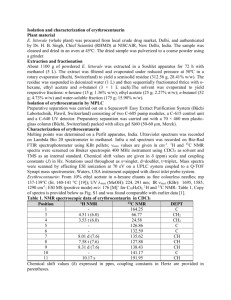
AN ABSTRACT OF THE THESIS OF June 3. 2005.
advertisement

AN ABSTRACT OF THE THESIS OF Bingnan Han for the degree of Doctor of Philosophy in Pharmacy presented on June 3. 2005. Title: Natural Products Studies of the Marine Cyanobacteria Lynqbya semiplena and Lyngbya majuscula Abstract Redacted for Privacy This thesis details my investigations of marine cyanobacterial natural products that resulted in the discovery of eighteen new secondary metabolites. Isolation and characterization of these unique molecules were carried out using different chromatographic techniques and by careful analyses of ID and 2D NMR data, respectively. Preliminary bioassays of the crude organic extract of the marine cyanobacterium Lyngbya semiplena collected from Papua New Guinea in 1999, showed good activity in the brine shrimp toxicity model at 10 ppm. Guided by this assay, seven new anandamide-like fatty acid amides, semiplenamides A to G, together with four cyclic depsipeptides, wewakpeptins A to 0, were identified. Due to the structural resemblance of the novel ethanolamide derivatives (semiplenamide A-G) with anandamide, (N-arachidonoyl-ethanolamine), an endogenous agonist of cannabinoid receptors compounds, semiplenamide A-G were tested on the well characterized proteins of the endocannabinoid system: 1) the "central" cannabinoid GB1 receptors; 2) the anandamide membrane transporter (AMT), which is responsible for anandamide cellular uptake; and 3) the fatty acid amide hydrolase (FAAH), which catalyses anandamide hydrolysis. Three showed modest potency in displacing radiolabeled anandamide from the cannabinoid receptor (GB 1), and one was a modest inhibitor of the anandamide membrane transporter (AMT). The wewakpeptins were tested for cytotoxicity to NCI-H460 human lung tumor and neuro-2a mouse neuroblastoma cells. Intriguingly, wewakpeptin A and B were approximately 10-fold more toxic than C and 0 to these cell lines. Lyngbya majuscula has been recognized as a chemically and biologically rich strain. Five new lyngbyabellin analogs, lyngbyabellins E-1, along with the known compound dolabellin, originally isolated from sea hare Dolabella auricularia, were identified from a 2002 Papua New Guinea collection of the marine cyanobacterium Lyngbya majuscula. The lyngbyabellins were tested for cytotoxicity to NCI-H460 human lung tumor and neuro-2a mouse neuroblastoma cells and had LC50 values between 0.2 and 4.8 pM. Intriguingly, lyngbyabellin E and H appeared to be more active against the H460 cell line with 0.4 pM and 0.2 pM, respectively, compared to LC50 LC50 values of values of 1.2 and 1.4 pM in the neuro-2a cell line. Lynbyabellin I was the most toxic to neuro-2a cells (LG50 0.7 pM), whereas lyngbyabellin G, was the least cytotoxic of all compounds to either cell line. On the basis of this limited screening, it appears that lung tumor cell toxicity is enhanced in the cyclic representatives, and overall potency is increased in those containing an elaborated side chain. Additionally, two new cytotoxins, aurilides B and C, which are closely related to aurilide, originally isolated from the sea hare Do/abe/Ia auricularia, have been identified from the same extract where the lyngbyabellins E-1 were isolated. Aurilides B and C were tested for cytotoxicity to NCI-H460 human lung tumor and neuro-2a mouse neuroblastoma cells. Interestingly, aurilide B was approximately 4-fold more toxic than C to these cell lines. The LC50 for Aurilide B was 0.01 pM and 0.04 pM for neuro-2a and H460 cells, respectively, and 0.05 pM and 0.13 pM, respectively, for aurilide C. Aurilide B was evaluated in the NCI 60 cell line panel and found to exhibit a high level of growth inhibition in leukemia, renal, and prostate cancer cell lines with a Gl less than 10 nM. Aurilide B showed net tumor cell killing activity in the NCI's hollow fiber assay, an in vivo model for assessing a chemical's anticancer activity. ©Copyright by Bingnan Han June 3, 2005 All Rights Reserved NATURAL PRODUCTS STUDIES OF THE MARINE CYANOBACTERIA LYNGBYA SEMIPLENA AND LYNGBYA MAJUSCULA by Bingnan Han A THESIS submitted to Oregon State University in partial fulfillment of the requirements for the degree of Doctor of Philosophy Presented June 3, 2005 Commencement June, 2006 Doctor of Philosophy thesis of Bingnan Han presented on June 3, 2005. APPROVED: Redacted for Privacy Major Professor, representing Pharmacy Redacted for Privacy Dean of the CoIIge of Pharmacy Redacted for Privacy Dean of the' ràdi1ate School I understand that my thesis will become part of the permanent collection of Oregon State University libraries. My signature below authorizes release of my thesis to any reader upon request. Redacted for privacy Bingnan Han, Author ACKNOWLEDGMENTS I would like to express my gratitude to my major advisor, Dr. William Gerwick, and my committee members, Dr. Philip Proteau, Dr. George Constantine, Dr. Michael Schimerlik, and Dr. Claudia Maier for their support and academic guidance throughout my graduate career at Oregon State University. I would also like to acknowledge Brian Arbogast and Jeff Morre for their extensive mass spectral work, and Roger Kohnert for his valuable NMR assistance. My appreciation also goes to Dr. Vincenzo Di Marzo (Endocannabinoid Reserch Group, Istituto di Chimica Biomolecolare, Italy), and Dr. Susan Mooberry (Southwest Foundation for Biomedical Research, San Antonio, Texas). I am very grateful to my current and former group colleagues for their mentorship, insight, and most importantly, friendship. I would especially like to acknowledge Dr. Kenneth Milligan, Dr. Patricia Flatt, Dr. Kerry McPhail, Dr. Dougals Goeger, and Dr. Harald Gross for their continual inspiration and motivation, Dr. Omar Mohamed Sabry, and Eric Andrianasolo for their friendship and encouragement throughout the years. Completion of this degree would have been impossible without them all. Finally, I would like to express my gratitude to my wonderful family, especially My Mother, Ruoli Wang, for their support and encouragement. I am forever indebted to them for their patience and love through these challenging years. CONTRIBUTION OF AUTHORS Anandamide cellular uptake assay, CB1 receptor binding assay, and fatty acid amide hydrolase assay were carried out by Dr. Vincenzo Di Marzo (Endocannabinoid Reserch Group, Istituto di Chimica Biomolecolare, Italy). Cytotoxicity bioassays against human lung cancer and mouse neuro-2a cell lines were carried out by Dr. Douglas Geoger in our laboratory. Actin-disrupting assay was performed by Dr. Susan Mooberry (Southwest Foundation for Biomedical Research, San Antonio, Texas). Collision induced ESI-MS/MS experiments in chapter three of my thesis were performed by Dr. Claudia Maier at the Department of Chemistry, OSU. Dr. Kerry McPhail and Dr. Harald Gross provided various assistance in chapter four and five of my thesis. TABLE OF CONTENTS Page CHAPTER ONE: GENERAL INTRODUCTION HIGHLIGHTS OF MARINE NATURAL PRODUCTS .................................. I BIOACTIVE SECONDARY METABOLITES FROM MARINE CYANOBACTERIA............................................................... 14 THE SYMBIOSIS OF MARINE INVERTEBRATES AND MARINE CYANOBACTERIA ........................................................................... 22 GENERAL THESIS CONTENTS ......................................................... 26 REFERENCES ................................................................................ 30 CHAPTER TWO: SEMIPLENAMIDES A G, FATTY ACID AMIDES FROM A PAPUA NEW GUINEA COLLECTION OF THE MARINE CYANOBACTERIUM LYNGBYA SEMIPLENA ABSTRACT ...................................................................................... 38 INTRODUCTION .............................................................................. 38 RESULTS AND DISCUSSION ........................................................................ 40 EXPERIMENTAL ............................................................................................. 53 REFERENCES ................................................................................ 57 CHAPTER THREE: THE WEWAKPEPTINS, CYCLIC DEPSIPEPTIDES FROM A PAPUA NEW GUINEA COLLECTION OF THE MARINE CYANOBACTERIUM LYNGBYA SEMIPLENA ABSTRACT .................................................................................... 58 INTRODUCTION .................................................................................. 59 RESULTS AND DISCUSSION ............................................................ 60 EXPERIMENTAL ............................................................................. 76 REFERENCES ................................................................................ 80 CHAPTER FOUR: ISOLATION AND STRUCTURE OF LYNGBYABELLIN DERIVATIVES FROM A PAPUA NEW GUINEA COLLECTION OF THE MARINE CYANOBACTERIUM LYNGBYA MAJUSCULA ABSTRACT.................................................................................... 82 TABLE OF CONTENTS (Continued) Page INTRODUCTION .............................................................................. 83 RESULTS AND DISCUSSION ............................................................. 85 EXPERIMENTAL .............................................................................. 102 REFERENCES ................................................................................. 106 CHAPTER FIVE: ISOLATION, STRUCTURE, AND BIOLOGICAL ACTIVITY OF AURILIDE B AND C FROM A PAPUA NEW GUINEA COLLECTION OF THE MARINE CYANOBACTERIUM LYNGBYA MAJUSCULA ABSTRACT .................................................................................... 108 INTRODUCTION .............................................................................. 109 RESULTS AND DISCUSSION ............................................................ 110 EXPERIMENTAL .............................................................................. 120 REFERENCES ................................................................................. 123 CHAPTER SIX: CONCLUSION ................................................................... 125 BIBLIOGRAPHY ........................................................................................ 129 APPENDICES ........................................................................................... 136 APPENDIXA ................................................................................... 137 APPENDIXB ................................................................................... 148 LIST OF FIGURES Page 1.1 Marine toxins 1.2 Largest non-proteinaceous marine toxin .................................................. 4 1.3 Marine toxins ...................................................................................... 5 1.4 Anticancer marine natural products ........................................................ 7 1.5 Anticancer marine natural products ....................................................... 10 1.6 Antiinflammatory marine natural products ............................................... 11 1.7 Anti-Alzheimer marine natural products .................................................. 12 1.8 Antimalarial marine natural products...................................................... 13 1.9 Anti-HIV marine natural products .......................................................... 14 1.10 Chemical structures of natural products 30-35 ......................................... 20 1.1 1 Chemical structures of natural products 36-46 ......................................... 21 1.12 Chemical structures of natural products 47-54 ......................................... 26 1.13 Chemical structures of natural products 55-63 ......................................... 27 11.1 1H and 13C NMR spectra of semiplenamide A (1) ...................................... 42 11.2 COSY spectrum of semiplenamide A (1) ................................................. 43 11.3 HSQC spectrum of semiplenamide A (1) ................................................. 44 11.4 HMBC spectrum of semiplenamide A (1) ................................................. 45 11.5 Determination of the geometry at C6-C7 by a 1H-1H .3 decoupling experiment ........................................................................ 46 11.6 Determination of the geometry at C2-C3 by a 1 D NOE experiment............... 46 111.1 1H NMR spectrum of wewakpeptin A (1) ................................................. 61 111.2 13C NMR spectrum of wewakpeptin A (1)................................................ 61 111.3 HSQC spectrum of wewakpeptin A (1) .................................................... 62 111.4 HMBC spectrum of wewakpeptin A (1) ................................................... 63 111.5 COSY spectrum of wewakpeptin A (1) ................................................... 64 111.6 HSQC spectrum of wewakpeptin A (1) ................................................... 65 111.7 Key fragments from collisionally induced ESI-MS/MS experiments with wewakpeptin A (1).and wewakpeptin C (3) ......................................... 68 111.8 1H NMR spectrum of wewakpeptin B (2) .................................................. 70 111.9 13C NMR spectrum of wewakpeptin B (2) ................................................. 70 111.10 1H NMR spectrum of wewakpeptin C (3) ................................................. 71 LIST OF FIGURES (Continued) Page 111.11 13C NMR spectrum of wewakpeptin C (3) ................................................ 72 111.12 1H NMR spectrum of wewakpeptin D (4) ..................................................72 111.13 13C NMR spectrum of wewakpeptin D (4) .................................................... 72 111.14 Cytotoxicity of wewakpeptins A-D (1-4) ..................................................... 76 IV.1 H and 13C spectra of Iyngbyabellin E (1) .................................................... 87 IV.2 HSQC spectrum of IV.3 HMBC spectrum of lyngbyabellin E (1) .................................................... 90 lV.4 HSQC-TOCSY spectrum of lyngbyabellin E (1) ......................................... 91 IV.5 COSY spectrum of IV.6 Diagram of all possible rotamers for the two possible epimers lyngbyabellin lyngbyabellin E (1) ..................................................... 89 E (1) .................................................... 92 26S,27S (A series) and 26R,27S (B series) of lyngbyabellin E (1) ................. 94 IV.7 1H and 13C spectra of lyngbyabellin F (2) ................................................. 96 lV.8 1H and 13C spectra of lyngbyabellin G (3) ................................................. 96 IV.9 1H and 13C spectra of Iyngbyabellin H (4) .................................................. 99 IV.10 1H and 13C of spectra of IV. 11 Effect of Iyngbyabellin E (1) on the actin V.1 1H and 13C spectra of aurilide B (1) ........................................................ 111 V.2 COSY spectrum of aurilide B (1) ........................................................... 113 V.3 HSQC spectrum of aurilide B (1) ........................................................... 114 V.4 HMBC spectrum of aurilide B (1) ........................................................... 115 V.5 HSQC-TOCSY spectrum of aurilide B (1) ................................................ 116 V.6 Key COSY, HMBC, and NOE correlations for 1 V.7 '5 V.8 1H and 13C spectra of aurilide C (2) ......................................................... 119 (s-R) lyngbyabellin 1(5) ................................................. 99 cytoskeleton of A-I 0 cells .............. 102 ........................................ 117 values (ppm) of MTPA esters of aurilide B (1) obtained in CD3CN ....... 118 LIST OF TABLES Page 1.1 Selected marine natural products currently in clinical trials ......................... 7 11.1 1H and 13C NMR data for semiplenamides A (I) and B (2) in CDCI3 ............ 47 11.2 1H and 13C NMR data for semiplenamide C (3) in CDCI3 ........................... 49 11.3 1H and 13C NMR data for semiplenamides D (4) and E (5) in CDCI3 ............ 50 11.4 1H and 13C NMR data for semiplenamides F (6) and G (7) in CDCI3 ............ 51 111.1 NMR data for wewakpeptins A (1) and B (2) in CDCI3 ..............................66 111.2 NMR data for wewakpeptins C (3) and D (4) in CDCI3 .............................. 73 IV.1 NMR data for lyngbyabellin E (1) in CDCI3 ............................................. 88 IV.2 NMR data for lyngbyabellins F (2) and G (3) in Cod3 .............................. 97 IV.3 NMR data for Iyngbyabellins H (4) and 1(5) in CDCI3 .............................. 100 lV.4 Cytotoxicity of compounds 1-5 ........................................................... 101 V.1 NMR data for aurilides B (1) and C (2) in C6D6 ...................................... 112 LIST OF ABBREVIATIONS AEA Arachidonoylethanolamide (Anandamide) Ala Alanine AMT Anandamide membrane transporter Ara-A Adenine arabinoside Ara-C Cytosine arabinoside br Broad CB1 Central cannabinoid receptors Cl Chemical Ionization CNS Central Nervous System COSY 1H-1H Chemical Shift Correlation Spectroscopy d Doublet OCAMO 7 ,7-dichloro-3-acyloxy-2-methyloctanoate Dhiv a, 13-d ihydroxyisovaleric acid Dhoaa 2,2-Dimethyl-3-hydroxy-octanoic acid Dhoya 2,2-Dimethyl-3-hydroxy-7-octynoic acid Dmhha 2,2-Dimethyl-3-hydroxy-hexanoic acid OMSO Dimethylsulfoxide ElMS Electron-Impact Mass Spectrometry EtOAc Ethyl Acetate FAAH Fatty acid amide hydrolase FAB Fast Atom Bombardment FDAA 1 -fluro-2,4-dinitrophenyl-5-L-alaninamide GC Gas Chromatography Gly Glycine HETLOC Hetero half-filtered TOCSY Hila 2-hydroxyisoleucic acid Hiva 2-Hydroxy-isovaleric acid HIV Human Immunodeficiency Virus H MBC Heteronuclear Multiple-Bond Correlation Spectroscopy Hmha 5-Hydroxy-3-methyl-heptanoic acid HPLC High Performance Liquid Chromatography HSQC Heteronuclear Single Quantum Coherence LIST OF ABBREVIATIONS (Continued) IC Inhibitory Concentration lie Isoleucine lR Infrared Spectroscopy Lac Lactic acid LD Lethal Dose m Multiplet Maba 2-Methyi-3-amino-butanoic acid Mapa 2-Methyl-3-amino-pentanoic acid Me Methyl MeOH Methanol MS Mass Spectroscopy MIT 3-(4, 5-dimethylthiazol-2-yl)-2, 5-diphenyltetrazolium bromide NMR Nuclear Magnetic Resonance NOESY Nuclear Overhauser Exchange Spectroscopy NRPS Non-Ribosomal Peptide Synthetase Phe Phenylalanine PKS Polyketide Synthase PMSF Phenylmethylsuiphonylfluoride Pro Proline q Quartet ROESY Rotating Overhauser Exchange Spectroscopy RI Room Temperature s Singlet SCUBA Self-Contained Underwater Breathing Apparatus sPLA2 secreted Phospholipase A2 SRi 4171 6A I H-Pyrazole-3-carboxamide, 5-(4-chlorophenyl)- I -(2,4-dichlorophenyl)-4- methyl-N-i -piperidinyl-, monohydrochloride t Triplet TEA Trifluoroacetic acid TOCSY Total Correlation Spectroscopy tR Retention Time Tyr Tyrosine UV Ultraviolet Spectroscopy Val Valine VLC Vacuum liquid chromatography NATURAL PRODUCTS STUDIES OF THE MARINE CYANOBACTERIA LYNGBYA SEMIPLENA AND LYNGBYA MAJUSCULA CHAPTER ONE GENERAL INTRODUCTION Nature has continuously provided mankind with broad and structurally diverse natural products which have a variety of purposes including use as food, fragrances, pigments, insecticides, and medicines. The use of natural products for the treatment of human ailments has its beginnings dating back to ancient times in both Eastern and Western cultures. Traditionally, higher plants and, since the discovery of the penicillins, terrestrial microorganisms have proven to be the richest sources of natural drugs that are indispensable, especially for the treatment of fatal diseases such as cancer and microbial infections. Currently, there is a pressing need for new therapeutic agents and this is due mainly to the resurgence of pathogenic microorganisms and parasites that have developed resistance to traditional chemotherapies. However, due to the increasing rates of deforestation and rediscovery of known compounds, scientists are beginning to focus their attention on the marine realm as an alternative source of novel and useful natural products. Oceans comprise over 70% of our world's surface and harbor a tremendous variety of flora and fauna. Although still quite young by many standards, the field of marine natural products has already proven to be a productive source for biologically active natural products. Coupled with the development of SCUBA in the 1960s, and more recently submersible vehicles, these have allowed relatively easy accessibility of both shallow and deep-water marine organisms for studies by natural products chemists. In contrast to their terrestrial counterparts, many marine derived secondary metabolites are structurally complex with unique functionalities. This is due in part to the differences in the physicochemical nature of the two environments where higher pressures, lower temperatures, lack of light as well as high ionic concentrations present in sea water may account for the biosynthesis of highly functionalized and unusual molecules in these organisms. It is because of these structural novelties that natural products chemists hope to uncover new classes of therapeutic agents with unique pharmacophores which may lead to defining novel sites of action in proteins and enzymes that are involved in human illnesses. HIGHLIGHTS OF MARINE NATURAL PRODUCTS Historically, the first recorded research on marine natural products began in the early 1950s with the pioneering work of Bergman, which led to the discoveries of the nucleosides spongouridine (1) and spongothymidine (2) from marine sponges collected off the coast of Florida.1 This initial serendipitous discovery led to the development of Ara-A (3) and Ara-C (4), which are based on these sponge nucleosides and are clinically available today. Ara-C (4) is an anticancer drug used for the treatment of acute myelocytic leukemia and non-Hodgkin's lymphoma while the antiviral drug, Ara-A (3), is used for the treatment of herpes HO HO OH o HO HO OH Spongouridine (1) Spongothymidine (2) j HO OH Ara-A (3) infections.2'3 HO oJ HO OH Ara-C (4) Since the early 1950s, more then 10,000 secondary metabolites have been defined from marine organisms.4 At the early stage, pioneering research was mainly concerned with marine toxins, in part because of the numerous poisonings of American soldiers in the Pacific during World War II. Many of the marine toxins appeared to be macromolecules, belonging mainly to the polyether structural class. Some of the well known marine polyether toxins are: brevetoxin B (5), isolated from the dinofiagellate Gymnodinium breve;6 ciguatoxin (6), initially reported from moray eels and a toxic constituent implicated in ciguateric seafood poisoning;7 maitotoxin (7), isolated from Gambierdiscus toxicus;8'9 and okadaic acid (8), a potent tumor promoter and major causative agent of diarrhetic shellfish poisoning, found from two 3 species of dinoflageflates, Prorocentrum lima and Dinophysis species.10'11 Of these macromolecules, maitotoxin is the largest (MW 3422), and most likely, the most lethal non-proteinaceous toxin (LD of Ca. 50 ng/kg in mice) characterized from a marine microorganism to date. Palytoxin (9), a Complex polyol, is another well known toxic macromolecule isolated from the zoanthid, Palythoa toxicus.12 Many of these toxic macromolecules have also proven to be invaluable as molecular tools for dissecting cellular processes by molecular pharmacologists.13'14 Brevetoxin B (5) Figure I. 1. Marine toxins. Meanwhile, the increasing need for drugs able to control new illnesses or resistant strains of microorganisms stimulated the search for new biomedicals from marine organisms. Thus, a main objective of most marine natural products programs is to discover secondary metabolites with a broad spectrum of therapeutic utilities, such as: antibiotic, antifungal, cytotoxic, neurotoxic, antimitotic, antiviral, and antineoplastic activities. In more recent years, new targets have been added to the general screening, for example: AIDS, immunosuppression, anti-inflammation, Alzheimer's disease, aging processes and some tropical diseases.15 This is especially so in the area of cancer research where numerous marine natural products are being screened. Although none of the discoveries since the 1970's has led to a clinical product, there are a number of potential candidates currently under intense investigation. Maitotoxin (8) Figure I. 2. Largest non-proteinaceous marine toxin with a MW of 3422. 5 Among the more promising anticancer natural products are ecteinascidin 743 (10) and bryostatin 1 (11) (Tablel. I). The marine alkaloid ecteinascidin 743 (ET-743) was isolated from the Caribbean ascidian, Ectinascidia turbinata,16' 17 and is by far the most advanced compound in the late stages of phase II clinical trials as an anti- cancer drug. Ecteinascidin's structure is consistent with a natural microbial origin (e.g. the saframycins). Indeed, there are two patents for bacterial symbionts of the tunicate E. turbinata. The first focuses on the isolation of the ET-743 producing microbe,18 while the second uses 16S rDNA sequences to identify the endosymbiont as Endoecteinascidia frumentensis, the apparent producer of the ecteinascidins.19 Okadaic acid (8) I-lu ),, HO LJOH 'OH OH HO HO OH 0H Palytoxin (9) Figure I. 3. Marine toxins. More recently, ET-743 has been reported to bind in the minor groove of DNA to induce an unprecedented bend in the DNA helix towards the major groove.20 The multi-faceted mechanism of action of ET-743 includes interference with the cellular transcription-coupled nucleotide excision repair to induce cell death, and cytotoxicity which is independent of p53 status yet occurs with MDR elicitation.21'22 Overall, advanced ovarian, breast and mesenchymal tumors which had been heavily pretreated with platinum/taxanes showed greatest response to ET-743 in phase I trials.23'24 In phase II trials, ET-743 was most effective in patients with refractory soft tissue sarcoma (STS), ovarian and breast cancer. However, difficulties in establishing the drug's efficacy in STS prevented its approval in 2003. Meanwhile, EU's Committee for Proprietary Medicinal Products has granted ET-743 orphan drug status for the treatment of refractory ovarian cancer. It is still in Phase Ill trials in the US and the EU, and it will probably be the first marine-derived anti-tumor agent to be commercialized. Bryostatin 1 (11) is a polyketide first isolated in minute quantity from the bryozoan, Bugula neritina collected from the Gulf of California in the 1970's.25 It is currently in phase Il clinical trials for the treatment of various leukemias, lymphomas, melanoma, and solid tumors. The biological profiles of bryostatin 1 (11) are interesting in that not only does it inhibit tumor growth but it stimulates bone marrow growth, increasing red blood cell production.26 Recent research has found that this cytotoxic macrolide shows potential in the treatment of ovarian and breast cancer and it also enhances lymphocyte survival in patients undergoing radiation treatment.27 To date, eighteen bryostatins have been defined and some members of this family (e.g. bryostatin 5 and 8) exhibit biological activities equivalent to or better than bryostatin 1 (11).28 Bryostatin 1 (11) Ecteinascidin 743 (10) OHO 0 NH N 0 NH 0 0 OCH3 Dehydrodidemnin B (12) (= Aplidine) Figure I. 4. Anticancer marine natural products. One major obstacle that may prevent the development of bryostatin 1 (11) to a clinically useful anticancer agent is the supply issue. The yield of bryostatin 1 (11) from B. neritina is organism.29 low, in the range of about 1.4 pg per gram of wet weight of the Another complication is the unpredictable production of the molecule by the bryozoan. One of the ways to meet the demand issue is through aquaculture which was undertaken by CalBioMarine Technologies in California, and has since proven to be a commercially viable source of the molecule. However, a recent report presented evidence that suggests a symbiotic bacterial origin for bryostatin 1 (11) production.30 This finding may open the way for genetic manipulation of the biosynthetic genes and fermentation for increased production of this potent compound. Table I. 1. Selected marine natural products currently in clinical trials21'31'32 Compound (Figure number) Source Organism Ecteinascidin 743 (Yondelis®) Dolastatin 10 Ecteinascidia turbinate (tunicate) (possible bacterial source) Dolabella auricularia / Symploca sp. (mollusc/cyanobacterium) Bugula neritina (bryozoan) Bryostatin 1 Synthadotin (1LX651, Dolastatin 15 derivative) Kahalalide F Squalamine Dehydrodidemnin B (Aplidine®) Cemadotin (LU 103793, Dolastatin 15 derivative) Soblidotin (TZT-1027, Dolastatin 10 derivative) E7389 (Halichondrin B derivative) NVP-LAQ824 (Psammaplin derivative) Discodermolide HTI-286 (Hemiasterlin derivative) LAF-389 (Bengamide B derivative) KRN-7000 (Agelasphin derivative) Dolabella auricularia/ Symploca sp (synthetic analog) Elysia rufescens I Bryopsis sp. (mollusc/green alga) Squalus acanthias (shark) Current Status Phase Il/Ill Phase II Phase II Phase II Phase II Phase II Trididemnum solidum (tunicate, synthetic) (possible bacterial/cyanobacterial source) Dolabella auricularial Symploca sp (synthetic analog) Dolabella auricularia/ Symploca sp (synthetic analog) Halichondria okadal (sponge, synthetic) Phase I Psammaplysilla sp. (sponge, synthetic) Phase I Discodermia dissolute (sponge) Phase I Cymbastella sp. (synthetic analog of sponge metabolite) Jaspis digonoxea (sponge, synthetic) Phase I Agelas mauritianus (sponge, synthetic) Phase I Phase II Phase II Phase II Phase I The dolastatins are a series of cytotoxic peptides that were originally isolated in very low yield by Pettit's group as part of their work on marine invertebrates from the Indian Ocean mollusk, Do/abe/Ia auricuIaria.2lM. Due to the potency and mechanism of action of dolastatin 10 (13), a linear depsipeptide which was shown to be a tubulin interactive agent, the drug entered Phase I clinical trials in the 1990s under the auspices of the NCI.'37 Dolastatin 10 progressed through to Phase II trials as a single agent, but although tolerated at the doses used, which were high enough to give the expected concentrations in vivo to inhibit cell growth, it did not demonstrate significant antitumor activity in a Phase II trial against prostate cancer in men. As a result, many derivatives of the dolastatins have been synthesized with TZT-1027 (Auristatin PE or Soblidotin) (14) now in Phase II clinical trials in Europe, Japan and the USA. Two derivatives of dolastatin 15 (15), a peptide related to dolastatin 10 in structure and origin, are known as Cemadotin or LU-I 03793 (16) and Synthadotin or ILX65I (17). These were entered into Phase I clinical trials by Abbott GmBH under the Knoll division for treatment of breast cancer.21 By using tritiumlabeled dolastatin 15 as a bioprobe, Hamel's group at NCI recently reported that the ymca domain in tubulin may well be composed of a series of overlapping domains rather than being a single entity. Another important marine-derived compound, aplidine (= dehydrodidemnin B) (12), is a synthetic analog of a cyclic antiproliferative depsipeptide, didemnin B. Didemnin B was isolated from the Caribbean tunicate Trididemnum solidum,4° and was the first marine natural product to enter clinical trial as an antitumor agent.41 However, it was dropped due to a disappointing performance in phase II testing. The dehydro-derivative, dehydrodidemnin B (12) is ten times more effective than didemnin B, and is less toxic. Apladin®, a registered trademark of aplidine (12), was found to be more selective towards leukemia and lymphoma cells than towards normal cells. In addition, the activity of Apladin® was found to be independent of other anti-cancer drugs commonly used in leukemia and lymphoma, suggesting that Apladin® may be effective in cases that have proved unresponsive to other agents. The success of aplidine in phase I trials has led to its current evaluation in Phase II trials against solid tumors. 10 NNNN I H N I OCH N' S \=1 DolastaUn 10 (13) I OHO 0CH30 TZT-1027 (14) (Auristatin PE or Soblidotin) Nj OCH3 0 Dolastatin 15 (15) LU 103793 (16) (Cemadotin) 1LX651 (17) (Synthadotin) Figure I. 5. Anticancer marine natural products: the dolastatins. In addition to the discovery of marine natural products with anticancer properties, marine organisms have also yielded a number of important antiinflammatory agents. Many of these compounds act as phospholipase A2 inhibitors, preventing release of arachidonic acid from the lipids of cell membranes.42 11 Arachidonic acid is involved in the biosyntheses of many proinflammatory compounds via enzymes such as cyclooxygenase, lipoxygenase, or monooxygenase. The quintessential compound from marine sources exhibiting activity as an anti- inflammatory agent was the sponge metabolite manoalide (18). This compound was originally reported by Scheuer's group as an antibiotic isolated from the marine sponge Luffariella variabilis.43 The groups of Jacobs and Dennis independently established that this compound was a potent inhibitor of the enzyme phospholipase A2. The original compound was licensed to Allergan and placed into clinical trials as a topical antipsoriatic with a company code name of AGN-1 90093. It advanced to Phase II, but work on the natural product stopped as sufficient quantities of the compound would not pass through the skin using the formulation developed for the trials.21 In addition, pseudopterosins A (19) and E (20), from the Caribbean gorgonian, Pseudopterogorgia elisabethae, are two diterpene glycosides showing promising in vitro and in vivo activities as anti-inflammatory agents.46'47 LL1o\ 0 io Manoalide (18) OH Pseudopterosin A (19) Pseudopterosin E (20) Figure I. 6. Antiinflammatory marine natural products. In 1971, Kern et al. reported the isolation of hoplonemertine toxin, a compound that subsequently became known as anabaseine (21 )48 A variety of synthetic analogues of the basic structure were made by Kem's group, and one of 12 which, GTS-21 (22), has been shown to have cytoprotective and memory enhancing effects, perhaps due to the ability to displace the binding of nicotinic ligands and to affect the function of the a413 and a7 subtypes of this pharmacokinetic trials,50 receptor.49 Following GTS-21 was licensed to the Japanese company, Taiho, by the University of Florida for clinical trials as a potential anti-Alzheimer's agent and is currently in Phase I trials in both Europe and USA under the auspices of Taiho. (LN K) Anabaseine (21) GTS-21 (22) Figure I. 7. Anti-Alzheimer marine natural products. One of the emerging trends in many infectious organisms is their resistance to the standard drugs. This is especially so in malarial infections by the protozoan parasite, Plasmodium species. Many resistant strains of this parasite are being uncovered and treatments of such infections by traditional drugs are proving ineffective. However, recent research has suggested that the next generation of new antimalarial chemotherapeutic agents might come from marine organisms. In an initial screening of extracts from marine sponges for antimalarial properties, Acanthella klethra yielded an active component, axisonitrile 3 (23), which is a bicyclic spiro-sesquiterpene with an isonitrile group.51 This compound possesses potent in vitro antimalarial activity with no detectable cytotoxicity. It was further revealed that marine natural products containing NC, NCS, and NCO functionalities exhibited antimalarial properties.51 A new class of 13-carboline alkaloids from marine sponges, known as the manzamines, is also gaining attention as potent antimalarial agents. Four members of this series, manzamine A (24), its enanatiomer (26), 8hydroxymanzamine A (25), and a manzamine dimer, neo-kauluamine (27) gave 13 encouraging results in preliminary in vivo testing. These preliminary data also indicated that the manzamines are more active and less toxic than the current available drugs artemisinin and chloroquine.52'53 Axisonitnle 3 (23) Manzamine A (24) R = H 8-Hydroxymanzamine A (25) R = OH ent-8-Hydroxymanzamine A (26) neo-Kauluamine (27) Figure I. 8. Antimalarial marine natural products. In the area of antiviral research, especially anti-HIV (human immunodeficiency virus), two main leads have been found from marine sources. The marine natural compound ilimaquinone (28) is currently being investigated as a chemotherapeutic in HIV infection.M.55 This compound acts by targeting at the RNase function of the reverse transcriptase enzyme encoded by HIV. The first natural product that inhibits HIV integrase enzyme was reported from a species of marine sponge belonging to the genus Lame!!aria. The metabolite, lamellarin alpha 20-sulfate (29) was found to inhibit early steps of HIV replication.56 14 OCH3 NaO3SO 0 0 H3C0N\ H3CO Ilimaquinone (28) Figure I. H3COT'OCH3 Lamellarin a 20-sulfate (29) 9. Anti-H IV marine natural products. BIOACTIVE SECONDARY METABOLITES FROM MARINE CYANOBACTERIA The predominant marine organisms studied by natural products chemists are sponges, soft corals, marine algae, sea squirts, and bryozoans. Marine algae are rich in secondary metabolites proven to serve multiple roles through field and laboratory bioassays. They are loosely divided into two groups, macroalgae, which include red (Rhodophyta), brown (Phaeophyceae), and green algae (Chlorophyta) and microalgae, including cyanobacteria (blue-green algae) as well as dinoflagellates (Dinoflagellatea, Protozoa). However, cyanobacteria, the most ancient (ca. 2 x 1O9 years) of the microalgae, have been the richest producers of novel and bioactive secondary metabolites. Cyanobacteria, also known as blue-green algae, are ancient photosynthetic prokaryotes which inhabit a wide diversity of habitats including open oceans, tropical reefs, shallow water environments, terrestrial substrates, aerial environments such as in trees and rock faces, and fresh water ponds, streams and puddles.57 The rich elaboration of biologically-active natural products has assisted some of these organisms to survive in predator-rich tropical reef ecosystems. As a result, tropical marine cyanobacteria, particularly the filamentous forms such as Lyngbya sp. or Symploca sp., have been exciting sources of novel natural products with therapeutic and biotechnological potential. a. Tubulin-binding compounds Symplostatin 1(30) 15 In 1998, the first natural dolastatin 10 analog, symplostatin 1 (30) was discovered by direct isolation from the cyanobacterium Symploca hydnoides. Symplostatin I differs from dolastatin 10 by the addition of a single CH2 in the dolavaline moiety resulting in a terminal N,N-dimethylisoleucine versus the terminal N,N-dimethylvaline.58 This compound exhibited potent cytotoxicity (lC= 0.3 ng/mL) against a human nasopharyngeal carcinoma cell line (KB), compared to < 0.1 ng/mL for dolastatin 10. Symplostatin 1 was also shown to induce microtubule loss by 80% at I ng/mL when tested in A-I 0 cells. It has been concluded that the mechanism of action of symplostatin 1 must be similar if not identical to that determined for dolastatin 1 0. In further analysis, symplostatin I displayed efficacy against a number of cancer cell lines and caused the formation of abnormal mitotic spindles and accumulation of cells in metaphase. These effects were observed at concentrations that elicited only minor loss of interphase microtubules. Cell cycle analysis indicated that symplostatin 1 caused G2/M arrest, a finding consistent with its effects on mitotic spindles. Evaluation of this aspect revealed potent inhibition of both endothelial cell proliferation and invasion, with in vivo efficacy against murine colon 38 and murine mammary 16/C cell lines.59 In addition to the exciting biological activity, this discovery offered further evidence that these potent peptides and depsipeptides are of cyanobacterial origin. Curacin A (31) Not all antimitotic marine natural products have an overt peptidic nature and the highly lipophilic natural product curacin A (31) is a good example. Isolated from a Caribbean collection of the cyanobacterium L. majuscula, the structure of curacin A consists of an interesting thiazoline ring containing a 14-carbon alkyl chain with conjugated diene and terminal olefin, and a methyl substituted cyclopropyl moiety.6° Curacin A was originally shown to be active against a Vero cell line, and subsequent assessment in the NCI 60 cell line assay revealed potent antiproliferative and cytotoxicity with some selectivity for colon, renal, and breast cancer cell lines. It was later shown that treatment of curacin A resulted in the depolymerization of purified tubulin induced by either glutamate or microtubule-associated proteins with an IC= 4.0 pM and lC=6.0 pM. Further testing revealed that curacin A (31) binds tightly at the colchicine site of tubulin. Curacin A stimulates the uncoupled GTPase reaction 16 typical of colchicine site agents, and indirect observations were consistent with curacin A binding rapidly and dissociating slowly from tubulin.61 Additionally, curacin A was shown to inhibit formation of the Cys239-Cys354 cross-link in f3-tubulin.62 Furthermore, under conditions ideal for tubulin polymerization, high concentrations of curacin A induced formation of aberrant tubulin polymers appearing similar to a twisted cable of fine spiral filaments.63 Based on these results a series of semi- synthetic analogs were produced to explore the structure activity relations in this new drug class. The structural modifications included reduction and E-to-Z transitions of the otefinic bonds in the alkyl side chain; disruption of and configurational changes in the cyclopropyl ring, thiazoline moiety, and substituent modifications at the Cl 0 methyl- and Cl 3 methoxy groups. This work revealed that the most crucial portions of curacin A for tubulin interaction seems to be the thiazoline ring and the side chain at least through C4, and the portions of the side chain including the C9-Cl 0 olefinic bond, and the ClO methyl group.M Recently, some new directions have emerged with construction of synthetic combinatorial libraries using solution phase and fluorous scavenging techniques, resulting in new analogs with improved bioavailability and efficacy.65 b. Actin-binding compounds Hectochlorin (32) Hectochlorin (32), was isolated from the marine cyanobacterium L. majuscula collected from Hector Bay, Jamaica, and Boca del Drago Beach, Bocas del Toro, Panama.66 The planar structure was deduced by one- and two-dimensional NMR spectroscopy, and X-ray crystallography was used to determine the absolute stereochemistry. Hectochlorin was initially shown to be a potent antifungal agent in preliminary antimicrobial bioassays. Further testing showed that Ptk2 cells (derived from the rat kangaroo, Potorous tridactylus) treated with hectochiorin showed an increase in the number of binucleated cells as result of arrest of cytokinesis. This result is consistent with an interference with the actin cytoskeleton. Hectochlorin is similar in its activity to jasplakinolide (or jaspamide) (33) in its ability to promote hyperpotymerization of actin.67'68 The main biochemical difference between jasplakinolide and hectochioriri is that while the former can displace fluorescently labeled phalloidin from actin polymers, the latter is unable to do so, suggesting that 17 the two have distinct interactions with actin. Hectochlorin was also evaluated against the in vitro panel of 60 different cancer cell lines (National Cancer Institute) and showed strong potency towards cell lines in the colon, melanoma, ovarian and renal sub-panels. It had a flat dose-response curve against most cell-lines which is characteristic of compounds that are antiproliferative but not directly cytotoxic.66 Lyngbyabellins A (34) and B (35) The lyngbyabellins (34,35) were isolated from collections of L. majuscula from the South Pacific and Caribbean, and bear structural resemblance to hectochlorin and dolabellin (36).6971 The structure of lyngbyabellin A was determined using 20 NMR techniques and its absolute stereochemistry was determined by chiral HPLC analysis. Lyngbyabellin A (34) exhibited IC values of 0.03 pg/mL and 0.50 pg/mL against KB cells (human nasopharyngeal carcinoma cell line) and LoVo cells (human colon adenocarcinoma cell line), respectively. It was also shown to disrupt the microfilament network in fibroblastic A-10 cells at concentrations between 0.01-5.0 pg/mL. However, when treated with a higher concentration of lyngbyabellin A many cells became binucleate, an observation which is consistent for compounds inhibiting cytokinesis. The structure of Iyngbyabellin B (35) was determined using a combination of ID and 20 NMR spectroscopy and its stereochemistry was proposed using a combination of NMR and chiral GC/MS analysis. Lyngyabeflin B was found to be less toxic than Iyngbyabellin A with IC values of 0.10 pglmL and 0.83 pg/mL against KB and LoVo cell lines, respectively. Lyngbyabellin B had the same effect as hectochlorin on PtK2 cells, in that an increased number of binucleate cells were observed when the cells were treated with 1 0 pM of the agent. These findings suggest that actin is the likely cellular target of the lyngbyabellins. c. Neurotoxic compounds Antillatoxin (37) The crude extract of a Curaçao collection of L. majuscula was found to be highly ichthyotoxic and molluscicidal. Fractionation and subsequent purification led to the discovery of the potent lipopeptide ichthyotoxin, antillatoxin.72 The structure of antillatoxin was elucidated using several spectroscopic methods including HR FABMS, IR and NMR. Antillatoxin (37) is one of the most ichthyotoxic metabolites 18 isolated to date from a marine cyanobacterium (LD in potency only by the brevetoxins.73 = 0.05 pglmL), and is exceeded Initial pharmacological studies showed that antillatoxin was acutely neurotoxic and rapidly induced morphological changes in rat cerebellar granule neurons (CGC's), including blebbing of neurite membranes. However, the toxicity was remarkably reduced when cells were treated with NMDA receptor antagonists like dextrorphan and MK-801, indicating that the toxicity of antillatoxin was mediated through an NMDA receptor- dependent mechanism.74 Antillatoxin was later shown to be a powerful activator of voltage-gated sodium channels and resembled brevetoxin in this respect. But unlike brevetoxin which is known to bind site 5 on the a- subunit of voltage-gated sodium channels, the binding site of antillatoxin is unknown and remains to be identified.75 The unique biological activity of antillatoxin combined with its unusual structure spurred several synthetic efforts, and the total synthesis of antillatoxin was reported by two groups.778 However, spectroscopic analyses of the synthetic versions showed that they were different from "natural antillatoxin". Eventually, the synthetic 4R,5R stereoisomer, which was identical to natural antillatoxin was reported by the Shioiri group.78 In addition to resolving some initial stereochemical issues, the syntheses of several stereoisomers of antillatoxin facilitated SAR studies and a recent report showed that naturally occurring antillatoxin was about 25-fold more potent than any of its C-4 or C5 stereoisomers.79 Kalkitoxin (38) Kalkitoxin (38) was first isolated from a Caribbean collection of L. majuscula and exhibited potent brine shrimp and fish toxicity. Subsequently, the compound was re-isolated in very small quantities from various Caribbean collections of L. majuscula.6° Structural elucidation was accomplished by various 2D NMR methods and stereochemistry was resolved using Marfey's analysis as well as comparison with synthetic standards. Kalkitoxin was toxic to rat CGC's and had an LC =3.86 nM and this effect could be inhibited by the addition of NMDA receptor antagonists.74 There is also evidence indicating that kalkitoxin is a blocker of voltage-sensitive Na channel in mouse neuro-2a cells (EC = 1 nM). In an inflammatory disease model, kalkitoxin was found to inhibit the release of 1L43-induced sPLA2 (secreted phospholipase A2), a key enzyme in the inflammatory cascade (IC = 27 nM). 19 Jamaicamides A-C (39-41) A strain of L. majuscula collected from Hector's Bay, Jamaica yielded a series of novel and functionalized lipopeptides, jamaicamides A-C (39-41 ).81 The structure of the jamaicamides was deduced using a combination of NMR spectroscopy including an ACCORD-1,1 -ADEQUATE experiment. Jamaicamaide A was particularly unique in its presence of an acetylenic bromide functionality. The jamaicamides exhibited cytotoxicity against H-460 human lung cell line and mouse neuro-2A neuroblastoma cell lines with an LC50 15 pM for both cell lines and showed sodium channel blocking activity at a concentration of 5 pM. From a combined precursor feeding study and a molecular genetics approach a fascinating mixed NRPS and PKS pathway was deduced. d. Protein kinase C activators Lyngbyatoxins A-C (42-44) Lyngbyatoxin A (42) was isolated from a Hawaiian shallow-water variety of L. majuscula and is believed to be responsible for a condition known as 'swimmer's itch'. The structure of lyngbyatoxin A is closely related to the teleocidins, metabolites produced by several Streptomyces sp.82 The structure of lyngbyatoxin A was elucidated using high resolution mass spectrometry and NMR analyses and is the first indole alkaloid to be reported from a marine cyanobacterium. This highly inflammatory compound had an LD100 =0.3 mg/kg in mice which was comparable to the toxicity of the teleocidins. It was also found to be ichthyotoxic and exposure to 0.15 pg/mL resulted in death of fish in 30 minutes. Ensuing pharmacological studies showed that lyngbyatoxin A (42) and teleocidin were potent tumor promoters in a manner similar to that of the phorbol esters when tested in mice.83 To confirm whether lyngbyatoxin was produced by the cyanobacterium or merely associated with it, collections of L. majuscula from Kahala Beach, Hawaii were re-examined and this led to the isolation of two related metabolites, lyngbyatoxin B (43) and lyngbyatoxin C (44)84 In an effort to understand the interaction of lyngbyatoxin A with PKC, several analogs were synthesized and it was found that the lactam ring is essential for PKC activation and that the hydrophobic group attached to C-7 actually downregulates PKC production.85'86 Very recently, the Gerwick group has reported the molecular 20 cloning of the lyngbyatoxin biosynthetic gene cluster from a Kahala Beach, Oahu, collection of L. majuscula, as well as the biochemical characterization of a novel aromatic prenyltransferase that transfers a geranyl group as the final step in the biosynthesis of lyngbyatoxin A (42).87 e. Other bioactive cyanobacterial metabolites Apratoxin A (45) bouillonhi isolated from Apra Bay, Guam initially mis-identified as L. majuscula was found to produce a very potent cytotoxin, apratoxin A (45).88 The L. structure of this mixed polyketide-peptide was deduced using a variety of 2D-NMR techniques, and the amino acid configurations were determined using chiral HPLC analysis. Apratoxin exhibits an IC50 = 0.52 nM and 0.36 nM against KB and LoVo cells, respectively but its exact mode of action at present is uncharacterized. 0 o__-_... 0 H N:*,, OCH3 i-f' V 'Curacin A (31) Symplostatin 1 (30) OH Ly il__i HN 0 "S BrNZ)% H Hectochlorin (32) CICI Jasplakinolide (33) HN HN HC70 Lyngbyabeflin A (34) 0 CT8 0 HN N HN H(LQ Lyngbyabellin B (35) Figure I. 10. Chemical structures of natural products and derivatives 30-35 discussed in text. 21 cI CI 0 o OH 0 OCH3 Dolabelljn (36) N'f' AntiUatoxjn (37) Kalkitoxjn (38) OCH3 Jamaicamjde A (39) R=Br Jamaicamide B (40) R=H 'NO cO OCH3 NO (J 0 Jamaicamide C (41) NNOO HO"' 0 0 NJ.... N-2° c) Apratoxin A (45) I H Lyngbyatoxin A (42) Lyngbyatoxin B (43) Lyngbyatoxin C (44) OCH3: I N rrt N'S 'I Barbamide (46) Figure I. 11. Chemical structures of natural products and derivatives 36-46 discussed in text. The overall unusual structure of apratoxin A and its potent activity, prompted several synthetic efforts and the total synthesis of apratoxin was reported by Chen et al.89 22 Further pharmacological studies along with the synthesis of several structural analogs may enable the determination of the biological target of this potent molecule. Barbamide (46) Barbamide (46) was obtained from the lipid extract of a Curaçao collection of L. majuscula and possesses several unique structural elements including a trichioromethyl group.9° Standard spectroscopic techniques were used for the structural elucidation of barbamide and stereochemistry was deduced by a combination of degradation and biosynthetic methods. The presence of several structurally intriguing elements in barbamide led to a detailed examination of its biosynthesis including the cloning and characterization of the barbamide biosynthetic gene cluster.91'92 Preliminary bioassays indicated that barbamide possesses anti- molluscicidal activity (LC = 10.0 pg/mL). However, the compound was found to be inactive in other assays and the full extent of its biological properties remains unknown. THE SYMBIOSIS OF MARINE INVERTEBRATES AND MARINE CYANOBACTERIA Marine invertebrates are largely sessile, filter-feeding organisms that contain complex assemblages of symbiotic microorganisms. Thus, when biologically active compounds are isolated from marine invertebrate sources, it is always uncertain if the invertebrate host or an associated microorganism is the actual producer. Observation that some metabolites occur in unrelated genera of marine invertebrates, as well as the isolation of related compounds from microbial sources, provides evidence that some natural products are actually produced by symbionts rather than their invertebrate hosts.30'93 For example, the sea hare, D. auricularia has yielded several cytotoxic agents that share similarity with cyanobacterial counterparts, including the anticancer agent dolastatin 10 (13) and the cytotoxic agent, dolabellin (36). The isolation of a dolastatin-lO and related compounds, including symplostatin 1 (30), from the marine cyanobacteria Symploca spp., and the dolabellin-like compounds, 23 Iyngbyabellin A and B from L. majuscula, suggests that sea hares obtain and accumulate bioactive compounds as a result of their cyanobacterial diet. Similarities between structures of cyanobacterial origin have also been found in filter-feeding invertebrates as well. For example, the antifungal compound, majusculamide C (47) was originally isolated from a deep water collection of L. majuscula and subsequently re-isolated from the sponge, Ptilocau/is trachys.94'95 In parallel, metabolites closely related to majusculamide C were also isolated from the sea hare Do/abel/a auricularia, including dolastatin 11(48) and dolastatin 12 (49)97 Furthermore, a study of several Guam collections of L. majuscu/a and L. majuscula/Sty/ocheilus ca/cico/a yielded a re-isolation of dolastatin 12, its c-i 5 epimer (50), and two closely related compounds, lyngbyastatin 1 (51) and its C-i5 epimer (52) providing more direct evidence of their cyanobacterial origin. Many didemnid family ascidians have also been shown to produce biologically active cyclic peptides, including the patellamides (e.g. patellamide A (53))and trunkamide (54)9899 Because many of these cyclic peptides resemble cyanobacterial metabolites and it is known that ascidians house cyanobacterial symbionts of the genus Proch!oron, there is a high likelihood that Prochloron is the producer of these cyclic peptides. However, chemical localization studies have demonstrated that the patellamides accumulate in the tunic tissue of the ascidian and are not associated with the cyanobacterial symbiont.10° In light of these contradictory results, it cannot be ruled out that the compounds are in fact produced and secreted by the cyanobacterial symbiont where they would then accumulate in the ascidian tissues. Advancements in our understandings of the biosynthetic pathways required to produce cyclic pepetides and other secondary metabolites will undoubtedly help to resolve these conflicting issues. Experiments are quickly being developed to assess the biosynthetic capacities housed within hosts and their symbiotic partners. In the case of Lissoclinum pate//a, Schmidt et al, recently identified an NRPS gene sequence from a mixture of the ascidian and its symbiotic Prochioron spp. Based on BLAST sequence homology and overall G/C base content, the NRPS domain appears to be of cyanobacterial origin, providing genetic evidence that the Proch/oron symbiont possesses the biosynthetic capacity to make complex metabolites. Evidence for NRPS-Iike genes in the tunicates has yet to be found. 24 Perhaps the best studied sponge-symbiont relationship involves the sponge of the genus Dysidea. Dysidea sp. have an abundance of symbiotic cyanobacteria, predominantly Oscillatoria spongelliae.102 In fact, the resulting cyanobacterial biomass can account for up to 40% of the total sponge biomass! Furthermore, within the genus Dysidea, it appears that different species have an obligate requirement for specific populations of microbial symbionts. Thacker and Starnes (2003) found that three species of Dysidea from Guam (D. herbacea, D. chiorea, and D. granulosa) each contain a genetically distinct dade of 0. spongeiiae symbionts.102 Concurrently, biochemical analysis has shown that Dysidea spp. are renowned for their production of bioactive metabolites that share high structural similarity with cyanobacterial natural products. The Okinawan sponge D. arenaria yielded the cytotoxic compound arenastatin A (55) that shows high structural similarity with cryptophycin A (56), a cyclic peptide isolated from the terrestrial cyanobacterium Nostoc sp.10105 A related sponge, Dysidea (Lameiodysidea) herbacea and its 0. spongellieae symbiont produces several chlorinated metabolites, including dysidenin (57)1o61o8 The trichloromethyl group of barbamide and the related cyanobacterial metabolites, pseudodysidenin (58), dysidenamide (59), nordysidenin (60), and herbamide B (61), strongly resembles that found in these various Dysidea-derived metabolites.90'109 Cell separation studies using flow cytometry analysis has shown that the chlorinated metabolites are associated with the cyanobacterial symbiont. Most recently, molecular analysis of the sponge-symbiont biomass have resulted in the isolation of biosynthetic genes containing high similarity with the barbamide biosynthetic gene cluster (bar). Fluorescence in situ hybridization (FISH) techniques were used to localize the bar-like halogenase biosynthetic genes to the 0. sponge!!ieae symboint, demonstrating that the biosynthetic potential for compound production is housed within the cyanobacterial symbiont (P.M.Flatt et al, manuscript in press). Recognition that microbial symbionts are often important contributors to the biosynthesis of biologically active compounds opens up additional opportunities to develop sustainable methods for compound production. One could envision the isolation and culturing of the associated microorganism as a viable option. However, isolating and culturing a producing microorganism from an invertebrate host has proven to be a difficult challenge. The microbial communities residing within marine 25 invertebrates are often diverse and many appear to have an obligate requirement for the host. For example, one report characterized a total of 228 different bacterial species, 25 fungi, 3 actinomycetes, and 9 cyanobacterial strains from the sponge Candidaspongia f/abe/late'1° In addition, a limited screening discovered 64 bacterial symbionts from the sponge Aplysina aerophoba and 51 from Theonella swinhoe representing seven different bacterial divisions.111 In some cases, bacteria account for up to 40% of the total sponge biomass.2 Since the early 1990's, several research groups have explored the stable production of bioactive compounds through the isolation and culturing of marine invertebrate-derived symbiotic microorganisms, including unicellular bacteria, filamentous bacteria, and fungi. However, there are only a few limited examples where the same natural product substances have been isolated from both an invertebrate macroorganism and their associated prokaryotic microorganisms, and include makaluvamine A (62) from both Zyzzya sponges 113 and Didymium bahiense myxomycetes,'14 manzamine A (24) from several sponges and a sponge-derived actinomycete,5 the polychlorinated metabolites, neodysidenin (63)108 from D. herbacea and its epimer, pseudodysidenin109 from L. majuscula. In addition, the trichlorinated compound, herbamide B has also been isolated from both the sponge D. herbacea 116 and the cyanobacterium L. majuscula (unpublished, McPhail). Alternatively, the isolation and characterization of biosynthetic genes from host-symbiont populations is a promising new approach to address the compound supply problem. The development of genetic techniques to screen sponge-symbiont metagenomes for genes related to secondary metabolite biosynthesis and then the use of these for in-situ hybridization experiments is a critical first step in determining the identity of the producing organism. However, the abundance of different microbial symbionts within an invertebrate host makes the isolation and characterization of individual biosynthetic gene clusters extremely difficult. From a conservative estimate of only 50 symbiotic species per marine sponge, an estimated 125,000 cosmid-containing colonies (estimated insert size -40kb) would need to be screened from a total genomic library to fully cover the genomic sequences of each organism. However, through the careful selection of metabolites that are likely of microbial origin and a sponge- 26 symbiont system that is limited to a few abundant symbiotic associates, as in the case of the D. herbaceae - 0. sponge!Iiae system, the screening and isolation of biosynthetic gene clusters can be feasible. R1 N HN1 %%- Majusculamide C (47) Dolatstatin 11(48) Dolastatin 12(49) Epidolastatin 12(50) Lyngbyastatin 1(51) Epilyngbyastatm 1(52) R2 R3 R4 R5 CH3 H OCH3 CH3 H CH3 H OCH3 H CH3 CH3 H H H CH3 H Cl-I3 H H CH3 CH3 H OCH3 H CH3 H CH3 OCH3 H H - R R7 H CH3 CH3 CH3 CH3 CF!3 CF!3 CH3 CH3 CH3 CH3 Cl-I3 oo (NS NHN1 HN NI-I 0 Patellamide A (53) Trunkamide A (54) Figure I. 12. Chemical structures of natural products and derivatives 47-54 discussed in text. 27 O>1OHN OHN 0 J) 0 OCH3 Arenastatin A (55) I-1N 0 Cryptophycin A (56) CI3C,-. N-CCI3 H CI3C-IL(_(CCI3 OCH3 H CI3C-. N-.CCI3 0 o O/L N=( Pseudodysidenin (58) R = CH3 Nordysidenin (60) R = H Dysidenin (57) 0 GI3C1NL N NR H2N Dysidenamide (59) H 0N S '-I Herbamide B (61) Makaluvamine A (62) Neodysidenin (63) Figure I. 13. Chemical structures of natural products and derivatives 55-63 discussed in text. GENERAL THESIS CONTENTS Investigation of marine cyanobacteria for their production of novel and bioactive chemistry is the focus of our laboratory and of the research presented within this Ph.D. thesis. The chemistry of marine microalgae, particularly that of the cyanobacterium Lyngbya sp., is extremely rich and diverse. Our extensive collections of marine algae are the results of continual expeditions to different parts of the world, including the Caribbean, Indonesia, Japan, Papua New Guinea, Fiji, South Africa, Panama, and Madagascar. As such, the various chapters outlined in this thesis have unifying themes, which entail isolation, structure elucidation (both planar and stereochemical determinations), and biological characterization of novel secondary metabolites from marine cyanobacteria of the genus Lyngbya collected from Papua New Guinea. This thesis begins with an investigation of semiplenamides A to G, a series of new anandamide-like fatty acid amides, which were isolated from a 1997 Papua New Guinea collection of the marine cyanobacterium Lyngbya semiplena. The planar structures of these lipids were determined using standard 1 D and 2D NMR methods. The relative stereochemistry of the aliphatic portion of the new metabolites was deduced from ID NOE data and 1H-decoupling experiments, while the absolute stereochemistry of the amino alcohol moieties was assigned by chemical derivatization and chiral GC-MS methods. All of these new metabolites displayed toxicity in the brine shrimp model system, three showed modest potency in displacing radiolabeled anandamide from the cannabinoid receptor (CBI), and one was a modest inhibitor of the anandamide membrane transporter (AMT). Chapter three presents four new depsipeptides also isolated from the marine cyanobacterium Lyngbya semiplena collected from Papua New Guinea. The amino and hydroxy acid partial structures of wewakpeptin A-D were elucidated through extensive spectroscopic techniques, including HR-FABMS, 1 D 1H and 13C NMR, as well as 2D COSY, HSQC, HSQC-TOCSY, and HMBC spectra. The sequence of the residues was determined through a combination of multifaceted approaches including ESI-MS/MS, HMBC, ROESY, and a modified ID HMBC experiment. The absolute stereochemistry of each residue was determined by chiral HPLC and chiral GC-MS methods. The wewakpeptins represent an unusual arrangement of amino- and hydroxy-acid subunits relative to known cyanobacterial peptides, and possess a bisester, a 2,2-dimethyl-3-hydroxy-7-octynoic acid (Dhoya) or 2,2-dimethyl-3hydroxyoctanoic acid (Dhoaa) residue, and a diprolyl group reminiscent of dolastatin 15 (15). Wewakpeptin A and B were the most cytotoxic among these four 29 depsipeptides with an LCof approximately 0.4 pM to both the NCI-H460 human lung tumor and the neuro-2a mouse neuroblastoma cell lines. A total of five new lyngbyabellin analogues isolated from the marine cyanobacterium Lyngbya majuscula collected from Papua New Guinea, are presented in chapter four. The structures of lyngbyabellins E-1 were elucidated through extensive spectroscopic techniques, including HR-FABMS, ID 1H and 13C NMR, as well as 2D COSY, HSQC, and HMBC spectra. The absolute configuration of lyngbyabellin E has been ascertained by chiral HPLC and GC/MS analysis of degradation products, in combination with NMR experiments. The lyngbyabellins were tested for cytotoxicity to NCI-H460 human lung tumor and neuro-2a mouse neuroblastoma cells and had LC values between 0.2 and 4.8 pM. Intriguingly, lyngbyabellin E and H appeared to be more active against the H460 cell line with LC values of 0.4 pM and 0.2 pM, respectively, compared to LC values of 1.2 and 1.4 pM in the neuro-2a cell line. Lynbyabellin I was the most toxic to neuro-2a cells (LC 0.7 pM), whereas Iyngbyabellin G, was the least cytotoxic of all compounds to either cell line. On the basis of this limited screening, it appears that lung tumor cell toxicity is enhanced in the cyclic representatives, and overa potency is increased in those containing an elaborated side chain. In the fifth chapter of this thesis, the structural and stereochemical analyses of two new aurilides (B and C) isolated from the marine cyanobacterium Lyngbya majuscula collected from Papua New Guinea, along with their biological activities, will be discussed. The thesis ends with a concluding chapter, summarizing the results presented in the preceding chapters, as well as comments on structural trends that are emerging in cyanobacterial secondary metabolites. General thoughts on the future direction of natural products and the pursuit of the marine environment as a viable source for novel chemistry of medicinal value are also considered. 30 REFERENCE (1) Bergman, W.; Feeney, R. J. J. Org. Chem. 1951, (2) Bodey, G. P.; Freirich, E. J.; Monto, R. W.; Hewlett, J. S. Cancer 16, 981-987. Chemother. 1969, 53, 59-66. (3) Buchanan, R. A.; Hess, F. Science 1980, 127, (4) Jaspars, M. Haivey, A. (5) Halstead, B. W. US Government Printing Office, Washington DC. 1965, I, (6) L., Ed.; 143-171. Wiley: New York, 1998, 1-155. Lin, Y.-Y.; Risk, M.; Ray, S. M.; Van Engen, D.; Clardy, J.; Golik, J.; James, J. C.; Nakanishi, K. J. Am. Chem. Soc. 1981, (7) 65-84. 103, 6773-6775. Murata, M.; Legrand, A.-M.; Ishibashi, Y.; Yasumoto, T. J. Am. Chem. Soc. 1989, 111, 8927-8931. (8) Murata, M.; Nakoi, H.; Iwashita, T.; Matsunaga, S.; Sasaki, M.; Yokoyama, A.; Yasumoto, T. J. Am. Chem. Soc. 1993, 115, 2060- 2062. (9) Norimura, T.; Sasaki, M.; Matsumori, N.; Miata, M.; Tachibana, K.; Yasumoto, T. Angew. Chem. Int. Ed. (10) 1996, 35, 1675-1678. Yasumoto, T.; Oshima, Y.; Sugawara, W.; Fukuyo, Y.; Oguri, H.; Igarashi, T.; Fujita, N. Nip. (11) EngI. Sui. Gak. 1980, 46, 1405-1411. Murakami, Y.; Oshima, Y.; Yasumoto, T. Nip. Sui. Gak. 1982, 48, 69- 72. (12) Moore, R. E.; Scheuer, P. J. Science 1971, 172, 495-498. (13) Mayer, A. M. S.; Hamann, M. T. Marine Biotechnology 2004, (14) Poli, (15) Kelecom, A. An Acad Bras Cienc 1999, 71, 249-263. (16) Wright, A. E.; Forleo, D. A.; Gunawardana, G. P.; Gunasekera, S. P.; M. A. Re. Adv. Mar. Blot. 2002, 7, 6, 37-52. 1-30. Koehn, F. E.; McConnell, 0. J. J. Org. Chem. 1990, 55,4508-4511. (17) Rinehart, K. L.; Holt, T. G.; Fregeau, N. L.; Stroh, J. G.; Kiefer, P. A.; Sun, F.; Li, L. H.; Martin, D. G. J. Org. Chem. 1990, (18) Haygood, M. G.; Salomon, C. E.; Faulkner, D. J. EP1360337 2003. 55, 4512-4515. International Patent 31 (19) Perez, E.; Beatriz, A.; Perez, T.; Velasco, I.; Henriquez, P.; Munoz, M.; Moss, C.; Mckenzie, D. International Patent W02004015143 2004. (20) Zewail-Foote, M.; Hurley, L. J Med Chem 1999, 42, 2493-2497. (21) Newman, D. J.; Cragg, G.M. JNatProd2004, 67, 1216-1238. (22) van Kesteren, C.; de Vooght, M.; Mathot, R.; Schellens, J.; Jimeno, J. M.; Beijnen, J. H. Anti-Cancer Drugs 2003, 14, 487-502. (23) Amador, M. L.; Jimeno, J.; Paz-Ares, L.; Cortes-Funes, H.; Hidago, M. Ann Oncol 2003, 14, 1607-1615. (24) Dileo P, C. P., Bacci G, et al. Proceedings of the 2002 ASCO Annual Meeting. Abstr #1628. 2002. (25) Pettit, G. R.; Herald, C. L.; Doubek, D. L.; Herald, D. L.; Arnold, E.; Clardy, J. J. Am. Chem. Soc. 1982, 104, 6846-6848. (26) May, W. S.; Sharkis, S. J.; Esa, A. H.; Gebbia, V.; Kraft, A. S.; Petht, G. R.; Sensenbrenner, L. L. Proc. Nat!. Acad. Sc!. USA 1987, 84, 8483-8487. (27) Kraft, A. S. J. Nat!. Cancer !nst. 1993, 85, 1790-1792. (28) Kraft, A. S.; Woodley, S.; Pettit, G. R.; Gao, F.; Coil, J. C.; Wagner, F. Cancer Chemother. Pharmaco!. 1996, 37, 271-278. (29) Schaufelberger, D. E.; Pettit, G. R. J. Nat. Prod. 1991, 54, 1265-1270. (30) Haygood, M. G.; Schmidt, E. W.; Davidson, S. K.; Faulkner, D. J. J. Mo!. Microbiol. Biotechno!. 1999, 1, 33-43. (31) Salomon, C. E.; Magarvey, N. A.; Sherman, D. H. Nat Prod Rep 2004, 21, 105-121. (32) Simmons, T. L.; Andrianasolo, E.; McPhail, K.; Flatt, P.; Gerwick, W. H. Mo!. Cancer Ther. 2005, 4, 333-342. (33) Pettit, G. R.; kamano, Y.; Herald, C. L.; Tuinman, A. A.; Boettner, F. E.; Kizu, H.; Schmidt, J. M.; Baczynkyji, L.; Tomer, K. B.; Bontems, R. J. J. Am. Chem. Soc. 1987, 109, 7581-7582. (34) Bal, R.; Pettit, G. R.; Hamel, E. Biochem. Pharmacol. 1990, 39, 19419. (35) Bai, R.; Friedman, S. J.; Pettit, G. R.; Hamel, E. Biochem. Pharmacol. 1992, 43, 2637-45. 32 (36) Bai, R.; Pettit, G. R.; Hamel, E. J. Biol. Chem. 1990, 265, 17141-9. (37) Hamel, E. Biopolymers 2002, 66, 142-160. (38) Vaishampayan, U.; Glode, M.; Du, W.; Kraft, A.; Hudes, G.; Wright, J.; Hussain, M. C/in. Canc. Res. 2000, 6, 4205-8. (39) Cruz-Monserrate, Z.; Mullaney, J. T.; Harran, P. G.; Pettit, G. R.; Mullaney, J. T.; Harran, P. G.; Pettit, G. R. Eur. J. Biochem. 2003, 270, 3822-3828. (40) Rinehart, K.; Gloer, J.; Cook, J.; Carter, J.; Mizsak, S.; Scahill, T. JAm Chem Soc 1987, 103, 1857-9. (41) Nuijen, B.; Bouma, M.; Manada, C. Anti-Cancer Drugs 2000, 11, 793811. (42) Wylie, B. L.; Ernst, N. B.; Grace, K. J. S.; Jacobs, R. S. Prog. Surg. 1997, 24, 146-152. (43) de Silva, E. D.; Scheuer, P. Tet. Left. 1980, 21, 1611-14. (44) Jacobs, R. S.; Culver, P.; Langdon, R.; O'Brien, T.; White, S. Tetrahedron 1985, 41, 981-4. (45) Deems, R. A.; Lombardo, D.; Morgan, 0. P.; Mihelich, E. D.; Dennis, E. A. Biochim. Biophys. Acta. 1987, 917, 258-68. (46) Look, S. A.; Fenical, W.; Matsumoto, G. K.; Clardy, J. J. Org. Chem. 1986, 51, 5140-5145. (47) Roussis, V.; Wu, Z.; Fenical, W.; Strobel, S. A.; Van Duyne, G. D.; Clardy, J. J. Org. Chem. 1990, 55, 4916-4922. (48) Kern, W. R.; Abbott, B. C.; Coates, R. M. Toxicon 1971, 9, 15-22. (49) de Fiebre, C. M.; Meyer, E. M.; Henry, J. C.; Muraskin, S. I.; Kern, W. R.; Papke, R. L. Mo!. Pharmacol. 1995, 47, 164-171. (50) Mahnir, V.; Lin, B.; Prokai-Tatrai, K.; Kern, W. R. Biopharm. Drug Disposit. 1998, 19, 147-151. (51) Wright, A. 0.; Konig, G. M. J. Nat. Prod. 1996, 59, 71 0-716. (52) Ang, K. K. H.; Holmes, M. J.; Higa, 1.; Hamann, M. T.; Kara, U. A. K. Antimicrob. Agents Ch. 2000, 44, 1645-1649. 33 (53) El Sayed, K. A.; Kelly, M.; Kara, U. A. K.; Ang, K. K. H.; Katsuyama, I.; Dunbar, D. C.; Khan, A. A.; Hamann, M. T. J. Am. Chem. Soc. 2001, 123, 1804-1808. (54) Luibrand, R. T.; Erdman, T. R.; Vollmer, J. J.; Scheuer, P. J.; Finer, J.; Clardy, J. Tetrahedron 1979, 35, 609-12. (55) De Cercq, E. Res. Rev. 2000, 20, 323-349. (56) Reddy, M. V.; Rao, M. R.; Rhodes, D.; Hansen, M. S.; Rubins, K.; Bushman, F. D.; Venkateswarlu, Y.; Faulkner, D. J. J. Med. Chem. 1999, 42, 1901-1 907. (57) Whitton, B. A.; Potts, M. In The ecology of cyanobacteria. Their diversity in time and space., B.A. Whitton and M.Potts, eds. (Dordrecht: Kluwer Academic Publisher), 2000, 1-11. (58) Harrigan, G. G.; Luesch, H.; Yoshida, W. Y.; Moore, R. E.; Nagle, D. G.; Paul, V. J.; Mooberry, S. L.; Corbett, T. H.; Valeriote, F. A. J. Prod. 1998, 61, (59) 1075-1077. Mooberry, S. L.; Leal, R. M.; Tinley, 1. L.; Luesch, H.; Moore, R. E.; Corbett, T. H. Inter. J. Cancer (60) 2003, 104, 512-521. Gerwick, W. H.; Proteau, P. J.; Nagle, D. G.; Hamel, E.; Blokhin, A. V.; Slate, D. L. J. Org. Chem. (61) Nat. 1994, 59, 1243-1245. Blokhin, A. V.; Yoo, H. D.; Geralds, R. S.; Nagle, D. G.; Gerwick, W. H.; Hamel, E. Mol Pharmacol 1995, 48, 523-31. (62) Luduena, R. F.; Prasad, V.; Roach, M. C.; Banerjee, M.; Yoo, H. D.; Gerwick, W. H. Drug Deliv. Res. (63) 40, 223-229. Hamel, E.; Blokhin, A. V.; Nagle, D. G.; Yoo, H. 0.; Gerwick, W. H. Drug Deliv. Res. (64) 1997, 1995, 34, 110-120. Verdier-Pinard, P.; Lai, J. Y.; Yoo, H. D.; Yu, J.; Marquez, B.; Nagle, D. G.; Nambu, M.; White, J. D.; Falck, J. R.; Gerwick, W. H.; Day, B. W.; Hamel, E. Mo!. Pharmacol. (65) 1998, 53, 62-76. Wipf, P.; Reeves, J. T.; Day, B. W. Curr. Pharm. Des. 1437. 2004, 10, 1417- 34 (66) Marquez, B. L.; Watts, K. S.; Yokochi, A.; Roberts, M. A.; Verdier- Pinard, P.; Jimenez, J. I.; Hamel, E.; Scheuer, P. J.; Gerwick, W. H. J Nat Prod. 2002, 65, 866-71. (67) Zabriskie, 1. M.; Kiocke, J. A.; Ireland, C. M.; Marcus, A. H.; Molinski, T. F.; Faulkner, 0. J.; Xu, C. F.; Clardy, J. J. Am. Chem. Soc. 1986, 108, 3123-4. (68) Crews, P.; Manes, L. V.; Boehier, M. Tetrahedron Lett. 1986, 27, 2797-2800. (69) Luesch, H.; Yoshida, W. Y.; Moore, R. E.; Paul, V. J.; Mooberry, S. L. J. Nat. Prod. 2000, 63, 611-5. (70) Sone, H.; Kondo, T.; Kiryu, M.; Ishiwata, H.; Ojika, M.; Yamada, K. J. Org. Chem. 1995, 60, 4774-81. (71) Milligan, K. E.; Marquez, B. L.; Williamson, R. T.; Gerwick, W. H. J. Nat. Prod. 2000, 63, 1440-3. (72) Orjala, J.; Nagle, 0. G.; Hsu, V.; Gerwick, W. H. J. Am. Chem. Soc. 1995, 117, 8281-2. (73) Baden, D. G.; Bikhazi, G.; Decker, S. J.; Foldes, F. F.; Leung, I. Toxicon 1984, 22, 75-84. (74) Berman, F. W.; Gerwick, W. H.; Murray, T. F. Toxicon 1999, 37, 16458. (75) Li, W. I.; Berman, F. W.; Okino, T.; Yokokawa, F.; Shioiri, T.; Gerwick, W. H.; Murray, T. F. Proc. Nat!. Acad. Sd. USA 2001, 98, 7599-7604. (76) Yokokawa, F.; Shioiri, T. J. Org. Chem. 1998, 63, 8638-8639. (77) White, J. D.; Hanselmann, R.; Wardrop, D. J. J. Am. Chem. Soc. 1999, 121, 1106-1107. (78) Yokokawa, F.; Fujiwara, H.; Shioiri, T. Tetrahedron 2000, 56, 17591775. (79) Li, W. I.; Marquez, B. L.; Okino, T.; Yokokawa, F.; Shioiri, T.; Gerwick, W. H.; Murray, T. F. J. Nat. Prod. 2004, 67, 559-68. (80) Wu, M.; Okino, T.; Nogle, L. M.; Marquez, B. L.; Williamson, R. T.; Sitachitta, N.; Berman, F. W.; Murray, T. F.; McGough, K.; Jocobs, R.; 35 Colsen, K.; Asano, T.; Yokokawa, F.; Shioiri, T.; Gerwick, W. H. J. Am. Chem. Soc. 2000, 122, 12041-1 2042. (81) Edwards, D. J.; Marquez, B. L.; Nogle, L. M.; McPhail, K.; Goeger, D. E.; Roberts, M. A.; Gerwick, W. H. Chem. Biol. 2004, 11, 817-33. (82) Cardellina, J. H., 2nd; Marner, F. J.; Moore, R. E. Science 1979, 204, 193-5. (83) Fujiki, H.; Mori, M.; Nakayasu, M.; Terada, M.; Sugimura, T.; Moore, R. E. Proc. Nat!. Acad. Sd. USA 1981, 78, 3872-6. (84) Aimi, N.; Odaka, H.; Sakai, S.; Fujiki, H.; Suganuma, M.; Moore, R. E.; Patterson, C. M. J. Nat. Prod. 1990, 53, 1593-6. (85) Basu, A.; Kozikowski, A. P.; Lazo, J. S. Biochemistry 1992, 31, 382430. (86) Kozikowski, A. P.; Shum, P. W.; Basu, A.; Lazo, J. S. J. Med. Chem. 1991, 34, 2420-30. (87) Edwards, D. J.; Gerwick, W. H. J. Am. Chem. Soc. 2004, 126, 1143211433. (88) Luesch, H.; Yoshida, W. Y.; Moore, R. E.; Paul, V. J.; Corbett, T. H. J. Am. Chem. Soc. 2001, 123, 5418-23. (89) Chen, J.; Forsyth, C. J. Proc. Nat!. Acad. Sc!. USA 2004, 101, 1206712072. (90) Orjala, J.; Gerwick, W. H. J. Nat. Prod. 1996, 59, 427-30. (91) Sitachitta, N.; Rossi, J.; Roberts, M.; Gerwick, W. H.; Fletcher, M. D.; Willis, C. L. J. Am. Chem. Soc. 1998, 120, 71 31-7132. (92) Chang, Z.; Flatt, P.; Gerwick, W. H.; Nguyen, V. A.; Willis, C. L.; Sherman, 0. H. Gene 2002, 296, 235-47. (93) Osinga, R.; Armstrong, E.; Burgess, J. G.; Hoffmann, F.; Reitner, J.; Schumann-Kindel, G. Hydrobio!ogia 2001, 461, 55-62. (94) Carter, D. C.; Moore, R. E.; Mynderse, J. S.; Niemczura, W. P.; Todd, J. S. J. Org. Chem. 1984, 49, 236-241. (95) Williams, 0. E.; Burgoyne, 0. L.; Rettig, S. J.; Andersen, R. J.; FathiAfshar, Z. R.; Allen, 1. M. J. Nat. Prod. 1993, 56, 545-551. 36 (96) In, Y.; Doi, M.; Inoue, M.; Ishida, T.; Hamada, V.; Shioiri, T. Acta Ctystallogr. C. 1994, 50, 432-434. (97) McDonald, L. A.; Ireland, C. M. J. Nat. Prod. 1992, 55, 376-379. (98) Degnan, B. M.; Hawkins, C. J.; Lavin, M. F.; McCaffrey, E. J.; Parry, D. L.; van den Brenk, A. L.; Watters, D. J. J. Med. Chem. 1989, 32, 13491354. (99) Fu, X.; Do, T.; Schmitz, F. J.; Andrusevich, V.; Engel, M. H. J. Nat. Prod. 1998, 61, 1547-1 551. (100) Salomon, C. E.; Faulkner, 0. J. J. Nat. Prod. 2002, 65, 689-692. (101) Schmidt, E. W.; Sudek, S.; Haygood, M. G. J. Nat. Prod. 2004, 67, 1341-1345. (102) Thacker, R. W.; Starnes, S. Marine Biology 2003, 142, 643-648. (103) Trimurtulu, G.; Ohtani, I.; Patterson, G. M.; Moore, R. E.; Corbett, T. H.; Valeriote, F. A.; Demchik, L. J. Am. Chem. Soc. 1994, 116, 47294737. (104) Smith, C. D.; Zhang, X.; Mooberry, S. L.; Patterson, G. M.; Moore, R. E. Cancer Res. 1994, 54, 3779-3784. (105) Kobayashi, M.; Kitagawa, K. J. Nat. Toxins 1999, 8, 249-258. (106) Dumdei, E. J.; Simpson, J. S.; Garson, M. J.; Bryriel, K. A.; Kennard, C. H. L. Aus. J. Chem. 1997, 50, 139-144. (107) Harrigan, G. G.; Goetz, G. H.; Luesch, H.; Yang, S.; Likos, J. J. Nat. Prod. 2001, 64, 1133-1138. (108) MacMillan, J. B.; Trousdale, E. K.; Molinski, T. F. Org. Lett. 2000, 2, 2721-2723. (109) Jiménez, J. I.; Scheuer, P. J. J. Nat. Prod. 2001, 64, 200-203. (110) Burja, A. M.; Hill, R. 1. Hydrobiologica 2001, 461,41-47. (111) Hentschel, U.; Hopke, J.; Horn, M.; Friedrich, A. B.; Wagner, M.; Hacker, J.; Moore, B. S. App!. Enviro. Micro. 2002, 68, 4431-4440. (112) Vacelet, J.; Donadey, C. J. Exp. Mar. Ecol. 1977, 30, 301-314. (113) Barrows, L. R.; Radisky, D. C.; Copp, B. R.; Swaffar, D. S.; Kramer, R. A.; Warters, R. L.; Ireland, C. M. Anti-Cancer Drug Design 1993, 8, 333-347. 37 (114) Ishibashi, M.; Iwasaki, T.; lmai, S.; Sakamoto, S.; Yamaguchi, K.; Ito, A.; Iwasaki, T.; lmai, S.; Sakamoto, S.; Yamaguchi, K.; Ito, A. J. Nat. Prod. 2001, 64, 108-110. (115) Hill, R. T.; Hamann, M. T.; Peraud, 0.; Kasanah, N. Ref Type: Patent 2003, 4115-1 80 PCT. (116) Clark, W. D.; Crews, P. Tetrahedron Left. 1995, 36, 1185-8. CHAPTER TWO SEMIPLENAMIDES A - G, FATTY ACID AMIDES FROM A PAPUA NEW GUINEA COLLECTION OF THE MARINE CYANOBACTERIUM LYNGBYA SEMIPLENA ABSTRACT Semiplenamides A (1) to G (7), a series of new anandamide-like fatty acid amides, were isolated from a 1999 Papua New Guinea collection of the marine cyanobacterium Lyngbya semiplena. The planar structures of these lipids were determined using standard ID and 2D NMR methods. The relative stereochemistry of the aliphatic portion of the new metabolites was deduced from ID NOE data and 1Hdecoupling experiments, while the absolute stereochemistry of the amino alcohol moieties was assigned by chemical derivatization and chiral GC-MS methods. All of these new metabolites displayed toxicity in the brine shrimp model system, three showed modest potency in displacing radiolabeled anandamide from the cannabinoid receptor (CBI), and one was a modest inhibitor of the anandamide membrane transporter (AMT). INTRODUCTION Marine cyanobacteria have emerged over the past few years as one of the richest groups of marine organisms for their bioactive and structurally complex natural products. Particularly prevalent among these is a rich elaboration upon a cyclic peptide template using a diversity of standard as well as modified amino acids.1 However, a growing trend from our investigations of these life forms is the production of fatty acid amides that combine unusual fatty acids with a variety of "biogenic amines". Examples include the hermitamides A (8) and B (9)2 and grenadamide 39 (1O), two types of toxic malyngamide-type natural products from the marine cyanobacterium Lyngbya majuscula. In this regard, they structurally resemble anandamide (11) and other endocannabinoids (12, 13) of importance to mammalian physiology,3 a finding that has stimulated our investigation of their pharmacological properties in several relevant bioassays. OCH3 0 H Hermitamide A (8) Hermitamide B (9) Grenadamide (10) Anandamide (11) OH N H LN0H Palmitamide (12) NH2 Oleamide (13) 40 RESULTS AND DISCUSSION Collections of a shallow water (1-3 m) strain of L. semiplena were made in Wewak Bay, Papua New Guinea. Preliminary bioassay of the crude organic extract showed good activity in the brine shrimp toxicity model at 10 ppm.4 Guided by this assay, the natural products semiplenamide A (1) to G (7) were isolated and purified by sequential vacuum liquid chromatography (VLC) and HPLC in 0.1-1.5 mg/g yield. )OR semiplenamide A,B I R=H 2 R=Ac J.OH 1' 16 2' semiplenamide o 20 4 semiplenamide o 18 semiplenamide 5 E OR 18 semiplenamide F,G 19 6 R=H 7 R=Ac Semiplenamide A (1) showed an [M+H] peak at m/z 366.3372 for a molecular formula of C23HNO2 by HR FABMS (3 degrees of unsaturation). The structure of I 41 was established mainly from analysis of ID (1H NMR, 13C NMR, and NOE) and 2D NMR (COSY, HSQC, and HMBC) spectra (Figure II. 1-4). The 1H NMR spectrum of I was indicative of a fatty acid amide-type metabolite, with the presence of proton signals for a long aliphatic chain in the 6 1.25-1.35 envelope (1 8H), and a poorly defined terminal CH3 triplet (6 0.88), as well as a broad amide singlet (6 6.25) (Tablell. 1). The three degrees of unsaturation were accounted for by an amide carbonyl carbon resonance (6 171.5; IR absorption at 1659 cm1) and two olefinic bonds (6 130.7, 137.4, 129.2, 132.3). In the 1H NMR spectrum of I, a deshielded methyl singlet at 6 1.89 (C-21) was assigned to a vinyl methyl moiety because it showed two and three bond HMBC correlations to two olefinic carbons (6 130.7, C-2 and 137.4, C-3) as well as to the amide carbonyl carbon (6 171.5, C-I). An ethanolamine moiety could be deduced from two mutually coupled (COSY) midfield proton signals at 6 3.54 (dt, 6.0, 4.0 Hz) and 6 3.82 (t, 5.0 Hz), each of which integrated to 2 protons, and which were correlated by HSQC with two midfield carbon resonances at 6 43.6 and 63.8, respectively. The former signal was also coupled to a broadened amide NH proton (6 6.25). The relative position of the two olefinic bonds was determined by HMBC (Table I. 1) which showed 2- and 3- bond coupling from two mutually coupled methylene resonances (6 2.23, H2-4; 2.14, H2-5) to all four olefinic carbons. Additionally, the olefinic proton resonance at 6 6.45 (H-3) showed a three bond correlation to the amide carbonyl carbon resonance (6 171.5, C-i), thus positioning this partial structure relative to the amide motif. The linking of the ethanolamine moiety to this amide was shown by an HMBC connectivity between the midfield methylene protons (6 3.54, H2-i') and the C-i carbonyl. Combining these partial structures accounted for all atoms, and thus completed the planar structure of semiplenamide A (I). The geometry of the C-6, 7 disubstituted olefin was determined by a 1H NMR decoupling experiment (Figure Il. 5), in which the signals for the associated protons were simplified by decoupling H2-5 and H2-8 (6 2.14, 1.98). In this latter spectrum it was possible to measure JH-61H.7as 15.0 Hz, thus assigning the C-6 olefin as trans. The geometry of the C-2, 3 olefin in I was assigned using I D NOE experiments. Reciprocal NOE enhancements were seen from the vinyl methyl protons (6 1.88, H3- 42 21) to the methylene protons (6 2.23, H2-4) as well as to the amide NH proton, thus defining the geometry of the C-2 olefin as E (Figure II. 6). The 1H NMR data for semiplenamide B (2) were similar to those obtained for 1, except for an additional singlet methyl resonance at 6 2.08. HR FABMS data for 2 showed an [M + H] ion at m/z 408.3473 consistent with a molecular formula of C25H46NO3, indicating a structure with four degrees of unsaturation. In concurrence with the 1H NMR data (Table II. 1), the additional degree of unsaturation was attributed to a terminal acetate motif linked to the ethanolamine; this was confirmed by 1H, 13C and 2D NMR data (Table II. 1). 7.5 170 7.0 160 Figure II. 1. 150 6.5 140 6.0 130 ............ 5.5 120 5.0 110 4.5 4.0 100 90 3.5 3.0 80 70 2.5 60 2.0 50 1H and 13C NMR spectra of semiplenamide A (1). 1.5 40 30 I 0.5 ppm 1.0 20 ppm 43 ppm 8.0- 8.0 7.5 7.0 6.5 6.0 5.5 5.0 4.5 4.0 3.5 3.0 2.5 Figure II. 2. COSY spectrum of semiplenamide A (1). 2.0 1.5 1.0 0.5 ppm 44 ppm 0 10 20 30 "4 40 50 60 4 70 80 90 100 110 120 130 140 ........................................... 9 8 7 6 5 4 3 Figure II. 3. HSQC spectrum of semiplenamide A (1). 2 1 0 ppm 45 ppm 0 20 40 60 0 80 100 120 01 140 160 I 180 200 220 8.0 7.5 7.0 6.5 6.0 5.5 5.0 4.5 4.0 3.5 3.0 2.5 Figure II. 4. HMBC spectrum of semiplenamide A (1). 2.0 1.5 1.0 0.5 ppm 46 I Iis.I J'"l - .JJI 5.5 I. I U4 ..:. ,.. - 5.4 Figure Il. 5. Determination of the geometry at C6-C7 in semiplenamide A (1) by a 1H NMR decoupling experiment. 2.23ppm H 1rH / 2Oppm OH 1,5 1.5 E.D 5.5 5. 4.5 4.0 3.5 3.0 2.0 1.5 1,0 0.5 p,:li Figure II. 6. Determination of the geometry at C2-C3 in semiplenamide A (1) by a 1D NOE NMR experimen 47 Table II. 1. 1H and 13C NMR Assignments for Semiplenamides A (1) and B (2). nr Semiplenamide A (1) HMBC 1 2 6.45 (t, 7.0) 4 2.23 (dm, 7.0) 5 2.14 (dm, 7.0) 6 5.42 (m) 7 5.46 (m) 8 1.98 (dm, 7.5) 9 1.33 (m) 10-17 1.27(m) 18 1.27 (m) 19 1.30 (m) 20 0.88 (t, 6.5) 21 1.89 (s) 3 1' 2' (dd, 3.82 (t, 5.0) 6.0, 171.5 130.7 137.4 C-21 29.0 C-3, C-5, C-6 32.3 129.2 C-8 132.3 C-5 33.4 30.6 30.3 32.8 23.8 14.9 12.5 HMBC 6.36 (t, 7.0) 2.23 (dm, 7.0) 2.14 (dm, 7.0) 5.42 (m) 5.46 (m) 1.98 (dm, 7.5) 1.33 (m) 1.27(m) 1.27 (m) 1.30 (m) C-18, 19 0.88 (t, 6.5) C-I, C-2, C-3 1.84 (s) (dd, 43.6 C-i, C-2' 63.8 OAc N-H Semiplenamide B (2) 6.25 (br s) 4.21 (t, 5 .0) 2.08 (s) 169.8 131.2 136.4 29.0 32.3 129.2 132.3 33.4 30.6 30.3 32.8 c-i C-3, c-5 C-3, C-4, C-7 C-8 C-6, C-8 C-6, C-7, c-9 C-8, C-b C-9 23.8 14.9 6.0, 13.1 C-I, C-2, C-3 39.6 C-2', OAc 63.8 21.30 171.5 c-i', c-i 6.03 (br s) HR FABMS of semiplenamide C (3) gave an [M + H] peak at 326.3053, yielding a molecular formula of C20H40NO2with two degrees of unsaturation. As for 1, the planar chemical structure of 3 was elucidated using an assemblage of 1 D and 2D NMR (COSY, HSQC, and HMBC) experiments. The 1H NMR spectrum of 3 had several features strikingly similar to that of I (Table II. 2), including a deshielded olefinic 1H signal ( 6.38), a broad amide NH resonance ( 5.81) and a deshielded methyl singlet ( 1.84), as well as proton signals for a long aliphatic chain in the S 1.25-1.35 envelope and a terminal methyl triplet (5 0.88). However, semiplenamide C (3) lacked resonances for the double bond found in I and 2, and was therefore a more saturated derivative. An additional difference in the 1H NMR spectra for I and 3 48 was the presence of a CH resonance at 6 4.13 (H-i '), and CH2 resonances at 6 3.72 and 3.58 (H2-2') in 3 instead of the two CH2 resonances at 6 3.54 (H2-1') and 3.82 (H2-2') in the spectrum of 1. This methine resonance (84.13 m) was coupled to two mutually coupled methylene resonances (6 3.72 dd, 3.58 dd), a broad amide NH resonance (6 5.81) and a methyl resonance (6 1.22 d) in the COSY spectrum of 3. Two- and three- bond HMBC correlations (Table II. 2) from H3-3' to C-i' and C-2' confirmed the connectivity deduced from COSY, and thus established the structure of an alaninol moiety in 3 versus the ethanolamine moiety in 1. An L configuration for this group was determined by chiral GC-MS comparison of the corresponding PFPAAc ester derivative with similarly prepared alaninol standards.5 HR FABMS data for semiplenamide D (4) showed an EM + H] ion at m/z 424.3802 consistent with a molecular formula of C26HNO3, indicating a structure with three degrees of unsaturation. The 1H and 13C NMR spectra for 4 displayed signals very similar to those of 3 (Table II. 3), except for an additional singlet methyl resonance (6H 2.08, oc 21.2). Detailed analysis of the 13C and 2D NMR data assigned this additional singlet methyl signal to a terminal acetate motif linked to the ethanolamine (as in 2), which also accounted for the additional unsaturatiori degree. Consistent with HRFABMS data, compounds 3 and 4 also differed in the lengths of their 'fatty acid' chains (16 and 20 carbons, respectively). While the NMR data for semiplenamide E (5, Table Il. 3) were essentially identical to those obtained for 4, integration of the aliphatic chain proton envelope was slightly less for 5 than for 4 (24H vs 28H), suggesting that 5 had a shorter aliphatic chain. HR FABMS data for 5 showed an EM + H] ion at m/z 396.3584 consistent with a molecular formula of C24H46NO3 and an 18-carbon chain, in contrast to the 20-carbon chain of 4. HR FABMS of semiplenamide F (6) gave an EM + H] peak at 370.3803, yielding a molecular formula of C22HNO3 with two degrees of unsaturation. In the 1H NMR spectrum of 6 (Table II. 4), chemical shifts of the alaninol residue protons were apparent as well as a broad amide NH resonance (6 6.49). Notably, the deshielded olefinic proton resonance and vinyl methyl singlet in the spectra of 1-5 were replaced by an upfield methine triplet (6 2.89, J = 6.2 Hz) and methyl singlet (6 1.55). The appearance of two midfield signals (6 60.3, 64.4) in the 13C NMR 49 spectrum, taken together with the HMBC data for 6 (Table II. 4), led to the assignment of an epoxide ring at C-2 ( 60.3) and C-3 (ö 64.4). Table II. 2. 1H and 13C NMR Assignments for Semiplenamide C (3). Semiplenamide C (3) HMBC number 1 170.9 2 131.1 3 4 5 6-13 6.38 (t, 7.2) 2.13 (dm, 7.2) 1.41 (m) 1.25(m) 14 1.25(m) 15 1.31 (m) 16 17 0.88 (t, 7.0) 1.84 (s) 1' 4.13(m) 2a' 2b' 3' N-H 137.7 C-i, C-i7, C-4 29.1 C-3, C-5, C-2 29.4 C-4 29.7 32.0 22.8 14.8 12.8 48.7 3.58 (dd, 5.5, 3.5) 68.3 3.72 (dd, 5.5, 6.0) 68.3 1.22 (d, 7.0) 17.3 5.81 (brs) C-i4, C-15 C-I, C-2, C-3 C-i', C-3' C-i', C-3' C-i',C-2' 50 Table II. 3. 1H and 13C NMR Assignments for Semiplenamides D (4) and E (5). Semiplenamide 0 (4) nr HMBC 2 4 5 6-15 HMBC 169.3 1 3 Semiplenamide E (5) 169.3 130.9 130.9 6.33 (t, 7.0) 2.13 (dm, 7.0) 1.41 (m) 1.25 (m) 30.1 6.33 (t, 7.0) 2.11 (dm, 7.0) 1.41 (dm, 7.0) 1.25 (m) 137.1 28.8 29.2 137.1 C-I, C-2, c-ic 28.8 29.2 C-3, C-5 C-4 30.1 16 1.25(m) 32.3 1.25(m) 32.3 17 1.29 (m) 23.1 23.1 18 23.1 14.5 C-17, C-16 19 1.29 (m) 1.29 (m) 1.29 (m) 0.88 (t, 6.0) 23.1 1.82 (s) 13.0 C-i, C-3 20 0.88 (t, 6.0) 14.5 21 1.82(s) 4.33 (m) 13.0 C-i,C-2,C-3 4.33 (m) 4.20 (dd, 5.8) 4.05 (dd, 4.0) 1.20 (d, 7.0) 2.08 (s) 45.1 C-2', C-3' 1' (dd, 2a' 4.05 (dd, 4.0) 3' 1.20 (d, 7.0) QAc 2.08(s) 2b' C-19, C-18 45.1 67.5 67.5 17.8 C-i', C-2' 21.2 171.8 N-H 5.82 (br s) 67.5 C-i', C-3' 67.5 C'-1, C-3' 17.8 C-i', C-2' 21.2 171.8 5.82 (br s) The 1H NMR data for semiplenamide G (7) were quite similar to those obtained for 6 (Table II. 4), with the exception again of an additional singlet methyl resonance at 2.08. HR FABMS data for 7 showed an [M + HJ ion at m/z4i2.3427 consistent with a molecular formula of C24H46N04, indicating a structure with three degrees of unsaturation. From examination of the HMBC data as well as the 13C NMR spectrum (Table 4), the additional degree of unsaturation in 7 was attributed to a terminal acetate group linked to the alaninol motif. While in CDCI3 the H3-19 and H2-4 signals were overlapped, in MeOH-d4 they were resolved and a I D NOE irradiation of H3-i 9 produced a significant enhancement in the H2-4 resonance (ö 1.58). Therefore, a 51 Table II. 4. 1H and 13C NMR Assignments for Semiplenamides F (6) and G (7). Semiplenamide F (6) HMBC 6c number Semiplenamide G (7) HMBC 6c 2.89 (t, 6.2) 1.58 (m) 1.26 (m) 1.27 (m) 173.2 60.3 64.4 C-4 28.4 C-3 32.0 30.2 2.84 (t, 6.2) 1.58 (m) 1.26 (m) 1.27 (m) 60.4 64.4 C-i, C-2, C-1 28.4 C-3, C-5 32.0 C-4 30.2 16 1.30(m) 23.1 1.30(m) 23.1 17 1.41 (m) 0.88 (t, 6.0) 1.55 (s) 4.01 (m) 26.6 1.41 (m) 14.5 C-17 0.88 (t, 6.0) 13.4 C-I, C-2, C-3 1.51 (s) 48.1 C-I, C-2', C-34.20 (m) 1 2 3 4 5 6-15 18 19 1' 2a' 2b' 3' (dd, 6.0, (dd, 4.0, 1.16(d,7.2) 67.5 C-i', C-3' 67.5 C-i', C-3' 17.8 C-i',C-2' OAc N-H 6.49 (br s) 173.2 (&1, (dd, 26.6 14.5 C-17, C-16 13.4 C-i, C-3 45.1 C-2', C-3' 2.0, 2.0, 67.5 C-I', C-3' 67.5 C'-i, C-3' 1.13 (d, 7.2) 17.8 2.08(s) 21.2 171.8 C-1',C-2' 6.40 (br s) relative stereochemistry of 2S*, 3R* was assigned to the epoxide in 7. Because the optical rotation values for semiplenamides F and G (6, 7) were very similar (-5.0° and -3.0°, respectively), we conclude that they likely possess the same relative and absolute stereochemistry. However, while it is likely that the alaninol moiety of 6 and 7 are of the same L-stereochemistry as in semiplenamides C-E (3-5), the small amount isolated of these compounds precluded this assignment. The semiplenamides were evaluated for their biological activity in several systems. In the brine shrimp (Artemia sauna) toxicity assay,4 semiplenamides A (1) toG (7) showed LDvalues of 1.4, 2.5, 1.5, 18, 19, 1.4, and 2.4 pM respectively. Semiplenamides E (5) and G (7) are structurally very close except for the replacement of the olefin moiety in 5 by an epoxide ring in 7; however, the LD value 52 of 5 is eight fold higher than that of 7. Apparently, the epoxide ring produces an enhanced toxicity in this series of metabolites. Due to the structural resemblance of the novel ethanolamide derivatives with anandamide, (N-arachidonoyl-ethanolamine), an endogenous agonist of cannabinoid receptors compounds,6 we tested them on the best characterized proteins of the endocannabinoid system: 1) the "central" cannabinoid CB1 receptors; 2) the anandamide membrane transporter (AMT), which is responsible for anandamide cellular uptake; and 3) the fatty acid amide hydrolase (FAAH), which catalyses anandamide hydrolysis! When the semiplenamides were tested for their capability to displace the high affinity CB1 ligand [3H]SR141716A from cannabinoid CB1 receptors in rat brain membranes, three semiplenamides (A (1), B (2) and G (7)) exhibited some affinity. The K values were 19.5±7.8, 18.7±4.6, and 17.9±5.2 for compounds A, B and G, respectively (means±SD, n=3). Under the same conditions, anandamide exhibited a K =0.4 pM, and therefore, these three Lyngbya metabolties should be considered as weak CB1 agonists. This was not surprising because substrate recognition requires the presence of at least one cis double bond situated at the middle of the fatty acid carbon chain, indicating a preference for ligands whose hydrophobic tail can adopt a bent U-shaped conformation.8 Moreover, fatty acid ethanolamides need to contain at least three homoconjugated double bonds in order to interact in an optimal way with GB1 receptors.6 However, it was interesting to note that: 1) semiplenamide B (2) was as active as its non-acetylated analogue, semiplenamide A (1), in agreement with the previous finding that the 2'-hydroxy-group in acylethanolamides is not necessary for the interaction with CB1 receptors;6 2) semiplenamide G (7) was more potent than its non-epoxide analogue, compound E (5); this is the first time that the affinity for GB1 receptors of a 2,3-epoxide-acyl-ethanolamide derivative was investigated. Next, we tested the effect of the novel compounds on [14C]anandamide hydrolysis, by using membranes from mouse NI8TG2 neuroblastoma cells, which express high levels of FAAH (Km for anandamidel5 j.tM).9 No appreciable inhibitory effect was found for any of the tested compounds. This was not surprising since previous experiments had shown that both 2-methyl and 1'-methyl groups on 53 anandamide confer a certain refractoriness to become a substrate for or to competitively inhibit FAAH.6 Finally, we tested the effect of the semiplenamides on the uptake of [14C]anandamide by intact RBL-2H3 cells, a cell type in which an anandamide membrane transporter (AMT) has been partially characterized ranges from 9.3 to 33 pM).7'1° (Km for anandamide Only semiplenamide A inhibited the AMT (lC=18.1±3.2 tM). Although more potent AMT inhibitors have been described in the literature (with lC values ranging between 0.8 and 10 pM), compound A (1) is the first of such class of inhibitors to contain at the same time 6(E) and 2-methyl-2(Z)double bond. As these two features should confer increased metabolic stability, the of semiplenamide A widens finding the possible chemical features that acylethanolamide derivatives might possess in o rder to inhibit anandamide cellular uptake. In conclusion, these data support the importance of natural products as a possible reservoir for new molecules capable of interacting with proteins of the endocannabinoid system. EXPERIMENTAL General Experimental Procedures. Optical rotations were measured on a Perkin-Elmer 141 polarimeter. IR and UV spectra were recorded on Nicolet 510 and Beckman DU64OB spectrophotometers, respectively. NMR spectra were recorded on Bruker Avance DPX 400 MHz and Bruker Avance 300 MHz spectrometers with the solvent CDCI3 used as an internal standard (oH at 7.26, Oc at 77.2). Mass spectra were recorded on a Kratos MS5OTC mass spectrometer. Chiral GCMS analysis was accomplished on a Hewlett-Packard gas chromatograph 5890 Series II with a Hewlett-Packard 5971 mass selective detector using an Alltech capillary column (CHIRASIL-VAL phase 25 m x 0.25 mm). HPLC isolations were performed using a Waters 515 HPLC pump and Waters 996 photodiode array detector. Collection. The marine cyanobacterium Lyngbya semiplena (voucher specimen available from WHG as collection number PNGE12-7Dec99-3) was collected from shallow waters (1-3 m) in Wewak Bay, Papua New Guinea, on December 7, 1999. The material was stored in 2-propanol at -20°C until extraction. 54 Extraction and Isolation. Approximately 138 g (dry wt) of the alga were extracted repeatedly with CH2Cl2/MeOH (2:1) to produce 3.05 g of crude organic extract. The extract (3.0 g) was fractionated by silica gel vacuum liquid chromatography using a stepwise gradient solvent system of increasing polarity starting from 10% EtOAc in hexanes to 100% MeOH. Fractions eluting with 50%, 65%, 75%, and 100% EtoAc in hexanes were found to be the most active at 10 ppm in the brine shrimp toxicity assay. These fractions were further chromatographed on Mega Bond RP18 solid-phase extraction (SPE) cartridges using a stepwise gradient solvent system of decreasing polarity starting from 80% MeOH in H20 to 100% MeOH. The most active fractions after SPE (85% toxicity at 1 ppm to brine shrimp) were then purified by HPLC [Phenomenex Sphereclone 5 j ODS (250 x 10.00 mm), 9:1 MeOH/H20, detection at 211 nm] giving compounds 1 (1.3 mg), 2 (0.5 mg), 3 (4.5 mg), 4 (2.5 mg), 5 (0.3 mg), 6 (0.3 mg), and 7 (2.0 mg). Semiplenamide A (1): white amorphous solid; UV (MeOH) Amax 206 nm (c 5100); IR (neat) 3330, 2918, 2850, 1659, 1616, 965 cm1; 1H and 13C NMR data, see Table 1; HR FABMS m/z [M + H] 366.3372 (calcd for C23HNO2, 366.3375). Semiplenamide B (2): colorless oil; UV (MeOH) Amax 203 nm (c 5900); IR (neat) 3389, 2920, 2851, 1741, 1660, 966 cm1; 1H and 13C NMR data, see Table 1; HR FABMS m/z [M + H] 408.3473 (calcd for C25H46NO3, 408.3469). Semiplenamide C (3): white amorphous solid; [aJ26D -5.0° (c 0.3, CHCI3); UV (MeOH) Amax 213 nm (c 6500); IR (neat) 3282, 2915, 2847, 1658, 1622, 966 cm1; 1H and 13C NMR data, see Table 2; HR FABMS m/z [M + H] 326.3053 (calcd for C20HNO2, 326.3047). Semiplenamide D (4): white amorphous solid; [a]26D -10.6° (c 0.15, CHCI3); UV (MeOH) Amax 207 nm (s 5200); IR (neat) 3279, 2916, 2849, 1721, 1622, 955 cm1; 1H and 13C NMR data, see Table 3; HR FABMS m/z EM + H] 424.3802 (calcd for C26HNO3, 424.3814). Semiplenamide E (5): white amorphous solid; [a126D -7.1 ° (c 0.28, CHCI3); UV (MeOH) A 207 nm (c 4300); IR (neat) 3280, 2916, 2849, 1719, 1623, 966 cm1; 1H and 13C NMR data, see Table 3; HR FABMS m/z [M C24H46NO3, 396.3578). HJ 396.3584 (calcd for 55 Semiplenamide F (6): white amorphous solid; EU1D -5.0° (c 0.3, CHCI3); UV (MeOH) A, 205 nm (c 5500); IR (neat) 3295, 2916, 2849, 1646, 1535, 1080 cm1; 1H and 13C NMR data, see Table 4; HR FABMS m/z [M + H1 370.3803 (calcd C22HNO3,370.3808). Semiplenamide G (7): white amorphous solid; (MeOH) Amax [a]26D 3.00 (c 0.6, CHCI3); UV 207 nm (c 6100); IR (neat) 3283, 2917, 2849, 1723, 1650, 1539, 1273 cm1; 1H and 13C NMR data, see Table 4; HR FABMS m/z [M + H] 412.3427 (calcd for C24H46N04, 412.3428). Chiral GC-MS Analyses of the Alaninol Moiety in 3. Approximately 0.2 mg of compound 3 was hydrolyzed with 6 N HCI at 110°C (20 h). The hydrolysate was evaporated to dryness and resuspended in HCI (0.2N, 100 pL) and heated (110 °C, 5 mm). The residue was reduced to dryness using a stream of N2, and the reaction mixture was esterified with acetyl chloride (10 pL) at 100 °C (lh) and reduced to dryness. To the ice cooled esterified mixture, pyridine (0.5 pL), CH2Cl2 (10 pL), and pentafluoropropionic anhydride (1.5 pL) were added successively. Finally, the mixture was heated in a sealed vial (2 h, 100 °C), evaporated to dryness and resuspended in hexane. Racemic mixtures as well as optically pure L- or 0- alaninol standards were derivatized in similar fashion. Capillary GC-MS analyses were conducted using a Chirasil-Val column (Alltech, 25 m x 0.25 mm) with the following conditions: initial oven temperature of 50 °C (4 mm), a 5 °C 1mm ramp from 50 °C to 150 °C, and concluding with a 20 °Clmin ramp from 150 °C to 180 °C. The fragment derivatized from compound 3 and the derivatized L-alaninol standard both eluted at 13.02 mm. The derivatized D-alaninol standard eluted at 13.58 mm. Brine Shrimp Toxicity Bioassay. Brine shrimp Artemia sauna toxicity was measured as previously described.4 After a 24 h hatching period, aliquots of a 10 mglmL stock solution of compounds A-G were added to test wells containing 5 mL of artificial seawater and brine shrimp to achieve a range of final concentrations from 0.1 to 100 ppm. After 24 h the live and dead shrimp were tallied. Anandamide cellular uptake assay. The effect of compounds on the uptake of [14CJAEA by intact rat basophilic leukemia (RBL-2H3) cells was studied by using 5.0 pM (20,000 cpm) of [14C]AEA as described [14C]AEA for 5 mm at previously.7 Cells were incubated with 37 °C, in the presence or absence of varying concentrations of the inhibitors. Residual [14C]AEA in the incubation media after extraction with CHCI3/CH3OH 2:1 (by vol.), determined by scintillation counting of the lyophilized organic phase, was used as a measure of the AEA that was taken up by cells. Previous studies7 had shown that, after a 5 mm incubation, the amount of [14C]AEA disappeared from the medium of RBL-2H3 cells is found mostly (>90%) as unmetabolized [14C]AEA in the cell extract. Non-specific binding of [14CJAEA to cells and plastic dishes was determined in the presence of 100 pM AEA and was never higher than 30%. Data are expressed as the concentration exerting 50% inhibition of AEA uptake (lC). CB, Receptor Binding Assay. Displacement assays for CB1 receptors were carried out by using [3H]SR141716A (0.4nM, 55Ci/mmol, Amersham) as the highaffinity ligand, and the filtration technique described previously,11 on membrane preparations (0.4 mg/tube) from frozen male CD rat brains (Charles River, Wilmington, MA, U.S.A.), and in the presence of 100 pM PMSF. Specific binding was calculated with 1 pM SR141716A and was 84.0%. Data are expressed as the K1, calculated using the Cheng-Prusoff equation from the concentration exerting 50% inhibition of [3H]SR141716A specific binding (lC). Fatty acid amide hydrolase assay. The effect of compounds on the enzymatic hydrolysis of AEA was studied as described previously,9 using membranes prepared from mouse neuroblastoma NI8TG2 cells, incubated with the test compounds and [14C]AEA (10 pM, 40,000 cpm) in 50 mM Tris-HCI, pH 9, for 30 mm at 37 °C. [14C]Ethanolamine produced from [14C]AEA hydrolysis was measured by scintillation counting of the aqueous phase after extraction of the incubation mixture with 2 volumes of CHCI3/CH3OH 2:1 (by vol.). Data are expressed as the concentration exerting 50% inhibition of [14CJAEA hydrolysis (lC). 57 REFERENCES (1) Gerwick, W. H.; Tan, L. T.; Sitachitta, N. In Alkaloids: Chemistry and Biology, Cordell, G. A., Ed.; Academic Press: New York, 2001; Vol. 57, pp 75-184. (2) Tan, L. T.; Okino, T.; Gerwick, W. H. J. Nat. Prod. 2000, 63, 952-955. (3) Sitachitta, N.; Gerwick, W. H. J. Nat. Prod. 1998, 61, 681-684. (4) Meyer, B. N.; Ferrigni, N. R.; Putnam, L. B.; Jacobsen, L. B.; Nichols, D. E.; McLaughlin, J. L. Planta Med. 1982, 45, 31-3. (5) Gerard, J. M.; Haden, P.; Kelly, M. T.; Andersen, R. J. J. Nat. Prod. 1999, 62, 80-85. (6) Melck, 0.; Bisogno, T.; De Petrocellis, L.; Chang, H.; Julius, 0.; Bifulco, M.; Di Marzo, V. B iochem. Biophys. Res. Commun. 1999, 262, 27584. (7) Di Marzo, V.; Bisogno, T.; De Petrocellis, L.; Melck, D.; Martin, BR. Curr. Med. Chem. 1999, 6, 721-744 (8) Sheskin, T.; Hanus, L.; Stager, J.; Vogel, Z.; Mechoulam, R. J. Med. Chem. 1997, 40, 659-667. (9) Maurelli, S.; Bisogno, 1.; De Petrocellis, L.; Di Marzo, V. FEBS Left. 1995, 377, 82-86. (10) Fowler, C.J.; Jacobsson, SO. Prostaglandins Leukot. Essent. Fatty Acids 2002, 66, 193-200. (11) Bisogno, T. ; Maurelli, S. ; Melck, D. ; De Petrocellis, L. ; Di Marzo, V. J. Biol. Chem. 1997, 272, 3315-23. CHAPTER THREE THE WEWAKPEPTINS, CYCLIC DEPSIPEPTIDES FROM A PAPUA NEW GUINEA COLLECTION OF THE MARINE CYANOBACTERIUM LYNGBYA SEMIPLENA ABSTRACT Four new depsipeptides have been isolated from the marine cyanobacterium Lyngbya semiplena collected from Papua New Guinea. The amino and hydroxy acid partial structures of wewakpeptin A-D (1-4) were elucidated through extensive spectroscopic techniques, including HR-FABMS, ID 1H and 13C NMR, as well as 2D COSY, HSQC, HSQC-TOCSY, and HMBC spectra. The sequence of the residues was determined through a combination of multifaceted approaches including ESI- MS/MS, HMBC, ROESY, and a modified I D HMBC experiment. The absolute stereochemistry of each residue was determined by chiral HPLC and chiral GC-MS methods. The wewakpeptins represent an unusual arrangement of amino- and hydroxy-acid subunits relative to known cyanobacterial peptides, and possess a bisester, a 2,2-dimethyl-3-hydroxy-7-octynoic acid (Dhoya) or 2,2-dimethyl-3hydroxyoctanoic acid (Dhoaa) residue, and a diprolyl group reminiscent of dolastatin 15. Wewakpeptin A and B were the most cytotoxic among these four depsipeptides with an LC50 of approximately 0.4 pM to both the NCI-H460 human lung tumor and the neuro-2a mouse neuroblastoma cell lines. 59 INTRODUCTION Cyanobacteria are an ancient and diverse group of microorganisms which occupy a broad range of habitats from marine to terrestrial. Marine representatives, especially those belonging to the genus Lyngbya, are a prolific source of secondary metabolites with pharmaceutical potential. A particularly prevalent structural theme among these is a rich elaboration of cyclic peptides using a diversity of standard as well as modified amino acids.1 An additional and emerging theme is the production of acyl amides, as revealed by our recent isolation of a series of new anandamide-like fatty acid amides, semiplenamides A to G, from the marine cyanobacterium Lyngbya semiplena Gomont, collected in Papua New Guinea.2 These latter metabolites displayed modest potency in displacing radiolabeled anandamide from the cannabinoid receptor (CB1). In the course of this latter effort, we found the most polar fraction from this organic extract contained a series of four new cyclic depsipeptides, wewakpeptin A-D (1-4). Herein, we report the isolation and structure elucidation of this new series of L. semiplena peptides, as well as their cytotoxicity to tumor cells. Wewakpeptin A and B were the most potent among these four depsipeptides, with an LC of approximately 0.4 pM to both the NCI-H460 human lung tumor and the neuro-2a mouse neuroblastoma cell lines. 30 2 4 19 Ii wewakpeptin A (1) R= wewakpeptin C (3) R wewakpeptin B (2) R= wewakpeptin 0 (4) R= RESULTS AND DISCUSSION Collections of a shallow water (1-3 m) strain of L. semiplena were made in Wewak Bay, Papua New Guinea. The algae were extracted with CH2Cl2/MeOH (2:1) and fractioned by silica gel vacuum liquid chromatography. Preliminary bioassay of the MeOH-eluted fraction showed toxicity in the brine shrimp model (LD - I ppm). Guided by this assay, this fraction was further chromatographed over a Mega Bond RP18 solid-phase extraction (SPE) cartridge and then via reversed-phase HPLC to afford four new depsipeptides, wewakpeptin A-D (1-4). The molecular formula of wewakpeptin A (1) was determined as C52H65N7011 on the basis of HR-FABMS and NMR spectral analysis (Table III. 1). From the proton and carbon NMR data (Figure III. I and 2), nine carbonyls accounted for nine of the fourteen degrees of unsaturation implied by the molecular formula. The IR spectrum revealed that I contained both amide (1644 cm1) and ester bonds (1739 cm1), indicating that it was in fact a peptolide. Its peptidic nature was further supported by the presence of two amide NH signals (6 8.11, 6.91) and three N-methylamide signals 61 (6 3.02, 2.98, and 2.75) in the 1H NMR spectrum. Additionally, the 13C NMR spectrum showed two distinctive carbon signals at 6 83.6 and 6 69.3, consistent with a terminal acetylenic functionality. As previously observed, the carbon at 6 69.3 exhibited weak HSQC correlations to a methine proton at 6 1.93, but this proton exhibited a 2JcH HMBC correlation to the quaternary carbon at 6 83.6, confirming the presence of an acetylene (Figure III. 3 and 4)3 The above functionalities accounted for 11 degrees of unsaturation, indicating that wewakpeptin A (1) had three rings. 6.5 ,: Figure Ill. 1. 170 160 Figure III. 2. 6.0 5.5 5.0 a., 4.0 3.5 3.0 2.5 60 50 2.0 1.5 1.0 0.Sppm 1H NMR spectrum of wewakpeptin A (1). 150 140 130 120 110 100 90 80 70 13C NMR spectrum of wewakpeptin A (1). 40 30 20 10 ppm 62 pp 10 S 15 20 25 , 30 gc? 35 40 0 I 45 50 - 0 55 60 65 N 70 75 80 85 B.5 .0 7.5 7.0 6.5 6.0 5.5 5.0 4.5 4.0 i.5 i.0 4.5 Figure III. 3. HSQC spectrum of wewakpeptin A (1). 4.0 1.5 1.0 ppm 63 ppm 0 I S UI 2o 1,4 , I 3o 1 4 I 5: P . II Ih 6 o 'l. $b 0 p 80 U. ' C 9 10 0 11 12 13 14 o I 15 0 16 17 : .0 /. 1.0 b. b.0 .0 .3 .IJ .j.z i.0 I. Figure III. 4. HMBC spectrum of wewakpeptin A (1). .O 1. 1.1.1 u. ppm 64 ppm 0.5 1.0 1.5 2.0 ti'1 . f,.1 2.5 3.0 3.5 4.0 4.5 5.0 5.5. 'P '.4 6.0 6.5 I 7.0 + 7.5. 8.0- q * I 8.5/ 6 5 4 3 2 Figure III. 5. COSY spectrum of wewakeptin A (1). 1 ppm ppm .0- i5.. 0 4 0 e 4 00- 0 m 0 :: 4, ?% ' 40 I I '0 3 . .0 b. .IJ I. I.0 b. .0 U q. S. i.0 2.1) Figure III. 6. HSQC-TOCSY spectrum of wewakpeptin A (1). i. J-.0 u. ppm Table III. 1. NMR Data for Wewakpeptin A (1) and B (2) in CDCI3. Wewakpeptin A (1) Unit Dhoya Position 1 2 3 4 5 6 7 8 lie 9 10 NH 11 N-MeAla 12 13 14 15 16 17 174.2 46.4 79.4 29.8 24.7 18.2 83.6 69.3 26.2 23.1 54.9 39.5 17.6 22.9 12.0 172.3 30.6 4.94, dd (10.0, 2.0) 1.90, 1.38 1.40, 1.36 2, 4, 9, 10, 52 3, 5 4, 6 2.16,m 5,7,8 1.93, brt(2.6) 7 1.28, s 1.26, s 6.91, d (8.2) 5.01, dd (8.2, 2.0) 1.71, m 1.04 1.01, 1.36 0.88, t (7.0) 1,2, 10 1, 2,9 1, ii 12, 13, 16 11 11, 12, 14 15 3.02, s 16, 18 5.86, m 1.17, d (6.6) 16, 17, 19, 20, 21 21 49.2 14.9 170.6 30.3 2.98, 20, 22 22 23 24 25 26 27 59.4 27.6 18.4 20.6 170.4 30.2 5.12, d (10.6) 2.46, m 0.81, d (6.8) 0.95, d (6.8) 20, 21, 23, 25, 26 24, 25 2.75, s 26, 28, 32 28 29 30 65.2 28.0 19.5 20.6 169.7 77.4 29.5 4.40, d (10.6) 2.36, m 0.98, d (7.1) 1.24, d (7.1) 26, 27, 29, 32 32, 34, 35, 36, 37 20.1 4.96, d (5.0) 2.26, m 1.07, d (7.0) 17.6 1.04, d (7.0) 3.72, 3.45, q (8.4) 2.26, 1.92 2.10,2.01 4.64, dd (8.0, 3.7) 39, 41 3.82, 3.60, q (8.2) 2.23, 1.94 1.89, 1.37 4.54, dd (7.2, 3.6) 44, 45, 46 43, 45, 46, 47 31 32 Pro-i 33 34 35 36 37 38 39 40 41 Pro-2 54.8 39.6 17.6 22.9 12.1 S 42 43 44 45 46 47 166.8 47.3 25.6 28.9 58.4 169.9 47.4 25.0 29.4 59.7 172.7 4.92 1.72, 1.28 1.29, 1.15 1.26 1.28 0.85c 1.28 1.26 6.89 5.02 1.71 1.04 1.36, 1.01 0.86 3.02 49.3 14.9 170.6 30.3 2.97 59.5 27.5 5.12 2.46 22, 23,25, 18.4 0.81 22, 23, 24 20.6 170.4 30.2 0.94 65.2 28.0 19.5 20.6 169.7 77.6 29.5 4.39 2.36 0.98 33, 34, 20.1 33, 34,37 17.6 1.07 1.04 18,20 Vat-I Hiv 174.4 46.4 80.2 31.0 26.0 31.7 22.9 14.4 26.2 23.2 172.3 30.6 20 N-MeVai-2 WewakinB (2) 12, 14 18 19 N-Me- HMBCa 8, (J in I-li) 30, 31 28, 29, 31 28, 29, 30 33 38, 40,41 38,39,41,42 38, 39, 40, 42, 43 43, 44,46, 47 44, 45, 47 166.9 47.3 25.5 28.8 58.4 169.9 47.4 24.9 29.4 59.7 172.7 5.83 1.19 2.75 1.25 4.95 2.26 3.77, 3.45 2.26, 1.92 2.11,2.04 4.65 3.80, 3.62 2.25, 1.94 1.85, 1.34 4.57 67 NH Val 8.11,d(9.7) 46,47,48 59.0 4.59, dd (9.7,4.6) 47, 49, 50, 51,52 31.5 19.8 2.27, m 48, 51, 52 50 1.07, d (6.8) 48, 49, 51 51 19.1 1.09, d (6.8) 48, 49, 50 52 171.0 48 49 58.7 31.8 19.8 19.0 171.1 8.08 4.64 2.26 1.08 1.09 a Proton showing HMBC correlation to indicated carbon. b Coupling constants for wewakpeptin B (2) are identical to those given for A (1) except where noted. C Resonance H3-8 was a triplet with 3JHCCH = 7.2 Hz. Two oxygenated the 13C sp3 carbons were indicated by signals at 6 79.4 and 77.4 in NMR spectrum, suggesting the presence of two hydroxy acids in addition to several amino acids. Further NMR analysis revealed a 2-hydroxyisovaleric acid (Hiva) and 7 a-amino acids, including two N-methylvalines (MeVal), two prolines (Pro), one isoleucine (ILe), one valine (Val), and one N-methylalanine (MeAla) (Tablelll. 1). Additionally, a 2,2-dimethyl-3-hydroxy-7-octynoic acid (Dhoya) was evident based on COSY (Figure III. 5) and HMBC correlations (Figure III. 4). Geminal dimethyl protons (H3-9 and H3-IO) showed HMBC correlations to a carbonyl carbon at 174.2 ppm (C-I), a quaternary carbon at 46.4 ppm (C-2), and an oxymethine carbon at 79.4 ppm (C-3). Further, COSY and HSQC-TOCSY (Figure III. 6) allowed connection of protons H-3 to H-6, the later of which showed long range coupling to H- 8 (J = 2.6 Hz). In the HMBC spectrum, the quaternary carbon of the terminal acetylene at 6 83.6 ppm (C-7) showed a two-bond correlations with H-8 (6 1.93) and H-6 (6 2.16, Table Ill. I). Determination of the sequence and connection of amino acid residues and other units (Hiva, Dhoya) in I was achieved primarily by long range 13C-1H correlation experiments (HMBC) with different mixing times and a ROESY experiment. Despite several proton and carbon resonances in I having close or overlapping chemical shifts, two fragments accounting for all seven amino acid and two hydroxy acid units could be constructed from these data (MeVal-MeVal-MeAla-lle-Dhoya; Hiva-Pro-Pro- Val). However, neither the Dhoya nor the Hiva moiety showed the connectivity necessary to complete the sequence. Fortunately, a modified decoupled HMBC pulse sequence (ID HMBC) was effectively employed and showed the necessary correlations 6,7 By irradiation of C-52 (6 171.0), selective couplings were seen to protons at 6 8.11(NH), 4.59 (H-48), and 4.94 (H-3). Similarly couplings to protons at 6 4.40 (H-28) and 4.96 (H-33) were observed when C-32 (ö 169.75) was irradiated, thus completing the sequence of residues in wewakpeptin A (1). Further evidence supporting this sequence was developed from (MS) experiments.8'9 Collisionally induced ES I-MS/MS of the mlz 985 [C52H86N7011 + H] gave ions at m/z 899 (lIe-Dhoya-Val-Pro-Pro-Hiva-MeVaI-MeVal +H), mlz 787 (Ue-Dhoya-Val-Pro- Pro-Hiva-MeVal +H), mlz 673 (lle-Dhoya-Val-Pro-Pro-Hiva +H), m/z 573 (lIeDhoya-Val-Pro-Pro +H), m/z 460 (Dhoya-Val-Pro-Pro +H), and m/z 294 (Val-ProPro +H) (Figure III. 7). These assignments corroborate the NMR results and illustrate the power of (MS) experiments in assigning cyclic peptide structures. m/z 899 (+H) m/z 871 (+H) a m/z 787 (+H) m/z 673 (+1-1) m/z 573 (+H) m/z 460 (+H) m/z 294 (+H) H3 MeVaI MeVaIHivaPro ProVat DhoyaIIe Q wewakpeptin A (1) m/z 920 (+11) mlz 891 (+H)' m/z 835(+H) m/z 721 (+1-1) m/z 573 (+H) m/z 476 (+H) m/z 343 (+H) MeAla MeAlaMeVaIPIa---ProProVaI DhoyaIle wewakpeptin C (3) Figure Ill. 7. Key fragments from collisionally induced ESI-MS/MS experiments with a) wewakpeptin A (1) and b) wewakpeptin C (3). Dhoya = 2,2-dimethyl-3-hydroxy-7octynoic acid; Hiva = 2-hydroxyisovaleric acid; Pla = 3-phenyllactic acid; a,b Fragment ions (= a-type fragment), formed by loss of CO from the respective acylium ions, mlz 899 and m/z 920 (= b-type fragment). The absolute configuration of I was established by analysis of degradation products. A small sample was hydrolyzed with 6N HCI to its constituent amino and rJ hydroxy acid units. These were analyzed by chiral HPLC as well as chiral GC-MS and compared with the retention times of authentic standards. All of the amino acids as well as the Hiva unit were shown to possess L-configuration. While the Dhoya unit was not stable to acid hydrolysis,1° its stereochemistry was determined by comparing the retention time of authentic 2,2-dimethyl-3-hydroxyoctanoic acid (Dhoaa) with that obtained through hydrogenation of I followed by acid hydrolysis and chiral GC-MS. The hydrogenated Dhoya product from I possessed the same retention time as synthetic R-Dhoaa. Proline amide bonds are known to have cis/trans geometry, which correlates with the difference of the proline 13 and y 13C NMR values (.6 compound I, the small values of A6 )5 In (3.3 and 4.5 ppm) seen for Pro-i and Pro-2, respectively, is indicative of a trans geometry for both proline amide bonds in 1, a finding that was supported by ROESY interactions observed between H2-43/H-41, and H2-38/H-33. Wewakpeptin B (2) was isolated by RP-HPLC from the crude fraction containing wewakpeptin A (1). Its high structural homology to I was evident by nearly identical 1H (Figure III. 8) and 13C NMR (Figure III. 9) chemical shifts (Table Ill. 1). However, it displayed an obvious difference in the Dhoya unit. The acetylenic carbons were absent from the 13C NMR spectrum of 2 and were replaced by two additional high-field carbons at 6 22.9 and 14.4, suggesting that 2 was likely the tetrahydro equivalent of I. Indeed, comparison of 1H NMR spectra and FABMS after reducing the triple bond in I over Pd-C confirmed these assignments (Table III. 1). Hydrolysis and stereoanalysis of 2 were not undertaken due to its limited quantity; however, because of the comparable spectroscopic properties of I and 2, we propose they are of the same enantiomeric series. 70 8.5 8.0 7.5 7.0 6.5 6.0 5.5 5.0 4.5 4.0 3.5 3.0 2.5 2.0 1.5 1.0 ppm 60 50 40 30 20 10 pptn Figure III. 8. 1H NMR spectrum of wewakpeptin B (2). 170 160 150 140 130 120 110 100 90 80 70 Figure III. 9. 13C NMR spectrum of wewakpeptin B (2). High-resolution FABMS analysis of wewakpeptin C (3) revealed an [M+H] ion (m/z 1004.6053) consistent with a molecular formula of CMH82N7O11,thus requiring 18 degrees of unsaturation. The IR spectrum of 3 gave characteristic absorption bands for esters and amides at 1741 and 1650 cm1, and the peptidic nature of 3 was again indicated by two exchangeable NH proton resonances at 8.12 and 7.00 and three distinct NCH3 proton singlets at ö 3.00, 2.95 and 2.66. Of the 54 carbon resonances in its 13C NMR spectrum, nine amide/ester carbonyls in the ö 165-1 80 range as well as six 71 characteristic low-field aromatic carbon resonances were observed. Two sp carbons resonating at 6 83.6 and 69.6 suggested a terminal acetylenic functionality in 3, as in wewakpeptin A (1). Oxygenated sp3 carbons were suspected because of signals at 6 79.3 and 74.0 in the 13C NMR spectrum, suggesting the presence of two hydroxy acids. Detailed analysis of 1H (Figure Ill. 10) and 13C NMR (Figure III. II), COSY, HSQC, HSQC-TOCSY, and HMBC NMR experiments (CDCI3) showed that the structure of 3 was closely related to 1, with seven a-amino acid residues including two methylalanines (MeAla), two prolines (Pro), one isoleucine (ILe), one valine (Val), and one methylvaline (MeVal) (Table III. 2). By comparative NMR analysis, one of the non-amino acid moieties in 3 was the hydroxy acid Dhoya. The other hydroxy acid, with proton resonances at 6 7.26-7.33 (H-34 H-38), 6 3.21 and 2.99 (H2-32), and 6 5.37 (H-31), was similar to those of phenylalanine; however, the chemical shift of the a carbon in this residue (C-31, 74.0 ppm) was typical for an oxymethine, thus indicating that this residue was 3-phenyllactic acid (Pla). Pla is a substructure that has been observed in several other marine metabolites, including the molluscan metabolite kulolide-1 (5)11 and the cyanobacterial metabolite symplostatin 3 (6).12 (Table III. 2). 8.5 8.0 I 7.5 1.0 6.5 6.0 5.5 5.0 4.5 4.0 3.5 3.0 Figure UI. 10. 1H NMR spectrum of wewakpeptin C (3). 2.5 2.0 1.5 1.0 ppm 72 170 160 150 140 130 120 110 100 90 80 70 60 50 40 30 20 10 ppm Figure III. 11. 13C NMR spectrum of wewakpeptin C (3). 8.5 8.0 7.0 1.5 6.5 6.0 5.5 5.0 4.5 4.0 3.5 3.0 2.5 2.0 50 40 1.5 1.0 ppm Figure III. 12. 1H NMR spectrum of wewakpeptin D (4). 170 160 Figure III. 13. 150 130 140 130 120 110 100 90 80 70 60 NMR spectrum of wewakpeptin D (4). 30 20 10 ppm 73 TablellI. 2. NMR Spectral Data for Wewakpeptin C (3jand 0 (4) in Unit Dhoya Position 1 2 3 4 lie 5 6 7 8 9 10 NH 11 N-Me- 12 13 14 15 16 17 3H 174.4 46.4 79.3 29.9 HMBC (.J in Hz) N-MeVal 2,4,9, 10, 54 18.3 1.38, 1.33 2.12, m 4, 6 5, 7, 8 83.6 69.6 25.9 31.8 22.9 1.89, brt(2.5) 7 14.5 1.24, S 1,2, 10 1.22,s 1,2,9 26.2 23.2 7.00, d (8.2) 4.98, dd (8.0, 2.2) 1.66, m 0.99 10 1.30, 1.01 0.84 15 24.7 26.11 23.11 54.7 39.4 17.3 23.2 12.3 172.1 30.5 3, 5 12, 13, 16 11, 12, 14 12, 14 Pia 3.00, 49.0 14.5 5.82 1.30 2.95, S 20, 22 5.55, m 1.18, d (6.7) 20, 21, 23, 24 2.66, s 24, 26 26 27 28 29 64.9 28.7 4.19, d (10.0) 2.23, m 0.83 24, 25, 27, 30 28, 29 26, 27, 29 1.01 26,27,28 5.37, t (7.0) 3.21, 2.99, dd (14.4, 3.7) 30, 32, 33, 34,39 31, 33, 34, 39 7.26 7.33 7.28 7.33 7.26 33, 35, 36, 32, 34, 36, 37, 38 35, 37, 38 34, 36, 36, 37 40 41 Pro-2 Vai 42 43 44 45 46 47 48 49 NH 50 51 52 53 54 170.1 22,24 136.1 25.1 29.6 59.8 172.6 59.2 31.7 19.1 19.1 170.6 0.86 16, 17, 19, 20, 21 18, 20 49.7 15.5 171.3 30.6 129.9 129.0 127.6 129.0 129.9 166.6 47.2 25.7 28.9 58.4 170.0 47.6 1.68 1.00 1.30, 0.99 5.85, m 1.25 22 23 24 25 33 34 35 36 37 38 39 4.98 3.00 30.2 31 1.30 0.85c 1.27 1.24 6.95 30.5 21 19.8 20.3 169.7 74.0 37.8 1.27, 1.14 1.26 16, 18 S 14.6 170.1 32 Pro-i 49.0 54.7 39.4 17.3 23.1 12.3 4.88 1.73, 1.29 172.1 18 19 20 30 174.5 46.3 80.2 31.0 4.91, dd (7.0, 2.0) 1.37, 1.87 Ala-i N-MeAla-2 CDCI3. 30.3 2.94 49.8 15.5 171.2 30.7 5.54 64.9 28.8 19.8 20.4 169.7 74.0 37.8 1.21 2.65 4.17 2.22 0.85 1.03 5.34 3.21, 2.98 136.1 1.96,2.24 40,42,43 40,43,44 4.48 40,41,42, 44, 45 3.85, 3.59, q (8.0) 1.71, 2.09 1.89, 1.37 4.54 46, 47 129.9 129.0 127.6 129.0 129.9 166.6 47.3 25.6 28.9 58.4 170.0 47.6 45,47 25.1 8.12, d (9.8) 4.51, dd (9.8, 4.5) 2.20, m 1.02, d (6.8) 1.04, d (6.8) 49 49, 51, 52, 53, 54 50, 52, 53 3.58, 3.12, q (7.7) 1.95, 2.27 41, 45, 46, 48 44, 45, 46, 47, 49 50, 51,53 50, 51, 52 29.5 59.8 172.5 59.0 31.7 19.2 19.0 170.9 7.26 7.33 7.28 7.33 7.26 3.62, 3.10 1.96, 2.29 1.96, 2.24 4.49 3.63, 3.83 1.66, 1.96 1.87, 1.35 4.57 8.10 4.56 2.22 1.04 1.06 74 a Proton showing HMBC correlation to indicated carbon. bcoupling constants for wewakpeptin 0 (4) are identical to those given for C (3) except where noted. C Resonance H3-8 was a triplet with 3JHCCH = 7.2 Hz. I OCH OCH3rj O"OH II 0 Symplostatin 3 (6) Kulolide-1 (5) Nj 0 0 1OCH3 I 0 O,CH3,O 0 0 0 LU 103793(8) Dolastatin 15(7) The sequencing of residues in 3 was achieved primarily by long range 13C-1H correlation experiments (HMBC) with different mixing times as well as a ROESY experiment. The modified decoupled HMBC pulse sequence (1D HMBC) was also employed as in the structure elucidation of 1, and indicated two fragments (MeValMeAla-MeAla-lle-Dhoya; Pla-Pro-Pro-Val) which accounted for all seven amino acid and the two hydroxy acid units. Collisionally induced ESI-MS/MS of the m/z 1005 [CMHB2N7O11 + gave ions at m/z 920 (lle-Dhoya-Val-Pro-Pro-Pla-MeVal-MeAla +H)4, m/z 835 (lle-Dhoya-Val-Pro-Pro-Pla-MeVal +H), mlz 721 (Dhoya-Val-Pro-Pro- Pla-MeVal +H), mlz 573 (lle-Dhoya-Val-Pro-Pro +H), m/z 476 (lle-Dhoya-Val-Pro- +H), and m/z 343 (Pro-Pro-Pla +H)4 (Figure III. 7) whose assignment corroborated the NMR results and completed the linkages between the structural units. Chiral HPLC and GC-MS analysis of the acid hydrolysate showed the presence of L-MeAla, L-MeVal, L-Pro, L-Val, L-lle, and D-Pla. The configuration of Dhoya was determined 75 as R using the method as described above for 1. Curiously, the Pla residue in 3 has the opposite stereochemistry compared with the Hiva residue of 1. HR-FABMS of wewakpeptin D (4) indicated that the molecular weight was four mass units higher than that of wewakpeptin C (3), and NMR data (Figure III. 12-13) indicated that it was closely related to 3. However, it also displayed obvious differences in the Dhoya unit in that it lacked the triplet at 6 1.89 for H-6 and the acetylenic carbons at 6 83.6 and 69.3. Chemical reduction (Pd-C, H2) of the triple bond in 3 produced 4, confirming these assignments (Table Ill. 2). Stereoanalysis of 4 was not undertaken due to its limited quantity; however, due to the highly comparable spectroscopic properties of 4 and 3, we again propose that they also are of the same enantiomeric series. The wewakpeptins were tested for cytotoxicity to NCI-H460 human lung tumor and neuro-2a mouse neuroblastoma cells (Figure Ill. 14). Intriguingly, wewakpeptin A and B were approximately 10-fold more toxic than C and 0 to these cell lines. The LC for wewakpeptin A was 0.49 pM and 0.65 pM for neuro-2a and H460 cells, respectively, and 0.20 pM and 0.43 pM, respectively, for wewakpeptin B. The LC50 for wewakpeptin C was 10.7 pM and 5.9 pM for neuro-2a and H460 cells, respectively, and 1.9 pM and 3.5 pM, respectively, for wewakpeptin D. These cyclic peptides most likely derive from a nonribosomal polypeptide synthetase (NRPS) pathway,1 and thus, the structural variation of the wewakpeptins is intriguing and might suggest that adenylation domains with relaxed substrate specificity are involved in their biosynthesis. Hydrolysis of the two ester bonds in the wewakpeptins yields two linear peptides in each case, and these are structurally related to the dolastatins (e.g. dolastatin 15 (7), and LU 103793 (8)), and thus, it is conceivable that the wewakpeptins undergo these reactions in nature to release more potent fragments. 76 I 0 U 0 0 20 C.) C.) -8 Dose (log M) -7 -6 -5 Dose (log M) I 1' U U .2 .2 C) C.) 0 -7 -6 -5 .4 -7 Dose (log M) -6 -5 -4 Dose (log M) Figure III. 14. Cytotoxicity of wewakpeptins A-D, panels A-D respectively, to H460 cells, solid lines and neuro 2-a cells, broken lines. Cells were exposed to the indicated concentration of chemical for 48 hr before assessment of toxicity by MU reduction. Where error bars are not present, they are occluded by the data point. EXPERIMENTAL General Experimental Procedures. Optical rotations were measured on a Perkin-Elmer 141 polarimeter. IR and UV spectra were recorded on Nicolet 510 and Beckman DU64OB spectrophotometers, respectively. NMR spectra were recorded on Bruker Avance DPX 400 MHz and Bruker Avance 300 MHz spectrometers with the solvent CDCI3 used as an internal standard (6H at 7.26, & at 77.4). High resolution mass spectra were recorded on a Kratos MS-50 TC mass spectrometer. Tandem mass spectrometric data were obtained on an electrospray ionization (ESI) quadrupole ion trap mass spectrometer (Finnigan LCQ, San Jose, CA). For ESIMS/MS analysis, samples were injected onto a Cl 8 trap column for desalting and 77 introduced into the mass spectrometer by isocratic elution using 50% acetonitrile containing 0.1 % formic acid. For MS/MS investigations, the protonated molecular ion clusters were isolated in the ion trap and collisionally activated with different collision energies to find optimal fragmentation conditions. For the most intense fragment ions MS3 and MS4 experiments were performed. Chiral GC-MS analysis was accomplished on a Hewlett-Packard gas chromatograph 5890 Series II with a Hewlett-Packard 5971 mass selective detector using an Alltech capillary column (CHIRASIL-VAL phase 25 mx 0.25 mm). HPLC was performed using Waters 515 HPLC pumps and a Waters 996 photodiode array detector. Collection. The marine cyanobacterium Lyngbya semiplena (voucher specimen available from WHG as collection number PNG12-7Dec99-3) was collected from shallow waters (1-3 m) in Wewak Bay, Papua New Guinea, on December 7, 1999. Taxonomy was assigned by microscopic comparison with the description given byDesikachary.13 The material was stored in 2-propanol at -20°C until extraction. Extraction and Isolation. Approximately 138 g (dry wt) of the alga were extracted repeatedly with CH2Cl2/MeOH (2:1)to produce 3.05 g of crude organic extract. The extract (3.0 g) was fractionated by silica gel vacuum liquid chromatography using a stepwise gradient solvent system of increasing polarity starting from 10% EtOAc in hexanes to 100% MeOH. The fraction eluting with 100% MeOH was found to be active at I ppm in the brine shrimp toxicity assay. This fraction was further chromatographed on Mega Bond RP18 solid-phase extraction (SPE) cartridges using a stepwise gradient solvent system of decreasing polarity starting from 80% MeOH in H20 to 100% MeOH. The most active fractions after SPE (85% toxicity at I ppm to brine shrimp) were then purified by HPLC [Phenomenex Sphereclone 5 p ODS (250 x 10 mm), 9:1 MeOH/H20, detection at 211 nm] giving compounds 1 (5.0 mg), 2 (0.7 mg), 3 (2.0 mg), and 4 (0.7 mg). Wewakpeptin A (1): colorless amorphous solid; UV (MeOH) A [aJo -45° (c 0.40, CHCI3); 216 nm (log c 4.6); IR (neat) 3327, 2926, 2875, 1739, 1644, 1454, 1243, 979 cm; 1H and 13C NMR data, see Table 1; HR FABMS m/z[M + H] 984.6262 (calcd for C52H86N7011, 984.6302). Wewakpeptin B (2): colorless amorphous solid; UV (MeOH) Amax 215 nm (log [a]26D -53° (c 0.47, CHCI3); 4.6); IR (neat) 3316, 2928, 2873, 1739, 1650, 1461, 1242, 976 cm; 1H and 13C NMR data, see Table 1; HR FABMS m/z[M HJ 988.6843 (calcd for C52HN7O11, 988.6701). Wewakpeptin C (3): colorless amorphous solid; [a]260 -56° (c 0.27, CHCI3); UV (MeOH) Amax 220 nm (log E 4.5); IR (neat) 3340, 2932, 2875, 1741, 1650, 1446, 1242, 980 cm'; 1H and 13C NMR data, see Table 2; HR FABMS m/z EM + H] 1004.6053 (calcd for CH82N7O11, 1004.6072). Wewakpeptin D (4): colorless amorphous solid; [a]26D -65° (c 0.60, CHCI3); UV(MeOH)Amax 221 nm(log E4.5); IR (neat) 3343, 2924, 2851, 1741, 1654, 1457, 1244, 989 cm1; 1H and 13C NMR data, see Table 2; HR FABMS m/z [M + H] 1008.6276 (calcd for CH86N7O11, 1008.6380). Absolute stereochemistry of 1. Wewakpeptin A (1, 500 pg) was hydrolyzed in 6 N HCI at 105 °C for 16 h, then dried under a stream of N2 and further dried under vacuum. The residue was reconstituted with 300 pL of H20 prior to chiral HPLC analysis [Phenomenex Chirex 3126 (D), 4.6 x 250 mm; UV 254 nm detector], mobile phase I: 100% 2 mM CuSO4 in H20, flow rate 0.7 mL/min; [column, Phenomenex chirex 3126 (0), 4.6 x 50 mm; UV 254 nm detector], mobile phase II: 2 mM CuSO4 in MeCN/H20 (15:85), flow rate 0.8 mL/min. Mobile phase I elution times (tR, mm) of authentic standards: L-MeAla (16.7), D-MeAla (17.2), L-MeVal (24.0), D- MeVal (39.0), L-Pro (28.4), D-Pro (63.4), L-Val (38.5), D-Val (68.9). Mobile phase II elution times (tR, mm) of authentic standards: L-Hiv (9.2), D-Hiv (14.5). The hydrolysate was chromatographed alone and co-injected with standards to confirm assignments of L-MeAla, L-MeVal (2 eq), L-Pro (2 eq), L-Val, and L-Hiv. The presence of L-lle was confirmed by chiral GC-MS using established methods.14 Determination of the configuration of the Dhoya unit was established through hydrogenation (10% Pd-C, H2), acid hydrolysis (6 N HCI, 105 °C for 14 h), and chiral GC-MS analysis of the 2,2-dimethyl-3-hydroxyoctanoic acid (Dhoaa) residue as direct hydrolysis of wewakpeptin A did not yield a sufficient amount of 2,2-dimethyl-3hydroxy-7-octynoic acid (Dhoya) for analysis. For chiral GC-MS analysis of methyl-Dhoaa, portions of each standard R- and S-Dhoaa (obtained as the gift from professor T. Ye at Hong Kong Polytechnic University) were separately diluted in 50 pL of MeOH and treated with diazomethane for 10 mm. Excess diazomethane and solvent were removed with a stream of N2, 79 and the residues were resuspended in CH2Cl2. Capillary GC-MS analysis was conducted using a Chirasil-Val column (Altech, 25 m x 0.25 mm) using the following conditions: column temperature held at 40 °C to 100 °C at a rate of 3 °Clmin, then from 100 °C to 150 °C at a rate of 15 °Clmin. The retention time of the methylated Dhoaa residue derived from hydrogenation and hydrolysis of I and 3, followed by methylation (CH2N2) matched that of the methylated R-Dhoaa standard (34.6 mm) but not the methylated S-Dhoaa standard (35.2 mm). Absolute stereochemistry of 3. The wewakpeptin C (3, 500 pg) hydrosylate (6 N HCI, 105 °C, 16 h) was worked up and analyzed as described above for the wewakpeptin A hydrolysate. The following residues co-eluted with wewakpeptin C hydrosylate peaks; mobile phase I: L-MeAla (16.3 mi, L-MeVal (25.9 mm), 2 equivalent L-Pro (28.3 mm), L-Val (38.2 mm); Mobile phase II: D-Pla (66.0 mm). The presence of L-lle and R- Dhoya were confirmed by chiral GC-MS as described above for the wewakpeptin A. Biological Activity. Brine shrimp (Artemia sauna) toxicity was measured as previously described.15 After a 24 h hatching period, aliquots of a 10 mg/mL stock solution of compounds A-D were added to test wells containing 5 mL of artificial seawater and brine shrimp to achieve a range of final concentrations from 0.1 to 100 ppm. After 24 h the live and dead shrimp were tallied. Cytotoxicity was measured in NCI-H460 human lung tumor cells and neuro-2a mouse neuroblastoma cells using the method of Alley et. determined by MU reduction.17 al'6 with cell viability being Cells were seeded in 96-well plates at 6000 cells/well in 180 p1 medium. Twenty-four hours later, the test chemical was dissolved in DMSO and diluted into medium without fetal bovine serum and then added at 20 pg/well DMSO was less than 0.5% of the final concentration. After 48 hr, the medium was removed and cell viability determined. REFERENCES (1) Gerwick, W. H.; Tan, L. T.; Sitachitta, N. In The Alkaloids; Cordell, G. A., Ed.; Academic Press: San Diego, 2001; Vol. 57, pp 75-184. (2) Han, B.; McPhail, K. L.; Ligresti, A.; Di Marzo, V.; Gerwick, W. H. J. Nat. Prod. 2003, 66, 1364-1 368. (3) Sitachitta, N.; Williamson, R. T.; Gerwick, W. H. J. Nat. Prod. 2000, 63, 197-200. (4) Wan, F.; Erickson, K. L. J. Nat. Prod. 2001, 64, 143-146. (5) Siemion, I. Z.; Wieland, T.; Pook, K. Angew. Chem., Int. Ed. Engi. 1975, 14, 702. (6) Meissner, A.; Sorensen, 0. W. Magn. Reson. Chem. 2001, 39, 49-52. (7) Nogle, L. M.; Marquez, B. L.; Gerwick, W. H. Org. Left. 2003, 5, 3-6. (8) Kuroda, J.; Fukal, T.; Nomura, 1, 1. J. Mass. Spectrom. 2001, 36, 3037. (9) Ngoka, L. C. M.; Gross, M. L. J. Am. Soc. Mass Spectrometry 1999, 10, 732-746. (10) (a) Nakao, Y. ; Yoshida, W. Y. ; Szabo, C. M.; Baker, B. J.; Scheuer, P. J. J. Org. Chem. 1998, 63, 3272-3280. (b) Luesch, H. ;Pangilinan, R. ; Yoshida, W. Y.; Moore. R. E.; Paul, V. J. J. Nat. Prod. 2001, 64, 304-307. (11) Reese, M. T.; Gulavita, N. K.; Nakao, Y.; Hamann, M. T.; Yoshida, W. V.; Coval, S. J..; Scheuer, P. J. J. Am. Chem. Soc. 1996, 118, 11081-11084. (12) Luesch, H. ; Yoshida, W. V.; Moore, R. E.; Paul, V. J.; Mooberry, S. L. ; Corbett, T. H. J. Nat. Prod. 2002, 65, 12-20. (13) Desikachary, T.V. Cyanophyta; Indian Council of Agricultural Research: New Delhi, India, 1959; p686. (14) Trimurtulu, G.; Ohtani, I.; Patterson, G. M. L.; Moore, R. E.; Corbett, T. H.; Valeriote, F. A.; Demchik, L. J. Am. Chem. Soc. 1994, 116, 472937. 81 (15) Meyer, B. N.; Ferrigni, N. R.; Putnam, L. B.; Jacobsen, L. B.; Nichols, 0. E.; McLaughlin, J. L. Planfa Med. 1982, 45, 31-3. (16) Alley, M. C., Scudiero, D. A. CancerRes. 1988, 48, 589-601. (17) Manger, R. L., Leja, L. S. J. AOAC. mt. 1995, 78, 521-527. CHAPTER FOUR ISOLATION AND STRUCTURE OF LYNGBYABELLIN DERIVATIVES FROM A PAPUA NEW GUINEA COLLECTION OF THE MARINE CYANOBACTERIUM LYNGBYA MAJUSCULA ABSTRACT Five new lyngbyabellin analogs along with the known compound, dolabellin, have been isolated from the marine cyanobacterium Lyngbya majuscula collected from Papua New Guinea. The structures of lyngbyabellins E-I (1-5) were elucidated through extensive spectroscopic analysis, including HR-FABMS, 1D and 2D NMR experiments. The absolute configurations of 1 and 4 were ascertained by chiral HPLC and GC/MS analysis of degradation products, in combination with NMR experiments. All five lyngbyabellins showed cytotoxicity to NCI-H460 human lung tumor and neuro-2a mouse neuroblastoma cell lines with LC50 values between 0.2 and 4.8 j.tM. INTRODUCTION Cyanobacteria are phenomenal producers of structurally intriguing and biologically active secondary metabolites.1'2 The pantropical marine cyanobacterium Lyngbya majuscula Gomont (Oscillatoriaceae) is one of the more prolific producers of interesting secondary metabolites, yielding no fewer than 150 reported compounds.1 As part of an effort to discover new and biologically active natural products, we now report the isolation and structure elucidation of a series of new lyngbyabellin analogs, lyngbyabellins E-1 (1-5), and the known compound dolabellin (6), originally isolated from the sea hare Do/abel/a auricularia.3 The sea hare, D. auricularia has yielded several cytotoxic agents structurally similar to cyanobacterial counterparts, including the anticancer agent dolastatin-1 0 and the cytotoxic agent, dolabellin.3'4 The isolation of dolastatin-lO (7) and related compounds, including symplostatin-1 (8), from the marine cyanobacteria Symploca spp.,5 and the dolabellin-like compounds, lyngbyabellins A-D (9-12) and hectochlorin(13) 6° from L. majuscula, suggests that these sea hares sequester bioactive metabolites from their cyanobacterial diet. Lyngbyabellin G (3) R = OH Lyngbyabeilin E (1) R = OH Lyngbyabellin H (4) R = H Lyngbyabellin F (2) R = OH Lyngbyabellin I (5) R = H Dolabellin (6) N ,,... I OCH3 o OCH3 o \=J Dolastatin 10 (7) XAN1AS -' N I 0 OCH3O OCH3O Symplostatin 1 (8) Lyngbyabellin A (9) Lyngbyabellin B (10) Lyngbyabellln U (12) Lyngabyabellin C (11) Hectochlorin (13) Results and Discussion A shallow water (1-3 m) strain of L. majuscula was collected by hand from Alotau Bay, Papua New Guinea, in 2002. The alga was extracted with CH2Cl2/MeOH (2:1) and fractionated by silica gel vacuum liquid chromatography. Preliminary bioassay of the EtOAc/hexanes (9:1) eluted fraction showed toxicity in the brine shrimp model (LD - I ppm). Guided by this assay, this fraction was further chromatographed over a Mega Bond RP18 solid-phase extraction (SPE) cartridge and then via reversed-phase HPLC to afford five new lyngbyabellin analogs E-1 (1-5), plus dolabeHin (6). The molecular formula of Iyngbyabellin E (1) was determined as C37H51C12N3012S2 on the basis of HR-FABMS and NMR spectral data (Table 1). The ratio of [M+H1 isotope peaks, 5:4:1, at m/z 862/864/866, clearly indicated the presence of two chlorine atoms. From the 1H and 13C NMR data (Figure IV. 1), six carbonyls, two carbon-carbon double bonds, and two additional carbon-heteroatom double bonds accounted for ten of the thirteen degrees of unsaturation implied by the molecular formula. Two downfield 1H singlets at 6 8.12 (H-12) and 8.27 (H-18), HSQC-correlated to carbons at 6 128.7 and 130.1 (Figure IV. 2), were consistent with the presence of two 2, 4-disubstituted thiazole rings, which assigned two of the remaining three degrees of unsaturation. The final degree of unsaturation could be accounted for by an additional ring within the structure of 1. The two thiazole ring structures were confirmed by HMBC correlations from H- 12 to C-Il (8c 146.4) and C-13 (6c 165.9), and H-18 to C-17 (3c 145.9) and C-19 (6c 165.0), respectively (Table IV. 1, Figure IV. 3). Furthermore, three-bond correlations from H-12 and H-18 to conjugated carbonyl carbons at 6c 161.2 (C-b), and 161.9 (C16), respectively, were indicative of carboxylic acid derived functionalities attached to these heterocycles. HMBC correlations from a 1H methine triplet at 6 6.45 (H-14) to the thiazole C-I 3 (öc 165.9) and an oxymethylene carbon at 6 64.4 (C-I 5) established a 2-( I ,2-dihydroxyethyl)thiazole-4-carboxylate unit. The second thiazole ring formed a 2-(1 ,2-dihydroxy-2-methylpropyl)thiazole-4-carboxylate unit as well based on HMBC correlations from an oxymethine singlet at 6 5.65 (H-20) to thiazole C-19 (6c 165.0) and an oxygenated quaternary carbon at 6 72.1 (C-21), which was in turn showed 3-bond correlations to two methyl singlets (6 1.18 and 1.34, H3-22 and H3-23, respectively). Inspection of the 1H NMR spectrum of I revealed a series of upfield and highly coupled resonances indicative of an aliphatic chain. A downfield methyl singlet at 6H 2.09 (H3-8) showed HMBC correlations to a quaternary carbon at 6c 90.4 (C-7) and a methylene carbon at 6c 49.3 (C-6). The chemical shift of C-7 was indicative of a gem-dichioro subtituent as observed in dolabellin (6), hectochlorin, and lyngbyabellins A-D. HSQC-TOCSY was used to extend this moiety to include an additional six carbons (C-2 - C-6, and C-8), identifying this unit as 7,7-dichloro-3-acyloxy-2methyloctanoate (DCAMO) (Figure IV. 4). Further analysis of 2D NMR data identified acetate and butyric acid moieties as well. The remaining unassigned C8H14NO2 was clearly attached to C-14 via an ester linkage, based on a 3JCH correlation from H-14 to C-24. A 1JCH from H-26 (6 5.17) to a downfield carbon at 72.6 ppm indicated that this carbon was oxygenated, while COSY correlation to the methylene protons (H2-25) and two HMBC correlations from H-26 to carbonyls at 169.2 and 170.6 (C-24, C-36) established the acetylated structure depicted. An exchangeable amide proton signal at 6 5.60 (27-NH) showed a strong COSY correlation to H-27 (Figure IV. 5), which was expanded into a modified leucine unit (C-24 to C-31) based on the extensive 2D NMR correlations. The HMBC data summarized in Table 1 allowed us to connect the partial structures and functional groups described above. Although the ester linkage between C-3 and C-b was not evidenced by the HMBC experiments, the 1H and 13C data were strongly supportive of this remaining linkage, and were needed to account for the final degree of unsaturation calculated from the molecular formula. The planar structure of lyngbyabellin E is thus represented by I, closely related to the recently reported structure of lyngbyabellin D.9 'I 8.5 8.0 7.5 7.0 6.5 6.0 170 160 150 140 130 120 5.5 110 5.0 100 4.5 90 4.0 80 3.5 70 Figure IV. 1. 1H and 13C spectra of Iyngbyabellin E (1). 3.0 60 2.5 50 2.0 40 1.5 3i 1.0 20 ppm ppm Table IV. 1. NMR Data for Lyngbyabellin E (1) in CDCI3. Lyngbyabeuin E (1) H (J in Hz) Position 1 3.00, dq (9.5, 7.5) 5.26, m 1, 3, 9 2, 4 1.80,1.75,m 3,5 1.75, 1.73, m 2.23, 2.15, m 6 5, 7 2.09, s 6, 7 1.25,d(7.5) 1,2,3 8.12,s 10,11,13 6.45, dd (5.4, 7.3) 4.99, dd (11.4, 5.4) 4.50, dd (11.4, 7.3) 13, 15, 24 13, 14, 16 130.1 8.27, s 16, 17, 19 20 165.0 76.0 5.65, s 1, 19,21 21 72.1 3.63, brs 1.18,s 1.34,s 20, 21,22 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18 19 21-OH 22 23 24 25 26 27 27-NH 28 29 30 31 32 33 34 35 36 37 a 174.4 43.3 74.6 31.4 20.9 49.3 90.4 37.6 14.4 159.6 146.4 128.7 165.9 70.6 64.4 HMBCa 160.5 145.9 27.4 26.1 169.1 36.5 72.6 49.2 40.0 25.0 21.6 23.8 2.81, dd (16.0, 4.9) 2.74, dd (16.1, 7.7) 5.17, ddd (4.9, 5.5, 7.7) 4.32, m 5.60, d (9.4) 1.28, m 1.65, m 0.90, d (6.5) 0.93, d (6.3) 28, 30, 31 28, 29, 31 28, 29, 30 2.12, t (7.2) 1.62, m 0.89, t (7.2) 32, 34, 35 32, 33, 35 33, 34, 2.05,s 36 24, 26,27 27, 36 26, 28 27, 29,30 173.1 38.9 19.4 13.9 170.6 21.2 Proton showing HMBC correlation to indicated carbon. ppm 'C 2C I 30 'I 40 50 60 70 4 ' 80 90 100 110 120 44 130 ...................................... 9 8 7 6 5 4 3 Figure IV. 2. HSQC spectrum of Iyngbyabellin E (1). 2 1 ppm ppm D .. I * 10- I . I 4* 1 0. 10- 0- I, I 1 C p I. 9.0 .5 8.0 1.5 7.0 b.5 b.0 5.5 5.0 4.5 4.0 3.5 3.0 2.5 2.0 Figure IV. 3. HMBC spectrum of Iyngbyabellin E (1). 1.5 1.0 ppm 91 pp." 1. 1, 1. .5 B.0 7. 7.0 6.5 6.0 5.5 5.0 4.5 4.0 3.5 3.0 2.5 2.0 1.5 Figure IV. 4. HSQC-TOCSY spectrum of Iyngbyabellin E (1). 1.0 0.5 ppm ppm 1 4 - 2 - 3 4 . - S V 5 I. 6 7 + 8 9 8 9 7 6 5 4 3 2 1 ppm Figure IV. 5. COSY spectrum of lyngbyabellin E (1). Compound I posed a number of interesting problems in assignment of stereochemistry. (S)- and/or (R)- a,13-dihydroxyisovaleric acid (Dhiv) could not be detected in the acid hydrolyzate by comparison with the retention times of synthetic standards.11 It appeared that the a,3-dihydroxy acid was unstable under the conditions of acid hydrolysis. However, ozonolysis and subsequent base hydrolysis at 90 °C afforded (S)- aj3-dihydroxyisovaleric acid, without any detectable racemization, as demonstrated by chiral GC-MS analysis of both synthetic (S)- and 93 (R)- a,13-dihydroxyisovaleric methyl esters and the hydrolyzed natural product. Thus, C-20 in compound I possessed an S configuration, which is identical to that found in the aj3-dihydroxyisovaleric acid of lyngbyabellin A6 and hectochlorin.1° Chiral HPLC firmly established the stereochemistry of the glyceric acid moiety, and thus lyngbyabellin E possessed an R configuration at C-14. There are few reports of and no general method for determining the configuration of y-amino--hydroxy acids. However, reports in the literature have indicated that 4-amino-3-hydroxy-5-methylhexanoic acid, such as found in the lyngbyabellin D, undergoes an epimerization at C-3 via an acid-catalyzed dehydration/hydration sequence, and that significant quantities of the intermediate aj3-unsaturated acid is produced upon prolonged hydrolysis.9'12 This suggested the absolute configuration of C-27 could be determined by acid hydrolysis, subsequent ozonolysis, and oxidative workup to D- or L- leucine. Indeed, this reaction sequence gave leucine, and chiral HPLC showed unambiguously that C-27 was derived from L- leucine. Knowing the absolute configuration at C-27, the stereochemistry at C-26 was defined using homonuclear coupling constant information and selective I D NOE experiments in combination with a systematic analysis of all appropriate energetically favored (staggered) rotamers (Figure IV. 6). Enhancement of the resonances for H26 and H2-25 was observed following low power irradiation at ö 4.32 (H-27), ruling out model A2, A3, B2 and B3 (Figure IV. 6). The 'H-1H coupling constant of 5.5 Hz between H-26 and H-27 suggested a gauche relationship between the two protons, consequently supported ruling out again model A3 and B2. Irradiation at 5.60 (NH27) led to enhancement of the signals for H-26 (ruling out model Al and again B3), H27, H2-28, H-29 and H2-33. The remaining model BI fulfilled all criteria, suggesting a 26R, 27S configuration for the y-amino-3-hydroxy acid residue of lyngbyabellin E (I). 94 35 3LOAc A C-28 Ac0,-5H HN H C-25 > C-28 C-25-0Ac C32-N HH Al H C-28 H3C-25 C-32-.N H I H OAc A2 A3 35 3Q B HN OAc ZJ26 125 C28 Ac0..A..C-25 C32NAJH HH BI C28 H5ç0AC C-32-...N i H C2I C 25 C-32NH H C-25 H OAc B2 B3 Figure IV. 6. Diagram of all possible rotamers for the two possible epimers 26S,27S (A series) and 26R,27S (B series) of Lyngbyabellin E (1). The relative stereochemistry of the polyketide-derived 3-hydroxy acid (DCAO) was assigned from NOE and hetero half-filtered TOCSY (HETLOC) experiments. NOE correlations were observed between H-3 and CH3-9, and between CH3-9 and H2-4. In combination with a large 3JH coupling of 9 Hz between H-2 and H-3, these data defined an anti relationship between H-2 and H-3, in an S,S or R,R configuration. In the HETLOC spectrum, a small heteronuclear coupling of 3JH3-C9 = 3.0 Hz was consistent with a gauche arrangement of H-3 and CH3-9, while a larger 2JH2-C3 (5.2 Hz) also implied a gauche relationship between H-2 and the oxygen of the ester linked (dihydroxyethyl)thiazole carboxylate moiety. Other JCH values for I were 0.3 Hz (H-3/C-2), -3.0 Hz (H-3/C-9), 2.6 Hz (H-9/C-2) and -5.3 Hz (H-9/C-3). All of these data were comparable to those for hectochlorin (13), the 2S, 3S configuration of which was determined by X-ray.13 Conformational analysis of both I and 7 by standard calculation methods (MM2*) based on the crystal structure of 7 resulted in good structural overlay of this energetically favored 2S, 3S configuration. Therefore, given also that all other members of this family possess a 3S configuration,3' 610 these data support the stereochemical assignment of lyngbyabellin E (I) as 2S, 3S, 14R, 20S, 26R, and 27S. 95 Lyngbyabellin F (2) was isolated by RP-HPLC from a slightly more polar fraction than that containing Lyngbyabellin E (1). Its high structural homology to I was evident from nearly identical 1H and 13C NMR chemical shifts (TablelV. 2, Figure IV. 7). However, the molecular formula of 2 was established as CH55Cl2N3O13S2 by HR-FABMS, indicating twelve degrees of unsaturation. These could be ascribed to six carbonyls, two carbon-carbon double bonds, two additional carbon-heteroatom double bonds, and two rings. The presence of a methoxy singlet at 395 (H-38) in conjunction with a free hydroxyl group at C-14 (Table IV. 2), suggested the linear nature of 2. Moreover, C-24 (169.4 ppm) was clearly attached to the 3-hydroxyl group of the glyceric acid-derived moiety via an ester linkage, rather than to the a- hydroxyl group in the structure of I, based on an HMBC correlation from H-15 to C24. Therefore 2 was a linear lipopeptide, structurally similar to the previously isolated lyngbyabellin D. High-resolution FABMS analysis of lyngbyabellin G (3) revealed a [M + H1 at 595.0733, consistent with a molecular formula of C23H29C12N2O8S2, thus requiring 10 degrees of unsaturation. Careful examination of ID and 2D NMR data for 3 indicated that 3 possessed the same structure as I from C-I through C-23, but lacked the acyclic moiety from C-24 to C-37 (Figure IV. 8). In this respect, the planar structure of 3 was almost identical to previously isolated lynbyabellin C, which has an aj3dihydroxy43-methylpentanoic acid unit instead of the aj3-dihydroxyisovaleric acid present in 3. Intriguingly, one ester linkage of lyngbyabellin E (1) appeared to be particularly prone to methanolysis and hydrolysis, as treatment of I with MeOH and H20 caused it to convert slowly to 3 by regioselective ester cleavage at C-14. Hence, lyngbyabellin G may be an artifact of I from storage of the collection in ethanol and seawater. Hydrolysis and stereoanalysis of 2 and 3 were not undertaken due to their limited quantity; however, because of the comparable spectroscopic properties of I, 2, and 3, we propose that they are of the same enantiomeric series. ......................................................... 6.5 6.0 5.5 5.0 4.5 4.0 3.5 3.0 2.5 2.0 1.5 1.0 -.-I-. 8.5 8.0 7.5 7.0 140 140 140 4o 90 0 70 60 50 40 30 I ppm 20 ppa Figure IV. 7. 1H and 13C of spectra of Iyngbyabellin F (2). 170 160 150 140 130 120 110 100 90 80 70 60 Figure IV. 8. 1H and 13C of spectra of Iyngbyabellin G (3). 50 40 30 20 ppm Table IV. 2. NMR Data for Lyngbyabellin F (2) and G (3) in Lyngbyabellin F (2) Position 1 2 3 4 5 6 7 8 9 10 11 12 13 14 14-OH 15 16 17 18 19 20 21 22 23 24 25 26 27 27-NH 28 29 30 31 32 33 34 35 36 37 38 CDCI3. Lyngbyabellin G (3) (J in Hz) 171.3 43.9 75.1 31.5 21.8 49.3 90.4 37.6 13.7 161.2 146.7 129.3 173.4 70.0 68.7 161.9 146.4 128.8 167.7 78.7 72.2 29.9 27.0 169.4 36.9 174.1 3.02, dq (4.8, 7.0) 5.44, m 1.80, m 1.78, m 2.25, 2.22, m 2.12, s 1.29,d(7.0) 8.21, s 5.32, m 5.82, d (5.4) 4.60, dd (11.3, 3.2) 4.46, dd (11.3, 8.7) 69.2 3.16, dq (9.5, 7.3) 5.35, m 1.95, 1.77, m 1.78, m 2.29, 2.17, m 2.12, s 1.30,d(7.3) 8.17, s 5.43, m 6.06, d (10.0) 4.81, dd (11.6,2.3) 4.56, dd (11.6, 2.9) 161.1 8.17, s 6.14, s 1.26,s 1.38, s 2.76, dd (15.5, 6.1) 2.62, dd (15.7, 8.0) 5.20, ddd (3.9, 6.2, 7.9) 73.2 49.1 4.30,m 38.1 5.52, d (9.3) 1.40, 1.35, m 24.9 21.5 23.9 173.6 39.0 19.4 13.9 170.9 21.2 52.7 43.3 75.2 30.9 20.7 49.7 90.5 37.8 15.2 160.3 146.0 128.5 170.5 70.2 1.61,m 0.90, d (6.5) 0.95, d (6.5) 2.17, t (7.2) 1.65,m 0.94, t (7.2) 2.09, s 3.95, s 146.2 130.0 166.2 78.3 72.2 27.2 25.9 8.22, s 5.59, s 1.32, s 1.36, s HR-FABMS of lyngbyabellin H (4) indicated that its molecular formula of C37H51C12N3011S2, which is one less oxygen than in lyngbyabellin E (1). 1D NMR data indicated that lyngbyabellin H (4) was also closely related to I (Figure IV. 9, Table IV. 3). However, it displayed obvious differences in the a,f3-dihydroxyisovaleric acid (Dhiv) unit in that the singlet at 6 5.65 for H-20 in I was replaced by a doublet at 6 5.52 in the spectrum for 4. Moreover, the oxygenated quaternary carbon C-21 (6 72.1) in I was replaced by an upfield methine carbon (6 32.5) in 4, thus indicating that this residue was 2-hydroxyisovaleric acid (Hiva). From extensive 2D NMR analysis, the planar structure of lyngbyabellin H (4) was otherwise the same as lyngbyabellin E (I), and conceptually, also a precursor of dolabellin (6), the original compound in the series discovered from the sea hare Do/abe/Ia auricularia. Ester bond cleavage at C24 in 4 may occurr in the acidic digestive glands of the sea hare to first yield the cyclized form of dolabellin, and subsquential methanolysis at any stage of the storage or isolation procedure would yield dolabellin (6). Chemical conversion of diet-derived metabolites in the digestive glands of sea hares has been observed previously (e.g. laurinterol into aplysin in Aplysia californica).12 In partial support of this proposal, lyngbyabellin I (5), the methyl ester derivative of lyngbyabellin H (4), was also isolated from this L. ma] uscula crude extract. From NMR data in conjunction with HR FABMS m/z [M + H] 880.2639 (calcd for C38H56C12N3O12S2, 880.2682), the planar structure of 5 appeared to be nearly identical to lyngbyabellin F (2) (Figure IV. 10), except for a residue replacement of Dhiv by Hiva as was the case for the structure of 4. The absolute configurations of 4 at C-14, 20, 26 and 27 were determined as described above for Iyngbyabellin E (I) by chiral HPLC and GC/MS analysis of degradation products, in combination with NMR analysis, which assigned the stereochemistry as 14R, 20S, 26R, and 27S. The absolute configurations at C-2 and C-3 of 4 were determined as 2S, 3S by comparison of coupling constants and chemical shifts with that of compound I. Due to the limited amount of compound 5, the absolute stereochemistry was not established by chemical methods; however, we propose that it belongs to the same enantiomeric series as these other co-occurring metabolites. 8.5 8.0 170 160 7.5 7.0 150 140 6.5 6.0 130 120 5.2 5.0 110 4.5 100 4.0 90 3.5 60 2.5 2.0 3.0 70 60 50 40 ppm 1.0 1.5 30 20 p Figure IV. 9. 1H and 13C of spectra of Iyngbyabellin H (4). 8.5 8.0 7.5 10 10 7.0 6.5 .... 330 6.0 10 5.5 ill 5.0 100 4.5 90 4.0 3.5 . 0 3.0 2.5 60 Figure IV. 10. 1H and 13C of spectra of Iyngbyabellin I (5). 2.0 1.5 1.0 ...... ppm 100 Table IV. 3. NMR Data for Lyngbyabellin H (4) and I (5) in Lyngbyabellin H (4) Position 1 2 3 Lyngbyabellin 1(5) 8H 174.2 43.5 74.9 4 5 6 7 8 31.1 9 10 15.1 20.8 49.3 90.4 37.6 (J in Hz) 3.02, dq (9.5, 7.4) 5.35, m 1.79,1.75, m 1.75, 1.73, m 2.26, 2.15, m 2.10, s 1.27,d(7.4) 172.3 43.3 75.0 30.4 21.6 49.5 90.5 37.5 13.1 159.8 146.6 128.6 165.5 70.4 8.09, s 6.39, dd (5.4, 7.3) 70.1 15 64.2 4.77, m 68.8 16 17 18 19 160.7 146.3 129.2 11 12 13 14 160.8 147.1 128.6 172.9 14-OH 20 22 23 24 25 26 72.6 27 27-NH 28 29 30 49.3 31 32 33 34 35 36 37 38 8.20, s 168.1 77.3 32.5 19.2 18.5 169.2 36.5 21 40.0 25.0 21.8 23.7 5.52, d (8.1) 2.47, m 0.92, d (6.5) 1.06,d(6.5) 161.9 146.9 127.7 170.3 77.9 33.6 18.9 17.4 3.12, dq (6.0, 7.0) 5.45, m 1.81, m 1.78, m 2.24, m 2.12, s 1.31,d(7.1) 8.06, s 5.29, m 5.82, d (5.3) 4.58, dd (11.4, 2.7) 4.50, dd (11.4, 8.7) 8.12, s 6.02, d (5.7) 2.44, m 0.95, d (6.5) 0.98,d(6.5) 169.1 2.82, dd (16.0, 4.8) 2.74, dd (16.1, 7.8) 5.18, ddd (4.9, 5.9, 7.8) 4.32, m 5.86, d (9.4) 1.30, m 1.62, m 0.88, d (6.4) 0.94, d (6.4) 173.1 38.9 19.4 13.9 170.6 21.2 CDCI3. 2.14, t (7.2) 1.62, m 0.91, t (7.2) 2.05,s 36.9 73.2 49.1 37.9 24.9 21.5 23.9 173.7 39.0 19.4 13.9 170.9 21.3 52.7 2.80, dd (15.2, 5.7) 2.61, dd (15.4, 8.3) 5.21, ddd (3.9, 5.7, 8.3) 4.32, m 5.56, d (9.4) 1.37, m 1.62, m 0.91, d (6.7) 0.96, d (6.7) 2.19, t (7.3) 1.66, m 0.96, t (7.2) 2.11,s 3.95, s 101 The lyngbyabellins were tested for cytotoxicity to NCI-H460 human lung tumor and neuro-2a mouse neuroblastoma cells and had LC values between 0.2 and 4.8 pM (Table 4). Intriguingly, lyngbyabellin E (1) and H (4) appeared to be more active against the H460 cell line with LC50 values of 0.4 pM and 0.2 pM, respectively, compared to LC values of 1.2 and 1.4 pM in the neuro-2a cell line. Lynbyabellin I (5) was the most toxic to neuro-2a cells (LC50 0.7 pM), whereas lyngbyabellin G (3), was the least cytotoxic of all compounds to either cell line. On the basis of this limited screening, it appears that lung tumor cell toxicity is enhanced in the cyclic representatives, and overall potency is increased in those containing an elaborated side chain. Because lygbyabellin A and hectochiorin appeared to be strong actindisrupting agents as previously reported,6' 10 the cytoskeletal-disrupting effects of I were also tested in this work. Lyngbyabellin E (I), at concentrations of 0.01-6.0 pM, disrupted the cellular microfilament network in A-b cells (Figure IV. 11). Additionally, at the higher concentrations tested, many cells contained two nuclei, consistent with the inhibition of cytokinesis that often occurs after disruption of the microfilament network. The effects were specific for microfilaments, as there was no evidence of microtubule loss at these concentrations. Table IV. 4. Cytotoxicity of Compounds 1-5 Compound (pM) 1 2 3 4 5 H460 LC (pM) 0.4 1.0 2.2 0.2 1.0 Neuro-2a LC50 1.2 1.8 4.8 1.4 0.7 Along with lyngbyabellins E (1) 1(5), we also isolated the known compound dolabellin (6), which was originally obtained from sea hare Do/abel/a auricularia. Due to the very minute amount (0.08 mg) of 6 isolated, the identity of dolabellin (6) was established by direct comparison of 1H NMR spectrum and HR-TOFMS with literature data.3 102 Biosynthetically, these lyngbyabellins may derive from an assembly by nonribosomal polypeptide synthetases (NRPS) and polyketide synthases (PKS),1 and thus, the structural variation of the lyngbyabellins is intriguing and might suggest that adenylation biosynthesis. domains with relaxed substrate specificity are involved in their Recently, the gene cluster involved in the biosynthesis of a related compound hectochlorin has been characterized, in which the adenylation domain involved in incorporation of a,3-dihydroxyisovaleric acid has exhibited relaxed substrate specificity.13 The isolation of dolabellin (6) and the related compounds lyngbyabellins A-I (1-5) from the marine cyanobacterium Lyngbya majuscula suggests that sea hares are obtaining and accumulating dolabellin and perhaps some other compounds (eg, dolastatin 10) as a result of their cyanobacterial diet. Figure IV. 11. Effect of lyngbyabellin E (1) on the actin cytoskeleton of A10 cells. After 24 h cells were processed and exposed to the microfilament staining reagent TRITC-phalloidin (visualized as red in the figure) and to the DNA-reactive compound DAPI (visualized as blue). (A) Control cells. (B) Treatment of the cells with lyngbyabellin F (1) at 60 nM, which caused complete loss of the cellular microfilament network and generated binucleated cells. Experimental General Experimental Procedures. Optical rotations were measured on a Perkin-Elmer 141 polarimeter. IR and UV spectra were recorded on Nicolet 510 and Beckman DU64OB spectrophotometers, respectively. NMR spectra were recorded on Bruker Avance DPX 400 MHz and Bruker Avance DPX 300 MHz spectrometers with the solvent CDCI3 used as an internal standard (6H at 7.27, at 77.2). High 103 resolution mass spectra were recorded on a Kratos MS-50 TO mass spectrometer. For chiral GC-MS, analysis was accomplished on a Hewlett-Packard gas chromatograph 5890 Series II with a Hewlett-Packard 5971 mass selective detector using an Alltech capillary column (CHIRASIL-VAL phase 25 m x 0.25 mm). HPLC was performed using Waters 515 HPLC pumps and a Waters 996 photodiode array detector. Asymmetric dihydroxylation reagents, AD-mix-a and AD-mix-13, were purchased from Aldrich. Collection. The marine cyanobacterium Lyngbya majuscula (voucher specimen available from WHG as collection number PNG5-27-02-1) was collected by hand from shallow waters (1-3 m) in Alotau Bay, Papua New Guinea, on May 7, 2002. The material was stored in 2-propanol at -20°C until extraction. Extraction and Isolation. Approximately 150 g (dry wt) of the alga were extracted repeatedly with CH2Cl2/MeOH (2:1)to produce 3.30g of crude organic extract. The extract (3.2 g) was fractionated by silica gel vacuum liquid chromatography using a stepwise gradient solvent system of increasing polarity starting from 10% EtOAc in hexanes to 100% MeOH. The fraction eluting with 100% MeOH was found to be active at I ppm in the brine shrimp toxicity assay. This fraction was further chromatographed on Mega Bond RP18 solid-phase extraction (SPE) cartridges using a stepwise gradient solvent system of decreasing polarity starting from 80% aqueous MeOH to 100% MeOH. The most active fractions after SPE (85% toxicity at I ppm to brine shrimp) were then purified by HPLC [Phenomenex Sphereclone 5 u ODS (250 x 10 mm), 9:1 MeOH/H20, detection at 211 nm] giving compounds 1 (7.0 mg), 2 (0.7 mg), 3 (0.7 mg), 4 (0.7 mg), 5 (0.5 mg), and 6 (0.08 mg). Lyngbyabellin E (1): colorless amorphous solid; [ctJ26D -31 ° (c 0.70, MeOH); UV (MeOH) A 240 nm (log c 4.18); IR (neat) 3365, 2926, 2875, 1739, 1644, 1454, 1243, 1030 cm1; 1H and 13C NMR data, see Table 1; HR FABMS mlz EM + H] 864.2373 (calcd for C37H52C12N3012S2, 864.2370). Lyngbyabellin F (2): colorless amorphous solid; [a]260 -6.5° (c 0.20, MeOH); UV (MeOH) 1232, 1098 Amax 238 nm (log 4.0); IR (neat) 3365, 2928, 2873, 1731, 1650, 1455, 104 cm1; 1H and 13C NMR data, see Table 2; HR FABMS mlz [M + H]4 896.2633 (calcd for C38H56C12N3O13S2, 896.2631). Lyngbyabellin 0 (3): colorless amorphous solid; UV (MeOH) Amax [a]26D -26° (c 0.20, MeOH); 238 nm (log g 3.75); IR (neat) 3398, 2925, 2853, 1732, 1484, 1233, 1097 cm1; 1H and 13C NMR data, see Table 2; HR FABMS m/z [M + H]4 595.0733 (calcd for C23H29C12N2O8S2, 595.0742). Lyngbyabellin H (4): colorless amorphous solid; UV (MeOH) Amax [a]260 53° (c 0.08, MeOH); 242 nm (log c4.15); IR (neat) 3305, 2959, 2852, 1735, 1649, 1470, 1238, 1073 cm1; 1H and 13C NMR data, see Table 3; HR FABMS m/z [M + H] 848.2458 (calcd for C37H52C12N3O11S2, 848.2420). Lyngbyabellin 1(5): colorless amorphous solid; UV (MeOH) Amax [a]26D -25° (C 0.04, MeOH); 238 nm (log 4.28); IR (neat) 3309, 2960, 2852, 1740, 1650, 1461, 1233, 1096 cm1; 1H and 13C NMR data, see Table 3; HR FABMS m/z [M + H] 880.2639 (calcd for C36HCl2N3O12S2, 880.2682). Absolute stereochemistry of Lyngbyabellin E (1). A stream of ozone was bubbled through 0.8 mg of I dissolved in 2 mL of CH2Cl2 at -78 °C until the solution turned pale blue (Ca. I mm). The solvent was removed under a stream of N2, and 0.3 mg of the sample was hydrolyzed with 2N NaOH at 90 °C for 7 h. This was analyzed by chiral HPLC [column Phenomenex Chirex 3126 (D), 4.6 x 50 mm; 0.8 mI/mm; detection at 254 nm]. The retention times (mm) of the commercially available Dglyceric and L-glyceric acid were 8.2 and 6.2 mm, respectively, with a solvent system of 0.5 mM CuSO4 in MeCN/H20 (5:95). The base hydrolyzate was found to contain D- glyceric acid (8.2 mm), which was confirmed by coinjection of the appropriate standard. The presence of S-a,3-dihydroxyisovaIeric acid in I was determined by chiral GC-MS analysis. Standard R- and S-Dhiv methyl esters were synthesized by asymmetric dihydroxylation of methyl 3,3-dimethylacrylate using AD-mix-a and ADmix-13, respectively.9'11 Capillary GC-MS analysis was conducted using a Chirasil-Val column (Altech, 25 m x 0.25 mm) using the following conditions: column temperature was set at 70 °C for 3 mm, and was increased from 70 °C to 100 °C (3 °C/min), then 100 °C to 200 °C (15 °C/min). The retention time of the methylated Dhiv residue derived from ozonolysis and saponification of 1, followed by methylation (CH2N2) 105 matched that of the methylated S-Dhiv standard (25.0 mm) but not the methylated RDhiv standard (24.7 mm). 0.5 mg of ozonized sample of I was hydrolyzed at 118 °C for 24 h in 6N HCI. The acid was removed under a stream of N2 and the residue ozonized for 30 mm in 2 mL of methanol at -78°C. After removal of the solvent, the residue was dissolved in 2:1 98% formic acid and 30% H202, and left to stir overnight before refluxing for I h at 100 °C. The solvent was removed and the sample was analyzed by chiral HPLC [column Phenomenex Chirex 3126 (D), 4.6 x 250 mm; 0.8 mI/mm; detection at 254 nm]. The retention time (mm, % CH3CN:2 mM CuSO4) of the standards, were L-Leu (15.2, 15%), D-Leu (16.5, 15%), while the hydrolysate showed a peak for the former at 15.2 mm. Absolute stereochemistry of Lyngbyabellin H (4). 0.4 mg of 4 was ozonized as described above for the lyngbyabellin E (1). Portion of the sample was hydrolyzed with 2N NaOH, and followed by chiral HPLC analysis to confirm assignments of D-glyceric acid (8.2 mi, L-Hiva (9.2 mm). The rest of sample was determined by acid hydrolysis, subsequent ozonolysis, and oxidative workup to give L- leucine (15.2 mm) from chiral HPLC analysis. Biological Activity. Brine shrimp (Artemia sauna) toxicity was measured as previously described.14 After a 24 h hatching period, aliquots of a 10 mg/mL stock solution of compounds 1-5 were added to test wells containing 5 mL of artificial seawater and brine shrimp to achieve a range of final concentrations from 0.1 to 100 ppm. After 24 h the live and dead shrimp were tallied. Cytotoxicity was measured in NCI-H460 human lung tumor cells and neuro-2a mouse neurablastoma cells using the method of Alley et. a115 with cell viability being determined by MIT reduction.16 Cells were seeded in 96-well plates at 6000 cells/well in 180 pL medium. Twenty-four hours later, the test chemicals were dissolved in DMSO and diluted into medium without fetal bovine serum and then added at 20 p1./well DMSO was less than 0.5% of the final concentration. After 48 hr, the medium was removed and cell viability determined. Microfilament Disrupting Assay. Lyngbyabellmn E (I) was tested for microfilament-disrupting activity using rhodamine-phalloidin. A-b cells (rat aortic smooth muscle cell line) were grown on glass coverslips in Basal Medium Eagle 106 (BME) containing 10% fetal calf serum. The cells were incubated with the test compound for 24 h and then fixed with 3% paraformaldehyde for 20 mm, permeabilized with 0.2% Triton X-100 for 2 mm, and chemically reduced with sodium borohydride (1 mg/mL in PBS) three times for 5 mm each. Following a 45 mm incubation with 100 nM TRITC-phalloidin in phosphate buffered saline (to visualize the actin cytoskeleton), the coverslips were washed, stained with 4,6-diamidino-2phenylindole (DAPI) to visualize DNA, mounted on microscope slides, and examined and photographed using a Nikon E800 Eclipse fluorescence microscope with a Photometrics Cool Snap FX3 camera. The images were colorized and overlayed using Metamorph® software. REFERENCES (1) Gerwick, W. H.; Tan, L. T.; Sitachitta, N. In The Alkaloids; Cordell, G. A., Ed.; Academic Press: San Diego, 2001; Vol. 57, pp 75-1 84. (2) Burja, A. M.; Hill, R. T. Hydrobiologia 2001, 461, 41-47. (3) Sone, H.; Kondo, T.; Kiryu, M.; Ishiwata, H.; Ojika, M.; Yamada, K. J. Org. Chem. 1995, 60,4774-4781. (4) Pettit, G. R.; Kamano, Y.; Herald, C. L.; Tuinman, A. A.; Boettner, F. E.; Kizu, H.; Schmidt, J. M.; Baczynkyji, L.; Tomer, K. B.; Bontems, R. J. J. Am. Chem. Soc. 1987, 109, 7581-2. (5) Harrigan, G. G.; Luesch, H.; Yoshida, W.Y.; Moore, R. E.; Nagle, D. G.; Paul, V. J.; Mooberry, S. L.; Corbett, T. H.; Valeriote, F. A. J. Nat. Prod. 1998, 61, 1075-1077. (6) Luesch, Hendrik; Yoshida, W. V.; Moore, R. E.; Paul, V. J.; Mooberry, S. L. J. Nat. Prod. 2000, 63, 611-615. (7) a. Luesch, H.; Yoshida, W. Y.; Moore, R. E.; Paul, V. J. J. Nat. Prod. 2000, 63, 1437-1439; b. Milligan, K. E.; Marquez, B. L.; Williamson, R. 1.; Gerwick, W. H. J. Nat. Prod. 2000, 63, 1440-1443. (8) Luesch, H.; Yoshida, W. Y.; Moore, R. E.; Paul, V. J. Tetrahedron, 2002, 58, 7959-7966. 107 (9) William, P. G.; Luesch, H.; Yoshida, W. Y.; Moore, R. E.; Paul, V. J. J. Nat. Prod. 2003, 66, 595-598. (10) Marquez, B. L.; Watts, K. S.; Yokochi, A.; Roberts, M. A.; VerdierPinard, P.; Jimenez, J. I.; Hamel, E.; Scheuer, P. J.; Gerwick, W. H. J. Nat. Prod. 2002, 65, 866-871. (11) The standards (R)-(-)-a,13-dihydroxyisovaleric acid and (S)-(+)-aj3- dihydroxyisovaleric acid were synthesized by asymmetric dihydroxylation of methyl 3,3-dimethylacrylate (analogous to: Sharpless, K. B.; Amberg, W.; Bennani, Y. L.; Crispino, G. A.; Hartung, J.; Jeong, K. S.; Kwong, H. L.; Morikawa, K.; Wang, Z. M.; Xu, D.; Zhang, X. L. J. Org. Chem. 1992, 57, 2768- 2771) using AD-mix-a and AD-mix-f3, respectively. (12) Stallard, M. 0.; Faulkner, D. J. Comp. Biochem. Physiol, Part B: Biochemistry & Molecular Biology 1974, 49, 37-41. (13) Ramaswamy, A. V.; Gerwick, W. H. (manuscript submitted) (14) Meyer, B. N.; Ferrigni, N. R.; Putnam, L. B.; Jacobsen, L. B.; Nichols, D. E.; McLaughlin, J. L. Planta Med. 1982, 45, 31-33. (15) Alley, M. C., Scudiero, D. A. CancerRes. 1988, 48, 589-601. (16) Manger, R. L.; Leja, L. S. J. AOAC. mt. 1995, 78, 521-527. CHAPTER FIVE ISOLATION, STRUCTURE, AND BIOLOGICAL ACTIVITY OF AURILIDES B AND C FROM A PAPUA NEW GUINEA COLLECTION OF THE MARINE CYANOBACTERIUM LYNGBYA MAJUSCULA ABSTRACT The bioassay-guided fractionation of the cytotoxic constituents of the marine cyanobacterium Lyngbya majuscula collected from Papua New Guinea led to the isolation of aurilides B (1) and C (2). The planar structures of I and 2 were established by spectroscopic analysis, including HR-FABMS, ID 1H and 13C NMR, as well as 2D COSY, HSQC, HSQC-TOCSY, and HMBC spectra. The absolute stereochemistry was determined by spectroscopic analysis and chiral HPLC analysis of acid hydrolysates of I and 2. Both aurilide B and C showed in vitro cytotoxicity toward NCI-H460 human lung tumor and the neuro-2a mouse neuroblastoma cell lines, with LCvalues between 0.01 and 0.13 pM. Aurilide B (I) was evaluated in the NCI 60 cell line panel and found to exhibit a high level of growth inhibition in leukemia, renal, and prostate cancer cell lines with a GI less than 10 nM. Aurilide B (I) showed net tumor cell killing activity in the NCI's hollow fiber assay, an model for assessing a chemical's anticancer activity. in vivo 109 INTRODUCTION Cyanobacteria are phenomenal producers of structurally intriguing and biologically active secondary metabolites including such important molecules as the curacins1 and cryptophycins.2 In our ongoing program to explore these organisms as sources of novel anticancer leads, we recently discovered dolabellin3'4 and lyngbyabellins E-14 from a Papua New Guinea collection of the marine cyanobacterium Lyngbya ma] uscula Gomont (Oscillatoriaceae). In addition, we have identified two new cytotoxins from this collection, aurilides B (1) and C (2), which are closely related to aurilide (3),5 originally isolated from the sea hare Do/abe/Ia auricularia. Herein, this paper describes the isolation, structure determination, and biological activities of aurilides B and C. JL NH 0 NH OH Aurilide B (1) Aurilide C (2) O' Aurilide (3) 110 RESULTS AND DISCUSSION Collections of a shallow water (1-3 m) strain of L. majuscula were made in Alotau Bay, Papua New Guinea, 2002. The alga was extracted with CH2Cl2/MeOH (2:1) and fractionated by silica gel vacuum liquid chromatography. Preliminary bioassay of the relatively polar EtOAc-hexanes eluted fraction showed toxicity in the brine shrimp model (LD - I ppm). Guided by this assay, this fraction was further chromatographed over a Mega Bond RP18 solid-phase extraction (SPE) cartridge and then reversed-phase HPLC to afford two new metabolites, aurilides B (1) and C (2). The molecular formula of aurilide B (1) was established as CH75N5O10 on the basis of HR-FABMS [mlz 834.5603 (M + H) (A -1.1 mmu)J. The 1H NMR data showed the presence of two amide NH groups at ö 7.69 and 6.75, and three Nmethylamide groups at 63.23, 2.88, and 2.63, suggesting the peptidic nature of 1. Detailed analysis of the 2D NMR data enabled us to assign all signals from the proton and carbon NMR spectra (Table V. 1, and Figure V. 1), and revealed a structural framework consisting of peptide and polyketide sections (substructures a and b, respectively). Substructure a was composed of six amino acid residues based on interpretation of 2D NMR data (Figure V. 2-4). Elucidation of three N-methyl amides (N-methylglycine, N-methylalanine, and N-methylisoleucine) was started with HMBC correlations from the N-methyl carbon to the a-proton of each amino acid residue (Figure V. 4), and followed by interpretation of 1H-1H COSY and HSQC-TOCSY NMR spectra to complete construction of each unit within one spin system (Figure V. 5-6). The presence of two valine residues was confirmed by 1H-1H correlations of their amide NH protons at 6 7.69 and 6.75 to the a-protons at 6 5.12 and 4.78, respectively, which in turn showed 1H-1H correlations to the appropriate remaining protons within each unit in the HSQC-TOCSY NMR spectrum (Figure V. 6). A 2hydroxyisoleucic acid residue (Hila) was determined by a 1JCH from H-26 (6 4.90) to a downfield carbon at 78.5 ppm (C-26), while a TOCSY correlation was observed from H-26, via the methine proton H-27, to a methyl group at C-30, and the methylene protons H2-28, which in turn showed correlation to the methyl groups at C-29. The sequence of these residues (two valines, N-methylglycine, N-methyaIanine, N- methylisoleucine, and 2-hydroxyisoleucic acid residue) was deduced from HMBC 111 correlations between H3-4/C-5, NH(1 )/C-1 0, H-1 2/C-I 3, H3-I 9/C-20, and NH(2)/C-25 (Table V. 1) to generate substructure a. Substructure b was elucidated as follows. COSY analysis connected proton signals from the olefinic proton H-33, via the allyllic methylene protons H2-34 and H- 35 oxymethine signal, to the methine proton H-36, which in turn showed correlations to the methyl group at C-43 and the oxymethine proton H-37. A second spin system could be traced from CH3-41 to the allyllic methylene protons at H2-40 and then to the other olefinic proton H-39. HMBC long range correlations between the resonance of H3-44 and those of C-37, C-38 and C-39 revealed that CH3-44 was attached to C-38, and provided the connectivity of the two COSY fragments in substructure b (Figure V. 6). In a similar manner the HMBC correlations observed between H3-42 and C-31, C-32 and C-33 completed substructure b (Table V. 1). The geometry of both the double bonds A3233 and was assigned as E on the basis of the 13C NMR chemical shifts of CH3-42 ( 12.7) and CH3-44 ( 11.3). 7.5 7.0 6.5 6.0 5.5 5.0 4.5 4.0 3.3 Figure V. 1. 1H and 13C spectra of aurilide B (1). 3.0 2.3 2.0 1.3 0:5 112 Table V. 1. NMR Data of Aurilide B (1) and C (2) in C6D6. Position 1 2 3 4 5 6 7 8 9 10 11 öc 170.2 58.9 13.8 36.1 172.1 54.3 31.0 20.1 17.3 169.9 51.8 12 13 14 15 16 17 18 19 36.8 170.0 58.6 33.9 27.4 20 173.1 21 54.7 31.7 22 23 24 25 26 27 28 29 30 4.40, d (18.0) 3.80, d (18.0) 3.23, s 5.24, d (10.0) 2.48, m 1.86, 1.30, m 18.1 26.1 11.8 14.9 14.1 12.7 10.2 11.3 H-6, 8,9 H-9 H-6, 8 NH(1),H-11 H-12 H-li Aurilide C (2) H (J in Hz) 170.2 59.6 14.0 36.8 54.4 32.0 20.4 17.5 170.11 51.9 37.1 170.14 58.7 7.74, t (9.0) 2.19, m 3.97, m 2.07, m 5.18,d(1l.2) H-39,43,44 132.1 H-37, 40,44 H-40, 41,44 134.6 21.4 14.3 12.8 4.78, dd (8.8, 8.8) 1.98, m 0.89, d (6.0) 0.90, d (6.0) 4.90, d (6.1) 2.17, m 1.50, 1.14, m 0.83, t (7.7) 1.03, d (6.0) 5.61, t (7.7) 1.95, 1.92, m 0.89 1.95, s 0.64, d (7.0) 1.54,s 7.69 br, d (9.1) 6.75 br, d (8.8) H-41 H-40 H-33, 34 H-36 3.08, m 1.25, d (7.1) 2.55, s 172.1 H-15, 16 H-16, 18 H-15, 18 H-16, 17 H-14, 15 H-14 H-14, 19, 21 H-23 H-21, 23 H-24 H-23 NH(2), H-26 H-27, 30 H-26, 29, 30 H-29, 30 H-28 H-27 H-26, 33, 42 H-34, 42 H-34, 42 H-35 H-43 H-37, 43 0.85, d (7.0) 2.88, s 20.2 170.3 78.5 37.2 HMBC H-2, 3, 37 H-3, 4 H-2 H-2 H-2, 4, 6 H-8, 9 H-il, 12,14 14.8 30.7 41 42 43 44 NH (1) NH (2) 5.12, dd (9.0, 7.4) 1.97, m 1.15, d (7.0) 1.25, d (7.0) 1.03 40 32 33 34 35 36 37 38 39 3.23, m 1.21, d (7.1) 2.63, s 12.1 169.3 128.0 145.3 30.9 71.0 41.1 82.5 131.4 134.2 21.4 31 Aurilide B (1) H (J in Hz) 34.1 27.6 12.2 15.1 30.6 173.2 54.9 31.0 18.9 20.3 170.3 80.4 30.5 18.7 18.4 169.7 128.3 146.0 30.9 71.2 41.2 82.6 10.1 11.4 5.15, dd (9.0, 5.0) 1.98, m 1.17, d (7.0) 1.28, d (7.0) 4.39. d (18.0) 3.80, d (18.0) 3.22, s 5.26, d (10.0) 2.49, m 1.89, 1.30, m 1.03 0.86, d (7.0) 2.85, s 4.75, dd (8.6, 7.5) 1.95, m 0.88 0.90 4.54, d (7.5) 2.36, m 1.00, d (7.0) 0.88, d (7.0) 7.75, t (9.0) 2.14, m 3.98, m 2.02, m 5.17, d (11.2) 5.62, t (7.7) 1.95, 1.92, m 0.89 1.95, s 0.66, (7.0) 1.54, s H-37,39 7.66 br, d (9.1) 6.70 br, d (8.8) 113 ppal 0 .5 II 1 .0 HI 1. 1 2. a, 2. 3, 3. 4. . $., 4. .1 5. : 5. 6. 6. .7 ..: .., ,. .0 .i. J.0 Figure V. 2. COSY spectrum of aurilide B (1). 2.5 2.0 1.5 1.0 0.5 ppm 114 pp 10 20 30 S. 40 50 0 o S 60 0 80 p S 90 100 110 120 130 0 140 4 150 160 b.0 1.0 l.0 0.3 0.11 3.0 3.11 %.3 1.0 0.3 Figure V. 3. HSQC spectrum of aurilide B (1). 0.0 4.3 4.11 1.3 1.0 0.3 ppm 115 ppm b.0 I. 7.0 6.5 6.0 5.5 5.0 4.5 4.0 3.5 3.0 Figure V. 4. HMBC spectrum of aurilide B (1). 2.5 2.0 1.5 1.0 0.5 ppm 116 p o4 i I ' : à.d' : C 0 I 4 0 II o 0 o .1 II' 'I P1 1 1 1 1 . 1 C 8.0 7.5 7.0 6.5 6.0 5.5 5.0 4.5 I 4.0 3.5 3.0 2.5 Figure V. 5. HSQC-TOCSY spectrum of aurilide B (1). 2.0 1.5 1.0 0.5 ppm 117 substructure a cosy HMBC -i---- NOE 44 42 substructure b Figure V. 6. Key COSY, HMBC, and NOE correlations for 1. Substructures a and b were connected on the basis of HMBC data. The aproton (H-26) of Hila showed a cross-peak to the C-31 carbonyl carbon of substructure b, and H-37 of b correlated with the C-I carbonyl carbon of substructure a to complete a 26-membered ring. Diagnostic NOEs from CH3-43 to H-35 and H-37, as well as the close similarity of 1H and 13C NMR shifts and JH,H values between I and the known compound (3), were indicative of the relative stereochemistry of the polyketide portion of I as 35S*, 36S*, 37S*. To deduce the absolute stereochemistry of C-35 through C-37, S- and R-MTPA esters were introduced to the C-35 hydroxyl group of I, respectively.6 The LA5(SR)values for H-33, 34, 36, 39, 40, 41, 42, 43, and 44 were indicative of a 5S, 6S, 7S configuration (Figure V. 7), which is identical to that of aurilide (3). 118 To assign the absolute configuration of the amino acid residues (substructure a), I was acid hydrolyzed and analyzed by chiral HPLC and chiral GC-MS analysis. The absolute configurations of the three components, Val, Melle, and isoleucic acid, were determined to be L, allo-L, allo-D, respectively. A portion of hydrolysate of I was used to determine the absolute configuration of MeAla by Marfey analysis;7 this residue was shown to be L, thus completing the structure as shown. 0 OMTPA o 002 +008 -0.02 +0.26 -0.03 -0.11 -0.01 Figure V.7. M(SR)values (ppm) of MTPA esters of aurilide B (1) obtained in CD3CN. Aurilide C (2) was isolated by RP-HPLC from the same fraction containing aurilide B (1). High-resolution FABMS analysis of aurilide C (2) revealed an [M+H] ion (m/z 820.5449) consistent with a molecular formula of C43H74N5010. Aurilide C had high structural homology to I as evidenced by nearly identical 1H and 13C NMR chemical shifts for most of the molecule (Table V. 1, and Figure V. 8). However, it displayed differences for the isoleucic acid unit of 1. 2D NMR and mass spectroscopy analysis revealed that the isoleucic acid residue has been replaced by a 2-hydroxyisovaleric acid (Hiva). Hydrolysis and stereoanalysis of the peptide portion of 2 were undertaken as described above for aurilide B (I). The absolute configurations of the four components, Val, MeAla, Melle, and Hiva, were determined to be L-, L-, allo-L-, and D-, respectively. We propose that the polyketide portion is of the same configuration as that of I because of their comparable spectroscopic properties. Aurilide B (I) and C (2) were tested for cytotoxicity to NCI-H460 human lung tumor and neuro-2a mouse neuroblastoma cells. Intriguingly, aurilide B was 119 approximately 4-fold more toxic than C to these cell lines. The LC50 for aurilide B was 0.01 pM and 0.04 pM for neuro-2a and H460 cells, respectively, and 0.05 pM and 0.13 pM, respectively, for aurilide C. Aurilide B (1) was evaluated in the NCI 60 cell line panel and found to exhibit a high level of growth inhibition in leukemia, renal, and prostate cancer cell lines with a Gl less than 10 nM. Aurilide B (1) showed net tumor cell killing activity in the NCI's hollow fiber assay, an in vivo model for assessing a chemical's anticancer activity.8 In the microfilament disruption assay compound I caused at 2.5 pm/mL partial loss of the microfilament network when tested against A-I 0 cells (smooth muscle cells). However the compound might possess a multimodal mode of action taking into account its general toxicity observed at much lower concentration. 7.5 7.0 6.5 6.0 5.0 5.5 4.5 4.0 3.5 90 80 3.0 2.5 2.0 60 50 1.5 0.5 ppn 1.0 I...... 170 160 150 140 130 120 110 100 70 Figure V. 8. 1 H and I 3C spectra of aurilide C (2). 40 30 20 pt 120 Biosynthetically, the aurilides likely derive from an assembly by nonribosomal polypeptide synthetases (NRPS) and polyketide synthases (PKS),9 and thus, the structural variation of the aurilides might be due to adenylation domains with relaxed substrate specificities. Recently, another cytotoxic depsipeptid, kulokekahilide-2,1° was isolated from a cephalaspidean mollusk, which is also closely related to aurilide (3) obtained from the sea hare D. auricularia. Observation that some metabolites occur in unrelated genera of marine invertebrates, as well as the isolation of related compounds from microbial sources, provides further evidence that many natural products are actually produced by symbionts rather than their invertebrate hosts.11'12 For example, the isolation here of aurilide B (1) and C (2) from the marine cyanobacterium Lyngbya majuscula suggests that marine invertebrates obtain and accumulate the related metabolites aurilide (3) and kulokekahilide-2 as a result of their cyanobacterial diet. Experimental General Experimental Procedures. Optical rotations were measured on a Perkin-Elmer 141 polarimeter. IR and UV spectra were recorded on Nicolet 510 and Beckman DU64OB spectrophotometers, respectively. NMR spectra were recorded on Bruker Avance DPX 400 MHz and Bruker Avance 300 MHz spectrometers with the solvent C6D6 used as an internal standard (H at 7.16, 6c at 128.4). High resolution mass spectra were recorded on a Kratos MS-50 TC mass spectrometer. Chiral GCMS analysis was accomplished on a Hewlett-Packard gas chromatograph 5890 Series II with a Hewlett-Packard 5971 mass selective detector using an Alltech capillary column (CHIRASIL-VAL phase 25 m x 0.25 mm). HPLC was performed using Waters 515 HPLC pumps and a Waters 996 photodiode array detector. Collection. The marine cyanobacterium Lyngbya majuscula (voucher specimen available from WHG as collection number PNG5-27-02-1) was collected from shallow waters (1-3 m) in Alotau Bay, Papua New Guinea, on May 7, 2002. The material was stored in 2-propanol at -20°C until extraction. 121 Extraction and Isolation. Approximately 150 g (dry wt) of the alga were extracted repeatedly with CH2Cl2/MeOH (2:1) to produce 3.30 g of crude organic extract. The extract (3.2 g) was fractionated by silica gel vacuum liquid chromatography using a stepwise gradient solvent system of increasing polarity starting from 10% EtOAc in hexanes to 100% MeOH. The fraction eluting with 90% EtOAc was found to be active at I ppm in the brine shrimp toxicity assay. This fraction was further chromatographed on Mega Bond RP15 solid-phase extraction (SPE) cartridges using a stepwise gradient solvent system of decreasing polarity starting from 80% MeOH in HO to 100% MeOH. The most active fractions after SPE (85% toxicity at 1 ppm to brine shrimp) were then purified by HPLC [Phenomenex Sphereclone 5 p ODS (250 x 10 mm), 9:1 MeOH/H20, detection at 211 nm] giving compounds 1 (80 mg), and 2 (5 mg). Aurilide B (1): colorless amorphous solid; (MeOH) Amax [cLJ24D -17° (c 0.34, MeOH); UV 222 nm (log 64.65); IR (neat) 3481 (br), 3353 (br), 2964, 1740, 1687, 1646, 1251,1205 cm1; 1H and 13C NMR data, see Table 1; HR FABMS m/z[M + H] 834.5603 (calcd for CH76N5O10, 834.5592). Aurilide C (2): colorless amorphous solid; (MeOH) Amax [a]240 -19° (c 0.39, MeOH); UV 222 nm (log c 4.56); IR (neat) 3487 (br), 3350 (br), 2962, 1740, 1687, 1647, 1251, 1207 cm1; 1H and 13C NMR data, see Table 1; HR FABMS m/z EM + H] 820.5449 (calcd for C43H74N5010, 820.5436). MTPA Esters of 1. Aurilide B (1, 0.5 mg each) was reacted with R- or SMTPACI (10 pL) in 300 pL of CH2Cl2 containing 10 mg of DMAP. The reaction mixtures were partitioned with EtOAcIO.1 M NaHCO3, and the EtOAc layers were washed with 0.1 M HCI and H20. The EtOAc layer was evaporated and then separated by ODS HPLC [Phenomenex Sphereclone 5 pODS (250 x 10mm), 9:1 MeOH/H20, detection at 220 nmj to yield S- and R- MTPA esters. S-MTPA ester: 1H NMR (CD3CN) ö 7.20 (H-33), 2.55 (H-34a), 2.19 (H-34b), 5.83 (H-35), 2.38 (H-36), 5.05 (H-37), 5.51 (H-39), 2.05 (H-40), 0.95 (H-41), 1.58 (H- 42), 0.85 (H-43), 1.61 (H-44); FABMS m/z 1050.7 (M + H). R-MTPA ester: 1H NMR (CD3CN) ö 7.28 (H-33), 2.67 (H-34a), 2.24 (H-34b), 5.80 (H-35), a 122 2.37 (H-36), 5.05 (H-37), 5.49 (H-39), 2.04 (H-40), 0.93 (H-41), 1.83 (H-42), 0.73 (H- 43), 1.60 (H-44); FABMS mlz 1050.7 (M + H). Absolute stereochemistry of the peptide portion of 1. Aurilide B (1, 0.3 mg) was hydrolyzed in 6 N HCI at 110 °C for 24 h, then dried under a stream of N2 and further dried under vacuum. The residue was reconstituted with 300 pL of H20 prior to chiral HPLC analysis. Mobile phase I: 2 mM CuSO4 in MeCN/H20 (5:95), flow rate I mL/min [Phenomenex Chirex 3126 (D), 4.6 x 250 mm; UV 254 nm detector]. Mobile phase I elution times (tR, mm) of authentic standards: allo-L-Melle (16.8), L-Melle (17.4), allo-D-Melle (22.2), D-MeIIe (23.0). Mobile phase II: 2 mM CuSO4 in MeCN/H20 (15:85), flow rate I mL/min [column, Phenomenex chirex 3126 (D), 4.6 x 50 mm; UV 254 nm detector]. Mobile phase II elution times (tR, mm) of authentic standards: allo-L-isoleucic acid (8.2), L- isoleucic acid (11.5), allo-D-isoleucic acid (14.2), D-isoleucic acid (18.2). The hydrolysate was chromatographed alone and co-injected with standards to confirm assignments of allo-L-Melle, allo-D-isoleucic acid. The presence of L-Val (2 eq) was confirmed by chiral GCIMS using established methods.13 The configuration of MeAla was determined by Marfey's analysis.7 A portion of the hydrolysate was evaporated to dryness and resuspended in H20 (100 p.L). A 0.1% 1-fluoro-2,4-dinitrophenyl-5-L-alaninamide solution in acetone (L-Marfey's reagent, 20 p.L) and IN NaHCO3(10 1tL) were added to a portion of the hydrolysate, and the mixtures were heated at 40°C for I h. The solutions were cooled to room temperature, neutralized with 2N HCI (5 j.tL) and evaporated to dryness. The residues were resuspended in H20 (50 jiL) and analyzed by reversed-phase HPLC [Microsob-MV C18, 4.6 x 250 mm, UV detection at 340 nm] using a linear gradient of 9:1 50mM triethylamine phosphate (TEAP) buffer (pH 3.1)/CH3CN to 1:1 TEAP/CH3CN over 60 mm. The derivatized MeAla residue in the hydrolysate of I eluted at the same retention time as the derivatized standard L-MeAla (12.0 mm) but not that of D-MeAIa (13.7 mm). The absolute stereochemistry of aurilide C (2) was analyzed as described above for the aurilide B (1) hydrolysate. Allo-L-Melle (16.8 mm), and D-Hiva (9.2 mm) were assigned to aurilide C (2) by chiral HPLC; L-MeAla (12.0 mm) was determined 123 based on Marfey's analysis. The presence of L-Val (2 eq) was confirmed by chiral GC/MS using established methods.13 Biological Activity. Brine shrimp (Artemia sauna) toxicity was measured as previously described.14 After a 24 h hatching period, aliquots of a 10 mg/mL stock solution of compounds A-D were added to test wells containing 5 mL of artificial seawater and brine shrimp to achieve a range of final concentrations from 0.1 to 100 ppm. After 24 h the live and dead shrimp were tallied. Cytotoxicity was measured in NCI-H460 human lung tumor cells and neuro-2a mouse neurablastoma cells using the method of Alley et al,15 with cell viability being determined by MU reduction.16 Cells were seeded in 96-well plates at 6000 cells/well in 180 p1 of medium. Twenty-four hours later, the test chemicals were dissolved in DMSO and diluted into medium without fetal bovine serum and then added at 20 pg/well DMSO was less than 0.5% of the final concentration. After 48 hr, the medium was removed and cell viability determined. REFERENCES (1) Gerwick, William H.; Proteau, Philip J.; Nagle, Dale G.; Hamel, Ernest; Blokhin, Andrei; Slate, Doris L. J. Org. Chem. 1994, 59, 1243-5. (2) Trimurtulu, Golakoti; Ohtani, Ikuko; Patterson, Gregory M. L.; Moore, Richard E.; Corbett, Thomas H.; Valeriote, Frederick A.; Demchik, Lisa. J. Am. Chem. Soc. 1994, 116, 4729-37. (3) Sone, H.; Kondo, T.; Kiryu, M.; Ishiwata, H.; Ojika, M.; Yamada, K. J. Org. Chem. 1995, 60, 4774-4781. (4) Han, B.; McPhail, K. L; Gross, H.; Goeger, D.; Moobery, S. L.; Gerwick, W. H. (manuscript in preparation). (5) Suenaga, K.; Mutou, T.; Shibata, T.; Itoh, 1.; Fujita, 1; Takada, N.; Hayamizu, K.; Takagi, M.; Irifune, T.; Kigoshi, H.; Yamada, K. Tetrahedron 2004, 60, 8509-8527. 124 (6) Ohtani, Ikuko; Kusumi, Takenori; Kashman, Yoel; Kakisawa, Hiroshi. J. Am. Chem. Soc. 1991, 113, 4092-6. (7) Marfey, Peter. Carlsberg Res. Commun. 1984, 49, 591-6. (8) Plowman, J.; Dykes, D. J.; Hollingshead, M.; Simpson-Herren, L; Alley, M. C. In Anticancer Drug Development Guide: Preclinical Screening, Clinical Trials, and Approval. B. Teicher, Ed.; Humana Press, Inc. Totowa, NJ. 1997, pp 101-125. (9) Gerwick, W. H.; Tan, L. T.; Sitachitta, N. In The Alkaloids; Cordell, G. A., Ed.; Academic Press: San Diego, 2001; Vol. 57, pp 75-1 84. (10) Nakao, Yoichi; Yoshida, Wesley Y.; Takada, Yuuki; Kimura, Junji; Yang, Liu; Mooberry, Susan L.; Scheuer, Paul J. J. Nat. Prod. 2004, 67, 13321340. (11) Haygood, M. G.; Schmidt., E. W.; Davidson, S. K.; Faulkner, D. J. J. Mo!. Microbiol. Biotechno!. 1999, 1, 33-43. (12) Osinga, R.; Armstrong, E.; Burgess, J. G.; Hoffmann, F.; Reitner, J.; Schumann-Kindel, G. Hydrobiologia 2001, 461, 55-62. (13) Trimurtulu, G.; Ohtani, I.; Patterson, G. M. L.; Moore, R. E.; Corbett, T. H.; Valeriote, F. A.; Demchik, L. J. Am. Chem. Soc. 1994, 116, 4729-37. (14) Meyer, B. N.; Ferrigni, N. R.; Putnam, L. B.; Jacobsen, L. B.; Nichols, D. E.; McLaughlin, J. L. Planta Med. 1982, 45, 31-3. (15) Alley, M. C.; Scudiero, D. A. CancerRes. 1988, 48, 589-601. (16) Manger, R. L.; Leja, L. S. J. AOAC. !nt. 1995, 78, 521-527. 125 CHAPTER SIX CONCLUSIONS As is quite evident from the findings reported in the thesis, marine cyanobacteria are extraordinary prolific in their production of elaborate and complex bioactive secondary metabolites. This investigation provided a good representation of the diversity of organic molecules produced by marine cyanobacteria of the genus Lyngbya. A total of eighteen new compounds were identified from the organic extracts of two strains of Lyngbya from Papua New Guinea. The isolation and purification of these molecules was achieved by different chromatographic techniques, including VLC and HPLC, while their structures were established by extensive ID and 2D NMR experiments and mass spectrometry. All of the secondary metabolites found in this work are nitrogen-containing compounds with molecular sizes ranging from 325 MW (semiplenamide C) to 1008 MW (wewakpeptin D). Preliminary bioassays of the crude organic extract of the marine cyanobacterium Lyngbya semip!ena collected from Papua New Guinea in 1997 showed good activity in the brine shrimp toxicity model at 10 ppm. Guided by this assay, seven new anandamide-like fatty acid amides, semiplenamides A to G, together with four cyclic depsipeptides, wewakpeptins A to D, were identified. Due to the structural resemblance of the novel ethanolamide derivatives (semiplenamide AG) with anandamide, (N-arachidonoyl-ethanolamine), an endogenous agonist of cannabinoid receptors, semiplenamide A-G were tested on the well characterized proteins of the endocannabinoid system: 1) the "central" cannabinoid CB1 receptors; 2) the anandamide membrane transporter (AMT), which is responsible for anandamide cellular uptake; and 3) the fatty acid amide hydrolase (FAAH), which catalyses anandamide hydrolysis. Three showed modest potency in displacing radiolabeled anandamide from the cannabinoid receptor (CBI), and one was a modest inhibitor of the anandamide membrane transporter (AMT). The wewakpeptins were tested for cytotoxicity to NCI-H460 human lung tumor and neuro-2a mouse neuroblastoma cells. Intriguingly, wewakpeptin A and B were approximately 10-fold more toxic than C and D to these cell lines. 126 Lyngbya majuscula has been recognized as a chemically and biologically rich strain. Five new lyngbyabellin analogs, lyngbyabellins E-I, along with the known compound dolabellin, originally isolated from sea hare Do/abel/a auricularia, were identified from a 2002 Papua New Guinea collection of the marine cyanobacterium Lyngbya majuscula. The lyngbyabellins were tested for cytotoxicity to NCI-H460 human lung tumor and neuro-2a mouse neuroblastoma cells and had LC50 values between 0.2 and 4.8 pM. Intriguingly, Iyngbyabellin E and H appeared to be more active against the H460 cell line with LC values of 0.4 pM and 0.2 pM, respectively, compared to LC values of 1.2 and 1.4 pM in the neuro-2a cell line. Lynbyabellin I was the most toxic to neuro-2a cells (LC 0.7 pM), whereas lyngbyabellin G, was the least cytotoxic of all compounds to either cell line. On the basis of this limited screening, it appears that lung tumor cell toxicity is enhanced in the cyclic representatives, and overall potency is increased in those containing an elaborated side chain. Additionally, two new cytotoxins, aurilides B and C, which are closely related to aurilide, originally isolated from the sea hare Dolabella auricularia, have been identified from the same extract where the lyngbyabellins E-1 were isolated. Aurilides B and C were tested for cytotoxicity to NCI-H460 human lung tumor and neuro-2a mouse neuroblastoma cells. Interestingly, aurilide B was approximately 4-fold more toxic than C to these cell lines. The LC for Aurilide B was 0.01 pM and 0.04 pM for neuro-2a and H460 cells, respectively, and 0.05 pM and 0.13 pM, respectively, for aurilide C. Aurilide B (1) was evaluated in the NCI 60 cell line panel and found to exhibit a high level of growth inhibition in leukemia, renal, and prostate cancer cell lines with a Gl50 less than 10 nM. Aurilide B (1) showed net tumor cell killing activity in the NCI's hollow fiber assay, an in vivo model for assessing a chemical's anticancer activity. The principle biogenetic theme in the natural products chemistry of marine cyanobacteria is the integration of nonribosomal polypeptide synthetase (NRPS) and polyketide synthesase (PKS) pathways, in a variety of configurations, so as to produce a great structural diversity. Study of the biosynthetic pathways employing traditional isotope-labeled precursor incorporation studies will multiply as knowledge of the culture requirements of these organisms increases. In turn, knowledge of 127 these unique structures and their biosynthetic pathways will intensify studies at the molecular genetic level. At present, the mechanisms and pathways that are used to regulate secondary metabolite expression in these creatures are wholly unknown, and breakthroughs in this area could provide the proverbial "golden key" for unlocking the full biosynthetic potential of these organisms. Given the prevalent antitubulin/antiactin theme in the mechanism of action of cyanobacterial metabolites, the discovery of new compounds of utility to the chemotherapy of neoplastic diseases can be anticipated with considerable certainty. However, similar to the natural products of other taxonomic groups, the biochemical mechanisms of action of many marine cyanobacterial compounds are largely unknown, and their continued study will provide a rich collection of tools for molecular pharmacologists. It is likely that new biochemical targets for toxins and other bioactive compounds will be revealed in the mechanism of action studies. Additional insight into the mechanism of action of marine cyanobacterial toxins will also come from continued studies of the chemical ecology of these organisms. Debate also continues on the true source of these bioactive marine natural products. The accumulated experimental and circumstantial evidence is in favor of the hypothesis that numerous nonribosomal peptides from marine invertebrates are actually produced by symbiotic microorganisms. For instance, both dolabellin and aurilide were originally isolated from the sea hare Dolabella auricularia with isolated yields of I O%. However, we have isolated both dolabellin and two aurilides from a single collection of marine cyanobacteria Lyngbya majuscula from Papua New Guinea. Moreover, the yield of aurilide B is over I O% of the wet weight of the sample. Isolation and cultivation of the suspected microbial producers of bioactive natural products either from the surrounding sea water or from the tissue of invertebrates through careful design of special media could provide a much needed answer to the pressing supply problem that is currently presented by many pharmacologically interesting compounds are hampered from further development as a drug due to limited supply problems. If these microorganisms are indeed the producers of bioactive metabolites of interest, we believe the investigation of the chemical ecology associated with these natural product-producing microbes is 128 essential, and represents the marine biologist's 'why' to go along with the chemist's 'what' and the bioprocess engineer's 'where' and 'how'. Novel compounds are discovered and reported on a daily basis. The marine environment is highly complex, however, and thus a highly interdisciplinary approach is required in order to realize its potential. As one of the two remaining environments (marine and extraterrestrial) which is yet to be fully explored, a greater understanding could be achieved by a close collaboration between marine biologists, microbiologists, geneticists, bioprocess engineers and natural product chemists. 129 BIBLIOGRAPHY Aimi N, Odaka H, Sakai S, Fujiki H, Suganuma M, Moore RE and Patterson GM (1990) Lyngbyatoxins B and C, two new irritants from Lyngbya majuscula. J Nat Prod 53:1593-1596. Amador ML, Jimeno J, Paz-Ares L, Cortes-Funes H and Hidago M (2003) Ann Onco! 14:1607-1615. Ang KKH, Holmes MJ, Higa T, Hamann MT and Kara UAK (2000) Antimicrobial Agents Chemothera. 44:1645-1649. Baden DG, Bikhazi G, Decker SJ, Foldes FF and Leung 1(1984) Neuromuscular blocking action of two brevetoxins from the Florida red tide organism Ptychodiscus brevis. Toxicon 22:75-84. Bai R, Friedman SJ, Pettit GR and Hamel E (1992) Biochem. Pharmacol. 43:2637-45. Bai R, Pettit GR and Hamel E (1990a) Biochem. Pharmaco!. 39:1941-9. Bai R, Pettit GR and Hamel E (1990b) J. Biol. Chem. 265:17141-9. Barrows LR, Radisky DC, Copp BR, Swaffar OS, Kramer RA, Warters RL and Ireland CM (1993) Anti-Cancer Drug Design 8:333-347. Basu A, Kozikowski AP and Lazo JS (1992) Structural requirements of lyngbyatoxin A for activation and downregulation of protein kinase C. Biochemistry 31:3824-3830. Bergman W and Feeney RJ (1951) J. Org. Chem. 16:981-987. Berman FW, Gerwick WH and Murray IF (1999) Toxicon 37:1645-1648. Blokhin AV, Yoo HO, Geralds RS, Nagle DG, Gerwick WH and Hamel E (1995) Mo! Pharmacol 48:523-531. Bodey GP, Freirich EJ, Monto RW and Hewlett JS (1969) Cancer Chemother. 53:5966. Buchanan RAand Hess F (1980) Science 127:143-171. Burja AM and Hill RT (2001) Hydrobio!ogica 461:41-47. CardeHina JH, 2nd, Marner FJ and Moore RE (1979) Seaweed dermatitis: structure of lyngbyatoxin A. Science 204:193-195. Carter DC, Moore RE, Mynderse JS, Niemczura WP and Todd JS (1984) J. Org. Chem. 49:236-241. 130 Chang Z, Flatt P, Gerwick WH, Nguyen VA, Willis CL and Sherman DH (2002) The barbamide biosynthetic gene cluster: a novel marine cyanobacterial system of mixed polyketide synthase (PKS)-non-ribosomal peptide synthetase (NRPS) origin involving an unusual trichloroleucyl starter unit. Gene 296:235-247. Chen J and Forsyth CJ (2004) Proc. Nat!. Acad. Sd. USA 101:12067-12072. Clark WD and Crews P (1995) Tet. Lett. 36:1185-1188. Crews P, Manes LV and Boehler M (1986) Tetrahedron Letters 27:2797-2800. Cruz-Monserrate Z, Mullaney JT, Harran PG, Pettit GR, Mullaney JT, Harran PG and Pettit GR (2003) Eur. J. Biochem. 270:3822-28. De Clercq E (2000) Res. Rev. 20:323-349. de Fiebre CM, Meyer EM, Henry JC, Muraskin SI, Kern WR and Papke RL (1995) Mo!. Pharmacol. 47:164-171. de Silva ED and Scheuer P (1980) Tet. Left. 21:1611-14. Deems RA, Lombardo 0, Morgan DP, Mihelich ED and Dennis EA (1987) Biochim. Biophys. Acta. 917:258-68. Degnan BM, Hawkins CJ, Lavin MF, McCaffrey EJ, Parry DL, van den Brenk AL and Watters DJ (1989) J. Med. Chem. 32:1349-1 354. Dileo P CP, Bacci G, et al. (2002) Proceedings of the 2002 ASCO Annual Meeting. Abstr #1628. Dumdei EJ, Simpson JS, Garson MJ, Bryriel KA and Kennard CHL (1997) Australian Journal of Chemistiy 50:139-144. Edwards DJ and Gerwick WH (2004) JAm Chem Soc 126:11432-11433. Edwards DJ, Marquez BL, Nogle LM, McPhail K, Goeger DE, Roberts MA and Gerwick WH (2004) Structure and biosynthesis of the jarnaicamides, new mixed polyketide-peptide neurotoxins from the marine cyanobacterium Lyngbya majuscula. Chem Biol 11:817-833. El Sayed KA, Kelly M, Kara UAK, Ang KKH, Katsuyarna I, Dunbar DC, Khan AA and Hamann MT (2001) J. Am. Chem. Soc. 123:1804-1808. Fu X, Do T, Schmitz FJ, Andrusevich V and Engel MH (1998) J. Nat. Prod. 61:15471551. Fujiki H, Mon M, Nakayasu M, Terada M, Sugimura I and Moore RE (1981) proc. Nat!. Acad. Sd. USA 78:3872-3876. 131 Gerwick WH, Proteau PJ, Nagle DG, Hamel E, Blokhin AV and Slate DL (1994) J. org. Chem. 59:1243-1245. Halstead BW (1965) Posinous and Venomous Marine Animals of the World. US Government Printing Office, Washington DC. 1:1-155. Hamel E (2002) Biopolymers 66:142-160. Hamel E, Blokhin AV, Nagle DG, Yoo HD and Gerwick WH (1995) Drug Delivery Research 34:110-120. Harrigan GG, Goetz GH, Luesch H, Yang S and Likos J (2001) Journal of Natural Products 64:1133-1138. Harrigan GG, Luesch H, Yoshida WY, Moore RE, Nagle DG, Paul VJ, Mooberry SL, Corbett TH and Valeriote FA (1998) Journal of Natural Products 61:1075-1077. Haygood MG, Salomon CE and Faulkner DJ (2003) Ecteinascidin Family Compounds: Compositions and Methods. International Patent EPI 360337. Haygood MG, Schmidt EW, Davidson SK and Faulkner DJ (1999) J. Mol. Microbiol. Biotechnol. 1:33-43. Hentschel U, Hopke J, Horn M, Friedrich AB, Wagner M, Hacker J and Moore BS (2002) Applied and Environmental Microbiology 68:4431-4440. Hill RT, Hamann MI, Peraud 0 and Kasanah N (2003) Ref Type: Patent:41 15-4180 PCT. In Y, Doi M, lnoue M, Ishida T, Hamada Y and Shioiri T (1994) Acta Ciystallogr. C. 50:432-434. Ishibashi M, Iwasaki T, lmai S, Sakamoto S, Yamaguchi K, Ito A, Iwasaki T, Imai S, Sakamoto S, Yamaguchi K and Ito A (2001) J. Nat. Prod. 64:108-110. Jacobs RS, Culver P, Langdon R, O'Brien T and White S (1985) Tetrahedron 41:9814. Jaspars M (1998) Advances in Drug Discovery Techniques;. Haniey, A. L., Ed.; Wiley: New York,:65-84. Jiménez JI and Scheuer PJ (2001) J. Nat. Prod. 64:200-203. Kelecom A (1999) Chemistry of Marine Natural Products: Yesterday, Today and Tomorrow. An Acad Bras Cienc 71:249-263. Kem WR, Abbott BC and Coates RM (1971) Toxicon 9:15-22. Kobayashi M and Kitagawa K (1999) J Nat Toxins 8:249-258. 132 Kozikowski AP, Shum PW, Basu A and Lazo JS (1991) Synthesis of structural analogues of lyngbyatoxin A and their evaluation as activators of protein kinase C. Journal of medicinal chemistiy 34:2420-2430. Kraft AS (1993) J. Nat!. Cancer Inst. 85:1790-1792. Kraft AS, Woodley S, Pettit GR, Gao F, Coil JC and Wagner F (1996) Cancer Chemother. Pharmacol. 37:271-278. Li WI, Berman FW, Okino T, Yokokawa F, Shioiri T, Gerwick WH and Murray TF (2001) Proc. Nat!. Acad. Sci. USA 98:7599-7604. Li WI, Marquez BL, Okino T, Yokokawa F, Shioiri T, Gerwick WH and Murray TF (2004) Characterization of the preferred stereochemistry for the neuropharmacologic actions of antillatoxin. J Nat Prod 67:559-568. Lin Y-Y, Risk M, Ray SM, Van Engen 0, Clardy J, Golik J, James JC and Nakanishi K (1981) J. Am. Chem. Soc. 103:6773-6775. Look SA, Fenical W, Matsumoto GK and Clardy J (1986) J. Org. Chem. 51:51405145. Luduena RF, Prasad V, Roach MC, Banerjee M, Yoo HD and Gerwick WH (1997) Drug Delivery Research 40:223-229. Luesch H, Yoshida WY, Moore RE, Paul VJ and Corbett TH (2001) Total structure determination of apratoxin A, a potent novel cytotoxin from the marine cyanobacterium Lyngbya majuscula. JAm Chem Soc 123:5418-5423. Luesch H, Yoshida WY, Moore RE, Paul VJ and Mooberry SL (2000) Isolation, structure determination, and biological activity of Lyngbyabellin A from the marine cyanobacterium lyngbya majuscula. J Nat Prod 63:611-615. Luibrand RT, Erdman TR, Voilmer JJ, Scheuer PJ, Finer J and Clardy J (1979) Tetrahedron 35:609-612. MacMillan JB, Trousdale EK and Molinski TF (2000) Org. Lett. 2:2721-2723. Mahnir V, Lin B, Prokai-Tatrai K and Kem WR (1998) Biopharm. Drug Disposit. 19:147-151. Marquez BL, Watts KS, Yokochi A, Roberts MA, Verdier-Pinard P, Jimenez JI, Hamel E, Scheuer PJ and Gerwick WH (2002) Structure and absolute stereochemistry of hectochlorin, a potent stimulator of actin assembly. J Nat Prod 65:866-871. May WS, Sharkis SJ, Esa AH, Gebbia V, Kraft AS, Pettit GR and Sensenbrenner LL (1987) Proc. Nat!. Acad. Sd. USA 84:8483-8487. Mayer AMS and Hamann MT (2004) Marine Biotechnology 6:37-52. 133 McDonald LA and Ireland CM (1992) J. Nat. Prod. 55:376-379. Milligan KE, Marquez BL, Williamson RT and Gerwick WH (2000) Lyngbyabellin B, a toxic and antifungal secondary metabolite from the marine cyanobacterium Lyngbya majuscula. J Nat Prod 63:1440-1443. Mooberry SL, Leal RM, Tinley TL, Luesch H, Moore RE and Corbett TH (2003) International Journal of Cancer 104:512-521. Moore RE and Scheuer PJ (1971) Science 172:495-498. Murakami Y, Oshima Y and Yasumoto T (1982) Nippon Suisan Gakkaishi 48:69-72. Murata M, Legrand A-M, Ishibashi Y and Yasumoto 1 (1989) J. Am. Chem. Soc. 111:8927-8931. Murata M, Nakoi H, Iwashita 1, Matsunaga S, Sasaki M, Yokoyama A and Yasumoto T (1993) J. Am. Chem. Soc. 115:2060-2062. Newman DJ and Cragg GM (2004) J Nat Prod 67:1216-1 238. Norimura T, Sasaki M, Matsumori N, Miata M, Tachibana K and Yasumoto T (1996) Angew. Chem. mt. Ed. EngI. 35:1675-1678. Nuijen B, Bouma M and Manada C (2000) Anti-Cancer Drugs 11:793-811. Orjala J and Gerwick WH (1996) Barbamide, a chlorinated metabolite with molluscicidal activity from the Caribbean cyanobacterium Lyngbya majuscula. J Nat Prod 59:427-430. Orjala J, Nagle DG, Hsu V and Gerwick WH (1995) Antillatoxin: An Exceptionally lchthyotoxic Cyclic Lipopeptide from the Tropical Cyanobacterium Lyngbya ma] uscula. Journal of the American Chemical Society 117:8281-8282. Osinga R, Armstrong E, Burgess JG, Hoffmann F, Reitner J and Schumann-Kindel G (2001) Hydrobiologia 461:55-62. Perez E, Beatriz A, Perez T, Velasco I, Henriquez P, Munoz M, Moss C and McKenzie D (2004) Sequences from an Endosymbiont and their Uses. International Patent W02004015143. Pettit GR, Herald CL, Doubek DL, Herald DL, Arnold E and Clardy J (1982) J. Am. Chem. Soc. 104:6846-6848. Pettit GR, Kamano Y, Herald CL, Tuinman AA, Boettner FE, Kizu H, Schmidt JM, Baczynkyji L, Tomer KB and Bontems RJ (1987) J. Am. Chem. Soc. 109:7581-82. Poll MA (2002) Recent Advances in Marine Biotechnology 7:1-30. 134 Reddy MV, Rao MR, Rhodes D, Hansen MS, Rubins K, Bushman ED, Venkateswarlu Y and Faulkner DJ (1999) J. Med. Chem. 42:1901-1907. Rinehart K, GloerJ, Cook J, CarterJ, Mizsak Sand Scahill T (1987) JAm Chem Soc 103: 1857-1 859. Rinehart KL, Holt TG, Fregeau NL, Stroh JG, Kiefer PA, Sun F, Li LH and Martin DG (1990)J. Org. Chem. 55:4512-4515. Roussis V, Wu Z, Fenical W, Strobel SA, Van Duyne GD and Clardy J (1990) J. Org. Chem. 55:4916-4922. Salomon CE and Faulkner DJ (2002) J. Nat. Prod. 65:689-692. Salomon CE, Magarvey NA and Sherman DH (2004) Nat Prod Rep 21:105-121. Schaufelberger DE and Pettit GR (1991) J. Nat. Prod. 54:1265-1270. Schmidt EW, Sudek S and Haygood MG (2004) J. Nat. Prod. 67:1341-1 345. Simmons LT, Andrianasolo E, McPhail K, Flatt P and Gerwick WH (2005) Mo!. Cancer Ther. 4:333-342. Sitachitta N, Rossi J, Roberts M, Gerwick WH, Fletcher MD and Willis CL (1998) Biosynthesis of the marine cyanobacterial metabolite barbamide. 1. Origin of the trichloromethyl group. Journal of the American Chemical Society 120:7131-7132. Smith CD, Zhang X, Mooberry SL, Patterson GM and Moore RE (1994) Cancer Research 54:3779-3784. Sone H, Kondo T, Kiryu M, Ishiwata H, Ojika M and Yamada K (1995) Dolabellin, a Cytotoxic Bisthiazole Metabolite from the Sea Hare Dolabella auricularia: Structural Determination and Synthesis. Journal of Organic Chemistry 60:4774-4781. Thacker RW and Starnes S (2003) Marine Biology 142:643-648. Trimurtulu G, Ohtani I, Patterson GM, Moore RE, Corbett TH, Valeriote FA and Demchik L (1994) JAm Chem Soc 116:4729-4737. Vacelet J and Donadey C (1977) J. Exp. Mar. Eco!. 30:301-314. Vaishampayan H, Glode M, Du W, Kraft A, Hudes G, Wright J and Hussain M (2000) Clin. Canc. Res. 6:4205. van Kesteren C, de Vooght M, Mathot R, Schellens J, Jimeno JM and Beijnen JH (2003) Anti-Cancer Drugs 14:487-502. Verdier-Pinard P, Lai JY, Yoo HD, Yu J, Marquez B, Nagle DG, Nambu M, White JD, Falck JR, Gerwick WH, Day BW and Hamel E (1998) Mo! Pharmacol 53:62-76. 135 White JD, Hanselmann Rand Wardrop DJ (1999) JAm Chem Soc 121:1106-1107. Whitton BA and Potts M (2000) Introduction to cyanobacteria. In The ecology of cyanobacteria. Their diversity in time and space., B.A. Whitton and M.Potts, eds. (Dordrecht: KluwerAcademic Publisher),:1-1 I. Williams DE, Burgoyne DL, Rettig SJ, Andersen RJ, Fathi-Afshar ZR and Allen TM (1993) J. Nat. Prod. 56:545-551. Wipf P, Reeves JT and Day BW (2004) Curr. Pharm. Des. 10:1417-1437. Wright AD and Konig GM (1996) J. Nat. Prod. 59:710-716. Wright AE, Forleo DA, Gunawardana GP, Gunasekera SP, Koehn FE and McConnell OJ (1990) J. Org. Chem. 55:4508-4511. Wu M, Okino T, Nogle LM, Marquez BL, Williamson RT, Sitachitta N, Berman FW, Murray IF, McGough K, Jocobs R, Colsen K, Asano T, Yokokawa F, Shioiri T and Gerwick WH (2000) Structure, Synthesis, and Biological Properties of Kalkitoxin, a Novel Neurotoxin from the Marine Cyanobacterium Lyngbya ma] uscula. Journal of the American Chemical Society 122:12041-12042. Wylie BL, Ernst NB, Grace KJS and Jacobs RS (1997) Prog. Surg. 24:146-1 52. Yasumoto I, Oshima Y, Sugawara W, Fukuyo Y, Oguri H, Igarashi T and Fujita N (1980) Nippon Suisan Gakkaishi 46:1405-1411. Yokokawa F, Fujiwara H and Shioiri I (2000) Total synthesis and revision of absolute stereochemistry of antillatoxin, an ichthyotoxic cyclic lipopeptide from marine cyanobacterium Lyngbya majuscula. Tetrahedron 56:1759-1775. Yokokawa F and Shioiri I (1998) J. Org. Chem. 63:8638-8639. Zabriskie TM, Kiocke JA, Ireland CM, Marcus AH, Molinski TF, Faulkner DJ, Xu CF and Clardy J (1986) Jaspamide, a modified peptide from a Jaspis sponge, with insecticidal and antifungal activity. Journal of the American Chemical Society 108:3123-3124. Zewail-Foote M and Hurley L (1999) J Med Chem 42:2493-2497. 136 APPEN DICES 137 APPENDIX A ISOLATION AND STRUCTURE ELUCIDATION OF A NEW CYCLIC DEPSIPEPTIDE, WEWAKMIDE A, FROM A PAPUA NEW GUINEA COLLECTION OF THE MARINE CYANOBACTERIUM LYNGBYA SEMIPLENA In our continuing effort to discover new biologically active and structurally intriguing secondary metabolites from a Papua New Guinea collection of the marine cyanobacterium Lyngbya semiplena, a new cyclic depsipeptide, wewakpeptide A (1), was isolated from the same crude extract as the wewakpeptins had previously been identified wewakamide A (1) lie aba R,3R)-3-amino-2-m thylbutanoic acid Hiv Guineamide B (2) Hiv Dolastatin D (3) (maba) 138 While the IR spectrum displayed absorption bands at 1736 and 1657 cm1 for both ester and amide functionalities, the HRFABMS of compound I analyzed for C53H86N8010, indicating a molecule of peptide origin. 1H and 13C NMR spectra of I were well dispersed in COd3 (Figure A. 1, Table A. 1), allowing for construction of nine partial structures by 20 NMR and accounting for all atoms in the molecular formula of I (Figures A. 2 4). Seven standard amino acids were deduced as Proline (Pro), Valine (Val), Leucine (Leu), Phenylalanine (Phe), N-methyl-alanine (N- MeAla), N-methyl-isoleucine (N-Melle), and N-methyl-leucine (N-MeLeu). Two additional residues were assembled to form the a-hydroxy acid, 2-hydroxyisovaleric acid (Hiva) and the 13-amino acid, 3-amino-2-methylbutanoic acid (Maba). Amino acid sequencing was accomplished by long range 13C-1H correlation experiments (HMBC) with different mixing times and a ROESY experiment. Further evidence supporting this sequence was developed from (MS) experiments (Figure A. 5). Figure A. I. 1H and 13C NMR spectra of wewakamide A (I). 139 IiI 'II, e PS d ,01 U' *+' a 'I ri I Si I U S U I S $4 SI a I U S a I,..'. Figure A. 2. COSY spectrum of wewakamide A (1). 140 Is's IuI 5, t 0 I I 0 p 100 120 ppm pp. 6 6 Figure A. 3. HSQC spectrum of wewakamide A (1). 141 d UI U 0 25 . : U. I S 50 mc *0' J ii* 75 I 100 US 1 U '4 125 I 150 Figure A. 4. HMBC spectrum of wewakamide A (4). 142 m/z 586 (+H) ii m/z 439 (+H) m/z 340 (+1-1) m/z 312 (+H) m/z 227 (+H) iva ProMe Ala Me LeuMe lie Vat Mab aLeu Phe wewakamide A (1) m/z 882 (+H) m/z 783 (1-1) m/z 684(+H) NHPile Hiva Pro Me Ala MeLeuMe lie Vat Mail aLeu - 0 wewakamide A (1) Figure A. 5. Key fragments from collisionally induced ESI-MS/MS experiments with wewakamide A (1). 143 Table A. 1. NMR Spectral Data for Wewakamide A (1) in Unit Position Hiv 1 Pro 2 3 4 5 6 7 8 9 10 N-Me-Ala N-Me-Leu 11 12 13 14 15 16 17 18 19 20 21 N-Me-lie Val 22 23 24 25 26 27 28 NH 29 30 31 Maba Leu 32 33 NH 34 35 36 37 38 NH 39 40 41 42 43 (J in Hz) 5.07, d (9.2) 2.37 0.98 1.08 CDCI3. HMBCa H 75.8 30.6 18.3 19.3 172.6 47.1 23.1 31.6 59.8 2, 3, 4, 5, 53 1,5 1,2,4 1,2,3 3.36, 3.57, m 1.79, 1.59 1.81, 1.61 2.86 7, 9 3.02 10, 12 10, 11, 13, 14, 15 9 6,9 7, 8, 10, 11 169.1 30.3 50.4 15.0 171.3 30.8 55.3 38.3 25.5 24.3 24.4 176.4 33.2 61.3 36.7 16.4 28.1 13.2 5.66,q(7.0) 1.32,d(6.6) 12,14 2.32 5.53 1.76, 1.69 1.48 0.98, d (6.6) 1.09, d (6.6) 12, 14, 16 14, 15, 17,21 16, 18, 19,20 3.08 5.51, d (6.0) 2.29 1.04 1.56, 1.36 0.97 21,23 21, 22, 24, 25,26,28 5.85, d (7.0) 3.66, dd (8.5, 7.5) 2.01, m 1.20, d (6.8) 28, 29, 30 28, 30, 31, 32, 33 29, 31, 32, 29, 30, 32 29, 30, 31 8.30, d (9.8) 4.04, ddq (9.8, 2.8, 7.0) 0.90, d (7.0) 2.32, dq (2.8, 7.0) 1.07, d (7.0) 29, 33, 34 35, 36, 37 34, 36 33, 34, 35, 37, 38 34, 36, 38 7.12 d (7.8) 4.73, dd (6.7, 6.1) 1.76, 1.32 1.78 1.05 38, 39 36, 40, 41 39, 41, 42, 43, 44 39 17, 18, 20 17, 18, 19, 25, 26, 28 23, 24, 24,27 24 172.9 61.8 29.8 20.0 20.2 170.4 47.9 14.2 44.7 13.8 174.3 50.5 38.7 25.8 21.5 21.3 1.10 1.01 40, 41,43 40, 41,42 144 44 Phe 45 46 47 48 49 50 a 169.30 NH 54.4 40.9 137.4 129.1 51 129.6 127.6 129.6 52 53 129.1 170.1 6.45, d (7.1) 4.66, m 3.18, 2.89, dd (6.0, 3.2) 44, 45, 53 46, 53 45, 47, 48/52, 53 7.33 7.35 7.28 7.35 7.33 46,49 47, 48 49,51 47, 52 46, 50, 51 Proton showing HMBC correlation to indicated carbon. The absolute configuration of I was established by analysis of degradation products. A small sample was hydrolyzed with 6N HCI to its constituent amino and hydroxy acid units. These were analyzed by chiral HPLC as well as chiral GO-MS and compared with the retention times of authentic standards. All of the usual amino acids as well as the Hiva unit were shown to possess L-configuration, while the Maba unit was not determined due to unavailability of the standards. A search of the chemical literature indicated that the Maba unit has only been identified in two natural products, guineamide B (2) 2 and dolastatin D (3) Diagnostic NOEs from H-34 to H- 36 were indicative of the relative stereochemistry as 34S*, 36S*. Therefore, we propose that the Maba unit is of the same enantiomeric series as that of dolastatin D because of the comparable spectroscopic properties between I and 3. Wewakamide A (1) was screened for biological activity using brine shrimp toxicity sodium channel modulation and both the NCI-H460 human lung tumor and the neuro-2a mouse neuroblastoma cell lines. This compound, however, proved inactive in all of these assays, except for brine shrimp toxicity, with LC50 = 5 ppm. Interestingly, four other related metabolites, wewakpeptins A - D from the same extract of Lyngbya semiplena display potent cytotoxicity against both the NCI-H460 human lung tumor and the neuro-2a mouse neuroblastoma cell lines. 145 EXPERIMENTAL General Experimental Procedures. Optical rotations were measured on a Perkin-Elmer 141 polarimeter. IR and UV spectra were recorded on Nicolet 510 and Beckman DU64OB spectrophotometers, respectively. NMR spectra were recorded on Bruker Avance DPX 400 MHz and Bruker Avance 300 MHz spectrometers with the solvent CDCI3 used as an internal standard (H at 7.26, ö at 77.4). High resolution mass spectra were recorded on a Kratos MS-50 TC mass spectrometer. Tandem mass spectrometric data were obtained on an electrospray ionization (ESI) quadrupole ion trap mass spectrometer (Finnigan LCQ, San Jose, CA). For ESIMS/MS analysis, samples were injected onto a Cl 8 trap column for desalting and introduced into the mass spectrometer by isocratic elution using 50% acetonitrile containing 0.1 % formic acid. For MS/MS investigations, the protonated molecular ion clusters were isolated in the ion trap and collisionally activated with different collision energies to find optimal fragmentation conditions. For the most intense fragment ions MS3 and MS4 experiments were performed. Chiral GC-MS analysis was accomplished on a Hewlett-Packard gas chromatograph 5890 Series II with a Hewlett-Packard 5971 mass selective detector using an Alitech capillary column (CHIRASIL-VAL phase 25 m x 0.25 mm). HPLC was performed using Waters 515 HPLC pumps and a Waters 996 photodiode array detector. Collection. The marine cyanobacterium Lyngbya semiplena (voucher specimen available from WHG as collection number PNGI2-7Dec99-3) was collected from shallow waters (1-3 m) in Wewak Bay, Papua New Guinea, on December 7, 1999. Taxonomy was assigned by microscopic comparison with the description given by Desikachary.6 The material was stored in 2-propanol at -20°C until extraction. Extraction and Isolation. Approximately 138 g (dry wt) of the alga were extracted repeatedly with CH2Cl2/MeOH (2:1)to produce 3.05 g of crude organic extract. The extract (3.0 g) was fractionated by silica gel vacuum liquid chromatography using a stepwise gradient solvent system of increasing polarity starting from 10% EtOAc in hexanes to 100% MeOH. The fraction eluting with 100% MeOH was found to be active at I ppm in the brine shrimp toxicity assay. This fraction was further chromatographed on Mega Bond RP18 solid-phase extraction 146 (SPE) cartridges using a stepwise gradient solvent system of decreasing polarity starting from 80% MeOH in H20 to 100% MeOH. The most active fractions after SPE (85% toxicity at I ppm to brine shrimp) were then purified by HPLC [Phenomenex Sphereclone 5 p ODS (250 x 10 mm), 9:1 MeOH/H20, detection at 211 nmj giving compounds 1 (2.5 mg), Wewakamide A (1): glassy oil; [a] -83° (c 0.03, CHCI3); UV (MeOH) Amax 215 nm (loge 4.6); IR (neat) 3317, 2966, 2922, 2854, 1736, 1657, 1546, 1461, 1238 cm1; 1H and 13C NMR data, see Table Al; HRFABMS m/z [M + H] 995.6544 (calculated for C53H87N8010, 995.6545). Absolute stereochemistry of 1. Wewakamide A (1, 500 pg) was hydrolyzed in 6 N HCI at 105 °C for 16 h, then dried under a stream of N2 and further dried under vacuum. The residue was reconstituted with 300 pL of H20 prior to chiral HPLC analysis [Phenomenex Chirex 3126 (D), 4.6 x 250 mm; UV 254 nm detector], mobile phase I: 100% 2 mM CuSO4 in mM CuSO4 in MeCN/H20 (15:85), flow H20, flow rate 0.7 mL/min; mobile phase II: 2 rate 0.8 mL/min; mobile phase III: 2 mM CuSO4 in MeCN/H20 (5:95), flow rate 1 mL/min; [column, Phenomenex chirex 3126 (D), 4.6 x 50 mm; UV 254 nm detector], mobile phase IV: 2 mM CuSO4 in MeCN/H20 (15:85), flow rate 0.8 mLlmin. Mobile phase I elution times (tR, mm) of authentic standards: L-MeAla (16.0), D-MeAla (16.5), L-Pro (28.4), D-Pro (63.0), L-Val (38.4), D-Val (68.5). Mobile phase II elution times (tR, mm) of authentic standards: L-MeLeu (12.5), D-MeLeu (14.1), L-Leu (16.0), D-Leu (17.1), L-Phe (39.8), D-Phe (41.3). Mobile phase Ill elution times (tR, mm) of authentic standards: L-Allo-Melle (17.7), L- Melle (18.8), D-Allo-Melle (27.7), D-Melle (28.2). Mobile phase IV elution times (tR, mm) of authentic standards: L-Hiv (9.2), D-Hiv (14.5). The hydrolysate was chromatographed alone and co-injected with standards to confirm assignments (LMeAla, L-Pro, L-Val, L-MeLeu, L-Leu, L-Phe, L-Melle, L-Hiv). Biological Activity. Brine shrimp (Artemia sauna) toxicity was measured as previously described.4 After a 24 h hatching period, aliquots of a 10 mg/mL stock solution of compound A was added to test wells containing 5 mL of artificial seawater and brine shrimp to achieve a range of final concentrations from 0.1 to 100 ppm. After 24 h the live and dead shrimp were tallied. Modulation of the voltage-sensative 147 sodium channel in mouse neuro-2a neuroblastoma cells was also examined as previously described. Cytotoxicity was measured in NCI-H460 human lung tumor cells and neuro-2a mouse neurablastoma cells using the method of Alley et. al 7with cell viability being determined by MIT reduction.5 Cells were seeded in 96-well plates at 6000 cells/well in 180 p1 medium. Twenty-four hours later, the test chemical was dissolved in DMSO and diluted into medium without fetal bovine serum and then added at 20 pg/well DMSO was less than 0.5% of the final concentration. After 48 hr, the medium was removed and cell viability determined. REFERENCES 1. Han, B.; Goeger, D.; Maier, C. S.; Gerwick, W. H. J. Org. Chem. 2005, 70, 3 133-3 139. 2. Tan, L. T.; Sitachitta, N.; Gerwick, W. H. J. Nat. Prod. 2003, 66, 764-771. 3. Sone, H.; Nemoto, T.; lshiwata, H.; Ojika, M.; Yamada, K. Tet. Left. 1993, 34, 8449-52. 4. Meyer, B. N.; Ferrigni, N. R.; Putnam, L. B.; Jacobsen, L. B.; Nichols, D. E.; McLaughlin, J. L. Planta Med. 1982, 45, 31-3. 5. Manger, R. L.; Leja, L. S.; Lee, S. Y.; Hungerford, J. M.; Hokama, Y.; Dickey, R. W.; Granade, H. R.; Lewis, R.; Yasumoto, T.; Wekell, M. M. J. AOAC. mt. 1995, 78, 521-527. 6. Marfey, P. Carlsberg Res. Commun. 1984, 49, 591-596. 7. Alley, M. C., Scudiero, D. A. Cancer Res. 1988, 48, 589-601. 148 APPENDIX B ISOLATION AND STRUCTURE ELUCIDATION OF GUINEAMIDE G FROM A PAPUA NEW GUINEA COLLECTION OF THE MARINE CYANOBACTERIUM LYNGBYA MAJUSCULA Cyanobacteria are phenomenal producers of structurally intriguing and biologically active secondary metabolites such as curacins1 and cryptophycins.2 In our ongoing program to explore these organisms as sources for novel anticancer agents, our most recent investigations have led to the findings of dolabellin,3 ', lyngbyabellins E-1 and aurilides B and C from a collection of the pantropical marine cyanobacterium Lyngbya majuscula Gomont (Oscillatoriaceae). Herein, this paper describes the isolation, structure determination, and a discussion of biological activities of guineamide G (1). An [M + H] peak observed in HR FABMS for guineamide G (1) suggested a molecular formula of C42H55N507, accounting for 18 degrees of unsaturation. The peptidic nature of this molecule was again established from tell-tale exchangeable NH protons resonating at 6 8.84 and 6 5.86 in the 1H NMR data. Similarly to guineamide F (2),6 four singlet CH3 proton signals (6 3.02, 6 3.44, 6 1.23, and 6 1.20) were present in the 1H NMR data (Table B. 1). The two low field singlet proton signals at 6 3.02 and 6 3.44 suggested N-methylations. Six distinct low field carbon signals, due to ester/amide carbonyls, were observed in the 13C NMR data of guineamide G (1) (Figure B. 1). At least 12 olefinic carbon signals, with four peaks having two overlapping carbon signals, were present in the 128-132 ppm range. Taken together with low field aromatic proton signals (6.8-7.4 ppm) in the 1H NMR data, two monosubstituted phenyl groups in I were suggested. Additionally, the 13C NMR spectrum showed two distinctive carbon signals at 6 83.6 and 6 69.3, 149 N-MePhe-1 Dhoya 19 V0 15o10 Gly Pro 26 i=O 0 N 23 0 0 N HN 42 38 13N N-MePhe-2'34 Guineamide F (2) Guineamides G (1) 8.5 170 8.0 160 7.5 150 7.0 140 6.5 130 6.0 5.5 120 5.0 110 4.5 100 4.0 90 3.5 3.0 2.5 80 70 60 Figure B. 1. 1H and 13C NMR spectra of Guineamide G (1). 2.0 50 1.5 40 1.0 30 0.5 20 ppm ppm 150 ppm 0- ' . i * 10 .. : :. : 5P .0 I I. 6 7 B 9 0 1 0 3 1 Figure B. 2. COSY spectrum of Guineamide G (1). 1 U ppm 151 ppm 10 20 rii, 0. 30 S 40 S Co 50 0 60 S 70 S 80 90 100 110 120 130 140 150 9 8 7 6 5 4 3 Figure B. 3. HSQC spectrum of Guineamide G (1). 2 1 0 ppm 152 ppm 20 ' I I I I P I 0 40 60 Is, I La I LaO 80 b * 100 120 4 * 1 140 160 ... ,.. 9 8 7 6 5 4 3 Figure B. 4. HMBC spectrum of Guineamide G (1). 2 1 0 ppm 153 Table B. 1. NMR data for Guineamide G (1) at 400 MHz (1H) and 100 MHz (13C) in CDCI3. multf(Hz) Oc 1 175.8 2 3 brt 47.1 77.1 Position 4a 4b 5a 5b 6 7 8 9 10 HMBC 2,2-dimethyl-3-hydroxy-7-octynoic acid (Dhoya) oH 5.27 1.85 1.69 1.55 1.55 2.27 1.99 1.26 1.24 9.0 29.3 29.3 25.0 25.0 18.4 83.7 69.5 18.3 26.1 m m m m m s s 1,5,9, 10, 11 3,5 3,6 5,8 1,2 1,2 Gly 11 12a 12b NH 3.24 4.72 8.85 m dd d 16.9, 9.6 9.4 170.8 41.3 41.3 11,13 13 N-MePhe-1 13 14 15a 15b 3.89 2.90 3.70 16 17/21 7.10 7.23 19 7.23 22(N-CH3) 18/20 169.0 10.2, 3.1 64.2 13.7, 10.2 34.3 14.5, 3.1 34.3 138.5 129.7 m m 127.2 127.4 m 31.5 3.03s dd dd dd 13, 15, 16, 22, 23 13, 14, 16, 17 15,19 14,23 Pro 23 24 25a 25b 26a 26b 27a 27b 3.41 m 0.74 m -0.03 m 1.27 m 1.27 m 3.20 m 3.39 m 171.3 57.8 30.2 30.2 22.2 22.2 46.5 46.5 25, 26, 27 23, 26, 27 24,27 25, 26 N-MePhe-2 28 29 30a 30b 5.14 2.78 3.12 dd dd dd 7.00 7.23 m m 31 32/36 33/35 168.9 10.2, 4.9 54.2 12.6, 4.9 37.9 12.6, 10.3 137.2 129.3 129.2 28, 30, 31, 37, 38 28, 29, 31, 32 30, 33, 34 31 154 7.19 37(N-CH3) 34 38 39 40 41 42 NH 4.54 1.93 0.95 0.95 5.85 m 3.44s Val brt 7.4 m m m d 7.6 127.5 31.7 173.2 55.2 31.0 18.5 19.3 29,38 1, 38, 40, 41, 38, 39, 41 39, 40, 42 40, 41 1 consistent with a terminal acetylenic functionality. As previously observed, the carbon at 69.3 exhibited weak HSQC correlations but showed a a methine proton at ö 1.93 in the HMBC JCH spectrum.3'4 1JCK coupling of 249 Hz to This proton also exhibited a HMBC correlation to the quaternary carbon at ö 83.6, confirming the presence of an acetylene. An interesting feature of the 1H NMR spectrum of guinamide G was the presence of a shielded diastereotopic proton resonance at ö 0.03. Careful analyses of COSY, HSQC, and HMBC data of guineamide G (1) revealed that it was consisted of five usual amino acid residues as guineamide B (Figures B. 2-4), which are two units of N-MePhe, one Pro, one Val, one Gly, and the 2,2-dimethyl-3-hydroxy-hexanoic acid (Dmhha) residue in 4 was replaced by 2,2- dimethyl-3-hydroxy-7-octynoic acid (Dhoya) in 1. The placement of the shielded proton signal at 0.03 was assigned as one of the methylene protons on 13-C of the Pro unit. Such a shielding effect could arise from the influence of nearby aromatic amino acids, such as N-MePhe-1 and N-MePhe-2 in the molecule. The final planar structure of guineamide G (1) was determined from key proton correlations observed in the HMBC data (Table A 2), which provided the same sequence as presented in structure of 2. Although no HMBC correlations were detected for the N-MePhe-2-Pro sequence, it is the remaining linkage needed to account for the degrees of unsaturation calculated from the molecular formula. Hydrolysis and stereoanalysis of metabolite guineamide G (1) was not undertaken due to the limited time and the desire to preserve as much sample as possible for biological testing. Initially reported, some of the guineamides possess moderate cytotoxicity to a mouse neuroblastoma cell line with lC values of 15 and 16 pM, respectively.6 155 However, guineamide G has shown stronger cytotoxicity to a mouse neuroblastoma cell line with LC values of 2 pglmL. REFERENCES 1. Gerwick, W. H.; Proteau, P. J.; Nagle, D. G.; Hamel, E.; Blokhin, A.; Slate, D. L. J. Org. Chem. 1994, 59, 1243-5. 2. Trimurtulu, G.; Ohtani, I.; Patterson, G. M. L.; Moore, R. E.; Corbett, T. H.; Valeriote, F. A.; Demchik, L. J. Am. Chem. Soc. 1994, 116, 4729-37. 3. Sone, H.; Kondo, T.; Kiryu, M.; lshiwata, H.; Ojika, M.; Yamada, K. J. Org. Chem. 1995, 60,4774-81. 4. Han, B.; MacPhail, K.; Gross, H.; Goeger, D.; Moobery, S. L.; Gerwick, W. H. (manuscript in preparation) 5. Han, B.; Gross, H.; Goeger, D.; Gerwick, W. H. (manuscript in preparation) 6. Tan, L. T.; Sitachitta, N.; Gerwick, W. H. J. Nat. Prod. 2003, 66, 764-71.