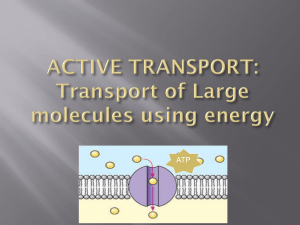
Molecular Circuitry of Endocytosis at Nerve Terminals Jeremy Dittman and Timothy A. Ryan
advertisement

Annu. Rev. Cell Dev. Biol. 2009.25:133-160. Downloaded from arjournals.annualreviews.org by WEILL MEDICAL COLLEGE OF CORNELL UNIVERSITY on 10/29/09. For personal use only. ANRV389-CB25-07 ARI 4 September 2009 17:25 Molecular Circuitry of Endocytosis at Nerve Terminals Jeremy Dittman and Timothy A. Ryan Department of Biochemistry, Weill Cornell Medical College, New York, NY 10065; email: jed2019@med.cornell.edu, taryan@med.cornell.edu Annu. Rev. Cell Dev. Biol. 2009. 25:133–60 Key Words First published online as a Review in Advance on July 8, 2009 synapse, vesicle cycle, trafficking, clathrin, pHluorin The Annual Review of Cell and Developmental Biology is online at cellbio.annualreviews.org This article’s doi: 10.1146/annurev.cellbio.042308.113302 c 2009 by Annual Reviews. Copyright All rights reserved 1081-0706/09/1110-0133$20.00 Abstract Presynaptic terminals are specialized compartments of neurons responsible for converting electrical signals into secreted chemicals. This selfrenewing process of chemical synaptic transmission is accomplished by the calcium-triggered fusion of neurotransmitter-containing vesicles with the plasma membrane and subsequent retrieval and recycling of vesicle components. Whereas the release of neurotransmitters has been studied for over 50 years, the process of synaptic vesicle endocytosis has remained much more elusive. The advent of imaging techniques suited to monitor membrane retrieval at presynaptic terminals and the discovery of the molecules that orchestrate endocytosis have revolutionized our understanding of this critical trafficking event. 133 ANRV389-CB25-07 ARI 4 September 2009 17:25 Contents Annu. Rev. Cell Dev. Biol. 2009.25:133-160. Downloaded from arjournals.annualreviews.org by WEILL MEDICAL COLLEGE OF CORNELL UNIVERSITY on 10/29/09. For personal use only. INTRODUCTION . . . . . . . . . . . . . . . . . . THE SYNAPSE . . . . . . . . . . . . . . . . . . . . . . THE SYNAPTIC VESICLE: A LOCAL CURRENCY FOR INFORMATION FLOW AT THE SYNAPSE . . . . . . . . . . . . . . . . . . . SYNAPTIC VESICLE RETRIEVAL AT NERVE TERMINALS . . . . . . . . THE TIMING OF ENDOCYTOSIS AT NERVE TERMINALS: EVIDENCE FOR MULTIPLE TIME SCALES OF RETRIEVAL . . . . . . . . . . . . . . . . . . . . . . MODULATION OF ENDOCYTOSIS: CALCIUM AND EXOCYTIC LOAD . . . . . . . . . KISS-AND-RUN: A LONG-RUNNING DEBATE . . . . . THE MOLECULAR MACHINERY OF SYNAPTIC VESICLE ENDOCYTOSIS. . . . . . . . . . . . . . . . . . A CLATHRIN-MEDIATED PATHWAY FOR VESICLE REBUILDING . . . . . . . . . . . . . . . . . . . . MECHANISMS FOR SORTING 134 134 135 136 137 140 141 142 143 INTRODUCTION SV: synaptic vesicle 134 Chemical communication between cells was harnessed for both distributing and controlling multicellular behavior early on in the development of complex life forms. In the nervous system, synapses encompass a transduction machine whereby propagating electrical signals, usually in the form of action potentials, are transformed into chemical messages in the form of packets of neurotransmitters that are secreted onto a neighboring cell, which in turn are transduced back into both chemical and electrical signals in this cell. The past 60 years have witnessed an intense scientific effort to understand how synapses work, how they form and are controlled, how their function changes over Dittman · Ryan THE APPROXIMATELY NINE CARGO PROTEIN TYPES . . . . . . . CURVING MEMBRANES: INITIATION OF SYNAPTIC VESICLE ENDOCYTOSIS . . . . . . . SECONDARY SCAFFOLDING FOR RECRUITING GENERAL ENDOCYTIC EFFECTORS . . . . . FINISHING THE JOB: MEMBRANE FISSION AND COAT DISASSEMBLY . . . . . . CALCIUM SENSORS FOR ENDOCYTOSIS? . . . . . . . . . . . . . . . . . A ROLE FOR ACTIN DURING SYNAPTIC VESICLE ENDOCYTOSIS? . . . . . . . . . . . . . . . . . REBUILDING SYNAPTIC VESICLES: A SINGLE-PASS SORTING AT THE PLASMA MEMBRANE? . . . . . . . . . . . . . . . . . . . . A MOLECULAR PICTURE FOR THE ENDOCYTIC MACHINE . . . . . . . . . . . . . . . . . . . . . . . NETWORK PROPERTIES OF THE ENDOCYTIC MACHINE . . . . . . . . CONCLUDING REMARKS . . . . . . . . . 144 145 149 149 151 152 152 152 153 154 time scales from milliseconds to months, and how they are built at the molecular level. Increasingly, as molecular detail has emerged, it has overlapped with the recognition that this information is crucial for understanding a variety of diseased states of neuronal function. Here we focus our attention on the first half of the synaptic transmission problem, the cell biology of the presynaptic nerve terminal and, in particular, the current understanding of endocytosis and synaptic vesicle (SV) recycling. THE SYNAPSE The basic structural organization of synaptic terminals appears to be relatively well conserved across evolution. Neurotransmitters are ANRV389-CB25-07 ARI 4 September 2009 17:25 Refilling, docking, priming Fusion Annu. Rev. Cell Dev. Biol. 2009.25:133-160. Downloaded from arjournals.annualreviews.org by WEILL MEDICAL COLLEGE OF CORNELL UNIVERSITY on 10/29/09. For personal use only. a Cargo mixing Lipid mixing f e b c Cargo capture Clathrin coat assembly Membrane curvature Membrane fission Uncoating d Vesicle budding Figure 1 The synaptic vesicle (SV) cycle. An SV fuses with the plasma membrane (a) and is subsequently rebuilt through the assembly of a clathrin-coated pit (b,c) followed by budding (d ), and fission and uncoating of the endocytic coat proteins (e). The vesicle is refilled with neurotransmitter and returned to the vesicle pool for further rounds of exocytosis ( f ). Several SV proteins are depicted including synaptobrevin/VAMP ( green), synaptotagmin ( purple), and the vacuolar ATPase (blue). Syntaxin is shown (red rods), and the assembled SNARE complex is represented (red-green coils). Other proteins depicted here are clathrin triskelia ( gray), adaptins (red ellipses), dynamin, and endophilin (red helices). packaged into small, clear vesicles with a diameter that ranges from ∼30 to 40 nm (depending on the species). Typically a few dozen to a few hundred SVs are maintained in a cluster near the active zone, a dedicated intracellular location on the presynaptic plasma membrane where SVs, upon the arrival of an action potential stimulus, will fuse with the plasma membrane and release their content through exocytosis (Figure 1). These preferential sites of exocytosis are typically found juxtaposed to a postsynaptic specialization of neurotransmitter-gated ion channels in the receiving cell. As nerve terminals are located along or at the end of axons very distal to the cell body and the total number of SVs is limited, ongoing synaptic transmission is maintained by local recycling of SVs. The mechanisms of how vesicles are rebuilt following exocytosis for reuse are the focus of this review. THE SYNAPTIC VESICLE: A LOCAL CURRENCY FOR INFORMATION FLOW AT THE SYNAPSE SVs act as key intermediates in converting electrical to chemical information at synapses www.annualreviews.org • Synaptic Vesicle Endocytosis 135 Annu. Rev. Cell Dev. Biol. 2009.25:133-160. Downloaded from arjournals.annualreviews.org by WEILL MEDICAL COLLEGE OF CORNELL UNIVERSITY on 10/29/09. For personal use only. ANRV389-CB25-07 ARI 4 September 2009 17:25 and represent a currency of information flow in the nervous system. Because their size is tightly regulated (see below), it has been possible to purify these organelles to near homogeneity. Such purified vesicles were the starting point for identifying SV proteins ( Jahn et al. 1985, Trimble et al. 1988, Perin et al. 1990) and provided the basis for generating the first draft of an SV proteome carried out by the lab of Reinhard Jahn (Takamori et al. 2006). SVs are ∼50% protein and 50% lipid, with a 1:1 ratio of phospholipid to cholesterol, similar to generic cellular plasma membranes. Following exocytosis, the transmembrane proteins of SVs become transiently incorporated into the plasma membrane and need to be endocytosed to rebuild the SV. Figure 2 illustrates the major transmembrane proteins associated with SVs as well as their average copy number deduced from quantitative proteomics. There are roughly nine types of transmembrane proteins that are specifically enriched in SVs and of these, only four types have been assigned clear-cut functions. These Synaptobrevin 2 / VAMP2 70 copies Vacuolar ATPase 1–2 copies are (a) the proton pump (vacuolar ATPase), which provides the proton-motive force for driving neurotransmitter uptake through (b) the vesicular neurotransmitter transporter (Edwards 2007), (c) synaptotagmin I, the calcium sensor for fast calcium-triggered exocytosis (Chapman 2008), and (d ) synaptobrevin 2/VAMP2, the vesicle-associated SNARE that provides one of the helices required to catalyze membrane fusion through formation of a fourhelix bundle with the specific plasma membrane SNARE proteins syntaxin and SNAP-25 (Rizo & Rosenmund 2008). It is noteworthy that the relative abundance of these different proteins varies significantly: Some proteins such as the proton pump, whose functional role is unequivocal and required for filling vesicles with transmitter, appear to be expressed at single-copy levels. In contrast, synaptophysin is at least one order of magnitude more abundant but is dispensable, at least in terms of animal viability (McMahon et al. 1996) and basic synaptic function (Abraham et al. 2006). Presently little is known about variability in the stochiometric relationship of these different proteins across individual vesicles. SYNAPTIC VESICLE RETRIEVAL AT NERVE TERMINALS Synaptotagmin I 15 copies Synaptogyrin 2 copies SV2 2 copies Neurotransmitter transporter 9–14 copies Synaptophysin 30 copies Scamp 1–2 copies Figure 2 The synaptic vesicle (SV) transmembrane proteins. Proteomic analysis of highly purified SV preparations allowed estimates of the abundance of each of the major transmembrane proteins associated with SVs. The protein copy numbers shown are taken from Takamori et al. (2006). 136 Dittman · Ryan Typical CNS synapses contain a limited number of SVs, usually a few hundred or less. As nerve terminals are typically located great distances from their cell bodies, the biosynthetic engines of cells, the proteins and lipids that make up a SV, must be recaptured from the plasma membrane following exocytosis, refashioned into a SV, and refilled with neurotransmitters (De Camilli et al. 2001). This currency of synaptic communication is in fact in relatively limited supply. The mechanism of vesicle retrieval in the SV cycle has been a subject of intense interest and debate for over 35 years. The first descriptions of vesicle recycling began with ultrastructural observations at neuromuscular junctions (NMJ), where uptake of extracellular horse radish peroxidase demonstrated conclusively the existence of the SV recycling Annu. Rev. Cell Dev. Biol. 2009.25:133-160. Downloaded from arjournals.annualreviews.org by WEILL MEDICAL COLLEGE OF CORNELL UNIVERSITY on 10/29/09. For personal use only. ANRV389-CB25-07 ARI 4 September 2009 17:25 pathway (Heuser & Reese 1973). These studies also provided the first evidence that vesicle retrieval occurred through a clathrin-mediated pathway. To visualize vesicle recycling at the EM level, it was necessary to provide a sufficiently rapid and intense stimulus while catching exo- or endocytic events in the act through rapid freezing. This approach led to the first kinetic description of endocytosis (Miller & Heuser 1984) because the time between stimulus and freezing could be varied (Figure 3a,b). The relatively long time scales observed (1 min for completion of vesicle endocytosis) fueled speculation that a faster, more direct route must be operational, and they provided grist for the idea that endocytosis may occur by reversal of fusion at the active zone, first proposed by Ceccarelli and colleagues (Ceccarelli et al. 1973); this pathway was later coined kiss-andrun. During the past 10–15 years, efforts to understand the mechanisms of endocytosis of SVs have been concentrated in two approaches. One is the examination and discovery of the molecular machinery responsible for endocytosis at nerve terminals; the other is the biophysical probing of the time scales and molecular behavior of the endocytic process at the synapse. THE TIMING OF ENDOCYTOSIS AT NERVE TERMINALS: EVIDENCE FOR MULTIPLE TIME SCALES OF RETRIEVAL Synaptic endocytosis kinetics measurements became possible with the advent of new technologies as well as suitable synaptic preparations. Electrical capacitance recordings, which, in principle, provide direct measurements of cell surface area, allow one to measure the net balance of exocytosis and endocytosis. This has been useful, however, only at select giant synaptic preparations because SVs have a very small surface area (3000–5000 nm2 ) and sites of fusion are usually quite distant from the cell body, where the recording electrode makes electrical contact. Three different types of giant synaptic preparations have proven useful in this regard. The first direct real-time measurements of vesicle retrieval following a burst of exocytosis were made in retinal bipolar cells (von Gersdorff & Matthews 1994a) and auditory hair cells (Parsons et al. 1994). Refinement of these measurements (Beutner et al. 2001, Neves & Lagnado 1999) revealed that, generally, two distinct kinetic components of endocytosis are apparent (with ∼1-s and ∼10-s time constants, respectively) where the relative proportion of retrieval via the faster component was sensitive to calcium influx (Beutner et al. 2001, Neves et al. 2001). Given that this calcium-driven fast component was sensitive to the calcium chelator BAPTA (and occurred even when calcium elevations were spatially confined), the authors concluded that the faster form was likely occurring near active zones. Similar observations were made in the giant auditory brainstem calyx of the Held synapse (Wu et al. 2005). The appearance of distinct kinetic components leads to the question of how each participates in regenerating SVs. Interestingly, the detailed ultrastructural kinetic study by Heuser and colleagues (Miller & Heuser 1984) also revealed ultrastructural components with distinct kinetic signatures following stimulation at the NMJ. Freeze-fracture views of presynaptic terminal membrane showed that following stimulation, non-clathrin-coated endocytic vacuole-like structures formed near the active zone without concentrating cargo molecules and disappeared within approximately 1 s (Figure 3a,b). In retrospect, it seems likely that the fast component seen with capacitance measurements could be the same fast pathway identified by Heuser at the NMJ. Consistent with this interpretation, this component was shown to be insensitive to clathrin perturbation ( Jockusch et al. 2005). At the calyx of Held, however, the appearance of fast endocytosis was independent of neurotransmitter release (Yamashita et al. 2005). Pretreatment of nerve terminals with botulinum toxin blocked neurotransmitter release without eliminating a capacitance transient with fast recovery. Thus, this rapid membrane response may represent a homeostatic response to acute calcium elevation independent of SV endocytosis. www.annualreviews.org • Synaptic Vesicle Endocytosis NMJ: neuromuscular junction 137 ANRV389-CB25-07 ARI 4 September 2009 17:25 a 8 b Endocytosis profiles (μm–2) 6 5 4 3 2 1 0 0 c 20 40 60 80 100 120 Time (s) tdwell ~ 0 s d tdwell ~ 4 s 0.20 Fraction of events Annu. Rev. Cell Dev. Biol. 2009.25:133-160. Downloaded from arjournals.annualreviews.org by WEILL MEDICAL COLLEGE OF CORNELL UNIVERSITY on 10/29/09. For personal use only. 7 tdwell ~ 10 s 5 0.10 0.05 tdwell > 22 s 0 0.15 10 15 Time (s) 20 0.00 0.1 5.6 11.1 16.6 22.1 Dwell time (s) Figure 3 Kinetics of endocytosis from 1984 to 2007 (a) The first measurements of the kinetics of synaptic vesicle (SV) endocytosis were obtained via pulse-chase freeze fracture experiments at the frog neuromuscular junction (NMJ) by Miller & Heuser (1984). An image that was obtained by quick freezing an NMJ 30 s after action potential stimulation in 4-AP is shown. The white circles (diameter of 110 nm) indicate membrane deformations that correspond to clathrin-coated pits. Image adapted courtesy of John Heuser. (b) The histogram of the accumulation and disappearance of the membrane invaginations obtained by examining freeze-fracture pictures taken at many different times with respect to the action potential stimulus. Redrawn from Miller & Heuser (1984). (c) Single-vesicle exocytosis and endocytosis as revealed by following the vesicular glutamate transporter (vGlut1) tagged with pHluorin in a luminal loop. Adapted from Balaji & Ryan (2007). Upon fusion, pHluorin becomes deprotonated and its fluorescence dequenched. After a variable delay (tdwell ), the SV protein is endocytosed and the fluorescence becomes quenched as the vesicle lumen reacidifies over a 3- to 4-s time scale. Four different examples displaying the range of tdwell are shown. (d ) The histogram of tdwell values obtained from approximately 200 events shows that the timing of endocytosis is dictated by an exponential process (red curve) whose mean time is approximately 13 s. 138 Dittman · Ryan ANRV389-CB25-07 ARI 4 September 2009 17:25 Endocytosis Annu. Rev. Cell Dev. Biol. 2009.25:133-160. Downloaded from arjournals.annualreviews.org by WEILL MEDICAL COLLEGE OF CORNELL UNIVERSITY on 10/29/09. For personal use only. Exocytosis V-ATPase proton pump Reacidification pKa ~7.1 Protonated (quenched) vGlut-pHluorin Deprotonated (fluorescent) vGlut-pHluorin Figure 4 pHluorin-based measurements of the vesicle cycle. pHluorin is a modified form of green fluorescent protein (GFP). GFPs, in general, are excellent proton sensors, as the fluorescence is quenched in an all-or-none fashion upon protonation, which is well described by a 1-proton binding equilibrium (Haupts et al. 1998). The pKa of pHluorin is ∼7.1, one log unit more alkaline than GFP and, when resident within the acidic lumen of a synaptic vesicle, is quenched ∼97% of the time. Redrawn from Sankaranarayanan et al. (2000). The introduction of optical tracers has proven particularly powerful for examining endocytosis at small nerve terminals. Pulse-chase application of the fluorescent amphipathic molecule FM 1-43 (Betz & Bewick 1992) allowed the first kinetic dissection of endocytosis in dissociated hippocampal neurons (Ryan et al. 1993, Ryan & Smith 1995). Vesicle retrieval following a large burst of activity indicated that endocytosis decayed over a 60-s time scale. Although useful for labeling recycling vesicles pools and measuring pool turnover kinetics, tracers such as FM 1-43 do not directly provide real-time endocytosis readouts. The advent of pHluorins (see Figure 4) provided higherfidelity readout where specific vesicle proteins fates can be followed. Initial measurements of VAMP2 endocytosis for relatively large stimuli showed that the recovery time scale depended on the amount of exocytosis (Sankaranarayanan & Ryan 2000). Improvements in these approaches allowed detection of endocytosis following single-action-potential stimuli (Balaji & Ryan 2007, Granseth et al. 2006), which showed endocytic recovery occurred with a time constant of approximately 15 s. Analysis of the delay time between exocytosis and endocytosis for individual vesicles showed that endocytosis times varied stochastically from vesicle to vesicle, ranging from instantaneous to tens of seconds with an exponentially distributed dwell time (Balaji & Ryan 2007), which indicates that a single rate-limiting stochastic step determines the time scale of endocytosis. www.annualreviews.org • Synaptic Vesicle Endocytosis 139 ANRV389-CB25-07 ARI 4 September 2009 17:25 MODULATION OF ENDOCYTOSIS: CALCIUM AND EXOCYTIC LOAD Annu. Rev. Cell Dev. Biol. 2009.25:133-160. Downloaded from arjournals.annualreviews.org by WEILL MEDICAL COLLEGE OF CORNELL UNIVERSITY on 10/29/09. For personal use only. A central goal of research in presynaptic function is to understand the regulation of different vesicle cycling steps. Prior to assigning specific molecular roles, one must understand how the conditions under which the measurements were performed impact the biophysical observations. Two main variables associated with driving exocytosis at synapses are the extent to which intracellular calcium becomes elevated and the total endocytic burden that ensues from successful exocytosis. These two variables are closely entwined as greater elevations in intracellular calcium generally lead to higher exocytic rates, more material that must be recaptured. Given the importance of intracellular calcium in signaling, determining calcium’s role in endocytosis is of significant interest. Early attempts to disentangle calcium elevations from exocytosis used α-latrotoxin, a potent secretagog from black widow spider venom, to stimulate exocytosis in the absence of extracellular calcium (Ceccarelli & Hurlbut 1980). After several hours of stimulation under these conditions, NMJs became severely depleted of SVs and the plasma membrane developed large enfoldings, indicating that endocytosis was blocked. Ceccarelli & Hurlbut (1980) concluded that intracellular calcium elevation was necessary for SV endocytosis. It was impossible, however, to distinguish the extent to which calcium might simply be modulating rates as opposed to playing a requisite role in the biochemistry of endocytosis. A number of different experiments have shown that, following exocytosis, calcium entry and endocytosis can be decoupled as the presence of extracellular calcium is no longer necessary for endocytosis to proceed (Gad et al. 1998, Ryan et al. 1996) and that bulk calcium levels typically decay faster than the completion of endocytosis (Wu & Betz 1996). At the other extreme, persistent photolysis of caged calcium led to complete arrest of endocytosis in giant bipolar terminals so long as bulk cytoplasmic calcium was greater than approximately 140 Dittman · Ryan 1 μM (von Gersdorff & Matthews 1994b). Although peak intracellular calcium levels near exocytosis sites transiently reach tens of micromolar, average sustained intracellular calcium levels remain well below 1 μM at most nerve terminals following a single-action potential or low-frequency stimulation. Thus, it is unclear if such inhibition plays a role under physiological conditions. Approaches that allowed direct measurements of endocytosis kinetics made it possible to examine the relationship between the amount of exocytosis and the speed of endocytosis, as well as how elevations in intracellular calcium might modulate this process. Experiments in hippocampal nerve terminals and at the calyx of Held showed that increases in exocytosis led to progressively longer endocytosis time scales (Sankaranarayanan et al. 2000, Sun et al. 2002), in spite of the fact that endocytosis was accelerated under conditions that led to elevated calcium in these preparations (Sankaranarayanan & Ryan 2001, Wu et al. 2005). This slowing of endocytosis with greater exocytosis may result from an underlying saturation of the total endocytic capacity of the synapse. It is notable that such saturation behavior is not observed in other synaptic preparations such as NMJs in mice (Tabares et al. 2007) and flies (Poskanzer et al. 2006), suggesting that it is not a fundamental property of endocytosis at nerve terminals. This paradox between acceleration of endocytosis due to intracellular calcium versus slowing due to increased accumulation, and the absence of either in certain preparations, was recently resolved. Taking advantage of single-vesicle sensitivity to monitor endocytosis kinetics systematically over a 40-fold range in exocytosis, Balaji et al. (2008) demonstrated that endocytosis recovery follows a single exponential time course (15 s) over a wide range of exocytosis but begins to slow significantly only above a threshold accumulation of SVs on the plasma membrane. These data imply that until this threshold is reached, endocytosis of different vesicles is likely happening in parallel, each with an Annu. Rev. Cell Dev. Biol. 2009.25:133-160. Downloaded from arjournals.annualreviews.org by WEILL MEDICAL COLLEGE OF CORNELL UNIVERSITY on 10/29/09. For personal use only. ANRV389-CB25-07 ARI 4 September 2009 17:25 average time scale of 15 s. Beyond this threshold, the synapse can no longer assemble sufficient endocytic machinery to handle greater endocytic loads, thereby defining the endocytic capacity of the synapse. Importantly, for sufficiently small amounts of exocytosis, the endocytosis time course was insensitive to manipulations of intracellular calcium. Balaji et al. (2008) observed that the endocytic capacity was increased by stimulus conditions that elevated residual calcium in the nerve terminal. These data thus help reconcile the original observations at the frog NMJ as well. Persistent stimulation in the absence of any calcium elevation (as achieved in α-latrotoxin in zero calcium for prolonged periods) could lead to a situation where the amount of exocytosis occurring greatly exceeds the endocytic capacity, eventually leading to persistent depletion of vesicle pools. Furthermore, this hypothesis offers an explanation as to why modulation of endocytosis by activity may not be observed: The detailed tuning of calcium entry, buffering, and extrusion, along with the specific abundance of the necessary endocytic proteins, may be such that one never operates in an exocytosis regime where the endocytic capacity becomes limiting. For example, at physiological temperatures, the time constant for endocytosis decreases to 6 s (Balaji & Ryan 2007) in hippocampal nerve terminals. This increase in clearance time coupled with the fact that release probability is lower at this temperature lead to a situation where the endocytic capacity is rarely exceeded. Thus, for three different central nervous system synapses examined (bipolar terminals, the calyx of Held, and dissociated hippocampal neurons), a common time scale of 10 to 15 s (at room temperature) appears to prevail in SV recovery unless significant accumulation of SV components can be driven to the cell surface. KISS-AND-RUN: A LONG-RUNNING DEBATE Beginning with the first observations of vesicle recycling, there has been speculation of a rapid recycling pathway whereby the vesicle maintains its identity and is retrieved intact at the active zone. This idea was originally fueled by disparity in the time scales of exo- and endocytosis observed at the NMJ. Endocytosis took approximately 1 min (Miller & Heuser 1984), whereas exocytosis took place on the submillisecond time scale. In comparing the time scale for exo- and endocytosis, however, one should consider the time for the entire recycling vesicle pool to fuse with the plasma membrane, not just a single vesicle. At hippocampal synapses at 37◦ C, exocytosis of the entire recycling pool requires approximately 20 s for stimulation at 10 Hz (Fernandez-Alfonso & Ryan 2004), which should be compared to the 6-s time constant for endocytosis under these conditions (Balaji et al. 2008). Thus, it is possible for exocytosis to outrun endocytosis, but only under relatively high-frequency stimulation. The most direct evidence for transient reversible fusion pores arose from capacitance recordings of single-vesicle fusion events in chromaffin cells coupled with simultaneous measurements of release of catecholamine using carbon fiber amperometry (Albillos et al. 1997). In the majority of events, the appearance of a small fusion pore was followed by an irreversible expansion and complete loss of catecholamine. However, in 5–10% of events, the pore closed without expanding. At conventional synapses, this dual approach of capacitance recordings and amperometry has not been possible, but single-vesicle fusion events have been recorded using capacitance measurements alone at the calyx of Held. Wu and colleagues reported that 17% of events appeared to be transient in nature (He et al. 2006). There are two important caveats in interpreting these results. First, this approach required examining exocytosis at random locations on the synaptic surface that were not necessarily active zones. For technical reasons, the events could be evoked only using nonphysiological stimuli. Second, the appearance of exocytosis followed soon after by endocytosis may simply represent a narrow sampling of the select fast events of an exponential distribution of time scales as predicted by a stochastic process (Balaji & Ryan www.annualreviews.org • Synaptic Vesicle Endocytosis 141 ANRV389-CB25-07 ARI 4 September 2009 Annu. Rev. Cell Dev. Biol. 2009.25:133-160. Downloaded from arjournals.annualreviews.org by WEILL MEDICAL COLLEGE OF CORNELL UNIVERSITY on 10/29/09. For personal use only. Qdot: quantum dot 142 17:25 2007). Verification that such events are insensitive to perturbations of elements in the clathrinmediated pathway will be necessary to assign them to a distinct endocytosis pathway. Less direct biophysical methods have also been used to ascertain the presence of kiss-andrun. One approach that has been used extensively at hippocampal synapses was to examine the efficiency of escape of lipophilic tracers from SVs during exocytosis. The idea is that a liphophilic dye potentially would dissociate from the vesicle lumen membrane too slowly to escape during a kiss-and-run event, but that it might fully dissociate during a full fusion event. This approach, first introduced more than a decade ago (Ryan et al. 1996), has been used in one variant or another to argue both for (Aravanis et al. 2003, Harata et al. 2006, Klingauf et al. 1998, Pyle et al. 2000, Richards et al. 2005) and against (Fernandez-Alfonso & Ryan 2004, Ryan et al. 1996, Zenisek et al. 2002) the presence of kiss-and-run. Examination of dye loss at the single-vesicle level, however, is quite challenging, as one is effectively asking whether losses in fluorescence are well quantized. A number of subtle artifacts can potentially lead to loss of fidelity of the fluorescence measurements. Chief among these is that lipophilic tracers are added to the outside of the synapse during activity to load recycling vesicles; however, other recycling membranes in neighboring glia and postsynaptic dendrites, as well as other presynaptic compartments, can become labeled. Such spurious labeling can strongly contribute to background fluctuations in fluorescence intensity. Recent experiments designed to control for some of these possible complications concluded that lipophilic dyes destain completely during action-potential stimuli (Chen et al. 2008) and suggest either that kiss-and-run does not occur at these synapses or that one cannot use these dyes reliably to detect such transient fusions. Recently, a novel form of molecular tracer was introduced for examining the frequency of full fusion during exocytosis in cultured neurons. In this new technique, a recycling vesicle endocytoses a quantum dot (Qdot) (Zhang et al. 2007) whose optical properties allow Dittman · Ryan continuous optical recordings without photobleaching. Because of its large size [hydrodynamic radius of nearly 15 nm (Larson et al. 2003)], the internalized Qdot escapes only during full-fusion exocytosis (Zhang et al. 2007). Zhang et al. (2007) found that stimulusdependent unloading of Qdots followed a slower time course than an FM dye loaded into the same synapses, suggesting that some fusion events release FM dye but not Qdots. A significant caveat with this approach comes from the large size of Qdots relative to the vesicles that contain them. If the Qdot interacts with the vesicle lumen and stabilizes the curvature of a fusing vesicle, perhaps an otherwise rare partialfusion event could be made more probable. Investigations into the molecules that underlie endocytosis have yielded little evidence in support of kiss-and-run mechanisms to date (see below). Furthermore, the observation that vesicle components exchange with counterparts resident on the cell surface during endocytosis over a wide range of stimulus conditions (Fernandez-Alfonso et al. 2006, Wienisch & Klingauf 2006) indicates that vesicles typically lose some of their components following exocytosis. Finally, detailed measurements of the time between exocytosis and endocytosis for individual vesicles showed that, although vesicles do occasionally endocytose quickly, the frequency of such events is well predicted by a stochastic (single-exponential) process with a mean endocytic time of 15 s (Balaji & Ryan 2007). Taken together, the biophysical evidence weighs largely against significant vesicle recycling via a kiss-and-run mechanism. Furthermore, as discussed below, most experiments aimed at explicitly targeting clathrin or associated machinery at the synapse support a major role for the classical endocytic pathway in vesicle retrieval. THE MOLECULAR MACHINERY OF SYNAPTIC VESICLE ENDOCYTOSIS Molecular and genetic insights over the past 25 years from a variety of model systems have Annu. Rev. Cell Dev. Biol. 2009.25:133-160. Downloaded from arjournals.annualreviews.org by WEILL MEDICAL COLLEGE OF CORNELL UNIVERSITY on 10/29/09. For personal use only. ANRV389-CB25-07 ARI 4 September 2009 17:25 culminated in a formidable list of potential proteins and lipids involved in endocytosis. Many of the proteins share common binding partners and exhibit overlapping functions within the endocytosis machinery. This property of interconnectivity may be important in explaining an emerging dilemma in the field of SV endocytosis: Gene knockouts and RNA interference knockdown of endocytic proteins generally do not eliminate SV endocytosis and, in many cases, have remarkably subtle effects (Di Paolo et al. 2002, Ferguson et al. 2007, Gu et al. 2008, Sato et al. 2009). This situation starkly contrasts with SV exocytosis, where removal of any one of a small collection of essential proteins eliminates SV fusion (Südhof 2004). If altered SV endocytosis is observed after a particular endocytic protein is removed, there are three possible considerations for the residual endocytosis. First, this perturbation may uncover a second, independent pathway with distinct underlying molecular mechanisms. Second, the measured endocytosis is identical to the wild-type process except that a functional paralog has taken the place of the deleted molecule, although it may not function with the same efficacy. Third, the impaired endocytosis proceeds via the identical molecular pathway but with lower efficacy because the deleted molecule played a contributory rather than obligatory role in the process. In this case, the network of endocytic proteins is robust to deletions of individual members. Consistent with the third scenario, experiments to date have demonstrated that no single protein appears to be absolutely essential to the process of SV endocytosis. Perhaps particular components of the synaptic endocytic network play a larger or smaller role in different animal phyla, but all the components are generally found in presynaptic specializations of metazoa. Furthermore, these components show a high degree of conservation, both at the sequence level and in terms of interactions with other proteins and phospholipids (Lloyd et al. 2000). In the following section of this review, we introduce 15 proteins/protein complexes that compose the SV endocytic network (Figures 6 and 7). These proteins are separated into three layers of the network on the basis of their function, binding interactions, and protein domain content. At the core of the endocytic network is the capacity to gather up cargo into a local patch of plasma membrane and deform this patch into a separate compartment destined for internalization. A second layer of proteins acts to stabilize the core proteins while recruiting additional effector molecules that both catalyze and terminate the process. These effectors make up the third layer, and they function in numerous cellular trafficking processes in addition to SV endocytosis. A CLATHRIN-MEDIATED PATHWAY FOR VESICLE REBUILDING The first details of the molecular basis of SV recycling arose with the pioneering studies of Heuser & Reese (1973), who noted that endocytosis following stimulation appeared to occur via the newly discovered clathrin coats. Since these early discoveries, progress in our understanding of the molecular basis of SV recycling has benefited from a number of approaches in different model systems. Although the relevance of the finding that clathrin-coated pits could be found at synapses originally was debated, the evidence that this pathway predominates as the route of vesicle recovery has become overwhelmingly strong in the past 10 years. Three types of experiments have been performed, all of which support a major role for the clathrin reuptake pathway. First, microinjections of peptides or antibodies that interfere with different steps of endocytosis were followed by electron microscopy and functional assays. Second, genetic ablations of endocytic proteins were followed by kinetic assays and electron microscopy. Third, the effects of photoinactivation of clathrin in Drosophila NMJs were followed by functional and ultrastructural assays. At synapses such as in the lamprey giant reticulospinal axons, injections of peptides and antibodies designed to interrupt dynamin function (see below), followed by activity-driven www.annualreviews.org • Synaptic Vesicle Endocytosis 143 ANRV389-CB25-07 ARI 4 September 2009 17:25 a Annu. Rev. Cell Dev. Biol. 2009.25:133-160. Downloaded from arjournals.annualreviews.org by WEILL MEDICAL COLLEGE OF CORNELL UNIVERSITY on 10/29/09. For personal use only. * 200 nm b Clathrin-coated pits SVs SVs Plasma membrane Figure 5 The accumulation of clathrin-coated pits at nerve terminals during activity in the absence of dynamin-1. Strong evidence for synaptic vesicle (SV) endocytosis normally proceeding through a clathrin-coated pit pathway has been obtained from a number of experiments in which molecular perturbations led to trapping vesicle endocytosis at an intermediate stage. Here, an example of the ultrastructure of a hippocampal synapse derived from a dynamin-1 knockout mouse taken from Ferguson et al. (2007) is shown. (a) Single section of a tomogram taken from a synapse where spontaneous activity led to the development of profound invaginations studded with clathrin-coated profiles. The small arrows indicate the presence of interconnected clathrin-coated buds, the asterisk shows an evagination of an adjacent cell into this nerve terminal, and the large arrow shows a small cluster of heterogeneously sized synaptic vesicles. (b) 3D tomographic reconstruction shows that these clathrin-coated profiles were associated with plasma membrane invagination. vGlut: vesicular glutamate transporter 144 exocytosis, led to accumulation of clathrincoated buds (Shupliakov et al. 1997). Similar accumulations were observed in hippocampal neurons when the abundant brain-specific isoform, dynamin-1, was genetically ablated (Figure 5). Additionally, peptide injections designed to interfere with clathrin assembly led to dramatic slowing of the kinetics of endocytosis ( Jockusch et al. 2005) as well as to activitydependent interruption of exocytosis (Gad et al. Dittman · Ryan 2000; Morgan et al. 2000, 2003), consistent with a depletion of available vesicles due to a block in endocytosis. At hippocampal synapses, the kinetics of endocytosis can be dramatically slowed when clathrin is removed by siRNA (Granseth et al. 2006). Finally, photoinactivation of fluorescently labeled clathrin, which offers the most temporally precise molecular interruption, showed that acutely inactivating clathrin’s inability to assemble in Drosophila NMJs undergoing exocytosis leads to the formation of large dead-end vacuoles (Heerssen et al. 2008, Kasprowicz et al. 2008) and the loss of any further vesicle recycling. Thus, data using several distinct approaches and examining multiple different types of synapses all argue that clathrin assembly normally plays a critical role in SV endocytosis. MECHANISMS FOR SORTING THE APPROXIMATELY NINE CARGO PROTEIN TYPES An important unsolved question in SV recycling is, how is each of the nine different types of SV transmembrane proteins resorted into SVs following exocytosis? As retrieval occurs through clathrin-mediated endocytosis, the expectation is that known mechanisms for receptor-mediated endocytosis will be manifest here. For instance, perhaps there are interactions of sorting motifs in vesicle protein cytoplasmic tails with the plasma membrane adaptor protein complex AP2. AP2, a heterotetrameric complex that additionally binds clathrin, is thought to coordinate clathrin assembly with cargo recognition during endocytosis (Keen 1987). At present, only a single putative sorting motif has been functionally identified in SV proteins. Mutation of a dileucine-like motif in the vesicular glutamate transporter (vGlut1) slows vGlut1 internalization at nerve terminals (Voglmaier et al. 2006). Synaptotagmin has also been identified as a binding partner to AP2 (Grass et al. 2004, Haucke & De Camilli 1999, Haucke & Krauss 2002, Haucke et al. 2000, Zhang et al. 1994). In the absence of Synaptotagmin 1 ( Jorgensen et al. 1995, Annu. Rev. Cell Dev. Biol. 2009.25:133-160. Downloaded from arjournals.annualreviews.org by WEILL MEDICAL COLLEGE OF CORNELL UNIVERSITY on 10/29/09. For personal use only. ANRV389-CB25-07 ARI 4 September 2009 17:25 Nicholson-Tomishima & Ryan 2004, Reist et al. 1998), or following either photoinactivation (Poskanzer et al. 2003) or mutation in residues in its C2B domain (Poskanzer et al. 2006), endocytosis of SVs is impaired. Surprisingly, deletion of the AP2 μ2 subunit fails to eliminate cargo retrieval and SV recycling in both Caenorhabditis elegans (Gu et al. 2008) and dissociated hippocampal neurons (Kim & Ryan 2009). In the absence of μ2 , however, there was a significant loss of SVs and endocytosis kinetics were measurably slower, implying that AP2 normally does play a role at synapses. These results imply that additional sorting mechanisms must be operating and open up the possibility that AP2 serves a more subtle role in SV endocytosis than originally anticipated. A leading candidate adaptor for dedicated SV protein sorting is stonin, first identified as a temperature-sensitive paralytic mutant in Drosophila (where it is named Stoned ) more than 35 years ago (Grigliatti et al. 1973, Kelly 1983). Stonin 2, which contains a μ2 homology domain (Figure 6), interacts genetically (Fergestad & Broadie 2001, Phillips et al. 2000) and biochemically ( Jung et al. 2007, Walther et al. 2004) with synaptotagmin. Removal of Stoned B in Drosophila NMJs leads to defects in endocytosis (Stimson et al. 2001) and, when coexpressed with synaptotagmin in nonneuronal cells, it facilitates synaptotagmin retrieval (Diril et al. 2006). Evidence for defects in cargo sorting in SV endocytosis has also emerged from examination of mutations in proteins associated with clathrin assembly. In particular, deletion of the monomeric adaptor protein AP180 led to mislocalization of VAMP2 but not synaptotagmin or synaptogyrin (Nonet et al. 1999, Zhang et al. 1998), suggesting a specific role in sorting VAMP2 during endocytosis. In general, these clathrin-associated sorting proteins usually associate with both AP2 and clathrin for proper function (Brett & Traub 2006). The fact that endocytosis and protein sorting can occur without AP2 suggests that redundant mechanisms that link sorting proteins, cargo, and coated pits must be operational. One proposed scenario for efficiently recapturing SV proteins is that they remain stably associated after exocytosis and directly nucleate clathrin-coat formation. Although this may occur for certain SV proteins, this is not the case for VAMP2 or synaptotagmin. At hippocampal nerve terminals, these proteins diffuse onto the axonal surface (Li & Murthy 2001, Sankaranarayanan & Ryan 2000), and they exchange with surface counterparts during endocytosis (Fernandez-Alfonso et al. 2006, Wienisch & Klingauf 2006). Currently, the issue of how SV proteins are retargeted into SVs during endocytosis remains largely unresolved. PIP2 : phosphoinositol-(4,5)-bisphosphate CURVING MEMBRANES: INITIATION OF SYNAPTIC VESICLE ENDOCYTOSIS In addition to recollecting and sorting SV proteins, the endocytosis machinery must locally curve the plasma membrane and eventually pinch off 3000 to 5000 nm2 of bilayer in the form of a 30- to 40-nm-diameter vesicle. This process can be broken into two parts: deformation of membrane into a vesicular or tubular shape and scission of the stalk connecting this structure to the plasma membrane. The process of membrane deformation begins with the organization of endocytic coat proteins on the inner leaflet of the synaptic membrane. This assembly is thought to be driven in part by the generation of phosphoinositol-(4,5)-bis-phosphate (PIP2 ), which recruits several endocytic proteins to the plasma membrane including AP2, epsin, dynamin, endophilin, amphiphysin, and AP180. At synapses, activity-dependent PIP2 synthesis is largely carried out by phospho-inositol-5kinase Iγ (Di Paolo et al. 2004), which presumably drives rapid recruitment of endocytic factors during repetitive exocytosis. In addition to providing a scaffold for concentrating appropriate cargo proteins destined for endocytosis, endocytic coat proteins are thought to regulate the area destined for internalization in a manner that leads to a precise SV size. www.annualreviews.org • Synaptic Vesicle Endocytosis 145 ANTH N NPF AP2/Clath Positive feedback + Avidity + + + + SH3 C C C MHD C + + + + + • Cargo binding • Curvature SHD Dynamic properties N-BAR Amphiphysin N AP2/Clath ENTH AP180 N Epsin N NPF NPF Stonin 2 NPF AP2/Clath Clathrin NPF EH EH EH EH N-BAR F-BAR EH EH Sac1p N Stonin 1 N ANTH EVH1 CALM N N-WASP N PH EH GBD/CRIB 5' p’ase Synaptojanin N GTPase Dynamin N Syndapin N Endophilin N Eps15 N Intersectin 1L N Intersectin 1S NPF Primary scaffold Primary effectors AP2/Clath SH3 GED NPF AP2/Clath NPF Core module C PRD C C-C SHD PRD PRD SH3 C-C C-C MHD C SH3 SH3 C VCA C BARS SH3 SH3 NPF AP2 NPF AP2/Clath C C C SH3 SH3 PH + + + + DH C C2 C + + + + + + + + + + Secondary effectors SH3 SH3 PRDs SH3 SH3 Secondary scaffolds Annu. Rev. Cell Dev. Biol. 2009.25:133-160. Downloaded from arjournals.annualreviews.org by WEILL MEDICAL COLLEGE OF CORNELL UNIVERSITY on 10/29/09. For personal use only. NPF Ryan NPF · NPF AP2/Clath Dittman NPF 146 AP2/Clath GDP + Pi GTP C-C SH3 EH C-C PRD Protein-protein interactions F-BAR C2 ENTH N-BAR PH ANTH Lipid-binding domains GTPase 5' p’ase PI(4)P + Pi Sac1p PIP2 Enzymatic domains Domain properties 4 September 2009 AP2/Clath ARI NPF ANRV389-CB25-07 17:25 Annu. Rev. Cell Dev. Biol. 2009.25:133-160. Downloaded from arjournals.annualreviews.org by WEILL MEDICAL COLLEGE OF CORNELL UNIVERSITY on 10/29/09. For personal use only. ANRV389-CB25-07 ARI 4 September 2009 17:25 On the basis of biochemical, structural, and genetic evidence, several protein domains have been implicated in the curving of membranes. ENTH (epsin N-terminal homology) domains found at the amino termini of epsin, AP180, and CALM are phospholipid-binding modules (Hurley 2006, Itoh & De Camilli 2006). ENTH domains interact preferentially with PIP2 -containing phospholipids and tubulate liposomes in vitro as well as plasma membranes of living cells when overexpressed (Ford et al. 2002, Itoh et al. 2001). Structural studies of ENTH domains suggest that an Nterminal amphipathic alpha helix inserts into the lipid bilayer upon binding to PIP2 (Itoh et al. 2001). The canonical example of an ENTH domain protein at the synapse is epsin. Thought to function as a clathrin adaptor, epsin binds clathrin heavy chain and AP2. These associations position epsin as part of the endocytic core module. Epsin contains several NPF (asparagine-proline-phenylalanine) motifs near its carboxy terminus that interact with Eps15 homology (EH) domain modules found in the secondary scaffolds Eps15 and intersectin/Dap160 (Figure 6). Epsin also contains ubiquitin-interacting motifs, which may function in the recognition of ubiquitinated cargo proteins. In yeast, expression of the ENTH domain is sufficient to rescue viability of the Ent1/2 double mutant (the two yeast epsin orthologs), suggesting that this domain has an essential role irrespective of its adaptor functions (Wendland et al. 1999). Under some conditions, epsin and clathrin triskelia, but not clathrin alone, can invaginate lipid monolayers (Ford et al. 2002). Disruption of epsin function by antibody binding in the lamprey giant synapse decreased SV numbers and led to an accumulation of large coated pits following stimulation ( Jakobsson et al. 2008). These data suggested that SV endocytosis was trapped at an early stage in the absence of functional epsin. In contrast, loss of the Drosophila epsin ortholog Liquid Facets did not appear to impair SV endocytosis, although synapse architecture was significantly affected (Bao et al. 2008). Epsin is also found in other cellular compartments, suggesting that it may have a general role in clathrin-mediated endocytosis (Horvath et al. 2007). AP180 and CALM share an ENTH-like module at their amino termini along with clathrin and AP2-interacting domains within their unstructured carboxy termini. However, interaction of these proteins with PIP2 is distinct from epsin’s ENTH domain interaction and the N-terminal domain contains additional alpha helices (Itoh & De Camilli 2006). These biochemical and structural differences led investigators to call these ANTH (AP180 N-terminal homology) domains. ANTH domains do not appear to deform lipids on their own, but they may influence curvature indirectly through interactions with other membrane-binding proteins. AP180 is predominantly found at presynaptic terminals, whereas CALM is ubiquitous and also possesses multiple NPF motifs (which are not found in AP180) near its carboxy terminus (Itoh & De Camilli 2006). C. elegans and Drosophila each have a single AP180/CALM ortholog ←−−−−−−−−−−−−−−−−−−−−−−−−−−−−−−−−−−−−−−−−−−−−−−−−−−−−−−−−−−−−−−−−−−−−−−−−−−−−−−−−−−−−−−−−− Figure 6 Categories of endocytosis proteins. Schematic diagrams of protein domain organization are shown for the core module (AP2, clathrin, stonin 2, epsin, AP180, and amphiphysin), secondary scaffolds (intersectin and Eps15), and secondary effectors (endophilin, syndapin, dynamin, synaptojanin, N-WASP, CALM, and stonin 1). The domain abbreviations are as follows: PH, pleckstrin homology domain; ANTH, AP180 N-terminal homology domain; ENTH, epsin N-terminal homology domain; BAR, bin amphiphysin rvs; EH, eps15 homology domain; PRD, proline-rich domain; C-C, coiled-coil domain. NPF represents any of five possible motifs that interact with EH: NPF, WW, FW, SWG, and H(ST)F. AP2/Clath represents two AP2-binding motifs [WXXF and DP(WF)] and three clathrinbinding motifs [D(LI)(LFQ), L(FWY)X(FWY)(DE), and PWXXW]. The diagram depicts these as one localized domain, but these motifs are actually distributed throughout the polypeptide. Two isoforms of intersectin are shown: long and short. Dynamic properties are depicted as either positive feedback (polymerization, enzymatic catalysis) or avidity (multiple binding sites linked together). www.annualreviews.org • Synaptic Vesicle Endocytosis 147 ARI 4 September 2009 17:25 that presumably fulfills the duties of both proteins. In both of these model systems, loss of AP180/CALM substantially broadened the distribution of SV sizes and increased the mean diameter by over 30% (Nonet et al. 1999, Zhang et al. 1998). In addition, AP180 and CALM regulate the plasma membrane abundance of synaptobrevin (Dittman & Kaplan 2006, Harel et al. 2008, Nonet et al. 1999, Zhang et al. 1998), suggesting that ANTH domain proteins play a role in both cargo recruitment and size of the endocytic area destined to undergo endocytosis. Another membrane binding module employed during SV endocytosis is the BAR (Bin, amphiphysin, Rvs) domain (Figure 6). BAR domains found in endophilin, amphiphysin, syndapin, and sorting nexin 9, among others, bind phospholipids as crescent-shaped dimers, and they are thought to generate or stabilize highly curved regions of the membrane (Gallop & McMahon 2005, Itoh & De Camilli 2006, Ren et al. 2006). In vitro, these domains can tubulate liposomes and, in some cases, appear to bind preferentially to particular liposome sizes, suggesting that the BAR domain can sense lipid curvature (Gallop & McMahon 2005). Amphiphysin and endophilin share amino-terminal domains that form amphipathic helices capable of penetrating the cytoplasmic bilayer leaflet and perturbing lipid packing, particularly in the presence of acidic phospholipids. Thus, their N-terminal helix and BAR motif are together termed N-BAR domains. Both amphiphysin and endophilin contain carboxy-terminal SH3 domains, which are thought to interact with the proline-rich domains of dynamin and synaptojanin (Itoh & De Camilli 2006). The combination of N-BAR and SH3 domains makes these proteins ideally suited for the dual task of deforming membranes and recruiting other endocytic molecules to the curved region. Amphiphysin additionally has clathrin- and AP2-binding motifs, linking it to the core endocytic machinery. The mouse knockout of amphiphysin I displayed a modest decrease in recycling SV pool size as well as a slower recycling rate (Di Paolo Annu. Rev. Cell Dev. Biol. 2009.25:133-160. Downloaded from arjournals.annualreviews.org by WEILL MEDICAL COLLEGE OF CORNELL UNIVERSITY on 10/29/09. For personal use only. ANRV389-CB25-07 148 Dittman · Ryan et al. 2002). Fly mutants lacking amphiphysin did not appear to have significant transmission defects at the NMJ, and nervous system function is largely intact (Razzaq et al. 2001, Zelhof et al. 2001). Similarly, neither deletion nor RNA interference knockdown of the worm ortholog AMPH-1 impacts nervous system function appreciably ( J. Kaplan and J. Bai, personal communication). However, loss of endophilin significantly impairs synaptic transmission in both flies and worms (Rikhy et al. 2002, Schuske et al. 2003, Verstreken et al. 2002). The SV pool is decreased, whereas SV diameter and quantal size are increased. Ultrastructural similarities between endophilin and synaptojanin mutants, such as accumulations of clathrin-coated pits and vesicles, queues of vesicles far from the active zone, and large cisternae, support the hypothesis that endophilin and synaptojanin act together at a similar stage during endocytosis (Schuske et al. 2003, Verstreken et al. 2003). Syndapin, a member of another class of BAR domain known as F-BAR (formerly known as extended FCH) domain proteins, may play a role during SV endocytosis, particularly during prolonged or intense stimulation (Andersson et al. 2008). Also known as PACSIN, syndapin possesses an N-terminal F-BAR domain, several NPF motifs, and a C-terminal SH3 domain, which interacts with dynamin and N-WASP (Anggono & Robinson 2007, Kessels & Qualmann 2004). Syndapin binds to EH domain proteins via its NPF motifs and participates in trafficking through the recycling endosome pathway (Braun et al. 2005). Fly syndapin mutants failed to display any significant defects in synaptic transmission or SV endocytosis (Kumar et al. 2009). However, antibodies against syndapin caused enhanced synaptic depression and large VAMP2-containing cisternae after sustained stimulation when injected into lamprey giant synapses (Andersson et al. 2008). Because no effect was observed at low stimulus frequencies, syndapin may be part of a secondary pathway for retrieving SV components through endocytic intermediates following prolonged bouts of activity. ANRV389-CB25-07 ARI 4 September 2009 17:25 Annu. Rev. Cell Dev. Biol. 2009.25:133-160. Downloaded from arjournals.annualreviews.org by WEILL MEDICAL COLLEGE OF CORNELL UNIVERSITY on 10/29/09. For personal use only. SECONDARY SCAFFOLDING FOR RECRUITING GENERAL ENDOCYTIC EFFECTORS The proteins described thus far can be organized either into a core endocytic module (clathrin, AP2, AP180, stonin 2, epsin, amphiphysin) or into secondary effectors (endophilin and synaptojanin; others mentioned below) based on their interactions (or lack thereof ) with vesicle cargo molecules or cargo-binding proteins. Two synaptic proteins bridge this gap between the core module and secondary effectors, acting as recruiters and organizers during SV endocytosis. Eps15 and intersectin/Dap160 are binding partners and each has highly conserved domain structures coupling the endocytic coat to secondary effectors (Montesinos et al. 2005). Eps15 is a 110-kDa protein containing three N-terminal EH domains, a central coiled-coil domain, and clathrin/AP2 binding motifs near its carboxy terminus (Morgan et al. 2003, Salcini et al. 2001). The EH domains mediate interactions with NPF motifs in epsin, stonins, AP180, and synaptojanin, whereas the coiled coil interacts with intersectin (Kelly & Phillips 2005, Morgan et al. 2003). Loss of Eps15 in C. elegans results in decreased SV number, particularly at elevated temperatures (Salcini et al. 2001). The Eps15 mutant ehs-1(ok146) genetically interacts with the worm temperature-sensitive dynamin mutant dyn-1(ky51) such that locomotion is severely impaired in the double mutant even at permissive temperatures (Salcini et al. 2001). Eps15 also appears to enhance AP180-mediated clathrin assembly in vitro, suggesting that it may have an early role in building clathrin pits (Morgan et al. 2003). The loss of Eps15 in Drosophila causes substantial decrease in synaptic intersectin, dynamin, stonin, synaptotagmin I, α adaptin, and endophilin (Koh et al. 2007). Intersectin/Dap160 is a synaptic protein containing two N-terminal EH domains, a central coiled-coil domain, and four to five SH3 domains (depending on the species) at its carboxy terminus (Koh et al. 2004, Marie et al. 2004). A long isoform in vertebrates also contains Dbl homology domain, a pleckstrin homology domain, and a C2 domain after the SH3 repeat. This module can act as a guanine nucleotide exchange factor for Cdc42, regulating actin dynamics through Cdc42 and N-WASP (Hussain et al. 2001). Interestingly, Drosophila and C. elegans intersectin orthologs do not contain this module. The EH domains interact with epsin; the coiled coil binds Eps15; and the SH3 domains bind to dynamin, synaptojanin, and N-WASP (Marie et al. 2004, Montesinos et al. 2005). The loss of intersectin in the fly results in diminished SV endocytosis; decreased synaptic levels of AP180, endophilin, synaptojanin, and dynamin; and increased synaptic depression during prolonged stimulation (Koh et al. 2004, Marie et al. 2004). Eps15 intersectin double mutants exhibit synaptic depression and decreased FM1-43 uptake to a similar degree as either single mutant (Koh et al. 2007). Microinjection of the intersectin SH3 domain in lamprey giant synapses inhibits SV recycling and traps a large number of coated pits in the periactive zone (Evergren et al. 2007). Furthermore, intersectin appears to negatively regulate recruitment of dynamin to periactive zones at this synapse. In C. elegans, ITSN-1 localizes to the periactive zone in NMJs, and loss of ITSN-1 reveals a substantial increase in large, irregular vesicular structures as well as a decrease in spontaneous transmitter release (Rose et al. 2007, Wang et al. 2008). FINISHING THE JOB: MEMBRANE FISSION AND COAT DISASSEMBLY The final step of endocytosis involves severing the endocytic bud from the plasma membrane. In vitro studies have shown that when artificial membrane necks are narrowed to sufficiently small diameter, thermal fluctuations result in a collapse of the neck, presumably the starting point for fission (Bashkirov et al. 2008, Israelachvili 1992). Dynamin, a mechanochemical GTPase, was originally identified as the gene product of the Drosophila shibire mutant, a temperature-sensitive paralytic that becomes www.annualreviews.org • Synaptic Vesicle Endocytosis 149 ANRV389-CB25-07 ARI 4 September 2009 17:25 Clathrin/AP2 binding motif SH3–PRD interaction EH–NPF interaction Other protein interactions PI(4,5)P2 Phospholipid binding Cytoskeleton Annu. Rev. Cell Dev. Biol. 2009.25:133-160. Downloaded from arjournals.annualreviews.org by WEILL MEDICAL COLLEGE OF CORNELL UNIVERSITY on 10/29/09. For personal use only. Second scaffold Scaffold ? Core module Clathrin ? Cargo selector Membrane binding Secondary scaffold Secondary effectors Eps15, Intersectin Stonin 1, CALM Syndapin AP2, stonin 2, AP180 Dynamin N-WASP Epsin, amphiphysin Synaptojanin Endophilin ? Figure 7 Synaptic vesicle endocytic network. The proteins that compose the endocytic machinery at the synapse are classified in two ways: location in the network (color) and functionality (shape). Blue proteins make up the core module, red proteins are secondary scaffolding, and green proteins are secondary effectors. Proteins found exclusively at the synapse are shown in bold type. Scaffolds are hatched lines, cargo binders are triangles, membrane benders are crescents, and secondary effectors are diamonds. Interactions between proteins are depicted as arrow connectors for clathrin/AP2 binding, SH3 to PRD binding, Eps15 homology domain to NPF binding, and other protein interactions (see figure key for arrow types). Membrane interactions are shown as downward gray arrows, and the PIP2 phosphatase synaptojanin is shown interacting with PIP2 (red arrow). Endophilin ( purple) has no known interactions with either the core module or the secondary scaffold and is therefore distinguished from the other secondary effectors. devoid of SVs during activity at the nonpermissive temperature (Koenig & Ikeda 1989). This protein, which is recruited to the endocytic bud neck, is the minimal essential fission machinery in vitro, provided the membranes undergoing fission are under tension (Roux et al. 2006). Dynamin interacts with a number of SH3-containing proteins such as endophilin, amphiphysin, syndapin, and intersectin via its proline-rich domain (Figures 6 and 7). Together with other BAR domain proteins, dynamin also appears to coordinate the shaping 150 Dittman · Ryan of endocytic buds (Itoh et al. 2005). The evidence for dynamin’s role in endocytosis at the synapse is very strong as many dominantnegative strategies targeting dynamin block SV endocytosis (Koenig & Ikeda 1999, Newton et al. 2006, Yamashita et al. 2005). Surprisingly, genetic ablation of dynamin-1, which encodes ∼85% of brain dynamin, resulted in only partial defects in SV endocytosis. Mice lacking dynamin-1 survive about two weeks (Ferguson et al. 2007). Under these conditions, endocytosis appeared to be completely arrested Annu. Rev. Cell Dev. Biol. 2009.25:133-160. Downloaded from arjournals.annualreviews.org by WEILL MEDICAL COLLEGE OF CORNELL UNIVERSITY on 10/29/09. For personal use only. ANRV389-CB25-07 ARI 4 September 2009 17:25 only during intense activity, presumably owing to the elevation of intracellular calcium. However, soon after cessation of action potential firing, endocytosis resumed with remarkably normal kinetics. Although synapses lacking dynamin-1 in mouse still contain a small amount of dynamin-3 and even less dynamin-2 (Ferguson et al. 2007), one possible conclusion from this work is that the fission step in endocytosis is not catalyzed by the action of dynamin alone, but it is simply accelerated by this protein. Once the endocytic vesicle is severed from the plasma membrane, the newly sculpted vesicle must shed all the machinery that was used in rebuilding prior to reuse. Clathrin uncoating occurs via the concerted action of the ATPase Hsc70 and auxilin, which helps recruit this enzyme to the vesicle (Eisenberg & Greene 2007). One important mystery is the identity of the molecular trigger to initiate uncoating, as it would be energetically costly and perhaps detrimental to launch this process prior to fission. A mechanistic clue has arisen from genetic ablation studies of synaptojanin: The loss of this lipid phosphatase leads to a pronounced accumulation of clathrin-coated vesicles at nerve terminals as well as a delay in the availability of newly endocytosed vesicles for additional rounds of exocytosis (Cremona et al. 1999, Mani et al. 2007, Schuske et al. 2003, Verstreken et al. 2003). Synaptojanin is stabilized at endocytic buds through multiple interactions including binding to the SH3 domains of intersectin and endophilin and the EH domains of intersectin and Eps15, as well as direct binding to clathrin and AP2 (Haffner et al. 1997, Marie et al. 2004, Montesinos et al. 2005, Schuske et al. 2003, Verstreken et al. 2003). The 5 phosphatase domain at the amino terminus of synaptojanin rapidly dephosphorylates PIP2. This depletion of PIP2 decreases the phospholipid binding affinity of the core module and BAR proteins, simultaneously destabilizing the coat assembly. Another possible coupling between membrane fission and synaptojanin arises from the theoretical consideration that local depletion of PIP2 selectively on the vesicle bud causes a transient gradient of PIP2 between the bud and the plasma membrane to which it is connected. If this gradient contributes to a lipid phase separation between these compartments, the interfacial line between the neck and bud tends to be minimized, spontaneously constricting the neck (Liu et al. 2006). Regardless of the details of fission and uncoating, these processes are quite rapid as evidenced by the difficulty of capturing a coated vesicle, even when rapid freeze EM is used. CALCIUM SENSORS FOR ENDOCYTOSIS? As described, presynaptic calcium affects numerous aspects of SV endocytosis. However, unlike SV exocytosis, in which a single protein (synaptotagmin 1) accounts for much of the calcium dependence of SV fusion (Chapman 2008), it is not clear where calcium acts during endocytosis. Nearly every endocytic protein interacts with the membrane or directly binds to a lipid-binding protein. Because elevated calcium can alter the energetics of these membrane interactions, it is plausible that presynaptic calcium functions to change the efficiency of endocytosis through general effects on protein-lipid interactions rather than through a dedicated calcium sensor. However, it is worth noting that synaptotagmin 1 interacts with the core module and its C2 domains may affect this interaction, at least in regimes where calcium is highly elevated (Diril et al. 2006, Jung et al. 2007, Mohrmann et al. 2008, Walther et al. 2001). Multiple lines of evidence support a role for synaptotagmin in endocytosis. Another candidate mechanism for calcium in endocytosis is through the dephosphins, endocytic proteins such as dynamin, synaptojanin, and amphiphysin, that are dephosphorylated in a calcium-dependent manner by the phosphatase calcineurin (Cousin & Robinson 2001). The significance of these dephosphorylation events has not been fully delineated, although dephosphorylation of dynamin promotes its interaction with syndapin (Anggono et al. 2006). Dephosphins seem to play a greater role www.annualreviews.org • Synaptic Vesicle Endocytosis 151 ANRV389-CB25-07 ARI 4 September 2009 17:25 during conditions that lead to bulk-endocytosis (Clayton et al. 2007), which may happen only rarely under physiological stimuli. A ROLE FOR ACTIN DURING SYNAPTIC VESICLE ENDOCYTOSIS? Annu. Rev. Cell Dev. Biol. 2009.25:133-160. Downloaded from arjournals.annualreviews.org by WEILL MEDICAL COLLEGE OF CORNELL UNIVERSITY on 10/29/09. For personal use only. The regulation of actin polymerization is a vital part of membrane remodeling and intracellular trafficking, and multiple endocytic proteins mentioned above have a direct impact on actin and the cytoskeleton. For example, the secondary effectors dynamin, syndapin, and N-WASP, as well as intersectin, all connect directly or indirectly to proteins that control actin polymerization (Hussain et al. 2001, Kessels & Qualmann 2004, Koh et al. 2004, Miki & Takenawa 2003, Shin et al. 2007). In yeast, actin plays an essential role in endocytosis (Kaksonen et al. 2006). In vitro membrane scission by dynamin can utilize actin as an anchor to transduce its force on tubules into longitudinal tension (Roux et al. 2006). However, actin does not appear to be important for SV endocytosis per se in hippocampal neurons (Sankaranarayanan et al. 2003), but it may contribute as a secondary scaffold (Figure 7). Interestingly, actin seems to have a more pronounced role in the endocytosis of SVs at a giant synapse (Brodin & Shupliakov 2006, Shupliakov et al. 2002). Perhaps the cytoskeletal connections to endocytosis depend on the geometry and size of a synaptic terminal, particularly because regulation of the cytoskeleton is likely to be a major determinant of synapse size. REBUILDING SYNAPTIC VESICLES: A SINGLE-PASS SORTING AT THE PLASMA MEMBRANE? One of the assumptions we have made in discussing vesicle recycling is that the sorting of SV proteins occurs in a single endocytic step, as opposed to having an intermediate organelle after endocytosis at the plasma membrane makes passage through a recycling 152 Dittman · Ryan endosome for further cargo sorting (Koenig & Ikeda 1996). The existence of such a recycling endosome remains a formal possibility, although very few experiments have provided definitive evidence for any functional significance. In the earliest studies of Heuser & Reese (1973), they identified a nonvesicular horse radish peroxidase–positive compartment that appeared only transiently following stimulation. More recent experiments in which serial-sectioning electron microscopy was performed demonstrated that, during intense stimulation, deep invaginations of the plasma membrane often form, which can easily be confused with internal organelles in any single section (e.g., see Figure 5), but, in fact, remain topologically connected to the cell surface (Ferguson et al. 2007). Nonetheless, an endosomal intermediate may play a role under some stimulus conditions, particularly following intensive stimulation, when bulk endocytosis has been reported to operate at a number of synapses (Clayton et al. 2008, Holt et al. 2003). A MOLECULAR PICTURE FOR THE ENDOCYTIC MACHINE Ideally, the molecules described here can be mapped to the phenomenology of SV endocytosis on the basis of their intermolecular interactions, genetics, structure, and cell biological roles in other forms of endocytosis. A host of studies published over the past decade have made apparent a rough version of this molecular map and the dynamics of the endocytic machinery. The core module couples nucleation of an endocytic coat with cargo recruitment. Polymerization of clathrin and membrane interactions with ENTH and N-BAR domains rapidly commence the process of curving the lipid bilayer at the site of endocytosis. Furthermore, assembly of the core may increase the avidity of cargo binding because a multiplicity of binding sites will be brought into a small region of the membrane. In this scenario, cargo capture, scaffold assembly, and membrane curvature are highly coupled coordinated events. Annu. Rev. Cell Dev. Biol. 2009.25:133-160. Downloaded from arjournals.annualreviews.org by WEILL MEDICAL COLLEGE OF CORNELL UNIVERSITY on 10/29/09. For personal use only. ANRV389-CB25-07 ARI 4 September 2009 17:25 The N-BARs endophilin and amphiphysin contribute to membrane bending and may initiate an additional positive-feedback cycle of binding to curved membrane and selfassembly, driving further membrane curvature. Amphiphysin has direct binding interactions with clathrin and AP2 (placing it in the core module), whereas endophilin has no known interaction with these or other proteins in the core module (qualifying endophilin as a secondary effector). The secondary scaffold molecules Eps15 and intersectin/Dap160 bind to the core module and provide additional binding sites for recruiting the secondary effector molecules: dynamin, synaptojanin, and N-WASP. Both dynamin and synaptojanin convey amplification of a sort within the endocytic complex. Dynamin self-assembles and can utilize some of the energy released by polymerizing to deform membranes. Synaptojanin contains a phospholipase enzymatic domain, and thus a small number of synaptojanins would be expected to deplete PIP2 rapidly from the membrane. These effectors terminate the endocytic process by irreversibly severing the membrane connection and releasing the endocytic coat with the help of Hsc70 and auxilin. The AP180 paralog CALM, the stonin 2 paralog stonin 1, and the F-BAR syndapin may also contribute to SV endocytosis, but their specific requirements are not well defined. We therefore include these components as potential secondary effectors (Figures 6 and 7). NETWORK PROPERTIES OF THE ENDOCYTIC MACHINE There are several advantages to considering the endocytic proteins as a network (Alon 2007, Hintze & Adami 2008, Schmid & McMahon 2007). First, fault-tolerant networks exhibit graceful degradation, because the failure of one component has little effect on the overall performance. Evidence to date suggests that SV endocytosis continues after the removal of any one protein. Second, weak interactions can be transiently strengthened and amplified by positive feedback within the network. Thus, the binding of cargo; polymerization of the core module; and membrane bending by N-BARs, epsins, and dynamin all collaborate to create a fast, high-fidelity endocytic process. Third, modularity within the network allows the basic endocytic machinery to be utilized in many separate types of trafficking processes within a neuron. For instance, most of the SV endocytosis machinery is likely to be involved in postsynaptic receptor trafficking, intracellular trafficking between organelles, and membrane remodeling during synapse development (Bushlin et al. 2008, Gong & De Camilli 2008, Itoh & De Camilli 2006, Koh et al. 2007, Montesinos et al. 2005). Another consequence of a densely connected network is that the removal of one protein can have repercussions on the stability of many others. For instance, in fly Eps15 mutant synapses, many of the core proteins have significantly decreased abundance along with the Eps15-binding partner intersectin/Dap160 (Koh et al. 2004, 2007). This interdependence of protein stability emphasizes the importance of surveying the abundance of all endocytic proteins following a molecular deletion. Parsing the endocytic proteins into three layers (Figure 7) is not absolute in that members of the core module and the N-BARs also assist in recruiting secondary effectors (Schuske et al. 2003, Takei et al. 1999, Verstreken et al. 2003), whereas Eps15 and intersectin also help stabilize the core module (Koh et al. 2007, Morgan et al. 2003). The relative impact of removing particular proteins does not necessarily correspond to the location within this endocytic network. For instance, the core module proteins AP2, clathrin, and amphiphysin all have a relatively small effect on SV endocytosis when removed individually (Di Paolo et al. 2002, Gu et al. 2008, Sato et al. 2009). These observations would be predicted in a redundant network such as this, because secondary scaffolds and effectors functionally compensate for the loss of these core proteins. Perhaps double deletions that specifically target redundant function may have a more dramatic effect on SV endocytosis. For example, removal of clathrin and intersectin simultaneously would be predicted www.annualreviews.org • Synaptic Vesicle Endocytosis 153 ANRV389-CB25-07 ARI 4 September 2009 17:25 to impact the scaffolding function of the network. Concomitant removal of ENTH- and BAR-domain proteins may eliminate the ability to deform the membrane. CONCLUDING REMARKS Annu. Rev. Cell Dev. Biol. 2009.25:133-160. Downloaded from arjournals.annualreviews.org by WEILL MEDICAL COLLEGE OF CORNELL UNIVERSITY on 10/29/09. For personal use only. In the half century following Katz’s quantum hypothesis at the nerve terminal (Del Castillo & Katz 1954), advances in molecular, imaging, and physiological techniques have conspired to produce a fairly detailed understanding of the SV cycle. The process of manufacturing, trafficking, and recycling SVs involves the coordinated efforts of hundreds of proteins discovered over the past few decades. As the list of molecular players grows, so does the need to synthesize and organize this collection. As with many aspects of modern cell biology, the detailed properties of SV endocytosis may be emergent properties of a network. One of the impediments to understanding this emergence is that we still lack in-depth information about the precise location of most of these molecules during various stages of the SV cycle. Quantitative studies of the dynamic interactions, as well as application of new super-resolution techniques, should provide a rich new level of detail to understand the functioning of the endocytic network. SUMMARY POINTS 1. Clathrin-mediated endocytosis accounts for the majority of SV endocytosis under normal conditions, whereas bulk endocytosis may contribute following high-frequency stimulation. 2. SV proteins are endocytosed stochastically such that their dwell time on the plasma membrane obeys an exponential distribution with a 6-s time constant at 37◦ C (13-s time constant at room temperature). 3. The proteins of endocytosis can be organized in a simple hierarchical network based on protein interactions, domains, and genetic ablation studies. 4. No single component of the SV endocytosis machinery appears to be essential: The network of endocytic proteins appears robust to these molecular perturbations. DISCLOSURE STATEMENT The authors are not aware of any biases that might be perceived as affecting the objectivity of this review. ACKNOWLEDGMENTS The authors apologize in advance to all the investigators whose research could not be appropriately cited owing to space limitations. We thank members of the Ryan and Dittman labs for useful discussions and suggestions. LITERATURE CITED Abraham C, Hutter H, Palfreyman MT, Spatkowski G, Weimer RM, et al. 2006. Synaptic tetraspan vesicle membrane proteins are conserved but not needed for synaptogenesis and neuronal function in Caenorhabditis elegans. Proc. Natl. Acad. Sci. USA 103:8227–32 Albillos A, Dernick G, Horstmann H, Almers W, Alvarez de Toledo G, Lindau M. 1997. The exocytotic event in chromaffin cells revealed by patch amperometry. Nature 389:509–12 154 Dittman · Ryan Annu. Rev. Cell Dev. Biol. 2009.25:133-160. Downloaded from arjournals.annualreviews.org by WEILL MEDICAL COLLEGE OF CORNELL UNIVERSITY on 10/29/09. For personal use only. ANRV389-CB25-07 ARI 4 September 2009 17:25 Alon U. 2007. An Introduction to Systems Biology: Design Principles of Biological Circuits. Chapman & Hall/CRC. 320 pp. Andersson F, Jakobsson J, Low P, Shupliakov O, Brodin L. 2008. Perturbation of syndapin/PACSIN impairs synaptic vesicle recycling evoked by intense stimulation. J. Neurosci. 28:3925–33 Anggono V, Robinson PJ. 2007. Syndapin I and endophilin I bind overlapping proline-rich regions of dynamin I: role in synaptic vesicle endocytosis. J. Neurochem. 102:931–43 Anggono V, Smillie KJ, Graham ME, Valova VA, Cousin MA, Robinson PJ. 2006. Syndapin I is the phosphorylation-regulated dynamin I partner in synaptic vesicle endocytosis. Nat. Neurosci. 9:752–60 Aravanis AM, Pyle JL, Tsien RW. 2003. Single synaptic vesicles fusing transiently and successively without loss of identity. Nature 423:643–47 Balaji J, Armbruster M, Ryan TA. 2008. Calcium control of endocytic capacity at a CNS synapse. J. Neurosci. 28:6742–49 Balaji J, Ryan TA. 2007. Single-vesicle imaging reveals that synaptic vesicle exocytosis and endocytosis are coupled by a single stochastic mode. Proc. Natl. Acad. Sci. USA 104:20576–81 Bao H, Reist NE, Zhang B. 2008. The Drosophila epsin 1 is required for ubiquitin-dependent synaptic growth and function, but not for synaptic vesicle recycling. Traffic 9:2190–205 Bashkirov PV, Akimov SA, Evseev AI, Schmid SL, Zimmerberg J, Frolov VA. 2008. GTPase cycle of dynamin is coupled to membrane squeeze and release, leading to spontaneous fission. Cell 135:1276–86 Betz WJ, Bewick GS. 1992. Optical analysis of synaptic vesicle recycling at the frog neuromuscular junction. Science 255:200–3 Beutner D, Voets T, Neher E, Moser T. 2001. Calcium dependence of exocytosis and endocytosis at the cochlear inner hair cell afferent synapse. Neuron 29:681–90 Braun A, Pinyol R, Dahlhaus R, Koch D, Fonarev P, et al. 2005. EHD proteins associate with syndapin I and II and such interactions play a crucial role in endosomal recycling. Mol. Biol. Cell 16:3642–58 Brett TJ, Traub LM. 2006. Molecular structures of coat and coat-associated proteins: function follows form. Curr. Opin. Cell Biol. 18:395–406 Brodin L, Shupliakov O. 2006. Giant reticulospinal synapse in lamprey: molecular links between active and periactive zones. Cell Tissue Res. 326:301–10 Bushlin I, Petralia RS, Wu F, Harel A, Mughal MR, et al. 2008. Clathrin assembly protein AP180 and CALM differentially control axogenesis and dendrite outgrowth in embryonic hippocampal neurons. J. Neurosci. 28:10257–71 Ceccarelli B, Hurlbut WP. 1980. Ca2+ dependent recycling of synaptic vesicles at the frog neuromuscular junction. J. Cell Biol. 87:297–303 Ceccarelli B, Hurlbut WP, Mauro A. 1973. Turnover of transmitter and synaptic vesicles at the frog neuromuscular junction. J. Cell Biol. 57:499–524 Chapman ER. 2008. How does synaptotagmin trigger neurotransmitter release? Annu. Rev. Biochem. 77:615–41 Chen X, Barg S, Almers W. 2008. Release of the styryl dyes from single synaptic vesicles in hippocampal neurons. J. Neurosci. 28:1894–903 Clayton EL, Evans GJ, Cousin MA. 2007. Activity-dependent control of bulk endocytosis by protein dephosphorylation in central nerve terminals. J. Physiol. 585:687–91 Clayton EL, Evans GJ, Cousin MA. 2008. Bulk synaptic vesicle endocytosis is rapidly triggered during strong stimulation. J. Neurosci. 28:6627–32 Cousin MA, Robinson PJ. 2001. The dephosphins: dephosphorylation by calcineurin triggers synaptic vesicle endocytosis. Trends Neurosci. 24:659–65 Cremona O, Di Paolo G, Wenk MR, Luthi A, Kim WT, et al. 1999. Essential role of phosphoinositide metabolism in synaptic vesicle recycling. Cell 99:179–88 De Camilli P, Slepnev VI, Shupliakov O, Brodin L. 2001. Synaptic vesicle endocytosis. In Synapses, ed. WM Cowan, TC Südhof, CF Stevens, pp. 217–74. Baltimore, MD: The Johns Hopkins Univ. Press Del Castillo J, Katz B. 1954. Quantal components of the end-plate potential. J. Physiol. 124:560–73 Di Paolo G, Moskowitz HS, Gipson K, Wenk MR, Voronov S, et al. 2004. Impaired PtdIns(4,5)P2 synthesis in nerve terminals produces defects in synaptic vesicle trafficking. Nature 431:415–22 Di Paolo G, Sankaranarayanan S, Wenk MR, Daniell L, Perucco E, et al. 2002. Decreased synaptic vesicle recycling efficiency and cognitive deficits in amphiphysin 1 knockout mice. Neuron 33:789–804 www.annualreviews.org • Synaptic Vesicle Endocytosis Measured kinetics of single-vesicle retrieval and described the single-mode stochastic nature of SV cargo dwell time on the surface. First demonstration that the regulation of phosphoinositides is important for the SV cycle through examination of a mouse synaptojanin knockout. 155 Annu. Rev. Cell Dev. Biol. 2009.25:133-160. Downloaded from arjournals.annualreviews.org by WEILL MEDICAL COLLEGE OF CORNELL UNIVERSITY on 10/29/09. For personal use only. ANRV389-CB25-07 ARI 4 September 2009 Described a single mode of SV endocytosis using two cargo molecule reporters and impaired retrieval by siRNA knockdown of clathrin. 156 17:25 Diril MK, Wienisch M, Jung N, Klingauf J, Haucke V. 2006. Stonin 2 is an AP-2-dependent endocytic sorting adaptor for synaptotagmin internalization and recycling. Dev. Cell 10:233–44 Dittman JS, Kaplan JM. 2006. Factors regulating the abundance and localization of synaptobrevin in the plasma membrane. Proc. Natl. Acad. Sci. USA 103:11399–404 Edwards RH. 2007. The neurotransmitter cycle and quantal size. Neuron 55:835–58 Eisenberg E, Greene LE. 2007. Multiple roles of auxilin and hsc70 in clathrin-mediated endocytosis. Traffic 8:640–46 Evergren E, Gad H, Walther K, Sundborger A, Tomilin N, Shupliakov O. 2007. Intersectin is a negative regulator of dynamin recruitment to the synaptic endocytic zone in the central synapse. J. Neurosci. 27:379–90 Fergestad T, Broadie K. 2001. Interaction of stoned and synaptotagmin in synaptic vesicle endocytosis. J. Neurosci. 21:1218–27 Ferguson SM, Brasnjo G, Hayashi M, Wolfel M, Collesi C, et al. 2007. A selective activity-dependent requirement for dynamin 1 in synaptic vesicle endocytosis. Science 316:570–74 Fernandez-Alfonso T, Kwan R, Ryan TA. 2006. Synaptic vesicles interchange their membrane proteins with a large surface reservoir during recycling. Neuron 51:179–86 Fernandez-Alfonso T, Ryan TA. 2004. The kinetics of synaptic vesicle pool depletion at CNS synaptic terminals. Neuron 41:943–53 Ford MG, Mills IG, Peter BJ, Vallis Y, Praefcke GJ, et al. 2002. Curvature of clathrin-coated pits driven by epsin. Nature 419:361–66 Gad H, Löw P, Zotova E, Brodin L, Shupliakov O. 1998. Dissociation between Ca2+ -triggered synaptic vesicle exocytosis and clathrin-mediated endocytosis at a central synapse. Neuron 21:607–16 Gad H, Ringstad N, Löw P, Kjaerulff O, Gustafsson J, et al. 2000. Fission and uncoating of synaptic clathrincoated vesicles are perturbed by disruption of interactions with the SH3 domain of endophilin. Neuron 27:301–12 Gallop JL, McMahon HT. 2005. BAR domains and membrane curvature: bringing your curves to the BAR. Biochem. Soc. Symp. 72:223–31 Gong LW, De Camilli P. 2008. Regulation of postsynaptic AMPA responses by synaptojanin 1. Proc. Natl. Acad. Sci. USA 105:17561–66 Granseth B, Odermatt B, Royle SJ, Lagnado L. 2006. Clathrin-mediated endocytosis is the dominant mechanism of vesicle retrieval at hippocampal synapses. Neuron 51:773–86 Grass I, Thiel S, Honing S, Haucke V. 2004. Recognition of a basic AP-2 binding motif within the C2B domain of synaptotagmin is dependent on multimerization. J. Biol. Chem. 279:54872–80 Grigliatti TA, Hall L, Rosenbluth R, Suzuki DT. 1973. Temperature-sensitive mutations in Drosophila melanogaster. XIV. A selection of immobile adults. Mol. Gen. Genet. 120:107–14 Gu M, Schuske K, Watanabe S, Liu Q, Baum P, et al. 2008. mu2 adaptin facilitates but is not essential for synaptic vesicle recycling in Caenorhabditis elegans. J. Cell Biol. 183:881–92 Haffner C, Takei K, Chen H, Ringstad N, Hudson A, et al. 1997. Synaptojanin 1: localization on coated endocytic intermediates in nerve terminals and interaction of its 170 kDa isoform with Eps15. FEBS Lett. 419:175–80 Harata NC, Choi S, Pyle JL, Aravanis AM, Tsien RW. 2006. Frequency-dependent kinetics and prevalence of kiss-and-run and reuse at hippocampal synapses studied with novel quenching methods. Neuron 49:243–56 Harel A, Wu F, Mattson MP, Morris CM, Yao PJ. 2008. Evidence for CALM in directing VAMP2 trafficking. Traffic 9:417–29 Haucke V, De Camilli P. 1999. AP-2 recruitment to synaptotagmin stimulated by tyrosine-based endocytic motifs. Science 285:1268–71 Haucke V, Krauss M. 2002. Tyrosine-based endocytic motifs stimulate oligomerization of AP-2 adaptor complexes. Eur. J. Cell Biol. 81:647–53 Haucke V, Wenk MR, Chapman ER, Farsad K, De Camilli P. 2000. Dual interaction of synaptotagmin with mu2- and alpha-adaptin facilitates clathrin-coated pit nucleation. EMBO J. 19:6011–19 Haupts U, Maiti S, Schwille P, Webb WW. 1998. Dynamics of fluorescence fluctuations in green fluorescent protein observed by fluorescence correlation spectroscopy. Proc. Natl. Acad. Sci. USA 95:13573–78 Dittman · Ryan Annu. Rev. Cell Dev. Biol. 2009.25:133-160. Downloaded from arjournals.annualreviews.org by WEILL MEDICAL COLLEGE OF CORNELL UNIVERSITY on 10/29/09. For personal use only. ANRV389-CB25-07 ARI 4 September 2009 17:25 He L, Wu X-S, Mohan R, Wu L-G. 2006. Two modes of fusion pore opening revealed by cell-attached recordings at a synapse. Nature 444:102–5 Heerssen H, Fetter RD, Davis GW. 2008. Clathrin dependence of synaptic-vesicle formation at the Drosophila neuromuscular junction. Curr. Biol. 18:401–9 Heuser JE, Reese TS. 1973. Evidence for recycling of synaptic vesicle membrane during transmitter release at the frog neuromuscular junction. J. Cell Biol. 57:315–44 Hintze A, Adami C. 2008. Evolution of complex modular biological networks. PLoS Comput. Biol. 4:e23 Holt M, Cooke A, Wu MM, Lagnado L. 2003. Bulk membrane retrieval in the synaptic terminal of retinal bipolar cells. J. Neurosci. 23:1329–39 Horvath CA, Vanden Broeck D, Boulet GA, Bogers J, De Wolf MJ. 2007. Epsin: inducing membrane curvature. Int. J. Biochem. Cell Biol. 39:1765–70 Hurley JH. 2006. Membrane binding domains. Biochim. Biophys. Acta 1761:805–11 Hussain NK, Jenna S, Glogauer M, Quinn CC, Wasiak S, et al. 2001. Endocytic protein intersectin-l regulates actin assembly via Cdc42 and N-WASP. Nat. Cell Biol. 3:927–32 Israelachvili JN. 1992. Intermolecular and Surface Forces: With Applications to Colloidal and Biological Systems. London: Academic. 450 pp. Itoh T, De Camilli P. 2006. BAR, F-BAR (EFC) and ENTH/ANTH domains in the regulation of membranecytosol interfaces and membrane curvature. Biochim. Biophys. Acta 1761:897–912 Itoh T, Erdmann KS, Roux A, Habermann B, Werner H, De Camilli P. 2005. Dynamin and the actin cytoskeleton cooperatively regulate plasma membrane invagination by BAR and F-BAR proteins. Dev. Cell 9:791–804 Itoh T, Koshiba S, Kigawa T, Kikuchi A, Yokoyama S, Takenawa T. 2001. Role of the ENTH domain in phosphatidylinositol-4,5-bisphosphate binding and endocytosis. Science 291:1047–51 Jakobsson J, Gad H, Andersson F, Löw P, Shupliakov O, Brodin L. 2008. Role of epsin 1 in synaptic vesicle endocytosis. Proc. Natl. Acad. Sci. USA 105:6445–50 Jahn R, Schiebler W, Ouimet C, Greengard P. 1985. A 38,000-dalton membrane protein (p38) present in synaptic vesicles. Proc. Natl. Acad. Sci. USA 82:4137–41 Jockusch WJ, Praefcke GJ, McMahon HT, Lagnado L. 2005. Clathrin-dependent and clathrin-independent retrieval of synaptic vesicles in retinal bipolar cells. Neuron 46:869–78 Jorgensen EM, Hartwieg E, Schuske K, Nonet ML, Jin Y, Horvitz HR. 1995. Defective recycling of synaptic vesicles in synaptotagmin mutants of Caenorhabditis elegans. Nature 378:196–99 Jung N, Wienisch M, Gu M, Rand JB, Müller SL, et al. 2007. Molecular basis of synaptic vesicle cargo recognition by the endocytic sorting adaptor stonin 2. J. Cell Biol. 179:1497–510 Kaksonen M, Toret CP, Drubin DG. 2006. Harnessing actin dynamics for clathrin-mediated endocytosis. Nat. Rev. Mol. Cell Biol. 7:404–14 Kasprowicz J, Kuenen S, Miskiewicz K, Habets RL, Smitz L, Verstreken P. 2008. Inactivation of clathrin heavy chain inhibits synaptic recycling but allows bulk membrane uptake. J. Cell Biol. 182:1007–16 Keen JH. 1987. Clathrin assembly proteins: affinity purification and a model for coat assembly. J. Cell Biol. 105:1989–98 Kelly LE. 1983. The regulation of phosphorylation of a specific protein in synaptosomal fractions from Drosophila heads: the effects of light and two visual mutants. Cell Mol. Neurobiol. 3:127–41 Kelly LE, Phillips AM. 2005. Molecular and genetic characterization of the interactions between the Drosophila stoned-B protein and DAP-160 (intersectin). Biochem. J. 388:195–204 Kessels MM, Qualmann B. 2004. The syndapin protein family: linking membrane trafficking with the cytoskeleton. J. Cell Sci. 117:3077–86 Kim SH, Ryan TA. 2009. Synaptic vesicle recycling at CNS synapses without AP-2. J. Neurosci. 29:3865–74 Klingauf J, Kavalali ET, Tsien RW. 1998. Kinetics and regulation of fast endocytosis at hippocampal synapses. Nature 394:581–85 Koenig JH, Ikeda K. 1989. Disappearance and reformation of synaptic vesicle membrane upon transmitter release observed under reversible blockage of membrane retrieval. J. Neurosci. 9:3844–60 Koenig JH, Ikeda K. 1996. Synaptic vesicles have two distinct recycling pathways. J. Cell Biol. 135:797–808 Koenig JH, Ikeda K. 1999. Contribution of active zone subpopulation of vesicles to evoked and spontaneous release. J. Neurophysiol. 81:1495–505 www.annualreviews.org • Synaptic Vesicle Endocytosis This seminal work employed electron microscopy to visualize exo- and endocytosis at the frog NMJ. The culmination of multiple studies on a temperature-sensitive fly mutant (dynamin) that blocked endocytosis. 157 Annu. Rev. Cell Dev. Biol. 2009.25:133-160. Downloaded from arjournals.annualreviews.org by WEILL MEDICAL COLLEGE OF CORNELL UNIVERSITY on 10/29/09. For personal use only. ANRV389-CB25-07 ARI 4 September 2009 This EM study provided the first direct measurement of synaptic vesicle endocytosis kinetics. 158 17:25 Koh TW, Korolchuk VI, Wairkar YP, Jiao W, Evergren E, et al. 2007. Eps15 and Dap160 control synaptic vesicle membrane retrieval and synapse development. J. Cell Biol. 178:309–22 Koh TW, Verstreken P, Bellen HJ. 2004. Dap160/intersectin acts as a stabilizing scaffold required for synaptic development and vesicle endocytosis. Neuron 43:193–205 Kumar V, Alla SR, Krishnan KS, Ramaswami M. 2009. Syndapin is dispensable for synaptic vesicle endocytosis at the Drosophila larval neuromuscular junction. Mol. Cell Neurosci. 40:234–41 Larson DR, Zipfel WR, Williams RM, Clark SW, Bruchez MP, et al. 2003. Water-soluble quantum dots for multiphoton fluorescence imaging in vivo. Science 300:1434–36 Li Z, Murthy VN. 2001. Visualizing postendocytic traffic of synaptic vesicles at hippocampal synapses. Neuron 31:593–605 Liu J, Kaksonen M, Drubin DG, Oster G. 2006. Endocytic vesicle scission by lipid phase boundary forces. Proc. Natl. Acad. Sci. USA 103:10277–82 Lloyd TE, Verstreken P, Ostrin EJ, Phillippi A, Lichtarge O, Bellen HJ. 2000. A genome-wide search for synaptic vesicle cycle proteins in Drosophila. Neuron 26:45–50 Mani M, Lee SY, Lucast L, Cremona O, Di Paolo G, et al. 2007. The dual phosphatase activity of synaptojanin1 is required for both efficient synaptic vesicle endocytosis and reavailability at nerve terminals. Neuron 56:1004–18 Marie B, Sweeney ST, Poskanzer KE, Roos J, Kelly RB, Davis GW. 2004. Dap160/intersectin scaffolds the periactive zone to achieve high-fidelity endocytosis and normal synaptic growth. Neuron 43:207–19 McMahon HT, Bolshakov VY, Janz R, Hammer RE, Siegelbaum SA, Südhof TC. 1996. Synaptophysin, a major synaptic vesicle protein, is not essential for neurotransmitter release. Proc. Natl. Acad. Sci. USA 93:4760–64 Miki H, Takenawa T. 2003. Regulation of actin dynamics by WASP family proteins. J. Biochem. 134:309–13 Miller TM, Heuser JE. 1984. Endocytosis of synaptic vesicle membrane at the frog neuromuscular junction. J. Cell Biol. 98:685–98 Mohrmann R, Matthies HJ, Woodruff E 3rd, Broadie K. 2008. Stoned B mediates sorting of integral synaptic vesicle proteins. Neuroscience 153:1048–63 Montesinos ML, Castellano-Muñoz M, Garcı́a-Junco-Clemente P, Fernández-Chacón R. 2005. Recycling and EH domain proteins at the synapse. Brain Res. Brain Res. Rev. 49:416–28 Morgan JR, Prasad K, Hao W, Augustine GJ, Lafer EM. 2000. A conserved clathrin assembly motif essential for synaptic vesicle endocytosis. J. Neurosci. 20:8667–76 Morgan JR, Prasad K, Jin S, Augustine GJ, Lafer EM. 2003. Eps15 homology domain-NPF motif interactions regulate clathrin coat assembly during synaptic vesicle recycling. J. Biol. Chem. 278:33583–92 Neves G, Gomis A, Lagnado L. 2001. Calcium influx selects the fast mode of endocytosis in the synaptic terminal of retinal bipolar cells. Proc. Natl. Acad. Sci. USA 98:15282–87 Neves G, Lagnado L. 1999. The kinetics of exocytosis and endocytosis in the synaptic terminal of goldfish retinal bipolar cells. J. Physiol. 515(Pt. 1):181–202 Newton AJ, Kirchhausen T, Murthy VN. 2006. Inhibition of dynamin completely blocks compensatory synaptic vesicle endocytosis. Proc. Natl. Acad. Sci. USA 103:17955–60 Nicholson-Tomishima K, Ryan TA. 2004. Kinetic efficiency of endocytosis at mammalian CNS synapses requires synaptotagmin I. Proc. Natl. Acad. Sci. USA 101:16648–52 Nonet ML, Holgado AM, Brewer F, Serpe CJ, Norbeck BA, et al. 1999. UNC-11, a Caenorhabditis elegans AP180 homologue, regulates the size and protein composition of synaptic vesicles. Mol. Biol. Cell 10:2343– 60 Parsons TD, Lenzi D, Almers W, Roberts WM. 1994. Calcium-triggered exocytosis and endocytosis in an isolated presynaptic cell: capacitance measurements in saccular hair cells. Neuron 13:875–83 Perin MS, Fried VA, Mignery GA, Jahn R, Südhof TC. 1990. Phospholipid binding by a synaptic vesicle protein homologous to the regulatory region of protein kinase C. Nature 345:260–63 Phillips AM, Smith M, Ramaswami M, Kelly LE. 2000. The products of the Drosophila stoned locus interact with synaptic vesicles via synaptotagmin. J. Neurosci. 20:8254–61 Poskanzer KE, Fetter RD, Davis GW. 2006. Discrete residues in the c(2)b domain of synaptotagmin I independently specify endocytic rate and synaptic vesicle size. Neuron 50:49–62 Dittman · Ryan Annu. Rev. Cell Dev. Biol. 2009.25:133-160. Downloaded from arjournals.annualreviews.org by WEILL MEDICAL COLLEGE OF CORNELL UNIVERSITY on 10/29/09. For personal use only. ANRV389-CB25-07 ARI 4 September 2009 17:25 Poskanzer KE, Marek KW, Sweeney ST, Davis GW. 2003. Synaptotagmin I is necessary for compensatory synaptic vesicle endocytosis in vivo. Nature 426:559–63 Pyle JL, Kavalali ET, Piedras-Renterı́a ES, Tsien RW. 2000. Rapid reuse of readily releasable pool vesicles at hippocampal synapses. Neuron 28:221–31 Razzaq A, Robinson IM, McMahon HT, Skepper JN, Su Y, et al. 2001. Amphiphysin is necessary for organization of the excitation-contraction coupling machinery of muscles, but not for synaptic vesicle endocytosis in Drosophila. Genes Dev. 15:2967–79 Reist NE, Buchanan J, Li J, DiAntonio A, Buxton EM, Schwarz TL. 1998. Morphologically docked synaptic vesicles are reduced in synaptotagmin mutants of Drosophila. J. Neurosci. 18:7662–73 Ren G, Vajjhala P, Lee JS, Winsor B, Munn AL. 2006. The BAR domain proteins: molding membranes in fission, fusion, and phagy. Microbiol. Mol. Biol. Rev. 70:37–120 Richards DA, Bai J, Chapman ER. 2005. Two modes of exocytosis at hippocampal synapses revealed by rate of FM1-43 efflux from individual vesicles. J. Cell Biol. 168:929–39 Rikhy R, Kumar V, Mittal R, Krishnan KS. 2002. Endophilin is critically required for synapse formation and function in Drosophila melanogaster. J. Neurosci. 22:7478–84 Rizo J, Rosenmund C. 2008. Synaptic vesicle fusion. Nat. Struct. Mol. Biol. 15:665–74 Rose S, Malabarba MG, Krag C, Schultz A, Tsushima H, et al. 2007. Caenorhabditis elegans intersectin: a synaptic protein regulating neurotransmission. Mol. Biol. Cell 18:5091–99 Roux A, Uyhazi K, Frost A, De Camilli P. 2006. GTP-dependent twisting of dynamin implicates constriction and tension in membrane fission. Nature 441:528–31 Ryan TA, Reuter H, Wendland B, Schweizer FE, Tsien RW, Smith SJ. 1993. The kinetics of synaptic vesicle recycling measured at single presynaptic boutons. Neuron 11:713–24 Ryan TA, Smith SJ. 1995. Vesicle pool mobilization during action potential firing at hippocampal synapses. Neuron 14:983–89 Ryan TA, Smith SJ, Reuter H. 1996. The timing of synaptic vesicle endocytosis. Proc. Natl. Acad. Sci. USA 93:5567–71 Salcini AE, Hilliard MA, Croce A, Arbucci S, Luzzi P, et al. 2001. The Eps15 C. elegans homologue EHS-1 is implicated in synaptic vesicle recycling. Nat. Cell Biol. 3:755–60 Sankaranarayanan S, Atluri PP, Ryan TA. 2003. Actin has a molecular scaffolding, not propulsive, role in presynaptic function. Nat. Neurosci. 6:127–35 Sankaranarayanan S, De Angelis D, Rothman JE, Ryan TA. 2000. The use of pHluorins for optical measurements of presynaptic activity. Biophys. J. 79:2199–208 Sankaranarayanan S, Ryan TA. 2000. Real-time measurements of vesicle-SNARE recycling in synapses of the central nervous system. Nat. Cell Biol. 2:197–204 Sankaranarayanan S, Ryan TA. 2001. Calcium accelerates endocytosis of vSNAREs at hippocampal synapses. Nat. Neurosci. 4:129–36 Sato K, Ernstrom GG, Watanabe S, Weimer RM, Chen CH, et al. 2009. Differential requirements for clathrin in receptor-mediated endocytosis and maintenance of synaptic vesicle pools. Proc. Natl. Acad. Sci. USA 106:1139–44 Schmid EM, McMahon HT. 2007. Integrating molecular and network biology to decode endocytosis. Nature 448:883–88 Schuske KR, Richmond JE, Matthies DS, Davis WS, Runz S, et al. 2003. Endophilin is required for synaptic vesicle endocytosis by localizing synaptojanin. Neuron 40:749–62 Shin N, Lee S, Ahn N, Kim SA, Ahn SG, et al. 2007. Sorting nexin 9 interacts with dynamin 1 and N-WASP and coordinates synaptic vesicle endocytosis. J. Biol. Chem. 282:28939–50 Shupliakov O, Bloom O, Gustafsson JS, Kjaerulff O, Löw P, et al. 2002. Impaired recycling of synaptic vesicles after acute perturbation of the presynaptic actin cytoskeleton. Proc. Natl. Acad. Sci. USA 99:14476–81 Shupliakov O, Löw P, Grabs D, Gad H, Chen H, et al. 1997. Synaptic vesicle endocytosis impaired by disruption of dynamin-SH3 domain interactions. Science 276:259–63 Stimson DT, Estes PS, Rao S, Krishnan KS, Kelly LE, Ramaswami M. 2001. Drosophila stoned proteins regulate the rate and fidelity of synaptic vesicle internalization. J. Neurosci. 21:3034–44 Südhof TC. 2004. The synaptic vesicle cycle. Annu. Rev. Neurosci. 27:509–47 www.annualreviews.org • Synaptic Vesicle Endocytosis Utilized SH3 and PRD peptides at a giant synapse to disrupt amphiphysin and dynamin interactions, blocking clathrinmediated endocytosis of SVs. 159 ARI 4 September 2009 17:25 Sun JY, Wu XS, Wu LG. 2002. Single and multiple vesicle fusion induce different rates of endocytosis at a central synapse. Nature 417:555–59 Tabares L, Ruiz R, Linares-Clemente P, Gaffield MA, Alvarez de Toledo G, et al. 2007. Monitoring synaptic function at the neuromuscular junction of a mouse expressing synaptopHluorin. J. Neurosci. 27:5422–30 Takamori S, Holt M, Stenius K, Lemke EA, Gronborg M, et al. 2006. Molecular anatomy of a trafficking organelle. Cell 127:831–46 Takei K, Slepnev VI, Haucke V, De Camilli P. 1999. Functional partnership between amphiphysin and dynamin in clathrin-mediated endocytosis. Nat. Cell Biol. 1:33–39 Trimble WS, Cowan DM, Scheller RH. 1988. VAMP-1: a synaptic vesicle-associated integral membrane protein. Proc. Natl. Acad. Sci. USA. 85:4538–42 Verstreken P, Kjaerulff O, Lloyd TE, Atkinson R, Zhou Y, et al. 2002. Endophilin mutations block clathrinmediated endocytosis but not neurotransmitter release. Cell 109:101–12 Verstreken P, Koh TW, Schulze KL, Zhai RG, Hiesinger PR, et al. 2003. Synaptojanin is recruited by endophilin to promote synaptic vesicle uncoating. Neuron 40:733–48 Voglmaier SM, Kam K, Yang H, Fortin DL, Hua Z, et al. 2006. Distinct endocytic pathways control the rate and extent of synaptic vesicle protein recycling. Neuron 51:71–84 von Gersdorff H, Matthews G. 1994a. Dynamics of synaptic vesicle fusion and membrane retrieval in synaptic terminals. Nature 367:735–39 von Gersdorff H, Matthews G. 1994b. Inhibition of endocytosis by elevated internal calcium in a synaptic terminal. Nature 370:652–55 Walther K, Diril MK, Jung N, Haucke V. 2004. Functional dissection of the interactions of stonin 2 with the adaptor complex AP-2 and synaptotagmin. Proc. Natl. Acad. Sci. USA 101:964–69 Walther K, Krauss M, Diril MK, Lemke S, Ricotta D, et al. 2001. Human stoned B interacts with AP-2 and synaptotagmin and facilitates clathrin-coated vesicle uncoating. EMBO Rep. 2:634–40 Wang W, Bouhours M, Gracheva EO, Liao EH, Xu K, et al. 2008. ITSN-1 controls vesicle recycling at the neuromuscular junction and functions in parallel with DAB-1. Traffic 9:742–54 Wendland B, Steece KE, Emr SD. 1999. Yeast epsins contain an essential N-terminal ENTH domain, bind clathrin and are required for endocytosis. EMBO J. 18:4383–93 Wienisch M, Klingauf J. 2006. Vesicular proteins exocytosed and subsequently retrieved by compensatory endocytosis are nonidentical. Nat. Neurosci. 9:1019–27 Wu LG, Betz WJ. 1996. Nerve activity but not intracellular calcium determines the time course of endocytosis at the frog neuromuscular junction. Neuron 17:769–79 Wu W, Xu J, Wu XS, Wu LG. 2005. Activity-dependent acceleration of endocytosis at a central synapse. J. Neurosci. 25:11676–83 Yamashita T, Hige T, Takahashi T. 2005. Vesicle endocytosis requires dynamin-dependent GTP hydrolysis at a fast CNS synapse. Science 307:124–27 Zelhof AC, Bao H, Hardy RW, Razzaq A, Zhang B, Doe CQ. 2001. Drosophila amphiphysin is implicated in protein localization and membrane morphogenesis but not in synaptic vesicle endocytosis. Development 128:5005–15 Zenisek D, Steyer JA, Feldman ME, Almers W. 2002. A membrane marker leaves synaptic vesicles in milliseconds after exocytosis in retinal bipolar cells. Neuron 35:1085–97 Zhang B, Koh YH, Beckstead RB, Budnik V, Ganetzky B, Bellen HJ. 1998. Synaptic vesicle size and number are regulated by a clathrin adaptor protein required for endocytosis. Neuron 21:1465–75 Zhang JZ, Davletov BA, Südhof TC, Anderson RG. 1994. Synaptotagmin I is a high affinity receptor for clathrin AP-2: implications for membrane recycling. Cell 78:751–60 Zhang Q, Cao YQ, Tsien RW. 2007. Quantum dots provide an optical signal specific to full collapse fusion of synaptic vesicles. Proc. Natl. Acad. Sci. USA 104:17843–48 Annu. Rev. Cell Dev. Biol. 2009.25:133-160. Downloaded from arjournals.annualreviews.org by WEILL MEDICAL COLLEGE OF CORNELL UNIVERSITY on 10/29/09. For personal use only. ANRV389-CB25-07 160 Dittman · Ryan AR389-FM ARI 14 September 2009 14:58 Contents Annu. Rev. Cell Dev. Biol. 2009.25:133-160. Downloaded from arjournals.annualreviews.org by WEILL MEDICAL COLLEGE OF CORNELL UNIVERSITY on 10/29/09. For personal use only. Chromosome Odds and Ends Joseph G. Gall p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p 1 Small RNAs and Their Roles in Plant Development Xuemei Chen p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p21 Annual Review of Cell and Developmental Biology Volume 25, 2009 From Progenitors to Differentiated Cells in the Vertebrate Retina Michalis Agathocleous and William A. Harris p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p45 Mechanisms of Lipid Transport Involved in Organelle Biogenesis in Plant Cells Christoph Benning p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p71 Innovations in Teaching Undergraduate Biology and Why We Need Them William B. Wood p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p93 Membrane Traffic within the Golgi Apparatus Benjamin S. Glick and Akihiko Nakano p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p 113 Molecular Circuitry of Endocytosis at Nerve Terminals Jeremy Dittman and Timothy A. Ryan p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p 133 Many Paths to Synaptic Specificity Joshua R. Sanes and Masahito Yamagata p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p 161 Mechanisms of Growth and Homeostasis in the Drosophila Wing Ricardo M. Neto-Silva, Brent S. Wells, and Laura A. Johnston p p p p p p p p p p p p p p p p p p p p p p p p p 197 Vertebrate Endoderm Development and Organ Formation Aaron M. Zorn and James M. Wells p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p 221 Signaling in Adult Neurogenesis Hoonkyo Suh, Wei Deng, and Fred H. Gage p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p 253 Vernalization: Winter and the Timing of Flowering in Plants Dong-Hwan Kim, Mark R. Doyle, Sibum Sung, and Richard M. Amasino p p p p p p p p p p p p 277 Quantitative Time-Lapse Fluorescence Microscopy in Single Cells Dale Muzzey and Alexander van Oudenaarden p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p 301 Mechanisms Shaping the Membranes of Cellular Organelles Yoko Shibata, Junjie Hu, Michael M. Kozlov, and Tom A. Rapoport p p p p p p p p p p p p p p p p p p p p 329 The Biogenesis and Function of PIWI Proteins and piRNAs: Progress and Prospect Travis Thomson and Haifan Lin p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p 355 vii AR389-FM ARI 14 September 2009 14:58 Mechanisms of Stem Cell Self-Renewal Shenghui He, Daisuke Nakada, and Sean J. Morrison p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p 377 Collective Cell Migration Pernille Rørth p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p 407 Hox Genes and Segmentation of the Hindbrain and Axial Skeleton Tara Alexander, Christof Nolte, and Robb Krumlauf p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p 431 Annu. Rev. Cell Dev. Biol. 2009.25:133-160. Downloaded from arjournals.annualreviews.org by WEILL MEDICAL COLLEGE OF CORNELL UNIVERSITY on 10/29/09. For personal use only. Gonad Morphogenesis in Vertebrates: Divergent Means to a Convergent End Tony DeFalco and Blanche Capel p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p 457 From Mouse Egg to Mouse Embryo: Polarities, Axes, and Tissues Martin H. Johnson p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p 483 Conflicting Views on the Membrane Fusion Machinery and the Fusion Pore Jakob B. Sørensen p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p 513 Coordination of Lipid Metabolism in Membrane Biogenesis Axel Nohturfft and Shao Chong Zhang p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p 539 Navigating ECM Barriers at the Invasive Front: The Cancer Cell–Stroma Interface R. Grant Rowe and Stephen J. Weiss p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p 567 The Molecular Basis of Organ Formation: Insights from the C. elegans Foregut Susan E. Mango p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p 597 Genetic Control of Bone Formation Gerard Karsenty, Henry M. Kronenberg, and Carmine Settembre p p p p p p p p p p p p p p p p p p p p p p 629 Listeria monocytogenes Membrane Trafficking and Lifestyle: The Exception or the Rule? Javier Pizarro-Cerdá and Pascale Cossart p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p 649 Asymmetric Cell Divisions and Asymmetric Cell Fates Shahragim Tajbakhsh, Pierre Rocheteau, and Isabelle Le Roux p p p p p p p p p p p p p p p p p p p p p p p p p p p 671 Indexes Cumulative Index of Contributing Authors, Volumes 21–25 p p p p p p p p p p p p p p p p p p p p p p p p p p p 701 Cumulative Index of Chapter Titles, Volumes 21–25 p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p 704 Errata An online log of corrections to Annual Review of Cell and Developmental Biology articles may be found at http://cellbio.annualreviews.org/errata.shtml viii Contents