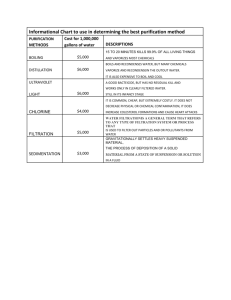
FLUME STUDY OF PARTICLE-SIZE-DEPENDENT FILTRATION RATES OF A
advertisement

FLUME STUDY OF PARTICLE-SIZE-DEPENDENT FILTRATION RATES OF A SOLITARY ASCIDIAN: THE INFLUENCE OF BODY SIZE, FLOW SPEED, AND DRAG Andrew N. Sumerel A Thesis Submitted to the University of North Carolina Wilmington in Partial Fulfillment of the Requirements for the Degree of Master of Science Center for Marine Science University of North Carolina Wilmington 2009 Approved by Advisory Committee Dr. Lynn A. Leonard Dr. Martin H. Posey Dr. Chris M. Finelli Chair Accepted by DN: cn=Robert D. Roer, o=UNCW, ou=Dean of the Graduate School & Research, email=roer@uncw.edu, c=US Date: 2010.01.21 14:49:23 -05'00' ___________________________________ Dean, Graduate School TABLE OF CONTENTS ABSTRACT . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . iv ACKNOWLEGEMENTS . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . v LIST OF TABLES . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . vi LIST OF FIGURES . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . vii LIST OF CONSTANTS AND VARIABLES . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . viii INTRODUCTION . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 1 Factors That Influence Suspension Feeding . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 3 Motivation For Study . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 8 MATERIALS AND METHODS . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 10 Animals . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 10 Flume . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 10 Experimental Protocol . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 11 Filtration Rate Measurements . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 12 Drag Measurements . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 15 Determination Of Additional Body Size Information . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 17 RESULTS AND DATA ANALYSIS . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 18 Relating Body Size Categories . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 18 Filtration Rates . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 19 Drag On Ascidians . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 22 DISCUSSION . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 26 Filtration Response To Increasing Body Size . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 26 Filtration Response To Increasing Flow Speed . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 28 ii Influence Of Drag . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 31 Summary . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 33 REFERENCES . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 35 iii ABSTRACT Ascidians are sessile, active suspension feeders, common in fouling communities around the world. Active suspension feeders create their own feeding current to filter suspended food-particles. Several factors can influence the filtration rates of active suspension feeders including body size, food-particle size, flow speed and the force of drag. It is well established in the literature that the relationship between filtration rate and body size of ascidians is allometric. The effects of food-particle size on filtration rate vary for active suspension feeders, and investigations into the effects of flow speed and drag on the filtration rates of active suspension feeders are rare. Moreover, flow speed and drag effects have not been quantified for ascidians. I have investigated the filtration rates of the solitary ascidian, Styela plicata, and found interacting effects among all of the previously mentioned variables. Consistent with the literature, I determined allometry with respect to body size, but my allometric exponents are comparatively lower than those found in other investigations. This is because of the confounding adverse effects of increasing flow speed, to which filtration rate responded in a non-linear, unimodal fashion. The likely mechanisms for flow-dependent suppression of filtration rate include deformation of internal feeding structures due to drag, and/or increased adverse pressure gradients between the siphons as a result of drag or flow itself. Interestingly, maximum filtration occurs at moderate flow speeds (~ 12 cm/s) when this ascidian filters particles in the range 1.25 to 6.03 µm. Maximum filtering of large particles (6.03 to 100 µm) occurs at low flow speeds (~ 0). The most likely reason for this shift in maximum filtering of different size particles is enhanced filtration of large particles at low speeds due to enhanced capture of large particles at the incurrent region of the inhalant siphon. This also enhances the allometric exponent relating ‘large-particle’ filtration rate to body size. iv ACKNOWLEDGEMENTS I would first like to thank Dr. Chris Finelli, Chair of my thesis advisory committee, for his patience, generosity, and support through this endeavor, and for being extremely approachable and available; a rarity among professors in my opinion. Additional thanks goes to the other members of my committee, Dr. Martin Posey and Dr. Lynn Leonard for their further insight into this research. I also wish to thank those in the Marine Biofluiddynamics and Ecology Lab including Lou Muzyczek, Tiffany Lewis, Kelly Woods and Morgan Church, without whom, life would have been more difficult. Additional thanks goes to Tse-Lynn Loh (and Lou again) for gracious help in the field. I also greatly appreciate Dr. Joan Willey for her guidance and patience with me in continuing to provide Teaching Assistantships as I worked through the Master of Science in Marine Science program. Additionally, this research was funded in part by grants OCE-0751753, OCE-0715271 and OCE-0715272 from the National Science Foundation. Finally, I am continually grateful for my family DeLane, Glenda, Rebecca and Grandma for their endless support, and for Ashley Bissette for her unending thoughtfulness and patience. v LIST OF TABLES Table Page I. Important Results From The Regressions Of Particle-size-dependent Filtration Rate vs. Flow Speed And Body Size . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 22 II. Comparison Of Allometric Exponents In The Literature Relating FR To Body Mass . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 27 vi LIST OF FIGURES Figure Page 1. Top and side cross-sectional drawing displaying structure of a typical ascidian . . . . . . . . . 1 2. Suspected dependence of filtration rate on flow speed and body size . . . . . . . . . . . . . . . . . 5 3. Theoretical dependence of drag on flow speed and body size . . . . . . . . . . . . . . . . . . . . . . . 6 4. Schematic of the flume tank used in these experiments . . . . . . . . . . . . . . . . . . . . . . . . . . . . 11 5. Volume concentration vs. particle-size range (bin) showing an initial sample (yellow) and a sample at four hours (red) for a particular biological trial . . . . . . . . . . . . . 12 6. Allometric relationship between ash-free, dry mass and original frontal area . . . . . . . . . . 18 7. Dependence of frontal area during flow on original area and flow speed . . . . . . . . . . . . . 19 8. Response of a.) ‘small-particle’ b.) ‘large-particle’ and c.) total filtration rate to increasing flow speed and frontal area during flow . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 20 9. Drag coefficient vs. Reynolds number for ‘bluff-body’ wax ascidians to determine drag correction . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 23 10. Corrected drag as a function of flow speed and frontal area for wax ascidians . . . . . . . . . 24 11. Corrected drag as a function of flow speed and frontal body area during flow for S. plicata . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 25 vii LIST OF CONSTANTS AND VARIABLES a . . . . . . . . . . . . . . . . . . . . . Empirically determined exponent in the potential power-law dependence of drag coefficient on Reynolds number (unitless) A . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . King’s Law constant (V2) b . . . . . . . . . . . . . . . . . . . . . . . . . . . Allometric exponent relating filtration rate to body size (unitless) B . . . . . . . . . . . . . . . . . . . . . . . . . King’s Law constant (typically with units V2 s¼ cm-¼ or V2 s½ cm-½) c . . . . . . . . . . . . . Empirical constant equivalent to body-size-dependent, filtration rate in still water (units dependent on body size category) CBo . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . Initial volume concentration during a biological trial (µL/L) CBt . . . . . . . . . . . . . . . . . . . . . . . . . . . Volume concentration at time, t, during a biological trial (µL/L) CD . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . The drag coefficient (unitless) CDo . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . Initial volume concentration during a control trial (µL/L) CDt . . . . . . . . . . . . . . . . . . . . . . . . . . . . . Volume concentration at time, t, during a control trial (µL/L) CL . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . Volume concentration of ‘large’ particles (µL/L) cr . . . . . . . . . . . . Empirical scaling constant relating drag coefficient to Reynolds number (unitless) CS . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . Volume concentration of ‘small’ particles (µL/L) CT . . . . . . . . . . . . . . . . . . . . . . . . . . Volume concentration across the entire particle-size range (µL/L) CTBo . . . . . . . . . . . . . . . . Initial volume concentration of all particles during a biological trial (µL/L) CTB4 . . . . . . . . . . . . . . . . Volume concentration of all particles at 4 h during a biological trial (µL/L) CTDo . . . . . . . . . . . . . . . . . . . Initial volume concentration of all particles during a control trial (µL/L) CTD4 . . . . . . . . . . . . . . . . . . . Volume concentration of all particles at 4 h during a control trial (µL/L) viii D . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . Force of drag on an object (Newtons) DR . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . Deposition rate (particles/h) E . . . . . . . . E-value indicating degree of reconfiguration over a specific flow speed range (unitless) F . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . Momentum flux (Newtons) FR . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . Filtration rate (L/h) FRL . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . Large-particle filtration rate (L/h) FRo . . . . . . . . . . . . . . . . . . . . . . . . . Empirical constant equivalent to filtration rate in still water (L/h) FRS . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . Small-particle filtration rate (L/h) FRT . . . . . . . . . . . . . . . . . . . . . . . . . . . . . Total filtration rate across the entire particle-size range (L/h) l . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . Characteristic length (cm) µ . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . Fluid kinematic viscosity (g cm-1 s-1) n . . . . . . . . . . . . . . . . . . . . . . . Empirically determined King’s Law exponent of flow speed (unitless) N . . . . . . . . . . . . . . . . . . . . . . Total number of 30 second, drag measurements during a trial (unitless) Pd . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . Voltage from a constant temperature thermistor (V) Re . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . Reynolds number (unitless) ρ . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . Fluid density (g/cm3) S . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . Body size (units dependent on body size category) Sa . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . Area of an imaginary plane through which, fluid flows (cm2) sc . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . Empirical scaling constant relating filtration rate to body size (units dependent on body size category) ix Sd . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . Area of an imaginary plane downstream of a body (cm2) Sdj . . . . . . . . . . . . . . . . . The jth area element of an imaginary plane downstream of an ascidian (cm2) Sf . . . . . . . . . . . . . . . . . . . . . . Frontal area of an object projected normal to fluid flow direction (cm2) SM . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . Body mass as wet mass, dry mass, or ash-free, dry mass (g) SO . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . Normally projected frontal area in still water (cm2) SSD . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . Speed-specific drag (Newtons s2 cm-2) Su . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . Area of an imaginary plane upstream of a body (cm2) t . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . Time (h) U . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . Fluid flow speed (cm/s) Ud . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . Flow speed downstream of a body (cm/s) Udj . . . . . . . . . . . . . . The jth downstream mean flow speed estimate over a 30 second interval (cm/s) Up . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . Flow speed where maximum filtration occurs (cm/s) Uu . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . Flow speed upstream of a body (cm/s) Uuj . . . . . . . . . . . . . . . . . The jth upstream mean flow speed estimate over a 30 second interval (cm/s) V . . . . . . . . . . . . . . . . . . . Volume of filterable seawater containing suspended particulate matter (L) w . . . . . . . . . Descriptor related to the full-width at half-maximum of a Gaussian distribution (cm/s) x INTRODUCTION Characterized by a diverse array of morphologies, including both solitary and colonial forms, ascidians are sessile, active suspension feeders that live fixed to a substrate and pump water through feeding structures to filter food particles. The basic structure of the solitary ascidian, Styela plicata, is roughly cylindrical with outer skin known as the tunic (Fig. 1). There are two openings, the inhalant and exhalant siphons, which permit water to enter and exit the body cavity where particles are removed. Tentacles at the inhalant siphon opening serve to select food particles of a certain size, mostly excluding large indigestible particles (Goodbody, 1974). The inhalant siphon leads to the pharynx where lateral cilia, lining stigmatal openings of the branchial basket (pharynx), pump water from the pharynx directly to the atrium from whence it is discharged through the exhalant siphon (Fig. 1). The endostyle lines the entire inner branchial basket and continuously secretes a mucus-net that captures food particles to be digested. Food particles are trapped in the net and then passed on (with the entire net) to the esophagus by frontal cilia also lining the endostyle (Fig. 1). In the esophagus, the food particles and net are digested, with wastes finally entering the atrium as effluent and discharging via the exhalant siphon. Figure 1: Top and side cross-sectional drawing displaying structure of a typical ascidian (Petersen, 2007) Ascidians feed on suspended particulate matter including phytoplankton and large bacteria (Petersen, 2007), and their diet depends in part, on the mucus net structure. The mucus net consists of transverse and longitudinal filaments (10 to 40 nm diameter) resulting in rectangular openings of 0.2-0.5 m by 0.5-2.2 m (Flood & Fiala-Médioni, 1981). Food-particles smaller than the rectangular openings are not necessarily passed through to the atrium undigested. As described by aerosol filtration theory (e.g. Rubenstein & Koehl, 1977), smaller particles can be captured by the filaments via several different mechanisms including direct interception, gravitational deposition, inertial impaction, Brownian diffusional deposition, and electrostatic attraction; and are likely retained after deposition due to van der Waals forces between the particle and sticky mucus filament (Rubenstein & Koehl, 1977; Shimeta & Jumars, 1991; Spielman, 1977; Pich, 1966). For this reason, particles in the 1-2 m range are captured with about 70% efficiency (Randløv & Riisgård, 1979). Particle retention studies have not been performed for sub-micron particles, but it is believed that if these particles are retained, they do not constitute a quantitatively important portion of ascidian diet (Petersen, 2007). Particles greater than 100 m can compound mucus net overloading and often lead to expulsion of pseudofaeces through the siphons via squirting (Robbins, 1984; Petersen, 2007). Styela plicata is a common solitary ascidian with a roughly cylindrical or oblong morphology. They are cloaked in an unstalked tunic that is typically described as tough, thick, leathery (Fuller, 2007), warty and ridged (Howey, 1998). Interestingly, the tunic is composed largely of cellulose compounds. For animals, this is common only to the tunicates (Kimura & Itoh, 2007). Their color is typically grayish or tannish-white, with red or purple stripes (usually four) on the siphons. They are abundant members of fouling communities throughout the world’s oceans in temperate sub-tropical and some tropical climate zones, and they 2 can grow on the sides of floating docks and project outward into flow, horizontal to the water’s surface with siphons typically oriented toward the water’s surface. S. plicata typically only live for less than a year (Kott, 1972), although some investigators suspect they may live between 2 and 3 years (Lambert & Lambert, 1998). Their abundance makes them important animals for ecological scientific inquiry. The connection between life and environment is progressively made clearer through studies of how feeding is affected by ambient hydrodynamic conditions. Such studies are rare for ascidians, yet theory and studies of other active suspension feeders indicate that their filtration rates should vary as a function of ambient flow speed, body size, and drag. Factors That Influence Suspension Feeding Ascidian feeding is very sensitive to a number of perturbing factors including temperature and food-particle concentration (Petersen, 2007). Filtration rate can also depend on body size, a term with variable meaning in the suspension-feeding literature used to describe one of several related terms including body mass (wet mass, dry mass, or ash-free dry mass) and body-area projected normal to flow (in still water or in flow). As body size increases, filtration rate also increases as the allometric relationship, Eq. 1 where FR is the filtration rate, sc is an empirical constant, S is body size, and b is a constant (Randløv & Riisgård, 1979). The exponent b, has been empirically determined to vary between 0.38 (Holmes, 1973) and 1.08 (Klumpp, 1984) for various species of ascidian under various experimental conditions. The 2/3 rule is used as the generally accepted comparison where b = 2/3 (0.667) for the relationship between filtration rate and body mass (Randløv & Riisgård, 1979; Klumpp, 1984) determined from dimensional scaling of squared dimensions of surface area 3 responsible for filtration (branchial surface) and cubed dimensions of body volume (Schmidt-Nielsen, 1997; Klumpp, 1984). In addition to body size, filtration rate can depend on ambient fluid flow. No investigation exists that explicitly concerns the relationship between ascidian filtration rate and varying ambient flow speed, but there have been some flow studies for other active suspension feeders. The giant scallop, Placopecten magellanicus, responded to increasing flow speed by increasing its filtration rate to a peak with a subsequent decrease up to flow speeds of 45 cm/s (Wildish et al., 1992; Wildish & Saulnier, 1993). Additionally, Wildish & Miyares (1990) found that the filtration rate of the blue mussel, Mytilus edulis, rapidly decreased after 6 cm/s and leveled off to near-zero above 25 cm/s. Filtration rates were not estimated for this mussel at flow speeds less than 6 cm/s, but if it was determined that the filtration rate of this mussel increases at zero flow speed to values observed at 6 cm/s, then its flow-dependent filtration rate would follow the same pattern observed for the giant scallop. Wildish & Kristmanson (1997) generally refer to the entire response as unimodal (peak function). The reasons for this general description are that there is no precise theoretical mathematical relationship between flow speed and filtration rate of active suspension feeders, and insufficient data prevent definition of the true form of the curve. Many mathematical descriptions of peak functions exist. In the interest of simplicity, I prefer to describe the response as the symmetrical Gaussian distribution such that filtration rate is a smooth, continuous peak-function of flow speed so Eq. 2 where, FRo is the filtration rate in still water, U is flow speed, Up is the position of the peak, and w describes the width of the “bell” curve. Multiplying the right-side terms of Equations 1 and 2, yields an estimate of filtration rate that is a function of both body size and flow speed 4 Eq. 3 shown in Fig. 2, where c = sc FRo. Figure 2: Hypothesized dependence of filtration rate on flow speed and body size Filtration rate increases directly with flow due to increasing particle flux across the feeding appendage; but does not decrease to zero in still water because active suspension feeders create their own feeding currents. At higher flow speeds, filtration rate is observed to be suppressed. The ultimate reason for this is still unknown, but several hypotheses include particle overloading (Wildish et al., 1992) due to overwhelming particle flux, the development of adverse pressure gradients between inhalant and exhalant siphons in excessive flow (Wildish et al., 1992; Wildish & Saulnier, 1993), and reconfiguration and body torsion due to drag, which could inhibit filtration rate as described below. Drag refers to force that resists the movement of a solid object through a fluid (or moving fluid about a fixed solid object). It consists of frictional forces between the fluid and object's surface and pressure forces, which act in a direction perpendicular to the object's surface. These 5 forces can be nearly impossible to resolve and can themselves depend on a large number of variables. Therefore to limit the degrees of freedom, the typical model for the drag on a body is described by Eq. 4, where drag depends largely on the square of the ambient fluid flow speed and on the frontal body area, Sf, projected normal to flow (Vogel, 1981) so Eq. 4 where D is drag, ρ is the density of the fluid, and CD is the drag coefficient. According to Vogel (1981), the drag coefficient is a function of the Reynolds number, Re, where Re = ρlU/µ. The new symbols here are l, the characteristic length of the object and µ, the dynamic viscosity. For the purposes of this study, the characteristic length is considered the greatest length of the ascidian in the direction of flow. The drag coefficient can be any function of Re, but typically for moderate to high Reynolds numbers, Eq. 5 where cr is an empirical constant and a is a number that may be constant or vary with Re. Figure 3: Theoretical dependence of drag on flow speed and body size 6 Fig. 3 represents the ideal case where the drag coefficient is always constant (if Eq. 5 is true for a body then this only happens when a = 0 or a = [ln CD/cr]/ln Re). This is largely true for what Vogel (1984) refers to as a “bluff” body; a non-streamlined, non-flexible object. One might then expect drag to behave differently for flexible bodies such as ascidians. Drag forces become a factor at high flow speeds, and this could have the effect of suppressing (or even completely inhibiting) filtration of flexible, active suspension feeders for several different possible reasons. First, drag forces can alter a body’s orientation to flow. With regard to ascidians, drag can perturb siphon orientation away from some ‘optimal’ feeding orientation. Knott et al. (2004) found that flow directly into the inhalant siphon was most optimal for feeding. Inhalant siphons oriented with the direction of flow resulted in greatly reduced feeding, and inhalant siphons oriented perpendicular to flow showed the least feeding for Pyura stolonifera. Each of these orientations were investigated at the same flow speed of 15.2 cm/s. It is unknown how the direct influence of increasing flow on orientation will affect feeding, but it is possible that when siphon orientation is sub-optimal, pressure differences increase between the inhalant and exhalant siphons so that active suspension feeders (such as ascidians) must pump harder to process water containing food (Wildish & Kristmanson, 1997; Knott et al., 2004). This scenario would not be energetically efficient, and the ascidian could not compensate. Second, drag forces bend and deform flexible creatures (reconfiguration), which may deform internal filtering structures and reduce the efficiency of food-particle capture and processing (Shimeta & Jumars, 1991). In contrast, reconfiguration could be beneficial where it facilitates shielding of external features responsible for feeding (siphons) or bringing these features closer to the boundary layer where speeds are not as harsh (Sponaugle & LaBarbera, 1991). This would allow some suppressed feeding as opposed to none at all. Finally, it is also possible that drag forces can become so high that complete 7 dislodgement occurs, thereby ending feeding altogether. Thus, it may be that the more flexible the organism, the better adapted they are to survive intense flow when dislodgement is likely (Vogel, 1984; Koehl, 1984; Denny et al., 1985; Koehl, 1999). Additionally, the response of filtration rates of active suspension feeders to varying flow speed over a distribution of particle-sizes varies. This author is unaware of any studies with regard to ascidians on this subject, but some exist concerning bryozoans. The bryozoans, Bugula neritina and B. stolonifera, showed increased ingestion of large-particles and decreased ingestion of small-particles around flow speeds of 10 to 12 cm/s while there was consistently higher selection of large particles at both flow speeds (Okamura, 1990). Each of the relationships mentioned previously are interrelated and underappreciated. Additionally, relationships among filtration rate, selectivity, flow speed, body size, and drag have not previously been quantified for ascidians. Quantifying and understanding these relationships is the intent of this investigation. Motivation For Study As inferred from the above discussion, quantitative uncertainty exists concerning ascidians’ response of particle-size-dependent filtration rate to fluid flow, body size and drag. Further understanding and quantification of these relationships is very important to theorists who use hydrodynamic theory to explain aspects of the biology and behavior of suspension feeders. Also, fouling due to ascidians has become an increasing nuisance for commercially important marine cultures (Petersen, 2007). A better understanding of how ascidians behave in different flow regimes may aid in reduction of fouling. It is possible that mussels can be given a competitive advantage over ascidians by adjustment of factors (temperature, food concentration, etc.) that affect feeding, because feeding response differs between mussels and ascidians (Petersen, 2007). It is also possible that mussels (and other active suspension feeders) can be 8 given a competitive advantage over fouling ascidians in certain flow regimes. Knowledge of how to reduce competitive pressure on commercially important species would be very valuable. Of further value is the possible use of S. plicata as bioremediators (Draughon, 2009). As coastal development has increased, natural habitats have changed and bacterial concentrations can rise to unhealthy levels (Aslan-Yilmaz, 2004). Because bacteria are a significant portion of ascidian diet, further understanding of the relationship between particle-size-dependent filtration and flow regime could prove useful for the bioremediation effort. Presented here are particle-size-dependent filtration rates of S. plicata as a response to varying flow speed and body size. I also present indirect measurements of drag on each solitary ascidian with varying flow speed and body size. These data provide a better understanding of how ascidian feeding is affected by ambient hydrodynamic conditions. 9 MATERIALS AND METHODS Animals Specimens of Styela plicata were collected from floating docks in the Atlantic Intracoastal Waterway (AIWW) near Wrightsville Beach NC (Latitude 34° 12' 31" N, Longitude 77° 47' 47" W). Individual ascidians were gently removed from the dock surface using a putty knife, and placed in plastic bags of ambient seawater for transport to the lab. Specimens were initially selected based on their maximum end-to-end length at minimum body size (attained after being gently disturbed in their habitat). Individuals were later grouped into a more formal body size category based on projected body area. Upon arrival at the lab, each asidian was cleaned to remove epibionts, blotted dry, and attached at the base to 6 cm by 8 cm Lucite Acrylic Sheet slides (2 mm thick) with cyanoacrylate glue (Elmer’s Instant Krazy Glue). Mounting on slides was necessary to facilitate immediate transfer to the flume for experimental use. Once secured in the flume, the frontal body area of an ascidian was determined by photographing the individual face-on in still water with a common digital camera (Samsung, 4.0 mega pixel, Digimax A402) and measuring the area with commonly available image analysis software (Broken Symmetry ImageJ, Vers. 1.4.3.67). Ascidians were grouped into three categories (for replication) based on frontal area: small = 4.9 to 8.6 cm2, medium = 9 to 12.2 cm2, and large = 13 to 17.7 cm2. Flume Measurements of filtration and drag were gathered for varying size specimens of S. plicata in a 16.4 L, laboratory paddle-flume tank (Fig. 4) at varying flow speeds. A Dayton gear motor (Model 2H577A) rotated a paddle wheel on one side of the annular, open-channel flume (raceway flume), and the rotation speed of the wheel was varied with a Dart Micro-drive II controller. The paddles on the wheel move water unidirectionally around the flume from the paddle channel to the observation channel. The flow speeds under consideration were approximately 3 cm/s, 15 cm/s, and 21 cm/s as determined using an acoustic Doppler velocimeter (ADV; SonTek 16-Mhz MicroADV) with the probe mounted in the center of the flume and sample volume located 5 cm above the flume bottom (e.g. Finelli et al. 1999) . This flume is described in more detail in Robinson et al. (2007) and Clarke et al. (2009). Figure 4: Schematic of the flume tank used in these experiments (modified from Robinson et al., 2007) Experimental Protocol Prior to the beginning of each experimental trial, the flume was filled to a height of 10 cm with seston-free artificial seawater (Crystal Sea) at a salinity of 35 ‰, and was constantly aerated. An ascidian (mounted on a plastic slide) was transferred to the flume and secured to the bottom of the flume channel with clear packing tape such that the inhalant siphon was oriented upward, perpendicular to flow, and the exhalant siphon was oriented horizontal to the water’s surface, also perpendicular to flow. The flume motor was started and allowed to come to speed. The ascidian was allowed to acclimate to flow without feeding for at least 12 hours. There were 45 total trials (3 different body sizes at 3 different flow speeds with 5 replicates), each with a corresponding 11 control trial using a simulated ascidian made from parafin wax (Boekel Tackiwax). Control wax ascidians did not require the 12 hour acclimation period. Filtration Rate Measurements After acclimation, 3 g of cultured and preserved algae paste (Innovative Aquaculture Products, LTD) was added to the flume. Filtration data were gathered over a four hour period by determining seston clearance. At each predetermined time (0, 2, 4 hrs), three replicate 180 mL water samples were obtained by syringe from the flume. Water samples were returned to the flume after being analyzed with a Sequoia LISST Portable Laser Diffraction Particle Size Analyzer. The LISST analyzer provides volume concentration of suspended particles (µL/L) in 32 logarithmically-increasing particle-size bins from 1.25 to 250 µm (Fig. 5). Because food-particles larger than 100 µm are not ingested by ascidians, I truncated the particle distribution to remove particles > 100 µm, resulting in 27 size bins. Figure 5: Volume concentration vs. particle-size range (bin) showing an initial sample (yellow) and a sample at four hours (red) for a particular biological trial Chlorophyll analysis, using a Turner Trilogy Fluorometer and following the methods of Welschmeyer (1994), showed that the concentration of Chl a always stayed above 12 µg/L during 12 an entire feeding period. This approach was necessary because filtration rates of ascidians do not appear to vary with concentration above this amount (Petersen, 2007). The volume concentration distribution of seston in the flume exhibited two distinct concentration peaks near 2 µm and near 22 µm (Fig. 5). Therefore for simplicity, the 27 particle ranges were grouped into two categories such that the ranges around the first peak (1.25 to 6.03 µm) were considered the ‘small’ particle group, and the ranges around the second peak (6.03 to 100 µm) were considered the ‘large’ particle group. The area under an entire curve over the particle-size distribution is the total volume concentration, CT. The area under a curve corresponding to the ‘small’ group is the ‘small-particle’ volume concentration, CS. The area under a curve corresponding to the ‘large’ group is the ‘large-particle’ volume concentration, CL. Filtration rates were obtained from the volume concentrations as outlined by Coughlan (1969). During a control trial, particles are assumed to only be deposited gravitationally in a moving fluid such that the volume concentration decays exponentially (Coughlan, 1969; Fries & Trowbridge, 2003), so Eq. 6 where CDt is the volume concentration at time t, CDo is the initial volume concentration, and DR is the deposition rate (particles per hour). Taking the natural log of both sides of Eq. 6, yields ln Eq. 7 which is the number of particles deposited by time t. Organisms feeding at a constant rate will reduce successive ambient concentrations such that the ambient concentration of food particles at any time decays exponentially (Coughlan 1969; Peters 1984). Therefore, during a biological trial with both filtration and gravitational deposition taking place, the volume concentration at time t is assumed to be 13 Eq. 8 where, CBt is the volume concentration at time t, CBo is the initial volume concentration, FR is the filtration rate, and V is volume of filterable seawater in the flume. Figure 5 is an example where CBo is represented in yellow and CBt is represented in red. Taking the natural logarithm of both sides of Eq. 8, yields ln Eq. 9 and substituting Eq. 7 into Eq. 9 to account for gravitational deposition, the filtration rate for a suspension feeding organism is ln ln ln Eq. 10 with units of liters per hour. Since there were three different classes of volume concentration (CT, CS and CL), and considering that each trial lasted 4.00 hours with V = 16.40 L, the total filtration rate, FRT, for example is 4.10 ln Eq. 11 with units of liters per hour where CTBo is the total, initial volume concentration during a biological trial, CTB4 is the total volume concentration at four hours during a biological trial, CTD4 is the total volume concentration at four hours during a control trial, and CTDo is the total, initial volume concentration during a control trial. Filtration rates (‘small-particle,’ ‘large-particle’ and total) were scatter-plotted (with SigmaPlot vers. 9.01) as a function of flow speed and each body size category. Regression analyses were performed for the 3-dimensional plots, with regressions of the surface described by Eq. 3. 14 Drag Measurements Drag data were gathered using a procedure proposed by Vogel (1981). First, it is necessary to introduce the concept of “momentum flux” of a fluid such that Eq. 12 where F is the momentum flux of a fluid through an imaginary plane of area Sa, ρ is the density of the fluid, and U is the velocity of the fluid. Notice that the momentum flux has units of force (Newtons). Applying Newton’s Third Law, if a fluid exerts a drag on a body, then that body must remove momentum from the fluid at a rate that balances its drag, or an incremental element of drag is equal to change in momentum flux across incremental area elements upstream and downstream of the body that is removing momentum so Eq. 13 where dD is an incremental element of drag, dSu and dSd are the incremental area elements of imaginary planes upstream and downstream of the body respectively, and Uu and Ud are the respective upstream and downstream velocities of the fluid through their respective incremental area elements. Applying the principle of continuity of an incompressible fluid (Vogel, 1981), Eq. 14 such that Eq. 13 becomes Eq. 15 and, integrating Eq. 15 yields the drag on a body such that D U d (U u U d )dS d Eq. 16 where D is drag, is the density of seawater, Ud is the downstream flow speed, Uu is the upstream flow speed, and dSd is the incremental area element of the imaginary plane downstream of the body. Therefore, drag on a body can be measured indirectly by measuring flow speeds at 15 infinitely many points on an imaginary plane downstream of the body while simultaneously measuring the corresponding flow speeds on an imaginary plane upstream of the body. The wake of typical ascidians for each of the three body size categories were observed with fluorescein dye (Sigma Chemical Co. A6877) to determine the position of the sampling plane where a constant-temperature, thermistor flowmeter (LaBarbera & Vogel, 1976) was placed. Using the thermistor, flow data was recorded for 30 seconds at a particular position in the downstream sampling plane (Fig. 4). The thermistor was moved to different positions in the sampling plane and the recording process repeated until an adequate array of measurements was determined across the imaginary plane downstream of the ascidian. Another series of corresponding measurements were made across an imaginary plane upstream of the ascidian in a similar way until there were sufficient measurements to determine drag during a trial. Flow data from the thermistor was recorded using a commercially available data logger (Campbell Scientific CR10X). Thermistor data was obtained from the data logger and reduced with common statistical analysis software (SAS Vers. 9.1). The data consisted of, at most, 120 groups of voltage values recorded every 0.25 seconds over 30 seconds. Using a calibration performed with the thermistor and the SonTek ADV, the voltage values were transformed to flow speed values using King’s Law (Brunn, 1996; Al-Deen et al., 1996; Al-Garni, 2007), Eq. 17 where U is the flow speed, Pd is the voltage, A and B are constants, and n is a constant that is usually considered to be ~ 0.5 but can depend on probe type, calibration method, and velocity range (Al-Garni, 2007; Brunn, 1996). More generally, n has been observed to vary between 0.4 and 1.3 in air-flow studies (Al-Garni, 2007), but smaller exponents have been observed in 16 water-flow studies where n is usually in the range 0.25 to 0.3 for flow velocities less than 20 cm/s and in the range 0.4 to 0.45 for flow speeds between 40 and 50 cm/s (Brunn, 1996). The constants A, B and n in this study were determined from calibrations to be -1.0159, 2.1443, and 0.25 respectively for water temperatures above 18.0°C and -1.038, 1.9226, and 0.25 respectively for water temperatures below 18.0°C. These King’s Law calibration constants lie in the reported range for the velocities in this study. Because measurements were obtained at 120 or fewer points on the plane (and not an infinite array of measurements), Eq. 16 was transformed to N D U dj (U uj U dj )S dj Eq. 18 j 1 where N is the total number of 30 second measurements for each area segment Sdj behind each ascidian, Udj is a downstream mean flow speed over the 30 second interval, and Uuj is an upstream mean flow speed over the 30 second interval. The mean of flow values were then found over each 30 second interval and input into Eq. 18 along with = 1.025 g/cm3 (Weast, 1967-1968) and Sdj = 0.25 cm2, which yields an adequate estimate of the drag on an ascidian of a particular body size at a particular upstream flow speed, Uu = (1/N)Uuj. Determination Of Additional Body Size Information After each experiment, the height of the ascidian (from tank bottom) was measured during flow. Because it was impossible to image the frontal area of the ascidian in flow, the ratio of final height to initial height was used to ‘vertically shrink’ the original face-on picture (if the ratio was less than one) in order to estimate frontal body area (also with ImageJ) during flow. In addition, after experimental use, each ascidian was dried in an oven at 60ºC for 24 hours, then combusted in a muffle furnace (550ºC for 3 hrs) to determine ash-free, dry mass (AFDM; range 0.15 and 1.05 g). 17 RESULTS AND DATA ANALYSIS Relating Body Size Categories The term, body size, is used with variable meaning in the suspension feeding literature. In this study, ascidian body was estimated size using ash-free, dry mass (AFDM) and frontal body area (both in still water and exposed to flow). These variables are closely related however, frontal body area during flow, Sf, directly links body size in both filtration and drag estimates. There was a strong power-law relationship between AFDM, SM, and original (i.e. in still water) frontal area (R2 = 0.8066, P < 0.0001), where SM increases with original frontal area, SO, according to the relation SM = 0.02420 SO1.304 (Fig. 6). This allometric exponent is not statistically different from the value of 1.47 (t-test, P = 0.1538) obtained by Sherrard & LaBarbera (2005) for the solitary ascidians, Corella inflata and Ciona savignyi. Figure 6: Allometric relationship between ash-free, dry mass and original frontal area (R2 = 0.8066, P < 0.0001) The relationship between original frontal area and frontal area during flow is more complex with the additional influence of flow speed (Fig. 7 where the planar regression is described by the equation Sf = 2.6938 + 0.8156SO – 0.1381U). The minus sign in this relationship indicates that as speed increases, ascidians increasingly ‘lay over’ in flow so that siphon orientation is initially perpendicular at slow flow but oriented (or nearly oriented) with flow at the highest speeds. Intermediate speeds result in intermediate orientations. Fig. 7 also shows that regardless of flow speed, frontal area in flow is nearly one-to-one with original frontal area. Understanding these relationships between body size categories (and flow speed) is useful for interpretation of filtration rate results. Figure 7: Dependence of frontal area during flow on original area and flow speed (R2 = 0.8833, P < 0.0001) Filtration Rates Roughly the same filtration pattern was seen for all three body size categories. Therefore, filtration rates are only presented as a function of flow speed and frontal area during flow because drag is also dependent on precisely these variables. Four of the biological trials yielded slightly negative filtration rates that were treated as zero filtration. Additionally, the choice of an asymmetrical peak distribution (or other form of unimodal curve) over the simple symmetrical peak distribution would have been inconsequential. In fact, a modified log-normal response of filtration rate to increasing flow speed was only a slight improvement of fit to ‘large-particle’ 19 filtration while also a slightly worse fit to ‘small-particle’ and total filtration. The resulting surface fits for ‘small-particle’ and total filtration rates yielded R-squared values less than 0.3. This is consistent with increased variation in the data for those scatter plots. Better fits occurred for ‘large-particle’ filtration rates (R2 ≈ 0.6). For both total and large-particle filtration, there was a highly statistically significant relationship (P < 0.01) between dependent and independent variables. This relationship was not statistically significant for small-particle filtration, although the P-value for the regression was less than 0.1. Specific R-squared values and P-values are given in Table I. Figure 8: Response of a.) ‘small-particle’ b.) ‘large-particle’ and c.) total filtration rate to increasing flow speed and frontal area during flow 20 Total filtration rate (FRT) seems to be largely driven by large-particle filtration rate (FRL) with some slight modification from small-particle filtration rate (FRS). Therefore FRT, like FRL, decreases with increasing flow speed, but FRT decreases less rapidly. The plotted data show a continual decrease of FRT and FRL with flow speed, which differs from the expected unimodal curve (Fig. 2). However if the Gaussian curve is a good representation of FR, then there are insufficient data to show the peak which would likely exist near the slow flow speed. Such is the case since the peak was constrained to be positive as seen in Table I. A unimodal response to flow speed is seen for FRS, with a peak near the moderate flow speed. Specific peak values are reported in Table I where σp is the standard error of the peak value. Particle selectivity is evident in Fig. 8. Upon comparison of the larger FRL values to the consistently smaller FRS values, Styela plicata appears to select large particles more than small particles at each flow speed. Flow appears to have differing effects on filtration of different size particles and can best be described by the peak position. As flow increases from slow to moderate speeds, large-particle filtration drops dramatically while small-particle filtration increases slightly. Both large- and small-particle filtration appears to be suppressed at high flow speeds. The relationship between FR and body size is consistent regardless of how body size is defined (AFDM vs. projected area in still water or in flow). The allometric exponents that describe the relationship between FR and the three measures of body size are reported in Table I where σb is the standard error of the exponent, b. In general, the allometric exponent, b, is lower for small-particle filtration and total filtration rates, than for large-particle filtration rates. Moreover, values of b are also lower for the relationship between FR and SM than for the same relationship using projected area to measure body size. This can be partially explained by the relationship between AFDM and projected area (SM = 0.02420 SO1.304). 21 Table I: Important Results From The Regressions Of Particle-size-dependent Filtration Rate vs. Flow Speed And Body Size Body Size Category FRS AFDM FRL FRT Original FRS frontal FRL area FRT Area FRS during FRL flow FRT R2 P-value* b ± σb** Up ± σp*** 0.152 0.579 0.270 0.0779 < 0.0001 0.0045 0.267 ± 0.188 0.579 ± 0.180 0.305 ± 0.182 11.9 ± 1.4 ~0 2.83 ± 9.16 0.160 0.585 0.286 0.0653 < 0.0001 0.0030 0.436 ± 0.278 0.812 ± 0.253 0.511 ± 0.254 12.3 ± 1.5 ~0 3.36 ± 9.00 0.145 0.567 0.254 0.0905 < 0.0001 0.0070 0.365 ± 0.280 0.745 ± 0.257 0.361 ± 0.267 12.8 ± 1.7 ~0 2.83 ± 11.6 *Probability that a random distribution will produce similar results as described by Eq. 3 **Allometric exponent (and associated standard error) of body size as it relates to filtration rate ***Flow speed where peak filtration occurs and the associated standard error Drag On Ascidians S. plicata are not bluff bodies. Therefore, drag on the organism does not increase precisely with the square of flow speed. Vogel (1984) introduced a way of comparing the drag coefficients of flexible organisms to those of bluff bodies. If drag is divided by the square of flow speed, Eq. 4 becomes Eq. 19 where SSD is what is known as speed-specific drag. Recall that for a bluff body, CD is typically constant, and a plot of SSD versus U will yield a horizontal line. However for a flexible body, a plot of SSD versus U should show deviations from the horizontal ‘base’ line of a bluff body. This ‘additional variation’ with flow speed appears in the drag coefficient because Eq. 5 is often true for flexible bodies, and substituting Eq. 5 into Eq. 19 with Re = ρlU/µ, then Eq. 20 and in short, SSD is proportional to Ua. For the purposes of comparison between flexible bodies, Vogel (1984) expressed the exponent, a using the variable E, because Eq. 5 might not always be 22 true for a body depending on flow regime and the body itself. These E-values are usually negative and therefore an indication of how well a flexible body can reduce the rate of increase in drag as speeds increase. Further variation in SSD due to body area can be accounted for by simply plotting the log of drag coefficient versus the log of Reynolds number. The slope of the resulting trend will be the exponent a, the E-value. Surprisingly, an E-value of -1.8 for the non-flexible wax ascidians was determined in the control trials. Because CD consistently decreased with increasing Reynolds number in the controls (Fig. 9), I suspected that there was an apparent drag-dampening effect due to my method and apparatus that would also be present in the biological trials. Therefore a correction factor of 9.4 x 10-7 Re1.8 was multiplied to the control drag data, and fit Eq. 4 to the corrected control data (Fig. 10) yielding a drag coefficient estimate of 0.98 ± 0.05. Figure 9: Drag coefficient vs. Reynolds number for ‘bluff-body’ wax ascidians to determine drag correction (R2 = 0.67, P < 0.0001) 23 Figure 10: Corrected drag as a function of flow speed and frontal area for wax ascidians (R2 = 0.86) The Reynolds number correction was determined for bluff bodies where the characteristic length does not vary with flow speed. Therefore, the Re correction could not be applied to the biological drag data until determination of what I termed the ‘bluff Reynolds number.’ The bluff Re is the Reynolds number a flexible object would have if it remained upright in flow so as not to change its characteristic length. After the bluff Re correction, drag on ascidians appears to increase somewhat linearly with the projected frontal area during flow, and drag increases with less-than-the-square of flow speed as expected (Fig. 11). Fitting D estimate of -0.43 ± 0.20 for the range 1000 < Re < 10000. 24 SfU2+E, yielded an E-value Figure 11: Corrected drag as a function of flow speed and frontal body area during flow for S. plicata (R2 = 0.83, P < 0.0001) 25 DISCUSSION I have presented measurements of drag and particle-size-dependent filtration rates for the ascidian Styela plicata as responses to varying body size and ambient flow speed in a flume. Below I discuss in further detail the possible reasons why drag may be producing the expected results and how these findings compare to those of other investigations of suspension feeders. Filtration Response To Increasing Body Size Reports of relationships between ascidian filtration rate and body mass are consistently allometric. Those exceptions not listed in Table II include the case of Draughon (2009) who found no relationship between filtration rate and total wet mass of Styela plicata, and the study by Petersen & Svane (2002) which showed no relationship between filtration rate and total dry mass of a conglomerate of species (Clavelina lepadiformis, Ciona intestinalis, Corella parallelogramma, Ascidia virginea, Boltenia echinata, Molgula manhattensis, and Pyura tessellata). Petersen & Svane (2002) suggest the lack of correlation to be due to their particular methods concerning mass determination. Additionally, differences in tunics among species could increase variation of total mass. The lack of correlation found by Draughon (2009) cannot yet be commented on for need of further details from the author including materials, methods and analysis. Allometric exponents in Table II approximately equal to one are surprising and considered to be due to a lack of low-mass specimens (Klumpp, 1984) or due to non-isometric growth of the tunic compared with total mass (Randløv & Riisgård, 1979). Holmes (1973) determined small allometric exponents in still water, and higher exponents in running water (with the exception of some small exponents in running water for Ascidiella aspersa). The flow speed of the running water regime cannot be precisely determined from Holmes’ data but is likely no more than 5 cm/s (estimate from volumetric flow). The low exponents were suspected to result from animal disturbance. This could be true for some of the results for Ascidiella aspersa, but the higher exponents in running water could be an indication of increased filtration from increased seston flux. Table II: Comparison Of Allometric Exponents In The Literature Relating FR To Body Mass Species Pyura stolonifera Ascidia atra Ciona intestinalis Animal Food-particle size range size range Allometric exponent 0.1-19 g 0.3-13 g 10-160 g 6-50 µm No feeding 0.7 1.08 0.87* 0.05-0.18 g 4.8-8 µm 0.68 Ciona intestinalis Ascidiella aspersa Ascidiella aspersa 0.03-0.4 g 1-7 µm 0.84 Styela clava 0.1-1.0 g Styela plicata 1.2-1.9 g 7-8 µm 0.415 0.617 0.646 0.934** Styela plicata 0.15-1.05 g 1.25-6.03 µm 6.03-100 µm 1.25-100 µm 0.267 0.579 0.305 0.04-0.8 g 0.1-0.4 g 0.1-1.0 g Methods/conditions Source Indirect, flow-through mL/min for g dry mass Indirect, flow-through cm3/min for g wet mass Indirect, static mL/min for g dry mass Indirect & direct, static mL/min for g dry mass 1.05 8-12 µm 0.45 0.383-0.607 Indirect, static Direct, flow-through mL/min for g dry mass Indirect, static Direct, flow-through mL/min for g dry mass Indirect, static L/h for g dry mass Indirect, flume L/h for g ash-free, dry mass Klumpp (1984) Hecht (1916) Petersen & Riisgård (1992) Randløv & Riisgård (1979) Holmes (1973) FialaMédioni (1978) Present study *Determined by Randløv & Riisgård (1979) **Calculated from the data of Fiala-Médioni (1978) The allometric exponent of 0.579, relating ‘large-particle’ filtration to body mass, is not statistically different from the expected value of 0.667 (t-test, P = 0.629). Estimated allometric exponents for small-particle filtration and total filtration were both significantly different than 0.667 (t-test, Small: P = 0.0388; Total: P = 0.0526). Indeed there is much variation in reported exponents in Table II, and some suggest that this variation not only reflects the differences among 27 species but also reflects differences in experimental methods used to determine allometry (Klumpp, 1984; Randløv & Riisgård, 1979; Riisgård, 2001). The reason for my low exponents would be that the allometric fit to varying body size is across the entire range of flow speeds, most of which tend to lower filtration rate. When the confounding influence of flow speed on the relationship between body size and FR was removed by calculating b for only the slow flow treatments, estimates of 0.519 for small particles, 0.646 for large particles, and 0.416 for total were obtained. These values are much closer to the expected 0.667; an indication that flow speed has a strong influence on the allometric filtration response to body size. Filtration Response To Increasing Flow Speed Particle-size-dependent filtration rates are clearly suppressed at high flow speeds. Particle overloading due to increased particle flux is an unlikely suppression mechanism, because this should be associated with increased siphon closure and squirting during intense flow, which was not observed. Also, no additional mucus strings were observed in the flume, which is also indicative of increased squirting. In fact, very little fecal matter was observed after the ascidians had acclimated to intense flow. This observation may indicate decreased digestion of material from the AIWW, although it is also possible for fecal matter to break up at high flow speeds. A more plausible explanation may be the development of adverse pressure gradients across the siphons during excessive flow. The Gaussian function of flow speed as a description of filtration rate, although arbitrary, proved to be quite useful. Interestingly, maximum filtration of small particles occurs at higher speeds (Up ≈ 12 cm/s) than does filtration of larger particles (Up ≈ 0 cm/s). I suspect enhanced filtration of large particles occurs at low speeds due to enhanced capture of large particles at the incurrent region of the inhalant siphon. This particle-size effect also enhances the allometry of 28 FRL with body mass as seen in Table II. Further, higher R-squared values for the surface fits to FRL indicate the influence of flow speed on filtration of large particles is far greater than filtration of small particles. Whether this change in particle-size-dependent filtration is due to active selection by the animal or changing particle dynamics in changing flow regimes, the result is that the total filtration response to increased flow gradually decreases over a wide range of speeds so that feeding remains somewhat steady until flow speeds become too great. Few studies investigate the response of filtration rate to increasing flow speed, so additional studies are discussed below describing the response of growth rate to increasing flow speed. The most commonly studied suspension feeders are bivalves and bryozoans. Bivalves The bay scallop, Argopecten irradians, showed similar responses of growth to increasing flow speed. Kirby-Smith (1972) found that growth rate falls off rapidly from ~ 0.5 cm/s to 12.5 cm/s after which, nearly no growth occurs. In another study, this bay scallop’s growth rate appears to increase steeply from 0 to 1 cm/s and then fall off gradually from 1 to 15 cm/s (Cahalan et al., 1989). Eckman et al. (1989) determined that growth rate decreased steadily from 0 to 17 cm/s after which, there was minimal growth. In contrast, the growth rate of the oyster, Crassostrea virginica, increased steadily within the flow range under investigation (0 to 7 cm/s), but is suspected to be inhibited with increasing flow speed at some point after 7 cm/s (Lenihan et al., 1996). The growth rate of the giant scallop, Placopecten magellanicus, appeared to increase steadily between flow speeds of 0 and 5 cm/s, after which growth rate dramatically fell off up to speeds of 18 cm/s (Wildish et al., 1987). Additionally, filtration rates of this giant scallop appeared to fall off steadily between flow speeds of 10 to 80 cm/s (Wildish et al., 1992; Wildish & Saulnier, 1993) depending on seston concentration, which led Wildish et al. to suspect that 29 inhibited growth is due to inhibited filtration. This may also be true of S. plicata based on the current investigation and personal observations along the floating docks of the AIWW. Where docks are parallel to flow and vary in distance from shore, the average size of S. plicata decreases with increasing distance from shore. The increased distance corresponds to increased flow speed (as determined by a Nortek Vector ADV) along the docks. Most of the natural flow speeds, created between tides, were replicated in the lab where filtration was inhibited in high flow. Supposing there is also a strong connection between filtration rate and growth rate for S. plicata, inhibited growth rate does not necessarily place a limit on body size. However, since S. plicata typically only live for nearly a year, inhibited growth rate could limit body size before natural mortality. It is therefore likely that body size is limited along the outer docks by an as-yet-undetermined mechanism (or mechanisms, discussed later) that limits filtration and thus growth and/or drag forces that lead to dislodgement. Bryozoans Bryozoans are active suspension feeders that are usually colonial. Membranipora membranacea showed steadily decreasing potential growth rate (temporal difference in circular area where radii were considered maximum distance from ancestrula to colony edge) over the flow speed range 0 to 16 cm/s (Eckman & Duggins, 1993). The encrusting cheilostome Parasmittina jeffreysi exhibited decreased growth rate between 4 cm/s to 18 cm/s, consistent with the findings of Eckman & Duggins (1993), but showed an increase around 24 cm/s (Genovese & Witman, 1999). If the response of filtration rate to flow speed is responsible for the response of growth rate to flow speed, this would be one of the only anomalous findings. Concerning particle-size selectivity with increasing flow speed, Okamura (1987) observed Bugula neritina ingested more large particles than small particles at slow flow (1 to 2 cm/s), and 30 while this pattern of selection did not change, ingestion of large particles decreased and ingestion of small particles increased around flow speeds of 10 to 12 cm/s. This observation concurs with the present study for S. plicata, but Okamura also found the opposite for a smaller species of bryozoan, B. stolonifera. This species ingested more small particles than large particles at slow flow, but at higher flow speeds, more large particles were ingested. Okamura suggested the most likely explanation for this anomaly is a switch from ciliary pumping to increased tentacular activity for the smaller species. These results were observed when bryozoans were fed single-sized latex beads during a trial. However, when fed evenly mixed solutions of the different-sized beads, both species showed increased ingestion of large-particles and decreased ingestion of small-particles around flow speeds of 10 to 12 cm/s while there was consistently more selection of large particles at both flow speeds (Okamura, 1990). In addition to different feeding structures and behavior between colonial bryozoans and solitary ascidians, one possible explanation for the difference between Okamura’s observations and those in this study is that I suspect large-particle filtration was greatly enhanced at slow flow speeds for S. plicata by enhanced capture of large particles at the incurrent region of the inhalant siphon, whereas Okamura observed no significant deposition of particles at either flow speed. Influence Of Drag At this point, it is necessary to separate the influence of drag on FR with increasing body size and increasing flow. Allometry dominates the filtration response to increasing body size at all flow speeds, but the height of the curve (as determined by the exponent, b) decreases in a flow dependent manner. This indicates that scope for increasing filtration rate in larger ascidians is less in higher flow than in slow flow. The influence of drag on FR, specifically with increasing flow, is more unfavorable (squared speed relationship to drag as opposed to a linear body size relation). 31 Therefore, drag could likely be responsible for the flow-dependent suppression of filtration in the following ways. Ascidians were clearly reconfigured in flow as a result of drag forces. Deformation of internal feeding structures is possible as the ascidian is bent, and this could reduce filtering efficiency. Reconfiguration also implies alteration of siphon orientation. Knott et al. (2004) found that model ascidians utilized ambient flow to filter more-so at the orientation with flow (laid-over) than at an upright orientation, perpendicular to flow. Further, this reliance on passive flow was shown to increase feeding in Pyura stolonifera (Knott et al., 2004). This finding is contrary to my results where feeding was increasingly suppressed from upright to laid-over orientations. However, because the orientation investigation of Knott et al. was performed at a single flow speed, and during this study, orientations were altered by varying flow, it is still unclear whether changing siphon orientation from upright to laid-over will increase adverse pressure gradients between the siphons or if the development of adverse pressure gradients is a result of simultaneously increasing flow and laying over. E-values describing flexibility of organisms in flow are rare in the literature. The flexibility of S. plicata compares best with that of a streamlined body in the same Re range (-0.5; Vogel, 1984). Other marine organisms in the same Re range are seemingly more flexible. The sea pen, Ptilosarcus gurneyi, was determined to have an E-value of -1.14 (Best, 1988), and E-values of -1.28 and -1.66 were respectively determined for the hydroid, Abietenaria rigida (Harvell & LaBarbera, 1985), and two species gorgonian, Pseudopterogorgia acerosa and P. americana (Sponaugle & LaBarbera, 1991). Nevertheless, it is reasonable that streamlined bodies are better adapted to ambient conditions than bluff bodies, and this adaptation may afford S. plicata some advantage over non-flexible suspension feeders (Vogel, 1984; Koehl, 1984; Denny et al., 1985; Koehl, 1999). 32 Summary The particle-size-dependent filtration rates of S. plicata are allometric with respect to body size and unimodal with respect to flow speed. Allometry with body mass is well established in the literature, and the allometric exponents reported in this study are comparatively lower than those found in other investigations. This was shown to be due to the confounding adverse effects of increasing flow speed. The likely mechanisms for flow-dependent suppression of filtration rate include deformation of internal feeding structures due to drag, and/or increased adverse pressure gradients between the siphons as a result of drag or flow itself. Interestingly, maximum filtration occurs at moderate flow speeds (~ 12 cm/s) when this ascidian filters particles in the range 1.25 to 6.03 µm. Maximum filtering of large particles (6.03 to 100 µm) occurs at low flow speeds (~ 0). The most likely reason for this shift in maximum filtering of different size particles is enhanced filtration of large particles at low speeds due to enhanced capture of large particles in the incurrent region of the inhalant siphon. These findings will be very useful to those interested in bioremediation since peak filtration of bacterial-size particles appears to occur at different flow speeds than peak filtration of particles the size of large phytoplankton. These experiments could be carried out again in more depth to gain a better understanding of the precise flow speeds where peak particle-size-dependent filtration occurs. Additionally, investigations into the relationship between filtration rate and growth rate with flow should also be performed for this ascidian and other suspension feeders. Optimal filtering conditions may differ in flow regime for fouling ascidians and other suspension feeders. If true, this knowledge could aid those who commercially culture marine organisms. The connection between life and environment is progressively made clearer through studies of how feeding is affected by ambient hydrodynamic conditions. Further understanding 33 of the feeding response to those conditions has been gained with additional regard to food-particle size and interactions among each of the biological and hydrodynamic variables. 34 REFERENCES Al-Deen, M. F. N., Samways, A. L., & Brunn, H. H., 1996, Water Flow Studies Using Split-film Anemometry, Measurement Science And Technology, Vol. 7, pp. 1529-1535 Al-Garni, A. M., 2007, Low Speed Calibration Of Hot-wire Anemometers, Flow Measurement And Instrumentation, Vol. 18, pp. 95-98 Best, B. A., 1988, Passive Suspension Feeding In A Sea Pen: Effects Of Ambient Flow On Volume Flow Rate And Filtering Efficiency, Biological Bulletin, Vol. 175, pp. 332-342 Brunn, H. H., 1996, Hot-film Anemometry In Liquid Flows, Measurement Science And Technology, Vol. 7, pp. 1301-1312 Aslan-Yilmaz, A., Okuş, E., & Övez, S., 2004, Bacteriological Indicators Of Anthropogenic Impact Prior To And During The Recovery Of Water Quality In An Extremely Polluted Estuary, Golden Horn, Turkey, Marine Pollution Bulletin, Vol. 49, pp. 951-958 Cahalan, J. A., Siddall, S. E., & Luckenbach, M. W., 1989, Effects Of Flow Velocity, Food Concentration And Particle Flux On Growth Rates Of Juvenile Bay Scallops, Argopecten irradians, Journal Of Experimental Marine Biology And Ecology, Vol. 129, pp. 45-60 Clarke, R. D., Finelli, C. M., & Buskey, E. J., 2009, Water Flow Controls The Distribution And Feeding Behavior Of Two Co-occurring Coral Reef Fishes: II. Laboratory Measurements, Coral Reefs, Vol. 28, pp. 475-488 Coughlan, J., 1969, The Estimation Of Filtering Rate From The Clearance Of Suspensions, Marine Biology, Vol. 2, pp. 356-358 Denny, M. W., Daniel, T., & Koehl, M., 1985, Mechanical Limits To The Size Of Wave-swept Organisms, Ecological Monographs, Vol. 55, pp. 69–102 Draughon, L., 2009, Nature Makes The Best Technology, <http://www.bioremediator.com/index.html>, Retrieval Date: 10/30/2009 Eckman, J., & Duggins, D., 1993, Effects Of Flow Speed On Growth Of Benthic Suspension Feeders, Biological Bulletin, Vol. 185, pp. 28-41 Eckman, J., Peterson, C., & Cahalan, J., 1989, Effects Of Flow Speed, Turbulence And Orientation On Growth Of Juvenile Bay Scallops, Argopecten irradians concentricus, Journal Of Experimental Marine Biology And Ecology, Vol. 132, pp. 123-140 Fiala-Médioni, A., 1978, Filter-feeding Ethology Of Benthic Invertebrates (Ascidians). IV. Pumping Rate, Filtration Rate, Filtration Efficiency, Marine Biology, Vol. 48, pp. 243-249 Finelli, C. M., Hart, D. D., & Fonseca, D. M., 1999, Evaluating The Spatial Resolution Of An Acoustic Doppler Velocimeter And The Consequences For Measuring Near-bed Flows, Limnology And Oceanography, Vol. 44, No. 7, pp. 1793-1801 Flood, P. R., & Fiala-Médioni, A., 1981, Ultrastructure And Histochemistry Of The Food Trapping Mucous Film In Benthic Filter-Feeders (Ascidians), Acta Zoologica, Vol. 62, pp. 53-65 Fries, J. S., & Trowbridge, J. H., 2003, Flume Observations Of Enhanced Fine-particle Deposition To Permeable Sediment Beds, Limnology And Oceanography, Vol. 48, No. 2, pp. 802-812 Fuller, P., 2009, Styela plicata, USGS Nonindigenous Aquatic Species Database, Gainesville, Florida, <http://nas.er.usgs.gov/queries/FactSheet.asp?speciesID=1293>, Retrieval Date: 10/15/2009 Genovese, S. J., & Witman, J. D., 1999, Interactive Effects Of Flow Speed And Particle Concentration On Growth Rates Of An Active Suspension Feeder, Limnology And Oceanography, Vol. 44, No. 4, pp. 1120-1131 Goodbody, I., 1974, The Physiology Of Ascidians, In: F. S. Russel & M. Yonge (Eds.), Advances In Marine Biology, Academic Press, New York, New York, USA, pp. 2-149 Harvell, D. C., & LaBarbera, M., 1985, Flexibility: A Mechanism For Control Of Local Velocities In Hydroid Colonies, Biological Bulletin, Vol. 168, pp. 312-320 Hecht, S., 1916, The Water Current Produced By Ascidia atra Lesueur, Journal Of Experimental Zoology, Vol. 20, No. 3, pp. 429-434. Holmes, N., 1973, Water Transport In The Ascidians Styela clava Herdman And Ascidiella aspersa (Müller), Journal Of Experimental Marine Biology And Ecology, Vol. 11, pp. 1-13 Howey, R., 1998, Tunicates with Salad on the Side, Micscape Magazine, <http://www.microscopy-uk.org.uk/mag/indexmag.html?http://www.microscopy-uk.org.uk/mag/ artnov98/rhtunic.html>, Retrieval Date: 10/15/2009 Kimura, S., & Itoh, T., 2007, Biogenesis And Function Of Cellulose In The Tunicates, In: R. M. Brown, Jr. & I. M. Saxena (Eds.), Cellulose: Molecular And Structural Biology - Selected Articles On The Synthesis, Structure, And Applications Of Cellulose, Springer, Dordrecht, The Netherlands, pp. 217-236 Kirby-Smith, W. W., 1972, Growth Of The Bay Scallop: The Influence Of Experimental Water Currents, Journal Of Experimental Marine Biology And Ecology, Vol. 8, pp. 7-18 Klumpp, D. W., 1984, Nutritional Ecology Of The Ascidian Pyura stolonifera: Influence Of Body Size, Food Quantity And Quality On Filter-feeding, Respiration, Assimilation Efficiency And Energy Balance, Marine Ecology – Progress Series, Vol. 19, pp. 269-284 36 Knott, N. A., Davis, A. R., & Buttemer, W. A., 2004, Passive Flow Through An Unstalked Intertidal Ascidian: Orientation And Morphology Enhance Suspension Feeding In Pyura stolonifera, Bilogical Bulletin, Vol. 207, No. 3, pp. 217-224 Koehl, M., 1984, How Do Benthic Organisms Withstand Moving Water?, American Zoologist, Vol. 24, pp. 57–70 Koehl, M., 1999, Ecological Biomechanics Of Benthic Organisms: Life History, Mechanical Design And Temporal Patterns Of Mechanical Stress, The Journal Of Experimental Biology, Vol. 202, pp. 3469-3476 Kott, P., 1972, Some Sublittoral Ascidians In Moreton Bay, And Their Seasonal Occurrence, Memoirs Of The Queensland Museum, Vol. 16, pp. 233-260 LaBarbera, M., & Vogel, S., 1976, An Inexpensive Thermistor Flowmeter For Aquatic Biology, Limnology And Oceanography, Vol. 21, pp. 750-756 Lambert, C. C., & Lambert, G., 1998, Non-indigenous Ascidians In Southern California Harbors And Marinas, Marine Biology, Vol. 130, pp. 675-688 Lenihan, H. S., Peterson, C. H., & Allen, J. M., 1996, Does Flow Speed Also Have A Direct Effect On Growth Of Active Suspension-feeders: An Experimental Test On Oysters, Limnology And Oceanography, Vol. 41, No. 6, pp. 1359-1366 Okamura, B., 1987, Particle Size And Flow Velocity Induce An Inferred Switch In Bryozoan Suspension-feeding Behavior, Biological Bulletin, Vol. 173, pp. 222-229 Okamura, B., 1990, Particle Size, Flow Velocity, And Suspension-feeding By The Erect Bryozoans Bugula neritina And B. stolonifera, Marine Biology, Vol. 105, No. 1, pp. 33-38 Peters, R. H., 1984, Methods For The Study Of Feeding, Grazing And Assimilation By Zooplankton, In: J. A. Downing, & F. H. Rigler (Eds.), A Manual On Methods For The Assessment Of Secondary Productivity In Fresh Waters, 2nd ed. IBP handbook 17, Blackwell Scientific Publications, Oxford, England, pp. 336-412 Petersen, J., 2007, Ascidian Suspension Feeding, Journal Of Experimental Marine Biology And Ecology, Vol. 342, pp. 127-137 Petersen, J., & Riisgård, H., 1992, Filtration Capacity Of The Ascidian Ciona intestinalis And Its Grazing Impact In A Shallow Fjord, Marine Ecology Progress Series, Vol. 88, pp. 9-17 Petersen, J., & Svane, I., 2002, Filtration Rate In Seven Scandinavian Ascidians: Implications Of The Morphology Of The Gill Sac, Marine Biology, Vol. 140, pp. 397-402 Pich, J., 1966, Theory Of Aerosol Filtration By Fibrous And Membrane Filters, In: C. N. Davies (Ed.), Aerosol Science, Academic Press, London, England, pp. 223-285 37 Randløv, A., & Riisgård, H., 1979, Efficiency Of Particle Retention And Filtration Rate In Four Species Of Ascidians, Marine Ecology – Progress Series, Vol. 1, pp. 55-59 Riisgård, H., 2001, On Measurement Of Filtration Rates In Bivalves – The Stony Road To Reliable Data: Review And Interpretation, Marine Ecology Progress Series, Vol. 211, pp. 275-291 Robbins, I. J., 1984, The Regulation Of Ingestion Rate, At High Suspended Particulate Concentrations, By Some Phleobranchiate Ascidians, Journal Of Experimental Marine Biology And Ecology, Vol. 82, pp. 1-10 Robinson, H. E., Finelli, C. M., & Buskey, E. J., 2007, The Turbulent Life Of Copepods: Effects Of Water Flow Over A Coral Reef On Their Ability To Detect And Evade Predators, Marine Ecology Progress Series, Vol. 349, pp. 171-181 Rubenstein, D., & Koehl, M., 1977, The Mechanisms Of Filter Feeding: Some Theoretical Considerations, American Naturalist, Vol. 111, pp. 981-994 Schmidt-Nielsen, K., 1997, Animal Physiology: Adaptation And Environment, 5th ed., Cambridge University Press, New York, New York, USA Sherrard, K. M., & LaBarbera, M., 2005, Form And Function In Juvenile Ascidians. II. Ontogenetic Scaling Of Volumetric Flow Rates, Marine Ecology Progress Series, Vol. 287, pp. 139-148 Shimeta, J., & Jumars, P., 1991, Physical Mechanisms And Rates Of Particle Capture By Suspension Feeders, Oceanography And Marine Biology: An Annual Review, Vol. 29, pp. 191-257 Spielman, L., 1977, Particle Capture From Low-speed Laminar Flows, Annual Review Of Fluid Mechanics, Vol. 9, pp. 297-319 Sponaugle, S., & LaBarbera, M., 1991, Drag-induced Deformation: A Functional Feeding Strategy In Two Species Of Gorgonians, Journal Of Experimental Marine Biology And Ecology, Vol. 148, pp. 121-134 Vogel, S., 1981, Life In Moving Fluids: The Physical Biology Of Flow, Willard Grant Press, Boston, Massachusetts, USA Vogel, S., 1984, Drag And Flexibility In Sessile Organisms, American Zoologist, Vol. 24, No. 1, pp. 37-44 Weast, R. C., & Selby, S. M., 1967-1968, Handbook Of Chemistry And Physics 48th Edition, Chemical Rubber Co., Cleveland, Ohio, U.S.A., F-3 Welschmeyer, N., 1994, Fluorometric Analysis Of Chlorophyll a In The Presence Of Chlorophyll b And Pheopigments, Limnology And Oceanography, Vol. 39, No. 8, pp. 1985-1992 38 Wildish, D. J., & Kristmanson, D. D., 1997, Benthic Suspension Feeders And Flow, Cambridge University Press, Cambridge, United Kingdom and New York, NY, USA Wildish, D. J., Kristmanson, D. D., Hoar, R. L., DeCoste, A. M., McCormick, S. D., & White, A. W., 1987, Giant Scallop Feeding And Growth Responses To Flow, Journal Of Experimental Marine Biology And Ecology, Vol. 113, pp. 207-220 Wildish, D. J., Kristmanson, D. D., & Saulnier, A. M., 1992, Interactive Effect Of Velocity And Seston Concentration On Giant Scallop Feeding Inhibition, Journal Of Experimental Marine Biology And Ecology, Vol. 155, pp. 133-143 Wildish, D. J., & Miyares, M. P., 1990, Filtration Rate Of Blue Mussels As A Function Of Flow Velocity: Preliminary Experiments, Journal Of Experimental Marine Biology And Ecology, Vol. 142, pp. 213-220 Wildish, D. J., & Saulnier, A. M., 1993, Hydrodynamic Control Of Filtration In The Giant Scallop, Journal Of Experimental Marine Biology And Ecology, Vol. 174, pp. 65-82 39