Document 11827247
advertisement

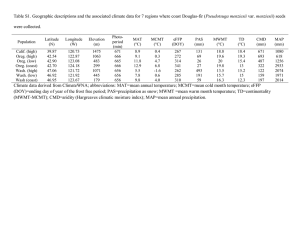
Quan%fying thermal constraints on carbon and water fluxes in a mixed-­‐conifer Sky Island ecosystem Zev Braun1, Rebecca L. Minor2, Daniel Po8s3, and Greg A. Barron-­‐Gafford2 1Grinnell College, Grinnell, IA; 2B2 Earthscience, Biosphere 2, University of Arizona, Tucson, AZ; 3Buffalo State College, Buffalo, NY E d d y IntroducPon Significance: * Understanding the consequences of climate change has been a growing concern for scienPsts and the public around the world. The rising temperatures and diminishing water availability associated with this trend are criPcally influenPal. Despite this issue, research surrounding the ramificaPons for higher temperatures in Southwestern high elevaPon ecosystems remains sparse. * One such ecosystem is the semi-­‐arid Sky Island Forest. Sky Islands are regions that could drasPcally change in the coming years, as this subalpine ecosystem evolved around -­‐ and now depends on -­‐ the winter snowpack. However, if in the future higher temperatures inhibit snow accumulaPon, this ecosystem will inevitably collapse whilst the aquifers that supply water resources to surrounding towns and ciPes will diminish further. * A drier climate could yield a host of alteraPons to the diversity and health of the ecosystem. More than likely, wildfire frequency will increase as seasonal soil moisture and annual precipitaPon levels decrease; drought tolerant tree species will dominate the forest, while many of the unique organisms that comprise Sky Island habitat will become scarce. IllustraPng the response of these isolated tree species to thermal stress will thus aid in modeling the enigmaPc repercussions of climate change within these elevated Southwestern areas. C o v a r i a n c e T o w e r Methods This experiment was conducted in the forest immediately surrounding the eddy covariance tower on Mt. Bigelow in the Santa Catalina Mountains, northeast of Tucson, Arizona. The forest consisted mainly of Pinus ponderosa (Ponderosa Pine), Pseudotsuga menziesii (Douglas Fir), Pinus leiophylla (Chihuahua Pine), and Pinus ayacahuite (Mexican White Pine). q Trees were selected based on their maturity, good health, and proximity to the eddy covariance tower in order to incorporate representaPve samples of each species into the study. Four P. ayacahuite, P. menziesii and Yellow Pine (both P. leiophylla and P. ponderosa) samples were assessed under the temperature curves. q Leaf samples were prepared to mimic the condiPon observed within the canopy itself. Healthy branches were clipped from each tree for each response curve (Figure 3), immediately placed in water (drawn from the same area), and re-­‐cut to eliminate the presence of air bubbles in the water column. Branches then acclimated in sunlight for one hour, before a twig was chosen and secured inside the lighted conifer chamber of the LI-­‐6400 portable photosynthesis system (LI-­‐COR, Lincoln, NE, USA) (Figure 2). q Temperature was adjusted using the LI-­‐6400 to the best of its ability. Measurements spanned a roughly 20°C range of air temperatures. Once stabilized, the two infrared gas analyzers within the instrument were matched, and the data logged. Block temperature increased in 5°C increments unPl a maximum potenPal temperature was reached, while allowing Pme for the leaf sample to stabilize at each increment. Measurements were conducted at constant CO2 concentraPon (400 ppm) and light intensity (1800 µmol m-­‐2 s-­‐1). q Topt was determined by locaPng the temperature at maximum photosynthesis, while Amax was derived from the magnitude of photosynthesis at this peak value. We calculated WUE using the formula: A/E , where A is photosynthesis and E is transpiraPon. Ω50 was found by locaPng the temperature at ½Amax and subtracPng that from Topt. Predic%ons: Due to the variety of parameters that affect plant funcPonality, concurrent with a lack of research concerning this ecosystem, it is difficult to predict which species will be best suited to extended periods of high temperatures (>30°C) and able to proliferate more readily. Some trees may a8ain higher rates of photosynthesis when water is abundant, expressing their maximum photosynthePc capability (Amax). AddiPonally, temperature opPma (Topt) may differ among species and through seasonal moisture changes, selecPng certain species over others. Furthermore, species that can remain highly producPve over a wider range of temperatures might assimilate more carbon than heat sensiPve species, and thus predominate when temperature fluctuaPons occur (Ω50). Temperature curves administered to samples will mimic diurnal fluctuaPons to idenPfy successful species under this condiPon. AlternaPvely, some trees may be able to assimilate more carbon for every water molecule lost (Water Use Efficiency [WUE]), allowing them to perform photosynthesis efficiently under drier condiPons. These possibiliPes necessitate wet and dry measurements to accurately predict forest stand dynamics when considering Amax, Topt, Ω50, and WUE. Figure 2. Experimental assembly with LI-­‐6400s Figure 3. Clipping branches from the canopy Figure 1. Preparing to clip a branch specimen Results: How do these tree species respond to soil moisture influx over the monsoon? Do some develop resistance to thermal stress due to this change? Figure 4. Response curves Temperature opPma for all species decreased when moisture levels under dry (lep) and wet increased (Table 1). Water abundance reduced plant sensiPvity to (right) condiPons temperature fluctuaPons and dramaPcally enhanced Amax. Figure 5. E. Darker TranspiraPon (E) rose with rising temperatures under both dry and wet condiPons, but increased more colors are wet measurements exponenPally during the monsoon. Each species saw 150% 140% higher rates of E with more soil moisture content (dark lines), and began at about the same level of E at the start of the response curve (~15ºC). * Yellow Pine had the most substanPal gain in E over the temperature spectrum, while P. menziesii had the most gradual rise. * P. menziesii maintained the lowest rates in E throughout the range of temperatures. 440% * P. ayacahuite performed the highest rates of maximum net photosynthesis (Amax) under pre-­‐monsoon condiPons, while P. menziesii expressed the lowest Amax. * Under monsoon condiPons, Yellow Pine outdid P. ayacahuite in photosynthePc producPvity, and wet condiPons boosted producPvity of P. menziesii by 590%. * P. menziesii presented the highest temperature opPmum, while Yellow Pine had the lowest temperature opPmum. Table 1. A representaPon of some parameters to photosynthesis Figure 6. WUE. Darker Under wet condiPons these species are more efficient at uPlizing water resources in high colors are wet measurements temperatures. However, WUE differs at colder temperatures. * WUE increased for both P. ayacahuite and Yellow Pine during the monsoon (36% and 23% respecPvely). * Overall, WUE diminished in P. menziesii during the monsoon (by approximately 13%). P. menziesii saw a decrease in WUE in colder weather, but an increase during warmer temperatures. Conclusions • Our findings support the predicPon that increased moisture content at the site allowed every species to tolerate higher temperatures and achieve a higher maximum photosynthePc rate (Amax increased) (Figure 4). • Moisture reduced forest sensiPvity to temperature extremes and fluctuaPons (Ω50), which substanPally magnified total photosynthePc producPvity (Table 1). • In parPcular, wet condiPons enhanced Amax the most dramaPcally for Pseudotsuga menziesii, elevaPng photosynthesis by 590%, while Ω50 grew most substanPally for Pinus ayacahuite (by 180%; Table 1). • Carbon sequestraPon decreased under high temperatures (>30°C) among all tree species that comprise this ecosystem, regardless of soil moisture status (Figure 4). • Interspecific differences in temperature opPma (Topt) elucidated possible species dominance predicPons for seasonal and gradual temperature changes (Figure 4). More than likely, P. menziesii may out-­‐perform Yellow Pine and P. ayacahuite in the event that temperatures rise. However, if monsoon rains fail to accumulate, P. menziesii may never undergo the 590% surge in Amax, and instead remain at subsistence levels of photosynthesis (Figure 4). • TranspiraPon rates (E) rose exponenPally in response to temperature during the monsoon, while WUE tended to elevate above the values seen in pro-­‐ monsoon condiPons (Figures 5 and 6). This trend supports the idea that as these species evolved where monsoon rains frequented the ecosystem, trees gained physiological and mechanical adaptaPons to exploit heavy rainfall. • Together, this data will enable the parameterizaPon of models to forecast the producPvity and composiPon of semi-­‐arid Sky island ecosystems, which may face future water deficits and thermal upheavals Acknowledgements: I sincerely appreciate the inspiraPon, encouragement, and generosity expressed by Dr. Greg Barron-­‐Gafford. Many thanks to Daniel Po8s for his anecdotes and advice regarding graduate school. Rebecca L. Minor, my trusted compatriot, was instrumental in gathering the data necessary for this project. Maggie and Ashley helped considerably to process leaf samples and keep laboratory work exciPng.