T cell development and self tolerance PPT
advertisement

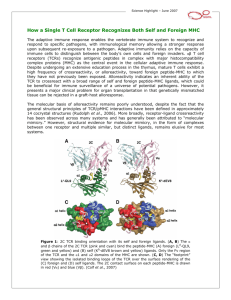
Topic 7 T Cell Development, Repertoire Selection and Immune Self Tolerance ©Dr. Colin R.A. Hewitt crah1@le.ac.uk Why is a mechanism for repertoire selection and self tolerance needed? T T T T T T T T T T T T T T T T T T T T T T T T T Generation of the TcR repertoire involves many random mechanisms The specificity of TcR in the immature repertoire is also random & will include cells with receptors that are: Self antigen recognition T T T T APC 1. Harmful 2. Useless 3. Useful Foreign antigen recognition Self proteins enter the endogenous and exogenous antigen processing pathways Self cellular proteins Self serum & cellular proteins Processing pathways do not distinguish self from non-self Self peptides load onto MHC class I & II molecules Purify stable MHCpeptide complexes Fractionate and microsequence peptides Acid elute peptides >90% of eluted peptides are derived from self proteins Yet self antigens do not usually activate T cells The immune system allows a limited degree of self recognition TcRs recognise the non-self peptide antigen and the self MHC molecule MHC molecules RESTRICT T cell activation But how do T cells learn how much self recognition is acceptable? T cells are only allowed to develop if their TcR recognise parts of self MHC Explains why T cells of MHC haplotype A do not recognise antigen specific presented by MHC haplotype B MHC A haplotype T CELL MHC A haplotype APC MHC B haplotype APC Wholly self-reactive and useless T cells are removed MHC-restricted are retained T T T T T T T Y Y Y Y Y Y Y Y Y Y Y YYYYY Y YY T T T T T Y T T Y ? Harmful Negatively select T THYMUS T T Y T T T Y Random TcR repertoire ensures diversity APC Useless Useful Neglect Positively select The thymus Lobulated structure with a STROMA of epithelial cells & connective tissue Stroma provides a microenvironment for T cell development & selection Lobules differentiated into an outer CORTEX & inner MEDULLA, both filled with bone-marrow-derived THYMOCYTES Thymocyte Cortex Cortical epithelial cell Dendritic cell Macrophage Medulla Medullary epithelial cell The thymus is required for T cell maturation Athymic mice (nude) and humans (DiGeorge syndrome) are immunodeficient due to a lack of T cells Neonatal thymectomy Thymus intact No mature T cells In adult Mature T cells In adult Roles of the bone marrow and thymus in T cell maturation Defective lymphocyte production Normal thymus scid/scid No mature T cells In adults Thymus defect Normal bone marrow nu/nu No mature T cells In adults Bone marrow supplies T cells, and they mature in the thymus Marrow defect Thymus defect Bone marrow transplant Thymus colonised by thymocytes from thymus defective, i.e. orange, mouse Thymus graft Thymus colonised by thymocytes from the thymus defective, i.e. orange, mouse The thymus matures T cells after birth, but early in life Remove Thymus Adult Neonate T cells not yet left thymus Mature T & B cells No T cells Mature B cells present The thymus is needed to generate mature T cells The thymus is most active in the foetal and neonatal period OVA Adult KLH Neonate T cells vs. OVA No T cells vs.OVA T cells vs. KLH T cells vs. KLH The thymus is needed for NEONATAL TOLERANCE T cells mature in the thymus but most die there. Mouse thymus Constant 1-2 x 108 cells 5 x 107 per day 2 x 106 per day 98% of cells die in the thymus without inducing any inflammation or any change in the size of the thymus. Thymic macrophages phagocytose apoptotic thymocytes. T cell development is marked by cell surface molecule changes As T cells mature in the thymus they change their expression of TcRassociated molecules and co-receptors. These changes can be used as markers of their stage of maturation Double negative Large double positive Small double positive Single positive CD3/TcRCD4-, 8- CD3+ TcR-chain + pre-TcR+ (pT CD4+, 8+ CD3+ TcR + CD4+ CD8+ CD3+ TcR + CD4+ TcR+ CD3+ CD4-, 8- Single positive 98% CD3+ TcR + CD8+ Different developmental stages of thymocytes are present in different parts of the thymus Cortex DN DN DN CD25CD44+ CD25+ CD44+ CD25+, CD44low DP DP DN CD3+ pT: CD3+ TcR+ CD3+ pT: CD25-, CD44- SP CD3+ TcR+ CD4+ SP CD3+ TcR+ CD8+ Immature double negative & positive thymocytes Medulla Mature single positive thymocytes TcR rearrangement CD25CD44+ DN V CD25+ CD44+ J C Germline configuration DN V CD25+, CD44low D DJ C V DJ C DN D-J fusion V-DJ fusion C region spliced to VDJ fusion and -chain protein produced in cytoplasm No TcR at cell surface Similarities in the development of T and B cells: A B cell reminder Large Pre-B Surrogate light chain is transiently expressed when VHDHJH CHm is productively rearranged 1. Triggers entry into cell cycle Expands pre-B cells with in frame VDJ joins 2. Suppresses further H chain rearrangement Allelic exclusion Similarities in the development of T and B cells: Pre T cell receptor preTcR -chain DN TcR -chain CD3+ very low pT: CD25- CD44- CD4 CD8 preTcR -chain DP TcR -chain CD3+ low pT: CD25- CD44CD4+ CD8+ 1. Cell proliferates rapidly to yield daughter cells with the same chain Expands only cells with in-frame TcR chains 2. Successful rearrangement shuts off rearrangement on 2nd chromosome Ensures only one specificity of TcR expressed per cell TcR rearrangement DP CD3+ low pT: CD25- CD44CD4+ CD8+ V When proliferation stops, the chain starts to rearrange J C Germline TcR V-J rearranged TcR 1° transcript Spliced TcR mRNA DP CD3+ TcR+ T cells can now recognise antigens and interact with MHC class I & II through CD4 & CD8 Selection can now begin Mouse thymus 5 x 107 per day 2 x 106 per day How does the thymus choose which of the cells entering the thymus are useful, harmful and useless Sorting the useful from the harmful and the useless Positive selection Retention of thymocytes expressing TcR that are RESTRICTED in their recognition of antigen by self MHC i.e. selection of the USEFUL Negative selection Removal of thymocytes expressing TcR that either recognise self antigens presented by self MHC or that have no affinity for self MHC i.e. selection of the HARMFUL and the USELESS MHC restriction Antigen can be seen by the TcR only in the context of an MHC molecule TcR will not bind to an MHC molecule unless there is an antigen in the groove In the presence of antigen, the TcR must have some affinity for the MHC molecule Experimental evidence for MHC restriction as a marker of positive selection CHIMERA Thymus defect Orange strain cells in a blue strain mouse Which MHC haplotype will restrict the T cells, Orange or blue? Bone marrow transplant Marrow defect Transplant reconstitutes marrow defective mouse Studies in bone marrow chimeras show that MHC restriction is learnt in the thymus Bone marrow donor MHC A MHC (AxB)F1 MHC B A Irradated bone marrow recipients A MHC (AxB)F1 B B B A T cell response of recipient T cells to antigen MHC haplotype of APC The MHC haplotype of the environment in which T cells mature determines their MHC restriction element Explanation of bone marrow chimera experiment: Mice of a particular MHC haplotype only make T cells restricted by that haplotype MHC A MHC B MHC (AxB)F1 Able to make T cells restricted by MHC A Able to make T cells restricted by MHC B Able to make T cells restricted by MHC A or B Bone marrow must contain potential to make T cells restricted by A and B MHC molecules Explanation of bone marrow chimera experiment: Irradiation prevents the bone marrow from generating lymphocytes MHC A MHC B Normal mice MHC A MHC B Irradiation destroys the immune system but has no effect on the epithelial or dendritic cells of the thymus MHC A MHC B Mice now have an intact, functional thymic stroma but have no thymocytes, T cells or bone marrow These mice are severely immunodeficient and can only be reconstituted by a bone marrow transplant Explanation of bone marrow chimera experiment: Reconstitution of irradiated mice with (AxB)F1 bone marrow MHC (AxB)F1 Bone marrow contains the potential to make T cells restricted by A and B MHC molecules Transplant bone marrow to reconstitute immune system of immunodeficient mice Irradiated bone marrow recipients MHC A MHC (AxB)F1 MHC B Explanation of bone marrow chimera experiment: MHC restriction is learnt in the thymus by positive selection A x B T cell precursors Mouse with an MHC A thymus, but A x B bone marrow MHC A Thymus A x B T cell precursors Mouse with an MHC B thymus, but A x B bone marrow MHC B Thymus Mature T cells restricted only by MHC A Mature T cells restricted only by MHC B Explanation of bone marrow chimera experiment: Peripheral T cells are restricted by the MHC type of the thymus that they mature in Bone marrow donor MHC (AxB)F1 Bone marrow recipients MHC A MHC B B A B A T cell response of recipient T cells to antigen MHC haplotype of antigen presenting cells Summary Bone marrow chimeras show that MHC restriction is learnt in the thymus T cells are ‘educated’ in the thymus to recognise antigens only in the context of self MHC MHC restriction is learnt in the thymus by positive selection The MHC haplotype of the environment in which T cells mature determines their MHC restriction element Negative Selection Removal of thymocytes expressing TcR that either recognise self antigens presented by self MHC or that have no affinity for self MHC i.e. selection of the HARMFUL and the USELESS Superantigens can be used to probe the mechanisms of negative selection Nominal antigens & superantigens Nominal antigens Superantigens Require processing to peptides Not processed TcR and chains are involved in recognition Only TcR chain involved in recognition >1 in 105 T cells recognise each peptide 2-20% of T cells recognise each superantigen Recognition restricted by an MHC class I or II molecule Presented by almost any MHC class II molecule Almost all proteins can be nominal antigens Very few antigens are superantigens Suggests a strikingly different mechanism of antigen presentation & recognition. Superantigens T cell e.g. Staphylococcal enterotoxins Toxic shock syndrome toxin I (TSST-1) Staphylococcal enterotoxins SEA, SEB, SEC, SED & SEE Do not induce adaptive responses, but trigger a massive burst of cytokines that may cause fever, systemic toxicity & immune suppression Severe food poisoning Toxic shock syndrome V V TcR from MHC A haplotype Class II from MHC A to Z haplotypes APC Interaction of SEB with MHC Class II molecules and the TcR MHC class II TcR beta chain MHC class II SEB TcR beta chain SEB Exogenous superantigen-V relationship Superantigen Human V region SEA 1.1, 5.3, 6.3, 6.4 6.9, 7.3, 7.4, 9.1 3, 12, 14, 15, 17, 20 12 12, 13.1, 13.2 5, 12 5.1, 6.3, 6.4, 6.9, 8.1 2 SEB SEC1 SEC2 SED SEE TSST-1 Explains why superantigens stimulate so many T cells Effect of TSST-1 on T cells expressing V2 Cell number Fresh PBMC unstained PBMC cultured with TSST-1 Stained with anti-V3 Cell number PBMC cultured with TSST-1 Stained with anti-V2 Fresh PBMC stained with anti-V2 Fluorescence intensity (i.e. amount of staining with anti-V antibody) Other exogenous superantigens Bacterial exoproteins Staphylococcal exfoliative toxins Streptococcus pyogenes erythrogenic toxins A & C (?Streptococcal M protein?) Yersinia enterocolitica superantigen Clostridium perfingens superantigen Mycoplasma arthritidis mitogen T cell Superantigens Mouse mammary tumour viruses (Mtv) V Vb V TcR from MHC A haplotype Cell-tethered superantigen encoded by the viral genome Class II from MHC A to Z haplotypes APC Endogenous superantigens Mouse mammary tumour viruses (MMTV) Retroviruses that contain an open reading frame in a 3’ long terminal repeat that encodes a superantigen associated with the cell surface of APC Most mice carry 2-8 integrated MMTV proviruses in their genome Integrated MMTV Mtv-1, 2, 3, 6, 7 (Mls-1a), 8, 9, 11, 13 & 43 Infectious and transmitted by milk MMTV (C3H) MMTV (SW) MMTV (GR) Endogenous superantigen V-relationship Mtv Mtv 8 Mtv 11 Mtv 9 Mtv 6 Mtv 1 Mtv 3 Mtv 13 Mtv 7 MMTV SW MMTV C3H MMTV GR Murine V region 11 11 5.1, 5.2, 11 3, 5.1, 5.2 3 3 3 6, 7, 8.1, 9 6, 7, 8.1, 9 14 14 Stimulate T cells in a similar manner to exogenous supernatigens Valuable tools in analysis of self tolerance Mtv act in a similar manner to exogenous superantigens in vitro STIMULATOR CELLS Mtv-7 +ve RESPONDING T CELLS Mtv-7 -ve Irradiated T APC Mtv-7 superantigen T T T T T T T Only T cells with TcR containing V6, V8.1 and V9 proliferate Mtv-7 interacts with V 6, V8.1 and V9 and activates only cells bearing those TcR Selective expansion of cells bearing certain V chains How do pathogens use superantigens? Unfocussed adaptive immune response activates cells of all specificities as well as those specific for the superantigens • Reduces the possibility that effective T cell clonal selection can eliminate the pathogen • Upon resolution, cells activated by the superantigen die, leaving the host immunosuppressed Transmission of infection Transmission of infection T 1. MMTV infected, MHC class II positive B cells 2. Massive T cell response to MMTV superantigen 3. Vigorous T cell help leads to B cell proliferation and differentiation to longlived B cells B 4. Infected cells traffic to mammary gland and infect young via milk Analysis of negative selection in vivo. Mtv Mtv-7 superantigen negative Immature CD4+8+ thymocytes expressing VV8.1 and V9 in the thymus Negative selection Mature CD4+ or CD8+ VV8.1 and V9 T cells in periphery T H Y M U S Mtv-7 superantigen positive Mtv-7 superantigen binds to V6, V8.1 and V9+ve thymocytes Immature CD4+8+ thymocytes expressing VV8.1 and V9 in the thymus Negative selection No mature CD4+ or CD8+ PERIPHERY VV8.1 and V9 T cells in periphery Analysis of negative selection in vivo. Milk transmissible superantigens - MMTV (C3H) V14 present? Male or female B10.BR Yes Male or female C3H No X Female B10.BR Male C3H V14 present? F1 offspring Yes X Female C3H F1 offspring Male B10.BR V14 present? No Deletion of V14 T cells in mice infected with MMTV by milk V14 present in fostered pups? + Young male or female C3H Foster female B10.BR + Young male or female C3H Or B10.BR Yes No Foster female C3H MMTV transmitted to fostered pups by infected B cells found in milk Are the signals that induce positive & negative selection the same, or different? SAME specificity T H Y M U S Immature thymocytes Positive selection Negative selection Peripheral T cells DIFFERENT specificity X Hypotheses of self-tolerance Avidity hypothesis Affinity of the interaction between TcR & MHC Density of the MHC:peptide complex on the cell surface Quantitative difference in signal to thymocyte. Differential signalling hypothesis Type of signal that the TcR delivers to the cell Qualitative difference in signal to thymocyte. Removal of useless cells Peptide is not recognised or irrelevant Thymocyte receives no signal, fails to be positively selected and dies by apoptosis. WEAK OR NO SIGNAL T cell TcR CD8 MHC Class I Thymic epithelial cell Positive selection Peptide is a partial agonist Thymocyte receives a partial signal and is rescued from apoptosis i.e. the cell is positively selected to survive and mature. PARTIAL SIGNAL T cell TcR CD8 MHC Class I Thymic epithelial cell Negative selection Peptide is an agonist Thymocyte receives a powerful signal and undergoes apoptosis i.e. the cell is negatively selected and dies. FULL SIGNAL T cell TcR CD8 CD8 MHC Class I Thymic epithelial cell The thymus accepts T cells that fall into a narrow window of affinity for MHC molecules Useless Neglect Useful Harmful Positively select Negatively select Number of cells Low High Affinity of TcR/MHC interaction Positive & negative selection occurs in distinct thymic microenvironments Cortex Proliferation CD3Positive selection CD3+ TcR+ Negative selection DN DP Cortical epithelial cells DP Medulla CD3+ TcR+ SP CD3+ TcR+ CD8+ or CD4+ Immature double negative & positive thymocytes Dendritic cells medullary Epithelial cells & Macrophages Mature single positive thymocytes How accurate are these models of positive and negative selection? Positive selection: Relied on very complex chimera experiments Relied on proof of MHC restriction as an outcome which is tested in an ‘unnatural’ response using MHC mismatched presenting cells Negative selection: Relied on exceptionally powerful superantigens operating outside the normal mechanisms of antigen recognition Illustration of selection using TcR transgenic mice Generation of transgenic mice T T cell clone with known TcR specificity and MHC restriction Rearranged chain cDNA construct Rearranged chain cDNA construct Re-implant } Analyse offspring for transgene expression. Inject into fertilised mouse ovum In TcR transgene-expressing mice almost all thymocytes express the transgenic TcR due to ALLELIC EXCLUSION. Cells that fail positive selection die in the thymus (neglect) In TcR transgenic mice expressing an MHC A restricted TcR, all thymocytes express the MHC A restricted TcR DN MHC B DP Transgenically express MHC A restricted TcR in an MHC B mouse CD3- CD3+ TcR+ SP No single +ve cells are present in the periphery CD3+ TcR+ CD8+ or CD4+ Thymocytes die at the double positive stage after failing +ve selection due to a lack of MHC A Positive selection determines the restriction element of the TcR AND the expression of CD4 or CD8 TcR transgenic mouse TcR transgenic mouse TcR from MHC class Irestricted T cell TcR from MHC class IIrestricted T cell Only CD8 cells mature Only CD4 cells mature Restriction element and co-receptor expression are co-ordinated Instructive model: Signal from CD4 silences the CD8 expression & vice versa? Stochastic/selection model: Cells randomly inactivate CD4 or CD8 gene, then test for matching of TcR restriction with co-receptor expression? Double positive to single positive transition Single CD4+ thymocyte Double positive thymocyte -ve TcR TcR 3 CD4 2 MHC Class I TcR √ X CD8 TcR 3 CD8 CD4 2 MHC Class II MHC Class I MHC Class II Thymic epithelial cell Instructive model: Signal from CD4 silences the CD8 expression & vice versa Double positive to single positive transition Single CD4+ thymocyte Double positive thymocyte TcR TcR 3 CD4 2 MHC Class I TcR √ X CD8 TcR 3 CD8 CD4 2 MHC Class II MHC Class I MHC Class II Thymic epithelial cell Stochastic/selection model: Cells randomly inactivate CD4 or CD8 gene, whilst testing a match of TcR restriction Deletion of cells in the thymus: differential effect on the mature and immature repertoire TcR transgenic mouse TcR from T cell specific for hen egg lysosyme (HEL) ~100% of T cells/thymocytes express anti-HEL TcR Immunise with HEL Analyse peripheral Analyse thymus: T cells: All transgenic T cells die All transgenic T cells by apoptosis proliferate Thymocytes activated by antigen in the thymic environment die T cells activated by antigen in the periphery proliferate How can the thymus express all self antigens – including self antigens only made by specialised tissues? How do we become self tolerant to these antigens? Nature Immunology November 2001 Promiscuous expression of tissue-specific genes by medullary thymic epithelial cells How is self tolerance established to antigens that can not be expressed in the thymus? • T cells bearing TcR reactive with proteins expressed in the thymus are deleted. • Some self proteins are not expressed in the thymus e.g. antigens first expressed at puberty • Self tolerance can be induced outside the thymus PERIPHERAL TOLERANCE or ANERGY A state of immunological inactivity caused by a failure to deliver appropriate signals to T or B cells when stimulated with antigen i.e. a failure of antigen presenting cells to deliver COSTIMULATION T helper cells costimulate B cells Two - signal models of activation Signal 1 antigen & antigen receptor B Y CD40 ACTIVATION Th Signal 2 - T cell help T cell antigen receptor Co-receptor (CD4) MHC class II and peptide CD40 Ligand (CD154) Antigen presentation - T cells are co-stimulated Signal 1 antigen & antigen receptor Th APC ACTIVATION Signal 2 B7 family members (CD80 & CD86) CD28 Costimulatory molecules are expressed by most APC including dendritic cells, monocytes, macrophages, B cells etc., but not by cells that have no immunoregulatory functions such as muscle, nerves, hepatocytes, epithelial cells etc. Mechanism of co-stimulation in T cells Low affinity IL-2 receptor Antigen 1 IL-2 IL-2 IL-2R IL-2R Resting T cells Express IL-2 receptor and chains but no chain or IL-2 Signal 1 NFAT binds to the promoter of of the chain gene of the IL-2 receptor. The chain converts the IL-2R to a high affinity form Mechanism of co-stimulation in T cells Costimulation Antigen Signal 2 1 2 Activates AP-1 and NFk-B to increase IL2 gene transcription by 3 fold Stabilises and increases the half-life of IL2 mRNA by 20-30 fold IL-2 IL-2R IL-2 production increased by 100 fold overall Immunosuppressive drugs illustrate the importance of IL-2 in immune responses Cyclosporin & FK506 inhibit IL-2 by disrupting TcR signalling Rapamycin inhibits IL-2R signalling Anergy Antigen Naïve T cell 1 Signal 1 only IL-2 IL-2R Epithelial cell Self peptide epitopes presented by a non-classical APC e.g. an epithelial cell The T cell is unable to produce IL-2 and therefore is unable to proliferate or be clonally selected. Unlike immunosupressive drugs that inhibit ALL specificities of T cell, signal 1 in the absence of signal 2 causes antigen specificT cell unresponsiveness. Arming of effector T cells Clonal selection and differentiation APC T Activation of NAÏVE T cells by signal 1 and 2 is not sufficient to trigger effector function, but….. the T cell will be activated to proliferate and differentiate under the control of autocrine IL-2 to an effector T cell. These T cells are ARMED IL-2 Effector T cell How can this cell give help to, or kill cells, that express low levels of B7 family costimulators? Effector function or Anergy? Clonally selected, proliferating and differentiated T cell i.e. ARMED sees antigen on a B7 -ve epithelial cell IL-2 Armed Effector T cell The effector programme of the T cell is activated without costimulation Armed Effector T cell This contrasts the situation with naïve T cells, which are anergised without costimulation Naïve T cell CD28 TcR Co-receptor Kill Epithelial cell Epithelial cell Epithelial cell Costimulatory molecules also associate with inhibitory receptors T cell Signal 1 + 2 2 Activated T cell -- -- CD28lo CTLA-4hi CD28 B7 B7 CD28 cross linked by B7 Co-stimulation induces CTLA-4 Cross-linking of CTLA-4 by B7 inhibits co-stimulation and inhibits T cell activation CTLA-4 binds CD28 with a higher affinity than B7 molecules The lack of signal 2 to the T cell shuts down the T cell response. The danger hypothesis & co-stimulation Full expression of T cell function and self tolerance depends upon when and where co-stimulatory molecules are expressed. Cell containing only self antigens No danger Apoptotic cell death. A natural, often useful cell death. No danger APC APC Innocuous challenge to the immune system fails to activate APC and fails to activate the immune system Fuchs & Matzinger 1995 The danger hypothesis Necrotic cell death e.g. tissue damage, virus infection etc APC DANGER Pathogens recognised by microbial patterns APC APC that detect ‘danger’ signals express costimulatory molecules, activate T cells and the immune response How the danger hypothesis suggests a review of immunological dogma • Antigens induce tolerance or immunity depending upon the ability of the immune system to sense them as ‘dangererous’, and not by sensing whether they are self or ‘non-self’. • There is no window for tolerance induction in neonates - if a ‘danger signal’ is received, the neonatal immune system will respond • Neonatal T cells are not intrinsically tolerisable but the natural antiinflammatory nature of the neonatal environment predisposes to tolerance • Apoptosis, the ‘non-dangerous’ death of self cells may prevent autoimmunity when old or surplus cells are disposed of. • Suggests that tolerance is the default pathway of the immune system on encountering antigens. • Explains why immunisations require adjuvants to stimulate cues of danger such as cytokines or costimulatory molecule expression. Doesn’t exclude self-nonself discrimination, but the danger hypothesis will be very hard to disprove experimentally.