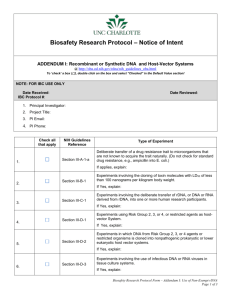
by
advertisement

Role of Topoisomerase I in the Stability of the Ribosomal DNA of Saccharomyces cerevisiae by Fred S. Dietrich B.S. Mathematics/Zoology, University of California - Davis (1985) Submitted to the Department of Biology in partial fulfillment of the requirements for the Degree of Doctor of Philosophy in Biology at the Massachusetts Institute of Technology February, 1996 © Massachusetts Institute of Technology All Rights Reserved Signature of Author: Fred S. Dietrich Thesis Supervisor: Dr. Gerald R. Fink - sor of Biology Certified by: Frank Solomon Professor of Biology Chairman, Committee on Graduate Students JUL 0 8 1996 Role of Topoisomerase I in the Stability of the Ribosomal DNA of Saccharomyces cerevisiae by Fred S. Dietrich Submitted to the Department of Biology on Feb 1, 1996 in partial fulfillment of the requirements for the Degree of Doctor of Philosophy in Biology ABSTRACT Mitotic recombination in the rDNA cluster is suppressed by DNA topoisomerase I and II. Strains containing either a null allele of the TOP1 gene, or a temperature sensitive allele (top2-1) of the TOP2 gene have a rate of mitotic recombination in the rDNA approximately 100 fold higher than do wildtype strains. These mutations have little, if any, effect on recombination at other loci. Using pulse field gel electrophoresis I have further investigated the role of topoisomerases in rDNA. Under a variety of running conditions chromosome XII DNA (containing the rDNA repeat) in a topI strain fails to enter the gel, where as this chromosome enters the gel normally in DNA made from an isogenic TOPi strain. This aberrant migration of chromosome XII is observed only in DNA preparations made from logarithmically growing cells. Thesis Supervisor: Dr. Gerald R. Fink Title: Professor of Biology Acknowledgments I am grateful to all the wonderful members of the Fink lab for their assistance and tolerance. I particularly want to acknowledge the assistance of Cora Styles and of Mike Christman in matters technical and Chris Dreft for many constructive discussions of the materials in this thesis. I would also like to thank Gerry Fink for his guidance and support. I also wish to thank my parents, Sam and Mary Dietrich for their unfaltering support. Table of Contents Title Page ............................................................................ 1 Abstract....................................................................................... ..... 2 Acknow ledgm ents.......................................................................... 3 Table of Contents............................................................................. 4 List of Figures........................................................ 5 List of Tables ................................................ 6 ......................... Chapter I - Introduction ............................ 7-43 The DNA Rotation Problem in Replication........ 8 The DNA Rotation Problem in Transcription.... 10 Topoisomerases .......................................................... 11 Biological Functions of Topoisomerases......... . 15 Topoisomerse Mutants................................... 17 Ribosomal DNA and Topoisomerases............ 18 Examination of Yeast Chromosomes............. 23 References 25 ............................... Chapter II - Mitotic Recombination in the rDNA of S. cerevisiae is suppressed by the combined action of DNA topoisomerase I and II................ 44-57 Chapter III - The ribosomal DNA array has an altered structure in topoisomerase I mutants of S. cerevisiae.......... ..................... 58-77 List of Figures Chapter I Figure 1 - Supercoiling by the DNA replication m achinery ............................................................. Figure 2 - Overall arrangement of the ribosomal DNA of S. cerevisiae ........................................................ Chapter II Figure 1 - Loss of a URA3 Insertion in the rDNA via Mitotic Recombination............................... Figure 2 - Segregation of Form I/Form II rDNA Polymorphisms after Recombination............. Figure 3 - DNA Probe Used to Distinguish between the Form I and Form II rDNA Polymorphisms....... Figure 4 - Southern Analysis of Tetrads to Follow the Segregation of Form I/FormII rDNA Polymorphisms in Mitosis and Meiosis in topl/topl and TOP1/TOP1 Backgrounds......... Figure 5 - Segregation of rDNA Markers at Meiosis following Unequl Sister Chromatid Exchange in the rDNA During Meiosis............................. Chapter III Figure 1 - Pulsed field gel electrophoresis of yeast chromosomal DNA................................ Figure 2 - Pulsed field gel electrophoresis of yeast restriction digested DNA..................................... Figure 3 - Construction of a strain containing rDNA on chrom osom e III................................. . ............ Figure 4 - Chromosome preparations from top1 and TOP1 strains grown to stationary phase....................... Figure 5 - Chromosomes from top2 strains............................. 10 19 46 48 48 49 52 63 65 66-67 69 71 List of Tables Chapter II Table 1 - Yeast Strains.................................................................. 46 Table 2 - Frequency and Rate of Mitotic Recombination in rDNA in TOP1 and top1 Strains...................... 47 Table 3 - Frequency of Mitotic Recombination in rDNA in rDNA in top2-1(ts) and TOP2 Strains................ 49 Table 4 - Mitotic Recombination at Non-rDNA Loci in top1 and TOP1 Strains........................ .............. 50 Table 5 - Mitotic Recombination at Non-rDNA Loci in top2-1(ts) and TOP2 Strains............................. 50 Table 6 - Frequency of Mitotic Recombination in rDNA in rad52 strains............................................. .......... 51 Table 7 - Meiotic Recombination in rDNA in topl/topl and TOP1/TOP1 Backgrounds................... 52 Chapter I Introduction Intoduction The subject of my thesis is how a specific class of yeast enzymes, the topoisomerases, interact with a specific portion of the yeast genome, the ribosomal DNA. Both of these subjects, topoisomerases and ribosomal DNA, have been extensively studied; the following introduction covers the more relevant aspects of these fields. The double helix structure of DNA imposes certain restrictions on the mechanisms of DNA replication, transcription, and recombination. This restriction, arising out of the necessity to unwind the strands, was first realized to be a problem soon after the elucidation of the structure of DNA (Watson and Crick 1953; Delburck 1954). Though early on it was realized that a DNA swivel was needed for unwinding the DNA, it was not until 1971 that such a swivel, in the form of an enzyme now referred to as a topoisomerase, was first reported (Wang 1971). The DNA Rotation Problem in Replication To replicate DNA it is first necessary to separate the two DNA strands in order for the replication complex to form. This process has been observed through sensitivity of the origin region, e.g. oriA (Schnos et al. 1988), to single strand nucleases such as S1 and P1. For DNA replication to proceed either the replication complex must rotate, following the rotation of the parental DNA, or a swivel must be introduced into the DNA to allow rotation around the DNA axis (See figure 1). Rotation of the DNA complex results in two highly intertwined daughter molecules which in the case of a circular DNA molecule is resolvable only by breaking one daughter molecule and passing the other daughter molecule through the break. A swivel necessitates at least one additional swivel after the replication fork to reintroduce the twists that were removed ahead of the replication fork. DNA molecules can be underwound (i.e. less tightly twisted than a relaxed molecule) or overwound (more tightly twisted) relative to the relaxed molecule. Underwinding or overwinding is generally referred to as supercoiling; an underwound molecule is said to be negatively supercoiled and an overwound molecule is said to be. positively supercoiled. Topoisomerases are important in the regulation of supercoiling. Supercoils can be introduced into DNA in a number of ways. In Escherichia coli DNA gyrase acts specifically to introduce negative supercoils (Gellert et al. 1976); in eukaryotes there is no known comparable enzyme that specifically introduces supercoils. A number of other processes, discussed below, can potentially affect the degree of supercoiling either by introducing or removing supercoils. I I OC13 B. C. D. Figure 1. Supercoiling by the DNA replication machinery(represented by the oval). As replication proceeds from (A) to (B) the DNA becomes underwound behind the point of replication and overwound ahead of the point of replication (assuming the polymerase does not rotate). Similarly, as the transcription machinery moves from (C) to (D) overwinding occurs ahead of the transcription and underwinding occurs after. The DNA Rotation Problem in Transcription The structure of DNA necessitates that transcription, like replication, can proceed only if there is rotation of the DNA or rotation of the transcription machinery around the DNA. Because of the size of the transcription complex its rotation has generally been considered implausible. The more widely favored model has been that transcription induces positive supercoiling in front of the transcription fork, and negative supercoiling after the transcription fork (Liu and Wang 1987). If the number of positive supercoils removed ahead of the transcription machinery does not exactly match the number of negative supercoils removed after the transcription machinery, then a net change in the degree of supercoiling will result from transcription. Evidence that transcription affects the supercoiling of circular plasmid DNA in Saccharomyces cerevisiae comes from experiments in top1 top2 double mutant strains where inducible transcription of a plasmid results in alterations in supercoiling (Brill and Sternglanz 1988; Giaever and Wang 1988). When multiple transcription complexes move in tandem through a region, the positive supercoil induced in front of the trailing transcription complex can cancel out the negative supercoils produced by the leading transcription complex. Topoisomerases Topoisomerases are classified into 2 types. Type I topoisomerases are those able to transiently break a DNA strand, pass another strand through the break, and reseal the break. Type II topoisomerases are able to break both strands of a DNA molecule, pass another double stranded DNA molecule through the break, and reseal the break. The type 1 topoisomerase of E. coli was the first topoisomerase characterized (Wang 1971). The type I topoisomerases can further be classified as "topA like" and "TOP1 like". While both of these groups relax DNA by producing and resealing a single strand nick and do not require ATP, there are several significant differences between them. The "topA like" enzymes , such as E. coli topA, only relax negatively supercoiled DNA (Wang 1971) or DNA molecules with single-stranded regions (Kirkegaard and Wang 1985) where as the "TOP1 like" enzymes, such as S. cerevisiae top1, can relax both positive and negative supercoils (Champoux and Dulbecco 1972). At the reaction level, E. coli topA attaches to the 5' end of the nicked DNA (Depew et al. 1978; Kirkegaard et al. 1984) where as the eukaryotic TOP1 attaches to the 3' end (Champoux 1977; Champoux 1978). An additional specialized type I topoisomerase differing in properties from both the "topA like" or the "TOP1 like" enzymes has been characterized from the thermophilic archaebacterium Sulfolobus acidocaldariuswhich is ATP dependent and can introduce positive supercoils (Kikuchi and Asai 1984; Mirambeau et al. 1984). The original assay for determining degree of supercoiling of closed circular DNA molecules was measurement of sedimentation rate of relaxed versus supercoiled DNA by ultracentrifugation(Wang 1971). Several more convenient assays involving filter binding (relying on the affinity of single stranded regions of negatively supercoiled DNA) and nuclease sensitivity have been used as topoisomerase assays (Wang and Kirkegaard 1981) though the standard assay now is gel electrophoresis (Keller 1975). This standard assay for topoisomerase I measures relaxation of supercoiled plasmid DNA by taking advantage of the fact that plasmids of different degrees of supercoiling can easily be resolved on an agarose gel. This assay is specific for type I topoisomerases because it is done without ATP which is required by type II topoisomerases. A number of type 1 topoisomerases have been cloned and sequencedl.In S. cerevisiae and S. pombe topoisomerase I has been shown to be non-essential though strains carrying deletions of these genes have slightly longer generation times (Thrash et al. 1985; Uemura et al. 1987). Deletions of the E. coli topA gene are lethal, and overexpression causes slow growth (Wang and Becherer 1983) but can be compensated for by second site mutations in the gyrase genes (DiNardo et al. 1982; Pruss et al. 1982), at the uncharacterized toc (topoisomerase one compensatory) locus (Raji et al. 1985), by enhanced expression of the gyrA and gyrB homologues parC and parE (Kato et al. 1990), or by an inhibitor of gyrase (Hammond et al. 1991). In addition the E. coli topA mutation can be complemented by the yeast topoisomerase I gene (Bjornsti and Wang 1987). Sequence comparisons of the type I topoisomerases have revealed that the "topA like" and the "TOP1 like" enzymes constitute two distinct classes on the basis of sequence similarity. In the "topA like" class in addition to E. coli topA and Salmonella typhimurium topA are thetopB gene of E. coli (DiGate and Marians 1989) and the S. cerevisiae TOP3 gene (Wallis et al. 1989). The topB gene product is able to decatenate concatenated double strand DNA circles which contain a single stranded region or a nick. The topB gene is also able to relax negatively supercoiled DNA circles as does topA. This relaxation activity can be detected in strains lacking topA (Srivenugopal et al. 1984; DiGate and Marians 1988). No biochemical activity has yet been ascribed to the TOP3 gene product from S. cerevisiae. 1 E. coli topA (Wang and Becherer 1983), Salmonella typhimurium (partial sequence only) (Ostrowski et al. 1987), yeast S. cerevisiae (Goto and Wang 1985; Thrash et al. 1985),Schizosaccharomyces pombe (Uemura et al. 1987), rat (Durban et al. 1988), vaccinia (Shuman and Moss 1987), Shope fibroma virus (Upton et al. 1990), and human (D'Arpa et al. 1988). Unlike the type I topoisomerase, all type II topoisomerases that have been sequenced so far 2 share extensive sequence homology. The main distinction among the various type II topoisomerases is between over- and underwinding enzymes. Bacterial gyrase is able to introduce negative supercoils (Gellert et al. 1976; Gellert et al. 1976; Liu and Wang 1978) whereas the eukaryotic type II topoisomerases found thus far cannot introduce negative supercoils but can relax either positively or negatively supercoiled DNA. The Eukaryotic type II topoisomerases are encoded by a single gene, where as the T4 equivalent is specified by three genes and the E. coli and B. subtilis equivalents by two. It has been shown that the Drosophila topoisomerase II can complement S. cerevisiae top2 mutations (Wyckoff and Hsieh 1988), that antibodies against purified Drosophila topoisomerase II can cross react with the S. cerevisiae Topoisomerase II (Heller et al. 1986), and that the TOP2 genes from S. cerevisiae and S. pombe can compliment in the heterologous yeast. In E. coli the product of the parC and parE loci are gyrA and gyrB homologues and can compensate for mutations in topA. The standard assay for topoisomerase II is the ATP dependent decatenation of kinetoplast DNA from trypanosomes (Marini et al. 1980). This assay is specific for type II topoisomerases because type I topoisomerases are unable to decatenate concatenated doubled stranded DNA circles. In addition to the general topoisomerases, a number of enzymes can act as site specific topoisomerases. Lambda int (Nash et al. 1981), and yeast S. cerevisiae 2 E. coli gyrase (Adachi et al. 1987; Swanberg and Wang 1987), phage T4 (Huang 1986; Huang 1986), Mycoplasma pneumoniae (Colman et al. 1990), Staphylococcus aureus (Hopewell et al. 1990), Haloferax sp.(Holmes and Dyall-Smith 1991), Neisseria gonorrheae (Stein et al. 1991), Bacillus subtilis (Moriya et al. 1985), Klebsiella pneumoniae (Dimri and Das 1990), Pseudomonas putida (Parales and Harwood 1990), Crithidiafasciculata (Pasion et al. 1992), S. cerevisiae (Giaever et al. 1986), Trypanosoma brucei (Strauss and Wang 1990), S. pombe (Uemura et al. 1986), Drosophila (Wyckoff et al. 1989), human (Tsai et al. 1988) flp protein (Beatty et al. 1986; Volkert and Broach 1986). both can alter the supercoiling of their respective substrates but differ from TOPI or TOP2 because they recognize specific sequences in the DNA. Similarly transposable element resolvases such as that of Tn3 (Krasnow and Cozzarelli 1983; Krasnow et al. 1983) can affect DNA supercoiling. Broken DNA strands either from DNA damage or excision repair mechanisms can also relax supercoiled DNA. Biological Functions of Topoisomerases Although a fair amount is known about the structure and biochemical activities of various topoisomerases, the question of their specific biological role is still open. One clue to the biological role of topoisomerases comes from sub-cellular localization studies. A number of studies have localized topoisomerase I to transcriptionally active regions. In Drosophila, antibodies against topoisomerase I stain the induced but not the uninduced heat shock loci (Fleischmann et al. 1984). One of the most heavily transcribed regions of a eukaryotic genome is the cluster of genes encoding the ribosomal RNA (rRNA), known as the ribosomal DNA (rDNA), found in the nucleolus. Recently in situ hybridization has shown that the S. cerevisiae nucleolus is as in other organisms composed of ribosomal DNA (Dvorkin et al. 1991). Topoisomerase I has been localized to the nucleolus in Drosophila (Fleischmann et al. 1984) a chicken lymphoblastoid cell line (Muller et al. 1985), rat (Thiry et al. 1991), and S. cerevisiae (Giroux et al. 1989). That DNA supercoiling can affect transcription as has been shown by studies in which the degree of supercoiling of the substrate is altered. In vitro studies of closed circular lambda DNA showed that more negatively supercoiled molecules have a greater rate of initiation of transcription (Botchan et al. 1973). In vivo synthesis of DNA gyrase has been shown to be higher when the template is relaxed than when it is negatively supercoiled(Menzel and Gellert 1983). Studies mapping DNA nicks resulting from interaction of the topoisomerase I specific inhibitor camptothecin have also shown a high level of topoisomerase I in transcriptionally active regions (Gilmour and Elgin 1987; Gilmour and Elgin 1987; Stewart et al. 1990). The transcription rate in the nucleolus is very high and in a number of systems topoisomerase I has been shown to localize to this region. The first work in this area was an examination of DNA-relaxing activity in isolated nucleoli compared with total DNA (Higashinakagawa et al. 1977) DNArelaxing activity co-segregated with RNA polymerase activity and appeared to be largely associated with the nucleoli. Because of the assay conditions this polymerase associated DNA-relaxing activity is most likely topoisomerase I. Transcription in vitro of rat rDNA appears to be reduced upon addition of the specific topoisomerase I inhibitor camptothecin. (Garg et al. 1987). This inhibition could be due to two different mechanisms. One possibility is that reduced transcription results from the decrease in topoisomerase activity. The other possibility, frequently not addressed, is that the camptothecin-DNAtopoisomerase complex blocks transcription directly. Blockage of transcription by this complex has been shown to occur under some conditions (Bendixen et al. 1990). This second possibility was ruled out in Garg et. al. by addition of purified topoisomerase I which restored the pre-camptothecin level of expression. A similar experiment in which anti-topoisomerase I antibodies were used instead of camptothecin also showed that inhibition of topoisomerase I inhibits ribosomal transcription in a way that can be bypassed by adding exogenous topoisomerase I (Egyhazi and Durban 1987). Topoisomerse Mutants In both S. cerevisiae (Brill et al. 1987) and S. pombe (Yamagishi and Nomura 1988) top1 top2-ts double mutant strains display greatly reduced ribosomal RNA synthesis at restrictive temperature for top2-ts. This phenotype is not seen in strains carrying either a top1 or top2-ts single mutation, suggesting that these topoisomerases have at least partially overlapping function. Additionally, topl top2-ts double mutant S. cerevisiae strains contain a large number of free rDNA circles (Kim and Wang 1989). These free rDNA circles are seen in large numbers only in the double mutant strains, not in strains carrying either a topl or a top2 mutation alone. In top1 top2-4 double mutant strains more than half of the rDNA was shown to be in the form of extrachromosomal rings, where as in top1 top2-1 double mutant strains few free rDNA circles were seen. In both S. cerevisiae and S. pombe top1 deletions are viable, although they grow more slowly than wildtype (Thrash et al. 1985; Uemura et al. 1987). One explanation for this phenotype is that topoisomerase I plays an important role, but one which can be substituted for by other topoisomerases. Topoisomerase II function appears to be essential for mitotic chromosome segregation, but not for DNA synthesis. Evidence for this comes from cell synchronization experiments in S. cerevisiae where temperature sensitive mutations lead to inviability at the time when the cells are undergoing mitosis, which is clearly after DNA synthesis (DiNardo et al. 1984; Holm et al. 1985). Additional evidence is that inhibition of topoisomerase II with VM-26, a specific inhibitor of topoisomerase II (Chen et al. 1984), does not affect DNA replication but delays formation of mitotic chromosomes (Charron and Hancock 1990). Topoisomerase I is also not required for DNA replication. If topoisomerase I plays an important role in DNA replication then it's function must be interchangeable with that of another gene product because yeast strains carrying topi deletions are viable. Although single mutations in either topoisomerase I or topoisomerase II do not block DNA replication, double mutant strains lacking both topoisomerases are blocked in mitotic DNA replication (Brill et al. 1987; Kim and Wang 1989). Similarly, in S. cerevisiae topoisomerase II appears to be required for meiotic chromosome segregation, but not pre-meiotic DNA replication (Rose et al. 1990). Likewise in surf clam oocytes the topoisomerase II inhibitor VM-26 can block both meiotic and subsequent mitotic chromosome condensation (Wright and Schatten 1990). The various models for the role of topoisomerases are somewhat clouded by the recent discoveries of additional topoisomerase genes, including topB (DiGate and Marians 1989) and parC-parE(Kato et al. 1990) in E. coli, and top3 (Wallis et al. 1989) and hprl (Aguilera and Klein 1990) in S. cerevisiae. Little is known about the roles of these enzymes, but their existence suggests that there may be additional mechanisms for regulating supercoiling. Ribosomal DNA and Topoisomerases The ribosomal DNA of S cerevisiae is composed of 50-200 (Retal and Planta 1968; Schweizer et al. 1969; Zamb and Petes 1982) tandemly repeated units of approximately 9Kb each (Cramer et al. 1976) located on chromosome XII (Petes 1979; Petes 1979; Petes and Smolik 1979; Szostak and Wu 1979) which behaves as a single Mendelian trait (Petes and Botstein 1977) and whose replication in meiosis and mitosis is similar to that of the rest of the genome (Brewer et al. 1980). Most or all of this region has been sequenced 3 though not all from the same strain and there remain a few ambiguities. Some of the main features of S. cerevisiae rDNA are shown in figure 2. A. Unit Repeat 5S 35S 5S 35S 35S B. NTS1 35S T E NTS2 5S A P 35S Figure 2. Overall arrangement of the ribosomal DNA of S. cerevisiae is 50-200 copies of the repeat shown in (A). 5S and 35S are the small and large ribosomal transcription units respectively. Details of the spacer regions are shown in in (B). Features are as follows: NTS1, nontranscribed spacer region 1; NTS2, nontranscribed spacer region 2; T, Putative Topoisomerase I binding site; E, 35S enhancer; A, ARS; P 35S promoter. Ribosomal DNA transcription in S. cerevisiae has been extensively studied. Each repeat contains two transcripts, the small 5S and the large 35S transcript which are divergently transcribed (Phillippsen et al. 1978). Promoter, enhancer, and terminator sequences necessary for transcription of the 35S (Bayev et al. 1980; Elion and Warner 1984; Elion and Warner 1986; Mestel et 3 (Bell et al. 1977; Olson et al. 1977; Valenzuela et al. 1977; Kramer et al. 1978; Skryabin et al. 1979; Skryabin et al. 1979; Bayev et al. 1980; Rubtsov et al. 1980; Veldman et al. 1980; Veldman et al. 1980; Bayev et al. 1981; Georgiev et al. 1981; Swanson and Holland 1983; McMahon et al. 1984; Piper et al. 1984; Skryabin et al. 1984; Nazar and Wong 1985; Swanson et al. 1985; Jemtland et al. 1986; Mankin et al. 1986; Vincent and Petes 1986; Riggs and Nomura 1990) al. 1989) ribosomal RNA have been characterized. Expression of the 5S RNA, which is transcribed by RNA polymerase III, is less well characterized, though appears to be independent of the RNA polymerase I enhancer (Neigeborn and Warner 1990). Each of the rDNA repeats contains an ARS element located between the 5' ends of the 5S and 35S transcription units (Skryabin et al. 1984). This ARS unit was first characterized by its ability to stabilize a circular plasmid (Szostak and Wu 1979; Kouprina and Larionov 1983), and has subsequently been verified to be a chromosomal origin of replication by electron microscopy (Saffer and Miller 1986) and by two dimensional agarose gel electrophoresis. Two separate two-dimensional gel electrophoresis techniques have been developed. One approach allows the characterization of the forked and bubble structures which identify a replication origin by separating the DNA in the native form in the first dimension, denaturing the DNA, and separating the single stranded DNA in the second dimension (Linskens and Huberman 1988). The second approach involves separating the DNA in the native form in both dimensions. The second dimension is run under conditions in which forked and bubble structures migrate slower relative to lineat molecules than they do in the first dimension (Brewer et al. 1988). The electron microscopic analysis of rDNA replication by Saffer and Miller reveals that replication occurs in transcriptionally active rDNA. It has been noted that in E. coli the replication and transcription of the rDNA proceed in the same direction (Nomura et al. 1977) and it has been hypothesized that this is necessary to avoid collisions between the DNA and RNA polymerases. This may also be the case in S. cerevisiae where it has been that DNA replication is blocked at the 3' end of the 35S transcription unit, as seen by the accumulation of replication forks at this point (Brewer and Fangman 1988) suggesting that the majority of the rDNA is replicated by movement of the replication fork in the same direction as transcription of the 35S rRNA. It is interesting to note that figures 4 and 5 in the paper by Saffer and Miller (Saffer and Miller 1986) show three fairly clear examples of actively transcribed rDNA units with replication forks located at their 3' ends. The presence of an ARS element within each rDNA repeat explains the existence of extrachromosomal rDNA circles which have been reported and are generally referred to as 3ýp circles (Clark-Walker and Azad 1980; Larionov et al. 1980). In wildtype cells these free circles appear to compose at most a small fraction of the total rDNA (Szostak and Wu 1979), though in the absence of topoisomerase they constitute a large fraction of the ribosomal DNA (Kim and Wang 1989). Tetrad analysis of a yeast strain carrying a restriction site polymorphism in the ribosomal DNA revealed of 14 tetrads no meiotic recombinants but two examples of mitotic recombination events that occurred before meiosis (Petes and Botstein 1977). The suggestion in these data that mitotic recombination occurs at a higher frequency than meiotic recombination is contrary to what is usually found for non-rDNA. A subsequent random screen for recombination stimulating sequences has shown that the rDNA fragments containing the 35S promoter and enhancer together can act as a mitotic (but not meiotic) recombination hot spot and are referred to as HOT1 (Keil and Roeder 1984; Voelkel-Meiman et al. 1987). It appears that HOT1 is cis-acting and that the stimulation of recombination is due to RNA polymerase I (Stewart and Roeder 1989). Analysis of HOT1 has been done at non-rDNA loci, so it is still not clear if RNA polymerase stimulates recombination within the rDNA. The recent development of a S. cerevisiae strain containing RNA polymerase II transcribed 35S RNA in which RNA polymerase I is dispensable (Nogi et al. 1991) should make it possible to determine if HOT1 stimulates mitotic recombination in the rDNA and if so if it is RNA polymerase I dependent. A feature of the ribosomal DNA of Tetrahymena and Dictyostelium is the presence of topoisomerase I binding sites (Bonven et al. 1985; Ness et al. 1988). Although topoisomerase I will bind and relax any DNA sequence, the enzyme has certain preferred cutting sites (Been et al. 1984; Champoux et al. 1984; Kirkegaard et al. 1984). These rDNA high affinity binding sites however have an even greater affinity for topoisomerase I, roughly 3 orders of magnitude greater than random DNA sequences (Busk et al. 1987; Thomsen et al. 1987). In S. cerevisiae there is a homologous site between the 3' end of the 35S transcript and the RNA polymerase I enhancer sequence. However, this sequence has not yet been shown to be a binding site. No role has been shown for any of these binding sites, nor is it known if localization of topoisomerase I to the nucleolus is due to the presence of these binding sites. One report suggests that S. cerevisiae topoisomerase I is capable of binding to the Tetrahymena high affinity binding sites (Bonven et al. 1985). Recombination of repeated units in S. cerevisiae has been studied in both mitosis and meiosis both between naturally occurring repeats and synthetic repeats (Petes 1980; Szostak and Wu 1980; Jackson and Fink 1981; Jackson and Fink 1985; Jinks-Robertson and Petes 1985; Jinks-Robertson and Petes 1986; Kupiec and Petes 1988). Recombination in repeated tandem arrays such as the rDNA can apparently occur by multiple mechanisms, including a RAD52independent pathway (Ozenberger and Roeder 1991). There appears to be an interesting relationship between topoisomerases and repeated DNA in S. cerevisiae. Mutations in several topoisomerases and topoisomerase homologues preferentially elevate the level of recombination between specific repeated elements. A mutation known as edrl (now called top3) was identified which enhanced recombination between repeated 8 elements flanking the SUP4-o marker (Rothstein 1984; Wallis et al. 1989). Another topoisomerase homologue was identified by a screen for mutations which increase recombination between synthetic direct repeats (Aguilera and Klein 1988; Aguilera and Klein 1989). This mutation, called hprl is in a gene homologous to TOPI (Aguilera and Klein 1990). Strains carrying either topl or top2 mutations show an elevated level of recombination in the rDNA of S. cerevisiae but not at a number of other loci (Christman et al. 1988). There also appears to be a relationship between recombination and transcription. In nonrDNA recombination has been shown to be enhanced by transcription from an RNA polymerase I promoter (Voelkel-Meiman et al. 1987) and also to be stimulated by RNA polymerase II transcription (Thomas and Rothstein 1989). It is possible that the affect of transcription on recombination may be due to changes in supercoiling arising from the transcription process. Examination of Yeast Chromosomes Yeast chromosomes are much smaller than those of higher eukaryotes and it is only within the last few years that techniques have been developed that allow entire chromosomes to be examined. One technique is the meiotic spread preparation (Dresser and Giroux 1988) in which the structure of condensed meiotic chromosomes can be examined. The other technique is that of pulse field gels (Schwartz and Cantor 1984; Carle and Olson 1985). These techniques provide a possible assay for mutations which affect long range structures in DNA. Supercoiling of DNA and the effects of topoisomerase on supercoiling are one of these long range affects. In my research I have tryed to address a number of questions about topoisomerases in yeast. What is the relationship between topoisomerases and recombination? Is the elevated level of recombination found in rDNA specific to rDNA or is it found elsewhere at other loci? In particular, do yeast strains carrying topoisomerases have an elevated level of recombination in other tandemly repeated genes such as CUP1. Another issue I have addressed is whether large scale changes in the rDNA are detectable by pulse field gel electrophoresis. References Adachi, T., M. Mizuuchi, E.Robinson, E.Appella, M. O'dea, M. Gellert and K. Mizuuchi (1987). "DNA sequence of the E. coli gyrB gene: Application of a new sequencing strategy." Nuc. Acids Res. 15: 771-783. Aguilera, A. and H. L. Klein (1988). "Genetic control of intrachromosomal recombination in Saccharomyces cerevisiae. I. Isolation and genetic characterization of hyper-recombination mutations." Genetics 119: 779790. Aguilera, A. and H. L. Klein (1989). "Genetic and molecular analysis of recombination events in Saccharomyces cerevisiae occurring in the presence of the hyper-recombination mutation hprl." Genetics 122: 503-517. Aguilera, A. and H. L. Klein (1990). "HPR1, a novel yeast gene that prevents intrachromosomal excision recombination, shows carboxy-terminal homology to the Saccharomyces cerevisiae TOP1 gene." Mol. Cell. Biol. 10: 1439-1451. Bayev, A., O. I. Georgiev, A. A. Hadjiolov, N. Nikolaev, K. G. Skryabin and V. M. Zakharyev (1981). "The structure of the yeast ribosomal RNA genes. 3. Precise mapping of the 18 S and 25 S rRNA genes and structure of the adjacent regions." Nuc. Acids Res. 9: 789-799. Bayev, A. A., O. I. Georgiev, A. A. Hadjiolov, M. B. Kermekchiev, N. Nikolaev, K. G. Skryabin and V. M. Zakharyev (1980). "The structure of the yeast ribosomal RNA genes. 2. The nucleotide sequence of the initiation site for ribosomal RNA transcription." Nuc. Acids Res. 8: 4919-4926. Beatty, L. G., C. D. Babineau, C. Hogrefe and P. D. Sadowski (1986). "FLP sitespecific recombinase of yeast 2-micron plasmid. Topological features of the reaction." J. Mol. Biol. 188: 529-544. Been, M. D., R. R. Burgess and J. J.Champoux (1984). "Nucleotide sequence preference at rat liver and wheat germ type 1 DNA topoisomerase breakage sites in duplex SV40 DNA." Nuc. Acids Res. 12: 3097-3114. Bell, G. I., L. J. Degennaro, D. H. Gelfand, R. J. Bishop, P. Valenzuela and W. J. Rutter (1977). "Ribosomal RNA genes of Saccharomyces cerevisiae: I. Physical map of the repeating unit and location of the regions coding for 5S, 5.8S, 18S, and 25S ribosomal RNAs." J. Biol. Chem. 252: 81188125. Bendixen, C., B. Thomsen, J. Alsner and 0. Westergaard (1990). "Camptothecin-stabilized topoisomerase I-DNA adducts cause premature termination of transcription." Biochemistry 29: 5613-5619. Bjornsti, M. A. and J. C. Wang (1987). "Expression of yeast DNA topoisomerase I can complement a conditional-lethal DNA topoisomerase I mutation in Escherichia coli." Proc. Natl. Acad. Sci. 84: 8971-8975. Bonven, B. J., E. Gocke and 0. Westergaard (1985). "A high affinity topoisomerase I binding sequence is clustered at DNAase I hypersensitive sites in Tetrahymena R-chromatin." Cell 41: 541-551. Botchan, P., J. C. Wang and H. Echols (1973). "Effect of circularity and superhelicity on transcription from bacteriophage lambda DNA." Proc. Natl. Acad. Sci. 70: 3077-3081. Brewer, B. J.and W. L. Fangman (1988). "A replication fork barrier at the 3' end of yeast ribosomal RNA genes." Cell 55: 637-643. Brewer, B. J., E. P. Sena and W. L. Fangman (1988). "Analysis of replication intermediates by two-dimensional agarose gel electrophoresis." Cancer Cells 6: 229-234. Brewer, B. J., V. A. Zakian and W. L. Fangman (1980). "Replication and meiotic transmission of yeast ribosomal RNA genes." Proc. Natl. Acad. Sci. 77: 6739-6743. Brill, S. J., S. DiNardo, M. K. Voelkel and R. Sternglanz (1987). "DNA topoisomerase activity is required as a swivel for DNA replication and for ribosomal RNA transcription." NCI Monogr. 1987: 11-15. Brill, S. J. and R. Sternglanz (1988). "Transcription-dependent DNA supercoiling in yeast DNA topoisomerase mutants." Cell 54: 403-411. Busk, H., B. Thomsen, B. J.Bonven, E. Kjeldsen, O. F. Nielsen and 0. Westergaard (1987). "Preferential relaxation of supercoiled DNA containing a hexadecameric recognition sequence for topoisomerase I." Nature 327: 638-640. Carle, G. F. and M. V. Olson (1985). "An electrophoretic karyotype for yeast." Proc. Natl. Acad. Sci. 82: 3756-3760. Champoux, J. J. (1977). "Strand breakage by the DNA untwisting enzyme results in covalent attachment of the enzyme to DNA." Proc. Natl. Acad. Sci. 74: 3800-3804. Champoux, J. J.(1978). "Mechanism of the reaction catalyzed by the DNA untwisting enzyme: Attachment of the enzyme to 3' -terminus of the nicked DNA." J. Mol. Biol. 118: 441-446. Champoux, J. J. and R. Dulbecco (1972). "An activity from mammalian cells that untwists superhelical DNA-A possible swivel for DNA replication." Proc. Natl. Acad. Sci. 69: 143-146. Champoux, J. J., W. J. McCoubrey and M. D. Been (1984). "DNA structural features that lead to strand breakage by eukaryotic type-I topoisomerase." CSH Symp. Quant. Biol. 49: 435-42. Charron, M. and R. Hancock (1990). "DNA topoisomerase II is required for formation of mitotic chromosomes in Chinese hamster ovary cells: studies using the inhibitor 4'-demethylepipodophyllotoxin 9-(4,6-0thenylidene-beta-D-glucopyranoside)." Biochemistry 29: 9531-9537. Chen, G. L., L. Yang, T. C. Rowe, B. D. Halligan, K. M. Tewey and L. F. Liu (1984). "Nonintercalative antitumor drugs interfere with the breakagereunion reaction of mammalian DNA topoisomerase II." J. Biol. Chem. 259: 13560-13566. Christman, M. F., F. S. Dietrich and G. R. Fink (1988). "Mitotic recombination in the rDNA of S. cerevisiae is suppressed by the combined action of DNA topoisomerases I and II." Cell 55: 413-425. Clark-Walker, G. D. and A. A. Azad (1980). "Hybridizable sequences between cytoplasmic ribosomal RNAs and 3 micron circular DNAs of Saccharomyces cerevisiae and Torulopsis glabrata." Nuc. Acids Res. 8: 1009-1022. Colman, S. D., P. C. Hu and K. F. Bott (1990). "Mycoplasma pneumoniae DNA gyrase genes." Mol. Microbiol. 4: 1129-1134. Cramer, J. H., F. W. Farrelly and R. H. Rownd (1976). "Restriction endonuclease analysis of ribosomal DNA from Saccharomyces cerevisiae." Mol. Gen. Genet. 148: 233-241. D'Arpa, P., P. S. Machlin, H. Ratrie 3d, N. F. Rothfield, D. W. Cleveland and W. C. Earnshaw (1988). "cDNA cloning of human DNA topoisomerase I: catalytic activity of a 67.7-kDa carboxyl-terminal fragment." Proc. Natl. Acad. Sci. 85: 2543-2547. Delburck, M. (1954). "On the the replication of deoxyribonucleic acid (DNA)." Proc. Natl. Acad. Sci. 40: 783-788. Depew, R. E., L. F. Liu and J. C. Wang (1978). "Interaction between DNA and Escherichia coli protein omega. Formation of a complex between single-stranded DNA and omega protein." T.Biol. Chem. 253: 511-518. DiGate, R. J. and K. J. Marians (1988). "Identification of a potent decatenating enzyme from Escherichia coli." I Biol Chem 263: 13366-73. DiGate, R. J. and K. J. Marians (1989). "Molecular cloning and DNA sequence analysis of Escherichia coli topB, the gene encoding topoisomerase III." J. Biol. Chem. 264: 17924-17930. Dimri, G. P. and H. K. Das (1990). "Cloning and sequence analysis of gyrA gene of Klebsiella pneumoniae." Nuc. Acids Res. 18: 151-156. DiNardo, S., K. Voelkel and R. Sternglanz (1984). "DNA topoisomerase II mutant of Saccharomyces cerevisiae: topoisomerase II is required for segregation of daughter molecules at the termination of DNA replication." Proc. Natl. Acad. Sci. 81: 2616-2620. DiNardo, S., K. A. Voelkel, R. Sternglanz, A. E.Reynolds and A. Wright (1982). "Escherichia coli DNA topoisomerase I mutants have compensatory mutations in DNA gyrase genes." Cell 31: 43-51. Dresser, M. E. and C. N. Giroux (1988). "Meiotic chromosome behavior in spread preparations of Yeast." J. Cell Bio. 106: 567-573. Durban, E., M. Bramucci and R. Cook (1988). "Partial amino acid sequence of rat topoisomerase I: comparison with the predicted sequences for the human and yeast enzymes." Biochem. Biophys. Res. Commun. 154: 358-364. Dvorkin, N., M. W. Clark and B. A. Hamkalo (1991). "Ultrastructural localization of nucleic acid sequences in Saccharomyces cerevisiae nucleoli." Chromosoma 100: 519-523. Egyhazi, E.and E. Durban (1987). "Microinjection of anti-topoisomerase I immunoglobulin G into nuclei of Chironomus tentans salivary gland cells leads to blockage of transcription elongation." Mol. Cell. Biol. 7: 4308-4316. Elion, E. A. and J. R. Warner (1984). "The major promoter element of rRNA transcription in yeast lies 2 kb upstream." Cell 39: 663-673. Elion, E.A. and J. R. Warner (1986). "An RNA polymerase I enhancer in Saccharomyces cerevisiae." Mol. Cell. Biol. 6: 2089-2097. Fleischmann, G., G. Pflugfelder, E.K. Steiner, K. Javaherian, G. C. Howard, J. C. Wang and S. C. Elgin (1984). "DrosophilaDNA topoisomerase I is associated with transcriptionally active regions of the genome." Proc. Natl. Acad. Sci. 81: 6958-6962. Garg, L. C., S. DiAngelo and S. T. Jacob (1987). "Role of DNA topoisomerase I in the transcription of supercoiled rRNA gene." Proc. Natl. Acad. Sci. 84: 3185-3188. Gellert, M., K. Mizuuchi, M. H. O'Dea and H. A. Nash (1976). "DNA gyrase: An enzyme that introduces superhelical turns into DNA." Proc. Natl. Acad. Sci. 73: 3872-3876. Gellert, M., M. H. O'Dea, T. Itoh and J. I. Tomizawa (1976). "Novobiocin and coumermycin inhibit DNA supercoiling catalyzed by DNA gyrase." Proc. Natl. Acad. Sci. 73: 4474-4478. Georgiev, O. I., N. Nikolaev, A. A. Hadjiolov, K. G. Skryabin, V. M. Zakharyev and A. A. Bayev (1981). "The structure of the yeast ribosomal RNA genes. 4. Complete sequence of the 25 S rRNA gene from Saccharomyces cerevisae." Nuc. Acids Res. 9: 6953-6958. Giaever, G., R. Lynn, T. Goto and J.C. Wang (1986). "The complete nucleotide sequence of the structural gene TOP2 of yeast DNA topoisomerase II." L Biol. Chem. 261: 12448-12454. Giaever, G. N. and J. C. Wang (1988). "Supercoiling of intracellular DNA can occur in eukaryotic cells." Cell 55: 849-856. Gilmour, D. S. and S. C. Elgin (1987). "Association of topoisomerase I with transcriptionally active loci in Drosophila." NCI Monogr. 1987: 17-21. Gilmour, D. S. and S. C. Elgin (1987). "Localization of specific topoisomerase I interactions within the transcribed region of active heat shock genes by using the inhibitor camptothecin." Mol. Cell. Biol. 7: 141-148. Giroux, C. N., M. E.Dresser and H. F. Tiano (1989). "Genetic control of chromosome synapsis in yeast meiosis." Genome 31: 88-93. Goto, T. and J.C. Wang (1985). "Cloning of yeast TOP1, the gene encoding DNA topoisomerase I, and construction of mutants defective in both DNA topoisomerase I and DNA topoisomerase II." Proc. Natl. Acad. Sci. 82: 7178-7182. Hammond, G. G., P. J. Cassidy and K. M. Overbye (1991). "Novobiocindependent topA deletion mutants of Escherichia coli." J. Bacteriol. 173: 5564-5567. Heller, R. A., E. R. Shelton, V. Dietrich, S. C. Elgin and D. L. Brutlag (1986). "Multiple forms and cellular localization of Drosophila DNA topoisomerase II." J. Biol. Chem. 261: 8063-8069. Higashinakagawa, T., H. Wahn and R. H. Reeder (1977). "Isolation of Ribosomal Gene Chromatin." Dev. Biol. 55: 375-386. Holm, C., T. Goto, J. C. Wang and D. Botstein (1985). "DNA topoisomerase II is required at the time of mitosis in yeast." Cell 41: 553-563. Holmes, M. L. and M. Dyall-Smith (1991). "Mutations in DNA gyrase result in novobiocin resistance in halophilic archaebacteria." J. Bacteriol. 173: 642-648. Hopewell, R., M. Oram, R. Briesewitz and L. M. Fisher (1990). "DNA cloning and organization of the Staphylococcus aureus gyrA and gyrB genes: Close homology among gyrase proteins and implications for 4quinolone action and resistance." J. Bacteriol. 172: 3481-3484. Huang, W. M. (1986). "The 52-protein subunit of T4 DNA topoisomerase is homologous to the gyrA-protein of gyrase." Nuc. Acids Res. 14: 73797390. Huang, W. M. (1986). "Nucleotide sequence of a type II DNA topoisomerase gene. Bacteriophage T4 gene 39." Nuc. Acids Res. 14: 7751-7765. Jackson, J. A. and G. R. Fink (1981). "Gene conversion between duplicated genetic elements in yeast." Nature 292: 306-311. Jackson, J. A. and G. R. Fink (1985). "Meiotic recombination between duplicated genetic elements in Saccharomyces cerevisiae." Genetics 109: 303-332. Jemtland, R., E. Maehlum, O. S. Gabrielsen and T. B. Oyen (1986). "Regular distribution of length heterogeneities within non-transcribed spacer regions of cloned and genomic rDNA of Saccharomyces cerevisiae." Nuc. Acids Res. 14: 5145-5158. Jinks-Robertson, S. and T. D. Petes (1985). "High-frequency meiotic gene conversion between repeated genes on nonhomologous chromosomes in yeast." Proc. Natl. Acad. Sci. 82: 3350-3354. Jinks-Robertson, S. and T. D. Petes (1986). "Chromosomal translocations generated by high-frequency meiotic recombination between repeated yeast genes." Genetics 114: 731-752. Kato, J., Y. Nishimura, R. Imamura, H. Niki, S. Hiraga and H. Suzuki (1990). "New topoisomerase essential for chromosome segregation in E. coli." Cell 63: 393-404. Keil, R. L. and G. S. Roeder (1984). "Cis-acting, recombination-stimulating activity in a fragment of the ribosomal DNA of S. cerevisiae." Cell 39: 377-386. Keller, W. (1975). "Characterization of purified DNA-relaxing enzyme from human tissue culture cells." Proc. Natl. Acad. Sci. 72: 2250-2554. Kikuchi, A. and K. Asai (1984). "Reverse gyrase--a topoisomerase which introduces positive superhelical turns into DNA." Nature 309: 677-681. Kim, R. A. and J. C. Wang (1989). "Function of DNA topoisomerases as replication swivels in Saccharomyces cerevisiae." I. Mol. Biol. 208: 257267. Kim, R. A. and J. C. Wang (1989). "A subthreshold level of DNA topoisomerases leads to the excision of yeast rDNA as extrachromosomal rings." Cell 57: 975-985. Kirkegaard, K., G. Pflugfelder and J. C. Wang (1984). "The cleavage of DNA by type-I DNA topoisomerases." CSH Symp. Quant. Biol. 49: 411-419. Kirkegaard, K. and J. C. Wang (1985). "Bacterial DNA topoisomerase I can relax positively supercoiled DNA containing a single-stranded loop." L. Mol. Biol. 185: 625-637. Kouprina, N. Y. and V. L. Larionov (1983). "The study of a rDNA replicator in Saccharomyces." Curr. Genet. 7: 433-438. Kramer, R. A., P. Philippsen and R. W. Davis (1978). "Divergent transcription in the yeast ribosomal RNA coding region as shown by hybridization to separated strands and sequence analysis of cloned DNA." J.Mol. Biol. 123: 405-416. Krasnow, M. A. and N. R. Cozzarelli (1983). "Site-specific relaxation and recombination by the Tn3 resolvase: recognition of the DNA path between oriented res sites." Cell 32: 1313-1324. Krasnow, M. A., A. Stasiak, S. J. Spengler, F. Dean, T. Koller and N. R. Cozzarelli (1983). "Determination of the absolute handedness of knots and catenanes of DNA." Nature 304: 559-560. Kupiec, M. and T. D. Petes (1988). "Meiotic recombination between repeated transposable elements in Saccharomyces cerevisiae." Mol. Cell. Biol. 8: 2942-2954. Larionov, V. L., A. V. Grishin and M. N. Smirnov (1980). "3 micron DNA - an extrachromosomal ribosomal DNA in the yeast Saccharomyces cerevisiae." Gene 12: 41-49. Linskens, M. H. K. and J.A. Huberman (1988). "Organization of replication of ribosomal DNA in Saccharomyces cerevisiae." Mol. Cell. Biol. 8: 49274935. Liu, L. F. and J. C. Wang (1978). "Micrococcus luteus DNA gyrase: active components and a model for its supercoiling of DNA." Proc. Natl. Acad. Sci. 75: 2098-2102. Liu, L. F. and J.C. Wang (1987). "Supercoiling of the DNA template during transcription." Proc. Natl. Acad. Sci. 84: 7024-7027. Mankin, A. S., K. G. Skryabin and P. M. Rubtsov (1986). "Identification of ten additional nucleotides in the primary structure of yeast 18S rRNA." Gene 44: 143-143. Marini, J. C., K. G. Miller and P. T. Englund (1980). "Decatenation of kinetoplast DNA by topoisomerases." J.Biol. Chem. 255: 4976-4979. McMahon, M. E., D. Stamenkovich and T. D. Petes (1984). "Tandemly arranged variant 5S ribosomal RNA genes in the yeast Saccharomyces cerevisiae." Nuc. Acids Res. 12: 8001-8016. Menzel, R. and M. Gellert (1983). "Regulation of the genes for E. coli DNA gyrase: homeostatic control of DNA supercoiling." Cell 34: 105-113. Mestel, R., M. Yip, J. P. Holland, E. Wang, J. Kang and M. J. Holland (1989). "Sequences within the spacer region of yeast rRNA cistrons that stimulate 35S rRNA synthesis in vivo mediate RNA polymerase Idependent promoter and terminator activities." Mol. Cell. Biol. 9: 12431254. Mirambeau, G., M. Duguet and P. Forterre (1984). "ATP-dependent DNA topoisomerase from the archaebacterium Sulfolobus acidocaldarius. Relaxation of supercoiled DNA at high temperature." 1. Mol. Biol. 179: 559-563. Moriya, S., N. Ogasawara and H. Yoshikawa (1985). "Structure and function of the replication origin of the Bacillus subtilis chromosome. III. Nucleotide sequence of some 10,000 base pairs in the origin region." Nuc. Acids Res. 13: 2251-2265. Muller, M. T., W. P. Pfund, V. B. Mehta and D. K. Trask (1985). "Eukaryotic type I topoisomerase is enriched in the nucleolus and catalytically active on ribosomal DNA." EMBO J. 4: 1237-1243. Nash, H. A., K. Mizuuchi, L. W. Enquist and R. A. Weisberg (1981). "Strand exchange in lambda integrative recombination: genetics, biochemistry, and models." CSH Symp. Quant. Biol. 1: 417-428. Nazar, R. N. and W. M. Wong (1985). "Is the 5S RNA a primitive ribosomal RNA sequence?" Proc. Natl. Acad. Sci. 82: 5608-5611. Neigeborn, L. and J.R. Warner (1990). "Expression of yeast 5S RNA is independent of the rDNA enhancer region." Nuc. Acids Res. 18: 41794184. Ness, P. J., T. Koller and F. Thoma (1988). "Topoisomerase I cleavage sites identified and mapped in the chromatin of Dictyostelium ribosomal RNA genes." J. Mol. Biol. 200: 127-139. Nogi, Y., R. Yano and M. Nomura (1991). "Synthesis of large rRNAs by RNA polymerase II in mutants of Saccharomyces cerevisiae defective in RNA polymerase I." Proc. Natl. Acad. Sci. 88: 3962-3966. Nomura, M., E.A. Morgan and S. R. Jaskunas (1977). "Genetics of bacterial ribosomes." Annu. Rev. Genet. 11: 297-347. Olson, M. V., D. L. Montgomery, A. K. Hopper, G. S. Page, F. Horodyski and B. D. Hall (1977). "Nucleotide sequence of the yeast 5S ribosomal RNA gene and adjacent putative control regions." Nature 267: 641-643. Ostrowski, J., G. Burdzy-Jagura and N. M. Kredich (1987). "DNA sequences of the cysB regions of Salmonella typhimurium and Escherichia coli." L Biol. Chem. 262: 5999-6005. Ozenberger, B. A. and G. S. Roeder (1991). "A unique pathway of doublestrand break repair operates in tandemly repeated genes." Mol. Cell. Biol. 11: 1222-1231. Parales, R. E.and C. S. Harwood (1990). "Nucleotide sequence of the gyrB gene of Pseudomonas putida." Nuc. Acids Res. 18: 5880-5880. Pasion, S. G., J. C. Hines, R. Aebersold and D. S. Ray (1992). "Molecular cloning and expression of the gene encoding the kinetoplast-associated type II DNA topoisomerase of Crithidiafasciculata." Mol. Biochem. Parasitol. 50: 57-67. Petes, T. D. (1979). "Meiotic mapping of yeast ribosomal deoxyribonucleic acid on chromosome XII." J. Bact. 138: 185-192. Petes, T. D. (1979). "Yeast ribosomal DNA genes are located on chromosome XII." Proc. Natl. Acad. Sci. 76: 410-414. Petes, T. D. (1980). "Unequal meiotic recombination within tandem arrays of yeast ribosomal DNA genes." Cell 19: 765-774. Petes, T. D. and D. Botstein (1977). "Simple Mendelian inheritance of the reiterated ribosomal DNA of yeast." Proc. Natl. Acad. Sci. 74: 5091-5095. Petes, T. D. and U. S. Smolik (1979). "Evidence that the ribosomal DNA genes of yeast are not on chromosome I." Mol. Gen. Genet. 175: 187-193. Phillippsen, P., M. J. Thomas, R. A. Kramer and R. W. Davis (1978). "Unique arrangement of coding sequences for 5S, 5.8S, 18S and 25S ribosomal RNA in Saccharomyces cerevisiae as determined by R-loop and hybridization analysis." J. Mol. Biol. 123: 387-404. Piper, P. W., A. Lockheart and N. Patel (1984). "A minor class of 5S rRNA genes in Saccharomyces cerevisiae X2180-1B, one member of which lies adjacent to a Ty transposable element." Nuc. Acids Res. 12: 4083-4096. Pruss, G. J., S. H. Manes and K. Drlica (1982). "Escherichia coli DNA topoisomerase I mutants: increased supercoiling is corrected by mutations near gyrase genes." Cell 31: 35-42. Raji, A., D. J. Zabel, C. S. Laufer and R. E. Depew (1985). "Genetic analysis of mutations that compensate for loss of Escherichia coli DNA topoisomerase I." J. Bact. 162: 1173-1179. Retal, J. and R. J. Planta (1968). "The investigation of the ribosomal RNA sites in yeast DNA by hybridization technique." Biochim. Biophys. Acta. 169: 416-429. Riggs, D. L. and M. Nomura (1990). "Specific transcription of Saccharomyces cerevisiae 35 S rDNA by RNA polymerase I in vitro." J. Biol. Chem. 265: 7596-7603. Rose, D., W. Thomas and C. Holm (1990). "Segregation of recombined chromosomes in meiosis I requires DNA topoisomerase II." Cell 60: 1009-1017. Rothstein, R. (1984). "Double-strand-break repair, gene conversion, and postdivision segregation." CSH Symp. Quant. Biol. 49: 629-637. Rubtsov, P. M., M. M. Musakhanov, V. M. Zakharyev, A. S. Krayev, K. G. Skryabin and A. A. Bayev (1980). "The structure of the yeast ribosomal RNA genes. I. The complete nucleotide sequence of the 18S ribosomal RNA gene from Saccharomyces cerevisiae." Nuc. Acids Res. 8: 57795794. Saffer, L. D. and 0. L. J. Miller (1986). "Electron microscopic study of Saccharomyces cerevisiae rDNA chromatin replication." Mol. Cell. Biol. 6: 1148-1157. Schnos, M., K. Zahn, R. B. Inman and F. R. Blattner (1988). "Initiation protein induced helix destabilization at the X origin: A prepriming step in DNA replication." Cell 52: 385-395. Schwartz, D. C. and C. R. Cantor (1984). "Separation of yeast chromosomesized DNAs by pulsed field gradient gel electrophoresis." Cell 37: 67-75. Schweizer, E., C. MacKechnie and H. O. Halvorson (1969). "The redundency of ribosomal and transfer RNA genes in Saccharomyces cerevisiae." Mol. Biol. 40: 261-277. Shuman, S. and B. Moss (1987). "Identification of a vaccinia virus gene encoding a type I DNA topoisomerase." Proc. Natl. Acad. Sci. 84: 74787482. Skryabin, K. G., M. A. Eldarov, V. L. Larionov, A. A. Bayev, J. Klootwijk, V. C. de Regt, G.M. Veldman, R. J. Planta, O. I. Georgiev and A. A. Hadjiolov (1984). "Structure and function of the nontranscribed spacer regions of yeast rDNA." Nuc. Acids Res. 12: 2955-2968. Skryabin, K. G., A. S. Kraev, P. M. Rubtsov and A. A. Baev (1979). "Complete nucleotide sequence of the spacer region situated between the 18S and 5.8S RNAs of yeasts." Dokl. Biol. Sci. 247: 278-281. Skryabin, K. G., V. M. Zakhararev, P. M. Rubtsov and A. A. Baev (1979). "Nucleotide sequence of the presumed region of initiation of transcription of the yeast ribosomal operon." Dokl. Biochem. 247: 288290. Srivenugopal, K. S., D. Lockshon and D. R. Morris (1984). "Escherichia coli DNA topoisomerase III: purification and characterization of a new type I enzyme." Biochemistry 23: 1899-906. Stein, D. C., R. Danaher and T. M. Cook (1991). "Characterization of a gyrB mutation responsible for low level nalidixic acid resistance in Neisseria gonorrheae." Antimicrob. Agents Chemother. 35: 622-626. Stewart, A. F., R. E. Herrera and A. Nordheim (1990). "Rapid induction of cfos transcription reveals quantitative linkage of RNA polymerase II and DNA topoisomerase I enzyme activities." Cell 60: 141-149. Stewart, S. E. and G. S. Roeder (1989). "Transcription by RNA polymerase I stimulates mitotic recombination in Saccharomyces cerevisiae." Mol. Cell. Biol. 9: 3464-3472. Strauss, P. R. and J. C. Wang (1990). "The TOP2 gene of Trypanosoma brucei: a single-copy gene that shares extensive homology with other TOP2 genes encoding eukaryotic DNA topoisomerase II." Mol. Biochem. Parasitol. 38: 141-150. Swanberg, S. L. and J. C. Wang (1987). "Cloning and sequencing of the Escherichia coli gyrA gene coding for the A subunit of DNA gyrase." L Mol. Biol. 197: 729-736. Swanson, M. E. and M. J. Holland (1983). "RNA polymerase I-dependent selective transcription of yeast ribosomal DNA. Identification of a new cellular ribosomal RNA precursor." J. Biol. Chem. 258: 3242-3250. Swanson, M. E., M. Yip and M. J. Holland (1985). "Characterization of an RNA polymerase I-dependent promoter within the spacer region of yeast ribosomal cistrons." J. Biol. Chem. 260: 9905-9915. Szostak, J. W. and R. Wu (1979). "Insertion of a genetic marker into the ribosomal DNA of yeast." Plasmid 2: 536-554. Szostak, J. W. and R. Wu (1980). "Unequal crossing over in the ribosmal DNA of Saccharomyces cerevisiae." Nature 284: 426-430. Thiry, M., U. Scheer and G. Goessens (1991). "Localization of nucleolar chromatin by immunocytochemistry and in situ hybridization at the electron microscopic level." Electron Microsc. Rev. 4: 85-110. Thomas, B. J. and R. Rothstein (1989). "Elevated recombination rates in transcriptionally active DNA." Cell 56: 619-630. Thomsen, B., S. Mollerup, B. J. Bonven, R. Frank, H. Blocker, O. F. Nielsen and 0. Westergaard (1987). "Sequence specificity of DNA topoisomerase I in the presence and absence of camptothecin." EMBO J. 6: 1817-1823. Thrash, C., A. T. Bankier, B. G. Barrell and R. Sternglanz (1985). "Cloning, characterization, and sequence of the yeast DNA topoisomerase I gene." Proc. Natl. Acad. Sci. 82: 4374-4378. Tsai, P. M., L. F. Liu, A. A. Liu, K. M. Tewey, P. J. Whang, T. Knutsen, K. Huebner, C. M. Croce and J. C. Wang (1988). "Cloning and sequencing of cDNA encoding human DNA topoisomerase II and localization of the gene to chromosome region 17q21-22." Proc. Natl. Acad. Sci. 85: 7177-7181. Uemura, T., K. Morikawa and M. Yanagida (1986). "The nucleotide sequence of the fission yeast DNA topoisomerase II gene: structural and functional relationships to other DNA topoisomerases." EMBO T.5: 2355-2361. Uemura, T., K. Morino, S. Uzawa, K. Shiozaki and M. Yanagida (1987). "Cloning and sequencing of Schizosaccharomyces pombe DNA topoisomerase I gene, and effect of gene disruption." Nuc. Acids Res. 15: 9727-9739. Upton, C., A. Opgenorth, P. Traktman and G. McFadden (1990). "Identification and DNA sequence of the Shope fibroma virus DNA topoisomerase gene." Virology 176: 439-447. Valenzuela, P., G. I. Bell, A. Venegas, E. T. Sewell, F. R. Masiarz, L. J. Degennaro, F. Weinberg and W. J. Rutter (1977). "Ribosomal RNA genes of Saccharomyces cerevisiae: II. Physical map and nucleotide sequence of the 5S ribosomal RNA gene and adjacent intergenic regions." J. Biol. Chem. 252: 8126-8135. Veldman, G. M., R. C. Brand, J. Klootwijk and R. J. Planta (1980). "Some characteristics of processing sites in ribosomal RNA of yeast." Nuc. Acids Res. 8: 2907-2920. Veldman, G. M., J. Klootwijk, P. de Jonge, R. J. Leer and R. J. Planta (1980). "The transcription termination site of the ribosomal RNA operon in yeast." Nuc. Acids Res. 8: 5179-5192. Vincent, A. and T. D. Petes (1986). "Isolation and characterization of a Ty element inserted into ribosomal DNA of the yeast Saccharomyces cerevisiae." Nuc. Acids Res. 14: 2939-2949. Voelkel-Meiman, K., R. L. Keil and G. S. Roeder (1987). "Recombinationstimulating sequences in yeast ribosomal DNA correspond to sequences regulating transcription by RNA polymerase I." Cell 48: 10711079. Volkert, F. C. and J. R. Broach (1986). "Site-specific recombination promotes plasmid amplification in yeast." Cell 46: 541-550. Wallis, J. W., G. Chrebet, G. Brodsky, M. Rolfe and R. Rothstein (1989). "A hyper-recombination mutation in S. cerevisiae identifies a novel eukaryotic topoisomerase." Cell 58: 409-419. Wang, J. C. (1971). "Interaction between DNA and an Escherichia coli protein omega." J. Mol. Biol. 55: 523-533. Wang, J. C. and K. Becherer (1983). "Cloning of the gene topA encoding for DNA topoisomerase I and the physical mapping of the cysB-topA-trp region of Escherichia coli." Nuc. Acids Res. 11: 1773-1790. Wang, J. C. and K. Kirkegaard (1981). "DNA topoisomerases." Gene Amplification & Analysis 2: 455-473. Watson, J. D. and F. H. C. Crick (1953). "Genetic implications of the structure of deoxyribonucleic acid." Nature 171: 964-967. Wright, S. J. and G. Schatten (1990). "Teniposide, a topoisomerase II inhibitor, prevents chromosome condensation and separation but not decondensation in fertilized surf clam (Spisula solidissima) oocytes." Dev. Biol. 142: 224-232. Wyckoff, E. and T. S. Hsieh (1988). "Functional expression of a Drosophila gene in yeast: genetic complementation of DNA topoisomerase II." Proc. Natl. Acad. Sci. 85: 6272-6276. Wyckoff, E., D. Natalie, J. M. Nolan, M. Lee and T. Hsieh (1989). "Structure of the Drosophila DNA topoisomerase IIgene. Nucleotide sequence and homology among topoisomerases II." J. Mol. Biol. 205: 1-13. Yamagishi, M. and M. Nomura (1988). "Deficiency in both type I and type II DNA topoisomerase activities differentially affect rRNA and ribosomal protein synthesis in Schizosaccharomyces pombe." Curr. Genet. 13: 305314. Zamb, T. J. and T. D. Petes (1982). "Analysis of the junction between ribosomal RNA genes and single-copy chromosomal sequences in the yeast Saccharomyces cerevisiae." Cell 28: 355-364. Chapter II Mitotic Recombination in the rDNA of S. cerevisiae is suppressed by the combined action of DNA topoisomerase I and II Cell, Vol. 55, 413-425, November 4, 1988, Copyright @ 1988 by Cell Press Mitotic Recombination in the rDNA of S. cerevisiae Is Suppressed by the Combined Action of DNA Topoisomerases I and II Michael F. Christman, Fred S. Dietrich, and Gerald R. Fink Whitehead Institute for Biomedical Research Nine Cambridge Center Cambridge, Massachusetts 02142 and Department of Biology Massachusetts Institute of Technology Cambridge, Massachusetts 02139 Summary We have found that mitotic recombination within the S. cerevisiae rDNA cluster (200 tandemly repeated 9.1 kb units) is strongly suppressed and that this suppression requires the combined action of DNA topoisomerases I and II. Strains with a null mutation in the TOP1 gene (encoding topoisomerase I) or a ts mutation in the TOP2 gene (encoding topoisomerase II) grown at a semipermissive temperature show 50- to 200-fold higher frequencies of mitotic recombination in rDNA relative to TOP + controls. Suppression of recombination is specific to the rDNA because the recombination frequency at another tandem array, the CUP1 locus, at a simple HIS4 duplication, or among dispersed repeats (MAT and HML or HMR) is not elevated in top1 or top2 mutants. The high frequency of mitotic recombination within the rDNA cluster in topoisomerase mutants shows that both TOP1 and TOP2 are required for suppression of recombination in this region of the genome. et al., 1969). If the meiotic recombination frequency in the rDNA cistrons were similar to that in other chromosomal regions, the distance from one end of the cluster to the other would be on the order of 800 map units. However, it has been estimated to be about 3 cM (Zamb and Petes, 1982). Two types of experiments have been used to measure the recombination frequency in the rDNA at meiosis. In one set of experiments two strains whose rDNA cistrons differ by a restriction site polymorphism were crossed and the progeny were examined for recombinants (Petes and Botstein, 1977). The frequency of recombination was less than 1% of that expected based on the number of recombination events per genome and the fraction of the genome that is rDNA. An independent measure of the frequency of meiotic recombination using markers at either end of the rDNA array (Zamb and Petes, 1982) indicated that the recombination frequency was at least 15-fold lower than expected. Both of these experiments suggested that meiotic recombination is suppressed in rDNA cistrons. Previous studies estimated the frequency of mitotic recombination inthe rDNA by following the loss of an indicator marker that has been introduced into the rDNA array on chromosome XII. The loss is presumed to result from mitotic recombination between the homologous rDNA regions flanking the marker. A marker in the rDNA is typically lost at a rate of roughly 5 x 10- 4 per cell division (Szostak and Wu, 1980). In this report we show that this level of mitotic recombination in rDNA is a result of a specific recombination suppression and that both topoisomerase I and topoisomerase II are required for the suppression. Introduction Results The homologous recombination system of the yeast Saccharomyces cerevisiae is extremely active in both meiosis and mitosis. The frequency of recombination at meiosis established for many chromosomal regions is about 1 map unit per 2 kb, or about 100 times greater than that found for human meiosis. The role of the homologous recombination system in controlling genome size is evident from studies on artificial duplications constructed by transformation. Transformation of yeast cells with a circular plasmid containing a yeast gene for which there is homology on the chromosome results in a duplication of that cloned segment (Hinnen et al., 1978). These duplications are unstable in both meiosis and mitosis. Loss of one of the copies occurs by a homologous recombination event that leaves a single copy of the gene on the chromosome (Hicks et al., 1979). The absence of substantial repeated DNA in yeast has been ascribed to the activity of this homologous recombination system. Despite the presence of this active recombination system, yeast cells are able to maintain the highly reiterated cluster of ribosomal DNA genes (rDNA) on chromosome XII. The 9.1 kb rDNA cistron encoding the ribosomal RNAs is tandemly repeated approximately 200 times (Schweizer Topoisomerase I Is Required for Suppression of Mitotic Recombination in rDNA The frequency of mitotic recombination in rDNA can be monitored by following the frequency of loss of a marker gene that has been inserted into the rDNA cluster by transformation. A marker gene in the rDNA array can be lost by any one of a variety of recombination events between rDNA repeats: unequal sister chromatid exchange, intrachromosomal excision, or gene conversion (either inter- or intrachromatid). Because loss of the marker gene will result from only a fraction of these recombination events (many events may not involve the particular rDNA repeat in which the marker resides), the frequency of loss of Ura + is a minimum estimate of the overall frequency of mitotic recombination in rDNA. A MATa strain containing a URA3 insertion in the rDNA (CY163) was crossed by a MATa strain containing the same URA3 insertion and a topl-8::LEU2 mutation (CY205, cross 50). The frequency of loss of Ura + (mitotic recombination in rDNA) was measured for the TOP1 and topl::LEU2 spores by scoring the frequency of 5-fluoroorotic acid (5FOA) resistance. In S. cerevisiae, URA3 cells Topoisomerases Suppress Mitotic rDNA Recombination 415 Table 2. Frequency and Rate of Mitotic Recombination in rDNA in TOP1 and topi Strains Strain Genotypea Frequency of Ura-3 Rate of Ura- CY117 CY119 TOPI rDNA::URA3-1 topl-7::LEU2 rDNA::URA3-1 1.5 x 10 (1) 2.1 x 10- 1 (140) 3.5 x 10- 4 (1) 1.9 x 10-2 (54) CY118 CY120 TOP1 rDNA::URA3-2 topl-7::LEU2 rDNA::URA3-2 1.6 x 10- 3 (1) 3.2 x 10-1 (200) 2.7 x 10- 4 (1) 1.4 x 10-2 (53) The frequency and rate of loss of Ura+ were determined as described in the text. Numbers in parentheses represent the fold increase in a top1 background as compared with TOP1. Each frequency represents the average of ten independent measurements. Rates were determined from the Ura+lUra - sectoring of at least 5000 colonies in each case. aOnly the relevant genotype for this experiment is shown for each strain. See Table 1 for complete genotypes. To be sure that the enhancement of URA3 loss in topi mutants was not a consequence of the presence of 5FOA, we also measured the frequency of segregation of Ura + to Ura- in these strains in the absence of 5FOA. Strains were first streaked selectively on complete medium lacking uracil. From this medium, independent Ura + colonies were then streaked nonselectively on complete medium (containing uracil) to permit loss of URA3 from the rDNA during mitotic growth of the colony. Colonies arising on the nonselective plates were then examined for the fraction of Ura + and Ura- cells in that colony. This type of quantitative analysis for both CY117/CY119 and CY118/ CY120 pairs is presented inTable 2. Ten independent colonies were examined from each isogenic pair for the fraction of Ura- cells, and the average frequency for each set of ten was used to compare the frequencies of mitotic rDNA recombination. The topi mutant strains show 140and 200-fold higher frequencies of mitotic recombination in rDNA compared with the isogenic TOP1 strains. Southern analysis of four independent Ura- clones derived from CY117 and CY119 and four from CY118 and CY120 indicate that in each case the lacZ DNA sequences present in CB32 were also lost from the rDNA array (data not shown). The co-loss of URA3 and lacZ excludes the possibility that the recombinant strains became Ura- by virtue of a gene conversion event between the URA3 gene in the rDNA and the ura3-52 allele at the resident URA3 locus, and shows that loss of Ura+ results from recombination between flanking rDNA sequences. The recombination rate in rDNA in topi mutants is so high that conventional methods of fluctuation analysis are difficult. We took advantage of this high rate to devise a simple fluctuation analysis of individual colonies that allows us to determine the fraction of cells that have not undergone a recombination event in the rDNA (i.e., the P(0) term of the Poisson; Luria and Delbrtck, 1943). Strains were grown overnight in complete medium lacking uracil to select for cells that retain the URA3 gene. Cells were diluted and plated for single colonies on rich medium (YPD). The colonies that grew on YPD were then printed to complete medium lacking uracil. Each colony was then examined for the presence of a Ura- sector. A colony that was one-half Ura ÷ and one-half Ura- was assumed to have incurred a recombination event in the rDNA at the first mitotic division in the growth of that colony. Unsectored colonies or colonies that were only partially sectored (less than one-half Ura-) were assumed to have had no recombination event at the first mitotic division in the growth of that colony. By determining which colonies had an event at the first division and which did not, we could determine the rate of Ura- segregants from the P(0) term of the Poisson. The data, presented in Table 2, show that top1 mutants have 53- to 54-fold higher rates of mitotic recombination in rDNA compared with their otherwise isogenic parents. Mitotic recombination in rDNA was measured by a third method that did not require an insertion in rDNA to signal the recombination event. Diploid strains were constructed that are heterozygous for an EcoRI restriction site polymorphism in rDNA. Most yeast strains contain seven EcoRI sites per rDNA cistron (form I strains), but some contain only six EcoRI sites per rDNA cistron (form II strains; Petes et al., 1978). If such a diploid were to undergo a recombination event at the four-strand stage in mitosis (and most mitotic recombination events in rDNA occur at the four-strand stage, since the primary mitotic event is sister chromatid exchange [Szostak and Wu, 1980]), then subsequent cell division would frequently produce diploid progeny that contained an unequal mixture of the two rDNA forms (Figure 2A). When such a recombinant was induced to undergo meiosis, four spores would be produced, two spores each with a mixture of forms I and II and two spores each with only form I(or with only form II; see Figure 2A). This analysis assumes that no recombination in rDNA occurs during meiosis (see below). Because recombination can take place anywhere within the rDNA array, an enormous variety of different form i/form II mixtures is possible for the four meiotic spores derived from a diploid that undergoes recombination at the four-strand stage of mitosis. However diverse these events may be, all mitotic events have one characteristic pattern: they generate only two patterns of form 11I mixtures among the four meiotic spores. By contrast, meiotic recombination events in rDNA could yield tetrads in which either three of the four spores or all four of the spores have different form I/I1patterns (Figure 2B). We have constructed form 1/11 diploids where the MATa parents are the same and the MATa parents differ by only the disruption of the TOPi gene (CY159, CY160, CY161, CY162; Table 1). The segregation of form I and form II rDNAs was analyzed by Southern analysis for 32 fourspored tetrads from CY159 and CY161 (TOP1/topl) and 26 Topoisomerases Suppress Mitotic rDNA Recombination 417 TOP 1/top1 a b c d topl/topi a b c Table 3. Frequency of Mitotic Recombination in rDNA in top2-1 (ts) and TOP2 Strains d ale M ---.. 40 - -`)- me 0 ý04M 404 -- q a 9 10 Frequency of UraSpores 0 0 24 C 30 C top2-1 (ts) 39-60 39-3D 39-18B 10101-14B 10102-78 10101-20 TOP2 1.7 5.1 1.3 9.7 3.8 1.7 x x x x x x 10- 3 10- 4 10- 3 10- 4 10- 3 10- 3 (1) (1) (1) (1) (1) (1) 4.0 2.3 3.2 6.7 1.7 1.0 x x x x x x 10-1 10-' 10-2 10-2 10-1 10-1 39-8D 39-4A 10101-1A 10102-8C 10101-4C 6.7 7.2 4.0 2.9 2.5 x x x x x 10- 4 10- 4 10- 3 3 1010- 3 (1) (1) (1) (1) (1) 6.8 2.9 1.2 1.6 3.8 x x x x x 10- 4 (1) 10- 3 (4) 10- 3 (0.3) 10- 3 (0.6) 10-4(0.2) (235) (451) (25) (69) (45) (59) Each frequency is the average of four independent measurements of the frequency of Ura- segregants. Numbers in parentheses represent the fold increase at 300C relative to 24 0C. 05 Figure 4. Southern Analysis of Tetrads to Follow the Segregation of Form IIForm IIrDNA Polymorphisms in Mitosis and Meiosis in toplltopl Backgrounds and TOP1/70TOP1 The bands marked X', B,and C in panel I correspond to the EcoRI fragments shown schematically in Figure 3. Panels 1-5 are tetrads from TOPIlTOP1 diploids, and panels 6-10 are tetrads from toplltopl diploids. Panels 1 and 2 are tetrads from CY161; panels 3, 4, and 5 are tetrads from CY159; panels 6, 7, and 8 are tetrads from CY160; and panels 8, 9, and 10 are tetrads from CY162. In each panel the probe was the 2.7 kb Hindlll fragment of yeast rDNA (see Figure 3). the same frequencies of mitotic recombination inrDNA at 240C as the five wild-type spores, as shown in Table 3. However, at 300C the top2-1 (ts)-containing spores have greatly ir,creased frequencies of mitotic recombination in rDNA relative to the TOP2 spores. The recombination frequency ,n rDNA in a top1 top2-1 (ts) double mutant is about the same as that in either topl or top2-1 (ts) single mutants (data not shown), although inall cases the recombination frequency is near the theoretical limit of 1.The interpretation of the results is not as clear as with the topi disruption because the top2-1 (ts) mutation affects viability (50%-80% of the cells in colonies grown at 30PC fail to form colonies on rich medium at 240C). However, the top21 (ts) mutation does not cause a general increase in all types of recombination events at 300C (see subsequent section). Mitotic Recombination at Other Loci Is Unaffected in top1 or top2 Mutants CUP1 Repeat The CUP1 locus in S. cerevisae encodes a metallothionein-like protein that mediates copper resistance (Fogel et al., 1983). Strains containing a single copy of CUP1 are sensitive to killing by copper, whereas strains containing ten tandemly repeated copies of CUPI are copper resistant. Most laboratory yeast strains contain about ten directly repeated copies of CUP1 on the left arm of chromosome VIII (Fogel and Welch, 1982). We inserted a URA3 marker into the CUP1 region of a strain (CY72) that has 10-12 tandemly repeated CUP1 genes (Southern analysis not shown) to monitor the frequency of mitotic recombination. Four independent URA3 insertions were generated at CUP1 by transformation of CY72 using plasmid FDp110. Southern analysis showed that each of the insertions was at a different site within the CUP1 repeat (data not shown). The strains carrying CUP1::URA3 were subsequently transformed with Hindllldigested CB25 to make disruptions of the TOP1 gene marked with LEU2. Each of the four isogenic pairs of TOP1 and topl insertions was tested for the loss of URA3 from the CUP1 repeat by assaying the frequency of 5FOAresistant colonies. As shown inTable 4, there is no difference inthe frequency of 5FOA-resistant colonies inTOP1 versus topl strains for any of the four different CUP1::URA3 insertions. One of the CUP1::URA3 insertion strains (FDY64) was crossed to a top2-1 strain (CY206, cross F88-24), and the appropriate spores were tested for the frequency of 5FOA resistance at both 240C and 300C. As shown inTable 5, the three different top2-1 (ts) spores tested at both 240C or 300C show no difference from TOP2 spores in the frequency of 5FOA-resistant segregants. Simple Duplications A simple duplication of the HIS4 region was constructed by transforming strain CY72 to Ura ÷ with plasmid CB30, which carries both HIS4 and URA3. The plasmid was first digested with Bglll inorder to direct integration to the HIS4 locus. Southern analysis of several transformants showed that the chromosomal arrangement in the transformants was a HIS4 duplication flanking a single copy of the URA3 gene (data not shown). One of these transformants, desig- Topoisomerases Suppress Mitotic rDNA Recombination 419 nated CY125 (Table 1), was then transformed to Leu + with the 5.0 kb Hindlll fragment of CB25 to disrupt the TOP1 gene with LEU2. Southern blots confirmed the structure of several transformants (data not shown). One of these, CY126 (HIS4::URA3::HIS4 topl), was compared with the isogenic CY125 (HIS4::URA3::HIS4 TOPI) for the frequency of loss of URA3 (5FOA resistance). As shown in Table 4, the frequency of 5FOA resistance is approximately the same for both CY125 (TOPi) and CY126 (topl::LEU2). A strain similar to CY125 was constructed (CY127, see Experimental Procedures) except that each of the duplicated HIS4 copies contained a 20 nucleotide oligomer representing the yeast version of the highly conserved topoisomerase I binding site from yeast rDNA. This experiment was designed to test whether the synthetic topoisomerase I binding site from rDNA was sufficient to cause topoisomerase I-mediated suppression of recombination. An isogenic topi derivative of CY127 (designated CY128) was constructed by transformation (see Experimental Procedures). As shown in Table 4, there is no difference in the frequency of 5FOA resistance between CY125 and CY127, and, therefore, no effect of top1 deficiency on recombination between these repeats. Southern analysis of four independent 5FOA-resistant derivatives of CY127 and four from CY128 demonstrate that the URA3 gene is missing from HIS4 in the recombinants and not simply gene-converted to Ura- from the resident ura3-52 allele (data not shown). Strains containing the simple HIS4 duplication and top2-1 (ts) (generated by crossing CY125 and CY206, cross F88-21) were assayed for their frequency of 5FOA resistance at both 240C and 300C. As shown in Table 5, there is no difference between top2-1 and wild-type strains in the frequency to 5FOA resistance at either 240C or 300C. The frequency of 5FOA resistance in three different top2-1 (ts) spores is essentially the same at both 240C and 300C, as shown in Table 5. MAT Switching Mitotic recombination between the silent mating type cassettes (HMR and HML) and the active cassette (MAT) is suppressed (Klar et al., 1981). Although the suppression is known to involve the SIR genes, the mechanism is unclear. We tested the ability of otherwise isogenic TOP1 (CY72 and CY66) and topi mutant strains (CY112 and CY64) to switch mating types spontaneously (Table 4). The frequency of switching is the same for both pairs of strains, indicating no role for topoisomerase I in suppression of mitotic recombination events between the repeated mating type cassettes. We have also tested several top2-1 strains for the ability to switch mating types spontaneously at both 240C and 300C. As shown in Table 5, there is no difference in either MATa-to-MATd or MATa-to-MATa switching frequency at either 240C or 300C in top2-1 versus wild-type strains. Mitotic Recombination in rDNA Is Not Dependent on RAD52 The RAD52 gene product is required for mitotic recombination events involving gene conversion (Jackson and Table 6. Frequency of Mitotic Recombination in rDNA in rad52 strains Strain Genotype Frequency of Ura- CY210 CY211 CY212 CY213 RAD52 TOP1 topl-7::LEU2 rad52::TRP1 rad52::TRP1 topl-7::LEU2 3 x 10 (1) 2 x 10-' (67) 8.5 x 10- 3 (2.8) 4.2 x 10-2 (14) 3 Numbers in parentheses represent the fold increase relative to the frequency in a RAD52 TOP1 background. Each frequency is the average of ten independent measurements. Fink, 1981). Aset of strains have been constructed (CY210, CY211, CY212, and CY213; see Experimental Procedures) consisting of otherwise isogenic pairs of RAD52 and rad52::TRP1 in both TOP1 and topl::LEU2 genetic backgrounds. Mitotic recombination in rDNA was measured in these strains as described above, and the results are presented in Table 6. A disruption of RAD52 does not decrease rDNA recombination in TOP1 strains. topi rad52 strains have an apparent 4.7-fold decrease in the frequency of rDNA recombination as compared with top1 RAD52. The interpretation of this result should be tempered by the observation that top1l rad52 strains grow much more slowly than either topi or rad52 single-mutant strains. Meiotic Recombination in rDNA in toplltopl Mutants Using strains polymorphic for an EcoRI restriction site in the rDNA cistron, Petes and Botstein (1977) showed that meiotic recombination in the rDNA cistrons is suppressed 300-fold relative to the rest of the nuclear genome. They observed no meiotic recombination in 12 tetrads despite the fact that the rDNA array is the size of a typical yeast chromosome and, therefore, should show an average of five meiotic recombination events per meiosis. We have constructed diploids that are heterozygous for an EcoRI site polymorphism in rDNA and are either TOPlltopl (CY159, CY161) or toplltopl (CY160, CY162; see Experimental Procedures and Table 1). We were unable to determine whether topoisomerase II is required for suppression of rDNA recombination during meiosis because recent data indicate that topoisomerase II is required for the first meiotic division (Connie Holm, personal communication). The toplltopl diploids were induced to sporulate, and their meiotic progeny were examined by Southern analysis to monitor the segregation of the rDNA polymorphism. A meiotic event is characterized by either three or four different form 1/11 patterns among the four meiotic progeny (see Figure 28). In the homozygous toplitopl mutants (CY160 and CY162), seven of the 32 tetrads examined gave meiotic patterns of this type (Figure 4, panels 6 and 7). In each of the seven tetrads that display meiotic recombination events, the event appears to have been nonreciprocal. For example, in Figure 4, panel 7, spore d appears to be a form I spore that has acquired-some form II rDNA. However, neither of the form IIspores (a or b)has acquired any additional form I DNA or appears to have lost Topoisomerases Suppress Mitotic rDNA Recombination 421 (Petes, 1980). The frequencies of these events in TOP11 TOP1 (15%, 10/65) and toplltopl (6%, 6/98) strains issimilar to that reported previously (10%; Petes, 1980). Reciprocal meiotic crossovers also occur at equal frequencies in TOP1lTOP1 and toplltopl diploids. Reciprocal recombination is highly repressed in rDNA relative to the rest of the nuclear genome. Petes and Botstein (1977) did not observe a reciprocal event in 12 tetrads using the EcoRI restriction polymorphism as a marker. Our data show that reciprocal recombination occurs in both TOP11 TOP1 and toplltopl backgrounds at a frequency of 8% (5/65) and 5% (5/98), respectively. This level of recombination is still low because a segment of DNA the size of the rDNA cistrons should have an average of five crossover events per meiosis. We observed four gene conversion events in the topll topi diploid and none in the TOPI/TOP1 diploid. We also observed an apparent increase in gene conversions in toplitopl strains using the EcoRI restriction polymorphism (see above). Clearly, more tetrad data are required to determine whether a deficiency of topoisomerase I causes any significant difference in meiotic gene conversion frequency. In agreement with the results from analysis of haploid cells and the form I/form II polymorphism, the toplltopl strain (relative to the TOPIlTOP1 control) shows a great increase in mitotic recombination events in rDNA preceeding meiosis. Most of the patterns that are presumed to be mitotic could also arise via double meiotic events. However, the absence of a large number of single meiotic events argues that these patterns represent mitotic events. We have noticed a slight growth defect that segregates 2:2 in 16 tetrads from CY182 (toplltopl). In each tetrad, the two slow-growing colonies have the same rDNA genotype as determined by the indicator genes (that is, the two slow growers are both HIS3 or both URA3). This result can be explained if the growth defect occurs via an event in mitosis that is linked to the rDNA cistrons. Since no growth defect is generated in TOP1ITOP1 diploids (CY180), we ascribe the slow-growth phenotype to the enhanced rDNA mitotic recombination in toplltopl cells. Perhaps the loss of amplification resulting from rDNA recombination is the cause of the slower growth rate. Discussion Yeast strains defective in either topoisomerase I or topoisomerase II activity (topi and top2-1 (ts) mutants) have much higher levels of mitotic recombination in the rDNA than do TOP + controls. Mitotic recombination in other duplicated regions of the genome is not affected by top1 or top2-1 (ts). We interpret this enhancement of rDNA recombination in top1 and top2 strains to mean that one function of topoisomerases in yeast is to suppress recombination specifically inthe rDNA. A large number of genes have been identified in yeast that are involved in the recombination process (Orr-Weaver and Szostak, 1985); however, most of these affect recombination throughout the genome, whereas the effects of the topi and top2 mutations appear to be restricted to the rDNA. The existence of a system to suppress mitotic recombination in the rDNA is somewhat unexpected because mitotic recombination is usually considered a rare event. However, in the absence of topoisomerase I or II,the frequency of rDNA recombination jumps as high as one recombinant in every three cells. At these high frequencies, successive amplification and deletion of the rDNA array by recombination could be deleterious to growth. Although we have no direct evidence for deletion or amplification, we have observed a growth defect that could result from an alteration in the number of rDNA cistrons. Tetrad analysis of toplltopl diploids revealed a growth defect linked to the rDNA cistrons. This slow growth could be a consequence of the increased frequency of deletion or amplification of the rDNA array in the toplltopl background. The slow-growing colonies gave rise spontaneously to faster-growing cells at high frequency, making analysis difficult. Presumably, repeated cycles of unequal sister chromatid exchange could give rise to fastergrowing segregants by restoring the array to its original size (Szostak and Wu, 1980). Since diploids as well as haploids require topoisomerase function for suppression of rDNA recombination, any defect in growth because of recombination has an added consequence for meiosis. Since each mitotic diploid is developmentally capable of undergoing meiosis, deleterious alterations of the rDNA can be transmitted to meiotic progeny. It is known that meiotic recombination in the rDNA is strongly suppressed relative to the rest of the nuclear genome in both yeast and Neurospora (Petes and Botstein, 1977; Russell et al., 1988). Our results indicate that tcpoisomerase I is not required for this suppression in yeast. Although the rDNA array is the size of a typical yeast chromosome (1000 kb) and should have about five crossovers per meiosis, homozygous toplltopl mutants gave only one crossover in 20 meioses as assayed with rDNA marked with URA3 and HIS3. Furthermore, no reciprocal crossovers were obtained in 32 tetrads from topl/topl diploids heterozygous for an rDNA restriction site polymorphism. We conclude that an alternative system is required for suppressing meiotic reciprocal recombination in the rDNA. Although the data are not conclusive, our results indicate a possible differential effect of topoisomerase Ion meiotic gene conversion as compared with reciprocal recombination. Using the rDNA polymorphism, we detected 7/32 tetrads with conversions in topl/topl mutants as compared with 0/26 in TOP1/topl strains. And using the URA31HIS3 markers, we detected 4/98 tetrads with conversions in toplltopl versus 0/65 in TOPI/TOP1 diploids. The overall interpretation of the top2 results is not as straightforward as that for topi. For example, we could not determine the meiotic effect of the top2 mutation, because top2/top2 diploids fail to undergo normal meiosis. Moreover, it is not possible to study the effects of a null allele of TOP2 (as was done for TOP1) because TOP2 is required for life. Although the top2-1 (ts) allele that we used is deficient in topoisomerase II activity at the semipermissive Topoisomerases Suppress Mitotic rDNA Recombination 423 suppression of recombination between the directly repeated termini of the Ty retrotransposon (Rothstein, 1984). The mechanism by which the SIR genes and the EDR1 gene suppress recombination is not known. However, our data show that neither topoisomerase I nor topoisomerase IIis required for recombination suppression between the mating type cassettes, and recent experiments show that they are not required for delta-delta recombination (unpublished observations). In many organisms meiotic recombination is suppressed incentromeric regions. InS.cerevisiae there is some evidence for suppression of recombination in centromeric regions (Lambie and Roeder, 1988). Perhaps this suppression is mediated by a topoisomerase activity that is among the ensemble of proteins that constitute the centromere. Experimental Procedures Strain Constructions CY119 and CY120 These strains are single-step disruptions of the TOP1 gene marked by LEU2. Plasmid CB25 was digested with Hindill and then used to transform CY117 and CY118 to Leu + . The large Hindill fragment of CB25 contains the entire TOP1 coding region from which 849 internal nucleotides have been deleted and into which the LEU2 gene was inserted (Thrash et al., 1985). CY159, CY160, CY161, and CY162 CY159 and CY160 are diploids that are heterozygous for an EcoRi restriction site polymorphism in rDNA. The MATa parents are the same for the two strains (CY167), and the MATa parents (derived from CY72) are isogenic except that the TOP1 gene has been disrupted using CB25 in the case of CY160. CY159 and CY160 contain the same URA3 insertion in the rDNA. CY161 and CY162 have the same MATa form II parent (CY168) and MATa parents that are isogenic except for a top1 disruption (CY64 for CY162 and CY66 for CY161). CY161 and CY162 contain no foreign insertions in the rDNA. CY125, CY126, CY127, and CY128 + CY125 was made by transformation of CY72 to Ura with CB30 (which contains an internal 2.2 kb Xbal-Xhol fragment of the HIS4 gene cloned into YIP5; see Donahue et al. 119821 for a restriction map). The TOP1 gene was disrupted in CY125 using CB25 digested with Hindlli to generate CY126. CY127 was made in two steps, as follows: First, CY72 was cotransformed with YEp13 and CB26 (which had been digested with Xbal and Xhol). CB26 contains an internal 2.2 kb Xbal-Xhol fragment from the HIS4 gene into which a single 20 nucleotide oligomer containing the conserved topoisomerase I binding site from yeast rDNA had been cloned. The sequence of the oligonucleotide was 5S'CGATTTCTTTCTAAGTGGAT-3'. The complementary oligonucleotide was made such that, upon annealing, the double-stranded molecule contains a 2 nucleotide overhang at each 5' end, forming Clal-compatible ends. Leu + transformants were screened for His-. + Two His- colonies were found among 2300 Leu transformants. These were grown nonselectively in order to lose the cotransforming Leu+ plasmid. Both of the His- transformants had polar his4A mutations as determined by complementation. This is expected, since the oligonucleotide was cloned into the Clal site in the HIS4A region. One of these was designated CY113. In the second step, CY113 was transformed to Ura+ with CB31. CB31 contains the Xbal-Xhol fragment from CB26 + cloned into YIP5. Therefore, Ura transformants arise via duplication of the his4 region flanking the URA3 gene. Each his4 copy contains a topoisomerase I rDNA olionucleotide at the Clal site. CY128 was de+ rived from CY127 by tranformation to Leu using CB25 that had been digested with Hindill such that the TOP1 gene was disrupted and marked by LEU2. CY210, CY211, CY212, CY213 CY210 is a TOP1 spore derived from cross 39. It contains a single URA3 insertion in the rDNA. CY211 is isogenic to CY210 except for the disruption of the RAD52 locus. The disruption was made using the 1.97 kb BamHI fragment from CB15, which contains the TRP1 marker. CY212 is a topl-8::LEU2 spore from cross 39. CY213 is isogenic to CY212 except for the same rad52::TRP1 disruption. FDY62-FDY69 CY72 was transformed to Ura+ with Kpnl-digested FDp110. Four independent transformants were designated FDY62, FDY64, FDY66, and FDY68. The URA3 marker was shown to have integrated at the CUP1 locus by both tetrad analysis and Southern blot analysis (data not shown). The Southern analysis showed that each of the four transformants contained a URA3 insertion at a different location in CUP1. Each of these strains was then transformed to Leu+ with Hindlll-digested CB25 in order to introduce the topl-7::LEU2 mutation. These transformants were designated FDY63, FDY65, FDY67, and FDY69. CY180 and CY182 CY180 is a diploid made by mating CY143 and CY147. CY143 is a Ura+ transformant of CY141 using CB32 (which contains rDNA sequences and the URA3 gene). CY147 isa His + transformant of CY142 (an otherwise isogenic MATd derivative of CY141) using CB39 (which contains rDNA sequences and the HIS3 gene). Therefore, CY180 is a diploid with a URA3 insert in the rDNA of one parent and a HIS3 insert in the rDNA of the other. CY182 was made by mating CY151 and CY155. CY151 is the top1 derivative of CY143, and CY155 is the top1 derivative of CY147 (both made by single-step disruption using Hindlll-digested CB25). Therefore, CY182 is isogenic to CY180 except that it is toplltopl whereas CY180 is TOP1/TOP1. Plasmid Constructions CB32 This plasmid contains the 6.4 kb Hindlll fragment from yeast rDNA cloned into pMC303 (Crabeel et al., 1985), which was partially digested with Hindlll. pMC303 is 11kb in size and carries an ARG3::IacZ fusion, the URA3 gene, and pBR322 backbone. CB25 This plasmid carries the 5.1 kb HindIll fragment containing the TOP1 gene from which 849 internal nucleotides have been deleted and the LEU2 gene has been inserted (Thrash et al., 1985). The disrupted topi7::LEU2 HindIll fragment is 5.1 kb long and is cloned into pUC18 to make CB25 (also designated pCT80; Thrash et al., 1985). CB21 This plasmid carries a 2.2 kb Xbal-Xhol fragment containing an internal portion of the HIS4 gene cloned into the Xbal-Xhol sites in the pUC18 polylinker. CB30 This plasmid carries a 2.2 kb Xbal-Xhol fragment containing an internal portion of the HIS4 gene cloned into the Nhel-Sall sites in YIP5 (which carries the URA3 gene). CB26 This plasmid was constructed by digesting CB21 with Clal and cloning into that site a 20 nucleotide oligomer containing the putative topoisomerase I binding site from rDNA (see DNA sequence above). The oligomer was made with Clal ends. CB31 This plasmid was constructed by cloning the 2.2 kb Xbal-Xhol fragment from CB26 (which contains an internal portion of the HIS4 gene with the 20 nucleotide oligomer carrying the putative yeast rDNA topoisomerase I site cloned into the Clal site) into the Nhel-Sall sites in YIP5. CB15 This plasmid carries a disrupted copy of the RAD52 gene in the BamHI site of pBR322. The 2 kb BamHI fragment of RAD52 (Schild et al., 1983) is disrupted by a BamHI-Bglll fragment containing TRP1 from YRp7. Thus, digestion of CB15 with BamHI releases a fragment containing the rad52::TRP1 disruption and a pBR322 backbone. CB39 This plasmid carries a pBR322 backbone into which the 6.4 kb Hindill fragment from yeast rDNA has been cloned in the Hindill site. It also carries a 1.8 kb BamHI fragment containing the HIS3 gene in the BamHI site of pBR322. FDpl04 The Kpni fragment of JW6 (see Fogel and Welch 11982] for a restriction map) was cloned into the Kpnl site in pUC18. Topoisomerases Suppress Mitotic rDNA Recombination 425 post division segregation. Cold Spring Harbor Symp. Quant. Biol. 49, 629-637. Russell, P.J., Petersen, R.C., and Wagner, S. (1988). Ribosomal DNA inheritance and recombination in Neurospora crassa. Mol. Gen. Genet. 211, 541-544. Schild, D., Konforti, B., Perez, C., Gish, W., and Mortimer, R. (1983). Isolation and characterization of yeast DNA repair genes. Curr. Genet. 7,85-92. Schweizer, E., MacKechnie, C., and Halvorson, H. 0. (1969). The redundancy of ribosomal and transfer RNA genes in Saccharomyces cerevisiae. J. Mol. Biol. 40, 261-277. Szostak, J. W., and Wu, R. (1980). Unequal crossing over in the ribosomal DNA of Saccharomyces cerevisiae. Nature 284, 426-430. Thrash, C., Voelkel, K., DiNardo, S., and Sternglanz, R.(1984). Identification of Saccharomyces cerevisiae mutants deficient in DNA topoisomerase I activity. J. Biol. Chem. 259, 1375-1377. Thrash, C., Bankier, A. T., Barrell, B. G., and Sternglanz, R. (1985). Cloning, characterization, and sequence of the yeast DNA topoisomerase I gene. Proc. Natl. Acad. Sci. USA 82, 4374-4378. Voelkel-Meiman, K., Keil, R. L., and Roeder, G. S. (1987). Recombination-stimulating sequences in yeast ribosomal DNA correspond to sequences regulating transcription by RNA polymerase I. Cell 48, 1071-1079. Wang, J. C. (1985). DNA topoisomerases. Annu. Rev. Biochem. 54, 665-697. Zamb, T. J., and Petes, T. D. (1982). Analysis of the junction between ribosomal RNA genes and single-copy chromosomal sequences in the yeast Saccharomyces cerevisiae. Cell 28, 355-364. Chapter III The ribosomal DNA array has an altered structure in topoisomerase I mutants of S. cerevisiae The ribosomal DNA array has an altered structure in topoisomerase I mutants of S. cerevisiae (Pulse field gel/topl) Fred S. Dietrich*t, Michael F. Christman*, and Gerald R. Fink*t *Whitehead Institute for Biomedical Research, 9 Cambridge Center, Cambridge MA 02142 and tDepartment of Biology, Massachusetts Institute of Technology, Cambridge, MA 02139 Yeast mutants lacking the only type I topoisomerase activity detectable in crude extracts (topl mutants) display only a minor slow growth phenotype. We have shown previously that topl mutants have an elevated rate of mitotic recombination in the ribosomal DNA (rDNA) array but not at other loci. We now report that in strains deleted for TOP1, chromosome XII (the rDNA containing chromosome) specifically fails to migrate into a pulsed field gel. The other 15 chromosomes do not migrate differently in top1 mutant versus wild-type strains. The aberrant migration of chromosome XII is caused by the presence of rDNA. When chromosome XII from top1 strains is cut into large fragments with a restriction enzyme, the fragment containing rDNA fails to enter the gel, whereas those without rDNA migrate normally. Furthermore, when rDNA is transferred to chromosome III, chromosome III migrates aberrantly in topl strains. These data demonstrate that the rDNA array has an unusual structure in topl mutants and suggest that topoisomerase I may have a specific role in rDNA metabolism. The TOP1 gene of Saccharomyces cerevisiae encodes a type I topoisomerase (1,2). Strains carrying a deletion of the TOP1 gene are viable, exhibiting only a minor growth defect. (2). This is surprising because the TOP1 gene encodes the only biochemically detectable type I topoisomerase activity in yeast; an activity that has been proposed to be required for several crucial processes in the cell including transcription and DNA replication. Recently we have shown that strains carrying a topi deletion have a 50-200 fold higher level of mitotic recombination in the ribosomal DNA (rDNA) than do wildtype strains (3). This elevated level of recombination occurs only in the rDNA, which consists of about 200 copies of a 9.1 kb unit encoding all four mature ribosomal RNAs (25, 18, 5.8, and 5S), and not in other repeated or single copy genes (3). Furthermore, topoisomerase I has been shown recently to be localized to the nucleolus (the site of rDNA transcription and ribosome biogenesis) in both yeast (4) and in animal cells (5). MATERIALS AND METHODS Preparation of Yeast Chromosomal DNA. Yeast chromosomes were prepared from logarithmically growing cells using the method of Carle and Olson (6). DNA to be cut with restriction enzymes was washed extensively in 1X restriction enzyme buffer (approximately 10 washes in 5 ml buffer) while the DNA was still imbedded in low melt agarose The DNA plug was then digested in a minimal amount of 1X buffer overnight at 370 with 50 - 100 units of enzyme. Running Pulse Field Gels. Pulse field gels were run using 0.5X TBE, 1% agarose in a CHEF apparatus (7). Gel buffer was cooled by circulation through a refrigerating water bath. Gels were stained with ethidium bromide and blotted to nitrocellulose by standard techniques (8). Southern Blots were probed using hexamer labeled probes (9). RESULTS In addition to the high frequency of recombination that occurs in the rDNA in topi mutants we report here that there is a physical alteration in the rDNA array. When chromosomes are isolated from isogenic TOP1 and top1 strains and separated by pulsed field electrophoresis (10), chromosome XII from the topi strain fails to enter the gel. The 15 other chromosomes that make up the chromosome set migrate normally in topl mutants and are present at levels equal to those found in the TOP1 strain (Fig. 1A). This phenomenon is found in many different DNA preparations, in strains from several different genetic backgrounds, and by two different DNA separation methods (7,11). Even when the DNA was subjected to conditions that separate very large chromosomes (12) or lig. I lulsed field gel ele~trophoresis of yea'st chromosomal DNA made from isogenic T( )1'I and tl,1p strains. (A) Pulsed field electrophoresis performed at 12 ( ,121) volts, with a 5 minute switching time for 32 hours (standard conditions).The gel was stained with ethidium bromide. (B) Pulsed field electrophoresis performed at 1() C(, 5() volts, with a 30) minute switching time for 122 hours. The gel was stained with ethidium bromide. A a WELL XII IV B XII- was extensively treated with RNase , chromosome XII from topi strains failed to enter the gel (Fig. 1B). A number of different experiments indicate that the rDNA repeat on chromosome XII is responsible for the aberrant migration of chromosome XII. When the chromosome preparations are cut with NotI , a restriction enzyme that fails to cut in the rDNA array but cuts chromosome XII into several large fragments, the rDNA containing NotI fragment fails to migrate into the gel but a non-rDNA fragment of chromosome XII containing the ILV5 gene migrates into the gel normally (Fig. 2A). The failure to observe chromosome XII on CHEF gels is not a consequence of preferential degradation of rDNA because cleavage of the same chromosome preparations with an enzyme that cuts once in each of the 200 rDNA repeats (Pstl) reveals equal amounts of rDNA in TOPI and topI strains (Fig. 2B). Furthermore, the amount and migration of rDNA isolated by standard procedures for Southern analysis on standard agarose gels was identical in TOP1 and topI strains (3).Thus, the aberrant migration of rDNA in topi strains is not observed when the DNA is cut into single repeat units (9kb). This result also demonstrates that the physical alteration in the array requires the presence of more than a single repeat unit, as individual units migrate similarly in topl mutant and TOP1 strains. Fig 2 (A) Chromosome preparations were digested with Notl, run under standard conditions., and transferred to nitrocellulose filters. The filters were then hvbridized to a mixture of 32P labelled DNA containing both rDNA and the ILV5 gene.(B) Chromosome preparations were digested with Pstl, run under standard conditions, and probed with the h.3 kb Hindll fragment of rDNA. A WELL- · .. B ' ýI -IIF & rDNA NotI -W fragment rDlNA repeat ILV5 NotI i - 4iaiLnc III 65 Fig.3 Construction of a strain containing rDNA on chromosome II and its behavior in pulsed field gels (A) The initial strain contains two different leu2 alleles, one on chromosome I and one inserted into rDNA on chromosome XII.(13) A strain carrying a Ieu2 fragment integrated into the rDNA can undergo a crossing over between leu2 in the rDNA and the resident leu2 allele giving rise to a Leu + strain carrying a reciprocal translocation (first arrow, 13). A second recombination event (second arrow) between the rDNAs on chromosome III::XII and chromosome XII::III reconstituted chromosome XII and generated a chromosome III containing a duplication of Ieu2 flanking the rDNA. (B)Chromosome preparations from logarithmically growing cultures were from strains CY215 which carries a III::rDNA chromosome(TOPl)and CY216 (isogenic to CY215 except that it contains a topl-7 mutation introduced by transformation). Pulsed field gel electrophoresis, transfer to nitrocellulose, and hybridization to 32 p labelled rDNA were performed under the same conditions as described in Figure 1. A rDNA leu2 rDNA XII III- leu2 -------- --- --leu2 XII::III III::XII XII' LEU2 -----, , -rDNA rDNA 66 w c,. ell Wcl - - !• XII Ilil::rDNA - 67 When rDNA is translocated to chromosome III, a chromosome that does not normally contain any rDNA, chromosome III now fails to enter a pulsed field gel in a topl strain (Fig. 3B). The structure of the rDNA repeats permits the transfer of an rDNA segment to chromosome III by genetic means (see Figure 3A for details). A chromosome III reconstituted by this event will have an rDNA array at the site of the original translocation (LEU2 locus). It should be noted that no other segment of chromosome XII should be transposed to chromosome III by this event (Fig 3A). Chromosome III containing rDNA transposed to the LEU2 locus by this method fails to enter the gel in a top1 mutant. The rDNA cistrons on chromosome III also show increased recombination in top1 strains- an ADE2 marker inserted into the rDNA on chromosome III shows and approximately 80-fold increased loss in a top1 strain as compared with a isogenic TOP1 strain. The aberrant migration of chromosome XII in pulsed field gels is observed in top1 strains only when they are growing exponentially (Fig. 4). Chromosome XII from top1 cells that have entered stationary phase migrates into the gel normally compared to chromosome preparations from a TOP1 strain. This result is important in view of the high frequency of recombination in top1 mutants because an explanation for the unusual behavior of chromosome XII might be Fig 4 Chromosome preparations from topi and TOP1 strains grown to stationary phase. DNA was run under standard conditions, blotted to nitrocellulose, and probed with the 6.3 kb Hindl fragment of rDNA. O 0 WELLXII- 69 that the excessive recombination produces a great degree of size heterogeneity in chromosome XII, some copies containing more and some containing less rDNA. These chromosomes might be spread out over the gel so that no distinct band is observed. The restoration of a single chromosome XII of normal size in stationary phase makes this explanation unlikely. Mutants defective in DNA topoisomerase II do not show aberrant migration of chromosome XII in pulsed field gels although they exhibit an elevated rate of recombination in the rDNA (Fig 5). This result suggests that the elevated rate of recombination in rDNA in topi and top2 mutants occurs through different mechanisms. Both of these experiments were performed with a ts allele of TOP2 (top2-1) at a semi-permissive temperature (300C), because the TOP2 gene is essential for viability. At 300C top2-1 strains exhibit elevated recombination in the rDNA. However, the interpretation of this experiment is not simple, because the top2-1 allele may provide sufficient activity at 300C to mask a migration defect. DISCUSSION Our data demonstrate that the rDNA array has an altered structure in strains carrying the top1 mutation.This structure must be transient because chromosome XII migrates normally when isolated from stationary phase cells. The migration defect seems unlikely to be Fig 5 Chromosomes were prepared from strain MFC39-6D (top2-1) growing logarithmically at 24 C or 30°C.and electrophoresed under standard conditions Southern blot was probed with the 6.3 kb Hindlll fragment of rDNA. 0 WELL XII 4 0 caused by unresolved recombination intermediates because both topi and top2 mutants show elevated levels of recombination in the rDNA (3), but only top1 mutants show altered mobility of chromosome XII. Another possible explanation for the observed phenotype is that the absence of topoisomerase I specifically interferes with replication of the rDNA array. Topoisomerases have been proposed to act as swivels to relieve the torsional stress that accompanies the movement of a replication fork. Perhaps a slower replication of the rDNA in top1 mutants allows replication bubbles to persist through much of the cell cycle. DNA containing these replication bubbles might fail to enter the pulsed field gel. In stationary phase when the cells are no longer replicating their DNA, chromosome XII would enter the gel normally because it would no longer have these bubbles. This explanation is attractive because it has been shown that during any given round of rDNA replication only one in every 3-5 replication origins is used (14,15) and the altered structure that we observe in topl mutants requires more than one rDNA unit. Perhaps the migration defect is cause by an aberrant migration of the 3-5 unit replication bubbles, whereas when the DNA is cut into single unit pieces these altered structures are destroyed. Alternatively, the altered mobility could result from a transcriptional defect in the topl mutant. Ribosomal DNA is highly transcribed as compared to most single copy genes. Transcription occurs divergently with RNA polymerase I generating a 35S transcript and RNA polymerase III producing a 5S transcript from the opposite strand (16). Divergent transcription has been postulated to introduce positively and negatively supercoiled domains (17), which can be relaxed by the actions of DNA topoisomerases. In the absence of topoisomerase I accumulation of supercoiled regions may lead to the rDNA becoming physically altered in such a way that large rDNA regions are unable to migrate into a pulsed field gel. In addition to the specific elevation of rDNA recombination in topl mutants (3) and the localization of the TOPI protein to the nucleolus (4) the physical alteration of the rDNA in top1 mutants further suggests that topoisomerase I has some specific role in rDNA metabolism. Determination of precisely what that role is will require further experiments. We thank Tom Petes from the translocation strain, and Nikki Levin, David Pellman, and F. Scott Kieff for critical reviews of the manucript M.F.C. was supported by a grant from the Jane Coffin Childs Memorial Fund for Medical Research. The work was supported by grant GM35010. G.R.F. is an American Cancer Society Professor of Genetics. REFERENCES AND NOTES 1. Goto, T., Laipis, P. & Wang, J. C. (1984) J. Biol. Chem. 259, 1042210429. 2. Thrash, C., Voelkel, K., DiNardo, S. & Sternglanz, R. (1985) Proc. Natl. Acad. Sci. U.S.A. 82, 4374- 4378. 3. Christman, M. F., Dietrich, F. S. & Fink, G. R. (1988) Cell 55, 413425. 4. Giroux, C. N., Dresser, M. E. & Tiano, H. F. (1989) Genome 31, 8894. 5. Muller, M. T., Pfund, W. P., Mehta, V. B. & Trask, D. K. (1985) EMBO J. 4, 1237- 1243 6. Carle, G. F. & Olson, M. V. (1987) Methods in enzymology 155, 468482. 7. Chu, G., Vollrath, D. & Davis, R. W. (1986) Science 234, 1582- 1585. 8. Maniatis, T., Fritsch, E. F. & Sambrook, J. (1982) Molecular Cloning (CSHL, Cold Spring Harbor, NY) 9. Feinberg, A. & Vogelstein, B. (1983) Analytical Biochemistry 132, 613. 10. Schwartz, D. C. & Cantor, C. R. (1984)Cell 37, 67- 75. 11. Carle, G. F. & Olson, M. V. (1985) Proc. Natl. Acad. Sci. U.S.A. 82, 3756- 3760. 12. Smith, C. L., Matsumoto, T., Niwa, O., Klco, S., Fan, J. B., Yanagida, M. & Cantor, C. R. (1987) Nucleic Acids Research 15, 4481- 4489. 13. Mikus, M. D. & Petes, T. D. (1982)Gentetics 101, 369-404. 14. Walmsley, R. M., Johnston, L. H., Williamson, D. H. & Oliver, S. G. (1984) Mol. Gen. Genet. 195, 260- 266. 15. Saffer, L. D. & Miller, O.L. Jr. (1986) Mol. Cell. Biol. 6, 1148 - 1157. 16. Bell, G. I., DeGennaro, L. J., Gelfand, D. H., Bishop, R. J., Valenzuela, P. & Rutter, W. J. (1977) J. Biol. Chem. 252, 8118- 8125. 17. Liu, L. F. & Wang, J. C. (1987) Proc. Natl. Acad. Sci. U.S.A. 84, 7024- 7027.