Effect of Hydrostatic Pressure on the Egg Development of Marine... Copepods 海産カラナス目カイアシ類の卵発生に与える圧力の影響
advertisement

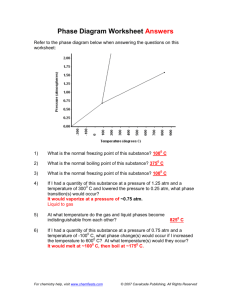
Effect of Hydrostatic Pressure on the Egg Development of Marine Calanoid Copepods 海産カラナス目カイアシ類の卵発生に与える圧力の影響 05D5504 吉木 朝子 指導教員 戸田 龍樹 SYNOPSIS 動物プランクトンの分布深度と卵の圧力耐性との関係を調べるために、生活史や産卵深度また、分布深度が異なるカイア シ類 3 属 5 種の Acartia steueri, Calanus sinicus, Neocalanus cristatus, N. flemingeri, N. plumchrus の卵発生に与える圧力の影響 を調べた。まず実験室で圧力環境をつくるため、(1)プランクトンが現場で経験するような、緩やかな圧力上昇が可能で あり、(2) 圧力容器の内部に 2 重構造を有することで、海水による圧力容器の腐食、また金属による生物への影響をなく し、(3)動物プランクトンの卵から成体までの幅広いサイズ(100-10000μm)の試料を観察することができる、圧力装置を開 発した。この装置を用い、異なる圧力、水温条件下においてカイアシ類の卵発生速度、卵の孵化率を調べ、卵発生に与え る圧力の影響を見た。また、圧力の変化が卵発生に及ぼす影響を見るために、圧力上昇速度を急激に上昇させた場合と、 緩やかに上昇させた場合における孵化率、奇形率の違いを観察した。卵発生速度はすべての種において、温度の変化に依 存し、圧力条件間における違いは見られなかった。孵化率においては、A. steueri では 1atm と 10atm で有意差が見られな かったのに対し、C. sinicus の卵は圧力条件の上昇と共に孵化率が低下する様子が観察された。また C. sinicus では、10atm まで圧力を徐々に上昇させた場合、急激に上昇させた場合よりも、孵化率が顕著に低い結果となり、緩やかな圧力変化で よりダメージを受けやすいことが明らかとなった。Neocalanus 属 3 種の卵の孵化率は、全ての実験条件において 1atm における孵化率と有意な差はなく、1atm から 100atm までの広い範囲の圧力に耐性を持っていることが明らかとなった。 孵化した幼生において、A. steueri と Neocalanus 属では奇形個体は観察されなかったのに対して、C. sinicus では奇形個体 が観察された。Neocalanus 属と C. sinicus はどちらも生活史を通して、表層から深度 1000m 以深までの生活史に伴う鉛直移動 OVM を行っているが、卵の圧力耐性は異なった。Neocalanus 属カイアシ類は深層 1000m 以深で産卵された後に、孵化したノープ リウスは表層まで上昇するため、初期生活史において 0~1000m の幅広い圧力範囲を経験する。それに対して、 C. sinicus の卵は 表層付近で産卵され沈降し、深度 100m よりも浅い深度で孵化し、ノープリウス幼生は表層へと上昇する。そのため、C. sinicus の 卵からノープリウス幼生の時期は 0m から深くても 100m までの圧力しか経験しない。このような Neocalanus と C. sinicus の初期生 活史において経験する圧力範囲の違いが、卵の圧力耐性と関係があると考えられた。 Key words: hydrostatic pressure, egg development, egg hatching success, vertical distribution, Acartia steueri, Calanus sinicus, Neocalanus cristatus, N. plumchrus, N. flemingeri relation to spawning in zooplankton. (1) Adult is usually distributed and spawns at the surface layer. Spawned eggs sink and hatch in the water column. Hatched nauplii ascend to feed and develop at the surface. Nauplii grow to copepodid V stage (CV) near the surface layer. CV copepods descend to deeper depth and molt to adult. Adult copepods return to the surface layer to feed and reproduce. (2) Adult is distributed at deep layers, and they spawn at the depth. Spawned eggs float and hatched nauplii ascend to the surface. They develop to copepodid V stage at the surface and descend to the depth again. Thus, copepods experience wide pressure range from surface to deep by undergoing vertical migration, and they reproduce eggs at specific depth range by according to species. Eggs spawned into the water column float at certain depth or sink to deeper waters. These eggs experience cleavage and environmental changes during sinking and/or ascending. In the cleavage process, different blastomere stages appear and they are subsequently exposed to different environmental factors such as variation in temperature and pressure in the water column. Egg development time of krill was examined in various pressure increasing conditions. The cleavage process was accelerated under moderate pressure of 1 to 5-20 atm than at 1 atm (George 1984). On the other hand, abruptly increasing pressure from 1 to 40-60 atm inhibited the cleavage process of krill eggs (George and Stromberg 1985). The pressurizing rate, abruptly or gradually, is considered to have some influence on the development of embryos. Also, the sensitivity of embryos to environmental factors may be variable among blastomere stages. The hydrostatic pressure appears to be an important Introduction Copepods are one of the most prime important animals in marine ecosystems because of their large biomass and trophic importance as prey organisms of other animals such as fishes and whales at higher trophic levels. In order to understand population dynamics and biomass of copepods in nature, it is necessary to investigate recruitment rate of copepods, Recruitment of copepod is determined by the egg production rate, egg hatching success and egg mortality rate. On the recruitment process, spawned eggs sink or ascend, and thus these eggs are exposed to variation in temperature and pressure. Although effect of temperature on egg hatching success and/or egg mortality has been studied quite a lot, pressure effect on eggs almost has never been considered in the recruitment. Because there are not many pressure apparatus which can mimic in situ pressure environment for zooplankton. A system producing in situ pressure conditions is essential for examining the effect of pressure on eggs and recruitment rate. Copepods occur from surface to sea bottom and they are exposed to various pressures throughout their lifetime. Most of copepods undergo diel vertical migration (DVM) and/or ontogenetic vertical migration (OVM). DVM is characteristic for later stages of copepod, probably for females (Conover 1988). Many calanoid copepods usually spawn eggs at night time when they migrate to the surface (e.g. Uye et al. 1990, Pagano et al. 2004). OVM is called as seasonal vertical migration, because large migration of zooplankton is related to boreal- or arctic- area where seasonal variations in temperature and phytoplankton production. Zooplankton has been evolved OVM as a survival strategy to get over seasonal environmental variations (Sekiguchi 1975). There are two patterns of OVM in 1 different areas from shallow coastal waters to open waters of West Pacific Ocean near Japan. These species differ in body size, habitat depth range, life cycle and spawning depth. A. steueri is dominant in near shore waters such as embayment (c.a. 5 ~ 20 m depth). C. sinicus and Neocalanus copepods are distributed from surface layer to deeper layer than 1000 m. All five species undergo DVM. C. sinicus and Neocalanus copepods also migrate ontogenetically from surface to deeper than 1000m. C. sinicus belong to OVM pattern (1) mentioned above. While Neocalanus copepods spawn at the depth, and thus these species belongs to OVM pattern (2). factor in egg development of marine animals, whereas pressure effect on eggs has not been understood well yet. The purposes of the present study were firstly (1) to develop new hydrostatic pressure apparatus which can increase pressure gradually for marine zooplankton egg study, (2) to select species which have different habitat and spawning depth range and to examine the effects of hydrostatic pressure and also pressurizing rate on eggs of selected zooplankton, and (3) to examine the effects of hydrostatic pressure between different egg development stages which change during egg sinking and/or ascending in the water column. Materials and Methods Study 1. Development of hydrostatic pressure apparatus The hydrostatic pressure apparatus developed in the present study can function at pressures of up to 200 atm and at temperatures ranging from -20 to 80 oC (Yoshiki et al. 2006). The pressure apparatus system is composed of a plunger pump, a pressure gauge, a pressure chamber, a specimen holder which is placed in the pressure chamber, a temperature-controlled bath, a CCD camera, a monitor and a video-recorder. Depth (m) 0 ri Whole life cycle Female distribution Spawning depth 500 1500 (Modified from Shimode 2003; Kobari and Ikeda 1999, 2000 a,b) Fig. 2. Distribution ranges of Acartia steueri, Calanus sinicus, Neocalanus spp. in Sagami Bay and Oyashio region. Spawning is conducted at female distributed depth. Pressure gauge ◆Sampling Acartia steueri Acartia steueri were collected in Manazuru Port (35◦ 09’49” N, 139◦ 10’ 33” E; depth, 5.5 m) where is located at near the north coast of Sagami Bay, Japan in November, December 2006, April, November 2007. Samplings were carried out by oblique tows using a 180 μm mesh net from the bottom to the surface. Adult females were individually incubated in a 10 ml beaker filled with a suspension of 105-106 cells mL-1 of Thalassiosira weissflogii mixed with Isochrysis galbana as food at 15.0 ◦C. Eggs spawned by the females were collected and used for pressure experiments. Calanus sinicus Adult females of C. sinicus were collected on board R. V. “Tachi-bana” of Yokohama National University, at a fixed location off the coast of the Manazuru Peninsula (35°09’55”N; 139°10’78”E, depth 300m), Sagami Bay, Japan. Samplings were carried out from June to November 2004. C. sinicus was obtained by gently towing a vertical zooplankton net with 330 μm mesh opening fitted with a 2 L cod-end from depth of 200 m to surface. Females of C. sinicus sorted with a wide-mouth pipette and individually placed in incubation chambers filled with suspension of 105-106 cells mL-1 of Thalassiosira weissflogii to obtain fresh eggs. The beakers were placed at 14 °C in temperature controlled incubators. Eggs were checked every morning and harvested for pressure experiments. Neocalanus copepods Adult females of N. cristatis, N. flemingeri, N. plumchrus were collected on board the R. V. "Hokko-maru" of Hokkaido National Fisheries Research Institute, in the Oyashio region, Western Subarctic Pacific, in January 2006 and January 2007. Sampling was conducted by vertical hauls of a ring net with 330 μm mesh opening fitted with a 2 L cod-end from depth of 1500 m to surface. Females of Neocalanus copepods were sorted and transferred in to 1-L or 250ml bottles filled with filtered seawater. Bottles were kept Pressure chamber Distilled water CCD Light Specimen holder Monitor & video recorder icus . cristatus . plumchrus . fleminge N N C. sin N 1000 Plunger pump camera eri A. steu Temperature Fig. 1 Structure of hydrostatic controlled pressurebath apparatus. Hydrostatic pressure in the chamber is controlled by sending distilled water into the chamber with a plunger pump (AC-L9, Shimadzu). The increasing velocity of pressure in the chamber is controlled by the flow rate of distilled water from the plunger pump (0.001 – 9.999 ml/min). The flow rate is adjusted so as to maintain a constant increase in pressure. The pressure chamber enables us to control the pressure increasing rate by adjusting a plunger pump which changes the flow rate of water into the pressure chamber. The pressure inside the pressure chamber is monitored by a pressure gauge (1~200 atm pressure range). A CCD camera (STC-R640, Sensor Technology Corporation) is placed in front of an observation window of the chamber and a light source is placed at the opposite side of the chamber. Specimen of image is displayed on a monitor and recorded by a video-recorder connected to the CCD camera. Study 2. Effects of hydrostatic pressure on copepods eggs ◆Selected species In the present study, clanoid copepods: Acartia steueri, Calanus sinicus, Neocalanus cristatis, N. plumchrus and N. flemingeri were selected. These species are the dominant in 2 at 4 °C in cold room and incubator. Bottles were checked every morning and spawned eggs were used to pressure experiments. ◆Pressure experiments For A. steueri, experiments were conducted at two pressure conditions, 1 and 10 atm and five temperatures, 4, 10, 15, 20 and 25 °C, for A. steueri. At 10 atm, two pressurizing patterns, abruptly (A) at 10 atm/min (10atm(A)) and gradually (G) at 0.1 atm/min to 10 atm (10 atm(G)), were used. In the gradual increasing experiment, the pressure was increased in a stepwise fashion during the first 100 min of the experiment and then maintained at constant during the remainder of the experiment. For C. sinicus, experiment was conducted at four pressures, 1, 10, 50 and 100 atm, and six temperatures, 5, 8, 10, 15, 20 and 25 °C. At 10 atm, two pressurizing patterns, 10 atm (A) and 10 atm (G), were used as mentioned above. For Neocalanus copepods, experiments were conducted at four pressures, 1, 10, 50 and 100 atm at 4 °C. At 10, 50 and 100 atm, two pressurizing patterns, abruptly and gradually, were used. Egg development times, egg hatching successes and deformation frequencies at each condition were examined. same as that at 1 atm between different pressure conditions and pressurizing rates, at all temperature conditions. These egg development times were changed depending on temperature which has been thought the most important factor for the egg development time. On the basis of the difference of the life histories and the spawning depths of examined copepods, probably it is thought that the egg development time of calanoid copepods is not influenced by the hydrostatic pressure. Egg development time of Euphausia superba is changed by pressure increasing rates (George 1984, George and Stromberg 1985). To clarify the difference related to taxa, copepods and krill, examining the response of egg development to hydrostatic pressure between copepods and E. superba in detail is needed. Egg hatching successes of A. steueri and C. sinicus at 1 atm were generally higher than 70 and 80 % at all temperature conditions, respectively (Fig. 3). At 10 atm (A; abruptly pressure increasing) condition, egg hatching successes of A. steueri and C. sinicus were not significantly different to 1 atm. Egg hatching success of A. steueri at 10 atm (G; gradually pressure increasing) condition was not significantly different to 1 atm. On the other hand, at 10 atm (G) condition, egg hatching successes of C. sinicus were significantly lower than 1 atm at all temperature conditions. Although A. steueri habitat is shallow coastal waters where depth is around 5-20 m, egg hatching successes were high even at 10 atm (G) which is as high as pressure of 100 m depth. Study 3. Hydrostatic pressure effect on egg development stages of C. sinicus Effects of pressure between blastomere stages were examined on C. sinicus eggs. Sampling method was same as Study 2. Samplings were conducted on December 2006, April, May, June and July 2007. The 1-, 2-, 4-, 8-, 16-32 cell, blastula and limb-bud stages were collected respectively. The eggs were divided to two aliquots. One aliquot was used in the pressure experiment and the other was used in the control experiment where pressure was kept constant at 1 atm. Two pressure increasing conditions were experimented; abruptly at 10 atm/min (10 atm (A)) and gradually at 0.1 atm/min (10 atm (G)) to a final pressure of 10 atm. Hatching success (%) 100 80 40 20 0 Hatching success (%) 100 Results and Discussions Study 1. Development of hydrostatic pressure apparatus The hydrostatic pressure apparatus developed in the present study makes it possible to (1) increase pressure gradually, (2) prevent corrosion of the pressure chamber by introducing a specimen holder that separates the pressure chamber from the surrounding seawater, and (3) observe a wise size range of zooplankton including eggs (100-10000μm size range). The present apparatus has made it possible for examining the effects of hydrostatic pressure on zooplankton, for instance egg development time and larval behavior by the response to pressure change. Hand pump has been previously adopted by most hydrostatic pressure apparatuses to increase pressure, but this method exposes zooplankton to abrupt increase in pressure. In the present study, the rate of pressure increased in the chamber is controlled by adjusting the flow rate of distilled water from the plunger pump. This was possible to increase pressure gradually (0.1 atm/min) and also simulate the pressure in the field such as that observed when zooplankton migrates vertically (from 1 to 150 atm). A. steueri 60 4 10 15 20 25 80 60 1 atm 10 atm (A) 10 atm (G) 50 atm (G) 100 atm (G) C. sinicus 40 20 0 000 5 00 8 10 15 20 Temperature (ºC) 25 Fig. 3. Egg hatching successes of A. steueri and C. sinicus. A: abrupt increasing, G; gradual increasing pressure conditions. 0; Hatching was not observed. Egg hatching success of C. sinicus decreased with gradual increasing pressure condition, while that of Neocalanus copepods have tolerate to wide pressure range from 1 atm to 100 atm. Both species are exposed to wide depth range throughout their life time by undergoing OVM. However spawning depths are different between C. sinicus and Neocalanus copepods. Adult females of C. sinicus are distributed from 0 to around 150 m depth and spawn at surface layer. Spawned eggs sink and hatch around 100 m depth. Hatched nauplii ascend to surface. Thus, eggs and nauplii are exposed to narrow pressure range from 1 atm to 10 atm at the most. On the other hand, females of genus Neocalanus are distributed from 250 m to 2000 m depth and spawn eggs at the depth. The spawned eggs float at the depth Study 2. Effects of hydrostatic pressure on copepods eggs Egg development times of all examined species were 3 or ascend slowly. Also, hatched nauplii ascend to the surface. Thus, eggs and nauplii of Neocalanus copepod exposed to large pressure ranges from spawned deeper depth to surface at early larval stage. The exposing pressure range of their early life stage of egg and nauplii might be related to the difference of pressure tolerance between C. sinicus and Neocalanus copepods. Deformation of A. steueri and genus Neocalanus were not found. However, deformed nauplii hatched under pressure conditions were observed only on C. sinicus. northern limit of C. sinicus distribution is the Oyashio front that an average temperature of annual maximum at 20-30m of its spawning depth during year 1990-1998 is 13.0±1.0 °C (Kidachi 1979; Hulsemann 1994; Saito et al. 2002). The egg development time of C. sinicus at 13 °C is estimated at 1.2 days. Considering the egg sinking rate of 68 m/d, these eggs will sink to around 100 m depth in 1.2 days. At this depth, the hydrostatic pressure will cause increasing in the egg hatching success rate of C. sinicus. Hydrostatic pressure, coupled with low temperatures in this region, may limit the recruitment of C. sinicus in the Oyashio region (Yoshiki et al. 2006). In other words, northern limit of C. sinicus distribution may be determined by combination of egg sinking, embryonic development rates and tolerance to hydrostatic pressure. Study 3. Hydrostatic pressure effect on egg development stages of C. sinicus Effects of high pressures on different cleavage stages of C. sinicus which appears deformation were examined. Deformity frequencies were from 0 to 54 % between 1-cell and limb-bud stages at abrupt pressure increasing condition (10 atm/min), and from 8.3 to 91 % at gradual pressurizing condition (0.1 atm/min) (Fig. 3) (Yoshiki et al. 2008). The deformity frequency between pressurizing rates on each blastomere stage was significantly different at the blastula stage (p < 0.05), while there were no differences between the other stages. In the result of 2-way ANOVA of deformity frequencies, there were significant differences among pressurizing conditions (p < 0.05) and blastomere stages (p < 0.01), though there was no interaction between them (Fig. 3). This indicates that the eggs of Calanus sinicus were damaged more easily by gradual increasing than abrupt increasing pressure. Moreover, the sensitivity of the blastomere stage increased progressively with cleavage development, and the blastula stage was the most delicate in the early phases of egg development. Control Deformity frequency frequency (%) (%) Deformity 10 atm A Implications Egg of Acartia steueri can tolerate to 10 atm while that habitat is very shallow coastal area. Why eggs of A. steueri could tolerate to 10 atm pressure? All examined species are classified to Calanoida copepods which are supposed to be a monophyletic group (Park 1986). During the hundreds of millions of years spanning the evolutionary history of the subclass Copepoda (Huys and Boxshall 1991), the habitat shift from benthos to plankton performed by the Calanoida. This can be viewed as the major and probably earliest attempt by copepod to colonize the pelagic biome (Ohtsuka and Huys 2001). On the basis of morphological characters, Andronov (1974) divide the Calanoida into nine superfamilies and proposed phylogenetic relations of them. Calanoids are highly adapted to the planktonic mode of life and dominate zooplankton communities, though members of the most primitive families have retained their close association with the sediment-water interface, the ancestral hyperbenthic habitat where most copepod orders probably originated (Huys and Boxshall 1991; Ohtsuka and Huys 2001). That is to say, primitive families of Calanoida copepods are supposed to tolerate benthic environment which is including high pressure environment, and to lose the pressure tolerance after these got into planktonic habitats. Genus Acartia is classified to primitive group. The pressure tolerance of A. steueri eggs appears to be related to the evolutionary history of Calanoida copepods. The relationship between pressure tolerance of copepods and lineage is clearly an area that warrants significant research in future. 10 atm 10atm G 100 80 60 40 20 0 00 00 0 0 1 4 8 2 0 16 Blastomere Blastomere stage stage B 0 L 0 0 1 0 2 4 0 0 8 16 0 B 0 L Blastomere Blastomere stage stage Fig. 4. Deformation frequency between blastomere stages of Calanus sinicus. A; abrupt pressure increasing, G; gradual pressure increasing. B; Blastula, L; Limb-bud stage. The embryonic sensitivity of C. sinicus to hydrostatic pressure increased with egg development, and eggs were more fragile to gradual pressure change than abrupt increasing pressure. This means that egg sinking is supposed to be detrimental effect for C. sinicus. At higher pressure conditions at 10, 50 and 100 atm, highly significant decreases in egg hatching successes of C. sinicus occurred as pressure increased. These eggs did not have tolerance to even 10 atm, this suggests that eggs would not survive if they did not hatch within the upper 100m. Upward spawning behavior of females of C. sinicus at night in the DVM process has been known. By the female’s upward behavior, eggs are spawned at lower pressure and warmer temperature environments in the surface. Harmless low pressure and warm temperature lead eggs to hatch early and to recruit. Egg hatching success rate of C. sinicus decreased with increasing hydrostatic pressure. This implicated that the egg pressure tolerance is related to distribution of C. sinicus. The References Andoronov (1974) Zool. Zh. 53: 1002-1012 Conover (1988) Hydrobiol. 167/168: 127-142 Forward, Wellis & Buswell (1989) Mar. Ecol. Prog. Ser. 57: 267-277 Goerge (1984) Crustac. Biol. 4: 252-262 Goerge & Stromberg (1975) Poler. Biol. 4: 125-133 Huys & Boxshall (1991) The Ray Soc. Lond.: 468pp. Kobari & Ikeda (1999) Mar. Biol. 134: 683-696 Kobari & Ikeda (2001a) J. Plank. Res. 23(3): 287-302 Kobari & Ikeda (2001b) Mar. Ecol. Prog. Ser. 209: 243-255 Lincoln (1972) J. Exp. Biol. 54: 677-688 Ohtsuka & Huys (2001) Hydrobiol. 453/454: 441-466 Pagano, Kouassi. Arfi, Bouvy & Saint-Jean (2004) Zool. Studies. 43(2): 244-254 Park (1986) Syllogeus 58: 191-196 Sekiguchi (1975) Bull. Fac. Fish. Mie. Univ. 2: 29-38 Uye, Huang & Onbe (1990) Mar. Biol. 104: 389-396 Yayanos (1981) Comp. Biochem. Physiol. 69: 563-565 Yoshiki,Toda, Yoshida & Shimizu (2006) J. Plank. Res. 28(6): 563-570. Yoshiki, Yamanoha, Kikuchi, Shimizu & Toda (2008) Mar. Biol. 156: 97-106 Young, & Tyler (1993) Limnol. & Oceanogr. 38: 178-181 Young, Tyler & Gage (1996) J. Mar. Biol. Assn. UK. 76: 749-757 4