2,32,3-diaminopropionate
diaminopropionate
Hollenhorst et al.
(2010)
FK506
TcsC
ACPTcsA Reduction/
carboxylation
E-pent-2-enoic
acid
2-Propylmalonic acid
Mo et al. (2011)
Kutzneride
KtzD
KtzC
Chlorination
L-isoleucine
g-Chloro-L-isoleucine
Neumann and Walsh
(2008)
Kutzneride
KtzA
KtzC
Cyclization
g-Chloro-Lisoleucine
Neumann and Walsh
(1S,2R)-1-amino-2ethylcyclopropanecarboxylic (2008)
acid
Kutzneride
KthP
KtzC
Chlorination
Piperazate
(3S, 5S)-5-chloropiperazate Jiang et al. (2011)
Kutzneride
KtzO
PCPKtzH Hydroxylation
L-glutamic
acid
Vaillancourt, Yeh,
Vosburg, O’Connor,
and Walsh (2005)
L-threo-b-hydroxy-glutamic Strieker, Nolan,
acid
Walsh, and Marahiel
(2009)
Continued
Table 16.1 Tailoring enzymes acting on carrier protein-tethered substrates that have been biochemically characterized from natural
product biosynthetic pathways.—cont'd
Tailoring Carrier
Natural products
enzyme protein Type of reaction Substrates
Products
Reference
Kutzneride
KtzP
PCPKtzH Hydroxylation
L-glutamic
Nikkomycin
NikQ
PCPNikP1 Hydroxylation
Novobiocin
NovI
PCPNovH Hydroxylation
Novobiocin
acid
L-erythro-b-hydroxyglutamic acid
Strieker et al. (2009)
Histidine
b-Hydroxy-histidine
Chen, Hubbard,
O’Connor, and
Walsh (2002)
L-tyrosine
b-Hydroxy-L-tyrosine
Chen and Walsh
(2001)
NovJ/K PCPNovH Oxidation
b-OH-L-tyrosine
b-Ketotyrosine
Pacholec, Hillson,
and Walsh (2005)
Pacidamycin
PacV
PCPPacP Methylation
L-2,3diaminobutyrate
L-3-N-methyl-2,3diamniobutyrate
Zhang et al. (2011)
Pyochelin
PchG
PCPPchF Reduction
Hydroxyphenylbisthiazolinic acid
Des-N-methyl-pyochelinic
acid
Reimmann et al.
(2001)
Pyoluteorin
PltA
PltL
Chlorination
Pyrrole-2carboxylic acid
4,5-Dichloropyrrole-2carboxylic acid
Dorrestein, Yeh,
Garneau-Tsodikova,
Kelleher, and Walsh
(2005)
Pyoluteorin
PltE
PltL
Dehydrogenation L-proline
Pyrrole-2-carboxylic acid
Thomas, Burkart,
and Walsh (2002)
Sibiromycin
SibG
PCPSibE Hydroxylation
3-hydroxy-4methylanthranilic
acid
3,5-dihydroxy-4methylanthranilic acid.
Giessen, Kraas, and
Marahiel (2011)
Syringomycin E
SyrB2
PCPSyrB1 Chlorination
L-threonine
g-Chloro-L-threonine
Vaillancourt, Yin,
and Walsh (2005),
Blasiak, Vaillancourt,
Walsh, and Drennan
(2006)
Syringomycin E
SyrP
PCP8SyrE Hydroxylation
L-aspartic
L-threo-b-hydroxy-aspartic
acid
Singh, Fortin,
Koglin, and Walsh
(2008)
pyrrole-2-carboxylic acid
Thomas et al. (2002)
(R)-b-hydroxy-tyrosine
Cryle, Meinhart, and
Schlichting (2010)
Undecylprodigiosin RedW
ORF9
Vancomycin
PCPBpsD Hydroxylation
a
OxyD
acid
Dehydrogenation L-proline
(R)-tyrosine
See Fig. 16.2 for structures of the natural products with moieties (highlighted in gray) that were modified by the tailoring enzymes acting on carrier protein-tethered
substrates.
330
Shuangjun Lin et al.
NH2
N
Armentomycin
(dichloroaminobutanoic acid)
O
S
O
N
H
HO
O
NH
N
H
O
C-1027
Chlorobiocin (R = Cl)
Novobiocin (R = CH3)
HH
N
CO2H
O
O
O
O
N
H
O
OH
O
O
Pacidamycin 1
N
H
OH O
N
O
HO
H
N
H19C9
OH
O
O
HO
Sibiromycin
O
O
O
HO2C
H
N
O
OH
Cl
OMe
N
H
N
N
H
Undecylprodigiosin
C11H23
O
N
H
HN
OH NH
2 HN
O
H
N
N
H
O
Syringomycin E
Cl
N
H
Pyoluteoin
NH2
O
O
R
Cl
OH
CO2H
OH O
H
N
O
S
N
Pyochelin
HO
H
N
O
H OH
Nikkomycin (I, R = Glu)
Nikkomycin (X, R = OH)
S
N
HN
H
N
H2N
NH2
N
H
OH
O
Cl
OH
N
O
HO
N NH
O
O
N
N
NH
HO
Kutzneride 2 (3S)
Kutzneride 8 (3R)
O
N
N HO
H
O
H O
O
OH O H
N
HN
O O
N
N
O
O
H OH
O
O O
3 HN
Cl
O
HN
Cl
H
N
OH
O
OH
HO
O
O
O
H
N
O
Dapdiiamide E
OH
HN
O
NH2
H
N
O
N
H
O
FK506
O
CDA
H2N
O
Coronatine
O
N
N
H
O
O
HO
O
HO
O
OH NH
H
O
OH
O
O
CO2H
N
H
NH
O
H NOC
O 2
H
N
N
H
OH
O
N
H
O
HN
HO2C
HO2C
O
NH2
O
O
H
N
N
H
Cl
O
O HO
R
Chloramphenicol
NH
O
O
O
OH OH
Cl
O
OH
O
O
H
N
OH O
O
NO2
HO
O
Barbamide
O O
CO2H
O
O
N
Cl
N
H
O
CCl3
OH
H
N
OMe
N
OH
Cl
O
O
O
Cl
HO
NH2
NH
N
H
O
HO
NH2
OH
O
O
O
O
HO
O
O
HN
HO2C
HO
N
H
OH
Cl OH
O
OH
Cl
O
H
N
O
OH
OH
O
N
H
H2N
O
H
N
O
O
N
H
H
N
Vancomycin
Figure 16.2 Structures of natural products whose biosynthetic pathways feature tailoring
enzymes that have been biochemically characterized to act on carrier protein-tethered
substrates. Moieties resulted from tailoring enzymes acting on carrier protein-tethered
substrates are highlighted in gray. See Table 16.1 for specific tailoring enzymes, ACP- or
PCP-tethered substrates and their corresponding products, and the types of modification.
incorporated directly into the C-1027 enediyne chromophore by the SgcC5
condensation enzyme (Lin, Huang, Horsman, Huang, Guo, & Shen, 2012;
Lin, Van Lanen, & Shen, 2009; Fig. 16.3).
In this chapter, we describe protocols for in vitro biochemical characterization of SgcC3 and SgcC that catalyze chlorination and hydroxylation of
SgcC2-tethered (S)-b-tyrosine and analogues. They include: (i) preparation
331
Tailoring Enzymes Acting on Carry Protein-Tethered Substrates
SgcC2
SgcC2
H
H2N
S
O
SgcC3
H
H2N
O
SgcC
Cl
OH
OH
(S)-b-Tyr
O
OH
OH
Å
H3N H
O
O
S
SgcC4
O
SgcC5
O
Enediyne core
Benzoxazolinate
Deoxy aminosugar
OH
O
O
O O
OH OH
O
O
N
H
O
N
SgcC1
ÅH
H3N
H
H2N
Cl
OH
O
SgcC2
S
O
R1
O
R2
NH2
C-1027 (R1 = OH, R2 = Cl)
20-Deschloro-C-1027 (R1 = OH, R2 = H)
22-Deshydroxy-C-1027 (R1 = H, R2 = Cl)
20-Deschloro-22-deshydroxy-C-1027 (R1 = R2 = H)
L-Tyr
Figure 16.3 Biosynthesis of the (S)-3-chloro-5-hydroxy-b-tyrosine moiety of C-1027 and
engineered biosynthesis of C-1027 analogues. (S)-b-Tyrosine was first activated and
tethered to the SgcC2 PCP by the SgcC1 adenylation enzyme. (S)-b-Tyrosyl-SgcC2
was sequentially chlorinated by SgcC3 and hydroxylated by SgcC to afford (S)-3chloro-5-hydroxy-b-tyrosyl-SgcC2, which was directly incorporated into C-1027 by
SgcC5. Manipulation of SgcC3 or SgcC in C-1027 biosynthesis resulted in the production
of three C-1027 analogues, 20-deschloro-C-1027, 22-deshydroxy-C-1027, and
20-deschloro-22-deshydroxy-C-1027.
of the holo-SgcC2 PCP; (ii) preparation of SgcC2-tethered (S)-b-tyrosine
substrates; (iii) SgcC3-catalyzed chlorination of (S)-b-tyrosyl-SgcC2;
(iv) SgcC-catalyzed hydroxylation of (S)-3-chloro-b-tyrosyl-SgcC2; and
(v) exploitation of SgcC2-tethered (S)-b-tyrosine substrates for structural
diversification.
2. METHODS
2.1. In vitro characterization of SgcC3-catalyzed
chlorination of (S)-b-tyrosyl-SgcC2
SgcC3 is a FAD-dependent halogenase, acting only on SgcC2-tethered substrates and accepting both (S)- and (R)-b-tyrosyl-SgcC2. SgcC3 catalyzes
preferentially chlorination but also bromination, and it does not catalyze
fluorination or iodination. SgcC3 requires Cl (or Br), O2, and reduced
FAD. The latter can be supplied by the C-1027 pathway-specific flavin
reductase SgcE6 or Escherichia coli flavin reductase Fre from FAD and NADH
(Fig. 16.4B; Lin et al., 2007).
2.1.1 Expression in E. coli and overproduction and purification of
apo-SgcC2
1. Most ACPs or PCPs of Streptomyces origin, upon expression in E. coli, are
overproduced in apo-form. Follow the protocols provided in Methods
Enzymology, volume 459 (Cheng, Coughlin, Lim, & Shen, 2009;
332
Shuangjun Lin et al.
A
Svp
SgcC2
SgcC2
OH
CoA
SH
ADP
holo-ACP
apo-ACP
B
SgcC2
ÅH
H3N
H
H2N
SgcC1
O
O
ATP
+
SgcC2
OH
PPi
+
AMP
O
S
X
KOH
O
H2O
HO-X
ÅH
H3N
OH
OH
(S)-3-chloro-b-Tyr (X = Cl)
(S)-3-bromo-b-Tyr (X = Br)
FAD-OH FAD-OOH
H2O
O
O
X
X
OH
SH
(S)-b-Tyr
SgcC2
H
H2N
SgcC3
S
O2
FAD
FADH2
SgcE6
NADH
NAD
C
SgcC2
ÅH
H3N
H
H2N
SgcC1
O
O
X
OH
(S)-b-Tyr
ATP
+
SgcC2
SH
PPi
+
AMP
SgcC2
H
H2N
SgcC
S
O
FAD-OOH
S
O
OH
X
O2
H2O
FADH2
NAD
ÅH
H3N
O
O
FAD-OH
X
OH
KOH
FAD
SgcE6
NADH
OH
OH
X
OH
(S)-3-hydroxy-b-Tyr (X = H)
(S)-3-fluoro-5-hydroxy-b-Tyr (X = F)
(S)-3-chloro-5-hydroxy-b-Tyr (X = Cl)
(S)-3-bromo-5-hydroxy-b-Tyr (X = Br)
(S)-3-iodo-5-hydroxy-b-Tyr (X = I)
(S)-3-methyl-5-hydroxy-b-Tyr (X = CH3)
Figure 16.4 In vitro characterization of SgcC3 as a FAD-dependent halogenase and
SgcC as a FAD-dependent hydroxylase that act on SgcC2-tethered (S)-b-tyrosine and
analogues: (A) Svp PPTase-catalyzed in vitro conversion of apo-SgcC2 into holo-SgcC2;
(B) SgcC1-catalyzed preparation of (S)-b-tyrosyl-SgcC2, SgcC3-catalyzed chlorination or
bromination of (S)-b-tyrosyl-SgcC2, and hydrolytic release from SgcC2 of the halogenated products (S)-3-chloro-b-tyrosine and (S)-3-bromo-b-tyrosine; and (C) SgcC1catalyzed preparation of SgcC2-tethered (S)-b-tyrosine and analogues, SgcC-catalyzed
hydroxylation of SgcC2-tethered (S)-b-tyrosine and analogues, and hydrolytic release
from SgcC2 of the hydroxylated products (S)-3-hydroxy-b-tyrosine, (S)-3-fluoro-5hydroxy-b-tyrosine, (S)-3-chloro-5-hydroxy-b-tyrosine, (S)-3-bromo-5-hydroxy-b-tyrosine, (S)-3-iodo-5-hydroxy-b-tyrosine, and (S)-3-methyl-5-hydroxy-b-tyrosine.
Horsman, Van Lanen, & Shen, 2009; Jiang, Rajski, & Shen, 2009) to
express sgcC2 in E. coli BL21 (DE3) and to purify the overproduced
apo-SgcC2 as an N-terminal His6-tagged fusion protein.
2. Dialyze the purified SgcC2 into 50 mM Tris–HCl (pH 7.5), containing
50 mM NaCl and 1 mM dithiothreitol (DTT), and concentrate using an
Amicon Ultra-4 (3K, GE Healthcare, Piscataway, NJ).
3. Check the purity of the isolated protein by SDS-PAGE on a 15% gel
(Fig. 16.5A), determine the concentration by Bradford assay (BioRad, Hercules, CA), and store in 40% glycerol at 20 C until use.
333
MW
S
Sgc tds
C
A
Sgc
E6
Sgc
C3
Sgc
C2
MW
Std
s
Sgc
C1
MW
Std
s
kD
Tailoring Enzymes Acting on Carry Protein-Tethered Substrates
97
66
45
31
21
14
B
C
mAU at 280 nm
D
mAU at 280 nm
I
I
I
II
II
II
III
III
III
15.0
20.0
Time (min)
18.0
20.0
22.0
Time (min)
24.0
mAU at 280 nm
15.0
20.0
25.0
Time (min)
Figure 16.5 Representative data from in vitro characterization of SgcC3 and SgcC with
SgcC2-tethered (S)-b-tyrosine and analogues as substrates. (A) SDS-PAGE analysis of
SgcC2, SgcC3, and SgcE6 on a 15% gel and SgcC1 and SgcC on a 12% gel. (B) HPLC
chromatograms of SgcC3-catalyzed chlorination of (S)-b-tyrosyl-SgcC2: (I) authentic
(S)-b-tyrosine standard (●), (II) assay solution, and (III) authentic (S)-3-chloro-b-tyrosine
standard (ç). (C) HPLC chromatograms of SgcC-catalyzed hydroxylation of (S)-b-tyrosylSgcC2: (I) authentic (S)-3-chloro-b-tyrosine standard (ç), (II) assay solution, (III) authentic
(S)-3-chloro-5-hydroxy-b-tyrosine standard (r), and 4,5-dihydroxy-1,2-dithiane (*)
presented in the assay. (D) HPLC chromatograms of SgcC3-catalyzed bromination of
(S)-b-tyrosyl-SgcC2: (I) authentic (S)-b-tyrosine standard (●), (II) assay solution, and
(III) authentic (S)-3-bromo-b-tyrosine standard (◊).
2.1.2 In vitro preparation of holo-SgcC2 by Svp and of
(S)-b-tyrosyl-SgcC2 by SgcC1
1. Follow the protocols provided in Methods Enzymology, volume 459 (Cheng
et al., 2009; Horsman et al., 2009; Jiang et al., 2009) to convert apo-SgcC2
into holo-SgcC2 using the Svp PPTase (Sanchez et al., 2001; Fig. 16.4A).
Mix 0.8 mL of solution containing 160 mM apo-SgcC2, 0.8 mM
CoA, 12.5 mM MgCl2, and 2 mM tris(2-carboxyethyl)phosphine
hydrochloride (TCEP) in 100 mM Tris–HCl (pH 7.5), initiate the
reaction by adding 5 mM Svp, and incubate at 25 C for 45 min.
334
Shuangjun Lin et al.
2. Express sgcC1 in E. coli BL21 (DE3) and purify the overproduced SgcC1
adenylation enzyme as an N-terminal His6-tagged fusion protein
according to the literature procedure (Fig. 16.5A; Van Lanen et al.,
2006); use SgcC1 to catalyze the tethering of (S)-b-tyrosine to
holo-SgcC2 (Fig. 16.4B). Add 0.8 mL of solution, containing 4 mM
(S)-b-tyrosine, 8 mM ATP, 2 mM TCEP, and 12.5 mM MgCl2, to
the holo-SgcC2 solution from step 1. Initiate the reaction by adding
2 mM SgcC1, and incubate at 25 C for 1 h.
3. Purify (S)-b-tyrosyl-SgcC2 by ion exchange chromatography on a 5-mL
HiTrap Q column (GE Healthcare). Preequilibrate the column with
50 mM Bis–Tris–HCl (pH 7.0), load the reaction mixture from step 2
to the column, and wash it with five column volumes of the same buffer.
Elute the column with a linear gradient from 0% to 100% 1 M NaCl in
50 mM Bis–Tris–HCl (pH 7.0), in 25 column volumes at a flow rate of
3 mL/min. (S)-b-Tyrosinyl-SgcC2 is typically eluted between 0.35 and
0.4 M NaCl.
4. Desalt b-tyrosyl-S-SgcC2 from step 3 using a Superose 12 column (GE
Healthcare) in 20 mM sodium phosphate, pH 7.0, and concentrate using
an Amicon Ultra-4 (3K, GE Healthcare) prior to use in SgcC3 assay.
2.1.3 Expression in E. coli and overproduction and purification of SgcC3
1. Prepare PCR primers for amplification of sgcC3 from cosmid pBS1005
(Liu et al., 2002), clone the PCR product into the pET-30Xa/LIC vector
(Novagen, Madison, WI) using a ligation-independent cloning procedure
to yield the expression plasmid pBS1041, and sequence the construct to
confirm PCR fidelity. With this construct, SgcC3 will be overproduced
as an N-terminal His6-tagged fusion protein (Lin et al., 2007).
2. Introduce pBS1041 into E. coli BL21 (DE3) by transformation, and select
transformants on LB agar plates containing 50 mg/mL kanamycin.
3. Pick a single colony to grow in 3 mL of LB containing 50 mg/mL kanamycin overnight at 37 C, and transfer 0.5 mL into 50 mL of LB containing 50 mg/mL kanamycin to grow again overnight at 37 C to
prepare the seed culture. Inoculate 500 mL of LB containing 50 mg/
mL kanamycin with 5 mL of the seed culture, and incubate at 18 C until
it reaches an OD600 of 0.6.
4. Induce sgcC3 expression by adding IPTG to 0.1 mM and continue incubation at 18 C for 15–20 h.
5. Harvest the cells by centrifugation at 4 C, resuspend the cells in buffer A
(100 mM sodium phosphate, pH 7.5, 300 mM NaCl) supplemented
Tailoring Enzymes Acting on Carry Protein-Tethered Substrates
335
with a complete protease inhibitor tablet, EDTA-free (Roche Applied
Science, Indianapolis, IN), lyse the cells by sonication (4 30 s pulse
cycle), and centrifuge the lysate at 4 C and 15,000 rpm for 30 min to
collect the clear supernatant.
6. Load the supernatant to a preequilibrated Ni-NTA agarose column
(Qiagen, Valencia, CA) with buffer B (buffer A plus 10% glycerol), and
wash the column sequentially with five column volumes of buffer B
and five column volumes of buffer B containing 20 mM imidazole. Elute
the column with five column volumes of buffer B containing 250 mM
imidazole, and pool fractions containing SgcC3.
7. Desalt the purified SgcC3 using a PD-10 column (GE Healthcare) into
50 mM Tris–HCl (pH 7.5), containing 50 mM NaCl and 1 mM DTT,
and concentrate using an Amicon Ultra-4 (10K, GE Healthcare).
8. Check the purity of the isolated protein by SDS-PAGE on a 15% gel
(Fig. 16.5A), determine the concentration by Bradford assay (Bio-Rad),
and store in 40% glycerol at 20 C until use.
2.1.4 Expression in E. coli and overproduction and purification of SgcE6
1. Prepare PCR primers for amplification of sgcE6 from cosmid pBS1006
(Liu et al., 2002) and clone the PCR product into the pET-30Xa/LIC
vector (Novagen) using a ligation-independent cloning procedure to
yield the expression plasmid pBS1042, in which SgcE6 will be overproduced as an N-terminal His6-tagged fusion protein. Sequence the
construct to confirm PCR fidelity.
2. Follow steps 2–8, Section 2.1.3, to afford pure SgcE6 (Fig. 16.5A), and
store in 40% glycerol at 20 C until use.
2.1.5 In vitro assay of SgcC3-catalyzed chlorination of
(S)-b-tyrosyl-SgcC2
1. Set up the SgcC3-catalyzed halogenation of (S)-b-tyrosinyl-SgcC2 in
200 mL of reaction solution, containing 50 mM (S)-b-tyrosyl-SgcC2,
5 mM NADH, 0.10 mM FAD, 100 mM NaCl, 1 mM TCEP, and 5 mM
SgcE6 in 50 mM sodium phosphate buffer (pH 6.0), at 37 C (Fig. 16.4B).
2. Initiate the reaction by adding 20 mM SgcC3, and incubate at 37 C
for 1 h.
3. Terminate the reaction by adding 35 mL of 70% trichloroacetic acid
(TCA), and incubate on ice for 15 min to precipitate all proteins.
336
Shuangjun Lin et al.
4. Pellet the proteins by centrifugation at 4 C and 14,000 rpm for 15 min,
wash the protein pellet twice with 200 mL of 5% cold TCA and once
with 200 mL of ice-cold ethanol, and dry the pellet in a speed-vac for
10 min.
5. Redissolve the protein pellet in 150 mL of 0.1 N KOH solution, and incubate at 70 C for 15 min to hydrolyze the SgcC2-tethered substrate
(S)-b-tyrosine and product (S)-3-chloro-b-tyrosine (Fig. 16.4B).
6. Adjust the solution with 2 N HCl to pH 6, cool on ice for 10 min, and
remove the precipitated proteins by centrifugation at 4 C and
14,000 rpm for 15 min. Collect the supernatant, concentrate to dryness
in a speed-vac, and redissolve the residue in 50 mL of H2O.
7. Subject 20 mL of the sample from step 6 to HPLC analysis on an Apollo
C18 column (5 mM, 250 4.6 mm, Alltech Associates Inc., Deerfield,
IL) with UV detection at 280 nm. Elute the column at a flow rate of
1 mL/min with a 24-min linear gradient from 0% to 40% acetonitrile
in 0.1% TFA.
8. Determine the peaks corresponding to (S)-b-tyrosine and (S)-3-chlorob-tyrosine by comparison to authentic standards (see Fig. 16.5B for a
representative HPLC chromatogram) and confirm their identity by
ESI-MS analysis.
2.2. In vitro characterization of SgcC-catalyzed hydroxylation
of (S)-b-3-chloro-tyrosinyl-SgcC2
SgcC is a FAD-dependent monooxygenase, acting only on SgcC2-tethered
substrates, and requiring O2 and reduced FAD. The latter can be generated
by the C-1027 pathway-specific flavin reductase SgcE6 or E. coli flavin
reductase Fre from FAD and NADH. While (S)-3-chloro-b-tyrosyl-SgcC2
is the natural substrate for SgcC in C-1027 biosynthesis (Fig. 16.3), both
(S)-3-bromo-b-tyrosyl-SgcC2 and (S)-3-iodo-b-tyrosyl-SgcC2 are better
substrates, with (S)-3-fluoro-b-tyrosyl-SgcC2, (S)-3-methyl-b-tyrosylSgcC2, and (S)-b-tyrosyl-SgcC2 also serving as substrates albeit significantly
poorer ones (Fig. 16.4C; Lin et al., 2008).
2.2.1 Expression in E. coli and overproduction and purification of SgcC
1. Follow steps 1–8, Section 2.1.3, to clone sgcC from pBS1005 (Liu et al.,
2002), construct expression plasmid pBS1092, overproduce SgcC in
E. coli BL21 (DE3), and purify SgcC as an N-terminal His6-tagged fusion
protein (Lin et al., 2008).
Tailoring Enzymes Acting on Carry Protein-Tethered Substrates
337
2. Check the purity of the isolated SgcC protein by SDS-PAGE on a
12% gel (Fig. 16.5A), determine the concentration by Bradford assay
(Bio-Rad), and store in 40% glycerol at 25 C until use.
2.2.2 In vitro assay of SgcC-catalyzed hydroxylation of
(S)-3-chloro-b-tyrosyl-SgcC2
1. Prepare (S)-3-chloro-b-tyrosyl-SgcC2 from (S)-3-chloro-b-tyrosine
and holo-SgcC2 by taking advantage of the substrate promiscuity of
SgcC1 (Van Lanen et al., 2006; Fig. 16.4C). Steps 2–4, Section 2.1.2,
provide a protocol to prepare (S)-b-tyrosyl-SgcC2 from purified
holo-SgcC2 using SgcC1. An alternative protocol is provided in this section for the preparation of (S)-3-chloro-b-tyrosyl-SgcC2 from apoSgcC2 directly by coupled assay using both Svp and SgcC1. The two
protocols afford comparative yields with >90% of the free (S)-b-tyrosine
or analogues tethered to SgcC2 (Fig. 16.4).
2. Set up the in vitro 40 -phosphopantetheinylation of apo-SgcC2 in 1.8 mL
of reaction solution containing 200 mM apo-SgcC2, 1.0 mM CoA,
12.5 mM MgCl2, and 2.0 mM TCEP in 100 mM Tris–HCl (pH 7.5),
at 25 C. Initiate the reaction by adding 10 mM Svp, and incubate at
25 C for 45 min.
3. Prepare a loading solution containing 7.0 mM (S)-3-chloro-b-tyrosine,
8 mM ATP, 2.0 mM TCEP, and 12.5 mM MgCl2 in 100 mM Tris–HCl
(pH 7.5), and mix it with an equal volume of the holo-SgcC2 reaction
solution from step 2. Initiate the loading reaction by adding 5 mM SgcC1,
and incubate at 25 C for 1 h. Follow steps 3 and 4, Section 2.1.2, to
purify (S)-3-chloro-b-tyrosyl-SgcC2.
4. Set up the SgcC-catalyzed hydroxylation of (S)-3-chloro-b-tyrosylSgcC2 in 200 mL of reaction solution containing 250 mM (S)-3chloro-b-tyrosyl-SgcC2, 5 mM NADH, 10 mM FAD, 1 mM TCEP,
50 mM NaCl, and 5 mM SgcC in 50 mM sodium phosphate (pH 6.0),
at 25 C.
5. Initiate the reactions by adding 1.5 mM SgcE6 and incubate at 25 C for 1 h.
6. Terminate the reaction and recover (S)-3-chloro-b-tyrosyl-SgcC2 and
its hydroxylated product (S)-3-chloro-5-hydroxy-b-tyrosyl-SgcC2 by
following the steps 3 and 4, Section 2.1.5.
7. Redissolve the protein pellet from step 6 by adding first 5 mL of 1.5 M
DTT and then 150 mL of 0.1N KOH, and incubate at 50 C for
15 min to hydrolyze the SgcC2-tethered substrate (S)-3-chloro-btyrosine and product (S)-3-chloro-5-hydroxy-b-tyrosine (Fig. 16.4C).
338
Shuangjun Lin et al.
8. Follow steps 6–8, Section 2.1.5, for sample preparation and HPLC analysis. Determine the peaks corresponding to (S)-3-chloro-b-tyrosine and
(S)-3-chloro-5-hydroxy-b-tyrosine by comparison to authentic standards (see Fig. 16.5C for a representative HPLC chromatogram), and
confirm their identity by ESI-MS analysis.
2.3. Exploitation of SgcC2-tethered (S)-b-tyrosine analogues
for structural diversification
2.3.1 SgcC3-catalyzed bromination of (S)-b-tyrosyl-SgcC2
1. Prepare SgcC3 according to Section 2.1.3 and SgcE6 according to Section 2.1.4 with the exception of excluding NaCl in all buffers used for
their purification.
2. Prepare the (S)-b-tyrosyl-SgcC2 according to Section 2.1.2.
3. Desalt the (S)-b-tyrosyl-SgcC2 sample from step 2 using a Superose 12
column (GE Healthcare) in 20 mM sodium phosphate (pH 7.0), and run
the sample twice to ensure the complete removal of residual NaCl.
4. Set up the SgcC3-catalyzed bromination of (S)-b-tyrosyl-SgcC2 reaction in an identical condition to that of chlorination with the exception
of replacing NaCl with 0.1 M NaBr and excluding TCEP from the assay
solution, and follow the steps in Section 2.1.5 to carry out the reaction
and analyze the product (Fig. 16.4B). Determine the formation of (S)-3bromo-b-tyrosine by HPLC analysis and comparison with authentic
standard (see Fig. 16.5D for a representative HPLC chromatogram),
and confirm its identity by ESI-MS analysis.
2.3.2 SgcC-catalyzed hydroxylation of SgcC2-tethered (S)-b-tyrosine
analogues
1. Prepare SgcE6 according to Section 2.1.4 and SgcC according to
Section 2.2.2.
2. Prepare SgcC2-tethered b-tyrosine analogues of (S)-3-fluoro-b-tyrosylSgcC2, (S)-3-bromo-b-tyrosyl-SgcC2, (S)-3-iodo-b-tyrosyl-SgcC2,
and (S)-3-methyl-b-tyrosyl-SgcC2 according to Section 2.1.2 with
the exception of replacing (S)-b-tyrosine with corresponding analogues
(Fig. 16.4C).
3. Since SgcC hydroxylates SgcC2-tethered (S)-b-tyrosine analogues with
varying rates, the assay condition described for (S)-3-chloro-b-tyrosylSgcC2 in Section 2.2.2 needs optimization for each of the analogues
to ensure efficient formation of the hydroxylated products.
Tailoring Enzymes Acting on Carry Protein-Tethered Substrates
339
4. Set up the SgcC-catalyzed hydroxylation reaction in 200 mL of solution
containing 250 mM SgcC2-tethered (S)-b-tyrosine or analogues, 5 mM
NADH, 10 mM FAD, 1 mM TCEP, and 50 mM NaCl, in 50 mM sodium phosphate (pH 6.0) at 25 C. For (S)-3-bromo-b-tyrosyl-SgcC2
and (S)-3-iodo-b-tyrosyl-SgcC2, add 1.5 mM SgcC and 2 mM SgcE6
and incubate the reaction at 25 C for 20 min, while for (S)-3methyl-b-tyrosyl-SgcC2, (S)-3-fluoro-b-tyrosyl-SgcC2, and (S)-btyrosyl-SgcC2, add 6 mM SgcC and 2 mM SgcE6 and incubate the
reaction at 25 C for 1 h.
5. Terminate the reaction, recover SgcC2-tethered substrates and their
hydroxylated products, release them from SgcC2 by hydrolysis, and
determine their identities by HPLC and ESI-MS analyses by following
the steps 6–8, Section 2.2.2 (Fig. 16.4C). For maximal sensitivity, use
varying wavelengths to detect the formation of each of the hydroxylated
products: (S)-3-fluoro-b-5-hydroxy-tyrosine from (S)-3-fluoro-btyrosyl-SgcC2 at UV 272 nm, (S)-3-bromo-5-hydroxy-b-tyrosine from
(S)-3-bromo-b-tyrosyl-SgcC2 at UV 282 nm, (S)-3-iodo-5-hydroxyb-tyrosine from (S)-3-iodo-b-tyrosyl-SgcC2 at UV 284 nm, (S)-3methyl-5-hydroxy-b-tyrosine from (S)-3-methyl-b-tyrosyl-SgcC2 at
UV 278 nm, and (S)-3-hydroxy-b-tyrosine from (S)-b-tyrosyl-SgcC2
at UV 277 nm.
3. CONCLUSION
We highlighted in this chapter the emerging roles CPs play in precursor
biosynthesis and post-PKS or post-NRPS modifications and summarized tailoring enzymes that are known to act on CP-tethered substrates (Figs. 16.1
and 16.2; Table 16.1). By covalently tethering, CPs sequester the substrates
from endogenous metabolite pools, thereby increasing their concentration
at the active sites for catalysis. CPs also provide the critical protein–protein
recognitions among the various enzymatic partners, and this feature
provides a new opportunity to engineer natural product diversity by
exploiting CPs to increase substrate promiscuity for the tailoring steps.
Realization of the full potential of tailoring enzymes that act on CPtethered substrates in engineered biosynthesis of natural product structural
diversity depends on continued discovery of new members of this family
of enzymes, further expansion of the catalytic portfolio, fundamental characterization of their reaction mechanisms, and exploitation of their
340
Shuangjun Lin et al.
portability in the broad context of natural product biosynthetic machinery.
The protocols provided here were developed from our current effort to
characterize the SgcC3 halogenase and SgcC hydroxylase, acting exclusively
on SgcC2-tethered b-tyrosine and analogues, in the biosynthesis of the (S)3-chloro-5-hydroxy-b-tyrosine moiety of the antitumor antibiotic C-1027
(Van Lanen & Shen, 2008), but should be applicable to mechanistic characterization and engineered exploitation of other tailoring enzymes that act on
CP-tethered substrates in natural product biosynthesis and structural diversification. The ultimate goal would be to use the in vitro findings to guide
in vivo engineering to produce designer natural product analogues. For
example, it has already been demonstrated that variants of the b-tyrosine
moiety can be tolerated by the C-1027 biosynthetic machinery, resulting
in the production of several C-1027 analogues (Fig. 16.3; Kennedy et al.,
2007; Van Lanen et al., 2005). It would be fascinating to investigate if
the sets of b-tyrosine analogues that can be readily generated by SgcC3
and SgcC in vitro (Fig. 16.4) can be recapitulated in vivo to produce a
focused library of C-1027 analogues, some of which could be developed
into novel anticancer drugs.
ACKNOWLEDGMENT
This work was supported in part by National Institute of Health (NIH) grant CA078747.
REFERENCES
Blasiak, L. C., Vaillancourt, F. H., Walsh, C. T., & Drennan, C. L. (2006). Crystal structure
of the non-haem iron halogenase in syringomycin biosynthesis. Nature, 440, 368–371.
Chan, D. I., & Vogel, H. J. (2010). Current understanding of fatty acid biosynthesis and the
acyl carrier protein. The Biochemical Journal, 430, 1–19.
Chen, H., Hubbard, B. K., O’Connor, S. E., & Walsh, C. T. (2002). Formation of
b-hydroxy histidine in the biosynthesis of nikkomycin antibiotics. Chemistry & Biology,
9, 103–112.
Chen, H., & Walsh, C. T. (2001). Coumarin formation in novobiocin biosynthesis:
b-Hydroxylation of the aminoacyl enzyme tyrosyl-S-NovH by a cytochrome P450
NovI. Chemistry & Biology, 8, 301–312.
Cheng, Y.-Q., Coughlin, J. M., Lim, S.-K., & Shen, B. (2009). Type I polyketide synthases
that require discrete acyltransferases. Methods in Enzymology, 459, 165–186.
Christenson, S. D., Liu, W., Toney, M. D., & Shen, B. (2003). A novel
4-methylideneimidazole-5-one-containing tyrosine aminomutase in enediyne antitumor
antibiotic C-1027 biosynthesis. Journal of the American Chemical Society, 125, 6062–6063.
Christenson, S. D., Wu, W., Spies, M. A., Shen, B., & Toney, M. D. (2003). Kinetic analysis
of the 4-methylideneimidazole-5-one-containing tyrosine aminomutase in enediyne
antitumor antibiotic C-1027 biosynthesis. Biochemistry, 42, 12708–12728.
Cryle, M. J., Meinhart, A., & Schlichting, I. (2010). Structural characterization of OxyD,
a cytochrome P450 involved in b-hydroxytyrosine formation in vancomycin biosynthesis. The Journal of Biological Chemistry, 285, 24562–24574.
Tailoring Enzymes Acting on Carry Protein-Tethered Substrates
341
Dorrestein, P. C., Yeh, E., Garneau-Tsodikova, S., Kelleher, N. L., & Walsh, C. T. (2005).
Dichlorination of a pyrrolyl-S-carrier protein by FADH2-dependent halogenase PltA
during pyoluterin biosynthesis. Proceedings of the National Academy of Sciences of the United
States of America, 102, 13843–13848.
Du, L., Sanchez, C., & Shen, B. (2001). Biosynthesis of hybrid peptide and polyketide
metabolites: Prospects towards engineering novel molecules. Metabolic Engineering, 3,
78–95.
Fischbach, M. A., & Walsh, C. T. (2006). Assembly-line enzymology for polyketide and
nonribosomal peptide antibiotics: Logic, machinery, and mechanisms. Chemistry Review,
106, 3468–3496.
Fischbach, M. A., & Walsh, C. T. (2010). Natural products version 2.0: Connecting genes to
molecules. Journal of the American Chemical Society, 132, 2469–2493.
Flatt, P. M., O’Connell, S. J., McPhail, K. L., Zeller, G., Willis, C. L., Sherman, D. H., et al.
(2006). Characterization of the initial enzymatic steps of barbamide biosynthesis. Journal
of Natural Products, 69, 938–944.
Gago, G., Diacovich, L., Arabolaza, A., Tsai, S.-C., & Gramajo, H. (2011). Fatty acid biosynthesis in actinomycetes. FEMS Microbiology Reviews, 35, 475–497.
Galonić, D. P., Vaillancourt, F. H., & Walsh, C. T. (2006). Halogenation of unactivated carbon centers in natural product biosynthesis: Trichlorination of leucine during barbamide
biosynthesis. Journal of the American Chemical Society, 128, 3900–3901.
Garneau-Tsodikova, S., Dorrestein, P. C., Kelleher, N. L., & Walsh, C. T. (2005). Characterization of the formation of the pyrrole moiety during clorobiocin and coumermycin
A1 biosynthesis. Biochemistry, 44, 2770–2780.
Giessen, T. W., Kraas, F. I., & Marahiel, M. A. (2011). A four-enzyme pathway for
3,5-dihydroxy-4-methylanthranilic acid formation and incorporation into the antitumor
antibiotic sibiromycin. Biochemistry, 50, 5680–5692.
Hollenhorst, M. A., Bumpus, S. B., Matthews, M. L., Jr., Bollinger, J. M., Kelleher, N. L., &
Walsh, C. T. (2010). The nonribosomal peptide synthetase enzyme DdaD tethers Nbfumaramoyl-L-2,3-diaminopropionate for Fe(II)/a-ketoglutarate-dependent epoxidation by DdaC during dapdiamide antibiotic biosynthesis. Journal of the American Chemical
Society, 132, 15773–15781.
Horsman, G. P., Van Lanen, S. G., & Shen, B. (2009). Iterative type I polyketide synthases for
enediyne core biosynthesis. Methods in Enzymology, 459, 97–112.
Jiang, W., Heemstra, J. R., Jr., Forseth, R. R., Neumann, C. S., Manaviazar, S.,
Schroeder, F. C., et al. (2011). Biosynthetic chlorination of the piperazate residue in
kutzneride biosynthesis by KthP. Biochemistry, 50, 6063–6072.
Jiang, H., Rajski, S. R., & Shen, B. (2009). Tandem acyl carrier protein domains in polyunsaturated fatty acid synthases. Methods in Enzymology, 459, 79–96.
Kennedy, D. R., Gawron, L. S., Ju, J., Liu, W., Shen, B., & Beerman, T. A. (2007). Single
chemical modifications of the C-1027 enediyne core, a radiomimetic antitumor drug,
affect both drug potency and the role of ataxia-telangiectasia mutated in cellular responses
to DNA double-strand breaks. Cancer Research, 67, 773–781.
Kopp, F., Linne, U., Oberthür, M., & Marahiel, M. A. (2008). Harnessing the chemical
activation inherent to carrier protein-bound thioesters for the characterization of
lipopeptide fatty acid tailoring enzymes. Journal of the American Chemical Society, 130,
2656–2666.
Lambalot, R. H., Gehring, A. M., Flugel, R. S., Zuber, P., LaCelle, M., Marahiel, M. A.,
et al. (1996). A new enzyme superfamily—The phosphopantetheinyl transferase. Chemistry & Biology, 3, 923–936.
Lin, S., Huang, T., Horsman, G. P., Huang, S.-X., Guo, X., & Shen, B. (2012). Specificity of
the ester bond forming condensation enzyme SgcC5 in C-1027 biosynthesis. Organic Letters, 14, 2300–2303.
342
Shuangjun Lin et al.
Lin, S., Van Lanen, S. G., & Shen, B. (2007). Regiospecific chlorination of (S)-beta-tyrosylS-carrier protein catalyzed by SgcC3 in the biosynthesis of the enediyne antitumor antibiotic C-1027. Journal of the American Chemical Society, 129, 12432–12438.
Lin, S., Van Lanen, S. G., & Shen, B. (2008). Characterization of the two-component, FADdependent monooxygenase SgcC that requires carrier protein-tethered substrates for the
biosynthesis of the enediyne antitumor antibiotic C-1027. Journal of the American Chemical
Society, 130, 6616–6623.
Lin, S., Van Lanen, S. G., & Shen, B. (2009). A free-standing condensation enzyme catalyzing ester bond formation in C-1027 biosynthesis. Proceedings of the National Academy of
Sciences of the United States of America, 106, 4183–4188.
Liu, W., Christenson, S. D., Standage, S., & Shen, B. (2002). Biosynthesis of the enediyne
antitumor antibiotic C-1027. Science, 297, 1170–1173.
Makris, T. M., Chakrabarti, M., Münck, E., & Lipscomb, J. D. (2010). A family of diiron
monooxygenase catalyzing amino acid beta-hydroxylation in antibiotic biosynthersis.
Proceedings of the National Academy of Sciences of the United States of America, 107,
15391–15396.
Marahiel, M. A., & Essen, L.-O. (2009). Nonribosomal peptide synthetases: Mechanistic and
structural aspects of essential domains. Methods in Enzymology, 458, 337–351.
Mercer, A. C., & Burkart, M. D. (2007). The ubiquitous carrier protein—A window to metabolite biosynthesis. Natural Product Reports, 24, 750–773.
Mo, S. J., Kim, D. H., Lee, J. H., Park, J. W., Basnet, D. B., Ban, Y. H., et al. (2011).
Biosynthesis of the allylmalonyl-CoA extender unit for the FK506 polyketide synthase
proceeds through a dedicated polyketide synthase and facilitates the mutasynthesis of
analogues. Journal of the American Chemical Society, 133, 976–985.
Neumann, C. S., & Walsh, C. T. (2008). Biosynthesis of ()-(1S,2R)-allocoronamic acyl
thioester by an FeII-dependent halogenase and a cyclopropane-forming flavoprotein.
Journal of the American Chemical Society, 130, 14022–14023.
Pacholec, M., Hillson, N. J., & Walsh, C. T. (2005). NovJ/NovK catalyze benzylic oxidation
of a b-hydroxyl tyrosyl-S-pantetheinyl enzyme during aminocoumarin ring formation in
novobiocin biosynthesis. Biochemistry, 44, 2819–2826.
Reimmann, C., Patel, H. M., Serino, L., Barone, M., Walsh, C. T., & Haas, D. (2001).
Essential PchG-dependent reduction in pyochelin biosynthesis of Pseudomonas aeruginosa.
Journal of Bacteriology, 183, 813–820.
Sanchez, C., Du, L., Edwards, D. J., Toney, M. D., & Shen, B. (2001). Cloning and characterization of a phosphopantetheinyl transferase from Streptomyces verticillus
ATCC15003, the producer of the hybrid peptide-polyketide antitumor drug bleomycin.
Chemistry & Biology, 8, 725–738.
Shen, B. (2000). Aromatic polyketide biosynthesis. Topics in Current Chemistry, 209, 1–51.
Shen, B. (2003). Polyketide biosynthesis beyond the type I, II, and III polyketide synthase
paradigms. Current Opinion in Chemical Biology, 7, 285–295.
Singh, G. M., Fortin, P., Koglin, A., & Walsh, C. T. (2008). Beta hydroxylation of the
aspartyl residue in the phytotoxin syringomycin E: Characterization of two candidate
hydroxylases AspH and SyrP in Pseudomonas syringae. Biochemistry, 47, 11310–11320.
Staunton, J., & Weissman, K. J. (2001). Polyketide biosynthesis: A millennium review.
Natural Product Reports, 18, 380–416.
Strieker, M., Nolan, E. M., Walsh, C. T., & Marahiel, M. A. (2009). Stereospecific synthesis
of threo- and erythro-b-hydroxyglutamic acid during kutzneride biosynthesis. Journal of the
American Chemical Society, 131, 13523–13530.
Thomas, M. G., Burkart, M. D., & Walsh, C. T. (2002). Conversion of L-proline to pyrrolyl2-carboxyl-S-PCP during undecylprodigiosin and pyoluterin biosynthesis. Chemistry &
Biology, 9, 171–184.
Tailoring Enzymes Acting on Carry Protein-Tethered Substrates
343
Ueki, M., Galonić, D. P., Vaillancourt, F. H., Garneau-Tsodikova, S., Yhe, E.,
Vosburg, D. A., et al. (2006). Enzymatic generation of the antimetabolite g,
g-dichloroaminobutyrate by NRPS and mononuclear iron halogenase action in a streptomycete. Chemistry & Biology, 13, 1183–1191.
Vaillancourt, F. H., Yeh, E., Vosburg, D. A., O’Connor, S. E., & Walsh, C. T. (2005). Cryptic chlorination by a non-haem iron enzyme during cyclopropyl amino acid biosynthesis.
Nature, 436, 1191–1194.
Vaillancourt, F. H., Yin, J., & Walsh, C. T. (2005). SyrB2 in syringomycin E biosynthesis is a
nonheme FeII a-ketoglutarate- and O2-dependent halogense. Proceedings of the National
Academy of Sciences of the United States of America, 102, 10111–10116.
Van Lanen, S. G., Dorrestein, P. C., Christenson, S. D., Liu, W., Ju, J., Kelleher, N. L., et al.
(2005). Biosynthesis of the b-amino acid moiety of enediyne antitunor antibiotic C-1027
featuring b-amino acyl-S-carrier protein intermediates. Journal of the American Chemical
Society, 127, 11594–11595.
Van Lanen, S. G., Lin, S., Dorrestein, P. C., Kelleher, N. L., & Shen, B. (2006). Substrate
specificity of the adenylation enzyme SgcC1 involved in the biosynthesis of the enediyne
antitumor antibiotic C-1027. The Journal of Biological Chemistry, 281, 29633–29640.
Van Lanen, S. G., & Shen, B. (2008). Biosynthesis of enediyne antitumor antibiotics. Current
Topics in Medicinal Chemistry, 8, 448–459.
Walsh, C. T., Chen, H., Keating, T. A., Hubbard, B. K., Losey, H. C., Luo, L., et al. (2001).
Tailoring enzymes that modify nonribosomal peptides during and after chain elongation
on NRPS assembly lines. Current Opinion in Chemical Biology, 5, 525–534.
Weissman, K. J. (2009). Introduction to polyketide biosynthesis. Methods in Enzymology, 459,
3–16.
Zhang, W., Ntai, I., Bolla, M., Malcolmson, S. J., Kahne, D., Kelleher, N. L., et al. (2011).
Nine enzymes are required for assembly of the pacidamycin group of peptidyl nucleoside
antibiotics. Journal of the American Chemical Society, 133, 5240–5243.